Una mirada macroecológica del riesgo de pérdida evolutiva de los roedores de Chile
A macroecological perspective on the risk of evolutionary loss of Chilean rodents
Wendy C. Hernández-Mazariegos a, *, Felipe I. Torres b, c, Jennifer Catalán a, d y Christian M. Ibáñez d
a Universidad Andres Bello, Facultad de Ciencias de la Vida, Programa de Doctorado en Medicina de la Conservación, Avenida República 440, 8370251 Santiago, Chile
b Universidad Católica de la Santísima Concepción, Programa de Doctorado en Ciencias Mención Biodiversidad y Biorecursos, Alonso de Ribera 2850, Concepción, Chile
c Instituto Milenio en Socio-Ecología Costera, Avenida Libertador Bernardo O’Higgins 340, Santiago, Chile
d Universidad Andres Bello, Facultad de Ciencias de la Vida, Instituto One Health, Avenida República 440, 8370251 Santiago, Chile
*Autor para correspondencia: wendychm7@gmail.com (W.C. Hernández-Mazariegos)
Recibido: 21 abril 2025; aceptado: 25 agosto 2025
Resumen
La pérdida de biodiversidad es un fenómeno global, pero sus impactos y respuestas varían según las regiones y grupos taxonómicos. En este estudio, aplicamos el enfoque EDGE (distinción evolutiva y en peligro global) para evaluar la priorización de la conservación de los roedores de Chile, altamente diversos y con alto endemismo. Calculamos los valores EDGE a partir de datos filogenéticos y del estado de conservación de 61 especies y analizamos su relación con el tamaño corporal y el rango de distribución mediante regresión filogenética de mínimos cuadrados. Además, examinamos la distribución espacial de la distinción evolutiva (ED) y el índice EDGE a lo largo del gradiente latitudinal de Chile. Nuestros resultados muestran que las especies con baja ED, pero alto peligro global (GE) presentan altos valores EDGE y que existe una asociación negativa entre EDGE y el tamaño del rango de distribución. A nivel espacial, encontramos altos valores de ED en el norte y sur del país, y altos valores de EDGE en el norte y centro-sur. Estos patrones destacan la necesidad de considerar tanto la historia evolutiva como las amenazas actuales para guiar estrategias de conservación más eficaces, especialmente en regiones subrepresentadas en los esquemas globales.
Palabras clave: Biogeografía; Conservación; Filogenia; Mastozoología; Rasgos
Abstract
Biodiversity loss is a global phenomenon, but its impacts and responses vary across regions and taxonomic groups. In this study, we applied the EDGE (Evolutionarily Distinct and Globally Endangered) framework to assess priorities for the conservation of rodents in Chile, a highly diverse and largely endemic group. We calculated EDGE scores using phylogenetic data and conservation status of 61 species and analyzed their relationship with body size and geographic range through phylogenetic least squares regression. We also examined the spatial distribution of Evolutionary Distinctiveness (ED) and EDGE scores along Chile’s latitudinal gradient. Our results show that species with low ED but high Global Endangerment (GE) tend to have high EDGE values, and that EDGE is negatively associated with range size. Spatially, we found high ED values in northern and southern Chile, and high EDGE scores in the north and south-central regions. These patterns highlight the importance of considering both evolutionary history and current threats to inform more effective conservation strategies, particularly in regions underrepresented in global prioritization schemes.
Keywords: Biogeography; Conservation; Phylogeny; Mammalogy; Traits
Introducción
Actualmente, la biodiversidad enfrenta amenazas sin precedentes, por lo que entender los patrones y procesos que determinan la distribución de las especies se ha vuelto urgente (Johnson et al., 2024). La biogeografía y macroecología han avanzado significativamente en la comprensión de los patrones y procesos que determinan la distribución de las especies y sus interacciones a distintas escalas espaciales y temporales (Lomolino et al., 2017). Aplicadas a la conservación, estas disciplinas permiten estimar los factores que influyen en la distribución de las especies, la dinámica poblacional, el ensamblaje de comunidades y las tasas de especiación y extinción. Su integración facilita la conexión entre el conocimiento teórico sobre la biodiversidad y su aplicación en la toma de decisiones informadas para la conservación (Santini et al., 2021).
La macroecología y la conservación, aunque inicialmente disciplinas separadas, han convergido progresivamente debido a la creciente necesidad de comprender los efectos antropogénicos y el cambio climático global sobre la biodiversidad (Santini et al., 2021). En particular, la macroecología ha proporcionado un enfoque a gran escala que permite evaluar procesos ecológicos regionales y globales, contribuyendo así a estrategias de conservación más efectivas (Santini et al., 2021).
La extinción de las especies representa una gran amenaza para la diversidad de linajes filogenéticos, ya que implica la pérdida irreversible de estos grupos únicos, representada por especies en peligro crítico con alta distinción evolutiva (especies EDGE: Evolutionarily Distinct and Globally Endangered) (Gumbs et al., 2024). Las especies EDGE son especies en peligro de extinción, destacan por su singularidad morfológica (en apariencia), ecológica (comportamiento) y genética (pocas especies filogenéticamente cercanas) (EDGE, 2024; Isaac et al., 2007; Mooers et al., 2008). La conservación de los linajes de las especies en peligro crítico es crucial y fundamental, ya que su desaparición no solo reduce la biodiversidad, sino también la variedad de estrategias ecológicas y funciones ecosistémicas, esenciales para la estabilidad ambiental y el bienestar humano (Gumbs et al., 2023; Tucker et al., 2019; WWF, 2020).
El riesgo de extinción no es aleatorio dentro de los linajes evolutivos, ya que especies emparentadas suelen compartir estados de amenazas y rasgos biológicos que los hacen más vulnerables —e.g., tamaño corporal, distribución geográfica— (Maestri et al., 2016; Meiri et al., 2018; Smith et al., 2004). La susceptibilidad de la pérdida de algunos linajes podría deberse a una fuerte señal filogenética de estos rasgos (Fritz y Purvis, 2010). Es decir, la señal filogenética de rasgos podría permitir predecir la probabilidad de pérdida evolutiva en función de los atributos compartidos entre especies (Fritz y Purvis, 2010; Isaac et al., 2007).
Algunos rasgos biológicos como el tamaño corporal y el tamaño de distribución han sido reportados en mamíferos como predictores para el riesgo de extinción (Chichorro et al., 2022; Ripple et al., 2017), los cuales también han sido reportados con fuerte señal filogenética, aunque el tamaño de distribución ha presentado resultados variables y lábiles entre especies (Avaria-Llautureo et al., 2012; Pie y Meyer, 2017; Rosauer et al., 2009). Asimismo, se ha documentado que la variación del tamaño corporal y tamaño de distribución está asociada a gradientes geográficos, como la latitud (regla de Bergmann, Blackburn et al., 1999) y altitud (regla de Rapoport, Stevens, 1992). Estos rasgos podrían no solo predecir el riesgo de extinción de las especies, sino también el riesgo de pérdida evolutiva y mostrar un patrón de distribución influenciado por las variables geográficas.
Actualmente, el riesgo de pérdida evolutiva es medido a través del índice EDGE (Isaac et al., 2007). Este índice combina la distinción evolutiva (ED) (diversidad filogenética) (Faith, 2019; Isaac et al., 2007) y el peligro global (GE) de conservación, según la Unión Internacional para la Conservación de la Naturaleza (UICN, 2024). Estudios previos han aplicado este enfoque a diversos grupos de vertebrados, incluyendo anfibios, aves, peces y mamíferos (Cortés-Díaz et al., 2023; Cheng et al., 2014; Gumbs et al., 2018, 2024; Stein et al., 2018). Entre los vertebrados terrestres, los mamíferos presentan el mayor número de familias en riesgo de extinción (166 especies en riesgo) y ocupan el cuarto lugar con mayor porcentaje de especies EDGE (~ 10%, ~ 640 especies EDGE/5,674 especies), después de los crocodilios, testudines y anfibios (EDGE, 2024; Gumbs et al., 2024).
Dentro de los mamíferos, se reporta que los roedores (orden Rodentia) presentan más de 100 especies catalogadas como EDGE (Kennerley et al., 2021). A pesar de que los roedores constituyen el grupo más diverso y ampliamente distribuido, con 2,664 especies vivientes (Kennerley et al., 2021; Llobet et al., 2023), se reporta un alto índice de extinción (~ 50 especies en los últimos 5 siglos) y un alto número de especies en riesgo crítico de amenaza (324, 15%) (Kennerley et al., 2021; MacPhee y Flemming, 1999; Teta et al., 2014). Además, a pesar de su amplia distribución geográfica, se ha reportado que los roedores con rangos geográficos restringidos están asociados con estados de conservación vulnerables a la extinción (Fourcade y Alhajeri, 2023; Iriarte, 2007; Patton et al., 2015; UICN, 2024). La vulnerabilidad a la extinción se debe principalmente a la pérdida y modificación del hábitat, pero además, a la poca atención y enfoque en su conservación (Kennerley et al., 2021; Ripple et al., 2017).
En Sudamérica, los roedores chilenos están representados por el grupo Caviomorpha y Sigmodontinae, cuyos orígenes se remontan a hace más de 33 y 12 millones de años, respectivamente (Patton et al., 2015; Poux et al., 2006; Rivera et al., 2023; Upham y Patterson, 2015). Su historia evolutiva ha dado lugar a una gran diversidad morfológica, ecológica y de distribución (Rivera et al., 2023), y por lo tanto, a una gran diversidad para responder a las distintas amenazas. Sin embargo, en Chile los roedores son el grupo con mayor riesgo de amenaza dentro de los mamíferos terrestres (32% en categorías de vulnerable, en peligro, en peligro crítico) (Gaulke et al., 2019; UICN, 2024).
En general, para los mamíferos chilenos, tanto grandes (> 320 g) como pequeños (< 250 g), se ha documentado que el riesgo de amenaza se ve afectado por la variabilidad del tamaño corporal de las especies (Cofré y Marquet, 1999). Específicamente en los roedores, se reportan cambios en sus patrones de distribución, ya sea de ampliación y/o disminución (e.g., Abrothrix longipilis, Oligoryzomys longicaudatus) (Belmar-Lucero et al., 2009; Teta y Pardiñas, 2014). Sin embargo, se desconoce qué linajes son vulnerables al riesgo de pérdida evolutiva, la influencia de rasgos, como el tamaño corporal y de distribución, en la distinción evolutiva y el riesgo actual de amenaza, y dónde se localiza (geográficamente) el riesgo evolutivo de los roedores en Chile.
Dada la singular historia evolutiva de los roedores chilenos, variabilidad en rasgos morfológicos, amplia distribución latitudinal y altitudinal, y alto riesgo de amenaza, se espera que las especies con mayor riesgo de pérdida evolutiva (alto EDGE) sean aquellas de mayor tamaño corporal y con rangos de distribución reducidos. Esta expectativa se basa en que ambos rasgos han sido identificados como predictores del riesgo de extinción en mamíferos, y porque suelen presentar una señal filogenética (Pie y Meyer, 2017; Ripple et al., 2017). Asimismo, se espera que el riesgo acumulado de pérdida evolutiva sea mayor en latitudes australes y altitudes bajas, ya que estudios previos han demostrado que estos gradientes geográficos influyen en los rasgos biológicos antes mencionados (e.g., reglas de Bergmann y Rapoport), y que pueden reflejar patrones de distribución (Blackburn et al., 1999; Stevens, 1992).
En este sentido, el objetivo de este estudio fue evaluar la distinción evolutiva y riesgo de amenaza, a través del índice EDGE, de los roedores chilenos. Evaluar asociaciones entre el riesgo evolutivo y los rasgos biológicos (i.e., tamaño corporal y tamaño de distribución), y su distribución a lo largo del gradiente latitudinal de Chile. Para ello, utilizamos datos filogenéticos, morfológicos y de distribución, y del riesgo de amenaza según la lista roja nacional e internacional de las especies de roedores actualmente presentes en Chile.
Materiales y métodos
Se obtuvieron datos de especies de roedores pertenecientes a 7 familias distribuidas en Chile: Abrocomidae, Caviidae, Chinchillidae, Cricetidae, Ctenomyidae, Echimyidae y Octodontidae. Se tomó como lista base de especies de roedores chilenos la lista actualizada de mamíferos vivientes de Chile, reportada en D’Elía et al. (2020). A partir de esta lista se consideraron cambios taxonómicos y de nomenclatura reportados hasta 2024 (Iriarte, 2021; MDD, 2024). Los cambios considerados fueron los siguientes: no se incluyó a la especie Lagidium peruanum (Iriarte, 2021; MDD, 2024), se actualizó el nombre de Euneomys mordax a E. fossor (Teta et al., 2021) y se incluyeron 3 especies descritas recientemente: Oligoryzomys yatesi (Palma y Rodríguez-Serrano, 2018), Oligoryzomys flavescens (Quiroga-Carmona, González et al., 2023) y Punomys lemminus (Quiroga-Carmona, Storz et al., 2023) (material suplementario: tabla S1).
Los datos morfológicos del tamaño corporal, medidos como longitud total desde la nariz hasta la base del cuerpo (en milímetros), fueron obtenidos de la literatura (Duclos et al., 2024; Iriarte, 2007, 2021; Patton et al., 2015). La información sobre la distribución latitudinal (grados de latitud) se estimó a partir de los polígonos de distribución disponibles en la Lista Roja de la UICN (2024) y en Map of Life (Jetz et al., 2012). Estos polígonos fueron superpuestos en el mapa de Chile, que fue descargado de la Biblioteca del Congreso Nacional de Chile (BCN, 2024) y posteriormente divididos por grados de latitud para identificar los valores mínimos y máximos de cada especie. Este procedimiento fue realizado con el software QGIS, versión 3.38.3 (QGIS Development Team, 2024). Los datos de distribución altitudinal (m snm) se estimaron a partir de la literatura, identificando la altitud mínima y máxima registradas para cada especie (Duclos et al., 2024; Iriarte, 2007, 2021) (material suplementario: tabla S2). A partir de estos valores mínimos y máximos, tanto para la latitud como para la altitud, se calculó el tamaño del rango de distribución (valor máximo – valor mínimo), así como el promedio de distribución como (valor máximo + valor mínimo) / 2.
Para la reconstrucción filogenética, se descargaron las secuencias del marcador mitocondrial citocromo b [Cytb] de GenBank (2023) (material suplementario: table S2). Las secuencias de ADN fueron alineadas en MUSCLE (Edgar, 2004) implementado en el software MEGA X versión 10.2.6 (Kumar et al., 2018) utilizando parámetros predeterminados. El mejor modelo de sustitución nucleotídica se estimó utilizando el criterio de información bayesiano (BIC) en MEGA X.
Las relaciones filogenéticas de los roedores se examinaron mediante inferencia bayesiana (IB) para estimar las probabilidades a posteriori de los nodos en los árboles filogenéticos, utilizando MrBayes versión 3.2. (Ronquist et al., 2012). Se creó un bloque en el archivo nexus de partición por codón para genes codificantes incluyendo el modelo de sustitución seleccionado para el gen (i.e., GTR + G + I, BCI = 32,983.54). El análisis bayesiano se realizó utilizando 4 cadenas de Markov y Monte Carlo (MCMC), cada una con 10 millones de generaciones, muestreadas cada 1,000 generaciones en 2 corridas independientes. Los primeros 1,000 árboles de cada ejecución se descartaron como “burnin” y se calculó un consenso de los árboles restantes utilizando el criterio de la mayoría (50%). Por último, se hizo edición del árbol filogenético en FigTree versión 1.4 (Rambaut, 2009). Se usó a Rattus rattus, Rattus norvegicus y Mus musculus como grupos externos. Además, se verificó la presencia de politomías en el árbol filogenético mediante la función is.binary.tree() del paquete ‘ape’ (Paradis y Schliep, 2019) en el programa R versión 4.4.1 (R Core Team, 2024). En los casos de nodos no resueltos, estos fueron transformados en dicotomías mediante la función multi2di() del paquete ‘ape’.
La clasificación del estado de amenaza de cada especie se determinó según la clasificación de la Lista Roja de la UICN (2024). Las especies que no contaban con una clasificación dada por la UICN, se clasificaron según el Reglamento para la Clasificación de Especies – RCE, del Estado de Conservación de Chile, publicado por el Ministerio del Medio Ambiente (MMA, 2024) (material suplementario: tabla S3) y las que no contaban con clasificación por la UICN (2024) y RCE (MMA, 2024), se clasificaron como no evaluadas, siguiendo las Directrices para el Uso de Categorías y Criterios de la Lista Roja de la (UICN Standards and Petitions Committee, 2024).
Para la identificación de especies EDGE se requirieron 2 análisis: distinción evolutiva de las especies (ED, Evolutionarily Distinct) y peligro global (GE, Globally Endangered). Se determinó ED por “proporción justa” de cada especie usando la función evol.distinct del paquete ‘picante’ (Kembel et al., 2020), implementando en el programa R. La medida “proporción justa” consiste en la suma del largo de las ramas desde la raíz hasta cada punta dividida por el número de especies que descienden de cada rama (Isaac et al., 2007), lo cual fue determinado con el paquete ‘ape’ en R. Para determinar GE las clasificaciones de UICN y RCE, se transformaron en valores numéricos propuestos por Isaac et al. (2007): menor preocupación – LC = 0, casi amenazado – NT = 1, vulnerable – VU = 2, en peligro – EN = 3 y críticamente en peligro – CR = 4. Con los valores ED y GE se identificaron las especies EDGE a través de la fórmula propuesta por Isaac et al. (2007): EDGE = ln (1 + ED) + (GE × ln (2)). Para el análisis no se tomó en cuenta a las especies clasificadas como DD (datos deficientes) y NE (no evaluadas) dadas las clasificaciones numéricas propuestas por Isaac et al. (2007).
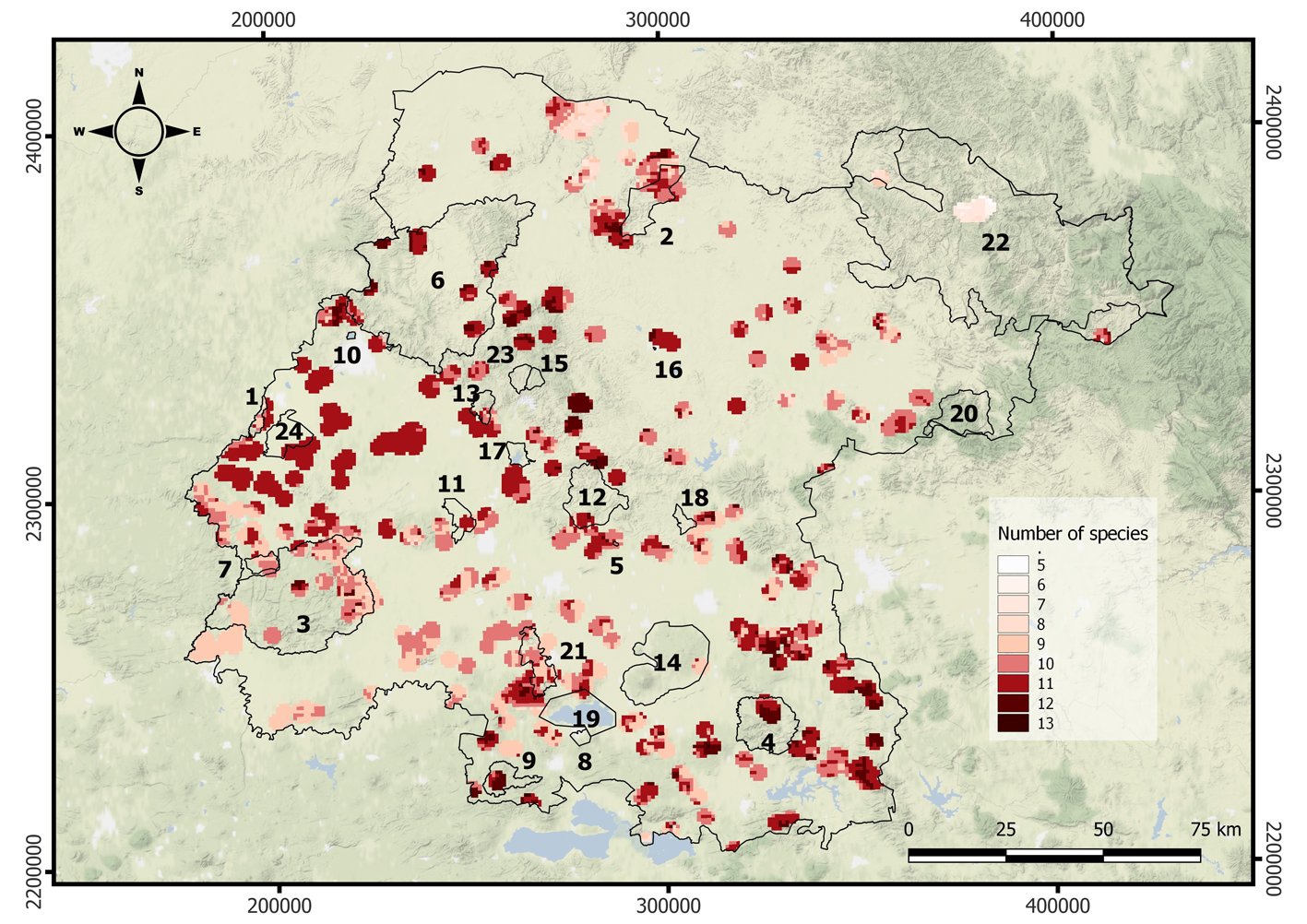
Con base en los resultados ED, EDGE y distribución latitudinal de las especies, se estimó su distribución en el país. Se construyó una matriz de presencia-ausencia en una escala de 1o de latitud para obtener el número total de ED y EDGE en cada segmento latitudinal. Se realizó un análisis de regresión lineal simple para evaluar la relación entre los EDGE y ED, y entre EDGE y GE, utilizando la función lm() del programa R. Además, se hizo una comparación geográfica visual de la distribución latitudinal de ED y EDGE con la distribución de las áreas terrestres protegidas de Chile. El mapa de las áreas protegidas fue obtenido de la página de la BCN (2024) y visualizado y editado en QGis.
Se estimó la señal filogenética para cada rasgo (tamaño corporal, rango y promedio de distribución) e índices (ED y EDGE) utilizando lambda de Pagel (λ), en el paquete ‘Phytools’ (Revell, 2012) implementado en R. El parámetro lambda varía entre 0 y 1 y cuantifica la cantidad de señal filogenética en el rasgo estudiado: cuando λ = 0, la distribución del rasgo entre las especies es independiente de su filogenia, mientras que λ = 1 indica que la distribución del rasgo coincide estrechamente con la expectativa del modelo de movimiento browniano (Pagel, 1999, 2002). Además, se realizó un mapa de calor filogenético para mostrar la distribución de rasgos de los roedores respecto a su filogenia, usando el paquete ‘Phytools’ en R.
Para determinar la asociación EDGE con los rasgos de tamaño corporal (mm) y de distribución latitudinal y altitudinal (rango y promedio), se realizaron análisis de mínimos cuadrados filogenéticos generalizados (PGLS: Pagel, 1999), usando los paquetes ‘phytools’ y ‘ape’ en R. Asimismo, se evaluó la asociación de ED con los rasgos. Se consideraron EDGE y ED como variables respuesta y el tamaño corporal y de distribución como predictoras.
Además, se hizo el análisis de regresión lineal simple para determinar relaciones entre los rasgos del tamaño corporal de los roedores con la latitud promedio de distribución y entre el tamaño de rango latitudinal con la distribución altitudinal promedio, en R. Para los análisis, cada rasgo se transformó a log 10 para disminuir el efecto de los valores atípicos e igualar la varianza.
Resultados
Se compilaron datos de 69 especies de roedores de Chile. El tamaño corporal presentó una variación que osciló entre 77 mm (i.e. Eligmodontia dunaris) y 500 mm (i.e. Myocastor coypus). La distribución de las especies estuvo comprendida entre los 18 y 55° de latitud, mostrando rangos de distribución latitudinal entre 1 y 35° de extensión, mientras que la distribución altitudinal varió entre 0 y 6,700 m snm (material suplementario: tabla S2).
El árbol de consenso evidenció altas probabilidades a posteriori (pp > 0.9), en la mayoría de los clados, considerando el grupo externo incluido para enraizar el árbol (fig. 1). El árbol filogenético mostró 2 clados: clado 1 correspondiente al grupo Caviomorpha, conformado por 27 especies de 6 familias (Caviidae, Chinchillidae, Abrocomidae, Ctenomyidae, Echimyidae, Octodontidae) (Poux et al., 2006); y un clado 2 correspondiente al grupo Sigmodontinae, conformado por 41 especies de una familia (Cricetidae) (Patton et al., 2015). Es decir, el árbol filogenético se construyó con 68 especies dado que fueron las que tenían información genética disponible (material suplementario: tabla S2).
Según la clasificación internacional UICN y nacional RCE de Chile, 45 especies se clasifican en LC, 5 en NT, 7 como VU, 3 como EN, 1 en CR, 3 como DD y 5 como NE (material suplementario: tabla S3).
La puntuación media de distinción evolutiva (ED) para los roedores fue 0.1684 (desviación estándar = 0.110) y clasificó a Reithrodon auritus (ED: 0.4916, LC), Chinchillula sahamae (ED: 0.4852, LC), Octodontomys gliroides (ED: 0.4594, LC), Abrocoma cinerea (ED: 0.4445, LC) y Myocastor coypus (ED: 0.4277, LC) con los valores más altos. Mientras que la vinculación de ED y GE clasificó a las especies Octodon pacíficus (EDGE: 2.84, CR), Chinchilla lanigera (EDGE: 2.24, EN), Chinchilla chinchilla (EDGE: 2.21, EN), Eligmodontia dunaris (EDGE: 2.18, EN) y Lagidium wolffsohni (EDGE: 1.55, VU) con los valores más altos. Para el análisis de ED se incluyeron las especies que tenían información genética (n = 68 especies) y para el de EDGE a las especies que tenían información genética y clasificación LC, NT, VU, EN y CR (n = 61) (material suplementario: tabla S3).
Respecto de la distribución latitudinal de la ED, se encontró un alto ED (> 0.15) en el norte (18 a 20o S) y en el sur del país (55o S). Mientras que el riesgo de pérdida evolutiva (EDGE, > 0.4) se identificó en el norte (28 a 29o S) y centro-sur (38o S) del país (fig. 2). No se encontró asociación entre EDGE y ED (r2 = -0.0025, p > 0.05), pero sí entre EDGE y GE (r2 = 0.9823, p < 0.05).
La señal filogenética de Pagel fue alta y significativa para ED (λ = 0.99) y para el tamaño corporal (λ = 0.89); y fue baja pero significativa para el promedio de distribución altitudinal (λ = 0.55) (tabla 1). Respecto al mapa de calor del árbol filogenético y rasgos de los roedores, se identificaron clados según similitud del tamaño corporal, los cuales corresponden a grupos según el género. Por ejemplo, el género Lagidium con longitudes entre 480 y 460 mm, Aconaemys entre 180 y 170 mm, Geoxus entre 120 y 110 mm, Eligmodontia entre 105 a 77 mm (fig. 3, tabla 2).
El análisis PGLS de la asociación entre ED y los rasgos no mostró una asociación significativa (p > 0.05). Sin embargo, el análisis de EDGE con los rasgos mostró una asociación negativa con el tamaño del rango de distribución latitudinal (b = -0.57, p < 0.05; tabla 2, fig. 4A), y con el promedio de distribución altitudinal (b = -0.60, p < 0.05; tabla 2, fig. 4B). No se observó asociación entre el tamaño corporal de los roedores con la latitud promedio de distribución (regla de Bergmann, p > 0.05) y tampoco entre el tamaño de rango latitudinal con la altitud promedio de distribución (regla Rapoport, p > 0.05).
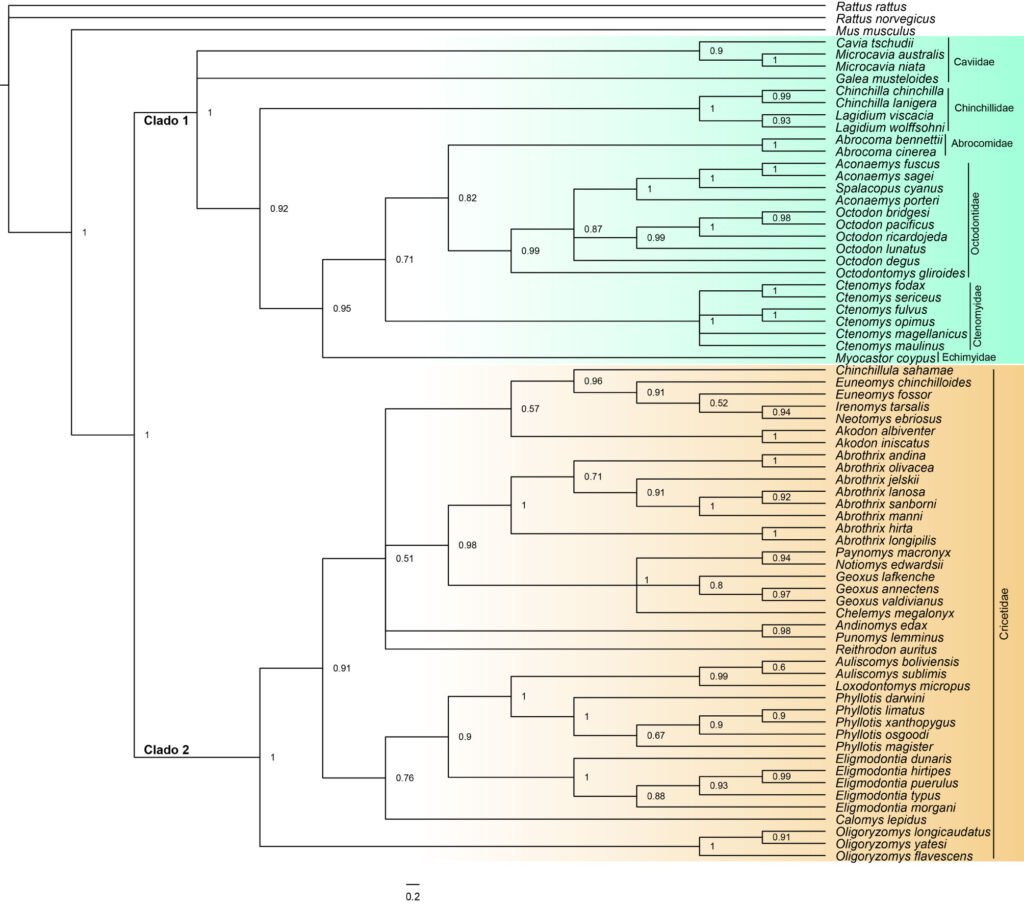
Figura 1. Relaciones filogenéticas de 68 especies de roedores chilenos, basadas en inferencia bayesiana utilizando marcador mitocondrial citocromo b [Cytb]. Clado 1 (color verde) representa al grupo Caviomorpha, clado 2 (color anaranjado) a los Sigmodontinae. Los valores de los nodos indican la probabilidad posterior bayesiana. Líneas verticales indican las familias a las que pertenece cada especie.
Discusión
Nuestros resultados muestran que, si bien la mayoría de las especies de los roedores de Chile presentan valores EDGE relativamente bajos y, por tanto, no se encuentran entre las prioridades de conservación a nivel global, existen excepciones críticas (e.g., Octodon pacificus y las chinchillas) que necesitan prioridades urgentes de conservación. Esto podría explicarse por la falta de información ecológica, genética y demográfica para muchas de las especies de roedores chilenos, lo que limita la precisión de su evaluación. En un contexto global de cambio ambiental acelerado, donde la incertidumbre sobre las tendencias reales de la respuesta de la biodiversidad persiste incluso en las especies con los conjuntos de datos más robustos (Johnson et al., 2024), es fundamental no interpretar la ausencia de señal como ausencia de riesgo. Esta advertencia cobra especial relevancia en regiones como Sudamérica, donde los vacíos de información son amplios y persistentes.
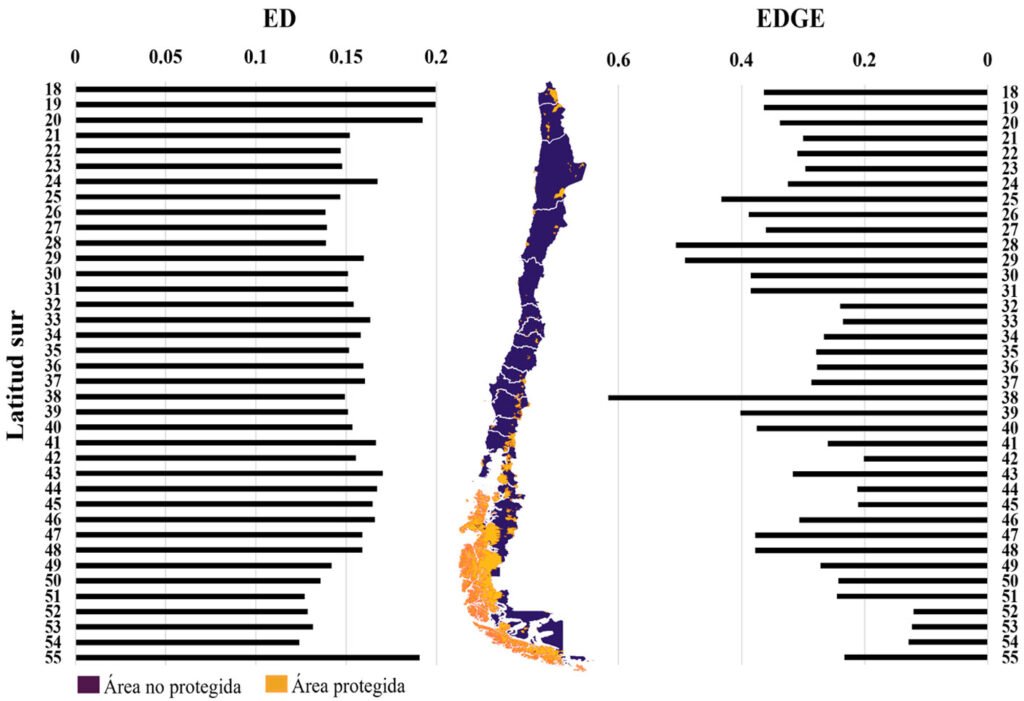
Figura 2. ED (distinción evolutiva) y EDGE (distinción evolutiva y peligro global) de los roedores a través del gradiente latitudinal de Chile.
Los resultados indican que las especies con los índices EDGE más altos son Octodon pacificus, Chinchilla lanigera, C. chinchilla, Eligmodontia dunaris y Lagidium wolffsohni; son especies categorizadas con un alto riesgo de amenaza (UICN, 2024; MMA, 2024; Spotorno et al., 2013). Esto concuerda con estudios previos que reportan que las especies con valores altos de EDGE suelen estar en mayor riesgo de extinción (Cortés-Díaz et al., 2023; Cheng et al., 2014; Gumbs et al., 2018; Owen et al., 2015; Stein et al., 2018).
Asimismo, nuestros resultados muestran una coincidencia parcial con el programa The EDGE of Existence (EDGE, 2024), la única iniciativa de conservación global enfocada en especies EDGE, ya que reconoce a C. lanigera, C. chinchilla y O. pacificus como especies EDGE, pero además incluye a Octodon bridgesi. Esta diferencia podría deberse, principalmente, a las discrepancias y a la falta de actualización en las clasificaciones del estado de amenaza de las especies a nivel nacional e internacional (UICN, 2024; MMA,
2024).
Tabla 1
Señal filogenética de los rasgos, ED (distinción evolutiva) y EDGE (distinción evolutiva y peligro global) de los roedores de Chile.
| Variables | Lambda | Valor p |
| Tamaño corporal | 0.8997 | < 0.001 |
| Rango latitudinal | < 0.001 | 1.0000 |
| Rango altitudinal | 0.0867 | 0.1569 |
| Promedio latitudinal | < 0.001 | 1.0000 |
| Promedio altitudinal | 0.5548 | 0.0154 |
| ED | 0.9918 | < 0.001 |
| EDGE | 0.0919 | 0.2744 |
Valor p < 0.05 fue considerado como significativo.
La discrepancia en las clasificaciones de estados de conservación es evidente: Octodon bridgesi ocupa el puesto 11 en el ranking EDGE, en Chile se considera en preocupación menor (MMA, 2024), pero a nivel internacional está categorizado como vulnerable (UICN, 2024). Lagidium wolffsohni que está clasificada como vulnerable a nivel nacional (MMA, 2024), internacionalmente se encuentra en la categoría de datos deficientes (UICN, 2024). La especie E. dunaris que no tiene una clasificación de amenaza a nivel nacional (MMA, 2024), ni internacional (UICN, 2024), ha sido sugerida como en peligro en el reporte de su descubrimiento como especie nueva en Chile (Spotorno et al., 2013). Esta discrepancia en las Listas Rojas puede influir en la clasificación numérica del riesgo de extinción seguida por Isaac et al. (2007) y, en consecuencia, en la priorización de especies EDGE.
En Chile, solo 25 especies de roedores tienen una clasificación oficial de amenaza a nivel nacional (MMA, 2024), mientras que a nivel internacional 17 especies están clasificadas como datos deficientes y no evaluadas (UICN, 2024). La falta de información en las clasificaciones y discrepancias del estado de conservación de las especies puede tener consecuencias directas en las estrategias de conservación locales. Si las decisiones de manejo se basan exclusivamente en los listados globales (UICN), especies que presentan amenazas significativas a nivel nacional podrían quedar fuera de las prioridades de conservación. Inversamente, especies clasificadas como amenazadas globalmente, pero no reconocidas en el contexto local, podrían no recibir atención ni medidas de protección. Esto pone en evidencia la necesidad de desarrollar estudios regionales de actualización de Listas Rojas para mejorar las evaluaciones del riesgo ecológico y evolutivo de las especies (Hidasi-Neto et al., 2015), fortalecer el monitoreo local y armonizar las evaluaciones nacionales e internacionales.
Tabla 2
Análisis de mínimos cuadrados filogenéticos generalizados (PGLS) entre ED (Distinción Evolutiva) y EDGE (Distinción Evolutiva y Peligro Global) y con los rasgos de los roedores de Chile.
| Variable | ED | EDGE | ||||
| Coeficiente | Error estándar | Valor p | Coeficiente | Error estándar | Valor p | |
| Intercepto | 0.0943 | 0.1963 | 0.6326 | 1.2636 | 2.0037 | 0.5309 |
| Tamaño corporal | 0.0757 | 0.0648 | 0.2472 | 0.6815 | 0.6049 | 0.2648 |
| Rango latitudinal | -0.0010 | 0.0124 | 0.9346 | -0.5713 | 0.2111 | 0.0091 |
| Rango altitudinal | -0.0021 | 0.0076 | 0.7788 | 0.0914 | 0.1382 | 0.5109 |
| Promedio latitudinal | 0.0129 | 0.0568 | 0.8212 | -0.1873 | 0.7045 | 0.7913 |
| Promedio altitudinal | 0.0142 | 0.0178 | 0.4283 | -0.6080 | 0.2754 | 0.0315 |
Valor p menor que 0.05 fue considerado como significativo.
Nuestros resultados indican que las especies EDGE identificadas pertenecen a los Caviomorpha, un linaje predominantemente sudamericano cuya diversificación se remonta al Eoceno-Oligoceno (~ 33 MA) (Poux et al., 2006; Rivera et al., 2023; Upham y Patterson, 2015). En comparación con los Sigmodontinae, los Caviomorpha presentan menor diversidad de especies (Maestri, 2020; Poux et al., 2006) y son los que han experimentado mayor número de extinciones a lo largo de su historia (Upham y Patterson, 2015). Específicamente en Chile, no se han registrado nuevas especies dentro de este grupo, lo que resalta la necesidad de medidas urgentes para su conservación.
Las especies EDGE identificadas en este estudio no presentaron los valores más altos de distinción evolutiva (ED). De hecho, las especies con mayor ED presentaron ramas filogenéticas más largas (e.g., R. auritus, C. sahamae, O. gliroides, A. cinerea, M. coypus), generalmente estaban representadas por una sola especie y clasificadas en menor preocupación (material suplementario: tabla S3). Esto contrasta con estudios previos que indican que las especies EDGE suelen ser las más distintas evolutivamente, con ramas largas, con pocas especies cercanas y con mayor riesgo de amenaza (Cortés-Díaz et al., 2023; Griffith et al., 2023).
Según nuestros resultados, el riesgo de pérdida evolutiva (alto EDGE) de las especies podría deberse más a las vulnerabilidades de las especies que a su distinción evolutiva (ED). Por ejemplo, la vulnerabilidad compartida con las especies con las que se agrupa, como el género Chinchilla, ambas especies (C. lanigera y C. chinchilla) están agrupadas en la filogenia y comparten un alto riesgo de amenaza (Roach y Kennerley, 2016a, b), dejando en evidencia el alto riesgo de perder el clado del género Chinchilla en Chile. La vulnerabilidad localizada: una especie puede tener muchas especies hermanas, ser evolutivamente distinta y vulnerable localmente (Huang et al., 2012). Esto podría explicar el riesgo evolutivo de O. pacificus, que filogenéticamente tiene varias especies cercanas, pero geográficamente es una especie aislada con riesgo crítico de amenaza (isla Mocha, Arauco; 38°22’ S, 73°55’ O) (Roach, 2016). Las especies con alto índice EDGE, identificadas en este estudio, presentaron bajo valor de ED y alto riesgo de amenaza (GE), sugiriendo que el alto riesgo de extinción (alto GE) las posiciona con alto riesgo evolutivo y prioritarias para la conservación. Este resultado sugiere que una especie con alto riesgo de amenaza (GE) y/o riesgo evolutivo (EDGE), debería ser priorizada para la conservación independientemente de si es o no distinta evolutivamente (ED).
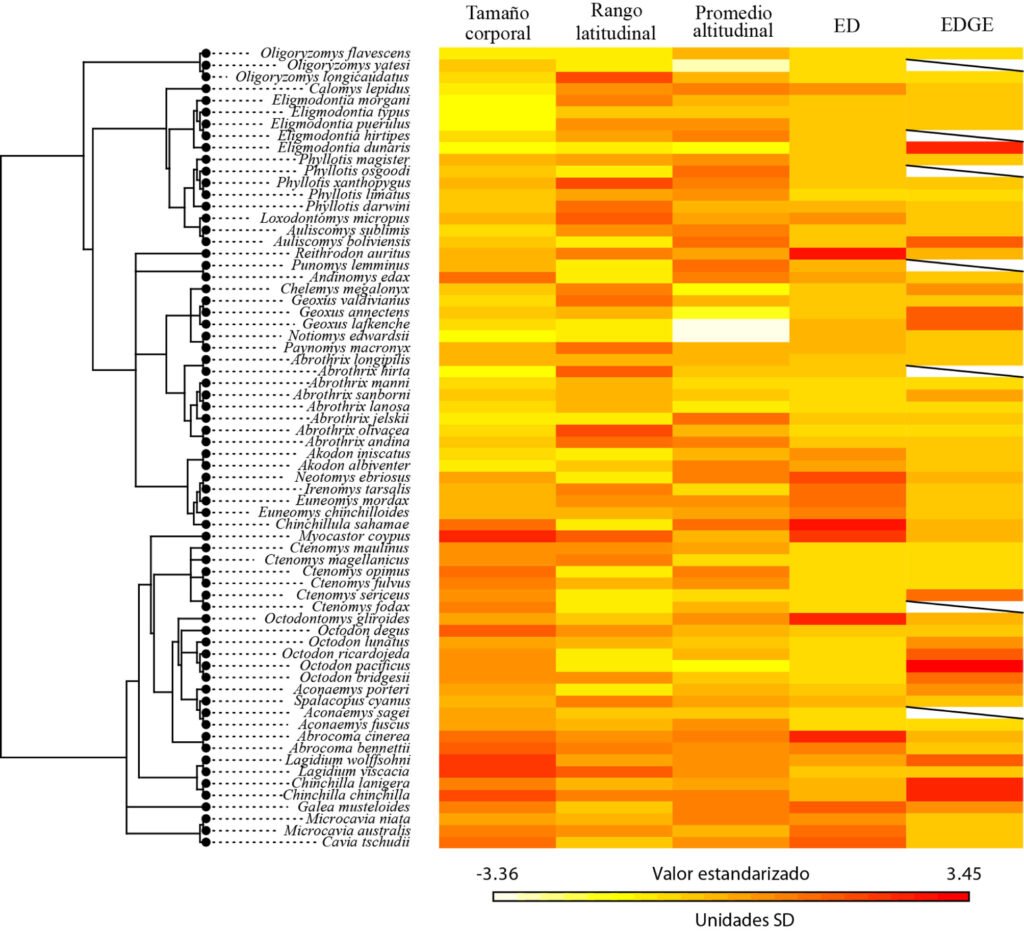
Figura 3. Mapa de calor filogenético de los roedores de Chile, que muestra los valores estandarizados (en unidades de desviación estándar, SD) de 3 rasgos (tamaño corporal, rango latitudinal y promedio altitudinal) junto con los índices de ED (distinción evolutiva) y EDGE (distinción evolutiva y peligro global). Los colores representan los valores estandarizados (media = 0, SD = 1) para cada especie, donde el amarillo indica valores bajos y el rojo valores altos. La línea diagonal en algunas celdas indica ausencia de datos para esa especie.
Asimismo, nuestros resultados sugieren que factores adicionales podrían estar contribuyendo a la situación de alto riesgo evolutivo de las especies. Estudios sugieren que la vulnerabilidad ecológica (e.g., especialización de hábitat, dietas restringidas, sensibilidad a perturbaciones), así como funciones biológicas (rasgos ecológicos inusuales o especializados) clave en los ecosistemas, están asociadas a la alta distinción evolutiva y mayor riesgo (Cooke et al., 2020; Redding et al., 2010). Estos resultados refuerzan la necesidad de incorporar métricas ecológicas junto con las evolutivas para una priorización de conservación más integral de las especies.
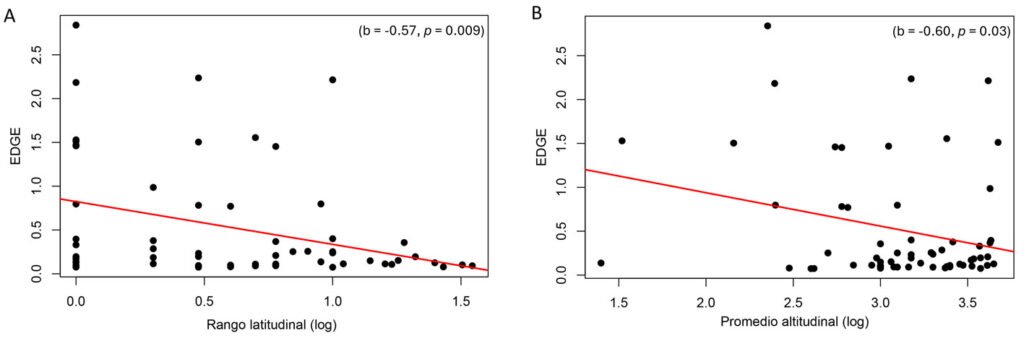
Figura 4. Análisis de mínimos cuadrados filogenéticos generalizados (PGLS) del índice EDGE (distinción evolutiva y peligro global) con el tamaño de rango latitudinal (A) y el promedio altitudinal (B) —valores transformados en logarítmicos. La línea roja representa la tendencia de PGLS.
Nuestros resultados indican que el tamaño corporal de los roedores tiene una alta señal filogenética (tabla 1), pero no hay asociación significativa con el índice EDGE (tabla 2), según el análisis PGLS. Estos hallazgos son consistentes con estudios previos que indican que los mamíferos, incluidos los roedores, tienden a agruparse filogenéticamente en función de su tamaño corporal, lo que probablemente responde a una tendencia evolutiva adaptativa (Avaria-Llautureo et al., 2012; Smith et al., 2004). Esto sugiere que las especies estrechamente relacionadas no necesariamente comparten probabilidades similares de riesgo de pérdida evolutiva en función de su tamaño corporal, aunque las similitudes de este rasgo entre especies se deban a sus ancestros compartidos.
Por otro lado, el tamaño de distribución, rango latitudinal y promedio altitudinal mostraron una baja señal filogenética (tabla 1), pero una asociación negativa con el índice EDGE (fig. 4, tabla 2). Los valores bajos de señal filogenética con los factores de distribución sugieren que no son rasgos heredados (Webb y Gaston, 2003, 2005). Mientras que la asociación del índice EDGE con latitud y altitud indica que el riesgo de pérdida evolutiva en los roedores chilenos podría depender de factores geográficos (fig. 4).
Los resultados son coincidentes con estudios que reportan que las variables geográficas se relacionan con los puntajes de EDGE y parcialmente con los rasgos heredables (e.g., tamaño corporal, rasgos reproductivos) (Redding et al., 2010). Lo anterior es inesperado ya que los rasgos heredables, como el tamaño corporal, reflejan la historia evolutiva (filogenia) de las especies, al contrario de los rasgos geográficos que son influenciados principalmente por procesos históricos, ecológicos o aleatorios (Avaria-Llautureo et al., 2012; Gaston, 2003; Smith et al., 2004). Probablemente la falta de relación del tamaño corporal con el índice EDGE se deba a que los subclados o clados menos inclusivos (es decir, clados por género) presentan muchas ramificaciones, lo que provoca un bajo ED. Al contrario de la relación de las variables geográficas con EDGE, que si bien las especies están cercanas en el árbol filogenético pueden estar en regiones o zonas geográficas completamente distintas (e.g., Octodon pacificus), y especies muy lejanas en el árbol pueden converger geográficamente. Estos resultados sugieren que los rasgos geográficos pueden estar más asociados con los valores EDGE que los rasgos hereditarios, como el tamaño corporal, convirtiéndolos en posibles indicadores para identificar especies EDGE.
El tamaño de rango de distribución latitudinal ha sido identificado como un predictor clave del riesgo de extinción en mamíferos, posiblemente debido a que no suele ser un rasgo taxón dependiente sino más bien dependiente de las condiciones del ecosistema y cambios climáticos antropogénicos (Chichorro et al., 2022; Smyčka et al., 2023). Por su parte, respecto a la distribución altitudinal, se observó que las especies a menor altitud presentan alto EDGE (fig. 4B). Se ha documentado que las especies que se distribuyen a mayores altitudes tienden a tener un rango altitudinal más amplio, lo que les permite tener una distribución geográfica más amplia y continua, y mejor resiliencia contra los impactos humanos (Keane et al., 2005). En este contexto, los factores geográficos (rango latitudinal y promedio de distribución altitudinal) son variables críticas respecto al riesgo evolutivo de los roedores de Chile.
Estudios previos han reportado asociaciones entre el tamaño corporal y variables geográficas en diversos taxones (Blackburn et al., 1999; Fourcade y Alhajeri, 2023; Maestri et al., 2016). La regla de Bergmann postula que el tamaño corporal de los animales homeotermos tiende a incrementarse con la latitud (Blackburn et al., 1999; Maestri et al., 2016). Mientras que la regla de Rapoport sugiere que el tamaño del rango de distribución aumenta con la altitud (Stevens, 1992). Sin embargo, nuestros resultados no evidenciaron una asociación entre el tamaño corporal de los roedores chilenos y la latitud, y tampoco entre el tamaño del rango de distribución y la altitud. Esto concuerda con estudios que indican que la relación entre el tamaño corporal y la latitud no siempre se ajusta en roedores de hemisferio sur; de hecho, el tamaño corporal de los roedores tiende a reducirse a medida que aumenta la latitud (Maestri et al., 2016). Asimismo, la falta de una correlación clara entre el tamaño del rango de distribución y la altitud podría deberse a la influencia de factores ambientales, lo que altera la relación esperada en elevaciones mayores (Maestri, 2020). Por ejemplo, en ecosistemas montañosos como la Cordillera de los Andes, la homogeneidad climática a gran escala podría influir en la distribución altitudinal de los roedores (Fourcade y Alhajeri, 2023; Maestri, 2020).
Áreas y riesgo de pérdida evolutiva. Identificamos áreas de altos valores de ED y EDGE en Chile que coinciden con regiones que han estado sujetas a procesos biogeográficos e históricos contrastantes. En el norte, la presencia del desierto de Atacama, uno de los más áridos del mundo, y la altitud de la Puna andina (~ 3,500-5,000 m snm) han generado condiciones ambientales extremas y aislamiento geográfico. Este aislamiento, sumado a la estabilidad climática, ha favorecido procesos de especiación y endemismo (MMA, 2018; Rivera et al., 2023). En contraste, el sur de Chile fue modelado por los avances y retrocesos glaciales del Pleistoceno, que fragmentan los hábitats a través de islotes, fiordos y canales de hielo (MMA, 2018). Estas condiciones también han favorecido la diferenciación filogenética y el endemismo local en especies que se han adaptado a ambientes fríos y húmedos (Cofré et al., 2007; Hernández-Mazariegos et al., 2023). La fragmentación y el aislamiento histórico de estos refugios glaciares pueden explicar los altos valores de ED observados en el extremo sur, conferidos por la historia evolutiva única de las especies que los habitan.
Asimismo, identificamos que las áreas con alto EDGE se concentran principalmente en el norte y centro-sur del país (fig. 2) y que geográficamente se superponen con aquellas que albergan la mayor cantidad de especies en estado de amenaza (casi amenazada, vulnerables, en peligro y en peligro crítico) y con las zonas más desprotegidas y perturbadas de Chile (Hernández-Mazariegos et al., 2025; Petit et al., 2018; UICN, 2024). Por ejemplo, el norte es el área con menor superficie bajo protección, mientras que el área central exhibe el mayor porcentaje de cambio de uso del suelo (Marquet et al., 2019; Miranda et al., 2017; Petit et al., 2018). Estos patrones refuerzan la idea de que el gradiente latitudinal de Chile no solo alberga una alta diversidad rica en historia evolutiva, sino además que las especies distribuidas en ciertas zonas están bajo más presión antropogénica, y posiblemente a un mayor riesgo futuro de extinción localizada.
Además, observamos que, al comparar la distribución latitudinal de los valores más altos de ED y EDGE con el mapa de áreas terrestres protegidas del país, se aprecia que algunas zonas coinciden parcialmente con regiones protegidas, en particular en el sur de Chile (fig. 2). Sin embargo, otras áreas con alto ED y EDGE, como el norte y parte del centro, están subrepresentadas en el sistema nacional de áreas protegidas (fig. 2). Esto sugiere vacíos importantes en la cobertura espacial de la conservación que podrían estar dejando fuera especies evolutivamente únicas y en riesgo. Esta discrepancia refuerza la necesidad de integrar métricas como ED y EDGE en la planificación de futuras áreas protegidas, considerando también la alta riqueza de especies en ciertas latitudes, lo que podría diluir la efectividad de protección para especies particulares. Proponemos que futuras estrategias de conservación territorial prioricen áreas donde se solapan altos valores de EDGE con baja cobertura de protección actual. Además, la cuantificación de la contribución individual de las especies a estas métricas puede orientar planes de manejo más específicos y eficaces.
Nuestros resultados subrayan que, aunque la mayoría de los roedores de Chile no figuran entre las especies distintamente evolutivas y con peligro global, algunos linajes presentan un alto índice EDGE que merece atención urgente (e.g., el género Chinchilla y Octodon pacificus). La distribución espacial de los valores ED y EDGE, particularmente en el norte y centro-sur del país, refuerza la necesidad de incorporar la dimensión geográfica en las estrategias de conservación. En un país megadiverso y con vacíos de información relevantes, integrar criterios evolutivos, ecológicos y espaciales puede fortalecer los esquemas regionales y prevenir pérdidas irreversibles de biodiversidad.
Este estudio es el primero en evaluar la distinción evolutiva y riesgo de amenaza de los roedores chilenos, por lo que no está exento de limitaciones. Entre ellas se encuentran las limitaciones metodológicas respecto a la filogenia generada, aunque la mayoría de los nodos presentan valores altos de probabilidad posterior (pp > 0.95), algunos nodos muestran soporte bajo (e.g., pp = 0.51). Al eliminar las politomías no cambió la señal filogenética. Esta incertidumbre filogenética podría afectar ciertas inferencias, como la estimación de la señal filogenética o la estructura de covarianza incorporada en los modelos PGLS. Idealmente, futuras evaluaciones podrían considerar la sensibilidad de los resultados utilizando un conjunto de árboles muestreados del posterior, para cuantificar cómo la incertidumbre topológica puede influir en los patrones detectados. También es importante considerar el posible sesgo en la estimación de la distribución de las especies, ya que los polígonos podrían estar sub o sobrerrepresentando la distribución latitudinal, además de considerar que algunas especies se distribuyen más allá de los límites de Chile (material suplementario: tabla S1). Asimismo, las discrepancias en las clasificaciones de estado de conservación de las especies de roedores (material suplementario: tabla S3). Dado que estas limitaciones pueden influir en los análisis de este estudio, es fundamental recalcularlos conforme se actualice la información genética disponible, se registren cambios en los rasgos de distribución y se reevalúe el estado de amenaza de las especies.
Agradecimientos
A la Agencia Nacional de Investigación y Desarrollo (ANID) [Núm. 21231791, Núm. 242240514 y Núm. 23240341] y a la Dirección General de Investigación de la Universidad Andrés Bello (DGI-UNAB) [Núm. DI-08-22/INI y Núm. DI-03-23/PASAN], por el apoyo financiero otorgado a WCHM durante los estudios de doctorado.
Referencias
Avaria-Llautureo, J., Hernández, C. E., Boric-Bargetto, D., Canales-Aguirre, C. B., Morales-Pallero, B. y Rodríguez-Serrano, E. (2012). Body size evolution in extant Oryzomyini rodents: Cope’s Rule or miniaturization? Plos One, 7, e34654. https://doi.org/10.1371/journal.pone.0034654
BCN. (2024). Mapas vectoriales. Biblioteca del Congreso Nacional de Chile. https://www.bcn.cl/siit/mapas_vectoria
les/index_html
Belmar-Lucero, S., Godoy, P., Ferrés, M., Vial, P. y Palma, R. E. (2009). Range expansion of Oligoryzomys longicaudatus (Rodentia, Sigmodontinae) in Patagonian Chile, and first record of Hantavirus in the region. Revista Chilena de Historia Natural, 82, 265–275. http://dx.doi.org/10.4067/S0716-078X2009000200008
Blackburn, T. M., Gaston, K. J. y Loder, N. (1999). Geographic gradients in body size: a clarification of Bergmann’s rule. Diversity and Distributions, 5, 165–174. https://doi.org/10.1046/j.1472-4642.1999.00046.x
Cheng, P., Yu, D., Liu, S., Tang, Q. y Liu, H. (2014). Molecular phylogeny and conservation priorities of the subfamily Acheilognathinae (Teleostei: Cyprinidae). Zoological Science, 31, 300. https://doi.org/10.2108/zs130069
Chichorro, F., Urbano, F., Teixeira, D., Väre, H., Pinto, T., Brummitt, N. et al. (2022). Trait-based prediction of extinction risk across terrestrial taxa. Biological Conservation, 274, 109738. https://doi.org/10.1016/j.biocon.2022.109738
Cofré, H. L. y Marquet, P. A. (1999). Conservation status, rarity, and geographic priorities for conservation of Chilean mammals: An assessment. Biological Conservation, 88, 53–68. https://doi.org/10.1016/S0006-3207(98)00090-1
Cofré, H. L., Samaniego, H. y Marquet, P. A. (2007). Patterns of small mammal species richness in mediterranean and temperate Chile. The Quintessential Naturalist Honoring the Life and Legacy of Oliver P. Pearson. University of California Press. https://doi.org/10.1525/california/978052
0098596.001.0001
Cooke, R. S. C., Eigenbrod, F. y Bates, A. E. (2020). Ecological distinctiveness of birds and mammals at the global scale. Global Ecology and Conservation, 22, e00970. https://doi.org/10.1016/j.gecco.2020.e00970
Cortés-Díaz, D., Buitrago-Torres, D. L., Restrepo-Cardona, J. S., Estellés-Domingo, I. y López-López, P. (2023). Bridging evolutionary history and conservation of new world vultures. Animals, 13, 3175. https://doi.org/10.3390/ani13203175
D’Elía, G., Canto, J., Ossa, G., Verde-Arregoitia, L. D., Bostelmann, E., Iriarte, A. et al. (2020). Lista actualizada de los mamíferos vivientes de Chile. Boletín Museo Nacional de Historia Natural, 69, 67–98. https://doi.org/10.54830/bmnhn.v69.n2.2020.6
Duclos, M., Silva-Pérez, C. P., Silva-Aránguiz, E. M. y Jaksic, F. M. (2024). Cráneos & Pelos. Guía de identificación de mamíferos de Chile. Santiago, Chile: Centro de Ecología Aplicada y Sustentabilidad, CAPES (ANID PIA/BASAL FB0002). https://capes.cl/craneosypelos/
Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5, 113. https://doi.org/10.1186/1471-2105-5-113
EDGE. (2024). The EDGE of Existence Programme. https://www.edgeofexistence.org/
Faith, D. P. (2019). EDGE of existence and phylogenetic diversity. Animal Conservation, 22, 537–538. https://doi.org/10.1111/acv.12552
Fourcade, Y. y Alhajeri, B. H. (2023). Environmental correlates of body size influence range size and extinction risk: a global study in rodents. Global Ecology and Biogeography, 32, 206–217. https://doi.org/10.1111/geb.13622
Fritz, S. A. y Purvis, A. (2010). Selectivity in mammalian extinction risk and threat types: a new measure of phylogenetic signal strength in binary traits. Conservation Biology, 24, 1042–1051. https://doi.org/10.1111/j.1523-1739.2010.01455.x
Gaston, K. J. (2003). The structure and dynamics of geographic ranges. Oxford. Oxford University Press. https://archive.org/details/structuredynamic0000gast/page/n9/mode/2up
Gaulke, S., Martelli, E., Johnson, L., Letelier, C. G., Dawson, N. y Nelson, C. R. (2019). Threatened and endangered mammals of Chile: Does research align with conservation information needs? Conservation Science and Practice, 1, e99. https://doi.org/10.1111/csp2.99
GenBank. (2023). National Library of Medicine. National Center for Biotechnology Information. https://www.ncbi.nlm.nih.gov/genbank/
Griffith, P., Lang, J. W., Turvey, S. T. y Gumbs, R. (2023). Using functional traits to identify conservation priorities
for the world’s crocodylians. Functional Ecology, 37, 112–124. https://doi.org/10.1111/1365-2435.14140
Gumbs, R., Gray, C. L., Böhm, M., Burfield, I. J., Couchman, O. R., Faith, D. P. et al. (2023). The EDGE2 protocol: advancing the prioritisation of Evolutionarily Distinct and Globally Endangered species for practical conservation action. Plos Biology, 21, e3001991. https://doi.org/10.1371/journal.pbio.3001991
Gumbs, R., Gray, C. L., Wearn, O. R. y Owen, N. R. (2018). Tetrapods on the EDGE: overcoming data limitations to identify phylogenetic conservation priorities. Plos One, 13, e0194680. https://doi.org/10.1371/journal.pone.0194680
Gumbs, R., Scott, O., Bates, R., Böhm, M., Forest, F., Gray, C. L. et al. (2024). Global conservation status of the jawed vertebrate Tree of Life. Nature Communications, 15, 1101. https://doi.org/10.1038/s41467-024-45119-z
Hernández-Mazariegos, Palma, R. E., y Escobar, L. E. (2023). Rodents of Chile: a brief appraisal of their conservation status and ecological significance. Zookeys, 1254, 107–129. https://doi.org/10.3897/zookeys.1254.148057
Hidasi-Neto, J., Loyola, R. y Cianciaruso, M. V. (2015). Global and local evolutionary and ecological distinctiveness of terrestrial mammals: identifying priorities across scales. Diversity and Distributions, 21, 548–559. https://doi.org/10.1111/ddi.12320
Huang, S., Davies, T. J. y Gittleman, J. L. (2012). How global extinctions impact regional biodiversity in mammals. Biology Letters, 8, 222–225. https://doi.org/10.1098/rsbl.
2011.0752
Iriarte, A. (2007). Mamíferos de Chile. Santiago, Chile: Ediciones.
Iriarte, A. (2021). Guía de los mamíferos de Chile, 2da edición. Santiago, Chile: Flora y Fauna.
Isaac, N. J. B., Turvey, S. T., Collen, B., Waterman, C. y Baillie, J. E. M. (2007). Mammals on the EDGE: conservation priorities based on threat and phylogeny. Plos One, 2, e296. https://doi.org/10.1371/journal.pone.0000296
Jetz, W., McPherson, J. M. y Guralnick, R. (2012). Integrating biodiversity distribution knowledge: toward a global map of life. Trends in Ecology & Evolution,, 27, 151–159. https://doi.org/10.1016/j.tree.2011.09.007
Johnson, T. F., Beckerman, A. P., Childs, D. Z., Webb, T. J., Evans, K. L., Griffiths, C. A. et al. (2024). Revealing uncertainty in the status of biodiversity change. Nature, 628, 788–794. https://doi.org/10.1038/s41586-024-07236-z
Keane, A., Brooke, M. L. y Mcgowan, P. J. K. (2005). Correlates of extinction risk and hunting pressure in gamebirds (Galliformes). Biological Conservation, 126, 216–233. https://doi.org/10.1016/j.biocon.2005.05.011
Kembel, S. W., Ackerly, D. D., Blomberg, S. P., Cornwell, W. K., Cowan, P. D., Helmus, M. R. et al. (2020). picante: Integrating Phylogenies and Ecology. R Package Version. https://CRAN.R-project.org/package=picante
Kennerley, R. J., Lacher, T. E., Hudson, M. A., Long, B., McCay, S. D., Roach, N. S. et al. (2021). Global patterns of extinction risk and conservation needs for Rodentia and Eulipotyphla. Diversity and Distributions, 27, 1792–1806. https://doi.org/10.1111/ddi.13368
Kumar, S., Stecher, G., Li, M., Knyaz, C. y Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution,
35, 1547–1549. https://doi.org/10.1093/molbev/msy096
Llobet, T., Velikov, I., Sogorb, M., Peacock, F., Jutglar, F., Mascarell, A. et al. (2023). All the mammals of the World. Barcelona, Spain: Lynx Nature Books. https://lynx
naturebooks.com/product/all-the-mammals-of-the-world/
Lomolino, M. V., Riddle, B. R. y Whittaker, R. J. (2017). Biogeography: biological diversity across space and time. 5a Ed. Oxford: Sinauer Associates.
MacPhee, R. D. E. y Flemming, C. (1999). Requiem Aeternam: the last five hundred years of mammalian species extinctions. En R.D.E. MacPhee (Ed.), Extinctions in near time: causes, contexts and consequenses (pp. 333–371). Kluwer Academic/ Plenum Publisher.
Maestri, R. (2020). A macroecological perspective on neotropical rodents. Mastozoología Neotropical, 27, 24–34. https://doi.org/10.31687/saremMN_SI.20.27.1.04
Maestri, R., Luza, A. L., De Barros, L. D., Hartz, S. M., Ferrari, A., De Freitas, T. R. O. et al. (2016). Geographical variation of body size in sigmodontine rodents depends on both environment and phylogenetic composition of communities. Journal of Biogeography, 43, 1192–1202. https://doi.org/10.1111/jbi.12718
Marquet, P. A., Fernández, M., Pliscoff, P., Smith-Ramírez, C., Arellano, E., Armesto, J. et al. (2019). Áreas protegidas y restauración en el contexto del cambio climático en Chile. Informe de la mesa de Biodiversidad. Santiago: Ministerio de Ciencia, Tecnología, Conocimiento e Innovación.
MDD. (2024). Mammal diversity database [Dataset]. Zenodo. https://www.mammaldiversity.org/
Meiri, S., Bauer, A. M., Allison, A., Castro-Herrera, F., Chirio, L., Colli, G. et al. (2018). Extinct, obscure or imaginary: the lizard species with the smallest ranges. Diversity and Distributions, 24, 262–273. https://doi.org/10.1111/ddi.12678
Miranda, A., Altamirano, A., Cayuela, L., Lara, A. y González, M. (2017). Native forest loss in the Chilean biodiversity
hotspot: revealing the evidence. Regional Environmental Change, 17, 285–297. https://doi.org/10.1007/s10113-016-1010-7
MMA (Ministerio del Medio Ambiente). (2018). Biodiversidad de Chile. Patrimonio y Desafíos. Tercera Edición. Tomo I. Santiago, Chile: Ministerio del Medio Ambiente.
MMA (Ministerio del Medio Ambiente). (2024).Lista de especies clasificadas desde el 1 al 19 proceso de clasificación RCE. Santiago, Chile: Ministerio del Medio Ambiente. https://clasificacionespecies.mma.gob.cl/
Mooers, A. Ø., Faith, D. P. y Maddison, W. P. (2008). Converting endangered species categories to probabilities of extinction for phylogenetic conservation prioritization. Plos One, 3, e3700. https://doi.org/10.1371/journal.pone.0003700
Owen, C. L., Bracken-Grissom, H., Stern, D. y Crandall, K. A. (2015). A synthetic phylogeny of freshwater crayfish: insights for conservation. Philosophical Transactions of the Royal Society B: Biological Sciences, 370, 20140009. https://doi.org/10.1098/rstb.2014.0009
Pagel, M. (1999). Inferring the historical patterns of biological evolution. Nature, 401, 877–884. https://doi.org/10.1038/44766
Pagel, M. (2002). Modelling the evolution of continuously varying characters on phylogenetic trees: the case of hominid cranial capacity. En N. MacLeod y P. Forey (Ed.). Morphology, shape and phylogenetics (pp. 269–286): Taylor y Francis.
Palma, R. E. y Rodríguez-Serrano, E. (2018). Systematics of Oligoryzomys (Rodentia, Cricetidae, Sigmodontinae) from southern Chilean Patagonia, with the description of a new species. Journal of Zoological Systematics and Evolutionary Research, 56, 280–299. https://doi.org/10.1111/jzs.12199
Paradis, E. y Schliep, K. (2019). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics, 35, 526–528. https://doi.org/10.1093/bioinformatics/bty633
Patton, J. L., Pardiñas, U. F. J. y D’Elía, G. (2015). Mammals of South America, Volume 2. Rodents. Chicago: University of Chicago Press. https://doi.org/10.7208/chicago/9780
226169606.001.0001
Petit, I. J., Campoy, A. N., Hevia, M.-J., Gaymer, C. F. y Squeo, F. A. (2018). Protected areas in Chile: Are we managing them? Revista Chilena de Historia Natural, 91, 1. https://doi.org/10.1186/s40693-018-0071-z
Pie, M. R. y Meyer, A. L. S. (2017). The Evolution of range sizes in mammals and squamates: heritability and differential evolutionary rates for low- and high-latitude limits. Evolutionary Biology, 44, 347–355. https://doi.org/10.1007/s11692-017-9412-0
Poux, C., Chevret, P., Huchon, D., De Jong, W. W. y Douzery, E. J. P. (2006). Arrival and diversification of caviomorph rodents and platyrrhine primates in South America. Systematic Biology, 55, 228–244. https://doi.org/10.1080/10635150500481390
Quiroga-Carmona, M., González, A., Valladares, P., Hurtado, N. y D’Elía, G. (2023). Increasing the known specific richness of living mammals in Chile. Therya, 14, 215–222. https://doi.org/10.12933/therya-23-2217
Quiroga-Carmona, M., Storz, J. F. y D’Elía, G. (2023). Elevational range extension of the Puna Mouse, Punomys (Cricetidae), with the first record of the genus from Chile. Journal of Mammalogy, 104, 1144–1151. https://doi.org/10.1093/jmammal/gyad064
R Core Team. (2024). r: a language and environment for statistical. R Foundation for Statistical Computing. https://www.R-project.org/
Rambaut, A. (2009). Molecular evolution, phylogenetics and epidemiology. https://tree.bio.ed.ac.uk/software/tracer/
Redding, D. W., DeWolff, C. V. y Mooers, A. Ø. (2010). Evolutionary distinctiveness, threat status, and ecological oddity in primates. Conservation Biology, 24, 1052–1058. https://doi.org/10.1111/j.1523-1739.2010.01532.x
Revell, L. J. (2012). phytools: an R package for phylogenetic comparative biology (and other things). Methods in Ecology and Evolution, 3, 217–223. https://doi.org/10.1111
/j.2041-210X.2011.00169.x
Ripple, W. J., Wolf, C., Newsome, T. M., Hoffmann, M., Wirsing, A. J. y McCauley, D. J. (2017). Extinction risk is most
acute for the world’s largest and smallest vertebrates. Proceedings of the National Academy of Sciences, 114, 10678–10683. https://doi.org/10.1073/pnas.1702078114
Rivera, R., Aldunate, C., Jerez, V., Berenguer, J., Lisón, F., Chamorro, S. et al. (2023). Fauna, un recorrido por el endemismo de Chile. Santiago, Chile: Banco Santander y Museo Chileno de Arte Precolombino.
Roach, N. (2016). Octodon pacificus. The UICN Red List of Threatened Species 2016: e.T15090A78321512. http://dx.doi.org/10.2305/UICN.UK.2016-2.RLTS.T15090A78321512.en
Roach, N. y Kennerley, R. (2016a). Chinchilla chinchilla. The UICN Red List of Threatened Species 2016: e.T4651A22191157. http://dx.doi.org/10.2305/UICN.UK.20
162.RLTS.T4651A22191157.en
Roach, N. y Kennerley, R. (2016b). Chinchilla lanigera, Long-tailed Chinchilla. The UICN
Red List of Threatened Species 2016: e.T4652A117975205.
http://dx.doi.org/10.2305/UICN.UK.20162.RLTS.T4652A22190974.en
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhna, et al. (2012). MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61, 539–542.
https://doi.org/10.1093/sysbio/sys029
Rosauer, D. F., Laffan, S. W., Crisp, M. D., Donnellan, S. C. y Cook, L. G. (2009). Phylogenetic endemism: a new approach for identifying geographical concentrations of evolutionary history. Molecular Ecology, 18, 4061–4072. https://doi.org/10.1111/j.1365-294X.2009.04311.x
Santini, L., Antão, L. H., Jung, M., Benítez-López, A., Rapacciuolo, G., Di Marco, M. et al. (2021). The interface between macroecology and conservation: existing links
and untapped opportunities. Frontiers of Biogeography, 13.4, e53025. https://doi.org/10.21425/F5FBG53025
Smith, F. A., Brown, J. H., Haskell, J. P., Lyons, S. K., Alroy, J., Charnov, E. L. et al. (2004). Similarity of mammalian body size across the taxonomic hierarchy and across space and time. The American Naturalist, 163, 672–691. https://doi.org/10.1086/382898
Smyčka, J., Toszogyova, A. y Storch, D. (2023). The relationship between geographic range size and rates of species diversification. Nature Communications, 14, 5559. https://doi.org/10.1038/s41467-023-41225-6
Spotorno, A. E., Zuletar R, C., Walker, L. I., Manriquez, G., Valladares F, P. y Marin, J. C. (2013). A small, new gerbil-mouse Eligmodontia (Rodentia: Cricetidae) from dunes at the coasts and deserts of north-central Chile: molecular, chromosomic, and morphological analyses. Zootaxa, 3683, 377-394. https://doi.org/10.11646/zootaxa.3683.4.3
Stein, R. W., Mull, C. G., Kuhn, T. S., Aschliman, N. C., Davidson, L. N. K., Joy, J. B. et al. (2018). Global priorities for conserving the evolutionary history of sharks, rays and chimaeras. Nature Ecology & Evolution, 2, 288–298. https://doi.org/10.1038/s41559-017-0448-4
Stevens, G. C. (1992). The elevational gradient in altitudinal range: an extension of rapoport’s latitudinal rule to altitude. The American Naturalist, 140, 893–911. https://doi.org/10.1086/285447
Teta, P., D’Elía, G., Lanzone, C., Ojeda, A., Novillo, A. y Ojeda, R. A. (2021). A reappraisal of the species richness of Euneo-
mys Coues 1874 (Rodentia, Cricetidae), with emendations of the type localities of Reithrodon fossor Thomas 1899 and Euneomys mordax Thomas1912. Mammalia, 85, 379–388. https://doi.org/10.1515/mammalia-2020-0157
Teta, P., Formoso, A., Tammone, M., De Tommaso, D. C., Fernández, F. J., Torres, J. et al. (2014). Micromamíferos, cambio climático e impacto antrópico: ¿Cuánto han cambiado las comunidades del sur de América del Sur en los últimos 500 años? Therya, 5, 7–38. https://doi.org/10.12933/therya-14-183
Teta, P. y Pardiñas, U. F. J. (2014). Variación morfológica cualitativa y cuantitativa en Abrothrix longipilis (Cricetidae, Sigmodontinae). Mastozoología Neotropical, 21, 291–309.
Tucker, C. M., Aze, T., Cadotte, M. W., Cantalapiedra, J. L., Chisholm, C., Díaz, S. et al. (2019). Assessing the utility of conserving evolutionary history. Biological Reviews, 94, 1740–1760. https://doi.org/10.1111/brv.12526
UICN (Unión Internacional para la Conservación de la Naturaleza). (2024). The Red List of threatened species. International Union for Conservation Nature. https://www.UICNredlist.org
UICN Standards and Petitions Committee. (2024). Guidelines for using the UICN Red List categories and criteria. https://www.UICNredlist.org/documents/RedListGuidelines.pdf
Upham, N. S. y Patterson, B. D. (2015). Evolution of the caviomorph rodents: a complete phylogeny and timetree of living genera. Mammalogical Research, 1, 63–120.
Webb, T. J. y Gaston, K. J. (2003). On the heritability of geographic range sizes. The American Naturalist, 161, 553–566. https://doi.org/10.1086/368296
Webb, T. J. y Gaston, K. J. (2005). heritability of geographic range sizes revisited: a reply to hunt. The American Naturalist, 166, 136–143. https://doi.org/10.1086/430726
WWF. (2020). Living planet report 2020-bending the curve of biodiversity loss (R.E.A. Almond, M. Grooten y T. Petersen (Eds). WWF, Gland, Switzerland.