Desde hace 40 años, el Simposio Nacional de Parasitología Forestal consolidó un espacio para la discusión científica y técnica en torno a los organismos que se relacionan con la salud de los ecosistemas forestales de México. En este periodo, el simposio promovió el intercambio de conocimientos entre diferentes sectores fortaleciendo el entendimiento de las interacciones entre insectos, patógenos y plantas hospederas bajo un enfoque centrado en sanidad forestal y, más recientemente, en el manejo integrado de plagas.
Los problemas contemporáneos en la gestión de los recursos forestales —aumento en la frecuencia e intensidad de brotes de insectos, expansión de especies invasoras, eventos extremos asociados al cambio climático y transformaciones socioecológicas en los territorios forestales— demandan una revisión profunda de los marcos conceptuales tradicionales. En este contexto, durante el XXI Simposio Nacional de Parasitología Forestal, celebrado en el Instituto de Biología de la Universidad Nacional Autónoma de México en la Ciudad de México, se tomó una decisión trascendental: la evolución del evento hacia el Primer Simposio de Salud Forestal, marcando un hito en la historia del foro. Este cambio representa el reconocimiento de que la integridad de los ecosistemas no puede comprenderse únicamente desde la perspectiva del daño o de sus agentes causales, sino desde una visión amplia, integral y sistémica que considere procesos ecológicos, socioambientales y culturales.
El concepto de salud forestal trasciende la noción de sanidad como control o mitigación de plagas; incorpora la resiliencia, la gobernanza comunitaria, la restauración ecológica y la comprensión de los factores climáticos, biológicos y humanos que modelan la estabilidad de los bosques. Así, esta transición abre un camino renovado para el análisis, la prevención y atención de las afectaciones forestales, alineado con los desafíos presentes y con las prácticas emergentes de manejo sustentable en México y el mundo.
Siguiendo la tradición iniciada en 2021 con la publicación del primer suplemento en la Revista Mexicana de Biodiversidad, este segundo volumen reúne investigaciones que reflejan la complejidad y diversidad de problemáticas abordadas en sistemas forestales, bajo el nuevo paradigma. Los trabajos aquí integrados abarcan enfoques taxonómicos, ecológicos, biogeográficos y socioambientales, todos vinculados por el interés común de comprender y promover la salud de los ecosistemas.
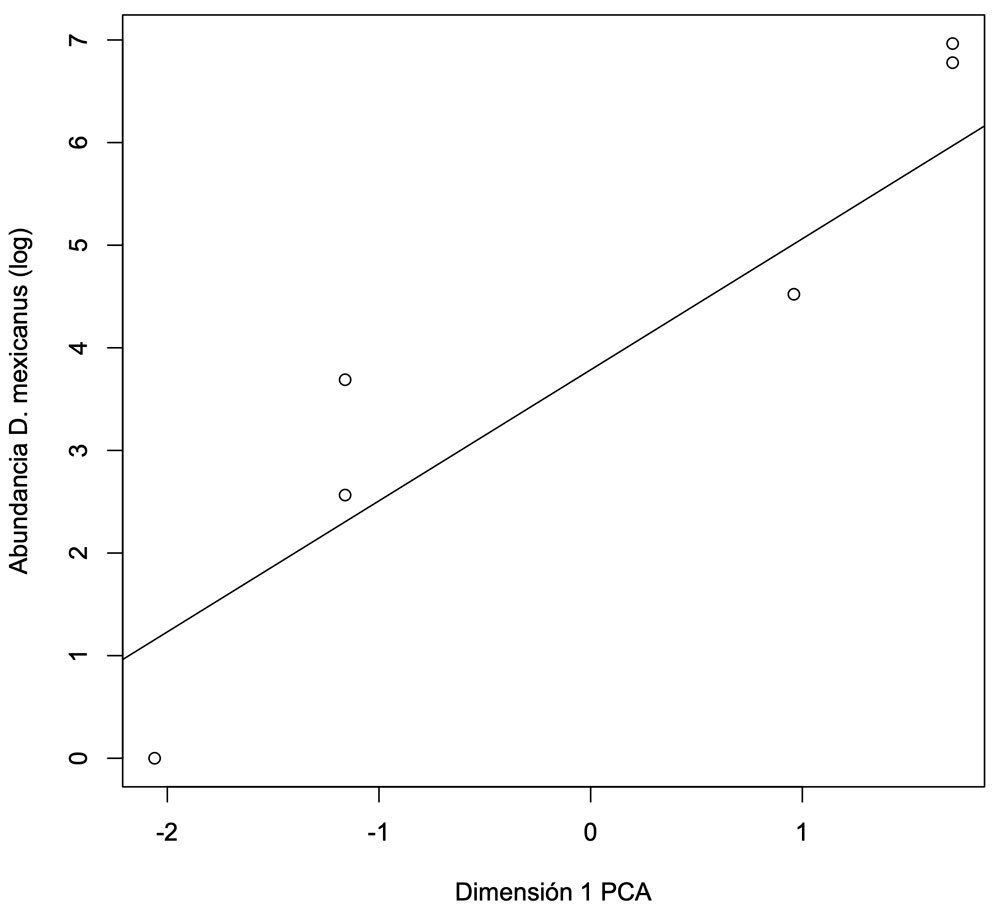
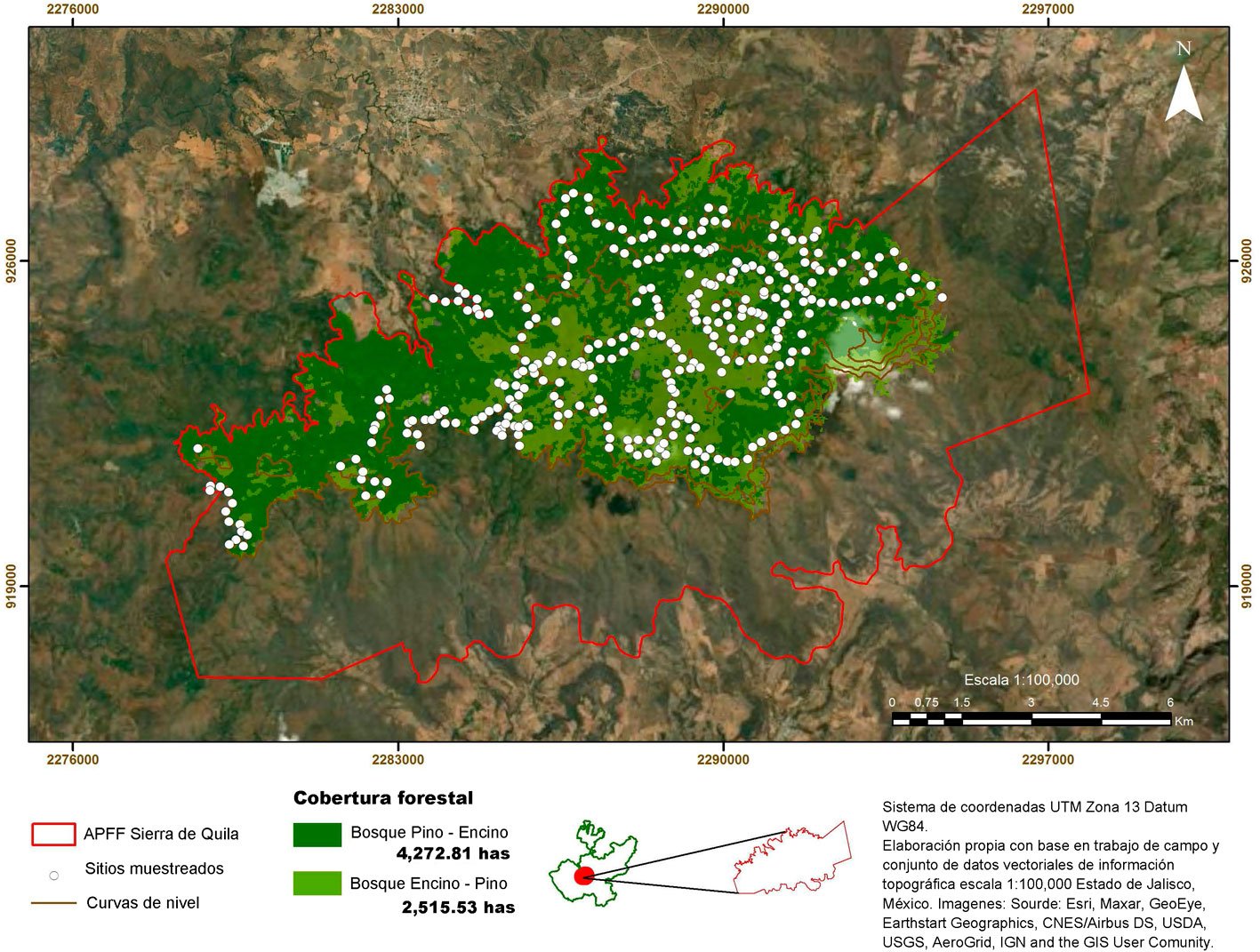
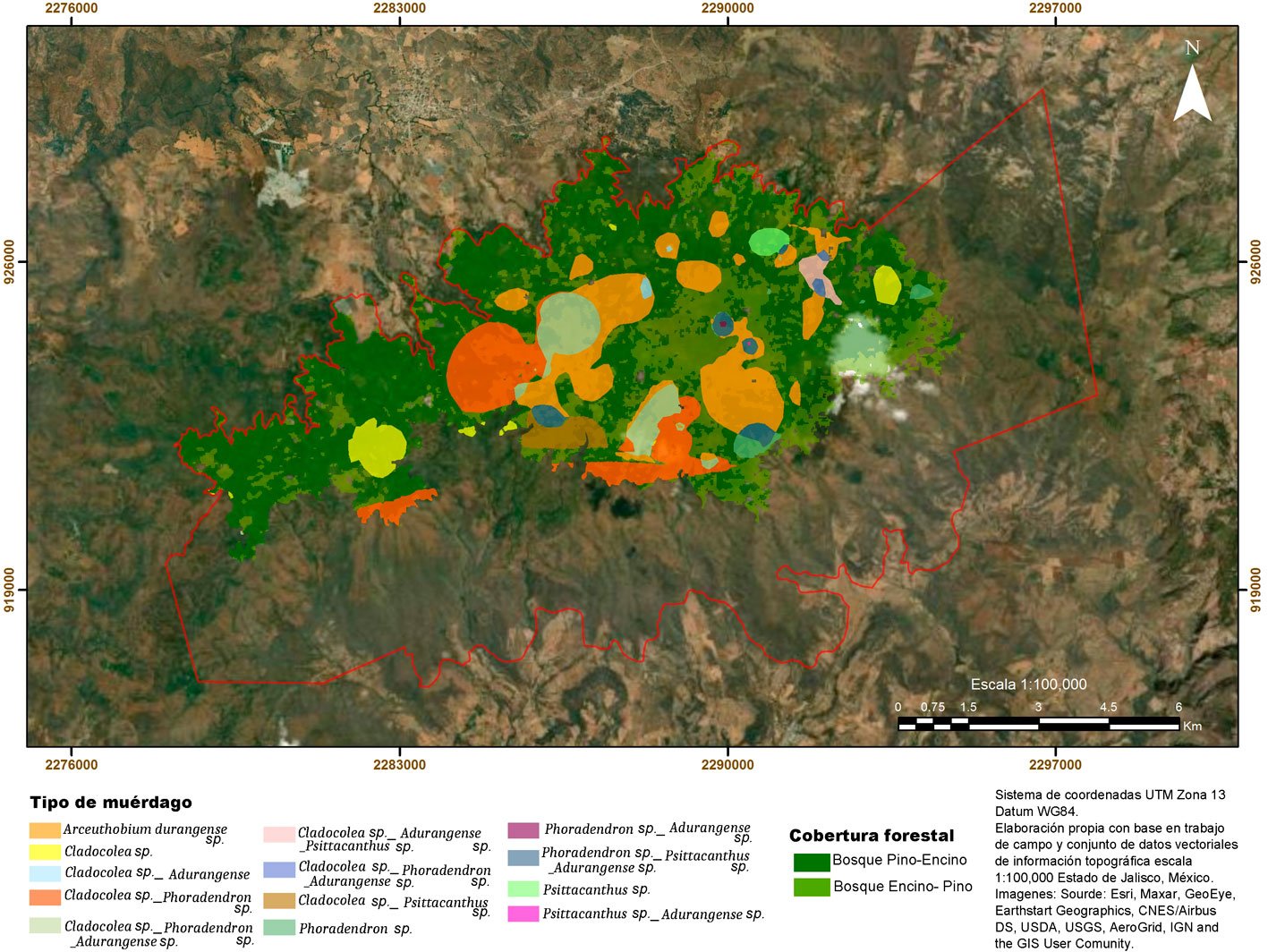
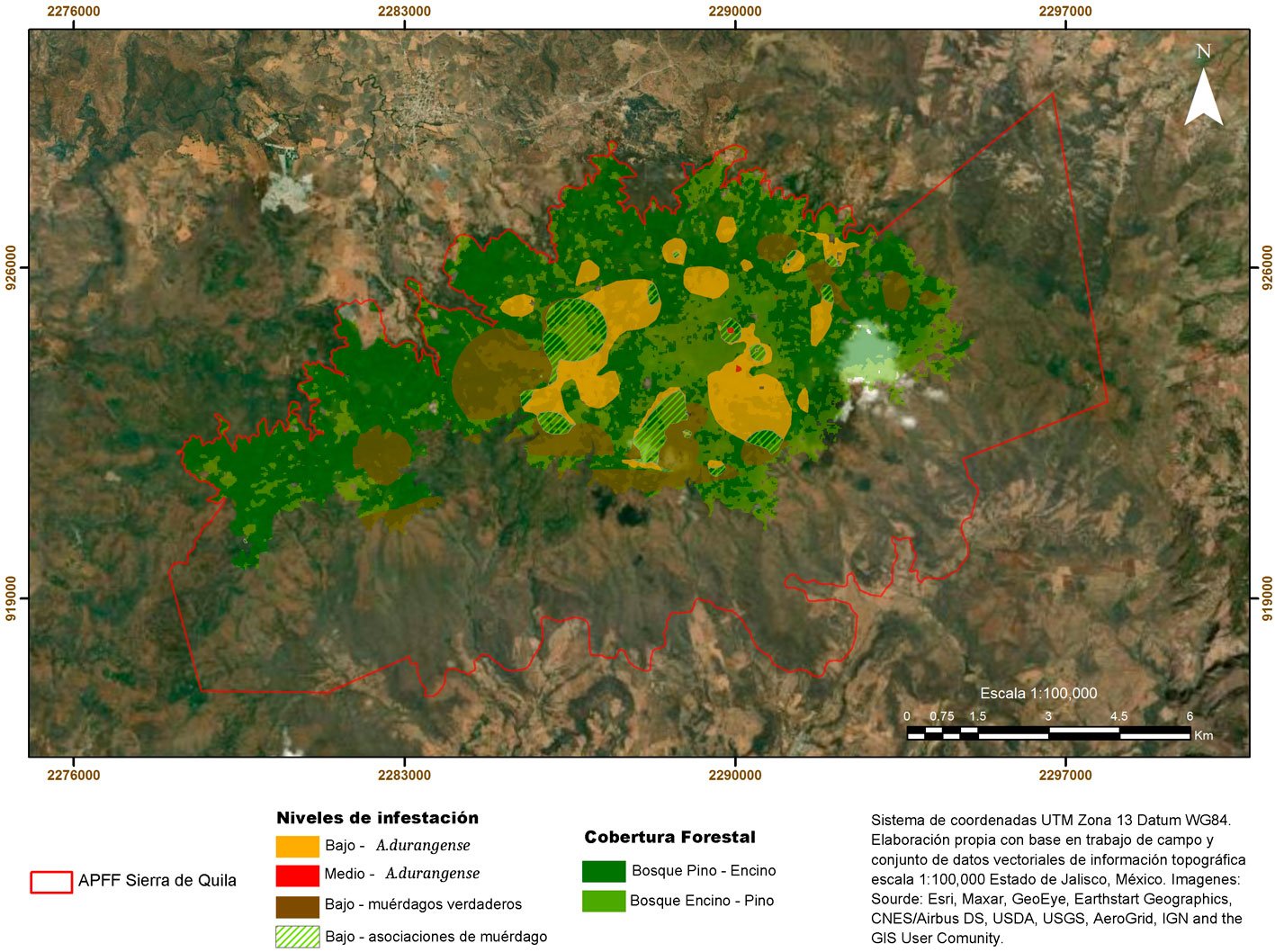
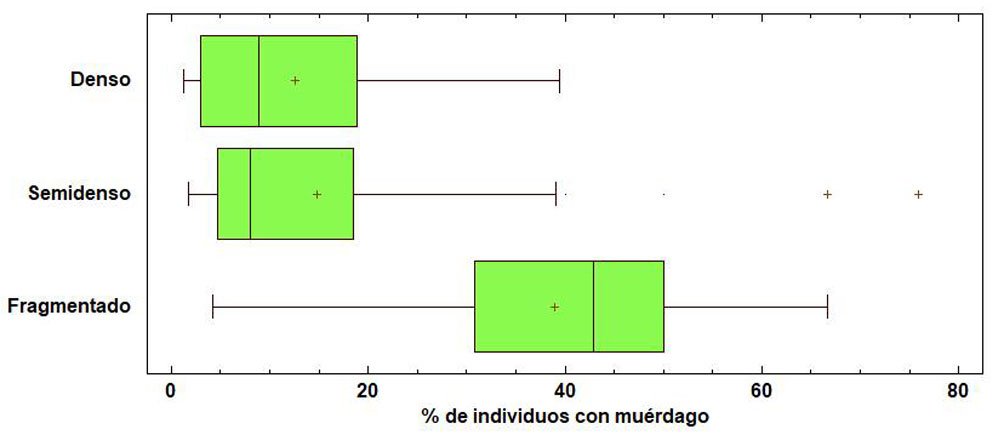
El fascículo inicia con un análisis detallado de la distribución espacial de plantas parásitas en la sierra de Quila, Jalisco. Se documenta que cerca de 30% del área boscosa presenta infestación por diversas especies de muérdagos, aunque aún con niveles bajos, lo que brinda una ventana de oportunidad para implementar medidas de manejo que prevengan afectaciones de mayor magnitud. La segunda contribución es un estudio realizado en Santa Catarina Lachatao, Oaxaca, donde se evalúan la diversidad de escarabajos del género Dendroctonus y el estado fitosanitario del bosque bajo un enfoque participativo. Los resultados muestran que el manejo forestal comunitario puede mantener bosques resilientes frente a los brotes favorecidos por el cambio climático, subrayando la relevancia de los modelos de gobernanza local. El tercer trabajo examina los efectos simultáneos de Dendroctonus adjunctus y del muérdago Arceuthobium vaginatum sobre la estructura del bosque de Pinus hartwegii en el Nevado de Colima. Los resultados muestran la alteración del dosel y la disminución de árboles sanos, con consecuencias en la regeneración natural, especialmente en la ladera occidental del volcán. El siguiente estudio reporta, por primera vez a nivel mundial, la asociación entre el áfido Pseudoregma panicola y su hospedante Arundo donax en la Ciudad de México. Mediante análisis morfológicos y moleculares se documenta la presencia histórica del insecto en el país, así como un proceso aún incipiente de integración de sus enemigos naturales, lo que resalta su potencial como un agente sujeto a manejo en las áreas urbanas. También se presenta una contribución sobre la diversidad de cicadélidos de la tribu Athysanini en bosques de pino y encino de México. Con más de 380 registros georreferenciados y la descripción de una especie nueva del género Eutettix, este trabajo amplía el conocimiento taxonómico y biogeográfico de un grupo clave en la dinámica de herbivoría en ecosistemas templados. Finalmente, la última obra analiza la biología, distribución e idoneidad de hábitat del gorgojo barbudo Rhinostomus barbirostris, un fitófago de alta relevancia para las palmas del Neotrópico. El estudio modela su distribución potencial en México y documenta un brote reciente en la Reserva de la Biosfera Tehuacán-Cuicatlán, evidenciando la necesidad de monitoreo continuo ante su creciente impacto ecológico y económico.
En conjunto, los trabajos de este suplemento reflejan la diversidad de agentes bióticos, ambientes, escalas y enfoques que constituyen hoy la agenda de la salud forestal en México. Esta publicación busca contribuir a la consolidación del nuevo paradigma, fomentando la integración del conocimiento científico, la experiencia comunitaria y las estrategias preventivas orientadas a conservar la funcionalidad y estabilidad de los ecosistemas forestales del país.
Editores responsables del suplemento: Francisco Armendáriz-Toledano (farmendariztoledano@ib.unam.mx), Colección Nacional de Insectos, Instituto de Biología-UNAM, Ek del Val de Gortari (Instituto de Investigaciones en Ecosistemas y Sustentabilidad, UNAM) y Adilson Pinedo Escatel (IB-UNAM)
Conociendo lo que se conoce: biología, distribución e idoneidad de hábitat del escarabajo barbudo Rhinostomus barbirostris (Curculionidae: Dryophthorinae) en México
Osiris Valerio-Mendoza a, b, Leticia Soriano-Flores c, Idalia Fabiola Lazaro-Lopez f, Miguel Murguía-Romero d, Enrique Ortiz e, Gerardo Cuellar-Rodrigueza, Francisco Armendáriz-Toledano b, *
a Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, Campus Linares, Km 145, Nacional 85, 67700 Linares, Nuevo León, Mexico
b Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Insectos, Tercer Circuito s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
c Programa de las Naciones Unidas para el Desarrollo, Montes Urales No. 440, Lomas de Chapultepec, 11000 Ciudad de México, Mexico
d Universidad Nacional Autónoma de México, Instituto de Biología, Unidad de Informática para la Biodiversidad, Tercer Circuito s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
e Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Botánica, Tercer Circuito s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
f Alianza de Comunidades Chocho-Mixtecas del Estado de Oaxaca, Cedros s/n, Col. Emiliano Zapata, Asunción Nochixtlán Oaxaca, Mexico
*Autor para correspondencia: farmendariztoledano@ib.unam.mx (F. Armendáriz-Toledano)
Recibido: 27 marzo 2025; aceptado: 24 septiembre 2025
Abstract
The bearded weevil, Rhinostomus barbirostris, is a significant damaging organism of palms in the Neotropics, heavily impacting ecologically and economically important species. This study details its biology, distribution, and habitat suitability in Mexico, focusing on a recent outbreak in the Tehuacán-Cuicatlán Biosphere Reserve (TCBR), where it affected the sweet palm, Brahea dulcis. Field surveys confirmed significant damage and palm death in the TCBR due to weevil larvae and adults. A database of historical and current records was used to model its potential distribution using bioclimatic variables. Results show wide habitat suitability for R. barbirostris across Mexico, especially in humid montane and tropical forests, although recent records center on the TCBR. This research provided crucial data on the weevil’s biology and distribution, emphasizing the need for monitoring and management to protect native palm populations and prevent economic and ecological harm.
El gorgojo barbudo, Rhinostomus barbirostris, es un organismo dañino para las palmas en el Neotrópico, con un fuerte impacto en especies de importancia ecológica y económica. Este estudio detalla su biología, distribución e idoneidad de hábitat en México, centrándose en un brote reciente en la Reserva de la Biosfera Tehuacán-Cuicatlán (RBTC), donde afectó a la palma dulce, Brahea dulcis. Los estudios de campo confirmaron daños significativos y la muerte de palmas en la RBTC debido a larvas y adultos del gorgojo. Se utilizó una base de datos de registros históricos y actuales para modelar su distribución potencial mediante variables bioclimáticas. Los resultados muestran una amplia idoneidad de hábitat para R. barbirostris en todo México, especialmente en bosques húmedos de montaña y tropicales, aunque los registros recientes se centran en la RBTC. Esta investigación proporcionó datos cruciales sobre la biología y distribución del gorgojo, enfatizando la necesidad de monitoreo y manejo para proteger las poblaciones nativas de palmas y prevenir daños económicos y ecológicos.
Dryophthorinae, identified by Schoenherr (1825), is recognized as one of the most distinctive and well-defined subfamilies of Curculionidae, notable for its unique synapomorphies (Kuschel, 1995; Luna-Cozar et al., 2024; Marvaldi & Morrone, 2000). These weevils, among the largest of the family, present a great diversity, with around 1,200 species grouped in more than 150 genera and 5 main tribes: Dryophthorinini, Stromboscerini, Cryptodermatini, Orthognathini and Rhynchophorini (Oberprieler et al., 2007).
This group of insects stands out for its specialization in feeding monocotyledons, and to a lesser extent, on dicotyledons. Their saprophagous habits, mainly focused on leaf litter and wood (Anderson & Marvaldi, 2014; Gardner, 1934, 1938; Grebennikov, 2018a-c; Vaurie, 1970a, b, 1971), allow them to adapt to a wide variety of ecosystems, from tropical forests to deserts and grasslands, on all continents. Their evolutionary success, evidenced by an extensive fossil record (McKenna et al., 2015; Chamorro et al., 2021) dating back between 65 and 115 million years, suggests great adaptability and a close relationship with the evolution of plants (O’Meara, 2001; McKenna et al., 2009).
Many species of the subfamily represent a serious threat to agriculture worldwide, because they feed on a wide range of crops, both tropical and temperate, including palms, pineapple, bamboo, banana, sugarcane, maize, wheat and rice (Van Huis et al., 2013). Among the most prominent and damaging members are the rice weevils of the genus Sitophilus, Schoenherr, and the palm weevils of the genera Rhynchophorus, Herbst, and Rhinostomus Rafinesque. These taxa cause significant economic losses and put food security at risk in numerous regions of the world (Rugman-Jones et al., 2013; Van Huis et al., 2013).
Rhinostomus Rafinesque, 1815 has 7 species: R. barbirostris, R. oblitus, R. quadrisignatus, R. scrutator, R. thompsoni from the Neotropics; R. niger from the Afrotropical region; and the Oriental R. meldolae (Vaurie, 1970a). The bearded weevil R. barbirostris is considered the third largest weevil on the planet, being surpassed in the same subfamily by Protocerius collosus from Australasia (60 mm) and Rhynchodynamis filirostris (Vaurie, 1970a; Zaragoza-Caballero et al., 2015). Adult individuals are characterized by having an average length of 2-4 cm. Males present an interesting appearance with a “brush of hairs” or setae on the head, that represent an advantage during courtship when they are rubbed on the dorsal surface of the female (Eberhard, 1983). Females lack a brush and prefer to oviposit on the intact bark of weakened or stressed palms (Cocos nucifera), both cultivated and wild (Bouchard, 2014; FAO, 2013; Zaragoza-Caballero et al., 2015). The bearded weevil is an important pest of palm trees across its distribution range, feeding on Attalea funifera Burret, A. maripa (Aubl.) Mart., A. pindobassu Bondar, A. piassabossu Bondar, Bactris gasipaes Kunth, Brahea dulcis (Kunth) Mart., Cocos botryophora Mart., C. coronata Mart, C. nucifera L., Elaeis guineensis Jacq., Euterpe oleracea Mart., Mauritia flexuosa L. F., Oenocarpa bacaba Mart., and Syagrus romanzoffiana (Cham.) Glassman (Araujo et al., 2018; Choo et al., 2009; Conafor, 2020; Conanp, 2015; De la Pava et al., 2019; Eberhard, 1983; Franqui-Rivera et al., 2003; Van Itterbeeck & Van-Huis, 2012; Lanteri et al., 2002; Pardo-Locamo et al., 2005, 2019; Salas, 1980; Vargas et al., 2013; Vaurie, 1970a; Vergara-Navarro et al., 2021; Zaragoza-Caballero et al., 2015). Like other dryophtorines, R. barbirostris shows a marked preference for plants in a state of stress, weakened or dead. These adverse conditions, commonly associated with intensive agricultural practices such as prolonged drought, monodominance, and the presence of diseases, create an environment conducive to the development of this weevil (Anderson & Marvaldi, 2014; Oberprieler et al., 2007; Zimmerman, 1968a, b, 1993). Its distribution includes humid climates from central Mexico, Central America, Venezuela, Ecuador, Guyana, Brazil, Peru, Suriname, Uruguay, and Argentina (Pardo-Locardo et al., 2019; Zaragoza-Caballero et al., 2015).
Rhinostomusbarbirostris is distributed in approximately one-third of the Mexican territory (Campeche, Chiapas, Guerrero, Jalisco, Nayarit, Oaxaca, Puebla, Tabasco, Quintana Roo, Veracruz, and Yucatán), and there are few records of damage and affectation by this beetle (Conafor, 2020). Basic aspects of biology, distribution, strategies, and control measures in R. barbirostris have not yet been addressed in Mexico; therefore, the potential damage to palm populations in the territory is unknown. In Central and South America, the damage caused to palms has led to establishing control methods, the most practiced is the early removal of infested plants, in addition to treating them with insecticides and destroying them once the disease is diagnosed (Franqui-Rivera & Medina-Gaud, 2003). Recently, in Oaxaca (2020) and Puebla (2015) records of the bearded weevil in the sweet palm Brahea dulcis have been reported in communities of the Tehuacán-Cuicatlán Biosphere Reserve. The sweet palmnative to Mexico and Central America is distinguished by its solitary stem, large leaves, and glaucous green color(Pavón et al., 2006; Rzedowski, 2006; SINAT & Semarnat, 2009). It lives in dry and semi-arid areas, mainly on limestone soils. It is found in a wide geographic region, from northern Veracruz to Central America, inhabiting diverse ecosystems such as dry forests, scrubland and oak forests (Quero, 1994). The main threat to the survival of this species is human activity. The unsustainable extraction of leaves for artisanal and construction purposes, together with extensive habitat degradation caused by agricultural expansion, livestock grazing, and urban development, has resulted in a pronounced reduction of its populations (Rangel-Landa et al., 2014; Rzedowski, 2006). In Mexico, the use of “Mexican sweet palm” is regulated by various laws and regulations, such as the General Wildlife Law and NOM-059-SEMARNAT-2001, which categorize species and establish protection measures (López-Serrano et al., 2021; Semarnat, 1997, 2002).
Given the limited information available, this study aimed to characterize the damage to the green palm (B. dulcis) and determine the incidence caused by R. barbirostris within the Tehuacán-Cuicatlán Biosphere Reserve to enhance understanding and management of this weevil in Mexico. From field surveys, we generated basic descriptive information about its biology and interaction with this plant. Likewise, we analyzed the temporal and spatial distribution of the weevil based on historical records of specimens deposited in entomological collections and online databases from Mexico. The gathered information will enhance the understanding of biological and behavioral variation, summarize the weevil’s distribution pattern, offer insights into its interactions, and help identify both sampled areas and potential monitoring sites.
Materials and methods
The Tehuacán-Cuicatlán Biosphere Reserve (TCBR) is located in southeastern Puebla and northwestern Oaxaca, it is the arid region with the greatest biodiversity in North America and 1 of the places with the greatest presence of cacti, flora, and fauna (Conanp, 2013; Conafor, 2023; Sinco-Ramos et al., 2021). With just over 4,900 km2, the Reserve is populated by the densest forests of columnar cacti with 45 of the 70 known species in the world (Pérez-Irineo & Mandujano-Rodríguez, 2020).
In February 2022, the Chochonteca zone of the Cañada Region covering Santa María Almoloyas, San Juan Tonaltepec, San Pedro Nodon, San Pedro Jocotipac, San Pedro Jaltepetongo, Santa María Ixcatlán, Santa María Cotahiuxtla, and Santiago Nacaltepec within the TCBR was visited. Agricultural areas were also surveyed due to the presence of the bearded weevil (R. barbirostris), which was observed to be affecting wild specimens of the green palm (B. dulcis), causing damage to their development and even threatening their survival. Guided tours were conducted by communal landholders, during which damage to the host plant was documented through photographs and field reports. Additionally, live specimens at various developmental stages were collected for analysis and taxonomic identification at the Collection Nacional de Insectos, Instituto de Biología, UNAM (CNIN-UNAM). Morphological identification of adults was carried out using the keys of Vaurie (1970a), based on the external morphological characters of the eyes, pronotum, antennal scape, and the sculpture of the tibia and femur. Identification was complemented with the additional morphological criteria of Morrone and Cuevas (2002). Photographs of specimens were taken with a Digital Rising Cam 16 MP attached to a RisingTech UrCMOS stereoscope.
To characterize the palms and quantify the degree of infestation, we conducted transects in the damaged areas. Affected palms were identified and documented with field notes and photographs. For each infested palm, we determined the following variables to assess the characteristics of the damage caused by R. barbirostris: height of the perforations on the trunk (HP), diameter of the perforations observed (DOP), and number of perforations present in 10 cm2 (NP).
The historical, current, and potential distribution of R. barbirostris were determined using data obtained from various databases. Records were consulted and downloaded from the Global Biodiversity Information Facility (GBIF 2023), iNaturalist (2023). The specimens examined in this study were obtained from the following collections: AMNH (American Museum of Natural History, New York, USA.), ASU (Arizona State University, Charles W. O’Brien Collection), CEAM (Centro de Entomología y Acarologia, Instituto de Fitosanidad, Colegio de Postgraduados, Montecillos), CECR (Centro de Referencia, SENASICA-SAGARPA), Conafor (Comisión Nacional Forestal, México), CNIN-UNAM (Colección Nacional de Insectos, Universidad Nacional Autónoma de México), DGSV (Dirección General de Sanidad Vegetal, SENASICA), ECOSUR (El Colegio de la Frontera Sur, Colección Entomológica San Cristóbal y Tapachula, Chiapas, México), MZFC (Museo de Zoología, Facultad de Ciencias, UNAM, Mexico), and UACH (Universidad Autónoma Chapingo, Colección del Departamento Forestal).
In total, 506 records were obtained, from which the following information was summarized: total registers with geographic coordinates, year range, collections, with maximum registers, countries, latitude range, longitude range, main collector, and main identifiers. All records were included in a database; records without geographic coordinates were geopositioned with Google Earth (2020). To determine the feeding spectrum of R. barbirostris, a table of the registered hosts was prepared and graphed according to the number of reports per host and species.
The potential distribution model for R. barbirostris was based on 655 georeferenced occurrence records. This final dataset is the result of a rigorous preprocessing process that ensured quality by eliminating errors and duplicates (Guisan & Zimmermann, 2000; Sillero, 2011; Warren, 2012). The records cover the entire known range of the species in Mexico, Central America, and South America and were the input data for Wallace package (Carretero & Sillero, 2016; Kass et al., 2023).
A database was compiled with presence records covering the entire range of the species. This strategy was adopted because niche models are highly sensitive to the spatial distribution of data. Using subsets of the range can violate key assumptions, such as equilibrium with the environment and niche homogeneity, potentially generating biased results. Therefore, including all available records ensures a more complete and robust representation of the species’ ecological niche. For the record, the values of the 19 bioclimatic variables of WorldClim 2 were considered (Fick & Hijmans, 2017). Extraction was done in QGIS 3.36.3 using the Point sampling tool plugin (https://github.com/borysiasty/pointsamplingtool). After the extraction, a multivariate normality test was performed on the values of the variables with the R package mvn (Korkmaz et al., 2014), which was negative. The distribution of environmental variables was examined to understand the species’ ecological niche. A normal distribution would suggest that the species has a narrow preference for certain conditions, while a non-normal distribution indicates that it can tolerate a wider range of environments. Since our species’ variables do not follow a normal distribution, it was decided to model the entire species distribution rather than a fraction of it. This decision avoids a bias in the model that could underestimate its adaptive capacity to a wider range of environmental conditions (Brown, 1984).
To evaluate the correlation among variables, a Spearman correlation analysis were performed using the R core function. Using the Wallace interface, next, the parameters specified are described (Kass et al., 2023). Those variables with a correlation value less than 0.85 were selected. Thus, the selected variables were: Bio01, Bio02, Bio03, Bio04, Bio05, Bio06, Bio08, Bio12, Bio14, Bio18, which were used to build the model at a resolution of 5 arcmin (~ 20km). The model calibration area (M) comprised Mexico, Central America, and South America, which includes all records of R. barbirostris. Records were thinned to 55.55 km, reducing the records to 220 (Aiello-Lammens et al., 2015; Boria et al., 2014).
A background of 5,000 points was generated. The selection of the number of background points was based on the common MaxEnt practice, which sets 10,000 points by default. Although there is no consensus on the optimal number, recent literature indicates that this value is an adequate baseline, depending on the study area and the resolution of the variables (Merow et al., 2013; Phillips et al., 2017). It is recognized that an excessive number of points can artificially inflate the AUC value (Sofaer et al., 2019); however, evaluating an optimal number of background points through metrics such as the Boyce index or True Skill Statistics (Valavi et al., 2022) is beyond the scope of this study. Therefore, the default value of 10,000 points was chosen as a justified balance between study area coverage and computational feasibility.
For model validation, a non-spatial Jackknife partition was used, applying the Maxent algorithm with a linear response (Elith et al., 2011; Phillips et al., 2006). This data partition was selected for model evaluation due to the small sample size (Shcheglovitova & Anderson, 2013). Three models were generated with a regularization multiplier (1, 1.5, and 2). The model with the highest AUC was selected (with a regularization multiplier of 1), whose value of 0.73 is considered acceptable. The “Minimum Training Presence” was used to generate the binary map. The map was exported to a TIFF file and imported to QGIS (QGIS.org, 2024) to calculate the percentage of area of the model in each of the 32 federal entities of Mexico.
In addition to the calculation by state, an analysis was conducted to determine how the potential distribution of R. barbirostris is distributed across the biomes of the national territory. Using the biome layer, a binary map of the species’ potential distribution was overlaid. Subsequently, the total area (km2) and the percentage of area projected by the model for each biome were quantified, allowing for a deeper understanding of the species’ preference for certain types of ecosystems.
Results
Rhinostomusbarbirostris specimens collected in the Tehuacán-Cuicatlán Biosphere Reserve were found associated with deteriorated B. dulcis palms. Species identifications were confirmed by the presence of a slightly raised frontal keel between the eyes (Fig. 1a-b), front femur virtually impunctate, front tibia on inner edge with long teeth; front tibia of male without hairs (Fig. 1c-e). Adults were collected with the respective larvae and pupae. The presence of R. barbirostris was recorded in wild populations of B. dulcis in Santa María Almoloyas, San Juan Tonaltepec, San Pedro Nodón, San Pedro Jaltepetongo, Santa María Ixcatlán, Santa María Cotahuixtla, and Santiago Nacaltepec, Oaxaca, Mexico. In addition to these sites, surveys were conducted in other agricultural centers such as San Pedro Jaltepetongo and San Pedro Jocotipac, where the presence of R. barbirostris was also documented.
In all colonized palms, the beetles caused notable damage to the base of stems and foliage, which disrupted their growth and, in some cases, led to the death of the affected specimens. The palm damage can be observed in individual or gregarious palms. In the gregarious stems, some of the palms display healthy foliage, meanwhile others exhibit signs of deterioration, such as browning and wilting leaves, indicating early damage from the insect infestation (Fig. 2a-c). In individual palms, the lateral shoots that grow on the main stem of the palm trees also presented these symptoms (Fig. 2d). The affected specimens showed the foliage dry and drooping, with the leaves appearing dead and lifeless. In the gregarious spots, it is evident that the damage is progressive, affecting the palms around the infested 1 (Fig. 2b). In the palms completely succumbed to the infestation, the stem is visibly blackened and decayed, standing as a stark remnant of the once-living tree, showing the lethal impact of the weevil. These palms are dry, and the foliage has collapsed (Fig. 2d). A close-up of the palm trunk revealed multiple boreholes and resin exudation; the damage is severe, with the bark visibly compromised by larval activity, showing extensive boring damage (Fig. 2e-f, 3d-f). The holes are located in the ground near the base of the stipe at the average height of 1.35 m, the holes secrete brown resin, with the smell of fermented tissue (Fig. 3b-c)
A single, distinct borehole in the trunk is created by an adult weevil for laying eggs or by larvae boring through the wood (Fig. 3a-b). The clean-cut appearance of the hole suggests recent activity (Fig. 3b). Numerous circular holes indicate the advance of colonization where multiple larvae have burrowed, compromising the structural integrity of the palm and contributing to its decline (Fig. 2f, 3a). Internal damage is characterized by a discolored and brittle texture of the bark, the texture of the wood appears spongy and degraded, illustrating the destruction caused by the feeding larvae (Fig. 3d-f).
The female lays her eggs on decomposing organic matter, such as fallen trunks or palm tree remains from previous crops, where she builds galleries. Larvae feed in the stipe-trunk, forming wide galleries toward the bulb of the palm. Larva built pupal chambers where the immature individuals develop until they emerge from the host (Fig. 3e-f). The different larval stages of the bearded weevil develop in these substrates, and the larvae can be found buried at a depth of 30-40 cm (Fig. 3f). Within the stipe and bulb tissues, different life stages of the weevil can be found including larvae, pupae, and adult insects (Fig. 3e). In only one specimen of the palm, dozens of beetles can be found (Fig. 3f). The variables HP, DOP and NP reported the following information obtained at the sampling sites: HP (0.85-1.35 m; µ = 1.10 m), DOP (45-158 mm, µ = 107.4 mm) and NP (1-17, µ = 7). Adult individuals and larvae are responsible for the damage (Fig. 3a-c), which begins with the excavation of tunnels in the soil until reaching the basal bulb, which is completely consumed. Subsequently, the insect ascends the plant to feed on the apical tissue, causing the death of the plant. The presence of mounds of fresh soil, like small hills built by the insect, reveals its presence, especially in areas with decaying trunks, which serve as incubators for future generations (Fig. 3e-f). Additionally, in B. dulcis palms with damage attributed to R. barbirostris, the presence of beetles from the Zopheridae family (Zopherus spp.) was recorded (Fig. 3a) in areas of plant tissue that were in an advanced state of deterioration and decomposition, confirming a direct association with the initial damage caused by the weevil.
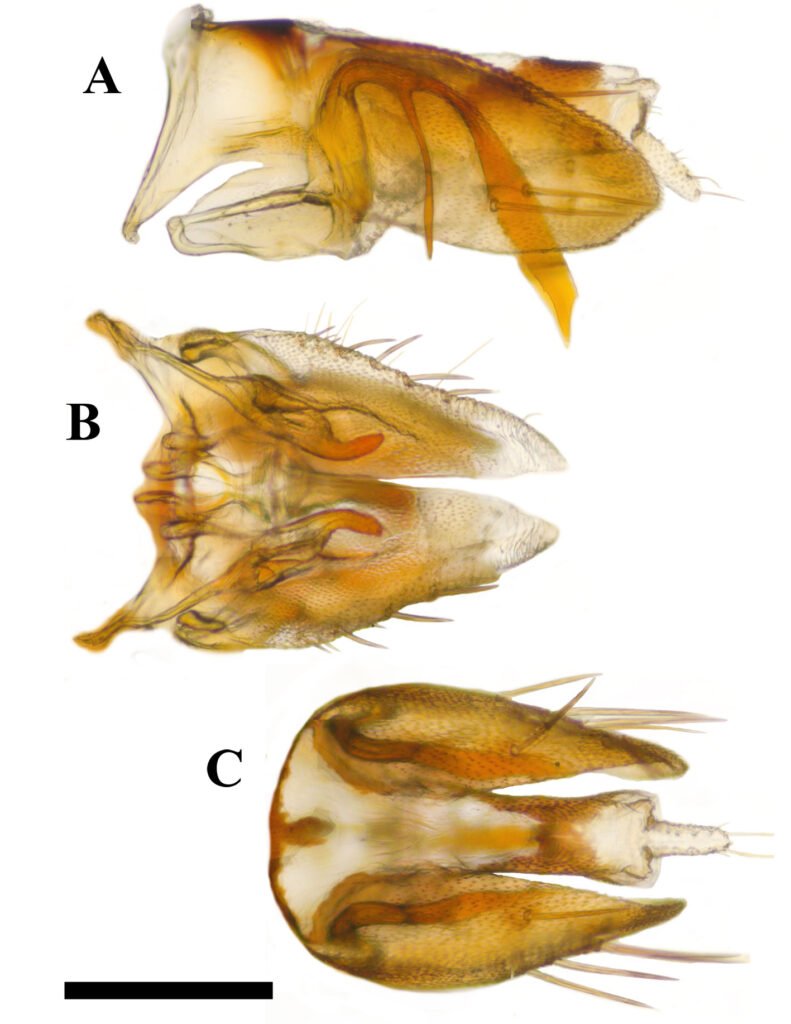
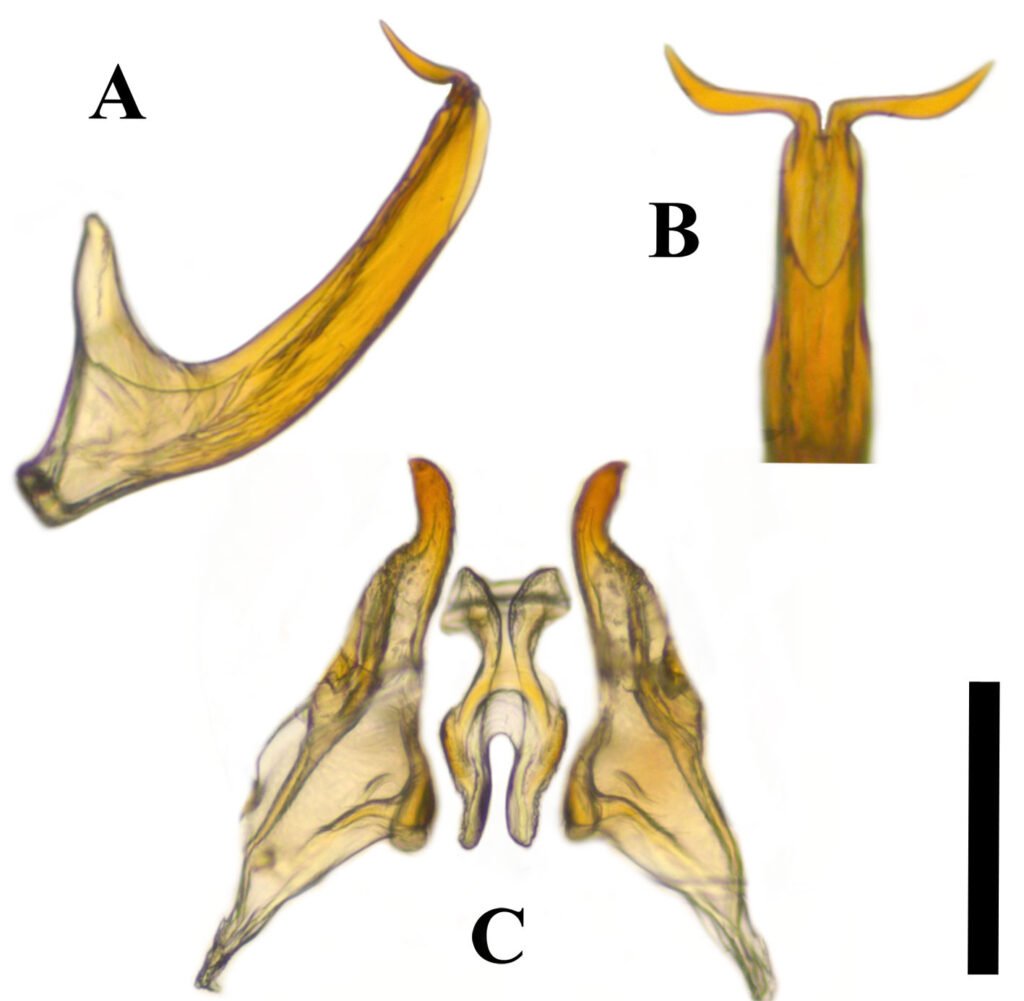
Figure 1. a) R. barbirostris, dorsal view; b) presence of a slightly raised frontal keel between the eyes; c) R. barbirostris in lateral view; d) tibia on inner edge with long teeth (male); e) front tibia without hairs (male). Photographs by Mauricio Ramírez Hernández.
A total of 506 geographic records of the bearded weevil were obtained in 2023 in Mexican territory, 3 from AMNH, 1 from ASU, 4 from CECR, 17 from CEAM, 67 from CNIN-UNAM, 123 from Conafor, 6 from DGSV, 2 from ECOSUR, 214 from iNaturalist, 4 from MZFC and 4 from UACH (Table 1). The oldest record for this species corresponds to Playa Vicente (17°49’46.98”N, 95°48’47.12”O) and Actopan (19°30’16.14”N, 96°36’56.82”O), Veracruz reported by Champion (1910) under the name Rhinabarbirostris. In Mexico, the number of records from 1927 to 2006 was 101 records; from 2007 to 2023 the collection of specimens and sightings increased to 337 records and 68 records without collection data (Table 1). In last decade (2013-2013), sampling efforts increased, mainly in the states of: Chiapas (23), Jalisco (24), Oaxaca (147), Quintana Roo (26), Sinaloa (24) and Veracruz (46). With a total of 2 records, the Sierra Negra de Puebla was the locality with the lowest number of records for the bearded weevil. The bearded weevil was most abundantly collected from march to december with a total of 251 specimens, corresponding to 49.59% of a total of individuals.
Based on the records of the collections and consulting the available literature it was possible to obtain information about the host plant of the adult individuals of R. barbirostris present in Mexico and throughout its distribution. These correspond to Attalea funifera, A. maripaA. pindobossu, A. piassabossu, Bactris gasipes, Brahea dulcis, Cocos botryophora, C. coronata, C. nucifera, Elaeis guineensis, Euterpe oleraceae, Mauritia flexuosa, Oenocarpa bacaba, Syagrusromanzoffiana and S. schizophylla (Table 2).
Figure 2. Bearded weevil host (B. dulcis) showing: a) healthy host; b) infested palm at RBTC; c-e) weakening of foliage and decay of the trunk; f) Ring on the trunk or exit hole of adult specimens of R. barbirostris. Photographs by Leticia Soriano Flores.
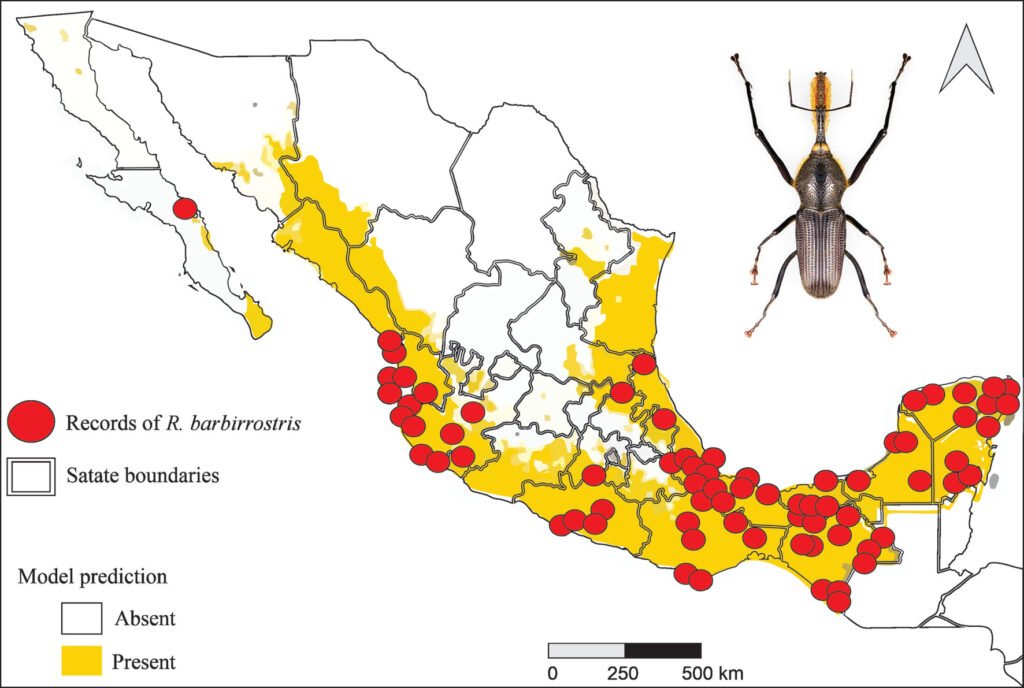
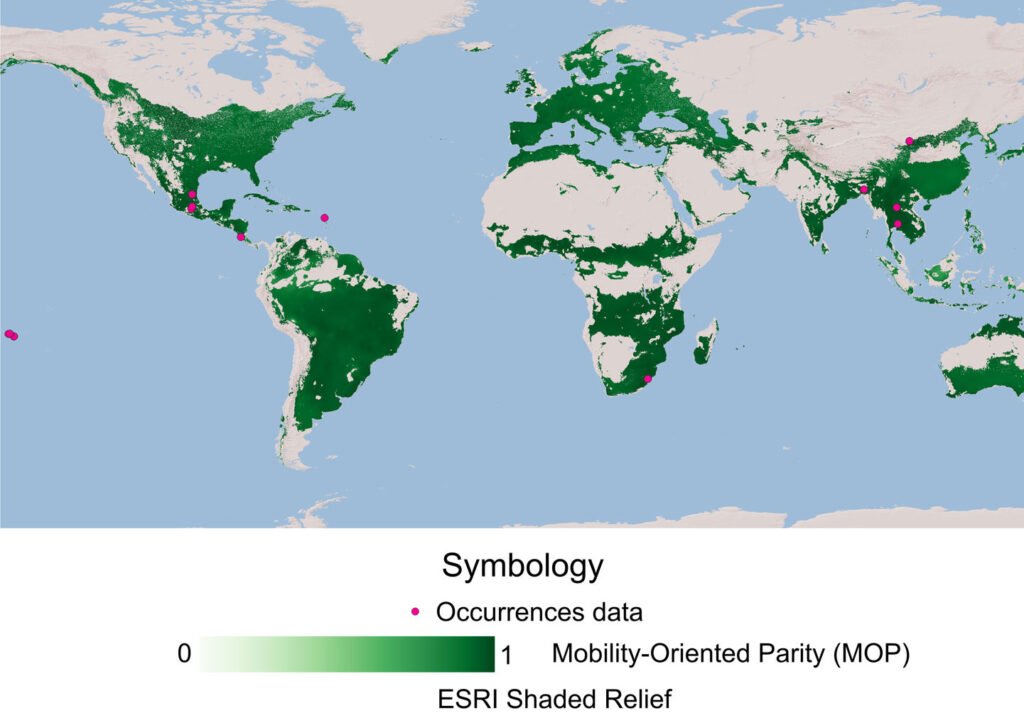
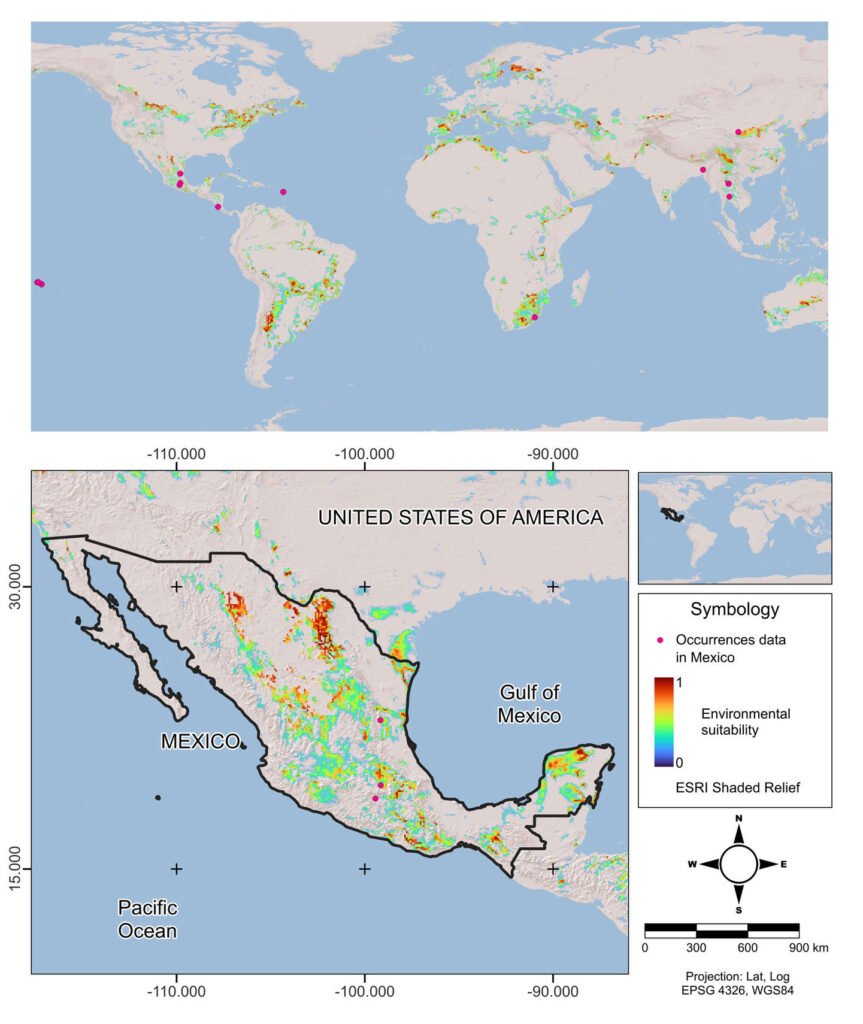
The potential distribution model developed with the Wallace platform revealed that this species has a surprisingly wide distribution in Mexico (Fig. 4). The model predicts the most likely areas for the species based on historical records of its presence (red dots on the map). To ensure the robustness of the analysis, we chose a model with a linear feature and a regularization multiplier of 1, allowing us to avoid overfitting and obtain more reliable results. Although AUC curves are a common metric, they are not included here due to differences in how Wallace calculates them compared to the MaxEnt Java application. Overall, the findings indicate that the species is capable of inhabiting a large portion of the national territory; although the species is present in many regions, a higher concentration of records was observed in certain areas, such as Sierra Madre Oriental (SMOR), Sierra Madre Occidental (SMOC), and Sierra Madre del Sur (SMAS). Although the distribution of this taxon is correlated with environmental factors such as vegetation, altitude, and climatic conditions, human intervention emerges as a determining factor in its dispersal pattern. Activities such as the movement of infested plant material through agricultural campaigns or the transport of goods act as key vectors, overcoming natural geographic and ecological barriers. This accelerated dispersal allows the species to colonize new territories with unprecedented speed and scope. Therefore, for a complete understanding of its current distribution, it is essential to complement the analysis of environmental factors with a thorough assessment of how human activities are shaping the geography of the species.
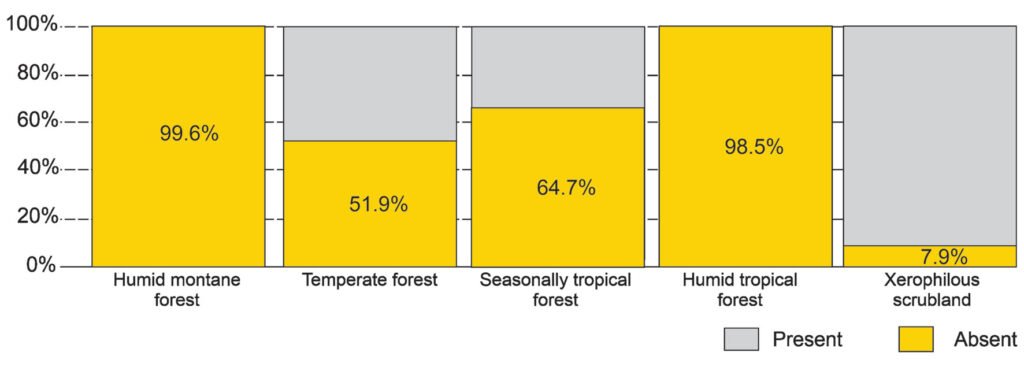
The potential distribution model covers almost the entire surface area in the national territory of 2 of the biomes: humid mountain forest (99.6%) and humid tropical forest (98.5%); in 2 others, close to half: temperate forest (51.9%) and seasonally dry tropical forest (64.7%); and only 7.9% in xerophilous scrubland (Fig. 5).
Figure 3. a) Damage caused by adult individuals of the bearded weevil on the green palm; b) other taxa associated with the damaged host; c) excavation tunnels in B. dulcis at TCBR; d, e) immature forms of the bearded weevil; f) galleries and pupal chambers where the immature individuals and presence of mounds of fresh earth, in areas with decomposing trunks for incubation. Photographs by Leticia Soriano Flores.
Discussion
Our field observations provide detailed evidence of the damage associated with R. barbirostris colonizing B. dulcis and allow for the description of key aspects of the biology of this weevil. As in Brazilian coconut plantations, we documented that the axils of older leaves of B. dulcis serve as a refuge for R. barbirostris (Ferreira, 2008). Although we did not record beetle eggs, the presence of adults in older axils suggests that oviposition may occur in the stem, and once hatched, larvae penetrate and form galleries (Ferreira, 2008; Lacerda-Moura et al., 2013). One of the characteristics associated with the colonization holes was the exudation of a dark reddish-brown liquid, which is consistent with gummosis disease or trunk bleeding caused by the fungus Ceratocystis paradoxa (Dade) C. Moreau anamorph in the coconut palm, which has been associated with the presence of R. barbirostris (Ferreira et al., 2007). Further studies are needed to determine whether the susceptibility of sweet palm is related to the presence of this fungus, as it is reported that C. paradoxa releases a semiochemical that attracts these beetles (Lacerda-Moura et al. 2013). Furthermore, that R. barbirostris is also mentioned as a vector of the nematode Bursaphelenchus cocophilus (Cobb.) (Goodey, 1933,1960) in the coconut groves (Franco, 1964).
Figure 4. Potential distribution and records of bearded weevil in Mexico.
Larval galleries in the bulb of B. dulcis, as well as the progressive destruction of apical tissues, reflect a strategy of underground colonization of R. barbirostris followed by upward expansion into the aerial parts of the plant. This behavior is consistent with that of R. barbirrostris in Brazil (Lacerda-Moura et al., 2013) and that of other palm-associated curculionids, in which the larval stage is the most destructive, feeding of internal tissues and compromising the structural integrity of the host (Faleiro, 2006; Soroker et al., 2015).
The coexistence of larvae, pupae, and adults within a single palm individual indicates that R. barbirostris exhibits a continuous life cycle inside its host, ensuring the local persistence of its populations. Similar patterns have been reported for the red palm weevil Rhynchophorus ferrugineus, where overlapping generations occur within the same palm and infestations are difficult to eradicate once established (Abraham et al., 1998; Dembilio & Jacas, 2011). The ability of R. barbirostris larvae to burrow 30-40 cm underground and construct pupal chambers in degraded tissues highlights a high degree of adaptation to concealed and protected habitats, reducing exposure to natural enemies and environmental fluctuations. Such cryptic development has also been documented in Rhynchophorus palmarum, a major pest of coconut and oil palms in the Neotropics (Griffith, 1987), and in Metamasius hemipterus, another palm-infesting weevil (Weissling & Giblin-Davis, 1993).
Table 1. Records of Rhinostomus barbirostris in Mexico by date, federal entity, geographical coordinates and site 556 of specimen consultation.
Date
Locality
Latitude
Longitude
Altitude (m asl)
Host
Number of specimens
Source
24/03/1973
35kms al sur de Mulejé, Baja california, Mexico
26°53’29.82”
111°59’0.79”
16
N/A
24
CNIN-UNAM
24/03/1973
25kms al sur de Mulejé, Baja California, Mexico
26°53’31.95”
111°59’0.91”
30
N/A
6
CNIN-UNAM
20/10/1971
Campeche, Mexico
19°49’48.15”
90°32’5.61”
3
N/A
1
CNIN-UNAM
16/12/2016
Calakmul Municipality, Campeche, Mexico
18°25’37.86”
89°42’1.62”
232
N/A
4
iNaturalist
10/02/2021
Municipio de Carmen, Campeche, Mexico
18°30’59.31”
91°26’36.71”
0
N/A
2
iNaturalist
24/02/2022
24327 Campeche, Mexico
18°39’8.16”
91°33’29.90”
0
N/A
2
iNaturalist
27/05/1981
Ejido 2 de Mayo, Chiapas, Mexico
15°1’46.44”
92°8’55.34”
698
N/A
1
DGSV
27/05/1981
Campo experimental Rosario Izapa, Chiapas, Mexico
14°58’30.32”
92°9’19.13”
445
N/A
1
DGSV
27/03/1982
Benemérito de las America, “s-palapa”, Chiapas, Mexico
16°30’51.36”
90°39’14.75”
141
N/A
1
MZFC
19/05/1984
Palenque, Chiapas, Mexico
17°30’39.36”
91°59’34.98”
71
N/A
1
CNIN-UNAM
28/04/1986
Ocosingo Chajul, Reserva Montes Azules, Chiapas
16°35’51.21”
91°9’9.44”
747
N/A
3
CNIN-UNAM
25/07/2013
Chiapas, Mexico
16°45’24.82”
93°7’45.25”
535
N/A
1
iNaturalist
10/10/2015
Las Nubes, Chiapas, Mexico
16°11’44.85”
91°20’20.73”
287
N/A
2
iNaturalist
25/07/2017
Tapachula, Chiapas, Mexico
14°54’19.82”
92°15’48.29”
173
N/A
2
iNaturalist
N/A
Tapachula, Chiapas, Mexico
14°54’19.82”
92°15’48.29”
173
N/A
4
N/A
12/12/2018
Ocosingo, Chiapas, Mexico
16°54’24.54”
92°5’39.53”
849
N/A
3
iNaturalist
03/01/2019
Villa Comaltitlán, Chiapas, Mexico
15°12’45.24”
92°34’34.50”
34
N/A
1
iNaturalist
29/05/2019
Finca La Alianza, Municipio Cacahoatán, Chiapas, Mexico
15°2’36.93”
92°10’52.66”
687
N/A
1
ECOSUR
09/06/2019
Ejido las nubes, Municipio Tapachula, Chiapas, Mexico
14°52’26.05”
92°16’4.11”
130
N/A
1
ECOSUR
02/08/2019
Calle San José, Tapachula de Córdova y Ordoñez, Chiapas, Mexico
14°52’39.33”
92°16’25.01”
121
N/A
5
iNaturalist
30/04/2020
Chiapas, Mexico
16°45’24.82”
93°7’45.25”
535
N/A
1
iNaturalist
05/01/2020
Tapachula, Chiapas, Mexico
14°54’19.82”
92°15’48.29”
173
N/A
2
iNaturalist
17/04/2021
Hotel Argovia Finca Resort, Chiapas, Mexico
15°7’31.20”
92°17’55.92”
625
N/A
4
iNaturalist
27/11/2022
29935 Ocosingo, Chiapas, Mexico
16°46’6.24”
90°56’37.83”
171
N/A
1
iNaturalist
N/A
Chiapas, Mexico
16°45’20.31”
93°7’45.41”
533
Coccos nucifera
1
CEAM
N/A
Colima, Mexico
19°14’42.27”
103°43’26.51”
503
N/A
1
iNaturalist
X/05/1948
Cacahuamilpa, Guerrero, Mexico
18°40’53.77”
99°30’19.96”
1,266
N/A
1
iNaturalist
19/09/1963
Guerrero, Mexico
17°26’21.02”
99°32’42.29”
2,136
N/A
1
DGSV
14/11/1971
Técpan de Galeana, Guerrero, Mexico
17°13’22.27”
100°37’42.16”
45
N/A
1
CNIN-UNAM
Table 1. Continued
Date
Locality
Latitude
Longitude
Altitude (m asl)
Host
Number of specimens
Source
14/11/1971
Rodecia, Guerrero, Mexico
17°12’24.68”
100°41’56.90”
22
N/A
3
CNIN-UNAM
03/11/1976
Ejido de Tenexpa, Tecpán de Galeana, Guerrero, Mexico
17°11’0.17”
100°40’16.97”
16
Coccos nucifera
7
CEAM
02/02/1976
Petatlán, Guerrero, Mexico
17°32’20.41”
101°16’12.70”
35
Coccos nucifera
2
CEAM
02/02/1976
Petatlán, Guerrero, Mexico
17°32’20.41”
101°16’12.70”
35
Coccos nucifera
2
CECR
24/02/1976
Petatlán, Guerrero, Mexico
17°32’20.41”
101°16’12.70”
35
Coccos nucifera
4
CEAM
24/02/1976
Petatlán, Guerrero, Mexico
17°32’20.41”
101°16’12.70”
35
Coccos nucifera
2
CECR
X/05/1985
Cacahuamilpa, Guerrero, Mexico
18°40’53.77”
99°30’19.96”
1,266
N/A
1
CNIN-UNAM
10/07/2007
Guerrero, Mexico
17°26’21.02”
99°32’42.29”
2,136
N/A
1
iNaturalist
07/10/2007
Reforma 4ta Etapa, Chilpancingo de los Bravo, Guerrero, Mexico
17°32’11.71”
99°28’49.61”
1,330
N/A
3
iNaturalist
28/12/2022
Guerrero, Mexico
17°26’21.02”
99°32’42.29”
2,136
N/A
1
iNaturalist
28/12/2022
Chilpancingo de los Bravo, Guerrero, Mexico
17°32’21.67”
99°29’4.65”
1,302
N/A
3
iNaturalist
06/10/2022
Puerto del gallo, Guerrero, Mexico
17°28’1.50”
100°10’32.63”
2,252
N/A
1
CNIN-UNAM
17/09/2016
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
1
iNaturalist
23/09/2016
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
2
iNaturalist
06/12/2017
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
2
iNaturalist
27/06/2018
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
2
iNaturalist
09/11/2019
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
1
iNaturalist
09/12/2019
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
2
iNaturalist
25/07/2020
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
1
iNaturalist
12/11/2021
Cabo Corrientes, Jalisco, Mexico
20°19’6.02”
105°19’17.70”
607
N/A
3
iNaturalist
13/06/2022
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
20
N/A
1
iNaturalist
09/09/2022
Yelapa, Jalisco, Mexico
20°29’16.29”
105°26’53.44”
19
N/A
2
iNaturalist
25/10/2022
Jalisco, Mexico
20°39’32.98”
103°20’57.86”
1,545
N/A
1
iNaturalist
25/03/2022
Paseo de las Conchas, Puerto Vallarta, Jalisco, Mexico
20°35’15.46”
105°14’29.19”
99
N/A
3
iNaturalist
26/10/2022
Cihuatlán, Jalisco, Mexico
19°14’15.50”
104°34’6.38”
42
N/A
2
iNaturalist
06/07/2023
48898 Jalisco, Mexico
19°16’47.08”
104°47’9.22”
66
N/A
1
iNaturalist
N/A
La Lima, Jalisco, Mexico
20°0’7.13”
103°59’5.06”
1,228
N/A
1
iNaturalist
18/02/1981
Cuernavaca, Morelos, Mexico
18°55’27.04”
99°13’17.61”
1,510
N/A
1
MZFC
12/06/2012
La puntilla, Nayarit, Mexico
22°28’8.45”
105°43’21.08”
8
N/A
1
DGSV
28/10/2013
Los Otates, Nayarit, Mexico
21°42’10.89”
105°22’29.99”
6
N/A
1
DGSV
08/11/2019
Av. Revolución 7, Sayulita, Nayarit, Mexico
20°52’7.35”
105°26’14.41”
9
N/A
2
iNaturalist
19/10/2020
Bahía de Banderas, Nayarit, Mexico
20°48’26.06”
105°14’53.25”
39
N/A
2
iNaturalist
30/12/2022
Bahía de Banderas, Nayarit, Mexico
20°48’26.06”
105°14’53.25”
39
N/A
2
iNaturalist
23/04/2023
Compostela, Nayarit, Mexico
21°14’11.66”
104°54’2.86”
879
N/A
2
iNaturalist
N/A
Acaponeta, Nayarit, Mexico
22°29’45.35”
105°21’46.40”
30
N/A
1
N/A
N/A
Compostela, Nayarit, Mexico
21°14’12.03”
104°54’2.61”
852
N/A
1
N/A
N/A
San Blás, Nayarit, Mexico
21°32’28.60”
105°17’4.98”
5
N/A
1
N/A
N/A
Tepic, El Cora, Nayarit, Mexico
21°30’14.30”
104°53’40.43”
932
N/A
1
N/A
1-4/09/1947
Tolosa, Oaxaca, Mexico
22°31’20.28”
101°21’23.68”
2,095
N/A
3
AMNH
30/05/1959
S. Carlos, Palomares N de Oaxaca, Mexico
17°4’22.64”
96°43’35.52”
1,604
N/A
1
CNIN-UNAM
X/07/1970
Sierra de Juárez, Oaxaca, Mexico
17°34’46.53”
17°34’46.53”
1,730
N/A
1
CNIN-UNAM
01/07/1996
Topiltepec, Oaxaca, Mexico
17°26’14.23”
97°20’41.66”
2,173
Brahea dulcis
4
UACH
27/05/2014
Oaxaca, Mexico
17°4’23.33”
96°43’35.80”
1,589
N/A
1
iNaturalist
02/04/2015
Oaxaca, Mexico
17°4’23.33”
96°43’35.80”
1,589
N/A
3
iNaturalist
16/01/2018
Loma Bonita, Oaxaca, Mexico
18°6’34.71”
95°52’49.39”
35
N/A
4
iNaturalist
16/01/2018
Oaxaca, Mexico
17°4’23.33”
96°43’35.80”
1,589
N/A
1
iNaturalist
19/06/2018
Tuxtepec-San Felipe Jalapa de Díaz, San Bartolo, Oaxaca, Mexico
16°57’13.90”
96°42’28.63”
1,520
N/A
2
iNaturalist
28/05/2018
Calle9 de Marzo, San José de las Flores, Oaxaca, Mexico
Northern South America in Colombia, Venezuela, Trinidad and Tobago (Caribbean), Peru, Guianas, Ecuador, Bolivia and Brazil.
Bactris gasipaes
Colombia
Pacific Colombian coast
Choo et al., 2009; Deloya & Gasca, 2018; Pardo-Locarno et al., 2005; Pardo-Locarno et al., 2019
Chontaduro palm
Bolivia, Colombia, Costa Rica, Ecuador and Panama.
Brahea dulcis
Mexico
Chochonteca in the Cañada region in Oaxaca; Reserva de la Biosfera Tehuacán-Cuicatlán Oaxaca
Conafor, 2020; Conanp, 2015; Zaragoza-Caballero et al., 2015
Mexican sweet palm, soyale or soyale palm
From Mexico to Peru
Cocos botryophora, C. coronata, C. nucifera
Colombia, Puerto Rico
Colombian western regions, Mayagüez
Vaurie,1970a; Eberhard,1983; Franqui-Rivera et al., 2003; Vergara-Navarro et al., 2021
Coconut palm
Pantropical
Elaeis guineensis
Ecuador, Costa Rica
La Loja; Quepos and Parrita
De la Pava et al., 2020; Lanteri et al., 2002; Vaurie, 1970a; Vergara-Navarro et al., 2021
Oil palm
Tropical Africa; grown in the tropics.
Euterpe oleraceae
Venezuela
N/D
Choo et al., 2009
Huasaí, huasaí palm
Northern Brazil, French Guiana, Suriname, Guyana, Peru, Bolivia, in Esmeraldas north of Ecuador, Venezuela, eastern Panama, Magdalena Medio and the Pacific and Amazon regions in Colombia
Mauritia flexuosa
Venezuela, Peru
N/D, Peruvian Amazone
Choo et al., 2009; Van Itterbeeck & Van Huis, 2012;
Moriche, moriche palm
Wide distribution in central and northern South America: Ecuador, Bolivia, Brazil, Peru, Colombia, Guyana, Venezuela, Suriname, Trinidad and Tobago
Oenocarpa bacaba
Venezuela
Venezuelan Amazon
Choo et al., 2009
Milpesillo
Guyana from Colombia to Brazil
Syagrus romanzoffiana. S. schizophylla
Brazil
Alto Paraná ecoregion (Paranaense Forest)
Vaurie, 1970a; Araujo et al., 2018
feathery coconut
It is native to southern Brazil, Paraguay, Uruguay, and the Argentine coast.
The infestation in gregarious palms, where damage spreads from a focal plant to adjacent individuals, mirrors the patch-level mortality reported in R. ferrugineus outbreaks in date palm plantations (Faleiro, 2006). Such localized dispersal may be driven by the physical proximity of palm stems, facilitating rapid expansion of infestations. In solitary palms, R. barbirostris was also observed attacking lateral shoots, indicating behavioral plasticity in selecting entry points. This opportunistic colonization strategy parallels that of Metamasius spp., which infest both primary stems and secondary shoots of diverse palm hosts (Weissling & Giblin-Davis, 1993).
The sequential progression of damage beginning with clean perforations indicative of recent oviposition or adult feeding, followed by multiple boreholes in advanced infestations, is consistent with the biology of other palm weevils. In R. ferrugineus, for example, early stages can only be detected through discrete entry holes or frass deposits, whereas advanced infestations produce widespread internal tunneling and crown collapse (Dembilio & Jacas, 2011). Similarly, in R. palmarum, larval feeding not only destroys host tissues but also facilitates the transmission of the nematode Bursaphelenchus cocophilus, the causal agent of red ring disease in coconut (Griffith, 1987). Although no pathogen association was observed in R. barbirostris, the fermentative odors and resin exudation recorded in this study are comparable to the secondary microbial colonization often reported in palms infested by Rhynchophorus.
The consistent presence of Zopherus spp. in highly degraded tissues further illustrates a successional process in which secondary insects exploit the conditions created by the primary borer. Comparable saproxylic successions have been reported in palms infested by species of Metamasius and Rhynchophorus, where decomposing tissues become niches for secondary beetles and fungi (Howard et al., 2001). This suggests that R. barbirostris not only functions as a primary pest but also plays a role in structuring arthropod communities associated with decaying palms.
The variables reported here, including perforation height, diameter, and number quantify the magnitude of damage and confirm that both larvae and adults actively contribute to palm mortality. In other palm weevils, similar diagnostic metrics (e.g., size of entry holes, pattern of galleries, crown symptoms) have been used as reliable indicators for early detection and management (Faleiro, 2006; Fiaboe et al., 2012). The construction of fresh soil mounds at the base of infested palms, recorded in this study, represents an additional diagnostic feature that could be incorporated into monitoring protocols, much like the frass deposits and oozing exudates used to detect R. ferrugineus.
Our findings indicate that R. barbirostris shares several biological and behavioral traits with major palm pests worldwide, such as Rhynchophorus ferrugineus, R. palmarum, and Metamasius spp. Its ability to establish overlapping generations, exploit multiple host tissues, and spread among neighboring palms underscores its potential as a pest of both ecological and economic concern. These parallels suggest that management strategies developed for other palm weevils such as pheromone trapping, sanitary removal of infested material, and cultural practices to reduce breeding sites, may serve as a useful foundation for developing targeted monitoring and control measures for R. barbirostris in Mexico.
Distribution and records. The geographical distribution of R. barbirostris from the lowlands of the Mexican Transition Zone (MTZ) to South America aligns with the classic Neotropical cenocron (Morrone, 2019). Its ecological niche characteristics, particularly temperature as a key environmental factor and its large amplitude, are also congruent with this pattern (Lizardo et al., 2025). This congruence supports the idea that cenocrons, as proposed by the MTZ theory, serve as evolutionary and ecological hypotheses. From these hypotheses, it is possible to make predictions about the ecological niches of species in relation to their patterns of richness and geographical distribution (Lizardo et al., 2025).
All records from entomological collections and databases allowed us to delimit the geographic distribution of R. barbirostris and suggested activity patterns throughout the year, although it is recognized that these data, not obtained from field observations, may present sampling biases. The higher frequency of records at certain times of the year (March to December 2007-2023), as reported in our findings, could be due to historical collecting efforts rather than the inherent biological activity of the species. However, when comparing these results with the existing literature, we found some similarities. For example, Sarwar (2016) reported the presence of weevil pupae (Rhyncophoruspalmarum) in infested palms during May and August, which aligns with the end of the activity window observed in our records. Similarly, Jaramillo-Vivanco’s (2023) fieldwork in Ecuador, conducted between March and December, coincides with the period when our records were most abundant. Unlike what has been documented for other insect groups, such as the genus Ocoaxo, which presents a marked seasonality and synchronizes its life cycle with the rainy season (Cid-Muñoz et al., 2020), our data do not allow us to confirm a similar biological pattern for R. barbirostris. The temporal analysis of the records of these insects suggests that they have a univoltine life cycle, since all the collection data were restricted to 4 months of the year, from the beginning of summer in May, until the beginning of Autumn in October, a period that corresponds to the season of greatest rainfall in Mexico.
The preference of these beetles for freshly fallen trunks, as noted by Eberhard (1983) on a palm trunk in May, could explain why specimen collection is higher in certain months. The presence of the species is strongly linked to resource availability, as found in the fieldwork of López-Zent and Zent (2020), who, over 21 months of direct observation, were able to document the species ecology over time. Their observations, obtained directly from fieldwork, complement and contrast our findings based on historical data. Although our records reflect an apparent peak in activity, it is crucial to consider that ecological context and collection patterns can influence data availability. This integration of historical data and field observations, such as those of the authors, is essential to a more complete and accurate understanding of the biology and ecology of R. barbirostris. Our data point to a concentration of activity, suggesting that this apparent periodicity could be, at least in part, a reflection of historical data collection methods, rather than a strictly biological pattern. For example, some studies indicate that this type of bias can influence the interpretation of distribution patterns and seasonality (Elith et al., 2006; Fourcade et al., 2014). It is inferred that the presence or absence of the species is linked to specific ecological factors, such as the type of ecosystem or the food sources of the adults, which require further field research in order to confirm. The analysis of these data allowed the formulation of assumptions about the spatial distribution and the estimation of the geographical limits of their sightings, as well as the identification of areas suitable for their surveillance.
Accurate taxonomic identification is critical for modeling species distributions (Elith et al., 2006; Elith & Leathwick, 2009; Pearman et al., 2010). Although the taxonomic history of the bearded weevil does not present the ambiguities common in other phytophagous insects, such as the bark beetles (Armendáriz-Toledano et al., 2014a, b; Valerio-Mendoza et al., 2019) of the genus Dendroctonus or the spittlebugs of the genus Ocoaxo (Armendáriz-Toledano et al., 2023; Castro-Valderrama et al., 2017, 2019; Cid-Muñoz et al., 2022), extensive record validation was performed to ensure the reliability of our data. To do so, we reviewed records and specimens from several key entomological collections in Mexico. The identification of each specimen was validated through its external morphological characteristics, allowing us to confirm the species. These findings were consistent with the descriptions and taxonomic keys of Vaurie (1970a, 1970b) and Morrone & Cuevas (2002), which served as the final validation criterion. The review of 506 specimens, representing 31.9% of the total records through 2023, revealed no inconsistencies, demonstrating accurate identifications of R. barbirostris throughout its distribution. This validation confirms that our model is based on taxonomically accurate data and supported by physical material, which strengthens the reliability of our conclusions about the species’ distribution.
Based on the above, and even though the taxon R. barbirostris was originally described by Fabricius (1775) and its formal name, as we know it today, has been in use since 1815, when the genus was established. This means that the species has maintained its nomenclatural stability for more than 2 centuries. This consistency over time is a critical factor, as its identification does not present significant problems, thereby reducing uncertainty in historical and recent records. This stability is largely attributed to the meticulous work of specialists, who have generated a series of reliable and consistent records over time (Alonso-Zaragaza et al., 1999; Jones et al., 2008; Morrone, 2014; Morrone & Cuevas, 2002, 2009; Oberprieler et al., 2007; Rafinesque, 1815; Vaurie, 1970a, b).
Since its first record in Veracruz in the 1910´s, approximately 400 records of R. barbirostris have been compiled in Mexico from entomological collections and online databases. The oldest records show a widespread geographic distribution, with presence in multiple states, including Baja California, Chiapas, Jalisco, Oaxaca, Quintana Roo, Sinaloa, and Veracruz. In recent years, new records have focused on a subset of these states. In 2006, it was revealed that concentrated infestations in a smaller area, particularly in the 9 communities sampled in this study, corresponding to the Chochonteca region and the agricultural zones of the Tehuacán-Cuicatlán Biosphere Reserve in Oaxaca, where an average number of 20 individuals per palm infested by the bearded weevil was reported. This localized concentration suggests a possible local adaptation of R. barbirostris to the environmental conditions and to the specific populations of Braheadulcis in this region (Conafor 2020, 2023; Conanp, 2023). However, focused monitoring in these affected areas may have introduced sampling bias, potentially overlooking other regions where the species may also be present. The bearded palm weevil causes significant damage to its host plant, affecting the stipe and foliage, which can lead to its death. This damage impacts communities that depend on the B. dulcis palm for handicrafts and as a source of income (Pavón et al., 2006; Pérez-Valladares et al., 2020). As with the palm weevil Rhynchophoruspalmarum, which is a disease vector and causes bud rot, the difficulty in controlling these pests has led to similar phytosanitary strategies. In the case of R. palmarum, high populations hamper plantation renewal (Aldana-de la Torre et al., 2011; Giblin-Davis et al., 1997). In both cases, traditional management strategies have recommended drastic control measures such as cutting and burning infested plants to eliminate the source of infestation (Wattanapongsiri, 1966). The discovery of Zopheridae specimens in infested B. dulcis palms suggests their role as secondary organisms in the decomposition process. Unlike weevils, which are often primary invaders of healthy or slightly weakened plant tissue, Zopheridae are typically associated with decaying wood or degraded plant tissue. Based on this, it is plausible that the activity of the bearded weevil, by drilling into the trunk and weakening the tissue, creates the favorable conditions for Zopheridae to colonize the palms. Therefore, the presence of these beetles does not indicate a primary infestation, but rather an advanced state of host deterioration, which reinforces the severity of the damage caused by the bearded weevil (Cebeci et al., 2018; Pezzi et al., 2022).
Adult bearded weevils have been recorded on 15 species of palm trees: Attalea funifera, A. maripa, A. pindobossu, A. piassabossu, Bactris gasipaes, Brahea dulcis, Cocos botryophora, C. coronata, C. nucifera, Elaeis guineensis, Euterpe oleraceae, Mauritia flexuosa, Oenocarpa bacaba, Syagrus romanzoffiana, and S. schizophylla (Bukkens 1997; Choo 2009; Eberhard 1983). When comparing the number of records per host between different geographical sites corresponding to the distribution of R. barbirostris, it is notable that individuals from other geographical sites have a broader diet, compared to Mexican specimens (Choo 2009; Eberhard 1983; Vaurie 1970a).
Regarding other members of the Dryophthorinae subfamily, they are characterized by presenting phytophagy as a feeding habit (Oberprieler et al., 2007) and colonizing a wide variety of hosts. The subfamily of these insects, although notable for its wide distribution in diverse monocotyledons with reports of infestations in grasses, sedges, orchids, and bromeliads in different biomes (Anderson, 2002; Bautista-Gallardo et al., 2020; Oberprielier et al., 2007), has been predominantly documented in association with palms of the genera mentioned here (Morrone, 2009, 2014). Although their role as agriculturally important pests is globally recognized (De la Pava et al., 2020; Sepulveda-Cano & Rubio-Gómez, 2009), this specificity in host choice in the context of palms not only justifies but also strengthens the relevance of our study by focusing attention on 1 of the most significant interactions for this subfamily.
In summary, the analysis of the geographic and potential distribution of R. barbirostris in Mexico revealed the phenotypic plasticity of the species, which can be inferred from its ability to thrive in a variety of biomes, from humid montane forests to dry tropical forests. Although current records are limited to certain areas, potential distribution models suggest that the palm weevil finds favorable environmental conditions in a large part of the national territory. The exploration of all available records for this taxon has not only contributed to identifying sites with ideal conditions for its presence but also underscores the need for monitoring and verification sampling to confirm these predictions. These findings support and suggest the inclusion of the palm weevil in the Mexican Official Standard.
Acknowledgements
We thank Fernando Reyes Flores, Director of Reserva de la Biosfera Tehuacán-Cuicatlán, Comisión Nacional de Áreas Naturales Protegidas Dirección and the following institutions for funding this research: PAPIIT-UNAM (IN223924) and Secretaría de Ciencias, Humanidades, Tecnología e Innovación (SECIHTI). O.VM. is a postdoctoral fellowship from SECIHTI at Universidad Autónoma de Nuevo León, Campus Linares. Francisco Armendáriz-Toledano and Gerardo Cuéllar-Rodríguez are members of Sistema Nacional de Investigadores-SECIHTI. We also thank Mauricio Ramírez Hernández for the photographs of R. barbirostris.
References
Abraham, V. A., Al-Shuaibi, M. A., Faleiro, J. R., Abozuhairahand, R. A., & Vidyasagar, P. S. P. V. (1998). An integrated management approach for red palm weevil, Rhynchophorus ferrugineus Oliv., a key pest of date palm in the Middle East. Journal of Agricultural and Marine Sciences, 3, 77–83. https://doi.org/10.24200/jams.vol3iss1pp77-83
Aiello-Lammens, M. E., Boria, R. A., Radosavljevic, A., Vilela, B., & Anderson, R. P. (2015). spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography, 38, 541-545. https://doi.org/10.1111/ecog.01132
Aldana- De la Torre, R. C., Aldana-De la Torre, J. A., & Mauricio-Moya, O. (2011). Manejo del picudo Rhyncophorus palmarum L. Publicación del Centro de Investigación en palma de aceite Cenipalma, realizada con fondos provenientes del Convenio 20110059 suscrito entre el Ministerio de Agricultura y Desarrollo Rural y el Instituto Colombiano Agropecuario, ICA. http://hdl.handle.net/20.500.12324/19497
Alonso-Zaragaza, M. A., & Lyal, C. H. C. (1999). A world catalogue of families and genera of Curculionoidea (Insecta: Coleoptera) (excluding Scolytidae and Platypodidae). Entomopraxis. Barcelona.
Anderson, R. S. (2002). Chapter 131. Curculionidae. In R. H. Jr. Arnett, M. C. Thomas, Skelley P. E., & J. H. Frank (Eds.), American beetles. Volume II: Polyphaga: Scarabaeoidea through Curculionoidea (pp. 722–815) Boca Raton: CRC Press. https://doi.org/10.1201/9781420041231
Anderson, R. S. & Marvaldi, A. E. (2014). Dryophthorinae Schoenherr, 1825. In R. A. B. Les-chen, & R. G. Beutel (Eds.), Handbook of Zoology, Arthropoda: Insecta:Coleoptera Volume3: morphology and systematics (Phytophaga) (pp. 477–487). Berlin: De Gruyter.
Araujo, J. J., Keller, H. A., & Hilgert, N. I. (2018). Management of pindo palm (Syagrus romanzoffiana Arecaceae) in rearing of Coleoptera edible larvae by the Guarani of northeastern Argentina. Ethnobiology and Conservation, 7, 1–18. https://doi.org/10.15451/ec2018-01-7.01-1-18
Armendáriz-Toledano, F., López-Posadas, M. A., Utrera-Vélez, Y., Romero-Nápoles, J. R., & Castro-Valderrama, U. (2023). More than 80 years without new taxa: analysis of morphological variation among members of Mexican Aeneolamia Fennah (Hemiptera, Cercopidae) support a new species in the genus. Zookeys, 1139, 71–106. https://doi.org/10.3897/zookeys.1139.85270
Armendáriz-Toledano, F., Niño-Domínguez, A., Macías-Sámano, J., & Zúñiga, G. (2014b). Review of the geographical distribution of Dendroctonus vitei (Curculionidae: Scolytinae) based on geometric morphometrics of the seminal rod. Annals of the Entomological Society of America, 107, 748–75. https://doi.org/10.1603/AN13176
Armendáriz-Toledano, F., Niño-Domínguez, A., Sullivan, B. T., Macías-Sámano, J., Víctor, J., Clarke, S. et al. (2014a). Two species within Dendroctonus frontalis (Coleoptera: Curculionidae): evidence from morphological, karyological, molecular, and crossing studies. Annals of the Entomological Society of America,107, 11–27. https://doi.org/10.1603/AN13047
Boria, R. A., Olson, L. E., Goodman, S. M., & Anderson, R. P. (2014). Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecological Modelling, 275, 73–77. https://doi.org/10.1016/j.ecolmodel.2013.12.012
Bouchard, P. (2014). The book of beetles. A life size-guide to six hundreds of nature’s gems. Chicago: The University of Chicago Press.
Brown, J. H. (1984). On the relationship between abundance and distribution of species. The American Naturalist, 124, 255–279. https://doi.org/10.1086/284267
Bukkens, S. G. F. (1997). The nutritional value of edible insects. Ecology of Food and Nutrition, 36, 287–319. https://doi.org/10.1080/03670244.1997.9991521
Carretero, M. A., & Sillero, N. (2016). Evaluating how species niche modelling is affected by partial distributions with an empirical case. Acta Oecologica, 77, 207–216. https: //doi.org/10.1016/j.actao.2016.08.014
Castro-Valderrama, U., Carvalho, G. S., Peck, D. C., Valdez-Carrasco, J. M., & Romero, J. N. (2019). Two new species of the spittlebug genus Ocoaxo Fennah (Hemiptera: Cercopidae) from Mexico, and keys for the groups, group three, and first subgroup. Neotropical Entomology, 48, 260–268. https://doi.org/10.1007/s13744-018-0629-0
Castro-Valderrama, U., Romero-Nápoles, J., Peck, D. C., Valdez-Carrasco, J. M., Llanderal-Cázares, C., Bravo-Mojica, H. et al. (2017). First report of spittlebug species (Hemiptera: Cercopidae) associated with Pinus species (Pinaceae) in Mexico. Florida Entomologist, 100, 206–208. https://doi.org/10.1653/024.100.0136
Cebeci, H. H., & Baydemir, M. (2018). Predators of bark beetles (Coleoptera) in the Balikesir region of Turkey. Revista Colombiana de Entomología, 44, 283–287. https://doi.org/10.25100/socolen.v44i2.7326
Champion, G. C. (1909-1911). Insecta, Coleoptera, Rhynchophora, Curculionidae, Curculioninae, (concluded) and Calandrinae. In R. H. Porter (Eds.), Biologia Centrali-Americana. Vol. IV (pp. 1–213). London: The Thomas Lincoln Casey Library.
Choo, J., Zent, E. L., & Simpson, B. B. (2009). The importance of traditional ecological knowledge for palm-weevil cultivation in the Venezuelan Amazon. Journal of Ethnobiology, 29, 113–128. https://doi.org/10.2993/0278-0771-29.1.113
Cid-Muñoz, R., Cervantes-Espinoza, M., Castro-Valderrama, U., Cuéllar-Rodríguez, G., Cibrián-Tovar, D., & Armendáriz-Toledano, F. (2022). Pasado y presente de los salivazos del complejo Ocoaxo de los pinos (Hemiptera: Cercopidae): Distribución espacial y temporal. Revista Mexicana de Biodiversidad, 93, e934130. https://doi.org/10.22201/ib.20078706e.2022.93.4130
Conafor (Comisión Nacional Forestal). (2020). Diagnóstico fitosanitario del estado de Oaxaca, primer semestre, año 2020. Gerencia Estatal, Oaxaca.
Conafor (Comisión Nacional Forestal). (2023). Programa Operativo de Sanidad Forestal 2023 del Estado de Oaxaca. Comité de Sanidad Forestal, Estado de México, México.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2013). Programa de Manejo Reserva de la Biosfera Tehuacán-Cuicatlán. Secretaría de Medio Ambiente y Recursos Naturales (Semarnat). México D.F.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2015). Reporte de presencia de Rhinostomus barbirostris en núcleos agrarios dentro de las Reserva de la Biosfera Tehuacán Cuicatlán. México D.F.
De la Pava, N., García, M. A., Brochero, C. E., & Sepúlveda-Cano, P. (2019). Registros de Dryophthorinae (Coleoptera: Curculionidae) de la costa Caribe Colombiana. Acta Biológica Colombiana, 25, 96–103. https://doi.org/10.15446/abc.v25n1.77797
Dembilio, Ó., & Jacas, J. A. (2011). Life cycle of the invasive Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in Phoenix canariensis under Mediterranean climate. Bulletin of Entomological Research, 101, 153–163. https://doi.org/10.1017/S0007485310000283
Eberhard, W. G. (1983). Behavior of adult bottle brush weevils (Rhinostomus barbirostris) (Coleoptera: Curculionidae). Revista de Biología Tropical, 31, 233–244. https://doi.org/10.15517/rbt.v31i2.24986
Elith, J., Graham, C. H., Anderson, R. P., Dudík, M., Ferrier, S., Guisan, A. et al. (2006). Novel methods improve prediction of species distributions from occurrence data. Ecography, 29, 129–15. https://doi.org/10.1111/j.2006.0906-7590.04596
Elith, J., & Leathwick, J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics, 40, 677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159
Elith, J., Phillips, S. J., Hastie, T., Dudík, M., Chee, Y. E., & Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 17, 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725
Faleiro, J. R. (2006). A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. International Journal of Tropical Insect Science, 26, 135–154. https://doi.org/10.1079/IJT2006113
FAO. (2013). Edible insects. Future prospects for food and feed security. FAO. Rome.
Ferreira, J. M. S. (2008). Manejo integrado de pragas docoqueiro. Ciência Agrícola, 8, 21–29.
Ferreira, J. M. S., Fontes, R. H., & Procopio, S. O. (2007). Resinose do coqueiro: como identificar e manejar. Aracaju: Embrapa Tabuleiros Costeiros.
Fiaboe, K. K. M., Peterson, A. T., Kairo, M. T. K., & Roda, A. L. (2012). Predicting the potential worldwide distribution of the red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) using ecological niche modeling. Florida Entomologist, 95, 559–673. https://doi.org/10.1653/024.095.0317
Fick, S. E. & Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37, 4302–4315. https://doi.org/10.1002/joc.5086
Fourcade, Y., Engler, J. O., Rödder, D., & Secondi, J. (2014). Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. Plos One, 9, e97122. https://doi.org/10.1371/journal.pone.0097122
Franco, E. (1964). Estudo sobre o Anel Vermelho do Coqueiro. Departamento de Defesa e Inspeção Agropecuária. Sergipe: Serviço de defesa sanitária vegetal. Inspetoria de defesa sanitária vegetal.
Franqui-Rivera, R., & Medina-Gaud, S. (2003). Identificación de insectos de posible introducción a Puerto Rico. San Juan: Departamento de Agricultura de Puerto Rico/ Universidad de Puerto Rico.
Gardner, J. C. M. (1934). Immature stages of Indian Coleoptera (14) (Curculionidae). Indian Forest Records, 20, 1–48.
Gardner. J. C. M. (1938). Immature stages of Indian Coleoptera (24 Curculionidae Contd.). Indian Forest Records, Entomology Series, 3, 227–261.
Giblin-Davis, R. M., Gries, R., Gries, G., Peña-Rojas, E., Pinzon, I., Peña, J. E. et al. (1997). Aggregation pheromone of the palm weevil, Dynamis borassi (F.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 23, 2287–2297. https://doi.org/10.1023/B:JOEC.0000006674.64858.f2
Goodey, T. (1933). Plant parasitic nematodes and the diseases they cause, with a foreword by R.T. Leiper. London: E.P. Dutton and Co.
Goodey, J. B. (1960). Rhadinaphelenchus cocophilus (Cobb 1919) n. comb, the nematode associated with ‘red ring’ disease of coconuts. Nematologica, 5, 98–102.
Google (2020). Google Earth Sofware libre Versión 9.140.0. http://earth.google.com/
Grebennikov, V. V. (2018a). Dryophthorinae weevils (Coleoptera: Curculionidae) of the forest floor in Southeast Asia: illustrated overview of nominal Stromboscerini genera. Zootaxa, 4418, 121–135. https://doi.org/10.11646/zootaxa.4418.2.2
Grebennikov, V. V. (2018b). Dryophthorinae weevils (Coleoptera: Curculionidae) of the forest floor in Southeast Asia: DNA analysis of two new Nephius from Vietnam and Taiwan suggests non monophyly of Stromboscerini. Zootaxa, 4500, 381–387. https://doi.org/10.11646/zootaxa.4500.3.5
Grebennikov, V. V. (2018c). Dryophthorinae weevils (Coleop- tera: Curculionidae) of the forest floor in Southeast Asia: three-marker analysis reveals monophyly of Asian Strombocerini and new identify of rediscovered Tasactes. European Journal of Entomology, 115, 437–444. https://doi.org/10.14411/eje.2018.044
Griffith, R. (1987). Red ring disease of coconut palm. Plant Disease, 71, 193–196.
Guisan, A., & Zimmermann, N. E. (2000). Predictive habitat distribution models in ecology. Ecological Modelling, 135, 147–186. https://doi.org/10.1016/S0304-3800(00)00354-9
Howard F. W., Moore D., Giblin-Davis R. M., & Abad, R. G. (2001). Insects on palms. New York: CABI Publishing.
Jaramillo-Vivanco, T., Cámara, R. M., Cámara, M., Tejera, E., Balslev, H., & Álvarez-Suárez, J. M. (2023). Ethnobiology of edible palm weevil larvae Rhynchophorus palmarum L. (Curculionidae, Coleoptera), a common food source in Amazonian Ecuador. Journal of Insects as Food and Feed, 10, 427–441. https://doi.org/10.1163/23524588-20230135
Jones, R. W., O’Brien, C. W., Ruiz-Montoya, L., & Gómez-Gómez, B. (2008). Insect diversity of tropical montane forests: diversity and spatial distribution of weevils (Coleoptera: Curculionidae) inhabiting leaf litter in Southern Mexico. Entomological Society of America, 101, 128–139. https://doi.org/10.1603/0013-8746(2008)101 [128:IDOTMF]2.0.CO;2
Kass, J. M., Pinilla-Buitrago, G. E., Paz, A., Johnson, B. A., Grisales-Betancur, V., Meenan, S. I. et al. (2023). Software note. Wallace 2: a shiny app for modeling species and distributions redesigned to facilitate expansion via module contributions. Ecography, 2023, e06547. https://doi.org/10.1111/ecog.06547
Korkmaz, S., Göksülük, D., & Zararsiz, G. (2014). MVN: an R package for assessing multivariate normality. The R Journal, 6, 151–162. https://doi.org/10.32614/RJ-2014-031
Kuschel, G. (1995). A phylogenetic classification of Curculionoidea to families and subfamilies. Memoirs of the Entomological Society of Washington, 14, 5–33.
Lacerda-Moura, J. I., Lopes de Oliveira, M., Martins-Newman Luz, E. D., Midlej-Silva, S. D., & Souza-Santos, R. (2013). Weevil associated to gummosis in oil palm (Elaeis guianeensis Jacq.) in Brazil. Revista de Agricultura, 88, 87–89.
Lanteri, A., Marvaldi, A., & Suárez, S. (2002). Gorgojos de la Argentina y sus plantas huéspedes. Tomo I: Apionidae y Curculionidae. La Plata: Sociedad Entomológica Argentina.
Lizardo, V., Ruggiero, A., & Morrone, J. J. (2025). Patrones de riqueza de especies y conservadurismo filogenético del nicho ecológico en la Zona de Transición Mexicana: evidencia y herramientas para su estudio. Revista Mexicana Biodiversidad, 96, e965347. https://doi.org/10.22201/ib.20078706e.2025.96.5347
López-Serrano, P. M., Hernández-Ramos, A., Méndez-González, J., Martínez-Salvador, M., Aguirre-Calderón, O., Vargas-Larreta, B. et al. (2021). Mejores prácticas de manejo y ecuaciones alométricas de biomasa de Brahea dulcis en el estado de Oaxaca y Guerrero. Proyecto: 2017-4-292674. México D.F.: Conafor/ Conacyt.
López-Zent, E., & Zent, S. (2004). Amazonian Indians as ecological disturbance agents: the Hotï of the Sierra de Maigualida, Venezuelan Guayana. Advances in Economic Botany, 15, 79–112.
Luna-Cozar, J., Jones, R. W., & Niño-Maldonado, S. (2024). Picudos (superfamilia Curculionoidea). In Conabio (Eds.), La biodiversidad en Tamaulipas. Estudio de estado. Vol. II (pp. 309–317).Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Marvaldi, A. E., & Morrone, J. J. (2000). Phylogenetic systematics of weevils (Coleoptera: Curculionoidea): a reappraisal based on larval and adult morphology. Insect Systematics and Evolution, 31, 43–58. https://doi.org/10.1163/187631200X00309
McKenna, D. D., Sequeira, A. S., Marvaldi, A. E., & Farrell, B. D. (2009). Temporal lags and overlap in the diversification of weevils and flowering plants. Proceedings of the National Academy of Sciences of the United States of America, 106, 7083–7088. https://doi.org/10.1073/pnas.081061810
McKenna, D. D., Wild, A. L., Kanda, K., Bellamy, C. L., Beutel, R. G., Caterino, M. S. et al. (2015). The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Systematic Entomology, 40, 835–880. https://doi.org/10.1111/syen.12132
Merow, C., Smith, M. J., & Silander Jr., J. A. (2013). A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography, 36, 1058–1069. 10.1111/j.1600-0587.2013.07872.x
Morrone, J. J. (2014). Biodiversidad de Curculionoidea (Coleoptera) en México. Revista Mexicana de Biodiversidad, 85 (Suppl.) 312–324. https://doi.org/10.7550/rmb.30038
Morrone, J. J. (2019). Regionalización biogeográfica y evolución biótica de México: encrucijada de la biodiversidad del Nuevo Mundo. Revista Mexicana de Biodiversidad, 90, e902980. https://doi.org/10.22201/ib.20078706e.2019.90.2980
Morrone, J. J., & Cuevas, P. I. (2002). Cladistics of the pantropical genus Rhinostomus (Coleoptera: Curculionidae: Dryophthorinae), with nomenclatural notes. Journal of the New York Entomological Society, 10, 76–388. https://doi.org/10.1664/0028-7199(2002)110[376:COTPGR]2.0.CO;2
Morrone, J. J., & Cuevas, P. I. (2009). On the status of the tribes Orthognathini and Rhinostomini (Coleoptera: Curculionidae: Dryophthorinae). Zootaxa, 2216, 57–63. https://doi.org/10.11646/zootaxa.2216.1.5
Oberprieler, R. G., Marvaldi, A. E., & Anderson, R. S. (2007). Weevils, weevils, weevils everywhere. Zootaxa, 1668, 491–520. https://doi.org/10.5281/zenodo.274039
Pardo-Locardo, L. C., Constantino, L. M., Agudelo, R., Alarcón, A., & Caicedo, V. (2005). Observaciones sobre el gualapán (Coleoptera: Chrysomelidae: Hispinae) y otras limitantes entomológicas en cultivos de chontaduro en el Bajo Anchicayá. Acta Agronómica, 54, 25–30.
Pardo-Locardo, L. C., Constantino, L. M., & Bustillo-Pardey, A. (2019). Las plagas más importantes del cultivo de palma Chontaduro Bactris gasipes en la costa Pacífica Colombiana. Gobernación Valle del Cauca. Memoria Técnica Convenio 1340-202.
Pavón, N. P., Escobar, R., & Ortiz-Pulido, R. (2006). Extracción de hojas de la palma Brahea dulcis en una comunidad otomí en Hidalgo, México: efecto sobre algunos parámetros poblacionales. Interciencia, 31, 57–61.
Pearman, P. B., D’Amen, M., Graham, C. H., Thuiller, W., & Zimmermann, N. E. (2010). Within-taxon niches structure: niche conservatism, divergence and predicted effects of climate change. Ecography, 33, 990–1003. https://doi.org/10.1111/j.1600-0587.2010.06443.x
Pérez-Irineo, G., & Mandujano-Rodríguez, S. (2020). Reserva de la Biosfera Tehuacán-Cuicatlán: un lugar increíble para la conservación de mamíferos. Ciencia y Mar, 24, 85–90.
Pérez-Valladares, C. X., Moreno-Calles, A. I., Casas, A., Rangel-Landa, S., Blancas, J., Caballero, J. et al. (2020). Ecological, cultural, and geographical implications of Brahea dulcis (Kunth) Mart. insights for sustainable management in Mexico. Sustainability, 12, 412. https://doi.org/10.3390/su12010412
Pezzi, M., Carlomagno, F., Mendicino, F., Bonelli, D., Pelle, R., Leis, M. et al. (2022). Pycnomerus italicus (Coleoptera: Zopheridae), an endemic endangered species: A new report on its presence in southern Italy. Forests, 13, 1838. https://doi.org/10.3390/f13111838
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E., & Blair, M. E. (2017). Opening the black box: an open-source release of Maxent. Ecography, 40, 887–893. https://doi.org/10.1111/ecog.03049
Phillips, S. J., Anderson, R. P., & Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190, 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
QGIS.org. (2024). QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://qgis.org
Quero, H. J. (1994). Flora del Valle de Tehuacán-Cuicatlán. Jardín Botánico. Instituto de Biología, UNAM. México D.F.
Rafinesque, C. S. (1815). Analyse de la nature, ou, Tableau de l’univers et des corps organisés. Instituto de Ciencias Naturales de Nápoles. Palermo.
Rangel-Landa, S., Rivera-Lozoya, E., & Casas, A. (2014). Uso y manejo de las palmas Brahea spp. (Arecaceae) por el pueblo ixteco de Santa María Ixcatlán Oaxaca, Mexico. Gaia Scientia, Volume Especial PopulaçõesTradicionais, 62–78.
Rugman-Jones, P. F., Hoddle, C. D, Hoddle, M. S., & Stouthamer, R. (2013). The lesser of two weevils: molecular-genetics of pest palm weevil populations confirm Rhynchophorus vulneratus (Panzer, 1798) as a valid species distinct from R. ferrugineus (Olivier, 1790) and reveal the global extent of both. Plos One, 8, e78379. https://doi.org/0.1371/journal.pone.0078379
Rzedowski, J. (2006). Vegetación de México. Edición digital. Conabio, Mexico City. https://www.biodiversidad.gob.mx/publicaciones/librosDig/pdf/VegetacionMxPort.pdf
Salas, J. A. (1980). El anillo rojo del cocotero en varias áreas afectadas de Costa Rica. Agronomía Costarricense, 4, 199–202.
Sarwar, M. (2016). Biological control of red palm weevil Rhynchophorusferrugineus (Coleoptera: Curculionidae) by the natural enemies. International Journal for Research in Biology & Pharmacy, 2, 22–35. https://doi.org/10.53555/bp.v2i7.131
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (1997). Norma Oficial Mexicana NOM-006-SEMARNAT-1997, que establece los procedimientos, criterios y especificaciones para realizar el aprovechamiento de hojas de palma. Diario Oficial de la Federación.
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (2002). Norma Oficial Mexicana NOM-059-SEMARNAT-2001, Protección ambiental-especies nativas de México de flora y fauna silvestre-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-lista de especies en riesgo. Diario Oficial de la Federación.
Sepúlveda-Canon, P. A., & Rubio-Gómez, J. D. (2009). Dryophthorinae species (Coleoptera: Curculionidae) associated to plantain and banana crops (Musa spp.) in Colombia. Acta Biológica Colombiana, 14, 49–72.
Shcheglovitova, M., & Anderson, R. P. (2013). Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecological Modelling, 269, 9–17. https://doi.org/10.1016/j.ecolmodel.2013.08.011
Sillero, N. (2011). What does ecological modelling model? a proposed classification of ecological niche models based on their underlying methods. Ecological Modelling, 222, 1343–1346. https://doi.org/10.1016/j.ecolmodel.2011.01.018
SINAT (Sistema Nacional de Información Ambiental y Recursos Naturales), & Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (2009). Informe preventivo de impacto ambiental para el aprovechamiento forestal no maderable “Hoja de Palma” “Brahea dulcis” y “Cryosophylla nana”. Ejido Luis Echeverría Álvarez. Municipio de Coquimatlan, Colima. SINAT-Semarnat.
Sinco-Ramos, P. G., García-Frapolli, E., Durand, L., & Porter-Bolland, L. (2021). Visiones de bienestar en la Reserva de la Biosfera Tehuacán-Cuicatlán, México. Región y Sociedad, 33, e1527. https://doi.org/10.22198/rys2021/33/1527
Sofaer, H. R., Hoeting, J. A., & Jarnevich, C. S. (2019). The area under the precision-recall curve as a performance metric for rare binary events. Methods in Ecology and Evolution, 10, 565–577. https://doi.org/10.1111/2041-210X.13140
Soroker, V., Harari, A., & Faleiro, J. R. (2015). The role of semiochemicals in date pest management. In W. Wakil, J. Romeno-Faleiro, & T. A. Miller (Eds.), Sustainable pest management in date palm: current status and emerging challenges (pp. 315–346). Cham, Switzerland: Springer International Publishing. https://doi.org/10.1007/ 978-3-319-24397-9_11
Valavi, R., Guillera-Arroita, G., Lahoz-Monfort, J. J., & Elith, J. (2022). Predictive performance of presence-only species distribution models: a benchmark study with reproducible code. Ecological Monographs, 92, e01486. https://doi.org/10.1002/ecm.1486
Valerio-Mendoza, O., Armendáriz-Toledano, F., Cuellar-Rodríguez, G., Negrón, J. F., & Zúñiga, G. (2019). Cryptic species discrimination in Western Pine Beetle, Dendroc- tonus brevicomis LeConte (Curculionidae: Scolytinae), based on morphological characters and geometric morphometrics. Insects, 10, 377. 10.3390/insects10110377
Van Huis, A., Van Itterbeeck, J. V., Klunder, H., Mertens, E., & Halloran, A. (2013). Edible insects: future prospects for food and feed security. Roma: FAO Forestry.
Van Itterbeeck, J., & Van Huis, A. (2012). Environmental manipulation for edible insect procurement: a historical perspective. Journal of Ethnobiology and Ethnomedicine, 8, 1–7. https://doi.org/10.1186/1746-4269-8-3
Vargas, G. E., Espinoza, G., Ruiz, C., & Rojas, R. (2013). Valor nutricional de la larva de Rhyncophoruspalmarum L.: Comida tradicional en la Amazonia Peruana. Revista de la Sociedad Química del Perú, 79, 64–70.
Vaurie, P. (1970a). Weevils of the tribe Sipalini (Coleoptera, Curculionidae, Rhynchophorinae). Part 1, the genera Rhinostomus and Yuccaborus. American Museum Novitates, 2419, 1–57.
Vaurie P. 1(970b). Weevils of the tribe Sipalini (Coleoptera, Curculionidae, Rhynchophorinae). Part 2, The genera Mesocordylus and Orthognathus. American Museum Novitates, 2441, 1–78.
Vaurie, P. (1971). Review of Scyphophorus (Curculionidae: Rhynchophorinae). Coleopterists Bulletin, 25, 1–8. https://doi.org/10.5962/p.372515
Vergara-Navarro, E. V., Montenegro-Silva, L. M., & Serna, F. (2021). Catálogo de Coleoptera de la Colección Taxonómica Nacional de Insectos Luis María Murillo (CINI), Cundinamarca, Colombia. Bogotá: Agrosavia editorial.
Warren, D. L. (2012). In defense of ‘niche modeling’. Trends in Ecology & Evolution, 27, 497–500. https://doi.org/10.1016/j.tree.2012.03.010
Wattanapongsiri, A. A. (1966). Revision of the genera Rhynchophorus and Dynamis (Coleoptera: Curculionidae) (Ph.D. Thesis). Oregon University State, USA.
Wesissling, T. J., & Gibling-Davis, R. M. (1993). Water loss dynamics and humidity preference of Rhyncophorus cruentatus (Coleoptera: Curculionidae) adults. Environ- mental Entomology, 22, 93-98. https://doi.org/10.1093/ee/22.1.93
Zaragoza-Caballero, S., Navarrete-Heredia, J. L., & Ramírez-García, E. (2015). Temolines.Los coleópteros entre los antiguos mexicanos. México D.F.: Universidad Nacional Autónoma de México.
Zimmerman, E. C. (1968a). The cosmopolites banana weevils (Coleoptera: Curculionidae: Rhynchophorinae). Pacific Insects, 10, 295–299.
Zimmerman, E. C. (1968b). Rhynchophorinae of southeastern Polynesia. Pacific Insects, 1, 47–77.
Zimmerman, E. C. (1993). Australian weevils, vol. III. Nanophyidae, Rhynchophoridae, Erirhinidae, Curculionidae: Amycterinae, literature consulted. Melbourne: CSIRO.
Forest management and bark-beetle control in Santa Catarina Lachatao, Oaxaca
Tania Alhelí Cruz-Mejía y Ek del-Val*
Universidad Nacional Autónoma de México, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Antigua Carretera a Pátzcuaro Núm. 8701, Col. Ex-Hacienda de San José de La Huerta, 58190 Morelia, Michoacán, México
*Autor para correspondencia: ekdelval@iies.unam.mx (E. del-Val)
Recibido: 09 noviembre 2024; aceptado: 09 abril 2025
Resumen
El cambio climático ha favorecido el incremento de las poblaciones de escarabajos descortezadores del género Dendroctonus en los bosques de pino debido al aumento de temperaturas e incidencia de sequías. En México, estos brotes han causado la muerte de millones de pinos, afectando la economía forestal y los ecosistemas silvestres. Algunas comunidades mitigan estos impactos mediante sus esquemas de manejo forestal. En este estudio, con un enfoque participativo, evaluamos la presencia de escarabajos descortezadores (Coleoptera: Curculionidae: Scolytinae) en el bosque de Santa Catarina Lachatao, Oaxaca, con el objetivo de describir su diversidad y los factores ambientales que la determinan, así como el estado fitosanitario del bosque. Además, se evaluó el contexto político y social en torno al manejo forestal. Se reporta una amplia diversidad de escarabajos del género Dendroctonus representada por 6 especies (D. adjunctus, D. frontalis, D. mexicanus, D. approximatus, D. parallelocollis y D. valens). Los resultados indican que el bosque de Santa Catarina Lachatao se encuentra en buenas condiciones fitosanitarias gracias al manejo forestal implementado, a pesar de los brotes de Dendroctonus. La participación activa de la comunidad en la investigación subraya la importancia de la gestión forestal comunitaria para conservar un bosque sano y resiliente.
Climate change has favored the increase of bark beetle populations of the genus Dendroctonus in pine forests due to rising temperatures and droughts. In Mexico, these outbreaks have caused the death of millions of pine trees, affecting both the forestry economy and natural ecosystems. Some communities mitigate these impacts through their forest management schemes. In this study, using a participatory approach, we assessed the presence of bark beetles (Coleoptera: Curculionidae: Scolytinae) in the forest of Santa Catarina Lachatao, Oaxaca, with the aim of describing their diversity and the environmental factors that determine it, as well as the forest’s phytosanitary condition. Additionally, we evaluated the political and social context surrounding forest management. We report a wide diversity of Dendroctonus species, represented by 6 species (D.adjunctus, D. frontalis, D. mexicanus, D. approximatus, D. parallelocollis, and D. valens). The results indicate that the forest of Santa Catarina Lachatao is in good phytosanitary condition thanks to the implemented forest management, despite the presence of Dendroctonus outbreaks. The active participation of the community in the research highlights the importance of community-based forest management for maintaining healthy and resilient forests.
Keywords: Temperate forests; Dendroctonus; Local management; Pinus; Scolytinae
Introducción
Los bosques templados tienen una gran importancia ecológica y económica debido a los servicios que proveen a las personas, como protección de suelos, regulación y provisión hídrica, mitigación de daños por fenómenos hidrometeorológicos extremos, hábitat de vida silvestre y conectividad biológica, preservación de áreas para recreación, valores rituales y del paisaje (Challenger y Soberón, 2008; Díaz et al., 2015). Asimismo, son reservorios de carbono por lo que son fundamentales en la mitigación del cambio climático (Challenger y Soberón, 2008; Chapela y Merino, 2019).
México es el centro de origen y diversificación del género Pinus, cuantificando 50% del total de especies conocidas del género (Challenger, 2003; Challenger y Soberón, 2008; Koleff et al., 2004). Los bosques mexicanos son relevantes en términos mundiales por la alta biodiversidad que albergan, así como por la experiencia de manejo forestal mediante esquemas comunitarios (Chapela y Merino, 2019).
En México los bosques de pino son los de mayor distribución entre los distintos tipos de bosques de coníferas en el país; cubren hoy alrededor de 75% de su distribución potencial, estimada en poco más de 10 millones de hectáreas, aunque los bosques bien preservados solo abarcan 5.2 millones de hectáreas en el país (INEGI, 2005; INEGI-Conabio-INE, 2007).
El cambio climático ha incidido sobre algunas poblaciones de animales y les ha permitido aumentar su intervalo de distribución latitudinal y altitudinal en los bosques templados (Bale et al., 2002; Gitai et al., 2002; Jepsen et al., 2008). En particular, las sequías y altas temperaturas que se han experimentado en los últimos años han favorecido a las poblaciones de las especies del género Dendroctonus especialistas del género Pinus (Cudmore et al., 2010; FAO, 2010; Marini et al., 2012; Safrayik y Wilson, 2006). Esto tiene mayor relevancia, ya que dentro de este género se encuentran especies con comportamiento agresivo, capaces de matar árboles sanos a diferentes escalas ecológicas, y por lo tanto considerarse plagas (Armendáriz-Toledano et al., 2018; Castellanos-Bolaños et al., 2013; Raffa et al., 2015).
Se han documentado incrementos poblacionales en varias especies agresivas de Dendroctonus,así como un aumento en el número de generaciones que pueden tener por año (Biedermann et al., 2019; Hart et al., 2014; Ning et al., 2021). Específicamente, en Norteamérica se están experimentando daños severos por incremento de la frecuencia de infestaciones en lugares donde estos escarabajos eran escasos y en áreas fuera de su distribución nativa (Creeden et al., 2014; Weed et al., 2013). En México, se han reportado casos en los que brotes recientes de escarabajos descortezadores han ocasionado importantes pérdidas en los bosques de coníferas (Conafor, 2014).
Aunque los pinos han interactuado históricamente con los escarabajos descortezadores, las disrupciones climáticas recurrentes han afectado su salud. El aumento de temperatura induce estrés fisiológico, debilitando sus defensas y favoreciendo los brotes de algunas especies de descortezadores (Boyd et al., 2013; Dukes et al., 2009; Weed et al., 2013). Cuando los árboles están debilitados, producen menos resina o dejan de hacerlo, al reducirse en los ductos del xilema, los insectos acceden al floema con facilidad, provocando la muerte de los pinos (Dukes et al., 2009; Gaylord et al., 2015; Raffa et al., 2008). Sin embargo, es evidente que aún se requieren investigaciones sobre la resiliencia de los ecosistemas ante el cambio climático, que permitan realizar predicciones más precisas. El aumento de brotes de Dendroctonus en México también ha impactado al estado de Oaxaca. En el periodo de 2004 a 2009 brotes de las especies primarias D. adjuntus, D. mexicanus y D. frontalis afectaron alrededor de 6,300 ha de bosque, cuantificando un volumen aproximado de madera de 957,400 m3 (Castellanos-Bolaños et al., 2013). Aunado a ésto, la precipitación anual e índice de infiltración disminuyeron considerablemente en los bosques de Oaxaca como consecuencia del cambio climático, lo que ocasionó incendios forestales con mayor frecuencia, disminución de la cubierta forestal y proliferación del escarabajo descortezador (Rivera-Jiménez et al., 2019).
Oaxaca es considerado como un centro de diversificación de pinos en el país, donde se reportan 14 especies de Pinus (Del-Castillo et al., 2004) y 7 especies de escarabajos del género Dendroctonus (Salinas-Moreno et al., 2010). Dadas estas características y aunado a las recientes sequías recurrentes, se ha considerado que varias regiones del estado presentan un alto riesgo de brotes de escarabajos descortezadores lo que significa una amenaza potencial para sus bosques (Cibrian et al., 1995; Durán y Poloni, 2014; Farjon, 1996; Salinas-Moreno et al., 2010). Aunque el impacto de estos escarabajos represente un gran riesgo para las zonas forestales, no se recomienda eliminarlos del bosque, pero es necesario mantener bajo control su potencial de contagio. A pesar de la información sobre el control de Dendroctonus y los avances tecnológicos para monitorear las plagas, la vía más exitosa para su control sigue siendo la remoción mecánica de los árboles infestados (Castellanos-Bolaños et al., 2013; Cibrián-Tovar et al., 1995, 2015; Durán y Polini, 2014).
En este contexto, el manejo forestal comunitario resulta fundamental para enfrentar dichos desafíos (Cibrián-Tovar y Macías-Sámano, 2021). En este estudio, este tipo de manejo forestal se entiende como un conjunto integral de prácticas comunitarias, basadas en conocimientos tradicionales y adaptadas a las condiciones ecológicas y culturales locales. Dentro de este enfoque, la recolección de leña, el ecoturismo y los saneamientos forestales se incorporan como componentes de un mosaico de actividades, lo que permite una gestión flexible y adaptativa del ecosistema forestal. Asimismo, el manejo forestal comunitario integra la conservación, el aprovechamiento sostenible y el uso diversificado de los recursos, facilitando la resiliencia y el mantenimiento de los bosques (Toledo, 2013; Toledo y Barrera-Bassols, 2008).
El 85% de los bosques de Oaxaca está en tierras de propiedad social, por ello, el saneamiento forestal y el manejo deben considerar aspectos socioculturales e institucionales, así como a las comunidades y actores involucrados (Bray, 2013). En el estado, los esquemas de gobernanza local sólidos han permitido experiencias exitosas de manejo, aprovechamiento maderero y conservación (Bray et al., 2012; Gasca-Zamora, 2014). En Oaxaca los sistemas de usos y costumbres regulan en gran medida las acciones de gobernabilidad y participación ciudadana a nivel local; es decir, las comunidades asumen su autonomía para ejercer el control de sus procesos de desarrollo y gestión de los recursos naturales (Bray et al., 2012; Mathus-Alonso et al., 2010). En las comunidades y ejidos forestales se ha optado por el control de plagas forestales bajo esquemas de gobernanza de múltiple escala, lo que implica la confluencia de distintos actores interesados en resolver dicha problemática. Los actores externos (local, gubernamental, académico, conservacionista) suman esfuerzos con las comunidades y ejidos forestales para aportar a través de capacitaciones, investigaciones, difusión de información y otras actividades que, en conjunto con el manejo forestal comunitario, fortalecen el conocimiento y las prácticas dirigidas a mantener bosques saludables (Durán y Polini, 2014; Gasca-Zamora, 2014; Mathus-Alonso et al., 2010).
Durante el periodo de 2004 a 2009 se registraron afectaciones importantes por ataques de escarabajos del género Dendroctonus en 98 comunidades. La región de Pueblos Mancomunados, conformada por los municipios de Santa Catarina Lachatao, Amatlán y Yavesía fue la más afectada. (Castellanos-Bolaños et al., 2013). Dado que los datos disponibles en la literatura científica y en los informes institucionales oficiales son muy generales (Pacheco-Aquino y Durán, 2023), se seleccionó a la comunidad de Santa Catarina Lachatao como estudio de caso para este proyecto. Esta comunidad manifestó un interés activo por disponer de información detallada que contribuyera a mejorar el manejo forestal y el control de plagas. En consecuencia, el presente proyecto se diseñó desde un enfoque participativo en el que la comunidad se involucró en el desarrollo de la investigación. El objetivo fue documentar y evaluar la eficiencia del manejo forestal comunitario para el control del escarabajo descortezador en Santa Catarina Lachatao en parcelas de bosque bajo diferente manejo, así como la cuantificación de la abundancia de Dendroctonus frontalis, D. mexicanus y D. adjunctus.
Materiales y métodos
Santa Catarina Lachatao (SCL) se encuentra al este del estado de Oaxaca entre las coordenadas 17°05’-17°17’ N, 96°27’-96°33’ O, en un gradiente altitudinal entre 1,800 y 3,200 m snm. La temperatura media promedio oscila entre 10 y 20 °C, con un rango de precipitación total anual de 800 a 1,200 mm. El clima es templado subhúmedo con lluvias en verano (95.84%), semicálido subhúmedo con lluvias en verano (2.31%) y templado húmedo con abundantes lluvias en verano (1.85%) (Fernández et al., 2013; INEGI, 2010). La cubierta de la comunidad es bosque templado, el cual se compone de asociaciones mixtas de pino-encino y encino-pino. La tenencia de la tierra es comunal y se rigen por el sistema de gobierno por usos y costumbres (Fernández et al., 2013; INEGI, 2010).
SCL pertenecía a Pueblos Mancomunados, que se estableció en 1957, junto con los municipios de Amatlán y Yavesía, las agencias municipales de Latuvi, Benito Juárez, Cuajimoloyas, y las agencias de policía de La Nevería y Llano Grande, todos forman parte del distrito de Ixtlán de Juárez. La superficie del Mancomún era de 29,430.86 ha y los comuneros de las 8 poblaciones eran poseedores de los recursos naturales dentro del área. Sin embargo, cada comunidad tenía un área determinada de la que tenía que hacerse responsable (DOF, 1961; Rojas-Serrano y Martínez-Corona, 2017).
Pueblos Mancomunados era considerado una comunidad indivisible; sin embargo, algunos municipios comenzaron a manifestar su desacuerdo con esta figura colectiva, debido a diferencias en la gestión y el manejo de los recursos naturales. Por ello, en 1961 se les reconocen los bienes comunales a Santa Catarina Lachatao, Amatlán y Yavesía, ésto ha derivado en tensiones y conflictos intracomunitarios recurrentes (DOF, 1961). Actualmente, se encuentran delimitando el territorio que correspondería a cada municipio, agencias municipales y agencias de policía.
En la comunidad de SCL, la tierra y los recursos naturales son propiedad comunal, lo que significa que todos tienen derecho de aprovecharlos, pero también la responsabilidad de conservarlos. Asimismo, tienen la obligación y el derecho de participar en los sistemas de cargos para la organización política y social de la comunidad, con el fin de decidir y contribuir en el destino de ésta. El principal órgano de toma de decisiones es la Asamblea Comunitaria, en la que participan todos los miembros de la comunidad (a partir de los 16 años), en ésta se distribuyen las tareas entre las personas de la comunidad y se asignan a las personas que ocuparán cargos comunales y cargos de autoridades (Bello-Cervantes, 2015; Rojas-Serrano y Martínez-Corona, 2017).
Con la finalidad de conocer el manejo forestal en la comunidad de SCL, así como las medidas para el control de las poblaciones de las especies de escarabajo descortezadores reportadas en la región, se realizaron 15 entrevistas semiestructuradas entre marzo y julio de 2019. Entre las personas entrevistadas se encontraba el encargado comunal del bosque, representantes comunales vigentes durante el periodo de muestreo, autoridades anteriores, guías turísticos y personas que realizaban alguna actividad en el bosque.
Los sitios para evaluar la presencia y abundancia de escarabajos descortezadores se eligieron en conjunto con la comunidad, seleccionando los sitios en los que los habitantes habían identificado brotes de descortezadores atípicos. Se seleccionaron 6 sitios separados entre sí por lo menos por 1,500 m y en un gradiente altitudinal entre 1,937 y 2,755 m snm (tabla 1).
Para el muestreo de las comunidades de escarabajos descortezadores, se siguió la metodología empleada por Rubín-Aguirre et al. (2015). En cada uno de los sitios seleccionados se colocaron trampas Lindgren de 12 unidades sobre una estaca de metal y en claros en donde el pino más cercano estuviera, mínimo, a 10 metros de distancia de la trampa, para evitar la invasión a pinos cercanos. En cada trampa se agregó frontalina (feromona) + alfa/beta-pineno (kairomona) y endo-brevicomina como atrayentes de escarabajos descortezadores, con especial énfasis en capturar especies agresivas de descortezadores (Macías-Sámano y Niño-Domínguez, 2016). En la base de cada trampa se colocó un contenedor plástico con anticongelante diluido al 50% con agua, con la finalidad de preservar los insectos capturados. Los insectos se recolectaron en conjunto con la comunidad, durante la época de secas cada 3 semanas por 5 meses, de marzo a julio de 2019, se preservaron en alcohol al 70% y se identificaron en el laboratorio con las claves de la Guía ilustrada para identificar a las especies del género Dendroctonuspresentes en México y Centroamérica (Armendáriz-Toledano et al., 2018) y The bark Ambrosia beetles of North and Central America (Coleoptera: Scolytidae): a taxonomic monograph (Wood, 1982).
En cada sitio de muestreo, se evaluó el estado fitosanitario de las especies de pino utilizando la metodología descrita por del Río-Mora (2011), la cual se fundamenta en los indicadores sugeridos por Innes (1993) y Berryman (1986). El diseño de muestreo consistió en marcar 2 parcelas circulares de 25 m2 cada una a 25 m de distancia de la trampa en cada sitio. Se optó por este diseño para maximizar la relevancia práctica, dada la realización del muestreo en conjunto con la comunidad.
Dentro de cada parcela se contaron e identificaron a nivel especie los individuos de pino presentes, el diámetro a la altura del pecho (DAP) y se evaluó el vigor de los pinos. Este último se calculó en una escala del 1 al 6, en donde el valor 1 significa que el árbol está en perfecto estado y el 6 es indicador de un árbol muerto. En el contexto del presente estudio participativo, se adoptó esta metodología por su facilidad de aplicación al permitir que las personas de la comunidad evaluaran el estado fitosanitario del bosque sin requerir conocimientos técnicos especializados. Se registró si los árboles presentaban síntomas de ataque por Dendroctonus en el momento de la evaluación (p. ej., grumos de resina sobre la corteza) o si presentaba marcas de haber sufrido un ataque anterior (p. ej., orificios con aserrín o galerías en la corteza sin escarabajos). Esta aproximación integrada facilita la interpretación de los datos y el seguimiento de la dinámica de infestaciones en un marco de manejo forestal comunitario.
Tabla 1
Localidades de estudio con su altitud correspondiente.
Nombre del sitio
Altitud (m snm)
El Llano
1,937
Sin nombre
2,200
Yalahui
2,263
Cuatro Caminos
2,385
Loma de Ocote
2,628
Lodiu
2,755
Para analizar si había diferencias en el vigor de los pinos entre sitios de muestreo, se realizó una prueba de X2 que comparó las categorías del arbolado. También se efectuó un análisis de componentes principales (PCA) como herramienta exploratoria para categorizar los tipos de manejo y observar si existía algún tipo de agrupación entre los sitios muestreados; las variables utilizadas en el PCA fueron: extracción de leña, actividades de ecoturismo, sin manejo actual, saneamiento oportuno y saneamiento inoportuno. Adicionalmente, se aplicó una regresión lineal utilizando las 2 primeras dimensiones del PCA con el fin de determinar si los distintos tipos de manejo forestal influían en la abundancia de los escolítidos del género Dendroctonus. Este enfoque combinado (PCA + regresión) brinda una manera de explorar patrones en los datos y probar si dichos patrones tienen un efecto estadísticamente significativo en la abundancia de los descortezadores.
Resultados
El 2003 fue el primer año en el que se identificaron los brotes de plaga de escarabajos descortezadores en Santa Catarina Lachatao. Derivado de las diferentes visiones e intereses de las comunidades que conformaban Pueblos Mancomunados en torno al manejo forestal, no se tomaron medidas inmediatas en cuanto al control de la plaga. Así, al siguiente año, en 2004, la Conafor trató de llegar a acuerdos con los municipios, las localidades y las agencias de policía para controlar la infestación por descortezadores, sin tener éxito por lo que la plaga continuó propagándose.
Fue hasta el 2009, debido a la preocupación general de la comunidad de Lachatao, cuando las mismas personas comenzaron a controlar la plaga sin intervención de expertos. Este saneamiento consistió en derribar los pinos plagados; no obstante, en la comunidad no se sintieron satisfechos con el resultado de esta intervención. Por lo anterior, para el 2010 en Cuatro Caminos (2,385 m snm) contrataron a una empresa privada para que realizara el saneamiento, debido a que la plaga estaba muy propagada. En este sitio, se consideró que el saneamiento fue inoportuno, ya que tuvieron que quemar y hacer derribos de una gran cantidad de árboles. En este sitio no se hizo extracción de la madera restante del saneamiento, pues fueron demandados por la Procuraduría Federal de Protección al Ambiente (Profepa), debido a que no se realizó el procedimiento establecido por la Ley General de Desarrollo Forestal Sustentable y por la Norma Oficial Mexicana NOM-019-RECNAT-2006, que establece los lineamientos técnicos para el combate y control de los insectos descortezadores de las coníferas, por lo que se les impidió hacer uso de esa madera. El sitio se dejó sin manejo y no se intentó reforestar en los años siguientes; para el 2019 era evidente el proceso de regeneración natural en el sitio y las personas afirmaron que el sitio comenzó a mejorar por sí solo después del saneamiento, a pesar de que en su momento fue considerado inoportuno.
En 2013, en Loma de Ocote (2,628 m snm), se detectó un rebrote de la plaga. Aprendiendo de su experiencia anterior, los habitantes de SCL iniciaron las acciones de control inmediatamente derribando los pinos afectados y notificando a Conafor. Esto se consideró un saneamiento oportuno sin pérdidas mayores. Ese mismo año, también fue el último en el que se tomaron acuerdos entre las comunidades que conformaban Pueblos Mancomunados para extraer madera sana y dañada del área correspondiente a SCL. Estos hechos confirmaron la importancia de una visión ya arraigada en la comunidad: en SCL, las personas consideran que es necesario que la comunidad se mantenga unida e informada sobre las dinámicas, tareas y problemas en torno al bosque, ya que la división al interior de la comunidad afecta el manejo y bienestar del mismo. Con respecto al manejo forestal comunitario, se identificaron 3 principales actividades de los sitios de muestreo establecidos para este estudio en SCL: extracción de leña, ecoturismo, saneamiento forestal de plaga del escarabajo descortezador (con 2 categorías: oportuno e inoportuno) y se agregó una cuarta categoría, sin manejo actual.
La extracción de leña se realiza en 3 sitios: El Llano, Yalahui y Lodiu (1,937, 2,263 y 2,755 m snm, respectivamente) y es para consumo local, solo extraen personas de la comunidad de Lachatao y de la comunidad de Yavesía. El Llano (1,937 m snm) fue el único sitio donde se realizan actividades de ecoturismo. Se le otorgó la categoría de saneamiento oportuno al control que se realizó inmediatamente después de identificar el brote en Yalahui (2,628 m snm). Por otro lado, el saneamiento inoportuno se definió como aquel que se demoró en controlar la plaga y se propagó al punto de tener que realizar un saneamiento más agresivo en Cuatro Caminos (2,385 m snm). A la fecha del muestreo se encontraron 3 sitios sin manejo, es decir, donde no se realiza ninguna actividad forestal: Sin nombre, Cuatro Caminos y Yalahui (2,200, 2,385 y 2,628 m snm, respectivamente).
Durante los 5 meses de muestreo se recolectaron 13,948 insectos que fueron clasificados a nivel de orden, de los cuales 4,477 individuos pertenecen a géneros de escarabajos descortezadores. El género más abundante y para el que se identificaron a una categoría de especie los individuos recolectados, fue Dendoctonus (n = 4,033).Se reportaron 3 especies primarias: D. mexicanus (n = 2,080), D.frontalis (n = 1,249)y D. adjunctus (n = 493), y la presencia de 3 especies secundarias: D.valens, D.parallelocollis y D. approximatus.
Con respecto a los diferentes sitios estudiados, la abundancia de Dendroctonus varió de entre 2 y 6 especies; en El Llano se encontraron únicamente 2 especies, en Sin nombre, Yalahui, Cuatro Caminos y Loma de ocote se encontraron 4 especies y en Lodiu 6 especies (tabla 2). Las especies que estuvieron presentes en más sitios fueron D. mexicanus, D. valens y D. approximatus (5 sitios), mientras que D. adjuctus estuvo presente en 4 sitios, D. frontalis en 3 y D. parallelicolis en un solo sitio. En términos de abundancia de escarabajos, Dendroctonus tuvo una variación significativa (X = 670 ± 945), el sitio con mayor abundancia presentó 2,433 individuos (Cuatro Caminos), mientras que en el sitio Sin nombre se registraron únicamente 57 individuos.
Se identificaron 3 especies de Pinus: P. teocote, P. pseudostrobus yP. patula. El sitio con mayor riqueza de especies fue Lodiu (2,755 m snm), donde se encontraron las 3 especies de Pinus, en el sitio Sin nombre (2,200 m snm) se encontraron 2 especies (P. teocote y P. pseudostrobus), al igual que en Cuatro Caminos (2,385 m snm) (P. teocote y P. patula), en El Llano, Yalahui y Loma de Ocote (1,937, 2,263 y 2,628 m snm, respectivamente) solo se identificó 1 especie (P. teocote). En las parcelas estudiadas encontramos un rango de abundancias de Pinus entre 32 y 12 individuos por parcela. El Llano (1,937 m snm) presentó la mayor abundancia mientras que Cuatro Caminos y Loma de Ocote (2,385 y 2,628 m snm, respectivamente) fueron los sitios con menor abundancia donde se realizó saneamiento para controlar la plaga por descortezador.
Tabla 2
Riqueza de especies de los sitios de estudio.
Nombre del sitio
Altitud (m snm)
Riqueza de Dendroctonus
Riqueza de Pinus
El Llano
1,937
2
1
Sin nombre
2,200
4
2
Yalahui
2,263
4
1
Cuatro Caminos
2,385
4
2
Loma de Ocote
2,628
4
1
Lodiu
2,755
6
3
En cuanto al vigor del arbolado, se observa que la mayoría de los pinos se encuentra dentro de las primeras 3 categorías de vigor, pero hubo diferencias significativas entre sitios (X2 = 86.3, gl = 25, p = 0.05), particularmente en Cuatro Caminos y Loma de Ocote que son los sitios con saneamiento (2,385 y 2,628 m snm) no hay árboles muy dañados (categorías de vigor 5 y 6; fig. 1).
Finalmente, los resultados del PCA muestran que 79% de la variación observada en los sitios muestreados está fuertemente asociada con el tipo de manejo forestal que se realiza. En este sentido, se identificó que las prácticas de extracción de leña y la ausencia de manejo actual son las principales variables que explican la diferenciación de los sitios, seguidas por el saneamiento forestal, mientras que el ecoturismo tuvo una contribución menor (fig. 2). Para evaluar si el manejo llevado a cabo en SCL se relaciona con las abundancias de escarabajos descortezadores, analizamos la abundancia total de Dendroctonus y la abundancia de D. mexicanus con respecto a la dimensión 1 del PCA. Encontramos que los análisis no evidenciaron una relación significativa entre el manejo forestal y la abundancia total de Dendroctonus cuando se consideran todas las especies juntas (D1 = F1,4 = 3.4, p = 0.14). En contraste, D. mexicanus,la especie más abundantesí muestra una relación positiva con la dimensión 1 del PCA; en particular, los sitios en donde se ha realizado manejo presentan una mayor abundancia de esta especie(D1 = F1,4 = 31.17, p = 0.005, R2 = 0.88; fig. 3).
Discusión
Los escarabajos descortezadores cumplen una función ecológica clave al eliminar árboles adultos y permitir la regeneración de los pinos al facilitar la entrada de luz al sotobosque. Sin embargo, algunas especies tienen un alto potencial de convertirse en plagas forestales, por lo que es crucial monitorear sus poblaciones para evitar crecimientos descontrolados (Castellanos-Bolaños et al., 2009; Durán y Poloni, 2014; Equihua-Martínez et al., 2011; Raffa et al., 2008).El manejo forestal es fundamental para la detección y control oportuno de plagas. Conocer estas especies permite identificarlas a tiempo, destacando la importancia del monitoreo y saneamiento en colaboración con comunidades y ejidos forestales (Castellanos-Bolaños et al., 2013; Conafor, 2018). En este estudio, en SCL se encontraron 6 de las 7 especies de Dendroctonus reportadas para Oaxaca (Cibrián-Tovar et al., 1995; Salinas-Moreno et al., 2010) y se registraron 3 especies de Pinus, por lo que se puede decir que hay una alta riqueza de estos escarabajos. Dentro de esta diversidad se encontraron 3 especies agresivas o capaces de matar árboles sanos (D. adjunctus, D. frontalis, D. mexicanus) y 3 que muestran una baja capacidad de agregación de adultos, una baja tolerancia a las defensas del hospedador y son incapaces de matar árboles sanos (Payne et al., 1973): D. approximatus, D. parallelocollis, D. valens; de tal manera que no todos los congenéricos registrados en SCL se deberían considerar como especies susceptibles a tratamientos fitosanitarios.
Figura 1. Vigor del arbolado por categorías en los 6 sitios de muestreo seleccionados en el bosque de Santa Catarina Lachatao, Oaxaca, donde 1 representa a un árbol en perfecto estado y 6 representa a un árbol muerto.Figura 2. Análisis de componentes principales, donde se observa la agrupación de los sitios de estudio. Cada sitio está representado por su altitud. El color de la flecha refleja la contribución de cada variable al análisis.Figura 3. Relación entre la dimensión 1 del PCA y la abundancia de Dendroctonus mexicanus por sitio (escala logarítmica). D1 = F1,4 = 31.17, p = 0.005, R2 = 0.88.
La abundancia total de escarabajos descortezadores encontrados y específicamente la abundancia de las especies de importancia fitosanitaria en SCL (4,033 individuos, de los que 3,822 son especies agresivas, de los cuales 1,249 eran D. frontalis, 2,080 eran D. mexicanus y 493 eran D. adjunctus recolectados en 5 meses) es menor en comparación con la reportada en otros sitios, ~ 50,000 en 9 meses en Zimapán, Hidalgo (Soto-Correa et al., 2019), ~ 134,000 en 10 meses en la Reserva de la Biosfera de la Sierra Gorda (López-Gómez et al., 2017), y similar a los ~ 40,000 descortezadores en 11 meses al norte del estado de Querétaro (Morales-Rangel et al., 2018). Esta comparación debe interpretarse con cautela, dado que las diferencias en las condiciones del bosque, los períodos de muestreo y la composición de los atrayentes utilizados dificultan establecer comparaciones directas. Debido a la falta de una línea base en el contexto local, estos resultados deben tomarse más a modo preliminar. En este contexto, pareciera que las abundancias actuales no están indicando un problema importante en ninguno de los sitios muestreados. Sin embargo, es fundamental mantener un monitoreo comunitario activo (Cibrián-Tovar y Macías-Sámano, 2021).
El brote masivo de escarabajos descortezadores reportado en las entrevistas realizadas en SCL coincide con los periodos reportados en los informes generales en que se presentaron grandes brotes en más comunidades en la sierra Norte de Oaxaca. En el período de 2004 a 2009, más de 6,300 ha de bosques de pinos de Oaxaca fueron atacadas por los insectos descortezadores D. adjuntus, D. mexicanus y D. frontalis (Castellanos-Bolaños et al., 2009). Además, en el periodo de 2009 a 2014, Pueblos Mancomunados fue reportado como la región con mayor devastación de bosques causada por la plaga del escarabajo descortezador (Castellanos-Bolaños et al., 2013; Salinas-Moreno et al., 2010; Sosa-Díaz et al., 2018). Con lo anterior y aunado a la percepción local, se confirma que en SCL las poblaciones de Dendroctonus fueron plaga y que las medidas que tomó la comunidad ante dicha problemática, a pesar de los desacuerdos intracomunitarios, evitaron que el brote epidémico siguiera propagándose. Existen reportes de que en varios casos, el manejo efectivo de esta plaga se ha visto menguado por la existencia de conflictos comunitarios, problemas de litigio en los terrenos plagados, priorización de la extracción de madera sana o incluso por la carencia de programas de manejo forestal. Todas estas barreras limitan las acciones para atender el saneamiento de los bosques, dando lugar a que las plagas se expandan con más facilidad (Castellanos-Bolaños et al., 2009; Conafor, 2018).
En varias comunidades con manejo forestal maderero donde se promueve el crecimiento de una o pocas especies maderables, en épocas recientes se han visto impactos más severos de los brotes de especies de descortezadores agresivas, debido a la poca diversidad tanto de especies como de generaciones de pinos (Castellanos-Bolaños et al., 2009; Conafor, 2014; Sánchez-Martínez et al., 2007). Sin embargo, el manejo forestal comunitario que se realiza en SCL tiene la intención de conservar el bosque de manera que la comunidad pueda seguir abasteciéndose de los bienes que brinda, únicamente para autoconsumo. Rojas-Serrano y Martínez-Corona (2017) encontraron que en la comunidad de SCL se cree que la supervivencia del pueblo depende de la conservación del bosque. Gracias al monitoreo constante de los pobladores, el manejo tiene un bajo impacto en las dinámicas del ecosistema, lo que permite que el bosque conserve su capacidad de resistencia y recuperación ante disturbios naturales.
El hecho de haber encontrado una relación entre las prácticas de manejo forestal y las abundancias del descortezador D. mexicanus pone de manifiesto que las áreas previamente saneadas deberían continuar bajo monitoreo comunitario activo, puesto que tienen el mayor potencial de generar nuevos brotes. Debido a que después de los saneamientos el bosque presenta árboles con tallas menores, estas áreas podrían ser menos susceptibles como se ha demostrado en otros estudios, donde comparan la susceptibilidad de ataque de árboles pequeños y grandes (Doak, 2004; Hutchinson y Reid, 2022; Koontz et al., 2021), no obstante, es fundamental continuar con el monitoreo comunitario para detectar los brotes nuevos de manera oportuna.
Debido a que la predicciones del cambio climático apuntan a un incremento en las temperaturas máximas y en las condiciones de sequía para México (Sáenz-Romero et al., 2010), y a que se ha demostrado que estas condiciones pueden influir en la dinámica poblacional de Dendroctonus —de forma que sus poblaciones tienen mayor tasa de supervivencia y reproducción (Billings, 2011; Sáenz-Romero et al., 2023)— a la par de un incremento en la susceptibilidad y estrés por parte de los pinos (Creeden et al., 2014; Raffa et al., 2008), es fundamental fortalecer y mantener un monitoreo de las poblaciones de descortezadores y evaluar continuamente la salud del bosque (Billings, 2011; Cibrián-Tovar y Macías-Sámano, 2021). Si bien el ecosistema posee mecanismos de resiliencia que pueden modular la respuesta de las poblaciones de insectos descortezadores ante cambios ambientales, los desafíos sin precedentes asociados al cambio climático requieren estrategias de manejo sustentable, adaptativo y comunitario. En este sentido, el enfoque participativo adoptado —que combinó la evaluación fitosanitaria con el conocimiento local— permitió una comprensión más integral de los efectos de los brotes de escarabajos descortezadores (Pacheco-Aquino y Durán, 2023). Asimismo, este enfoque resalta la importancia de la implementación de saneamientos oportunos (p. ej., en SCL) como una herramienta clave para mitigar el impacto de los descortezadores y contribuir a la conservación de los bosques en escenarios de variabilidad climática.
Los resultados de esta investigación nos indican que el bosque de Santa Catarina Lachatao presenta buenas condiciones fitosanitarias y refleja su resiliencia gracias al manejo comunitario y las acciones de control aplicadas tras el brote de escarabajos descortezadores entre 2003 y 2009, con un rebrote en 2013. Aunque las especies más abundantes del género Dendroctonus (D. mexicanus, D. frontalis y D. adjunctus) son consideradas las más agresivas para los sistemas forestales mexicanos, su tamaño poblacional en esta comunidad está muy alejado al de otras zonas que presentan brotes considerables de descortezadores. Sin embargo, es crucial mantener un monitoreo activo y participativo debido a su potencial epidémico, especialmente ante el aumento de sequías y estrés hídrico previsto por el cambio climático, lo que podría incrementar las infestaciones. Estos hallazgos subrayan la importancia de un manejo comunitario adaptativo para enfrentar futuros brotes en un contexto de cambio climático.
Agradecimientos
A los pobladores y autoridades de la comunidad de Santa Catarina Lachatao por permitirnos trabajar en su territorio y compartir sus experiencias de manejo con nosotras. En particular a la familia Santiago Hernández, que fue crucial para el buen desempeño de la investigación. Todas las personas entrevistadas dieron su consentimiento para realizar el estudio.
Referencias
Armendáriz-Toledano, F., Zúñiga, G., García-Román, L. J., Valerio-Mendoza, O. y García-Navarrete, P. G. (2018). Guía ilustrada para identificar a las especies del género Dendroctonus presentes en México y Centroamérica. Ciudad de México:Instituto Politécnico Nacional.
Bale, J. S., Masters, G. J., Hodkinson, I. D., Awmack, C., Bezemer, T. M., Brown, V. K. et al. (2002). Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Global Change Biology, 8, 1–16.
Bello-Cervantes, E. (2015). Evaluación del deterioro ambiental del territorio Pueblos Mancomunados, Oaxaca: estudio con miras de restauración(Tesis de maestría). Instituto de Geografía, Universidad Nacional Autónoma de México. México, D.F.
Biedermann, P. H. W., Müller, J., Grégoire, J., Gruppe, A., Hagge, J., Hammerbacher, A. et al. (2019). Bark beetle population dynamics in the Anthropocene: challenges and solutions. Trends in Ecology & Evolution, 34, 914–924. https://doi.org/10.1016/j.tree.2019.06.002
Billings, R. F. (2011). Mechanical control of southern pine beetle infestations. En R. N. Coulson y K. D. Klepzig (Eds.), Southern pine beetle II. Gen. Tech. Rep. SRS-140 (pp. 399-413). Asheville, NC: U.S. Department of Agriculture Forest Service, Southern Research Station.
Boyd, I. L., Freer-Smith, P. H., Gilligan, C. A. y Godfray, H. C. J. (2013). The consequence of tree pests and diseases for ecosystem services. Science, 342, 1235773. https://doi.org/10.1126/science.1235773
Bray, D. B. (2013). When the State supplies the Commons: origins, changes, and design of Mexico’s Common Property Regime. Journal of Latin American Geography, 12, 33–55. https://doi.org/10.1353/lag.2013.0003
Bray, D. B., Durán, E. y Molina-González, O. A. (2012). Beyond harvests in the commons: Multi-scale governance and turbulence in indigenous/community conserved areas in Oaxaca, Mexico. International Journal of the Commons, 6, 151–178.
Castellanos-Bolaños, J. F., Maldonado-Arango, O., Piñeiro-Márquez, F., Hernández-Hernández, J. y Gómez-Cárdenas, M. (2009). Condición de los bosques afectados por insectos descortezadores en el estado de Oaxaca, México. Memorias del XV Simposio Nacional de Parasitología Forestal. Oaxaca, México.
Castellanos-Bolaños, J. F., Ruiz-Martínez, E. O., Gómez-Cárdenas, M. y González-Cubas, R. (2013). Fundamentos técnicos para el control de insectos descortezadores de pinos en Oaxaca. Folleto Técnico Núm. 40. Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias/ Centro de Investigación Regional Pacífico Sur/ Campo Experimental Valles Centrales de Oaxaca. Villa de Etla, Oaxaca, México.
Challenger, A. (2003). Conceptos generales acerca de los ecosistemas templados de montaña de México y su estado de conservación. En O. Sánchez, E. Vega, E. Peters y O. Monroy-Vilchis (Eds.), Conservación de ecosistemas templados de montaña en México (pp. 17–44). Ciudad de México: Instituto Nacional de Ecología.
Challenger, A. y Soberón, J. (2008). Los ecosistemas terrestres. En J. Soberón, G. Halffter y J. Llorente-Bousquets (Eds.), Capital natural de México. Vol. I. Conocimiento actual de la biodiversidad (pp. 87–108). México D.F.: Conabio.
Chapela, G. y Merino, L. (2019). Hacia una política forestal sustentable e incluyente. Los bosques de México, problemas y propuestas. Crisis ambiental en México ruta para el cambio climático. Ciudad de México: Universidad Nacional Autónoma de México.
Cibrián-Tovar, D. y Macías-Sámano, J. E. (2021). Marco conceptual y desarrollo del manejo integrado de plagas MIPF. En D. Cibrián-Tovar (Ed.), Fundamentos para el manejo de plagas forestales MIPF (pp. 3–12). Texcoco: Universidad Autónoma Chapingo.
Cibrián-Tovar, D., Méndez, J. T., Campos, R., Yates III, O. y Flores, J. (1995). Insectos forestales de México. Estado de México: Universidad Autónoma Chapingo,
Cibrián-Tovar, D., Quiñonez-Favila, S. A., Quiñonez-Barraza, S., Olivo-Martínez, J. A., Robles-Uribe, S., Hernández-Villa, L. et al. (2015). Manual para la identificación y manejo de insectos descortezadores del pino: guía práctica para la identificación y manejo de los descortezadores del pino. Ciudad de México: Conafor.
Conafor (Comisión Nacional Forestal). (2014). Resultados del mapeo aéreo fitosanitario. Oaxaca: Gerencia Estatal Oaxaca, Comisión Nacional Forestal.
Conafor (Comisión Nacional Forestal). (2018). Programa Institucional de la Comisión Nacional Forestal 2014–2018. Avance y Resultados 2018. México D.F.: Comisión Nacional Forestal, Gobierno de México.
Creeden, E. P., Hicke, J. A. y Buotte, P. C. (2014). Climate, weather, and recent mountain pine beetle outbreaks in the western United States. Forest Ecology and Management, 312, 239–251.4 https://doi.org/10.1016/j.foreco.2013.09.051
Cudmore, T. J., Björklund, N., Carroll, A. L. y Staffan-Lindgren, B. (2010). Climate change and range expansion of an aggressive bark beetle: evidence of higher beetle reproduction in native host tree populations. Journal of Applied Ecology, 47, 1036–1043. https://doi.org/10.1111/j.1365-2664.2010.01848.x
Del-Castillo, R. F., Pérez-de la Rosa, J. A., Vargas-Amado, G. y Rivera-García, R. (2004). Coníferas. En A. J. García-Mendoza, M. J. Ordóñez y M. Briones-Salas (Eds.), Biodiversidad de Oaxaca (pp. 141–158). México D.F.: Instituto de Biología, UNAM/ Fondo Oaxaqueño para la Conservación de la Naturaleza/ World Wildlife Fund.
del Río-Mora, A. (2011). Manejo integrado de plagas forestales: un enfoque ecológico para la sustentabilidad de los ecosistemas forestales. México D.F.: INIFAP.
Díaz, S., Demissew, S., Carabias, J., Joly, C., Lonsdale, M., Ash, N. et al. (2015). The IPBES Conceptual framework – connecting nature and people. Current Opinion in Environmental Sustainability, 14, 1–16. https://doi.org/10.1016/j.cosust.2014.11.002
Doak, P. (2004). The impact of tree and stand characteristics on spruce beetle (Coleoptera: Scolytidae) induced mortality of white spruce in the Copper River Basin Alaska. Canadian Journal of Forest Research, 34, 810 –816 https://doi.org/10.1139/x03-256
DOF (Diario Oficial de la Federación). (1961, 20 de octubre). Tomo CCXLVIII, núm. 41. Gobierno de México. https://www.dof.gob.mx/
Dukes, J. S., Pontius, J., Orwig, D., Garnas, J. R., Rodgers, V. L., Brazee, N. et al. (2009). Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: what can we predict? Canadian Journal of Forest Research, 39, 231–248 https://doi.org/10.1139/X08-171
Durán, E. y Poloni, A. (2014). Escarabajos descortezadores: diversidad y saneamiento en bosques de Oaxaca. Conabio. Biodiversitas, 117, 7–12
Equihua-Martínez, A., Estrada-Venegas, E. G. y Burgos-Solorio, A. (2011). Descortezadores y barrenadores (Insecta: Coleoptera: Scolytidae). En A. Cruz Aragón (Ed.), La biodiversidad en Veracruz: estudio de estado (pp. 367–370). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Gobierno del Estado de Veracruz/ Universidad Veracruzana/ Instituto de Ecología, A.C.
FAO (Food and Agriculture Organization of the United Nations). (2010). Global forest resources assessment 2010(Vol. 163). Rome: FAO.
Farjon, A. (1996). Biodiversity Pinus (Pinaceae) in Mexico: speciation and palaeoendemism. Botanical Journal of the Linnean Society, 121, 365–384. https://doi.org/10.11 11/j.1095-8339.1996.tb00762.x
Fernández, E. A., Romero, C. R. y Zavala, H. J. (2013). Atlas climático digital de México. Centro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México. http://uniatmos.atmosfera.unam.mx
Gasca-Zamora, J. (2014). Gobernanza y gestión comunitaria de recursos naturales en la Sierra Norte de Oaxaca. Región y Sociedad, 26, 89–119. https://doi.org/10.22198/rys.2014.60.a11
Gaylord, M. L., Kolb, T. E. y McDowell, N. (2015). Mechanisms of piñon pine mortality after severe drought: a retrospective study of mature trees. Tree Physiology, 35, 806–816. https://doi.org/10.1093/treephys/tpv038
Gitai, H., Suárez, A., Watson, R. T. y Dokken, D. J. (2002). Climate change and biodiversity. Ginebra: IPCC Technical paper V. IPCC.
Hart, S. J., Veble, T. T., Eisenhart, K. S., Jarvis, D. y Kulakowski, D. (2014). Drought induces spruce beetle (Dendroctonus rufipennis) outbreaks across northwestern Colorado. Ecology, 95, 930–939. https://doi.org/10.1890/13-0230.1
Hutchinson, I. K. y Reid, M. L. (2022). Phloem and defense traits in relation to tree size and age: implications for host selection by bark beetles. Forest Ecology and Management, 513, 120183. https://doi.org/10.1016/j.foreco.2022.120183
INEGI (Instituto Nacional de Estadística, Geografía e Informática). (2005). Conjunto de datos vectoriales de la carta de uso del suelo y vegetación: escala 1 : 250 000. Serie III (continuo nacional). Aguascalientes: Instituto Nacional de Estadística, Geografía e Informática, INEGI (Instituto Nacional de Estadística, Geografía e Informática)-Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad)-INE (Instituto Nacional de Ecología). (2007). Ecorregiones de México, nivel IV, escala 1 : 1 000 000. México D.F.: Instituto Nacional de Estadística, Geografía e Informática-Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Instituto Nacional de Ecología.
INEGI (Instituto Nacional de Estadística, Geografía e Informática). (2010). Información Nacional por entidad federativa y municipios. México D.F.: Instituto Nacional de Estadística, Geografía e Informática.
Innes, J. L. (1993). Methods to estimate forest health. Silva Fenica, 27, 145–157.
Jepsen, J. U., Hagen, S. B., Ims, R. A. y Yoccoz, N. G. (2008) Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: evidence of a recent outbreak range expansion. Journal of Animal Ecology, 77, 257–264 https://doi.org/10.1111/j.1365-2656.2007.01339.x
Koleff, P., Soberón, J. y Smith, A. (2004). Madrean pine oak woodland. En R. A. Mittermeier, P. Robles-Gil, M. Hoffmann, J. D. Pilgrim, T. A. Brooks, C. G. Mittermeier et al. (Eds.), Hotspots (Biodiversidad amenazada II). Nuevas ecorregiones terrestres prioritarias del mundo (pp. 291–293). Monterrey, México: Cemex/ Agrupación Sierra Madre.
Koontz, M. J., Latimer, A. M., Mortenson, L. A., Fettig, C. J. y North M. P. (2021). Cross-scale interaction of host tree size and climatic water deficit governs bark beetle-induced tree mortality. Nature Communications, 12, 1–13. https://doi.org/10.1038/s41467-020-20455-y
López-Gómez, V., Torre-Huerta, B., Reséndiz-Martínez, J. F., Sánchez-Martínez, G. y Gijón-Hernández, A. R. (2017) Influence of climatic parameters on the population fluctuations of the complex Dendroctonus frontalis Zimmerman, 1869 and Dendroctonus mexicanus Hopkins, 1909. Revista Mexicana de Ciencias Forestales, 8, 7–29. https://doi.org/10.29298/rmcf.v11i59.668
Macías-Sámano, J. E. y Niño-Domínguez, A. (2016). Protocolo para monitoreo de descortezadores de coníferas mediante el uso de atrayentes y semioquímicos: para México y Centroamérica. San Cristóbal de Las Casas: ECOSUR.
Marini, L., Ayres, P. M., Battisti, A. y Faccoli, M. (2012). Climate affects severity and altitudinal distribution of outbreaks in an eruptive bark beetle. Climatic Change, 115, 327–341. https://doi.org/10.1007/s10584-012-0463-z
Mathus-Alonso, M., López-Pardo, G., Gasca-Zamora, J. y Villavicencio, B. P. (2010). La gestión comunitaria de recursos naturales y ecoturísticos en la Sierra Norte de Oaxaca. Ciudad de México: IIE, Universidad Nacional Autónoma de México.
Morales-Rangel, A., Cambrón-Sandoval, V. H., Soto-Correa, J. C., Jones, R. W. y Obregón-Zuñiga, J. A. (2018) Efecto de la temperatura en poblaciones de Dendroctonus frontalis Zimmerman y Dendroctonus mexicanus Hopkins (Coleoptera: Curculionidae: Scolytinae) bajo un escenario de cambio climático en la Sierra Gorda queretana. Acta Zoológica Mexicana (nueva serie), 34, 1–8. https://doi.org/10.21829/azm.2018.3412141
Ning, H., Tang, M. y Chen, H. (2021). Impact of climate change on potential distribution of chinese white pine beetle Dendroctonus armandi in China. Forests, 12, 544. https://doi.org/10.3390/f12050544
Pacheco-Aquino, G. y Durán, E. (2023). Tree canopy patterns in a temperate forest in Oaxaca, Mexico after bark beetle outbreaks: Implications for ecological resistance and resilience. Forest Ecology and Management, 542, 121099. https://doi.org/10.1016/j.foreco.2023.121099
Raffa, K. F., Aukema, B. H., Bentz, B. J., Carroll, A. L., Hicke, J. A., Turner, M. G. et al. (2008). Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. BioScience, 58, 501–517. https://doi.org/10.1641/B580607
Raffa, K. F., Grégoire, J. y Lindgren, B. S. (2015). Natural history and ecology of bark beetles. En F. E. Vega y R. W. Hofstetter (Eds.), Bark beetles (pp. 1–40). Oxford, UK: Academic Press. https://doi.org/10.1016/b978-0-12-417156-5.00001-0
Rivera-Jiménez, E., Regino-Maldonado, J., Pérez-Rosas, B., Luis-Santiago, M., García-Hernández, J., Lázaro-Juárez, E. et al. (2019). La plaga por descortezador en Oaxaca, México: análisis con ecuaciones estructurales. Contribución al Conocimiento Científico y Tecnológico en Oaxaca, 3, 27–35 https://doi.org/10.13140/RG.2.2.22272.17922
Rojas-Serrano, C. y Martínez-Corona, B. (2017). Transformaciones y continuidades en el manejo y concepción del bosque y las relaciones de género en Santa Catarina Lachatao, Oaxaca. Agricultura, Sociedad y Desarrollo, 14, 451–478. https://dialnet.unirioja.es/servlet/articulo?codigo=6151954
Rubín-Aguirre, A., Sáenz-Romero, C., Lindig-Cisneros, R., del Río-Mora, A. A., Tena-Morelos, C. A., Campos-Bolaños, R. et al. (2015). Bark beetle pest in an altitudinal gradient of a Mexican managed forest. Forest Ecology and Management, 343, 73–79 https://doi.org/10.1016/J.FORECO.2015.01.028
Sáenz-Romero, C., Cambrón-Sandoval, V. H., Hammond, W., Méndez-González, J., Luna-Soria, H., Macías-Sámano, J. E. et al. (2023). Abundance of Dendroctonus frontalis and D. mexicanus (Coleoptera: Scolytinae) along altitudinal transects in Mexico: Implications of climatic change for forest conservation. Plos One, 18, e0288067. https://doi.org/10.1371/journal.pone.0288067
Sáenz-Romero, C., Rehfeldt, G. E., Crookston, N. L., Duval, P. y Beaulieu, J. (2010). Spline models of contemporary, 2030, 2060 and 2090 climates for Mexico and their use in understanding climate-change impacts on the vegetation. Climatic Change, 102, 595–623. http://doi.org/10.1007/s10584-009-9753-5
Salinas-Moreno, Y., Ager, A., Vargas, C. F., Hayes, J. L. y Zúñiga, G. (2010). Determining the vulnerability of Mexican pine forests to bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). Forest Ecology and Management, 260, 52–61. https://doi.org/10.1016/j.foreco.2010.03.029
Sánchez-Martínez, G., Torres-Espinosa, L. M., Vázquez-Collazo, I., González-Gaona, E. y Narváez-Flores, R. (2007). Monitoreo y manejo de insectos descortezadores de coníferas. Libro Técnico No. 4. Aguascalientes: INIFAP/ CIRNOC/ Campo Experimental Pabellón.
Sosa-Díaz, L., Méndez-González, J., García-Aranda, M. A., Cambrón-Sandoval, V. H., Villarreal-Quintanilla, J. Á., Ruiz-González, C. G. et al. (2018). Distribución potencial de barrenadores, defoliadores, descortezadores y muérdagos en bosques de coníferas de México Librado. Revista Mexicana de Ciencias Forestales, 9, 187–208
Soto-Correa, J. C., Avilés-Carrillo, I., Girón-Gutiérrez, D. y Cambrón-Sandoval, V. H. (2019). Abundancia altitudinal de Dendroctonus frontalis (Coleoptera: Curculionidae) en relación a variables climáticas en Hidalgo, México. Revista de Biología Tropical, 67, 370–379. https://doi.org/10.15517/RBT.V67I3.34436
Toledo, V. M. (2013). Indigenous peoples and biodiversity conservation: the case of Mexico. Journal of Agricultural and Environmental Ethics, 26, 983–1000.
Toledo, V. M. y Barrera-Bassols, N. (2008). La memoria biocultural: la importancia ecológica de las sabidurías tradicionales. Barcelona: Icaria Editorial.
Weed, A. S., Ayres, M. P. y Hicke, J. A. (2013). Consequences of climate change for biotic disturbances in North American Forests, Ecological Monographs, 83, 441–470. https://doi.org/10.1890/13-0160.1
Wood, S. L. (1982). The bark Ambrosia beetles of North and Central America (Coleoptera: Scolytidae): a taxonomic monograph. Great Basin Naturalist, 6, 1–1359.
Relationship of the establishment of Dendroctonus adjunctus and Arceuthobium vaginatum with the structure of a Pinus hartwegii forest in Nevado de Colima, Mexico
Angel Rolando Endara-Agramont *, Rebeca Dennise Varo-Rodríguez, José Jonathan Aguirre-Zúñiga, Claudia Guadalupe Enríquez-Sánchez y Alma Abigail Luna-Gil
Universidad Autónoma del Estado de México, Instituto de Ciencias Agropecuarias y Rurales “El Cerrillo Piedras Blancas”, 50000 Toluca, Estado de México, México
*Autor de correspondencia: arendaraa@uaemex.mx (A.R. Endara-Agramont)
Recibido: 6 mayo 2024; aceptado: 29 julio 2025
Resumen
Las poblaciones de Pinus hartwegii se establecen por encima de 3,500 m snm, en torno a las montañas más altas del país, una de ellas es el Parque Nacional Volcán Nevado de Colima, mismo que está afectado por Arceuthobiumvaginatum y Dendroctonusadjunctus. El objetivo de la investigación fue determinar el daño provocado por ambos patógenos sobre la estructura y regeneración del bosque de P. hartwegii. Para ello, se instalaron 384 unidades de muestreo (UM de 1,000 m2 cada uno), donde se registró información dasonómica asociada a niveles de infestación de muérdagos y descortezadores. De acuerdo con los resultados obtenidos, 20% del bosque se encuentra infestado por muérdago y descortezador. Además, se presenta la eliminación del dosel dominante y reducción de árboles sanos > 40 DN en los sitios afectados. Por lo anterior, se concluye que los altos niveles de infestación de D. adjunctus y la superficie infestada por A. vaginatum afectan la estructura del bosque y, por tanto, comprometen el establecimiento de la regeneración natural, siendo la ladera oeste en torno al volcán la más afectada.
Palabras clave: Muérdago; Descortezador; Pinus hartwegii; Alta montaña; Estrato arbóreo
Abstract
The populations of Pinus hartwegii are established above 3,500 m asl, around the highest mountains in the country, one of them is the Nevado de Colima Volcano National Park, which is affected by Arceuthobium vaginatum and Dendroctonus adjunctus. The objective of the research was to determine the damage caused by both pathogens on the structure and regeneration of the P. hartwegii forest. To this, 384 sampling units were installed (MU of 1,000 m2 each), where forest information associated with mistletoe and bark beetle infestation levels was recorded. According to the results obtained, 20% are attacked by mistletoe and bark beetle. In addition, the elimination of the dominant canopy and reduction of healthy trees > 40 DN is presented in the affected sites. From the above, it is concluded that the high levels of infestation of D. adjunctus and the surface infested by A. vaginatum affect the structure of the forest and, therefore, compromise the establishment of natural regeneration, with the western slope around the volcano the most affected.
Keywords: Mistletoe; Bark beetle; Pinus hartwegii; High mountain; Tree stratum
Introducción
Los bosques templados de México cubren 15% de la superficie forestal, distribuidos en 5 regiones: Península de Baja California, Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur y el Sistema Volcánico Transmexicano (SVTM) (Sánchez-González, 2008). Se encuentran en un rango altitudinal de 1,800 a 4,000 m snm, donde se establecen más de 60 especies del género Pinus (Farjon et al., 1997; Styles, 1993) y hasta 8 especies del género Abies (Gernandt y Pérez-de la Rosa, 2014; Villaseñor, 2016).
En Jalisco convergen la Sierra Madre Occidental y el Sistema Volcánico Transmexicano, donde se establecen 34 especies de coníferas (Gernandt y Pérez-de la Rosa, 2014), es una de las entidades con mayor diversidad forestal. Sin embargo, para el periodo 2012-2021 se reportaron 345 notificaciones de saneamiento, lo que representa una superficie total de 54,157.70 ha, en 7,347.02 de ellas, las notificaciones se levantaron por insectos descortezadores y en 33,064.31 por plantas parásitas. El Parque Nacional Volcán Nevado de Colima abarca 2 de las principales regiones afectadas (Tuxpan y Zapotlán el Grande), cuenta con bosques de Pinus hartwegii y Abies religiosa (Sivicoff, 2022).
Las poblaciones de Pinus hartwegii se establecen sobre 3,500 m snm, en las laderas de los edificios volcánicos más altos del país (Endara et al., 2013). Estos bosques se ven afectados por plantas parásitas del género Arceuthobium (Endara-Agramont et al., 2022; Sáenz et al., 2020) e insectos descortezadores del género Dendroctonus (Endara et al., 2023).
México concentra la mayor diversidad de especies del género Arceuthobium (23) (Villaseñor, 2016); las afectaciones por plantas parásitas predisponen a los árboles al ingreso de descortezadores (Ferrenberg, 2020; Mathiasen, 2019), debido a la pérdida de vigor y reducción en el crecimiento de los árboles parasitados (Cibrián-Tovar et al., 1995) y, posteriormente, facilitando el ingreso de hongos ambrosiales que conducen a la muerte del hospedero (Farrell et al., 2001). En rodales de pino con múltiples agentes de perturbación (Dendroctonus ponderosa y Arceuthobium vaginatum subs. cryptopodum) se encontraron bajos porcentajes de copas vivas y una predisposición a la acumulación de combustible (Klutsch et al., 2014). Por su parte, en poblaciones de Pinus contorta con historiales de mortalidad en más de 70% del dosel causado por descortezador, se concluye que el manejo de los individuos muertos propicia la regeneración hasta 4 veces mayor que en rodales no tratados (Collins et al., 2011). Este escenario, podría influir negativamente en la combinación de muérdagos y descortezadores en la zona de estudio, puesto que la regeneración restante puede ser infectada por el muérdago.
México alberga 13 especies del género Dendroctonus (Armendáriz-Toledano y Zúñiga, 2017), que afectan, al menos, a 20 especies de coníferas de los géneros Pinus, Pseudotsuga, Abies y Cupressus (Cibrián-Tovar et al., 1995; Salinas-Moreno et al., 2004). La función de los descortezadores en ecosistemas naturales es inducir el recambio de especies vegetales a través de la infestación de árboles viejos o estresados hasta causarles la muerte, para posteriormente dar paso a nueva generación del componente arbóreo y, con ello, permitir el establecimiento de individuos con mejores condiciones de competitividad (Christiansen y Bakke, 1988).
Uno de los controles poblacionales de los descortezadores son las temperaturas invernales, que ayudan a mantener bajas las poblaciones de las larvas y a retrasar el desarrollo de los adultos (Raffa et al., 2008); sin embargo, en las últimas décadas el aumento de la temperatura en verano está acelerando el desarrollo de los adultos, al incrementar su capacidad reproductiva y fecundidad, lo que implica una mayor cantidad de generaciones de descortezadores al año (Lombardero et al., 2000; Sáenz et al., 2020).
El Parque Nacional Volcán Nevado de Colima (PNVNC) reporta la presencia de insectos descortezadores en sus bosques de pino desde 1963 (Conanp, 2006). De la misma manera, Arceuthobium vaginatum afecta 14% de la superficie de los bosques de pino establecidos por encima de 3,500 m snm. (Endara et al., 2023). Por lo anterior, el objetivo de la investigación fue determinar el daño provocado por el descortezador y el muérdago enano sobre la estructura y regeneración del bosque de P. hartwegii del Nevado de Colima.
Materiales y métodos
El PNVNC fue decretado en 1936 y cuenta con una superficie de 6,554 ha, alcanza una altitud de 4,270 m snm (fig. 1). El tipo de clima es semifrío con verano fresco corto, subhúmedo con precipitaciones en verano C (w2)(w)(x´)c(e). Los tipos de suelo más importantes son regosol, andosol, litosol y en menor escala, cambisol. En el límite altitudinal superior arbóreo se establece una población de Pinus hartwegii asociado con el zacatonal alpino (3,500-4,100 m snm), entre 3,300 y 3,500 se distribuyen los bosques de Alnus jorullensis y Abies religiosa, mientras que el bosque mesófilo de montaña entre 3,000 y 3,300 m snm. Estos ecosistemas albergan aproximadamente 172 especies de plantas vasculares,124 de mamíferos y 117 de aves (Conanp, 2006).
Figura 1. Área de estudio y sitios de muestreo.
Se establecieron 384 unidades de muestreo (UM) temporales en 3,086 ha del bosque de P. hartwegii del Parque Nacional Nevado de Colima, Jalisco, México. Los sitios se instalaron por encima de 3,500 m snm y hasta 3,900 m snm. Para ello se utilizó la intensidad mínima de muestreo del inventario forestal de reconocimiento, esto permitió dar la representatividad a la superficie muestreada; posteriormente, se adaptó el método de transectos, utilizando las curvas de nivel como líneas de acceso al bosque (Dauber, 1995); sobre cada línea se instalaron las UM con una equidistancia de 300 m y una separación altitudinal de 100 m, esto debido a la variación climática asociada a la altitud (Mayer y Ott, 1991).
Cada UM abarca una superficie de 0.1 ha (17.86 m de radio), en el levantamiento de información del sitio como altitud, pendiente y exposición se utilizaron instrumentos de precisión (brújula Suunto Tandem y GPS Garmin Rino 700). El registro de la regeneración (< 2.5 cm de DN), juveniles (≥ 2.5 < 7.5 de DN) y fustales (≥ 7.5 cm de DN) permitió cuantificar al arbolado (DN y altura total) y clasificar sus niveles de infestación por muérdagos enanos (Hawksworth, 1983) e insectos descortezadores (Billings y Espino, 2005).
Con base en la NOM-019, se determinó la presencia de descortezadores (< a 3 árboles con presencia de grumos blandos) y brotes (≥ a 3 árboles con presencia de grumos blandos) en cada UM (DOF, 2018). Para determinar la especie de descortezador se observaron la presencia de grumos de coloración transparente a blanca en la corteza de los árboles infestados, donde se realizaron colectas manuales de individuos adultos y se preservaron en alcohol etílico al 70% (Macías et al., 2004). Posteriormente, se llevaron a laboratorio y se observaron bajo un microscopio estereoscopio LEICA y un óptico MOTIC (40X) para su identificación utilizando las claves taxonómicas de Wood (1982) y Cibrián-Tovar et al. (1995), además de la observación de la varilla seminal (Lanier et al., 1988; Perusquía, 1978). En arbolado con presencia de muérdago enano se colectaron muestras botánicas, las cuales fueron prensadas y llevadas al herbario de la Facultad de Ciencias Agrícolas de la Universidad Autónoma del Estado de México, donde se identificaron mediante claves taxonómicas (Rzedowski y Calderón, 2011).
La información de sitios e individuos permitió generar valores numéricos por sitios (sanos e infestados), así como niveles de infestación (bajo, medio y alto); estos datos se visualizaron en el software ArcGIS ver.10.8, donde se utilizó la extensión para la interpolación de datos del vecino natural (Sibson, 1982), con ello se generaron mapas temáticos para la distribución espacial ambos patógenos. La densidad del arbolado se traduce en la sumatoria de individuos juveniles y fustales sanos (DN ≥ 7.5), así como el número de árboles muertos en pie por hectárea, para ello, se consideraron los rangos propuestos por Endara et al. (2013) y Rojas et al. (2019): denso (≥ 336 ind./ha), semidenso (< 336 ≥ 150 ind./ha) y fragmentado (< 150 ≥ 20 ind./ha).
La estructura horizontal se determinó mediante la agrupación de los individuos por categoría diamétrica, iniciando en la categoría de 10 (≥ 7.5 < 12.4 cm DN). Para la estimación de los estratos arbóreos (estructura vertical) se agruparon a todos los juveniles y fustales con sus alturas totales, los cuales se sometieron a una prueba de Anova para identificar diferencias mínimas significativas en el paquete estadístico MINITAB Release 12.21. (Endara et al., 2013). Para determinar el efecto de la afectación de muérdagos y descortezadores, se separaron los sitios sanos de los infestados (presencia de muérdago y descortezador) y así compararlos. Se cuantificó todo el renuevo clasificándolo en plántulas (< 30 cm de altura), brinzales (≥ 30 cm < 1.5 m de altura) y latizales (≥ 1.5 m de altura < 2.5 cm DN); a este grupo se sumaron los juveniles, ya que todavía son altamente vulnerables a incendios forestales y aún no garantizan su establecimiento en el bosque.
Resultados
El trabajo en laboratorio permitió identificar al escarabajo de los pinos de montaña Dendroctonus adjunctus (fig. 2) afectando al pino de las alturas (P. hartwegii), mismo que ha sido reportado en esta ANP desde 1963 (Conanp, 2006). En esta investigación se determinó que 24% de los sitios presentan brotes activos (52 sitios) y 15% solo con presencia (32 sitios).
Las colectas de material botánico permitieron identificar a la especie de muérdago enano Arceuthobium vaginatum, que para el PNVNC ya había sido reportado por Ruiz (1994) y Endara et al. (2023) y cubre una superficie de 417 ha (14%), mientras que el descortezador abarca 209 ha (7%), ambos parecen tener un patrón de distribución similar, ya que más de 70% de la superficie se concentra en la ladera oeste en torno al Volcán Nevado de Colima (fig. 3). Respecto a la altitud, tanto el muérdago como el descortezador están distribuidos desde 3,500 y hasta 3,900 m snm, el descortezador en polígonos grandes y continuos, el muérdago en parches más dispersos (de arbolado juvenil y con niveles de infestación: bajo 298 ha, medio 79 ha y alto 39 ha).
Figura 2. Izquierda, Arceuthobium vaginatum; derecha, Dendroctonus adjunctus en Pinus hartwegii.
La distribución de la estructura horizontal muestra una marcada diferencia entre los sitios con y sin muérdago y descortezador; en primer lugar, es notoria la reducción del número de árboles sanos, provocada por el establecimiento, tanto de muérdago como de descortezador, misma que se refleja en 48% menos en las categorías de regeneración y 59% menos en arbolado adulto (categorías 10-85 cm); en segundo lugar, la ausencia de arbolado con potencial semillero (> 35 cm) puede comprometer el establecimiento de la regeneración natural de la especie (fig. 4).
El Anova utilizado para conocer la distribución del arbolado agrupado por sus alturas en sitios sanos e infestados muestra la presencia de 3 estratos bien diferenciados en el bosque sin infestaciones y solo 2 en el bosque infestado (fig. 5); esto atribuido a la mortalidad de arbolado adulto en zonas con brotes activos de descortezador.
La representación gráfica del estrato arbóreo muestra que, el bosque sin infestación, pese a presentar 3 estratos en el análisis estadístico, el número de árboles del estrato superior no es representativo (4 ind./ha-1), ya que al graficar un bosque de 1,000 m2, éste desaparece, atribuido a la baja densidad de estos bosques (236 ind./ha-1). Por otra parte, los bosques infestados presentan una densidad mayor (263 ind./ha-1), esto se explica por la preferencia del descortezador por arbolado adulto, mismos que conforman los bosques densos del parque nacional (fig. 6).
Discusión
El descortezador de los pinos de montaña (Dendroctonus adjunctus) se distribuye en el centro de México sobre los principales sistemas montañosos (García-Navarrete et al., 2021), donde su impacto es mayor (Pérez-Miranda et al., 2021). La presencia de los escarabajos descortezadores de montaña está directamente relacionada con el establecimiento de muérdagos enanos; Endara et al. (2023) reportaron que más de 50% de los árboles enfermos en el Monte Tláloc y Nevado de Toluca presentan la sinergia de ambos patógenos. En el presente estudio, se reporta que 14% de la superficie del bosque de pino (Pinus hartwegii) está afectado por Arceuthobium vaginatum y 7% por D. adjunctus. En términos de superficie, el Nevado de Colima es la única montaña donde se pudo cuantificar la superficie infestada por descortezadores debido a la afectación en parches pequeños de bosques densos de arbolado adulto, esto parece favorecer su dispersión gradual hacia los árboles sanos en torno a los sitios de arbolado muerto en pie con brotes activos hacia los bordes. Al respecto, Cuellar et al. (2013) y Ruiz-González et al. (2018), reportan que, a mayor densidad de arbolado, aumenta el riesgo de la infestación por descortezadores del género Dendroctonus; esto concuerda con lo encontrado en este estudio, ya que los bosques densos son los que presentan mayor cantidad de arbolado muerto por descortezador. En contraste, Sosa et al. (2018) señalan que las altas densidades del arbolado no parecen ser los sitios favoritos para el escarabajo, aunque esto pueda deberse a que dicho estudio no se realizó en sitios de condición de infestación masiva.
Figura 3. Distribución espacial de muérdago enano y descortezador. Fuente: Endara et al. (2023) y trabajo de campo.
Lo anterior, aunado a que la presencia de Dendroctonus, es mayor en diámetros más grandes (Vázquez-Ochoa et al., 2022), provoca la reducción del número de árboles semilleros y, por lo tanto, puede comprometer el establecimiento de la regeneración natural. Por un lado, el descortezador está causando mortalidad en arbolado adulto (muchos de ellos semilleros), por el otro, el muérdago está atacando arbolado juvenil, esta combinación crea condiciones que puede contribuir a una mayor vulnerabilidad de los bosques (Aguirre et al., 2024), además de afectar en gran medida al establecimiento de la regeneración natural, ya que se ha registrado que la infección de muérdago enano reduce el éxito reproductivo del árbol y, a su vez, disminuye el reclutamiento (Queijeiro-Bolaños y Cano-Santana, 2015).
Dado que la homogeneidad en estructura y edad de los bosques son determinantes para la incidencia de insectos descortezadores (Raffa et al., 2008), la afectación de arbolado adulto y juvenil altera la distribución de la estructura vertical del bosque y ocasiona la desaparición del dosel dominante, ésto incrementa la vulnerabilidad de estos bosques ante incendios forestales y estrés de calor (Quintero-Gradilla et al., 2019), ya que no cuentan con un estrato superior bien definido, que cumple la función de dar sombra a los estratos medio e inferior. Además, dicho estrato protege a los árboles jóvenes de eventos climáticos como huracanes con rachas de viento de alta intensidad.
El clima es otra variable importante que limita la dispersión de los descortezadores, principalmente en la temporada de invierno (García-Navarrete et al., 2021). Sin embargo, las temperaturas en las zonas altas del país están incrementando (Sáenz et al., 2020), dados estos escenarios climáticos futuros, D. adjunctus probablemente alcanzará sitios altitudinales superiores (Estrada-Contreras et al., 2022). En el presente estudio, el escarabajo descortezador alcanza 3,900 m snm, mientras que, Salinas-Moreno et al. (2010) reportaron a D. adjunctus en intervalo de 3,100 y 3,500 m snm. Al mismo tiempo, el hospedero también migrará altitudinalmente, pues las zonas bajas del parque prácticamente carecen de arbolado adulto del pino de las alturas, esto probablemente debido a que el incremento actual del clima está reduciendo el vigor de P. hartwegii en las partes bajas, provocada por las altas temperaturas (Gallardo-Salazar et al., 2023).
Figura 4. Efecto de Arceuthobium vaginatum y Dendroctonus adjunctus sobre la estructura de Pinus hartwegii. Arriba, sitios sin muérdago descortezador; abajo sitios atacados por ambos patógenos.Figura 5. Estimación del estrato arbóreo en Pinus hartwegii (estructura vertical).
Las poblaciones del insecto descortezador han aumentado en años recientes, especialmente en la zona norte y centro de su distribución (Vázquez-Ochoa et al., 2022), además, los mayores brotes de descortezadores en México han ocurrido en el Eje Neovolcánico Transversal (del-Val y Sáenz-Romero, 2017). En este sentido y dado que, D. adjunctus presenta un comportamiento agresivo de colonización (Armendáriz-Toledano et al., 2018), produce alta mortalidad de pino, por lo que es necesario incorporar medidas de prevención para reducir sus afectaciones, principalmente sobre arbolado semillero y juvenil, y así disminuir los daños y cambios en la dinámica y estructura de los bosques.
El bosque de Pinus hartwegii del Parque Nacional Volcán Nevado de Colima está siendo afectado por Arceuthobium vaginatum y Dendroctonus adjunctus. La ladera oeste en torno al volcán Nevado de Colima es la más afectada, por un lado, el descortezador se distribuye en los bosques densos de arbolado adulto entre 3,600 y 3,800 m snm; mientras que el muérdago se establece en bosques fragmentados de arbolado juvenil entre 3,500 y 3,600 m snm, con pendientes ligeras y mayor exposición al sol. En ambos casos, los niveles de infestación afectan la estructura y comprometen la regeneración natural de P. hartwegii.
El 20% de la superficie muestreada presenta el establecimiento de uno u otro patógeno, por lo que, es necesario dar seguimiento a la dinámica de estas poblaciones, ya que pueden alterar el equilibrio natural de este ecosistema, principalmente en términos de estructura y regeneración del bosque.
Figura 6. Representación gráfica de la estructura vertical de poblaciones de Pinus hartwegii.
Agradecimientos
A los directivos del Parque Nacional Nevado de Colima y a la Conanp por su aprobación para la realización del estudio. Un agradecimiento especial al Grupo de Alta Montaña, quienes nos apoyaron en el trabajo de campo. Este estudio fue financiado por el proyecto Conacyt-Conafor/A3-S-130105.
Referencias
Aguirre, Z. J., Heredia, B. R., Franco, M. S, Fredericksen, T. S. y Endara, A. A. (2014). Occurrence and effect of dwarf mistletoe (Arceuthobium globosum and A. vaginatum) in high-elevation forests in México. Trees, Forests and People, 18, 100706. https://doi.org/10.1016/j.tfp.2024.100706.
Armendáriz-Toledano, F. y Zúñiga, G. (2017). Illustrated key to species of genus Dendroctonus (Coleoptera: Curculionidae) occurring in Mexico and Central America. Journal of Insect Science, 17, 34. https://doi.org/10.1093/jisesa/iex009
Armendáriz-Toledano, F., Zúñiga, G., García-Román, J., Valerio-Mendoza, O. y García-Navarrete, P. G. (2018). Guía ilustrada para identificar a las especies del género Dendroctonus presentes en México y Centroamérica. Ciudad de México: Red Temática de Salud Forestal/ Consejo Nacional de Ciencia y Tecnología/ Instituto Politécnico Nacional.
Billings, R. F. y Espino, V. (2005). El gorgojo descortezador del pino (Dendroctonus frontalis) en Centroamérica: cómo reconocer, prevenir y controlar plagas. Texas, USA: Texas Forest Service Publication.
Chávez-Salcedo, L. F., Queijeiro-Bolaños, M. E., López-Gómez, V., Cano-Santana, Z., Mejía-Recamier, B. E. y Mojica-Guzmán, A. (2018). Contrasting arthropod communities associated with dwarf mistletoes Arceuthobium globosum and A. vaginatum and their host Pinus hartwegii. Journal of Forestry Research, 29, 1351–1364. https://doi.org/10.1007/s11676-017-0544-y
Christiansen, E. y Bakke, A. (1988). The spruce bark beetle of Eurasia. En A. Berryman (Ed.), Dynamics of forest insect populations: patterns, causes, implications (pp. 479–503). Boston, MA: Springer US. https://doi.org/10.1007/978-1-4899-0789-9_23
Cibrián-Tovar, D., Méndez-Montiel, J. T., Campos-Bolaños, R., Tates III, H. O. y Flores-Lara, J. E. (1995). Insectos forestales de México. México D.F.: Universidad Autónoma Chapingo.
Collins, B. J., Rhoades, C. C., Hubbard, R. M. y Battaglia, M. A. (2011). Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine stands, Forest Ecology and Management, 261, 2168–2175. https://doi.org/10.1016/j.foreco.2011.03.016
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2006). Programa de Conservación y Manejo Parque Nacional Volcán Nevado de Colima. México: Comisión Nacional de Áreas Naturales Protegidas.
Cuéllar-Rodríguez, G., Equihua-Martínez, A., Villa-Castillo, J., Estrada-Venegas, E. G., Méndez-Montiel, T. y Romero-Nápoles, J. (2013). Análisis espacio-temporal de los bosques de Pinus cembroides Zucc. atacados por Dendroctonus mexicanus Hopkins. Revista Mexicana de Ciencias Forestales, 4, 42–49. https://doi.org/10.29298/rmcf.v4i17.419
Dauber, E. (1995). Guía práctica y teórica para el diseño de un inventario forestal de reconocimiento. Santa Cruz. BOLFOR.
del-Val, E. y Sáenz-Romero, C. (2017). Insectos descortezadores (Coleoptera: Curculionidae) y cambio climático: problemática actual y perspectivas en los bosques templados. TIP. Revista Especializada en Ciencias Químico-Biológicas, 20, 53–60. https://doi.org/10.1016/j.recqb.2017.04.006
DOF (Diario Oficial de la Federación). (2018). NORMA OFICIAL MEXICANA NOM-019-SEMARNAT-2017 lineamientos técnicos para la prevención, combate y control de insectos descortezadores. México D.F.
Endara, A. R., Calderón-Contreras, R., Nava-Bernal, G. y Franco, S. (2013). Analysis of fragmentation processes in high-mountain forests of the Centre of Mexico. American Journal of Plant Sciences, 4, 697–704. https://doi.org/10.4236/ajps.2013.43A088
Endara, A. R., Heredia, R. L., García, L. A., Luna, A. A. y Aguirre, J. J. (2023). Distribución espacial del descortezador Dendroctonus adjunctus Blandford, 1897 (Coleoptera: Curculionidae, Scolytinae) en dos bosques de alta montaña del centro de México. Acta Zoológica Mexicana, 39, e3911. https://doi.org/10.21829/azm.2023.3912569
Endara-Agramont, A. R., Heredia-Bodadilla R. L., García-Almaraz, L. A., Luna-Gil, A. A., Franco-Mass, S. y Cibrián-Llanderal, V. D. (2022). Factores asociados con la distribución espacial de muérdagos enanos en dos poblaciones de Pinus hartwegii del centro de México. Revista Mexicana de Biodiversidad, 93, e935008. https://doi.org/10.22201/ib.20078706e.2022.93.5008
Estrada-Contreras, I., Ruiz-Montiel, C., Ibarra-Zavaleta, S. P., Sánchez-Velásquez, L. R., Hoyos-Rivera, G. J., Cristóbal-Salas, A. et al. (2022). Predicting distribution overlaps between Dendroctonus adjunctus Blandford 1897 and six Pinus species in Mexico under global climate change. Canadian Journal of Forest Research, 52, 1201–1211. https://doi.org/10.1139/cjfr-2022-0022
Farjon, A., de la Rosa, J. A. P. y Styles, B. T. (1997). A field guide to the pines of Mexico and Central America. Kew, Reino Unido: Royal Botanic Gardens.
Farrell, B. D., Sequerira, A. S., O’Meara, B. C., Normark, B. B., Chung, J. H. y Jordal, B. H. (2001). The evolution of agriculture in beetles (Curculionidae Scolytinae and Platypodinae). Evolution, 55, 2011–2027. https://doi.org/10.1111/j.0014-3820.2001.tb01318.x
Ferrenberg, S. (2020). Dwarf mistletoe infection interacts with tree growth rate to produce opposing direct and indirect effects on resin duct defenses in lodgepole pine. Forests, 11, 222. https://doi.org/10.3390/f11020222
Gallardo-Salazar, J. L., Lindig-Cisneros, R. A., López-Toledo, L., Endara-Agramont, A. R., Blanco-García, A. y Sáenz-Romero, C. (2023). Analysis of the vigor of Pinus hartwegii Lindl. along an altitudinal gradient using uav multispectral images: evidence of forest decline possibly associated with climatic change. Forests, 14, 1176. https://doi.org/10.3390/f14061176
García-Navarrete, P. G., Soria-Ortiz, G. J. y González-Salazar, C. (2021). Interacciones potenciales parásito-hospedero entre el escarabajo Dendroctonus (Coleoptera: Scolytidae) y Pinus (Pinaceae) en México. Revista de Biología Tropical, 69, 1004–1022. https://doi.org/10.15517/rbt.v69i3.45910
Gernandt, D. S. y Pérez-de la Rosa, J. A. (2014). Biodiversidad de Pinophyta (coníferas) en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), S126–S133. https://doi.org/10.7550/rmb.32195
Hawksworth, F. (1983). Mistletoes as forest parasites. En M. Calder y P. Bernhardt (Eds.), The biology of mistletoes (pp. 320–329). Nueva York: Academic Press.
Klutsch, J. G., Beam, R. D., Jacobi, W. R. y Negrón, J. F. (2014). Bark beetles and dwarf mistletoe interact to alter downed woody material, canopy structure, and stand characteristics in northern Colorado ponderosa pine, Forest Ecology and Management, 315, 63–71. https://doi.org/10.1016/j.foreco.2013.12.024
Lanier, G. N., Hendrichs, J. P. y Flores, J. E. (1988). Biosystematics of the Dendroctonus frontalis (Coleoptera: Scolytidae) complex. Annals of the Entomological Society of America, 81, 403–418. https://doi.org/10.1093/aesa/81.3.403
Lombardero, M. J., Ayres, M. P., Lorio Jr., P. L. y Ruel, J. J. (2002). Environmental effects on constitutive and inducible resin defenses of Pinus taeda. Ecology Letters, 3, 329–339. https://doi.org/10.1046/j.1461-0248.2000.00163.x
Macías, S. J., Domínguez, A. N., López, J. C. y Mérida, R. A. (2004). Monitoreo de descortezadores y sus depredadores mediante el uso de semioquímicos. Manual operativo. Tapachula, Chiapas, México: ECOSUR/ Conafor/ Comisión Nacional de Áreas Naturales Protegidas/ USDA Forest Service.
Martínez, S. M., Madrigal, H. S., Vázquez, C. I., Velasco, B. E., Morales, N. C. y Villareal, G. F. (2015). Efecto de Arceuthobium vaginatum (Willd.) Presl. subsp. vaginatum en Pinus hartwegii Lindl. en Colima. Revista Mexicana de Ciencias Forestales, 6, 44–55. https://doi.org/10.29298/rmcf.v6i29.215
Martínez-Martínez, N., Ramírez-Dávila, J. F., Lara-Vázquez, F. y Figueroa-Figueroa, D. K. (2021). Distribución espacial de muérdago enano en la Reserva de la Biosfera Mariposa Monarca. Colombia Forestal, 22, 65–81. https://doi.org/10.14483/2256201X.17163
Mathiasen, R. L. (2019). Susceptibility of red fir and white fir to fir dwarf mistletoe (Arceuthobium abietiunum) in California. Forest Pathology, 2019, e12516. https://doi.org/10.1111/efp.12516
Mayer, H. y Ott, E. (1991). Gebirgswaldabau-Schutzwaldpflege: ein waldbauulicher Beitrag zur Landschaftsokologie und zum Umweltschutz (Silviculture in mountain forest-management of protection forest: a silvicultural contribution to landscape ecology and environmental protection). 2a Ed. Gustav Fischer, Stuttgart.
Pérez-Miranda, R., González-Hernández, A., Velasco-Bautista, E., Romero-Sánchez, M. E., Arriola-Padilla, V. J., Acosta-Mireles, M. et al. (2021). Análisis temporal de la distribución de Dendroctonus mexicanus Hopkins (1905) en México (2009-2018). Revista Mexicana de Ciencias Forestales, 12, 27–55. https://doi.org/10.29298/rmcf.v12i67.1079
Perusquía, O. J. (1978). The pine bark beetles, Dendroctonus spp.: taxonomy and distribution. Boletín Técnico Núm. 55. México D.F.: Instituto de Investigaciones Forestales, Secretaría de Agricultura y Recursos Hidráulicos.
Queijeiro-Bolaños, M. E. y Cano-Santana, Z. (2015). Dinámica temporal de la infestación por muérdago enano (Arceuthobium globosum y A. vaginatum) en Zoquiapan (Parque Nacional Iztaccíhuatl Popocatépetl), México. CienciaUAT, 9, 6–14. https://doi.org/10.29059/cienciauat.v9i2.705
Queijeiro-Bolaños, M., Cano-Santana, Z. y García-Guzmán, G. (2014). Incidence, severity, and aggregation patterns of two sympatric dwarf mistletoe species (Arceuthobium spp.) in Central Mexico. European Journal of Forest Research, 133, 297–306. https://doi.org/10.1007/s10342-013-0762-6
Quintero-Gradilla, S. D., Jardel-Peláez, E. J., Cuevas-Guzmán, R., García-Oliva, F. y Martínez-Yrizar, A. (2019). Cambio postincendio en la estructura y composición del estrato arbóreo y carga de combustibles en un bosque de Pinus douglasiana de México. Madera y Bosques, 25, 1–14. https://doi.org/10.21829/myb.2019.2531888
Raffa, K. F., Aukema, B. H., Bentz, B. J., Carroll, A. L., Hicke, J. A., Turner, M. G. et al. (2008). Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. BioScience, 58, 501–517. https://doi.org/10.1641/B580607
Rojas-García, F., Fredericksen, T.S., Vazquez-Lozada, S. y Endara-Agramont, A. R. (2019). Impact of timber harvesting on carbon storage in montane forests of central Mexico. New Forests, 50, 1043–1061. https://doi.org/10.1007/s11056-019-09714-z
Ruiz, M. M. A. (1994). Estudio taxonómico de los muérdagos(Loranthaceae)del Nevado de Colima y Volcán de Fuego, Jalisco-Colima, México(Tesis). División de Ciencias Biológicas y Ambientales, CUCBA. Universidad de Guadalajara, México.
Ruiz-González, C. G., Méndez-González, J., Cambrón-Sandoval, H., García-Aranda, A., Montoya-Jiménez, C. y Sosa-Díaz, L. (2018). Distribución altitudinal y estacional de Dendroctonus adjunctus Blandford y Dendroctonus brevicomis Leconte en Coahuila, México. Revista Fitotec- nia Mexicana, 41, 519–526. https://doi.org/10.35196/rfm.2018.4-A.519-526
Rzedowski, J. y Calderón, R. C. (2011). Flora del bajío y de regiones adyacentes. VISCACEAE, Fascículo 170. Instituto de Ecología, A.C. Centro Regional del Bajío, Pátzcuaro, Mich.
Sáenz-Romero, C., Mendoza-Maya, E., Gómez-Pineda, E., Blanco-García, A., Endara-Agramont, A., Lindig-Cisneros, R. et al. (2020). Recent evidence of Mexican temperate forest decline and the need for ex situ conservation, assisted migration, and translocation of species ensembles as adaptive management to face projected climatic change impacts in a megadiverse country. Canadian Journal of Forest Research, 50, 843–854. https://doi.org/10.1139/cjfr-2019-0329
Salinas-Moreno, Y., Mendoza, G. M., Barrios, M. A., Cisneros, R., Macías-Sámano, J. y Zúñiga, G. (2004). Areography of the genus Dendroctonus (Coleoptera: Curculionidae: Scolytinae) in Mexico. Journal of Biogeography, 31, 1163–1177. https://doi.org/10.1111/j.1365-2699.2004.01110.x
Salinas-Moreno, Y., Vargas, C. F., Zúñiga, G. J., Ager, A. y Hayes, J. L. (2010) Atlas de distribución geográfica de los descortezadores del género Dendroctonus (Curculionidae: Scolytinae) en México. Ciudad de México: Instituto Politécnico Nacional/ Comisión Nacional Forestal.
Sánchez-González, A. (2008). Una visión actual de la diversidad y distribución de los pinos de México. Madera y Bosques, 14, 107120. https://doi.org/10.21829/myb.2008.1411222
Sibson, R. (1982). A brief description of natural neighbor interpolation. En John Wiley y Sons (Eds.), Interpolating multivariate data (pp. 21–36). Nueva York.
Sivicoff (2022). Programa operativo estatal de sanidad forestal para el estado de Jalisco. Zapopan, Jalisco. Comité Técnico Estatal de Sanidad Forestal Jalisco, Comisión Nacional Forestal.
Sosa, D. L., Méndez, G. J., García, A. M., Cambrón, S. V., Villarreal, Q. J., Ruiz, G. C. et al. (2018). Distribución potencial de barrenadores, defoliadores, descortezadores y muérdagos en bosques de coníferas de México. Revista Mexicana de Ciencias Forestales, 9, 187–208. https://doi.org/10.29298/rmcf.v9i47.159
Styles, B. T. (1993). Genus Pinus: a Mexican purview. (pp. 397-420). En T. P. Ramamoorthy y A. Lot (Eds.), Biological diversity of Mexico: origins and distribution (pp. 397–420). New York: Oxford University Press.
Vázquez-Ochoa, M. F., Sánchez-Velásquez, L. F., Hernández-Vargas, G., Ibarra-Zavaleta, S. P., Ruiz-Montiel, C. y Pineda-López, M. (2022). La presencia de Dendroctonus es diferente entre especies de pinos y sus diámetros en la región del Parque Nacional Cofre de Perote, México. Revista Mexicana de Biodiversidad, 93, e934048. https://doi.org/10.22201/ib.20078706e.2022.93.4048
Villaseñor, J. L. (2016). Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidad, 87, 559–902. https://doi.org/10.1016/j.rmb.2016.06.017
Wood, D. L. (1982). The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annual Review of Entomology, 27, 411–446. https://doi.org/10.1146/annurev.en.27.010182.002211
Pseudoregma panicola (Hemiptera: Aphididae), un registro nuevo sobre Arundo donax y su potencial como plaga urbana en México
Ana Lilia Muñoz-Viveros a, Damaris Arce-Lara a, Rebeca Peña-Martínez b, Alfonso David Ríos-Pérez c, Álvaro de Obeso-Fernández del Valle c, Juan Manuel Vanegas-Rico a, *
a Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Iztacala, Unidad de Morfología y Función, Laboratorio de Control de Plagas, Av. de los Barrios No. 1, Los Reyes Iztacala, 54090 Tlalnepantla de Baz, Estado de México, Mexico
b Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas, Prolongación de Carpio y Plan de Ayala, Sto. Tomás, 11340 Ciudad de México, Mexico
c Tecnológico de Monterrey, Departamento de Bioingeniería, Escuela de Ingeniería y Ciencias, Av. Eugenio Garza Sada 2501, 64849 Monterrey, Nuevo León, Mexico
Received: 30 April 2024; accepted: 28 February 2025
Abstract
Aphids are phytophagous insects that can affect the health of wild and urban vegetation, their effect being enhanced when they are exotic. The study was conducted in the garden of the Vasconcelos library, Mexico City. Insects were collected on ornamental Poaceae during December 2019, February 2020, and continued post-pandemic in 2023 (May-August 2023). Organisms were quantified and searched for natural enemies; morphological and molecular analyses of insects and plants were carried out. The aphid was determined as Pseudoregma panicola and Arundo donax as its host and first world report. Pseudoregma panicola and Arundo donax have molecular similarities with populations from Yunnan, China. The review of the scarce material from Mexico shows that P. panicola was collected from 1979 to 1981, on 3 wild species of Poaceae: ca. Phragmites in Morelos, Paspalum sp. and another undetermined species in Tamaulipas. The entomophagous present were: scarce larvae of Chamaemyiidae, and adults of the coccinellids Harmonia axyridis and Hippodamia convergens. The absence of parasitoids during 3 years of research, in addition to the low population of entomophagous, indicates an unfinished process of adaptation to this urban pest.
Keywords: Exotic insects; Poaceae; Chamaemyiidae
Resumen
Los áfidos son insectos fitófagos que pueden afectar la salud de la vegetación silvestre y urbana, cuyo efecto se potencia cuando son exóticos. El estudio se realizó en el jardín de la biblioteca Vasconcelos, Ciudad de México. Se recolectaron insectos sobre Poaceae ornamentales durante diciembre 2019, febrero 2020 y se continuó postpandemia en 2023 (mayo a agosto 2023). Se cuantificaron los organismos y se buscaron enemigos naturales; se realizaron análisis morfológicos y moleculares de los insectos y la planta. Se determinó el áfido como Pseudoregmapanicola y Arundo donax como su hospedante, lo cual constituye el primer reporte mundial. Pseudoregma panicola y Arundo donax tienen cercanía molecular con poblaciones de Yunnan, China. La revisión del escaso material de colección en México muestra que P. panicola se recolectó de 1979 a 1981, sobre 3 especies silvestres de Poaceae:ca. Phragmites en Morelos, Paspalum sp. y otra especie sin determinar en Tamaulipas. Los entomófagos presentes fueron escasas larvas de Chamaemyiidae y adultos de los coccinélidos Harmonia axyridis e Hippodamia convergens. La ausencia de parasitoides durante 3 años de investigación, además de la baja población de entomófagos, indica un proceso inconcluso de adaptación de esta plaga urbana.
Aphids are among the most significant pest groups affecting urban vegetation (Tello et al., 2005). They damage a wide range of host plants by sucking the juices from leaves and stems and producing honeydew. The damage is evident through symptoms such as discolored, battered, and yellowed leaves, as well as stunted growth, which detracts from the aesthetics of green areas and could represent a disease reservoir (Peña-Martínez, 1992; Rozo-Lopez et al., 2023; Tubby & Webber, 2010). Phytosanitary inspections are crucial in urban areas to ensure healthy vegetation, which, in turn, contributes to thermal comfort and indirectly reduces the risks of diseases (Tochaiwat et al., 2023; Yang et al., 2023).
In Mexico City, one of the largest cities in the world, commercial and tourist activities may facilitate the introduction of insect pests. The incursion of exotic aphids to Mexico can lead to severe economic impacts, like those generated by M. sorghi (also cited as M. sacchari) in sorghum crops during 2015 and 2016 (Peña-Martinez et al., 2016; Rodríguez-del Bosque & Terán, 2015). Therefore, it is essential to promptly document the presence of these exotic phytophagous insects and their hosts to mitigate potential damage to agricultural activities (Barczak et al., 2021; Korányi et al., 2021). Consequently, this study aims to report the presence of the aphid Pseudoregma panicola in ornamental plants in Mexico City.
Materials and methods
The sampling site was the garden of the Vasconcelos Public Library, located in Mexico City (19°26’53.5” N, 99°09’01.8” W; 2,241 m asl). This garden was created in 2006 to refurbish the city’s train station and incorporates 168 plant species from different sites (garden staff, personal communication).
The sample period spans from December 2019 to February 2020, when we observed and collected pest insects weekly. Following the COVID-19 pandemic, sampling was restarted in January 2023 to follow up on maintenance staff observations about the pest on Poaceae specimens used as ornamentals. The fieldwork at each visit consisted of 2 steps: photographing leaves (adaxial and abaxial side) with aphid colonies, including a metal ruler as a size reference, and sampling of these leaves, preserving them in airtight plastic bags for subsequent processing (rearing, mounting, and identifying natural enemies) in the Laboratorio de Control de Plagas de la Facultad de Estudios Superiores, Iztacala, UNAM. At the same time, another part of the plant material was provided to the herbarium of the same institution for identification.
The photographs were edited with Gimp software ver. 2.10.32 (The GIMP Development Team, 2023) to obtain tricolor images (green = leaf area, red = aphid specimens, white = background), including a centimeter scale. Then, images were processed with Image J software ver. 11 (Rasband, 2018) to estimate the area covered by the aphids on the adaxial and abaxial sides of the leaves. The abundance was calculated and compared with the M. sorghi infestation counting scale proposed for sorghum leaves (Bowling et al., 2015), using the original photos and Image J. Means and standard errors were calculated using SPSS software ver. 24.0 (IBM, 2016).
Some aphid specimens were processed according to the technique described by Blackman and Eastop (1994) and were identified according to the Noordam keys (1991). Part of the specimen material was preserved in 96% ethanol and kept refrigerated. Later, it was sent to the Laboratorio de Estrategias Ambientales y Fitotecnologías, of the Departmento de Bioingeniería at the Tecnológico de Monterrey for DNA extraction.
Molecular analyses were performed using cytochrome c oxidase I (COI) as suggested by several publications for aphids (Corsini et al., 1999; Folmer et al., 1994; Nováková et al., 2013; Zhang et al., 2011). A PCR reaction to amplify a COI fragment was performed using the primers LCOI490 and HCO2198 (Folmer et al., 1994). The reaction was performed using a denaturing step of 95° for 3 min, followed by 35 cycles of 95° for 1 min, 45° for 1.5 min, and 72° for 1 min, and finishing with an extension of 3 min at 72°. The amplicon was purified using a QIAGEN PCR Purification Kit following the manufacturer’s instructions. Finally, the amplicon was sequenced using Sanger sequencing with the same primers, and the sequence was uploaded to NCBI with the accession number PQ196704. Once the sequence was obtained, a BLAST was performed to identify the organism (Altschul et al., 1990). Similar sequences were downloaded, cropped, and aligned (muscle algorithm) using MEGA XI when the organisms were identified. A phylogenetic tree was created using maximum likelihood (Tamura et al., 2021).
An ecological niche model was made with the field records in Mexico (by the authors) and from the Global Biodiversity Information Facility (GBIF.org, 2024) in Darwin Core format. The obtained data were subjected to 2 debugging processes: elimination of those records with coordinates 0 and 0, and elimination of duplicate data. Climatological variables were downloaded from WorldClim at a resolution of 5 arc minutes (≈ 10 km2) (Fick & Hijmans, 2017). The variables combining temperature and precipitation information, Bio8 – Mean temperature of the wettest quarter, Bio9 – Mean temperature of the driest quarter, Bio18 – Precipitation of the warmest quarter, and Bio19 – Precipitation of the coldest quarter, were removed due to the spatial anomalies they generate (Escobar et al., 2014).
The remaining 15 variables were used globally (except Antarctica) to avoid the risk of niche truncation (Chevalier et al., 2022). The variables were entered into the Niche A software (Qiao et al., 2016), and a principal component analysis (PCA) was performed. The 15 components were used to conduct ecological niche modeling based on the calibration zone and a 0.5-degree buffer (M, of the BAM model, Soberón et al., 2017). Using the ntbox package, more than 400 possible models were built within R, of which the one with the best omission and commission rates was kept (Osorio-Olvera et al., 2020). The resulting model was subjected to the MOP (Mobility-Oriented Parity) validation test, and only those regions that showed a low risk of extrapolation (near 1) were used (Owens et al., 2013).
Results
According to taxonomic identification and molecular test results, the specimens corresponded to Pseudoregma panicola (Takahashi) (Fig. 1). The host plant was determined by Izta Herbarium as Arundo donax L. The population density of P. panicola on this plant was estimated at 255 ± 65.4 individuals (range 75-750) per colony/leaf, with a more significant presence on the abaxial side, representing 46% of its cover. The leading group of associated entomophagous insects were silver fly larvae (Diptera: Chamaemyiidae), with an average abundance of 2.4 ± 0.8 (range 1-5) individuals/leaf over 3 years, besides a few adults of coccinellids (Coleoptera: Coccinellidae), Hippodamia convergens L., and Harmonia axyridis Pallas (0.12 ± 0.01 individuals per leaf/year). The population density of all predators was low compared to the pests. In the case of coccinellids, they were occasional, and immatures were not obtained.
Molecular sequence analysis indicated that the aphid P. panicola and its host A. donax, are most closely related to those sequenced from South Africa, Egypt, India, Italy, USA, and China (Fig. 2). Specimens of P. panicola from the latter country are most closely related to those sequenced from Poaceae in Yunnan Province, China, and, to a lesser extent, to specimens collected from Phyllostachys nigra Lodd. former Lindl. In the same region, the Fujian Province (Rui-Ling et al., 2013) (Fig. 3).
The model was only built with 60 occurrence points, of which 3 were obtained by the authors. The MOP achieved acceptable accuracy and a low risk of extrapolation values for Mexico, despite the fact that only 63 occurrence points were established (Fig. 4). For Mexico, regions such as the Yucatán Peninsula, the Valley of Mexico and northern and central Mexico seemed the most suitable to P. panicola distribution.
Discussion
The aphid P. panicola is reported in 4 continents: Africa —Argelia (Samia y Benoufella-Kitous, 2021), Asia —China (Holman, 2009; Riu-Ling, 2013), Japan (Holman, 2009), India (Holman, 2009; Singh & Singh 2018), Indonesia (Noordam, 1991), Nepal (Holman, 2009), and south of the Asian continent (Brumley, 2020), Oceania —Australia (Brumley, 2020) and New Zealand (Cottier, 1953), and America —Cuba, Costa Rica, Guadalupe, Puerto Rico, and Venezuela (Villalobos-Muller et al., 2010).
In Mexico, this species was initially reported by Peña-Martínez (1985), without specifying the host plant or location. Unedited documentary information from the Aphidomorpha collection of Mexico indicates that the aphid was collected by J. Butze at Coatlán del Río, Morelos, in 1979, on a host tentatively identified as Phragmites. The insects were identified by Dr. Remaudière in 1981, and the slide was subsequently donated to the former Dirección General de Sanidad Vegetal, today named Senasica. There are 2 unique slides in the Aphidomorpha collection: the first was surveyed from undetermined Poaceae on September 2nd, 1981, and the second on Paspalum sp. on September 4th, 1981; both collected by J. Holman and R. Peña-Martínez from Reserva de la Biosfera El Cielo, Gómez Farías, Tamaulipas, and corroborated in 2024 by the aphid specialists Ana Lilia Muñoz Viveros and Rebeca Peña Martínez.
Figure 1. Colonies and structures of Pseudoregma panicola. A, Leaf of Arundo donax infested by P. panicola colonies; B, live adults and nymph waxy covered, 1 mm scale; C, first instar nymph, 0.5 mm scale; D, habitus of aptera, 0.5 mm scale; E, antenna, 0.1 mm scale; F, frontal horns, 0.1 mm; G, ana plate bilobed, 0.1 mm scale; H, rostrum, 0.1 mm scale; I, cauda knobbed, 0.1 mm scale; J, K, sifunculus, lateral view and frontal view, 0.1 mm scale.Figure 2. Comparative dendrogram of Hormaphidinae aphids and comparative dendrogram of Arundo donax sequences, registered in Genbank and specimens from the study site.Figure 3. Mobility-oriented parity (MOP) values of the worldwide ecological niche model of Pseudoregma panicola.
Regarding P. panicola feeding, different genera of Poaceae were recorded as hosts: Andropogon, Arthraxon, Bambusa, Capillipedium, Cyrtococcum, Eragrostis, Ichnanthus, Indocalamus, Lasiacis, Oplismenus, Panicum, Paspalum, Phragmites, Phyllostachys, Pseudoechinolaena, Setaria, Stenthotheca, and Thysanolaena (Favret & Miller, 2012; Holman, 2009; Rui-Ling et al., 2013; Singh & Singh, 2018). Arundo donax is a new record of the host, which develops as secondary vegetation in the country, even as an ornamental, in various urban areas of Mexico City.
In Vasconcelos Library, A. donax occupies 80 m2 of the 2,600 m2 total garden area and is placed as a green barrier to hide the walls and give a feeling of thickness. The origin of purchase of this plant is uncertain; furthermore, no aphids or any other phytophagous have been recorded that have affected it since its placement (garden staff, personal communication). This green area is located near downtown Mexico City, where 3 public transportation systems converge, providing connections to the south, north, and east of the city, as well as to the airport, potentially facilitating the introduction of pest insects. Records on Phyllostachys nigra in Mexico indicate its presence in Nuevo León, Sinaloa, and San Luis Potosí (iNaturalist, 2024). Additionally, there are a few reports on the American continent (GBIF 2023), which also does not correspond to the continental distribution of P. panicola, suggesting that this plant is not the cause of its dispersal in America. A possible approach to discern the dispersal of P. panicola in Mexico would be the trade from Yunnan and other provinces of China to Central America and the USA, where Mexico is part of the naval route, and where plants with roots and foliage are sold for floral arrangements (Economic Complexity Observatory, 2023).
According to the Bowling’s scale infestation per leaf, the abundance of P. panicola on A. donax (up to 750 aphids) is higher than M. donacis (range 12 to 500 aphids) on the same host in Mexico (Vanegas-Rico et al., 2023), and M. sorghi on Sorghum (≈ 20-136 aphids) in Texas (Brewer et al., 2022). The low diversity of predators and the absence of parasitoids on P. panicola suggests that the potential natural regulators were in the process of adaptation during that period. Regarding exotic aphid populations, M. sorghi was detected in 2015, and more recently, M. donacis on A. donax in 2023. In the case of M. sorghi, at least 19 entomophagous (including 2 parasitoids) were recorded in northeastern Mexico sometime after its first report in the country (Rodríguez-del Bosque et al., 2018). The effect of natural enemies in Texas, 14 predators and 2 parasitoids, shows low abundance after 3-4 years of population studies (Brewer et al., 2022) compared to the initial report as a pest, where more than 1,000 aphids per leaf were observed (Bowling et al., 2015).
Figure 4. Ecological niche model of Pseudoregma panicola. The color bar depicts the environmental suitability of the species worldwide and Mexico.
In the case of M. donacis, coccinellids and silver flies were observed preying on aphids during their first report in Mexico (Vanegas-Rico et al., 2023). Recently, laboratory observations indicated that ladybugs and silver flies could consume all instars of M. donacis; in fact, a low percent of parasitism (˂ 5%) was observed in the Valley of Mexico and Ensenada, Baja California (pers. com. Vanegas-Rico).
The identity of silver flies remains unknown due to limited material and inconclusive molecular results. This Diptera group can feed on different species of adelgids, aphids, soft scales, and other hemipterans (Gaimari et al., 2024; Salas-Monzón et al., 2020; Satar et al., 2015; Vanegas-Rico et al., 2010), exhibiting a density-dependence with their prey (Vanegas-Rico et al., 2017). The last bioecological aspect is relevant because it influences the success of biological control programs (Gaimari, 1991; Havill et al., 2018), and has favored the selection of some silver fly species as biological control agents in Europe (Justesen et al., 2023), USA (Ross et al., 2011; Havill et al., 2018), and Israel (Mendel et al., 2020).
According to site management personnel statements, P. panicola is controlled through broad-spectrum insecticide and sanitary pruning. This last method is frequently used when dry, greyish leaves and white spots (aphid wax) are observed. As a result, the aphid population densities recorded since 2023 are low (18.5 ± 4.2) and scarcer in coccinellids (2.6 ± 1.4) and silver flies (0.4 ± 0.1). So far, there was no evidence of parasitism in additional surveys of 2024. In the 2023 and 2024 samples, predation was observed on the aphid colonies and coccinellid eggs, mainly by H. axyridis andsporadic Syphid and Scymninae larvae. This suggests the continued adaptation process of these entomophagous and a possible adverse effect of pest management. Therefore, a biological control strategy for conservation must be implemented to promote the permanence of beneficial organisms and increase their populations.
Arundodonax has been encouraged as an ornamental plant so that the first negative impact would be in this type of urban system. There is still no prospecting in the agricultural landscape, despite the wide distribution of A. donax in 27 states of Mexico, mostly as a riparian system weed (Goolsby et al., 2023). The tendency of the A. donax distribution model (Goolsby et al., 2023) is similar to the current climate modeling of the aphid proposed in this work. This shows the potential for the establishment of the aphid in different states of the republic, highlighting its suitability in the Basin of Mexico and northern and southwestern parts of the country where this plant develops as an invader in bodies of water.
Finally, the work confirms the presence of P. panicola in Mexico and its development as an urban pest of A. donax, representing a New World host record for this aphid, and the first report from North America. Molecular information suggests China as a possible place of origin for both the plant and the aphid. Although this process is not entirely clear, it is feasible that the economic routes for transporting live or forage plants from this Asian country are a significant element to consider for the potential introduction of Poaceae phytophagous insects. The impact on this family of plants is currently unknown, so future studies should focus in that direction.
Acknowledgments
Special thanks to Francisco Amador-Cruz, Professor of Ecology career at FES-Iztacala, for elaborating the maps and for providing suggestions for this manuscript.
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Barczak, T., Bennewicz, J., Korczyński, M., Błażejewicz-Zawadzińska, M., & Piekarska-Boniecka, H. (2021). Aphid assemblages associated with urban park plant communities. Insects, 12, 173. https://doi.org/10.3390/insects12020173
Blackman, R. L., & Eastop, V. F. (1994). Aphid’s of the World’s trees. An identification and information guide. Wallingford, UK: CAB International.
Bowling, R., Brewer, M., Knutson, A., Way, M., Porter, P., Bynum, E. et al. (2015). Scouting sugarcane aphids. Retrieved December 18, 2023 from http://agrilife.org/mid-coast-ipm/files/2015/05/Scouting-Sugarcane-Aphids-2015.pdf
Brewer, M. J., Elliott, N. C., Esquivel, I. L., Jacobson, A. L., Faris, A. M., Szczepaniec, A. et al. (2022). Natural enemies, mediated by landscape and weather conditions, shape response of the sorghum agroecosystem of North America to the invasive aphid Melanaphis sorghi. Frontiers in Insect Science, 2, 830997. https://doi.org/10.3389/finsc. 2022.830997
Brumley, C. (2020). A checklist and host catalogue of the aphids (Hemiptera: Aphididae) held in the Australian National Insect Collection. Zootaxa, 4728, 575–600. https://doi.org/10.11646/zootaxa.4728.4.12
Chevalier, M., Zarzo-Arias, A., Guélat, J., Mateo, R. G., & Guisan, A. (2022). Accounting for niche truncation to improve spatial and temporal predictions of species distributions. Frontiers in Ecology and Evolution, 10, 944116. https://doi.org/10.3389/fevo.2022.944116
Corsini, G., Manubens, A., Lladser, M., Lobos, S., Seelenfreund, D., & Lobos, C. (1999). AFLP analysis of the fruit fly Ceratitis capitata. Focus, 21, 72–73.
Cottier, W. (1953). Aphids of New Zealand. New Zealand Department of Scientific and Industrial Research Bulletin, 186, 1–382.
Economic Complexity Observatory. (2023). Retrieved December 14th, 2023 from: https://oec.world/es/profile/bilateral-country/chn/partner/usa?depthSelector=HS6Depth&dynamicBilateralTradeSelector=year2018&productSectionSelector=sectionID2#bi-trade-products
Escobar, L. E., Lira-Noriega, A., Medina-Vogel, G., & Peterson, A. T. (2014). Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: use of Maxent and NicheA to assure strict model transference. Geospatial Health, 9, 221–229. https://doi.org/10.4081/gh.2014.19
Favret, C., & Miller, G. L. (2012). AphID. Identification Technology Program, CPHST, PPQ, APHIS, USDA; Fort Collins, CO. Retrieved on January 8, 2024 from: http://AphID.AphidNet.org/.
Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37, 4302–4315. https://doi.org/10.1002/joc.5086
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
Gaimari, S. D. (1991). Use of silver flies (Diptera: Chamaemyiidae) for biological control of homopterous pests. Proceedings of the Entomological Society of Washington, 53, 947–950.
Gaimari, S. D., González, C. R., & Elgueta, M. (2024). A catalog of the Chamaemyiidae of Chile (Diptera: Lauxanioidea). Zootaxa, 5399, 555–569. https://doi.org/10.11646/zootaxa.5399.5.5
Goolsby, J. A., Moran, P. J., Martínez-Jiménez, M., Yang, C., Canavan, K., Paynter, Q. et al. (2023). Biology of Invasive Plants 4. Arundo donax L. Invasive Plant Science and Management, 16, 81–109. https://doi.org/10.1017/inp.2023.17
GBIF (Global Biodiversity Information Facility). (2023). Phyllostachys nigra. Retrieved September 14th, 2023. From: https://doi.org/10.15468/dl.y5xrtv
GBIF (Global Biodiversity Information Facility). (2024). Pseudoregma panicola. Retrieved August 20th, 2024. From https://doi.org/10.15468/dl.6n5574
Havill, N. P., Gaimari, S. D., & Caccone, A. (2018). Cryptic east-west divergence and molecular diagnostics for two species of silver flies (Diptera: Chamaemyiidae: Leucopis) from North America being evaluated for biological control of hemlock woolly adelgid. Biological Control, 121, 23–29. https://doi.org/10.1016/j.biocontrol.2018.02.004
Holman, J. (2009). Host plant catalog of aphids: Paleartic Region. Dordrecht, The Netherlands: Springer. https://doi.org/10.1007/978-1-4020-8286-3
IBM Corp. (2016). IBM SPSS Statistics for Windows, Version 24.0. Armonk, NY: IBM Corp.
iNaturalist community. (2024). Observaciones de Phyllostachys nigra de México. Retrieved on January 12, 2024. From: https://mexico.inaturalist.org/taxa/166766-Phyllostachys-nigra
Justesen, M. J., Seehausen, M. L., Havill, N. P., Kenis, M., Gaimari, S. D., Matchutadze, I. et al. (2023). Evaluation of Leucopis hennigrata (Diptera: Chamaemyiidae) as a classical biological control agent of Adelges nordmannianae (Hemiptera: Adelgidae) in northern Europe. Biological Control, 183, 105264. https://doi.org/10.1016/j.biocontrol. 2023.105264
Korányi, D., Szigeti, V., Mezőfi, L., Kondorosy, E., & Markó, V. (2021). Urbanization alters the abundance and composition of predator communities and leads to aphid outbreaks on urban trees. Urban Ecosystems, 24, 571–586. https://doi.org/10.1007/s11252-020-01061-8
Mendel, Z., Protasov, A., Vanegas-Rico, J. M., Lomeli-Flores, J. R., Suma, P., & Rodríguez-Leyva, E. (2020). Classical and fortuitous biological control of the prickly pear cochineal, Dactylopius opuntiae, in Israel. Biological Control, 142, 104157. https://doi.org/10.1016/j.biocontrol.2019.104157
Noordam, D. (1991). Hormaphidinae from Java (Homoptera: Aphididae). The Zoologische Verhandelingen, 270, 1–525.
Nováková, E., Hypša, V., Klein, J., Foottit, R. G., von Dohlen, C. D., & Moran, N. A. (2013). Reconstructing the phylogeny of aphids (Hemiptera: Aphididae) using DNA of the obligate symbiont Buchnera aphidicola. Molecular Phylogenetics and Evolution, 68, 42–54. https://doi.org/10.1016/j.ympev.2013.03.016
Osorio-Olvera, L., Lira-Noriega, A., Soberón, J., Peterson, A. T., Falconi, M., Contreras-Díaz, R. G. et al. (2020). ntbox: an R package with graphical user interface for modeling and evaluating multidimensional ecological niches. Methods in Ecology and Evolution, 11, 1199–1206. https://doi.org/10.1111/2041-210x.13452
Owens, H. L., Campbell, L. P., Dornak, L. L., Saupe, E. E., Barve, N., Soberón, J. et al. (2013). Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecological Modelling, 263, 10–18. https://doi.org/10.1016/j.ecolmodel.2013.04.011
Qiao, H., Peterson, A. T., Campbell, L. P., Soberón, J., Ji, L., & Escobar, L. E. (2016). NicheA: creating virtual species and ecological niches in multivariate environmental scenarios. Ecography, 39, 805–813. https://doi.org/10.1111/ecog.01961
Peña-Martínez, R. (1985). Ecological notes on aphids of the high plateau of Mexico with a check-list of species collected in 1980. In Proceedings of the International Aphidological Symposium (pp. 425–430). Jablonna. Polska Akademia Nauk, Warsaw.
Peña-Martínez, R. (1992). Biología de áfidos y su relación con la transmisión de virus. In C. Urias, R. Rodríguez, & T. Alejandre (Eds). Áfidos como vectores de virus en México: contribución a la ecología y control de áfidos en México (pp. 11–35). CEFIT-CP. Vol 1. Montecillo, Estado de México, México.
Peña-Martínez, R., Muñoz-Viveros, A. L., Bujanos-Muniz, R., Luevano-Borroel, J., Tamayo-Mejía, F., & Cortez-Mondaca, E. (2016). Sexual forms of sorghum aphid complex Melanaphis sacchari/sorghi in Mexico. Southwestern Entomologist, 41, 127–132. https://doi.org/10.3958/059.041.0114
Rasband, W. S. (2018). Software ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA. https://imagej.nih.gov/ij/
Rodríguez-del Bosque, L. A., & Terán, A. P. (2015). Melanaphis sacchari (Hemiptera: Aphididae): a new sorghum insect pest in Mexico. Southwestern Entomologist, 40, 433–434. https://doi.org/10.3958/059.040.0217
Rodríguez-del Bosque, L. A., Rodríguez-Vélez, B., Sarmiento-Cordero, M. A., & Arredondo-Bernal, H. C. (2018). Natural enemies of Melanaphis sacchari on Grain Sorghum in Northeastern Mexico. Southwestern Entomologist, 43, 277–279. https://doi.org/10.3958/059.043.0103
Ross, D. W., Gaimari, S. D., Kohler, G. R., Wallin, K. F., & Grubin, S. M. (2011). Chamaemyiid predators of the hemlock woolly adelgid from the Pacific Northwest. In Brad Onken, R. O., & R. Reardon (Eds.), Implementation and status of biological control of the hemlock woolly adelgid (pp. 97–106). USDA Forest Service.
Rozo-Lopez, P., Brewer, W., Käfer, S., Martin, M. M., & Parker B. J. (2023). Untangling an insect’s virome from its endogenous viral elements. BMC Genomics, 24, 636. https://doi.org/10.1186/s12864-023-09737-z
Rui-Ling, Z., Xiao-Lei, H., Li-Yun, J., & Ge-Xia, Q. (2013). Molecular phylogenetic evidence for paraphyly of Ceratovacuna and Pseudoregma (Hemiptera, Hormaphidinae) reveals late Tertiary radiation. Bulletin of Entomological Research, 103, 644–655. https://doi.org/10.1017/S0007485313000321
Salas-Monzón, R., Rodríguez-Leyva, E., Lomeli-Flores, J. R., & Vanegas-Rico, J. M. (2020). How two predators feed on Dactylopius opuntiae beneath its ventral side. Southwestern Entomologist,45, 823–826. https://doi.org/10.3958/059.045.0324
Samia, A. I. T., & Benoufella-Kitous, K. (2021). Diversity of aphids (Hemiptera: Aphididae) associated with potato crop in Tizi-Ouzou (North of Algeria), with new records. Acta Agriculturae Slovenica, 117, 1–9. https://doi.org/10.14720/aas.2021.117.1.1768
Satar, S., Raspi, A., Özdemir, I., Tusun, A., Karacaoğlu, M., & Benelli, G. (2015). Seasonal habits of predation and prey range in aphidophagous silver flies (Diptera, Chamaemyiidae), an overlooked family of biological control agents. Bulletin of Insectology, 68, 173–180.
Singh, G., & Singh, R. (2018). Updated checklist of Indian Hormaphidinae (Aphididae: Hemiptera) and their food plants. Journal of Entomology and Zoology Studies, 6, 1345–1352.
Soberón, J., Osorio-Olvera, L., & Peterson, T. (2017). Diferencias conceptuales entre modelación de nichos y modelación de áreas de distribución. Revista Mexicana de Biodiversidad, 88, 437–441. https://doi.org/10.1016/j.rmb.2017.03.011
Tamura, K., Stecher, G., & Kumar, S. (2021). MEGA11: Molecular Evolutionary Genetics Analysis V.11. Molecular Biology and Evolution, 38, 3022–3027. https://doi.org/10. 1093/molbev/msab120
Tello, M. L., Tomalak, M., Siwecki, R., Gáper, J., Motta, E., & Mateo-Sagasta, E. (2005). Biotic urban growing conditions-threats, pests and diseases. In C. Konijnendijk, K. Nilsson, T. Randrup, & J. Schipperijn (Eds.), Urban forests and trees: a reference book (pp. 325–365). Berlin, Heidelberg: Springer Berlin Heidelberg. https://doi.org/10. 1007/3-540-27684-X_13
The GIMP Development Team. (2023). GNU Image Manipulation Program (GIMP), Version 2.10.32. Community, Free Soft- ware. https://www.gimp.org/
Tochaiwat, K., Phichetkunbodee, N., Suppakittpaisarn, P., Rinchumphu, D., Tepweerakun, S., Kridakorn Na Ayutthaya, T.et al. (2023). Eco-efficiency of green infrastructure on thermal comfort of outdoor space design. Sustainability, 15, 2566. https://doi.org/10.3390/su15032566
Tubby, K. V., & Webber, J. F. (2010). Pests and diseases threatening urban trees under a changing climate. Forestry: An International Journal of Forest Research, 83, 451– 459. https://doi.org/10.1093/forestry/cpq027
Vanegas-Rico, J. M., Lomeli-Flores, J. R., Rodríguez-Leyva, E., Mora-Aguilera, G., & Valdez, J. M. (2010). Enemigos naturales de Dactylopius opuntiae (Cockerell) en Opuntia ficus-indica (L.) Miller en el centro de México. Acta Zoológica Mexicana, 26, 415–433. https://doi.org/10.21829/azm.2010.262718
Vanegas-Rico, J. M., Pérez-Panduro, A., Lomeli-Flores, J. R., Rodríguez-Leyva, E., Valdez-Carrasco, J. M., & Mora-Aguilera, G. (2017). Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae) population fluctuations and predators in Tlalnepantla, Morelos, México. Folia Entomológica Mexicana (nueva serie), 3, 23–31.
Vanegas-Rico, J. M., Peña-Martínez, R., & Muñoz-Viveros, A. L. (2023). First record of Melanaphis donacis (Passerini, 1861) (Hemiptera: Aphididae) in México. Entomological Communications, 5, ec05031. https://doi.org/10.37486/2675-1305.ec05031
Villalobos-Muller, W., Pérez-Hidalgo, N., Mier-Durante, P., & Nieto-Nafría, J. M. (2010). Aphididae (Hemiptera: Sternorrhyncha) from Costa Rica, with new records for Central America. Boletín de la Asociación Española de Entomología, 34, 145–182.
Yang, T., Gu, T., Xu, Z., He, T., Li, G., & Huang, J. (2023). Associations of residential green space with incident type 2 diabetes and the role of air pollution: a prospective analysis in UK Biobank. Science of the Total Environment, 866, 161396. https://doi.org/10.1016/j.scitotenv.2023.161396
Zhang, R., Huang, X., Jiang, L., & Qiao, G. (2011). Phylogeny and species differentiation of Mollitrichosiphum spp. (Aphididae, Greenideinae) based on mitochondrial COI and Cyt b genes. Current Zoology, 57, 806–815. https://doi.org/10.1093/czoolo/57.6.806
Leafhoppers of the tribe Athysanini associated to pine and oak forests in Mexico (Hemiptera: Cicadellidae), with description of a new species of the genus Eutettix
J. Adilson Pinedo-Escatel a, b, *
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Insectos, Tercer Circuito s/n, Ciudad Universitaria, 04510, Ciudad de México, México
b University of Illinois, Prairie Research Institute, Illinois Natural History Survey, 1816 S. Oak Street, 61820 Champaign, Illinois, EUA
*Autor para correspondencia: adilson.pinedo@ib.unam.mx (J.A. Pinedo-Escatel)
Recibido: 19 marzo 2024; aceptado: 28 febrero 2025
Resumen
Históricamente los insectos herbívoros de la tribu de cicadélidos Athysanini (Hemiptera: Cicadellidae) han sido ávidamente recolectados en bosques de pino y encino en México, además de contar con una notable distribución en el país, sin embargo, se desconoce cuáles géneros y especies están asociadas a estos ambientes boscosos. Esta es la primera contribución mediante múltiples eventos de recolección, especímenes revisados en colecciones y literatura especializada en la que se compiló un total de 385 georreferencias para 83 especies de 31 géneros, donde 90% de las especies son endémicas para el territorio mexicano y los datos demuestran presencia sobre 15 estados; Guerrero lidereó tanto en el número de registros como en número de especies. Además, se describe una especie nueva del género Eutettix Van Duzee, 1892, recolectada en el estado de Jalisco y se discute la distribución de las especies de Athysanini presentes sobre las pináceas y encinares mexicanos.
Historically, herbivorous insects of the leafhopper tribe Athysanini (Hemiptera: Cicadellidae) have been avidly collected in pine and oak forests in Mexico and have a remarkable distribution in the country. However, it is unknown which genera and species are associated with these forest environments. In this contribution, through multiple collection events, specimens reviewed in collections and specialized literature, a total of 385 georeferences were compiled for 83 species from 31 genera, where 90% of the species are endemic to the Mexican territory and its occurrences extend over 15 states; Guerrero led both, number of records and number of species. In addition, a new species of the genus Eutettix Van Duzee, 1892, collected in the state of Jalisco, is described and known distribution of the Athysanini species present in Mexican pine and oak forests is discussed.
Los bosques mexicanos cubren una significante extensión del territorio mexicano manteniendo una biota muy diversa y en algunos casos, hasta exclusiva (Rzedowski, 1991a, b, 2006). La cobertura vegetal forestal dentro del país se encuentra interconectada por límites de ecotonos diferenciándose por diversos factores estructurales, entre ellos, la composición de los bosques ya sean de coníferas, encino, tropical caducifolio, mesófilo de montaña o lluvioso (Rzedowski, 1991a, 2006). Algunos de estos últimos, son resilientes en áreas uniformes de vegetación, o en parches disruptivos con superficies fragmentadas, perturbadas o en constante presión ambiental por actividades antropogénicas (Parada-Aragón et al., 2019; Rzedowski, 1991b; Sosa y De-Nova, 2012; Steinmann, 2002). Las pináceas y encinares en México según Gernandt y Pérez-de la Rosa (2014), Tantray et al. (2017) y Valencia-Ávalos (2010) están ampliamente distribuidos desde 32º hasta 14º latitud norte en un rango altitudinal que oscila entre 200 y 3,700 m snm. A lo largo de la topografía mexicana se reportan 49 especies de pináceas, de las cuales 22 son endémicas y el resto están compartidas en gran parte con otros países del hemisferio norte (30%). En cambio, para los encinares se conocen 160 especies, de las cuales, 109 son endémicas (60%) y el resto se comparten en límites del norte o el sur del país (Gernandt y Pérez-de la Rosa, 2014; Tantray et al., 2017; Valencia-Ávalos, 2010, 2004).
La familia Cicadellidae (Hemiptera: Auchenorrhyncha) es un grupo de insectos que cuenta con una distribución cosmopolita albergando en la actualidad más de 23,000 especies en todos los ecosistemas terrestres conocidos, a excepción de los hielos perpetuos (Bartlett et al. 2018; Dietrich 2005). Los cicadélidos son organismos estrictamente obligados a consumir savia de las plantas vasculares superiores y, en consecuencia, su distribución está estrechamente sujeta a los gradientes tanto en latitud como en longitudes regidas por la vegetación asociada donde residen (Hamilton y Whitcomb, 2010; Pinedo-Escatel y Moya-Raygoza, 2015, 2018). A su vez, esta familia de insectos presenta altos grados de especialización y algunas especies particularmente consumen exclusivamente de una familia o especie vegetal, por lo cual, ciertas agrupaciones de especies se han considerado como organismos potenciales o vectores efectivos en la trasmisión de enfermedades fitopatogénicas a diferentes plantas ya sean cultivadas o silvestres alrededor del mundo (Hamilton y Whitcomb, 2010; Moya-Raygoza et al., 2019).
Por otro lado, la tribu Athysanini comprende a un grupo de cicadélidos muy diverso tanto en especies como en morfología. La tribu cuenta con 911 especies reportadas en el mundo y en México. Pinedo-Escatel, Aragón-Parada et al.(2021) y Pinedo-Escatel, Dietrich et al.(2021) documentaron 146 especies pertenecientes a 46 géneros, de las cuales 70% del total de especies son endémicas. Los hábitos alimenticios de estos organismos bajo esta categoría de tribu son variables y se encuentran desde pastizales o herbáceas, hasta árboles de tallas considerables. De 1930 a la fecha, se han descrito y reportado atisaninos asociados a sistemas boscosos en México (DeLong, 1939; Nielson, 1988; Pinedo-Escatel, Moya-Raygoza et al., 2021), sin embargo, no se ha detallado cuáles son las especies que exclusivamente residen en los bosques mexicanos de pinos y/o encinos, por consecuencia, el nombre de las pináceas hospederas aún sigue sin ser documentado; sin embargo, el rango altitudinal ha sido contundente para la distribución de las especies.
Los bosques de pinos y encinos, históricamente, albergan cicadélidos a lo largo del continente americano, sin embargo, poco o casi nada se conoce sobre su biología incluyendo aspectos como la alimentación o la reproducción (Taylor et al., 1993), a diferencia de otras familias dentro del suborden Auchenorrhyncha reportando ciclos biológicos a detalle (p. ej., Cercopidae en Castro-Valderrama et al. [2017]). En esa misma línea, poco o casi nada se ha profundizado para dar a conocer las especies de atisaninos sobre estos ecosistemas boscosos. Aunque Pinedo-Escatel, Aragón-Parada et al. (2021) ejemplificaron los patrones de distribución que la tribu exhibe sobre varios tipos de bosques mexicanos: seco, encinar, nuboso y de pino, no enlista en detalle las especies que residen exclusivamente en estos hábitats.
La presente contribución tiene como objetivo documentar las especies de la tribu Athysanini asociadas a los bosques de pino y encino en México con base en ejemplares recolectados por el autor, especímenes revisados en colecciones entomológicas y registros de la literatura. Además, se describe una especie nueva del género Eutettix Van Duzee, 1892 a partir de la recolecta en proximidades de la sierra madre del Sur en el estado de Jalisco.
Materiales y métodos
Se utilizaron todos los registros de especímenes recolectados por Pinedo-Escatel, Dietrich et al. (2021) sobre los bosques de pino y encino de México. Además de una serie de trabajos de campo entre 2022 a 2024, se implementó el uso de una red entomológica (35 cm de diámetro × 75 cm de profundidad) y una trampa de luz modificada de acuerdo con Aguilar-Pérez et al. (2019). Los ejemplares recolectados fueron preservados en frascos con etanol al 96% y depositados en refrigeración a -25 °C. Los segmentos terminales masculinos se examinaron siguiendo las técnicas de Triplehorn y Johnson (2005) y Oman (1949) para muestras de Auchenorrhyncha con las siguientes modificaciones: abdómenes sumergidos en solución caliente de KOH al 15%, enjuagados 3 veces en agua destilada y remojados con ácido acético para neutralizar remanentes de la potasa. Posterior al proceso de aclaración, los segmentos terminales se almacenaron en microtubos con glicerina debajo de las muestras correspondientes montadas en seco. Los especímenes fueron revisados en un microscopio estereoscópico Stemi DVA4 Carl Zeiss y digitalizados a través de una cámara óptica Olympus XLC. Las etiquetas de los especímenes estudiados incluyen el tipo de hábitat en el que fueron recolectados.
Para la especie nueva, se tomaron fotografías digitales de los organismos en vista dorsal, lateral y anterior, utilizando una cámara montada en un microscopio estereoscópico Olympus SZX12. La morfología interna de los machos fue fotografiada mediante un microscopio compuesto Olympus BX31 a través del software Gryphx. Todas las imágenes de múltiples planos focales se apilaron utilizando el software Helicon Focus (versión 8.2.18). La longitud del cuerpo se midió con un vernier electrónico desde el margen anterior de la corona, hasta el ápice de las alas anteriores en reposo. El color de los especímenes estudiados fue descrito con base en los ejemplares montados en seco. Los especímenes recolectados de la especie nueva se encuentran en la Colección Nacional de Insectos del Instituto de Biología, Universidad Nacional Autónoma de México, Ciudad de México, México (CNIN) y el Illinois Natural History Survey de la University of Illinois at Urbana-Champaign, Illinois, EUA (INHS).
La morfología externa sigue la esquematización de Dietrich (2005) y Pinedo-Escatel, Dietrich et al. (2021), la venación de las alas sigue el sistema propuesto por Anufriev y Emeljanov (1988) y la quetotaxia de los apéndices sigue a Rakitov (1998). Los cambios nomenclaturales y los nombres válidos siguieron la lista de verificación de Linnavuori (1959), Pinedo-Escatel, Dietrich et al. (2021), Oman et al. (1990), Omán (1949) y Zanol (2008). La descripción morfológica de la especie nueva sigue el modelo y circunscripción para la tribu Athysanini en México por Pinedo-Escatel, Dietrich et al. (2021).
Compilación de registros de Athysanini en pináceas y encinares mexicanos. Se generó una base de datos general que incluye todos los registros conocidos para las especies que se distribuyen específicamente en los bosques de pino y encino en México mediante: a) organismos recolectados por el autor entre 2010 y 2024, especialmente en áreas con bosque de pino y encino en el país, b) la revisión de especímenes tipos y otros adicionales en las siguientes colecciones entomológicas nacionales e internacionales: Colección de Insectos del Instituto de Fitosanidad, Colegio de Postgraduados, Estado de México, México (CEAM); Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México, Ciudad de México, México (CNIN); Centro de Estudios en Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Jalisco, Zapopan, México (CZUG); Illinois Natural History Survey, Champaign, Illinois, EUA (INHS); Ohio State University, C.A. Triplehorn Insect Collection, Columbus, Ohio, EUA (OSUC); United States National Museum of Natural History, Washington, DC, EUA (USNM), y c) los registros conocidos por Nielson (1988) y DeLong (1939, 1946). Todos los registros de presencia, de manera individual, fueron reunidos en un archivo único para cada especie de México. Se utilizaron los trabajos de Pinedo-Escatel, Aragón-Parada et al. (2021) y Pinedo-Escatel, Dietrich et al. (2021) como referencia para las especies endémicas y la homogenización entre los ambientes de pino y encino debido a que demostraron una preferencia por estos hábitats debido a su rango altitudinal.
Renderización de mapas. Los trazos vectoriales con todos los puntos georreferenciados provenientes de la literatura, especímenes en colecciones y eventos de recolección mediante trabajo de campo, fueron compilados y proyectados mediante ArcGis. Los polígonos de la vegetación en los mapas vectorizados siguen el sistema propuesto por Rzedowski (2006) y el mapa resultante incluye la distribución traslapada de los bosques de pino y de encino, además de los registros de atisaninos incluidos en los resultados de Pinedo-Escatel, Aragón-Parada et al. (2021) y Pinedo-Escatel, Dietrich et al. (2021). Únicamente los sitios con presencia de atisaninos fueron proyectados en el mapa. Todos los renderizados fueron elaborados por el autor.
Resultados
Se compiló un total de 385 georegistros de atisaninos para 83 especies de 31 géneros (120 de recolectas, 160 de revisión en colecciones y 105 de literatura) asociados a los bosques mexicanos de pino y encino. Del total de registros, 90% de las especies (74) y 53% de los géneros (17) son endémicos de México. En cambio, 27 especies (32%) residen exclusivamente sobre las pináceas y encinares mexicanos (tabla 1). Se presentan registros a lo largo de 15 estados del país, Guerrero ocupa el primer lugar (n = 80) seguido del Estado de México (n = 70) y Michoacán (n = 63) (fig. 1). El estado con más especies fue Guerrero (32 spp., 40%), seguido de Michoacán (28 spp., 35%) y el Estado de México (23 spp., 22%). Por otro lado, también Guerrero reportó 18 géneros (equivalente a 60% del total) y es el estado con el mayor número de taxones, tanto en registros como en especies endémicas (fig. 2). La distribución de los registros abarca latitudes desde el norte hasta el sur de México sobre la distribución reportada para ambos tipos de bosques (fig. 3).
Diagnosis. Color general amarillo blanquecino con múltiples manchas marrón. Dorso pigóforo fuertemente esclerotizado; apéndice en el interior fuertemente curvo dividido en 2 proyecciones, una delgada en longitud media y otra ensanchada sobresaliendo por el margen caudal. Edeago curvo dorsalmente con 1 par de procesos apicales lateralmente y gonoducto esclerotizado.
Descripción. Chicharritas con cuerpo alargado y moderadamente robustas, en vista dorsal semicilíndricas. Longitud media corporal 2.8 veces más larga que ancha. Dorso blanquecino con múltiples manchas marrones distribuidas disruptivamente. Cabeza con una banda no lineal de color naranja a distancia media entre el margen anterior y posterior dispuesta a manera transocular. Corona, 2.9 veces más ancha que larga; línea media ligeramente marcada; línea transocular convexa; superficie ligeramente estriada. Ojos blancos, 2.5 veces más anchos que largos, basolateral borde interno redondeado (fig. 4A, B). Ocelos negros, distancia al ojo menos de 0.3 veces del diámetro ocelar y 4 veces hacia la línea media. Rostro, 1.3 veces más largo que ancho; línea central blanca con 1 par de manchas naranjas transversales arriba y 4 pares de color marrón subsecuentes. Margen antenal de color marrón obscuro, sin formar una prolongación lateral y débilmente carinado. Antena igual a 1.1 veces en longitud respecto al rostro, color marrón y negro en ápice. Frontoclípleo con suturas laterales parcialmente subparalelas y con construcción media al costado de los pozos antenales, límite superior del frontoclípeo naranja claro y resto de la superficie con múltiples franjas transversales de color marrón delimitando una franja blanquecina sobre la línea media. Anteclípeo con construcción cerca de la base con márgenes laterales subparalelos, ápice expandido y sobrepasando ligeramente el margen posterior de la gena, superficie general de color marrón. Gena igual de amplia como el ancho del frontoclípeo y margen exterior débilmente angulado bajo el ojo (fig. 4C).
Tórax, pronoto blanquecino con manchas irregulares en color marrón, 2.2 veces más ancho que largo, margen anterior débilmente proyectado, margen lateral carinado y no extendido más allá del ancho de la base del ojo, margen posterior cóncavo y una muesca central; superficie trasversalmente surcada con algunas puntuaciones distribuidas sobre todo el disco. Escutelo blanquecino con margen anterior ancho (1.2 veces más que largo) y 1 par de manchas amarillas; margen posterior con 1 par de puntos negros sobre margen lateral; no protuberante en vista lateral. Tegminas. Membrana del primer par de alas translucida con patrones marrón sobre la sección claval y ápice de color negruzco, venas teñidas de color marrón; vena costal con margen blanco y líneas naranjas cercanas a la base alar; sin falsas venas trasversales, celda subapical externa sin falsas venas trasversales, celda subapical central traslucida, celda subapical interna sin venas trasversales adicionales y de color uniforme; 4 celdas apicales presentes, celda interior 1.2 veces más corta que la celda exterior, segunda y tercera celda igual en longitud pero la segunda presenta el doble de ancho que la tercera; región del clavo conectada con la vena Pcu y Cs mediante una vena trasversal en el primer tercio de la región y vena Pcu no conectada a A1. Segundo par alar, con esquema de venación típico para la tribu Athysanini. Patas blancas con máculas anaranjadas y patrón marrón, profémur fila AM con 1 macroseta a media altura, fila IC con 10 setas finas; protibia con 21 macrosetas sobre fila AD; mesotrocánter con 1 seta apical; metafémur con formula 2+2+1 sin setas adicionales.
Tabla 1
Listado de especies de la tribu Athysanini asociadas a bosques de pino y encino en México. Taxón endémico (*).
Género
Especie
Estado
Acunasus*
capitatus*
Guerrero, Jalisco
luteus*
Guerrero, Hidalgo
viridus*
Guerrero, Michoacán
Aligia
alvona*
Guerrero, Hidalgo, Ciudad de México, Veracruz
bicolor*
Ciudad de México
mexicana*
Ciudad de México, Veracruz, Hidalgo
Tabla 1. Continúa
Género
Especie
Estado
Alladanus*
mexellus*
Michoacán, Veracruz, Oaxaca
Angulanus*
incisurus*
Estado de México, Morelos, Jalisco, Oaxaca, Guerrero
Idiodonus
beamerellus*
Michoacán, Hidalgo
copulus*
Michoacán, Guerrero, Veracruz
edentulus*
Estado de México
excavatus*
Hidalgo, Estado de México
plummeri*
Estado de México, Morelos
wickhami*
Puebla, Estado de México, Ciudad de México, Puebla, Michoacán
Bardana*
depressa*
Estado de México, Morelos
Pseudaligia*
nigropunctata*
Hidalgo, Guerrero
Jaacunga*
spatulata*
Ciudad de México
vincula*
Hidalgo, Guerrero, Ciudad de México, Veracruz
Bonneyana
schwartzi
Coahuila, Nuevo León, Chihuahua
caldwelli*
Guerrero, Morelos, Michoacán, Puebla
Colladonus
anademus*
Estado de México
beameri*
Michoacán, Estado de México, Hidalgo, Puebla, Veracruz
claustrus*
Veracruz, Chiapas
bicinctus*
Ciudad de México
dampfi*
Veracruz, Estado de México, Michoacán
verecundus*
Estado de México, Michoacán, Morelos
albocinctus*
Estado de México, Hidalgo, Michoacán, Morelos, Ciudad de México
fasciaticollis*
Puebla, Michoacán, Estado de México
clathrus*
Puebla, Estado de México
titulus*
Ciudad de México, Veracruz, Estado de México
incidus*
Ciudad de México
Paracolladonus*
insculptus*
Estado de México
Paranurenus*
latidens*
Michoacán, Ciudad de México
Stoneana*
balli*
Guerrero
Crassana
marginella*
Guerrero, Oaxaca, Nuevo León
Paracrassana*
nigrifrons*
Puebla
Dampfiana*
deserta*
Guerrero
Conversana*
conversa*
Puebla, Morelos, Ciudad de México
Bandara
spinella*
Guerrero, Michoacán
mimica
Veracruz
Cahya
variabilis
Guerrero, Hidalgo, San Luis Potosí, Oaxaca
Eutettix
copula*
Ciudad de México
discapa*
Michoacán, Estado de México, Ciudad de México
placida*
Michoacán
ortegai*
Puebla
pedus*
Estado de México, Michoacán, Puebla, Morelos, Ciudad de México
Tabla 1. Continúa
Género
Especie
Estado
spinus*
Hidalgo, Ciudad de México, Estado de México
alvadus*
Hidalgo
chelatus*
Estado de México, Ciudad de México, Veracruz
contorqus*
Michoacán, Jalisco, Hidalgo
divergens*
Ciudad de México
harlani
Michoacán, Ciudad de México, Michoacán
krameri*
Hidalgo
lanceolatus*
Puebla
amixtlani sp. nov.*
Jalisco
Mesamia
alta*
Michoacán, Puebla, Guerrero
bifurcata*
Puebla, Michoacán, Ciudad de México, Estado de México, Morelos, Veracruz, Hidalgo
divisa*
Guerrero, Hidalgo, Veracruz, Michoacán, Jalisco
interrupta*
Ciudad de México
frigida*
Estado de México
orizaba*
Veracruz, Michoacán, Guerrero, Puebla, Ciudad de México, Hidalgo
puebla*
Puebla
montana*
Michoacán
interrupta*
Jalisco, Ciudad de México
separata*
Veracruz, Guerrero, Puebla, Hidalgo, Jalisco
Norvellina
cincta*
Puebla, Guerrero
denotata*
Ciudad de México, Guerrero, Michoacán, Estado de México
forficata*
Veracruz, Puebla
recepta*
Zacatecas
uncata*
San Luis Potosí, Guerrero, Puebla, Michoacán, Jalisco
Neodonus*
piperatus*
Michoacán, Hidalgo, San Luis Potosí
Renonus*
rubraviridis*
Jalisco, Guerrero
Retusanus*
luteus*
Guerrero
apicatus*
Guerrero
Cetexa
graecula
Coahuila
Cocrassana*
sexvarus*
Jalisco, Campeche, San Luis Potosí, Veracruz, Michoacán, Morelos
Figura 1. Número total de registros de presencia de la tribu Athysanini en bosques de pino y encino por entidades federativas de la República Mexicana.Figura 2. Número de géneros y especies de la tribu Athysanini presentes en bosques de pino y encino por entidades federativas de la República Mexicana que presentan registros.
Abdomen de color marrón, apodemas no evidentes del esternito I y apodemas del segundo esternito amplios basalmente, pero pobremente desarrollados.
Cápsula genital, macho. Pigóforo 3.0 veces más ancha que alta, dorso fuertemente esclerotizado e inciso a mediana distancia de la base, márgenes sin procesos; con un proceso interior supra desarrollado con 2 terminales, la primera delgada saliendo ligeramente de la cápsula genital y la segunda, robusta y extendida más allá del margen ventral; lóbulos laterales con 3 hileras de 3 a 4 macrosetas distribuidas sobre el tercer tercio del pigóforo; microsetas pronunciadas y puntuaciones desde la longitud media hasta el ápice del pigóforo (fig. 5A, C). Tubo anal 2 veces más largo que ancho con relación al largo total, margen lateral y dorsal esclerotizado. Valva y placas masculinas libres, articuladas con el pigóforo (fig. 5 B). Valva masculina triangular, 3 veces más corta que ancha, superficie numerosamente puntuada. Placa subgenital, con forma triangular; superficie punteada; margen exterior sinuoso con una fila de 10-12 setas finas y margen interior recto sin setas; 5 a 8 macrosetas dispuestas a lo largo del margen externo de la placa (fig. 5 B). Estilo fuertemente bilobulado en la base, con 6 setas en lóbulo preapical y textura corrugada; lóbulo medial corto, no extendido; lóbulo preapical pobremente desarrollado; apófisis curvo, simple y con ápice romo. Conectivo más corto que el estilo; 2.5 veces más corto que el tamaño del edeago en vista dorsal, con forma de Y, brazos anteriores 2 veces más largos que la longitud total del tallo (figs. 5B, 6C). Edeago moderado dentro del pigóforo cubriendo cerca de 2/4 del espacio interior, en dirección dorsal, atrio desarrollado, preatrio poco desarrollado y fuertemente esclerotizado; en vista lateral edeago curvo, ápice con 1 par de espinas laterales; procesos aedeagales sinuosos y direccionados dorsalmente; gonoducto tan amplio como el eje edeagal; gonoducto bien esclerotizado más allá del preatrio hasta el ápice; gonoporo en posición caudal y abierto tan amplio como el eje (fig. 6A, B).
Figura 3. Puntos georreferenciados de registros de atisaninos en bosques de pino y encino en México. Mapa elaborado por J.A. Pinedo-Escatel.
Medidas (mm). Cuerpo: longitud ♂ 4.18-4.55; ancho 1.60-1.67. Cabeza, ancho 1.20-1.28; longitud media 0.36-0.39; ancho previo a los ojos 0.63-0.69; ancho entre los ojos 0.50-0.53. Ojo, ancho 0.46-0.50 y longitud 0.20-0.22. Distancia entre ocelos 0.60-0.62. Frontoclípeo, ancho 0.56-0.57 y longitud 0.96-0.98. Anteclípeo, ancho 0.18-0.19 y longitud 0.26-0.29. Lora, ancho 0.19-0.20 y longitud 0.30-0.33. Gena, ancho 0.50-0.53 y longitud 0.28-0.30. Pronoto, ancho 1.30-1.33 y longitud 0.52-0.54. Escutelo, ancho 0.90-0.96 y longitud 0.72-0.75. Primer par de alas, longitud 3.59-3.62. Pigóforo, altura 0.55-0.59 y longitud 0.85-0.98. Valva, ancho 0.60-0.65 y longitud 0.40-0.53. Placa subgenital, ancho ápice 0.10-0.12, ancho medial 0.22-0.26, ancho base 0.32-0.33 y longitud 0.81-0.83. Estilo, longitud 0.50-0.52. Edeago, longitud 0.77-0.80.
Figura 4. Eutettix amixtlanus sp. nov., aspecto general macroscópico del holotipo macho. A, Hábito general en vista dorsal; B, hábito general en vista lateral; C, hábito del rostro en vista anterior. Escala 1 mm.
Resumen taxonómico
Holotipo. Macho (CNIN) – México: Jalisco, Zapotitlán de Vadillo, Centro Ecoturístico Amixtlan, 19°29’43.1” N, 103°43’16.6” O, 1,543m, 22 febrero 2019, Pinedo-Escatel Col., Trampa de Luz – MEXJAL158.
Paratipos. Dos hembras: 1 hembra (INHS) – México: Jalisco, Autlán de Navarro, Reserva de la Biosfera Sierra de Manantlán, Puerto Los Masos, 19°41’20.0” N, 104°23’48.2” O, 1,651m, 3 noviembre 2017, Pinedo-Escatel Col., Red Entomológica – MEXJAL19; 1 hembra (CNIN) – México: Jalisco, Zapotitlán de Vadillo, Centro Ecoturístico Amixtlan, 19°29’41.6” N, 103°43’17.1” O, 1,545m, 23 febrero 2019, Pinedo-Escatel Col., Trampa de Luz – MEXJAL154
Etimología. El epíteto de la especie, con género masculino, está compuesto por la palabra Amixtlán derivada de la lengua originaria Náhuatl; formada de los vocablos: atl (agua), mixtli (nube) y tan (lugar), que significa “lugar entre las nubes”, nombre que recibe la región por los pobladores locales donde fue encontrado el espécimen holotipo.
Figura 5. Eutettix amixtlanus sp. nov., capsula genital del holotipo. A, Pigóforo en vista dorsal; B, placas subgenitales, estilos y conectivo en vista ventral; C, pigóforo y segmento X y XI en vista dorsal. Escala 0.5 mm.
Hábitat y plantas huésped. La nueva especie se encuentra en bosque de encino en la provincia sierra Madre del Sur, en Jalisco. La caracterización de la localidad tipo cuenta principalmente con Quercus spp., parches de Pinus douglasiana y P. oocarpa (fig. 8A, B).
Anotaciones adicionales. La especie nueva es muy similar a E. apicalus Hepner, 1942, sin embargo, de esta última puede ser diferenciada fácilmente por la disposición de los apéndices de la cápsula masculina, la curvatura del edeago y por los procesos apicales del edeago dispuestos lateralmente.
Discusión
Del total de registros obtenidos de las especies de atisaninos, la mayoría son reportes inéditos para el país (60%) y de éstos, únicamente 8 especies se han registrado en Norteamérica (Oman, 1949). En cambio, la presente contribución registra 74 especies endémicas mexicanas asociadas con los bosques mexicanos de pino y encino. Del total de especies endémicas registradas, algunas se habían reportado anteriormente por Nielson (1988) y DeLong (1946, 1939), sin embargo, no especifican detalles o la especie de árbol hospedero.
Figura 6. Eutettix amixtlanus sp. nov., detalles internos de la cápsula genital del holotipo. A, Edeago en vista lateral; B, ápice del edeago en vista anterior; C, estilos y conectivo en vista ventral. Escala 0.5 mm.
De las 83 especies, 12 cicadélidos fueron colectados en 7 especies de pinos y 2 encinos (tabla 2), en cambio para el resto de las especies que se reportan en asociación con estos bosques, no fue posible documentar el hospedero. Los registros demostraron también que la distribución de los atisaninos recae sobre las principales cadenas montañosas de México, sin embargo, algunas presentan pocos hallazgos como es el caso de la sierra Madre Occidental, a pesar de la extensión de este complejo montañoso, lo cual refleja el hueco que existe en el esfuerzo de muestreo para dicha zona.
Algunas de las especies aquí compiladas han sido mencionadas por Nielson (1988) y DeLong (1946), quienes detallan que los organismos que habitan estos bosques de altitud cuentan con múltiples registros exclusivamente sobre las Pinofitas; sin embargo, se desconocen las especies a las que están asociadas, en contraparte los registros en encinares son inéditos. Así mismo, se ha observado que los patrones de distribución presentados por varios organismos son inespecíficos o por el contrario para algunas especies puntuales, por ejemplo, Colladonus albocinctus (DeLong, 1946), que a pesar de ser una especie endémica de México, podría contar con hospederos alternativos a causa de la cantidad de registros conocidos en altitudes superiores a 2,500 m en Morelos, Ciudad de México, Hidalgo, Michoacán y Estado de México, ésto con fundamento en la distribución que presentan las pináceas mexicanas, según Gernandt y Pérez-de la Rosa (2014). En cambio, C. trabilis Nielson, 1988 se conoce de una localidad en el Estado de México a una altitud de 2,900 m, lo cual podría sugerir un microendemismo en una única planta hospedera, que Nielson (1988) no mencionó. Por consiguiente, otra evidencia es la adaptabilidad de algunas especies de cicadélidos al maniobrar e incitar más actividad en otro tipo de vegetación, como por ejemplo los encinos, como es el caso del género Acunasus DeLong, 1945, que mayormente se ha registrado en varias especies de encinares cercanos a pinos. La complejidad de la biología de los atisaninos por residir en los doseles ha complicado comprender los patrones de distribución, y a su vez, las causas o razones de asociación directamente hacia una o varias especies de plantas hospederas (Pinedo-Escatel, Aragón-Parada et al., 2021; Pinedo-Escatel, Moya-Raygoza et al., 2021).
Figura 7. Distribución de Eutettix amixtlanus sp. nov. en el estado de Jalisco. Mapa elaborado por J.A. Pinedo-Escatel.
Tabla 2
Especies endémicas de la tribu Athysanini con referencia directa de asociación a pinos y encinos en México.
Cicadélidos
Hospederos
Acunasus capitatus
Pinus jaliscana; Pinus douglasiana
Acunasus luteus
Pinus douglasiana; Pinus patula; Quercus affinis
Acunasus viridus
Pinus herrerae; Quercus affinis
Aligia alvona
Pinus douglasiana
Idiodonus beamerellus
Pinus douglasiana; Quercus mexicana
Jaacunga spatulata
Pinus douglasiana
Bonneyana schwartzi
Pinus greggii
Colladonus beameri
Pinus patula
Colladonus incidus
Pinus douglasiana; Quercus mexicana
Eutettix amixtlani sp. nov.
Pinus douglasiana; Pinus oocarpa
Mesamia alta
Pinus lawsonii; Quercus mexicana
Comayagua taeniata
Pinus patula; Quercus affinis
Figura 8. Bosque de encino en localidades donde los especímenes fueron recolectados en la proximidad de la sierra Madre del Sur, Jalisco.
Por otro lado, estos organismos son indicadores de la calidad del hábitat o de hábitats en regeneración debido a la sensibilidad que presentan al adaptarse a las condiciones que requieren para desempeñar sus ciclos biológicos (Biedermann et al., 2005; Hollier et al., 2005). Varias especies de esta tribu también tienden a presentar distribuciones estrechas y no se desplazan más allá de los límites geográficos de su planta huésped (Pinedo-Escatel, Aragón-Parada et al., 2021; Pinedo-Escatel, Moya-Raygoza et al., 2021), lo cual ha generado evaluaciones sobre su conservación, proponiendo que varias de éstas se encuentran en un estatus vulnerable o amenazadas, paralelamente junto al ambiente donde se encuentran (Pinedo-Escatel y Dietrich, 2020). Una consecuencia o reflejo directo es la disminución de especies por causas antropogénicas y por 1 o múltiples factores que intervienen directamente en el número de poblaciones en términos de riqueza, por lo cual las colecciones, acervos y registros disponibles reflejan una oportunidad para realizar evaluaciones pertinentes en la toma de decisiones para su conservación (Pinedo-Escatel et al., 2024). La nueva especie, E. amixtlanus, presenta una distribución restringida ya que reside en 2 localidades aisladas, donde las actividades antropogénicas alrededor van en súbito aumento, limitando la dispersión fuera del hábitat para ambos, la chicharrita y la vegetación hospedera. El aumento de la deforestación no regulada, ganadería con sobrecarga y uso de pesticidas desmedidos se suman a ejercer presión en la región, además de las reportadas por Pinedo-Escatel, Moya-Raygoza et al. (2021). Por ello, para comprender mejor el estatus de conservación de la chicharrita y de los insectos, deben dedicarse esfuerzos futuros en marcos de referencias en acciones de protección de flora y fauna en áreas clave y que prioricen a las especies que residen en ecosistemas paralelamente amenazados.
Agradecimientos
A Cristina Mayorga por su apoyo técnico en la Colección Nacional de Insectos (CNIN) del Instituto de Biología, UNAM. A Edith Blanco Rodríguez (Colegio de Postgraduados) por la toma de fotografías durante su estancia en el Illinois Natural History Survey, Univeristy of Illinois en Urbana-Champaign. A Diego y Axel Pinedo por su apoyo en la recolecta de los organismos estudiados. Esta investigación recibió apoyo por PAPIIT-DGAPA-UNAM (Núm. IT200324).
Referencias
Aguilar-Pérez, J. G., Pinedo-Escatel, J. A. y Valdez-Quezada, B. C. (2019). Three new Mexican species of the endemic Athysanini leafhopper genus Devolana DeLong (Hemiptera: Cicadellidae) from the tropical dry forest. Journal of Natural History, 53, 2039–2056. https://doi.org/10.1080/00222933.2019.1683244
Anufriev, G. A. y Emeljanov, A. F. (1988). Suborder Cicadinea (Auchenorrhyncha). En P. A. Lehr (Ed.), Keys to the insects of the Far East of the USSR. Homoptera and Hemiptera (pp. 12-504). Leningrado: Nauka Publishing House.
Bartlett, C. R., Deitz, L. L., Dmitriev, D. A., Sanborn, A. F., Soulier-Perkins, A. y Wallace, M. S. (2018). The diversity of the true hoppers (Hemiptera: Auchenorrhyncha). En R. G. Foottit y P. H. Adler (Eds.), Insect biodiversity: science and society (pp. 501–590). Glasgow: John Wiley. https://doi.org/10.1002/9781118945582.ch19
Biedermann, R., Achtziger, R., Nickel, H. y Stewart, A. J. (2005). Conservation of grassland leafhoppers: a brief review. Journal of Insect Conservation, 9, 229–243.
Castro-Valderrama, U., Romero-Nápoles, J., Peck, D. C., Valdez-Carrasco, J. M., Llanderal-Cázares, C., Bravo-Mojica, H. et al. (2017). First report of spittlebug species (Hemiptera: Cercopidae) associated with Pinus species (Pinaceae) in Mexico. Florida Entomologist, 100, 206–208. https://doi.org/10.1653/024.100.0136
Dietrich, C. H. (2005). Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Florida Entomologist, 88, 502–517. https://doi.org/10.1653/024.100.0136
DeLong, D. M. (1939). Los phlepsidos (Phlepsius y Texananus) de México (Homoptera-Cicadellidae). Anales de la Escuela Nacional de Ciencias Biológicas, 1, 379–405.
DeLong, D. M. (1946). The Mexican species of Idiodonus (Homoptera-Cicadellidae). Ohio Journal of Science, 46, 13–30.
Gernandt, D. S. y Pérez-de la Rosa, J. A. (2014). Biodiversidad de Pinophyta (coníferas) en México. Revista Mexicana de Biodiversidad, 85 (Supl.), 126–133. https://doi.org/10.7550/rmb.32195
Hamilton, K. G. A. y Whitcomb, R. F. (2010). Leafhoppers (Homoptera: Cicadellidae): a major family adapted to grasslands habitats. En J. D. Shorthouse y K. D. Floate (Eds). Arthropods of Canadian grasslands: ecology and interactions in grassland habitats (pp. 169–197). Cambridge: Biological Survey of Canada, Cambridge University Press.
Hollier, J. A., Maczey, N., Masters, G. J. y Mortimer, S. R. (2005). Grassland leafhoppers (Hemiptera: Auchenorrhyncha) as indicators of habitat condition–a comparison of between-site and between-year differences in assemblage composition. Journal of Insect Conservation, 9, 299–307.
Linnavuori, R. (1959). Revision of the Neotropical Deltocephalinae and some related subfamilies (Homoptera). Annales Botanici Societatis Zoologicae-Botanicae Fenni- cae Vanamo, 20, 1–370.
Moya-Raygoza, G., Cuevas-Guzmán, R., Pinedo-Escatel, J. A. y Morales-Arias, J. G. (2019). Comparison of leafhopper (Hemiptera: Cicadellidae) diversity in maize and its wild ancestor teosinte, and plant diversity in the teosinte habitat. Annals of the Entomological Society of America, 112, 99–106. https://doi.org/10.1093/aesa/say053
Nielson, M. W. (1988). Colladonus and related genera of Mexico and Central America with new taxa and synonymy (Homoptera: Cicadellidae). Great Basin Naturalist Memoirs, 39, 103–134.
Oman, P. W. (1949). The Nearctic leafhoppers (Homoptera: Cicadellidae). A generic classification and check list. Memoirs of the Entomological Society of Washington, 3, 1–253.
Oman, P. W., Knight, W. J. y Nielson, M. W. (1990). Leafhoppers (Cicadellidae): a bibliography, generic check-list and index to the world literature 1956–1985. Wallingford, Inglaterra: CAB International.
Parada-Aragón, J., Reyes, P. C., Rodríguez, A. y Lino, G. M. (2019). Diversidad y distribución geográfica del género Sedum (Crassulaceae) en la Sierra Madre del Sur, México. Revista Mexicana de Biodiversidad, 9, e902921. https://doi.org/10.22201/ib.20078706e.2019.90.2921
Pérez-de la Rosa, J. A. y Gernandt, D. S. (2017). Pinus vallartensis (Pinaceae), a new species from western Jalisco, Mexico. Phytotaxa, 331, 233–242. https://doi.org/10.11646/phytotaxa.331.2.7
Pinedo-Escatel, J. A., Aragón-Parada, J., Dietrich, C. H., Moya-Raygoza, G., Zahniser, J. N. y Portillo, L. (2021). Biogeographical evaluation and conservation assessment of arboreal leafhoppers in the Mexican Transition Zone biodiversity hotspot. Diversity and Distributions, 27, 1051–1065. https://doi.org/10.1111/ddi.13254
Pinedo-Escatel, J. A. y Dietrich, C. H. (2020). Nomenclatural changes and two new species in the leafhopper genus Usanus DeLong (Hemiptera: Cicadellidae) with notes on conservation status. Zootaxa, 482, 567–576. https://doi.org/10.11646/zootaxa.4822.4.6
Pinedo-Escatel, J. A., Dietrich, C. H. y Aragón-Parada, J. (2024). Natural history collections as resources for assessing biodiversity hotspots and insect declines: case studies, opportunities, and challenges. En J. L. León-Cortés y A. Córdoba-Aguilar (Eds.), Insect decline and conservation in the Neotropics (pp. 275–299). The Netherlands: Springer International Publishing. https://doi.org/10.1007/978-3-031-49255-6_14
Pinedo-Escatel, J. A., Dietrich, C. H., Zahniser, J. N., Moya-Raygoza, G. y Portillo, L. (2021). A dichotomous key and checklist for Mexican Athysanini leafhopper genera (Hemiptera: Cicadellidae) with a new species from the Oaxacan dry tropical forest. European Journal of Entomology, 118, 255–278. https://doi.org/10.14411/eje.2021.027
Pinedo-Escatel, J. A. y Moya-Raygoza, G. (2015). Diversity of leafhoppers during the winter dry season on perennial grasses bordering harvested fields of maize. Southwestern Entomologist, 40, 263–272. https://doi.org/10.3958/059.040.0203
Pinedo-Escatel, J. A. y Moya-Raygoza, G. (2018). Diversity of leafhoppers (Hemiptera: Cicadellidae) associated with border grasses and maize during the wet and dry seasons in Mexico. Environmental Entomology, 47, 282–291. https://doi.org/10.1093/ee/nvx204
Pinedo-Escatel, J. A., Moya-Raygoza, G., Dietrich, C. H., Zahniser, J. N. y Portillo, L. (2021). Threatened Neotropical seasonally dry tropical forest: evidence of biodiversity loss in sap-sucking herbivores over 75 years. Royal Society Open Science, 8, 1–13. https://doi.org/10.1098/rsos.201370
Rakitov, R. A. (1998). On differentiation of cicadellid leg chaetotaxy (Homoptera: Auchenorrhyncha: Membracoidea). Russian Entomological Journal, 6, 7–27.
Rzedowski, J. (1991a). Diversidad y orígenes de la flora fanerogámica de México. Acta Botanica Mexicana, 14, 3–21.
Rzedowski, J. (1991b). El endemismo en la flora fanerogámica mexicana: una apreciación analítica preliminar. Acta Botanica Mexicana, 15, 47–64.
Rzedowski, J. (2006). Vegetación de México. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Recuperado el 07 de junio, 2014 de: https://www.biodiversidad.gob.mx/publicaciones/librosDig/pdf/VegetacionMx_Cont.pdf
Sosa, V. y De-Nova, J. A. (2012). Linajes de angiospermas endémicas en México: zonas de alto endemismo para la conservación. Acta Botanica Mexicana, 100, 293–315.
Steinmann, V. W. (2002). Diversidad y endemismo de la familia Euphorbiaceae en México. Acta Botanica Mexicana, 61, 61–93.
Tantray, Y. R., Wani, M. S. y Hussain, A. (2017). Genus Quercus: an overview. International Journal of Advance Research in Science and Engineering, 6, 1880–1886.
Taylor, R. A. J., Nault, L. R. y Styer, W. E. (1993). Experimental analysis of flight activity of three Dalbulus leafhoppers (Homoptera: Auchenorrhyncha) in relation to migration. Annals of the Entomological Society of America, 86, 655–667.
Triplehorn, C. A. y Johnson, N. F. (2005). Borror and DeLong’s introduction to the study of insects. Belmont, TN: Thomson.
Valencia-Ávalos, S. (2010). Notes on the genus Quercus in Mexico. International Oaks, 21, 100–120.
Valencia-Ávalos, S. (2004). Diversidad del género Quercus (Fagaceae) en México. Boletín de la Sociedad Botánica de México, 75, 33–53.
Zanol, K. M. R. (2008). Catalogue of the Neotropical Deltocephalinae (Hemiptera: Cicadellidae). Part III–Tribe Athysanini. Acta Biológica Paranaense, 37, 1–104.
Universidad Autónoma del Estado de México, Instituto de Ciencias Agropecuarias y Rurales, Instituto Literario Núm. 100, Colonia Centro, 50000 Toluca, Estado de México, México
*Autor para correspondencia: arendaraa@uaemex.mx (A.R. Endara-Agramont)
Recibido: 13 febrero 2024; aceptado: 11 julio 2025
Resumen
La extensión forestal de México es de 138.7 millones de ha, 42% de las cuales se encuentran afectadas por plantas parásitas. El Área de Protección de Flora y Fauna Sierra de Quila tiene 6,788 ha de bosques de pino-encino y encino-pino, que son susceptibles al ataque de plagas y enfermedades. El objetivo fue determinar su distribución espacial y niveles de infestación. Para ello, se establecieron 330 sitios de muestreo (SM) circulares de 1,000 m2, utilizando las curvas de nivel como transectos de acceso al bosque, con una separación altitudinal de 100 m entre curvas desde 2,000 hasta 2,600 m snm, con una equidistancia entre sitios de 300 m. En cada sitio se registraron datos físico-geográficos, dasonómicos y afectación por plantas parásitas. Más de 2,050 ha de sus bosques (30%) están afectadas por muérdago enano (Arceuthobium durangense) y diversos muérdagos verdaderos (Phoradendron bolleanum, P. brachystachyum, P. lanceolatum, P. reichenbachianum, Cladocolea cupulata, C. grahamii, C. mcvaughii, C. microphylla y Psittacanthus calyculatus), todos con niveles bajos de infestación, por lo que aún resulta oportuno establecer estrategias de manejo y control para evitar mayores daños al bosque.
The forested area of Mexico is 138.7 million ha, 42% of which are affected by parasitic plants. The Sierra de Quila Flora and Fauna Protection Area has 6,788 ha of pine-oak and oak-pine forests, which are susceptible to pests and diseases. The objective was to determine their spatial distribution and infestation levels. To this end, 330 circular sampling sites (SM) of 1,000 m2 were established, using contour lines as forest access transects. The elevations were separated by 100 m between contours from 2,000 to 2,600 m asl, and the distance between sites was 300 m. Physical-geographical and forestry data, as well as the impact on parasitic plants, were recorded at each site. More than 2,050 ha of its forests (30%) are affected by dwarf mistletoe (Arceuthobium durangense) and various true mistletoes (Phoradendron bolleanum, P. brachystachyum, P. lanceolatum, P. reichenbachianum, Cladocolea cupulata, C. grahamii, C. mcvaughii, C. microphylla, and Psittacanthus calyculatus), all with low levels of infestation, so it is still possible to establish management and control strategies to avoid further damage to the forest.
México posee una extensión forestal de 138.7 millones de ha (Conafor, 2021). Éstas son susceptibles a disturbios; particularmente los ocasionados por insectos fitófagos, plantas parásitas e incendios (USDA, 2013). En particular, en Jalisco, durante el periodo 2012-2021 se reportaron 54,158 ha de bosque con diversos problemas fitosanitarios: plantas parásitas (61%), enfermedades (cancro resinoso, pudrición de raíz y roya; 19%), insectos descortezadores (13%) e insectos defoliadores (6%) (Semadet, 2022). Un factor ambiental que incrementa la vulnerabilidad de las especies forestales a plagas o enfermedades es el cambio climático (Bentz et al., 2010).
En México, se tienen registros de más de 200 especies de insectos y patógenos que provocan daños en los ecosistemas forestales (Conafor, 2007). En Jalisco, las plantas parásitas que se registran con mayor incidencia en los bosques son 4 especies de muérdagos enanos (Arceuthobium spp.) en coníferas y 21 especies de muérdagos verdaderos en latifoliadas (Semadet, 2021). La mayor riqueza de plantas parásitas se encuentra en las sierras de Tapalpa, Quila, Primavera, Cacoma, Manantlán (Vázquez-García et al., 1995), Bolaños (Vázquez-García et al., 2004), del Halo, Nevado de Colima, Cerro Grande y volcán Cerro Viejo, con 16 especies de plantas parásitas, el mayor número de especies encontradas para un municipio (Vázquez-García et al., 1995).
Los muérdagos son plantas hemiparásitas que dependen parcialmente de otros organismos para poder subsistir (Conafor, 2007; Nickrent y Musselman, 2004). Cuando se establecen en el tallo de su hospedero, compiten con él por agua y nutrimentos, lo que dificulta su crecimiento (Espinoza-Zúñiga et al., 2019). Por otra parte, cuando los muérdagos son abundantes en el árbol hospedero pueden provocarle la muerte (Alvarado-Rosales et al., 2007). Abarcan especies del género Arceuthobium, conocidas como muérdagos enanos, así como de los géneros Cladocolea, Phoradendron, Psittacanthus y Struthanthus, todos ellos conocidos como muérdagos verdaderos (Nickrent y Musselman, 2004). Son plantas arbustivas que se establecen en la parte aérea, más frecuentemente en ramas (muérdagos verdaderos) o en las ramas y en el fuste (muérdagos enanos), que afectan el desarrollo de su hospedero a través del sistema endofítico (Hawksworth, 1961; Rey et al., 1991), lo que induce cambios fisiológicos y bioquímicos (Mathiasen et al., 2008). Endara-Agramont et al. (2022) encontraron que algunas de las variables que influyen en la presencia de muérdago son la altitud y la orientación de ladera, y que su existencia en un árbol hospedero lo predispone al establecimiento de insectos descortezadores y plantas parásitas debido a que hay pérdida del vigor y reducción en su crecimiento, lo que genera mortalidad y facilita la dispersión de semillas (Cibrián-Tovar et al., 1995; Klutsch et al., 2014).
Las plantas parásitas tienen un valor ecológico. Son indicadoras de la salud del bosque, pues a mayores niveles de disturbio su prevalencia se incrementa (Cházaro et al., 1992). En contraste, promueven la diversidad de los bosques al proporcionar recursos alimentarios para algunas especies animales debido a la disponibilidad de sus frutos en épocas en las que otros alimentos son escasos (Matula et al., 2015).
La información sobre la distribución espacial de muérdagos en la sierra de Quila es escasa, a pesar de haber sido reportada como una zona con alta riqueza de plantas parásitas (Vázquez-García et al., 1995), por lo que el objetivo de esta investigación fue identificar las plantas parásitas y sus hospederos, así como determinar la distribución espacial y los niveles de infestación a través de un muestreo intensivo en esta área.
Materiales y métodos
El estudio se realizó en el Área de Protección de Flora y Fauna (APFF) Sierra de Quila (fig. 1), en Jalisco entre los municipios de Tecolotlán, San Martín de Hidalgo y Tenamaxtlán, a 100 km al suroeste de la ciudad de Guadalajara, Jalisco (Villavicencio et al., 2005). Forma parte del Sistema Volcánico Transmexicano, incluida en la subprovincia “Sierras de Jalisco”, el relieve es montañoso con pendientes de 15 a 60° (Guerrero y Cházaro, 1995). La altitud oscila entre 1,350 y 2,560 m snm y de acuerdo con la clasificación de Köppen, el clima en la región es templado húmedo con lluvias en verano C(w2) (W) (Santiago et al., 2012). La alta variedad de condiciones geográficas y climáticas de esta sierra le permiten albergar una muestra representativa de la biodiversidad del occidente de México, con distintos tipos de bosques, que incluyen encino, pino-encino, encino-pino y selva baja caducifolia (Villavicencio etal., 2015) (fig. 1).
Este estudio se realizó en los bosques de pino-encino y encino- pino (6,788 ha); debido a que el mayor número de reportes de presencia de muérdago se encuentra en éstos (Cibrián-Tovar et al., 2007; Galván, 2016; Geils et al., 2002; Martínez-Ambriz, 2020; Ornelas, 2019; Rzedowski y Calderon de Rzedowski, 2011; Sandoval y Siqueiros, 2019). El muestreo se realizó de noviembre 2021 a noviembre 2022. Para ello, se establecieron 330 sitios de muestreo (SM) circulares de 0.1 ha (35.6 m de diámetro) establecidos sobre las curvas de nivel con una separación altitudinal de 100 m entre curvas (fig. 2); las cuales fueron ubicadas en 7 distintas altitudes fijadas entre 2,000 y 2,600 m snm. Lo anterior se hizo para considerar la variabilidad climática y los cambios de vegetación asociados a las diferencias de altitud (Mayer y Ott, 1991). Dentro de la misma cota altitudinal los sitios tuvieron una separación equidistante de 300 m, lo que permitió superar la intensidad mínima de muestreo recomendada de 0.28% (Dauber, 1995).
En cada SM se registraron las variables fisiográficas, las características de la vegetación (cobertura de pastos y arbustos) e información dasonómica de las especies arbóreas con un diámetro normal (DN) ≥ 2.5 (Endara-Agramont et al., 2022, 2023; Rojas-García et al., 2019) (tabla 1), así como, el nivel de infestación de los árboles con muérdago enano y verdadero (Alvarado-Rosales y Saavedra-Romero, 2005; Hawksworth, 1983) (tabla 2). Además, se recolectaron muestras botánicas de las plantas parásitas y del árbol hospedero, las cuales fueron identificadas mediante las claves taxonómicas de Alvarado-Cárdenas (2010), Martínez-Ambriz (2020) y Vázquez-Collazo et al. (2006), así como otros ejemplares en el Herbario CHAP, de la Universidad Autónoma Chapingo y el Herbario Nacional de México de la Universidad Nacional Autónoma de México, para poder rectificar que las muestras recolectadas coincidieran con otras previamente identificadas.
Se elaboró una base de datos que permitió generar los polígonos con la distribución espacial de muérdago a través del software ArGis ver. 10.8, en el cual se aplicó la metodología de interpolación de datos del vecino natural (Sibson, 1982), que consiste en generar polígonos con información específica (asignada a través de la generación de la base de datos) para formar superficies continuas (Childs, 2004; Pirotti y Tarolli, 2010).
Figura 1. Mapa de ubicación geográfica del Área de Protección de Flora y Fauna Sierra de Quila, Jalisco. Fuente: Elaboración propia con base en información de Conabio (2019) y Villavicencio et al. (2015).Figura 2. Distribución de los sitios de muestreo (SM). Fuente: elaboración propia, con base en Villavicencio et al. (2015).
Se realizó un Anova para determinar el efecto de la densidad del bosque sobre el porcentaje de cobertura y los individuos con muérdago con el programa Statgraphics Centurion XVI v.16.1.03 (StatPoint Technologies Inc., 2009). Finalmente, se realizó un gráfico descriptivo para analizar si existe relación entre la presencia de muérdago y la estructura arbórea.
Resultados
Se encontró que cerca de 40% de los SM tiene presencia de muérdagos, lo que corresponde a poco más de 2,050 ha afectadas. Las especies encontradas fueron: Arceuthobium durangense (A) (42%) sobre Pinus douglasiana y P. devoniana; Phoradendron bolleanum (B)(9%), P. brachystachyum (C)(2%), P. lanceolatum (D) (2%) y P. reichenbachianum (E) (2%) sobre Arbutus xalapensis, Quercus castanea, Q. laeta y Q. coccolobifolia; Cladocolea cupulata (F)(25%), C. grahamii (G) (5%), C. mcvaughii (H) (7%)y C. microphylla (I) (2%) sobre Pinus lumholtzii, P. oocarpa, Quercus coccolobifolia, Q. castanea, Q. resinosa, Q. rugosa y Q. crassipes; así como Psittacanthus calyculatus (J) (4%) sobre Quercus resinosa, Q. laeta y Q. rugosa (tabla 3), lo que muestra que estos bosques son dañados por una parte representativa de la diversidad de muérdagos presentes en Jalisco.
Asimismo, el muestreo señala que la distribución espacial de muérdagos enanos y verdaderos sobre los bosques de pino-encino y encino-pino de la sierra de Quila obedece patrones de distribución altitudinal para cada género de plantas parásitas: Cladocolea de 2,000 a 2,300 m, Arceuthobium de 2,100 a los 2,300 m, Phoradendron de 2,200 a 2,400 m y Psittacanthus únicamente en 2,400 m (fig. 3). Por otro lado, de acuerdo con la superficie afectada de la sierra de Quila, 35% presenta muérdago enano (A. durangense), 47% muérdago verdadero (Cladocolea, Phoradendron y Psitthacanthus) y 18% asociaciones de diferentes especies de muérdagos. Si bien, todas las especies de muérdago (A. durangense, Cladocolea, Phoradendron y Psitthacanthus) se encuentran en niveles bajos dentro de los hospederos (tabla 2), la distribución en términos de superficie es de 30% dentro de todo el bosque (fig. 4), pudiendo incrementar hacia otros hospederos y rodales.
Tabla 1
Variables de sitio e individuos; información recabada en los sitios de muestreo (SM).
Diámetro a la altura del pecho (DAP) Altura total Adaptación de la escala del nivel de infestación para muérdagos enanos (Hawksworth, 1983) Nivel de infestación de muérdagos verdaderos (Alvarado-Rosales y Saavedra-Romero, 2005) Observaciones (daños por rayo, sin ápice o muertos)
Tabla 2
Escalas de evaluación para muérdagos enanos y verdaderos.
Escalas de evaluación de muérdagos enanos Adaptada de Hawksworth (1983)
Escalas de evaluación de muérdagos verdaderos Alvarado-Rosales y Saavedra-Romero (2005)
0 Sano. No muestra rasgos de muérdago 1 Infestación ligera (50% o menos de las ramas infectadas) 2 Infestación severa (más de 50% de las ramas infectadas o en el fuste)
0 Sano. Sin infección aparente 1 infección ligera (1-30%) 2 Infección media (31-60%) 3 Infección severa (61-90%)
Otro factor que influye en la presencia de muérdagos es la densidad, lo que corrobora el Anova, donde se determinaron 3 categorías, denso, semidenso y fragmentado; el total de los árboles registrados fueron de 10,346 y se clasificaron de la siguiente manera: denso (≥ 361), semidenso (151-360) y fragmentado (30-150). (tabla 4). Estos resultados muestran que la presencia de muérdago se concentra en el bosque semidenso (13%) y fragmentado (34%) (fig. 5), observando estas afectaciones en el dosel dominante y codominante, donde se confirma que la reducción del número de árboles por unidad de superficie implica una mayor cantidad de luz al interior del bosque, relacionado con el incremento de las poblaciones de muérdago. Aun cuando el muérdago afecta generalmente a todas las categorías diamétricas, es más abundante entre los 5 y 40 cm de DAP (fig. 6). Del total de árboles registrados mencionados anteriormente, 10% está infestado por muérdago, lo que indica que los niveles se encuentran dentro de los límites del parasitismo (Mathiasen, 1998).
Figura 3. Mapa de la distribución espacial de tipo de muérdagos en APFF Sierra de Quila, Jalisco. Fuente: trabajo de campo.Figura 4. Mapa de los niveles de infestación por muérdagos enanos y verdaderos en el APFF Sierra de Quila, Jalisco. Fuente: trabajo de campo.
Discusión
El muestreo realizado indica un rango altitudinal que oscila entre 2,100 y 2,400 m snm, lo que coincide con Queijeiro-Bolaños et al. (2013) y Endara-Agramont et al. (2023), quienes mencionan que a mayor altitud existe menor número de las plantas parásitas; así mismo, la distribución de éstos obedece a la presencia de sus hospederos y a su capacidad de dispersión (Cibrián-Tovar et al., 2007). Estos registros muestran que Arceuthobium durangense es el único que presenta niveles de infestación medio, siendo más agresivo debido a que ataca en mayor medida al fuste en comparación con el muérdago verdadero (Hawksworth, 1961; Rey et al., 1991).
Cabe destacar que de los 122 SM con muérdago, 10% de los individuos se ve afectando principalmente a individuos juveniles de pinos y encinos (5 y 40 cm de DAP), lo que concuerda con Cibrián-Tovar et al. (2007), donde mencionan que el arbolado juvenil es el más susceptible al establecimiento de estas plantas parásitas. De acuerdo con la estructura y densidad del bosque, el dosel dominante y codominante compuesto por diversas especies de pinos y encinos y los rodales fragmentados son los más afectados, lo anterior coincide con lo reportado por Queijeiro-Bolaños y Cano-Santana (2015), quienes afirman que los muérdagos responden a presiones ambientales, donde la disminución de las densidad de los hospederos favorece la infestación debido a que estas condiciones son necesarias para la producción de estructuras vegetativas y reproductivas, teniendo menos competencia por recursos al tener una menor cantidad de árboles vecinos; por lo tanto, estos son directamente aprovechados por los muérdagos aumentando la probabilidad de infestación dentro de los individuos.
Figura 5. Comparación del porcentaje de individuos infestados por muérdagos en parcelas con distinta densidad de árboles (p < 0.0001).
Tabla 4
Asociaciones arbóreas encontradas en el APFF Sierra de Quila, Jalisco.
Asociaciones arbóreas
Densidad (núm./ha)
Especie
DAP Promedio (cm ± e.e.)
Altura promedio (m ± e.e.)
Dosel
Clethra sp. – Pinus sp. – Quercus sp.
310
Pinus sp.
34
20
Dominante
Quercus sp.
24
16
Codominante
Clethra sp.
17
11
Suprimido
Pinus sp. – Quercus sp.
287
Pinus sp.
24
15
Dominante
Quercus sp.
18
11
Codominante
Quercus sp. – Pinus sp.
292
Pinus sp.
24
14
Dominante
Quercus sp.
17
11
Codominante
Figura 6. Número de árboles en sitios sanos y enfermos de acuerdo con las categorías diamétricas.
En este estudio se encontró que la altitud, la densidad de árboles, y el DN favorecen la prevalencia de muérdagos (Ferrenberg, 2020; Mathiasen, 2019), como consecuencia reducen el vigor de los árboles infestados, ésto coincide con lo encontrado por Cibrián-Tovar et al. (1995) y Endara-Agramont et al. (2023), quienes reportan que la pérdida del vigor y reducción en el crecimiento de árboles parasitados genera mayor probabilidad de muerte en los hospederos y facilita la infestación por descortezadores y/o enfermedades (Klutsch et al., 2014).
Hawksworth (1983) encontró que otro de los factores que afecta inversamente la prevalencia de los muérdagos es la densidad de árboles, a mayor densidad arbórea menor presencia de muérdago. En relación con ésto, se ha reportado que las semillas de estos organismos proliferan bajo un dosel forestal semidenso o fragmentado (Endara-Agramont et al., 2023). Más de 30% de la superficie de la sierra de Quila está infestada por muérdagos de 4 géneros (Arceuthobium, Cladocolea, Phoradendron y Psittacanthus), distribuidos entre 2,000 y 2,400 m snm, mismos que afectan a todas las categorías diamétricas, sobre todo los que están establecidos en bosques semidensos y fragmentados.
Agradecimientos
Agradecemos el apoyo financiero del proyecto Conafor-Conacyt A-S-130105 “Identificación, evaluación y manejo integrado de plantas parásitas en cuatro regiones de México: Noroeste (Durango), Centro-Occidente (Michoacán y Jalisco), Oriente (Puebla y Veracruz), Centro-Sur (Estado de México y Tlaxcala)”; a las brigadas contra incendios de las localidades de Quila, El Cobre, Mesa del Cobre y Lagunillas y al grupo de investigación de Bosques de Alta Montaña del Instituto de Ciencias Agropecuarias y Rurales.
Referencias
Alvarado-Cárdenas, L. (2010). Flora del valle de Tehuacán-Cuicatlán, fascículo 75, VISCACEAE Bastch. México D.F.: Universidad Nacional Autónoma de México.
Alvarado-Rosales, D. y Saavedra-Romero, L. (2005). El género Cladocolea (Loranthaceae) en México: muérdago verdadero o injerto. Revista Chapingo. Serie Ciencias Forestales y del Ambiente, 11, 5–9.
Alvarado-Rosales, D., Saavedra-Romero, L. y Almaraz-Sánchez, A. (2007). Agentes asociados y su papel en la declinación y muerte de encinos (Quercus, Fagaceae) en el centro-oeste de México. Polibotánica, 23, 1–21.
Bentz, B. J., Régniére, J., Fettig, C.J., Hansen, E. M., Hayes, J. L., Hicke, J. A. et al. (2010). Climate change and bark beetles of the Western United Satates and Canada: direct and indirect effects. BioScience, 60, 602–613. https://doi.org/10.1525/bio.2010.60.8.6
Cházaro, B., Huerta, M. M., Patiño, B. R., Sánchez, F. R., Lomelí, M. E. y Flores, M. A. (1992). Los muérdagos (Loranthaceae) de Jalisco, parásitas poco conocidas. Ciencia y Desarrollo, 17, 70–85.
Childs, C. (2004). Interpolating surfaces in ArcGIS spatial analyst. In ArcUser, ESRI, California, pp. 32-35.
Cibrián-Tovar, D., Alvarado-Rosales, D. y Gracia-Díaz, S. E. (2007). Enfermedades forestales en México. Chapingo, Estado de México: Universidad Autónoma de Chapingo.
Cibrián-Tovar, D., Méndez-Montiel, J. T., Campos-Bolaños, R., Tates III, H. O. y Flores-Lara, J. E. (1995). Insectos forestales de México. Chapingo, Estado de México: Universidad Autónoma de Chapingo.
Conafor (Comisión Nacional Forestal). (2007). Manual de sanidad vegetal. México D.F.: Conafor.
Conafor (Comisión Nacional Forestal). (2021). Estado que guarda el sector forestal en México. Bosques para el bienestar social y ambiental 202. México D.F.: Conafor.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2019). División política estatal, escala 1:250000. Catálogo de metadatos geográficos. México D.F.
Dauber, E. (1995). Guía práctica y teórica para el diseño de un inventario forestal de reconocimiento. Santa Cruz. BOLFOR.
Endara-Agramont, A, R., Heredia-Bobadilla, R. L., García-Almaraz, L. A., Luna-Gil, A. A y Aguirre-Zúñiga, J. J. (2023). Distribución espacial del descortezador Dendroctonus adjunctus Blandford, 1897 (Coleoptera: Curculionidae, Scolytinae) en dos bosques de alta montaña del centro de México. Acta Zoológica Mexicana, 39, 1–17. https://doi.org/10.21829/azm.2023.3912569
Endara-Agramont, A. R., Calderón-Contreras, R., Nava-Bernal, G. y Franco-Mass, S. (2013). Analysis of fragmentation processes in high-mountain forests of the Centre of México. American Journal of Plant Sciences, 4, 697–704. https://doi.org/10.4236/ajps.2013.43A088
Endara-Agramont, A. R., Heredia-Bobadilla, R. L., García-Almaraz, L. A., Luna-Gil, A. A., Franco-Mass, S. y Cibrián-Llanderal, V. D. (2022). Factores asociados con la distribución espacial de muérdagos enanos en dos poblaciones de Pinus hartwegii del centro de México. Revista Mexicana de Biodiversidad, 93, e935008. https://doi.org/10.22201/ib.20078706e.2022.93.5008
Espinoza-Zúñiga, P., Ramírez-Dávila, J. F., Cibrián-Tovar, D., Villanueva-Morales, A., Cibrián-Llanderal, V. D., Figueroa-Figueroa, D. K. y Rivera-Martínez, R. (2019). Modelación de la distribución espacial del muérdago (Santalales: Loranthaceae) en las áreas verdes de la delegación Tlalpan, México. Bosque,40, 17–28. https://doi.org/10.4067/S0717-92002019000100017
Ferrenberg, S. (2020). Dwarf mistletoe infection interacts with tree growth rate to produce opposing direct and indirect effects on resin duct defenses in lodgepole pine. Forests, 11, 222. https://doi.org/10.3390/f11020222
Galván, G. L. (2016). El género Cladocolea Tiegh. (Loranthaceae) en México (Tesis de maestría). Universidad Autónoma Metropolitana, Unidad Iztapalapa. México D.F.
Geils, B. W y Vázquez, C. (2002). Loranthaceae and Viscaceae in North America. En B. W. Geils, J. C. Tovar y B. Modoy (Coords.), Mistletoes of North America conifers. (pp. 1–5). Gen. Tech. Rep. RMRS-GTR-98. U. S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, Utah.
Guerrero, N. J. y Cházaro, B. (1995). Datos generales sobre la Sierra de Quila, Jalisco. En M. Cházaro Basáñez, E. Lomelí-Mijes, R. Acevedo-Rosas y S. Ellerbracke-Román (Compiladores) Antología botánica del estado de Jalisco (pp. 70–73). Guadalajara: Universidad de Guadalajara.
Hawksworth, F. (1961). Dwarf mistletoe of ponderosa pine in the Southwest. Technical Bulletin, 1246. USDA, Washington DC.
Hawksworth, F. (1983). Mistletoes as forest parasites. En M. Calder y P. Bernhardt (Eds.), The biology of mistletoes (pp. 320–329). Nueva York: Academic Press.
Klutsch, J. G., Beam, R. D., Jacobi, W. R. y Negrón, J. F. (2014). Bark beetles and dwarf mistletoe interact to alter downed woody material, canopy structure, and stand characteristics in northern Colorado ponderosa pine. Forest Ecology and Management, 315, 63–71. https://doi.org/10.1016/j.foreco.2013.12.024
Martínez-Ambriz, E. (2020). Familia Loranthaceae, fascículo 214. Flora del Bajío y Regiones adyacentes. Pátzcuaro, Mich.: Instituto de Ecología, A.C.
Mathiasen, R. L. (1998). Comparative susceptibility of conifers to larch dwarf mistletoe in the Pacific Northwest. Forest Science, 44, 559–568. https://doi.org/10.1093/forest science/44.4.559
Mathiasen, R. L. (2019). Susceptibility of red fir and white fir to fir dwarf mistletoe (Arceuthobium abietiunum) in California. Forest Pathology, 2019, e12516.
Mathiasen, R. L., Nickrent, D., Shaw, D. y Watson, D. (2008). Mistletoes. Pathology, systematics, ecology, and management. The American Phytopathological Society, 92, 988–1006. https://doi.org/10.1094/pdis-92-7-0988
Matula, R., Svátek, M., Pálková, M., Volařík, D. y Vrška, T. (2015). Mistletoe infection in an oak forest is influenced by competition and host size. Plos One, 10, 1–11. https://doi. org/10.1371/journal.pone.0127055
Mayer, H. y Ott, E. (1991) Gebirgswaldabau-Schutzwaldpflege: ein waldbauulicher Beitrag zur Landschaftssokologie und zum Umweltschutz(Silviculture in Mountain Forest-Management of protection forest: A silvicultural contribution to landscape ecology and enviromental protection), 2nd revised Edn. Stuttgart: Gustav Fischer.
Nickrent, D. L. y Musselman, L. J. (2004). Introduction to parasitic flowering plants. The plant health instructor. Heide-Jorgensen HS (Ed.) Brill. https://doi.org/10.1094/PHI-I-2004-0330-01
Ornelas, J. F. (2019). Los muérdagos Psittacanthus en México: ecología, evolución, manejo y conservación. Biodiversitas, 146, 12–16.
Pirotti, F. y Tarolli, P. (2010). Suitability of LiDAR point density and derived landform curvature maps for channel network extraction. Hydrological Processes, 24, 1187–1197. https://doi.org/10.1002/hyp.7582
Queijeiro-Bolaños, M. E. y Cano-Santana, Z. (2015). Dinámica temporal de la infestación por muérdago enano (Arceuthobium globosum y A.vaginatum) en Zoquiapan (Parque Nacional Iztaccíhuatl Popocatépetl), México. CienciaUAT, 9, 6–14.
Queijeiro-Bolaños, M. E., Cano-Santana, Z. y Castellanos-Vargas, I. (2013). Does disturbance determine the prevalence of dwarf mistletoe (Arceuthobium, Santalales: Viscaceae) in Central Mexico? Revista Chilena de Historia Natural, 2, 181–190. https://doi.org/10.4067/s0716-078×2013000200007
Rey, L., Sadik, A., Fer, A. y Renaudin, S. (1991). Trophic relations of the dwarf mistletoe Arceuthobium oxycedri with its host Juniperus oxycedrus. Journal of Plant Physiology, 138, 411–416. https://doi.org/10.1016/S0176-1617(11)80515-8
Rojas-García, F., Fredericksen, T. S., Vázquez, L. S. y Endara, A. A. R. (2019). Impact of timber harvesting on carbon storage in montane forests of central Mexico. New Forest, 50, 1043–1061. https://doi.org/10.1007/s11056-019-09714-z
Rzedowski, J. y Calderón-de Rzedowski, C. (2011). Dos especies notables de Phoradendron (Viscaceae) de la mixteca Oaxaqueña (México), una nueva y una complementada. Acta Botanica Mexicana, 1, 3–10. https://doi.org/10.21829/abm96.2011.254
Sandoval, O. M. y Siqueiros, D. M. (2019). Cladocolea loniceroides, un nuevo registro para la flora de Aguascalientes, México. Investigación y Ciencia de la Universidad Autónoma de Aguascalientes, 27, 51–54. https://doi.org/10.33064/iycuaa2019782232
Santiago, P. A. L., Domínguez, M. L., Rosas, V. E. y Rodríguez, J. M. (Coords.). (2012). Anfibios y reptiles de las montañas de Jalisco: Sierra de Quila. Orgánica Editores/ Conabio. Guadalajara, Jal., México.
SEMADET (Secretaría de Medio Ambiente y Desarrollo Territorial). (2021). Diagnóstico y plan de acción para la atención a plagas y enfermedades forestales en el Estado de Jalisco. Recuperado el 23 de noviembre, 2023 de: http://sivicoff.cnf.gob.mx/ContenidoPublico/02%20Informes%20de%20acciones%20operativas/DiagnosticosEstatales/2021/Jalisco.pdf
SEMADET (Secretaría de Medio Ambiente y Desarrollo Territorial). (2022). Programa operativo estatal de sanidad forestal para el estado de Jalisco. Recuperado el 23 de noviembre, 2023 de: http://sivicoff.cnf.gob.mx/ContenidoPublico/02%20Informes%20de%20acciones%20operativas/DiagnosticosEstatales/2022/Jalisco.pdf
Sibson, R. (1982). A brief description of natural neighbor interpolation. En John Wiley y Sons (Eds.), Interpolating multivariate data (pp. 21–36). Nueva York: John Wiley & Sons.
StatPoint Technologies, Inc. (2009). Version, 16, 17. Statgraphics Centurion, X. V. I.
USDA (United States Departament of Agriculture). (2013). National strategy framework for invasive species management. Forest Service. Washington: United States Department of Agriculture. Recuperado el 14 de diciembre, 2023 de: https://www.fs.usda.gov/foresthealth/publications/Framework_for_Invasive_Species_FS-1017.pdf
Vázquez-Collazo, I., Villa, R. y Madrigal, H. (2006). Los muérdagos (Loranthaceae) en Michoacán. Libro TécnicoNúm. 2. Mich. Uruapan, Michoacán: CIRPAC/ INIFAP.
Vázquez-García, J. A., Cházaro B., Nieves H., Vargas-Rodríguez, M., Vázquez-García, A. y Flores, M. (2004). Flora del norte de Jalisco y etnobotánica huichola. Guadalajara: Universidad de Guadalajara.
Vázquez-García, J. A., Cuevas, G. R., Cochrane, T. S., Litis, H. H., Santana, M. F y Guzmán, H. L. (1995). Flora de Manantlán: plantas vasculares de la Reserva de la Biósfera Sierra de Manantlán Jalisco-Colima, México. Sida, Botanical Miscellany, 13, 1−312.
Villavicencio, G. R., Ávila, C. R., Treviño, G. E. y Muñiz, C. M. (2015). Cartografía de la cobertura vegetal y uso actual del suelo de la Sierra de Quila, Jalisco, México. Memorias de resúmenes en extenso SELPER-XXI-México-UACJ-2015.
Villavicencio, G. R., Bauche, P., Gallegos, R., Santiago, P. y Huerta, M. (2005). Característica estructural y diversidad de comunidades arbóreas de La Sierra de Quila. Boletín del Instituto de Botánica de la Universidad de Guadalajara, 13, 67−76.
Manuel Mendoza-Carranza a, *, Wendi Arévalo-Frías b, Alejandro Espinoza-Tenorio c, Chrystian C. Hernández-Lazo a, Arturo M. Álvarez-Merino a y Rocío Rodiles-Hernández d