
Spatial distribution of parasitic plants in Sierra de Quila, Jalisco
Miriam Alejandra Rosas-Sánchez, Sergio Franco-Mass, Angel Roberto Martínez-Campos, Angel Rolando Endara-Agramont *
Universidad Autónoma del Estado de México, Instituto de Ciencias Agropecuarias y Rurales, Instituto Literario Núm. 100, Colonia Centro, 50000 Toluca, Estado de México, México
*Autor para correspondencia: arendaraa@uaemex.mx (A.R. Endara-Agramont)
Recibido: 13 febrero 2024; aceptado: 11 julio 2025
Resumen
La extensión forestal de México es de 138.7 millones de ha, 42% de las cuales se encuentran afectadas por plantas parásitas. El Área de Protección de Flora y Fauna Sierra de Quila tiene 6,788 ha de bosques de pino-encino y encino-pino, que son susceptibles al ataque de plagas y enfermedades. El objetivo fue determinar su distribución espacial y niveles de infestación. Para ello, se establecieron 330 sitios de muestreo (SM) circulares de 1,000 m2, utilizando las curvas de nivel como transectos de acceso al bosque, con una separación altitudinal de 100 m entre curvas desde 2,000 hasta 2,600 m snm, con una equidistancia entre sitios de 300 m. En cada sitio se registraron datos físico-geográficos, dasonómicos y afectación por plantas parásitas. Más de 2,050 ha de sus bosques (30%) están afectadas por muérdago enano (Arceuthobium durangense) y diversos muérdagos verdaderos (Phoradendron bolleanum, P. brachystachyum, P. lanceolatum, P. reichenbachianum, Cladocolea cupulata, C. grahamii, C. mcvaughii, C. microphylla y Psittacanthus calyculatus), todos con niveles bajos de infestación, por lo que aún resulta oportuno establecer estrategias de manejo y control para evitar mayores daños al bosque.
Palabras clave: Muérdagos; Bosque pino-encino; Infestación
Abstract
The forested area of Mexico is 138.7 million ha, 42% of which are affected by parasitic plants. The Sierra de Quila Flora and Fauna Protection Area has 6,788 ha of pine-oak and oak-pine forests, which are susceptible to pests and diseases. The objective was to determine their spatial distribution and infestation levels. To this end, 330 circular sampling sites (SM) of 1,000 m2 were established, using contour lines as forest access transects. The elevations were separated by 100 m between contours from 2,000 to 2,600 m asl, and the distance between sites was 300 m. Physical-geographical and forestry data, as well as the impact on parasitic plants, were recorded at each site. More than 2,050 ha of its forests (30%) are affected by dwarf mistletoe (Arceuthobium durangense) and various true mistletoes (Phoradendron bolleanum, P. brachystachyum, P. lanceolatum, P. reichenbachianum, Cladocolea cupulata, C. grahamii, C. mcvaughii, C. microphylla, and Psittacanthus calyculatus), all with low levels of infestation, so it is still possible to establish management and control strategies to avoid further damage to the forest.
Keywords: Mistletoes; Pine-oak forest; Infestation
Introducción
México posee una extensión forestal de 138.7 millones de ha (Conafor, 2021). Éstas son susceptibles a disturbios; particularmente los ocasionados por insectos fitófagos, plantas parásitas e incendios (USDA, 2013). En particular, en Jalisco, durante el periodo 2012-2021 se reportaron 54,158 ha de bosque con diversos problemas fitosanitarios: plantas parásitas (61%), enfermedades (cancro resinoso, pudrición de raíz y roya; 19%), insectos descortezadores (13%) e insectos defoliadores (6%) (Semadet, 2022). Un factor ambiental que incrementa la vulnerabilidad de las especies forestales a plagas o enfermedades es el cambio climático (Bentz et al., 2010).
En México, se tienen registros de más de 200 especies de insectos y patógenos que provocan daños en los ecosistemas forestales (Conafor, 2007). En Jalisco, las plantas parásitas que se registran con mayor incidencia en los bosques son 4 especies de muérdagos enanos (Arceuthobium spp.) en coníferas y 21 especies de muérdagos verdaderos en latifoliadas (Semadet, 2021). La mayor riqueza de plantas parásitas se encuentra en las sierras de Tapalpa, Quila, Primavera, Cacoma, Manantlán (Vázquez-García et al., 1995), Bolaños (Vázquez-García et al., 2004), del Halo, Nevado de Colima, Cerro Grande y volcán Cerro Viejo, con 16 especies de plantas parásitas, el mayor número de especies encontradas para un municipio (Vázquez-García et al., 1995).
Los muérdagos son plantas hemiparásitas que dependen parcialmente de otros organismos para poder subsistir (Conafor, 2007; Nickrent y Musselman, 2004). Cuando se establecen en el tallo de su hospedero, compiten con él por agua y nutrimentos, lo que dificulta su crecimiento (Espinoza-Zúñiga et al., 2019). Por otra parte, cuando los muérdagos son abundantes en el árbol hospedero pueden provocarle la muerte (Alvarado-Rosales et al., 2007). Abarcan especies del género Arceuthobium, conocidas como muérdagos enanos, así como de los géneros Cladocolea, Phoradendron, Psittacanthus y Struthanthus, todos ellos conocidos como muérdagos verdaderos (Nickrent y Musselman, 2004). Son plantas arbustivas que se establecen en la parte aérea, más frecuentemente en ramas (muérdagos verdaderos) o en las ramas y en el fuste (muérdagos enanos), que afectan el desarrollo de su hospedero a través del sistema endofítico (Hawksworth, 1961; Rey et al., 1991), lo que induce cambios fisiológicos y bioquímicos (Mathiasen et al., 2008). Endara-Agramont et al. (2022) encontraron que algunas de las variables que influyen en la presencia de muérdago son la altitud y la orientación de ladera, y que su existencia en un árbol hospedero lo predispone al establecimiento de insectos descortezadores y plantas parásitas debido a que hay pérdida del vigor y reducción en su crecimiento, lo que genera mortalidad y facilita la dispersión de semillas (Cibrián-Tovar et al., 1995; Klutsch et al., 2014).
Las plantas parásitas tienen un valor ecológico. Son indicadoras de la salud del bosque, pues a mayores niveles de disturbio su prevalencia se incrementa (Cházaro et al., 1992). En contraste, promueven la diversidad de los bosques al proporcionar recursos alimentarios para algunas especies animales debido a la disponibilidad de sus frutos en épocas en las que otros alimentos son escasos (Matula et al., 2015).
La información sobre la distribución espacial de muérdagos en la sierra de Quila es escasa, a pesar de haber sido reportada como una zona con alta riqueza de plantas parásitas (Vázquez-García et al., 1995), por lo que el objetivo de esta investigación fue identificar las plantas parásitas y sus hospederos, así como determinar la distribución espacial y los niveles de infestación a través de un muestreo intensivo en esta área.
Materiales y métodos
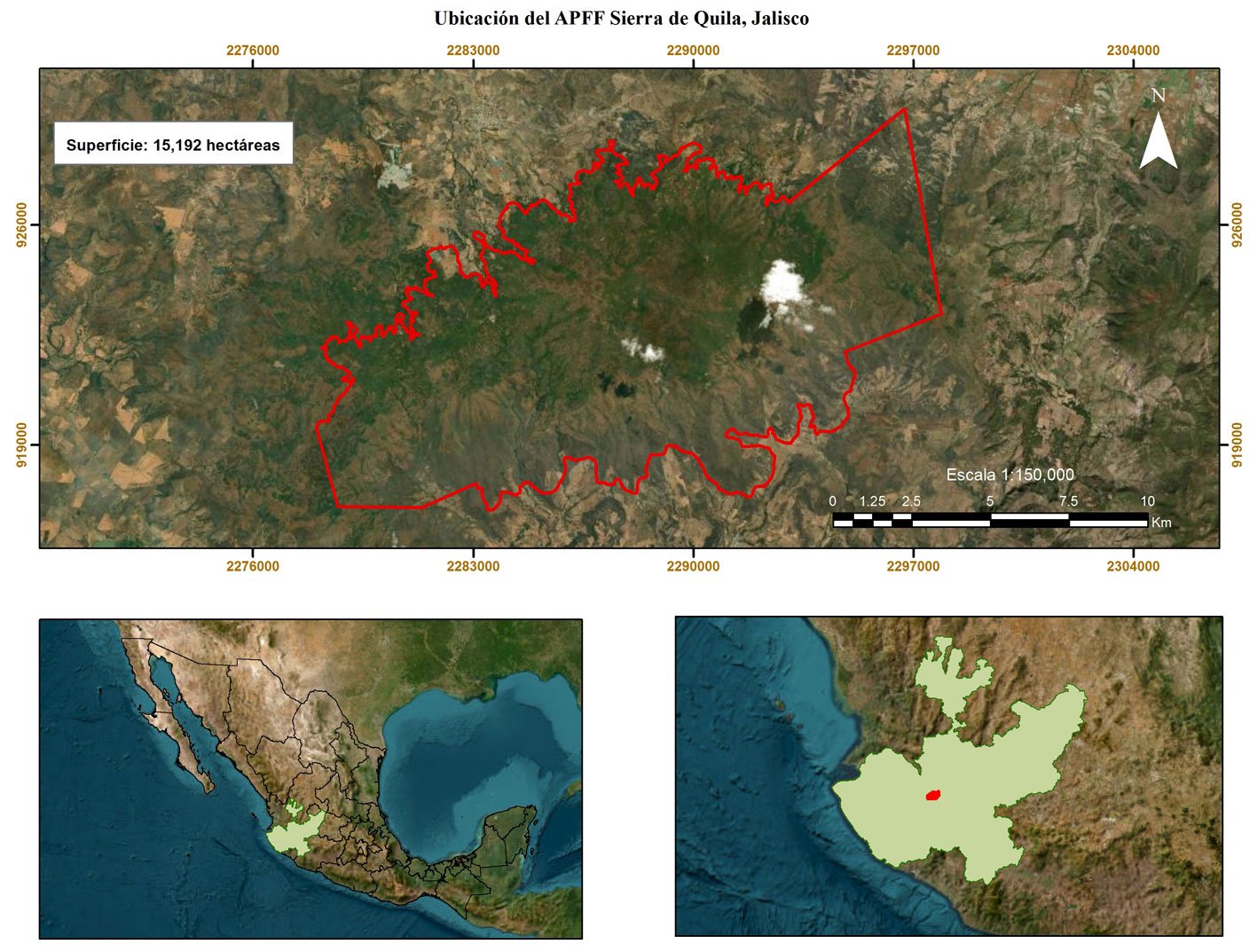
El estudio se realizó en el Área de Protección de Flora y Fauna (APFF) Sierra de Quila (fig. 1), en Jalisco entre los municipios de Tecolotlán, San Martín de Hidalgo y Tenamaxtlán, a 100 km al suroeste de la ciudad de Guadalajara, Jalisco (Villavicencio et al., 2005). Forma parte del Sistema Volcánico Transmexicano, incluida en la subprovincia “Sierras de Jalisco”, el relieve es montañoso con pendientes de 15 a 60° (Guerrero y Cházaro, 1995). La altitud oscila entre 1,350 y 2,560 m snm y de acuerdo con la clasificación de Köppen, el clima en la región es templado húmedo con lluvias en verano C(w2) (W) (Santiago et al., 2012). La alta variedad de condiciones geográficas y climáticas de esta sierra le permiten albergar una muestra representativa de la biodiversidad del occidente de México, con distintos tipos de bosques, que incluyen encino, pino-encino, encino-pino y selva baja caducifolia (Villavicencio etal., 2015) (fig. 1).
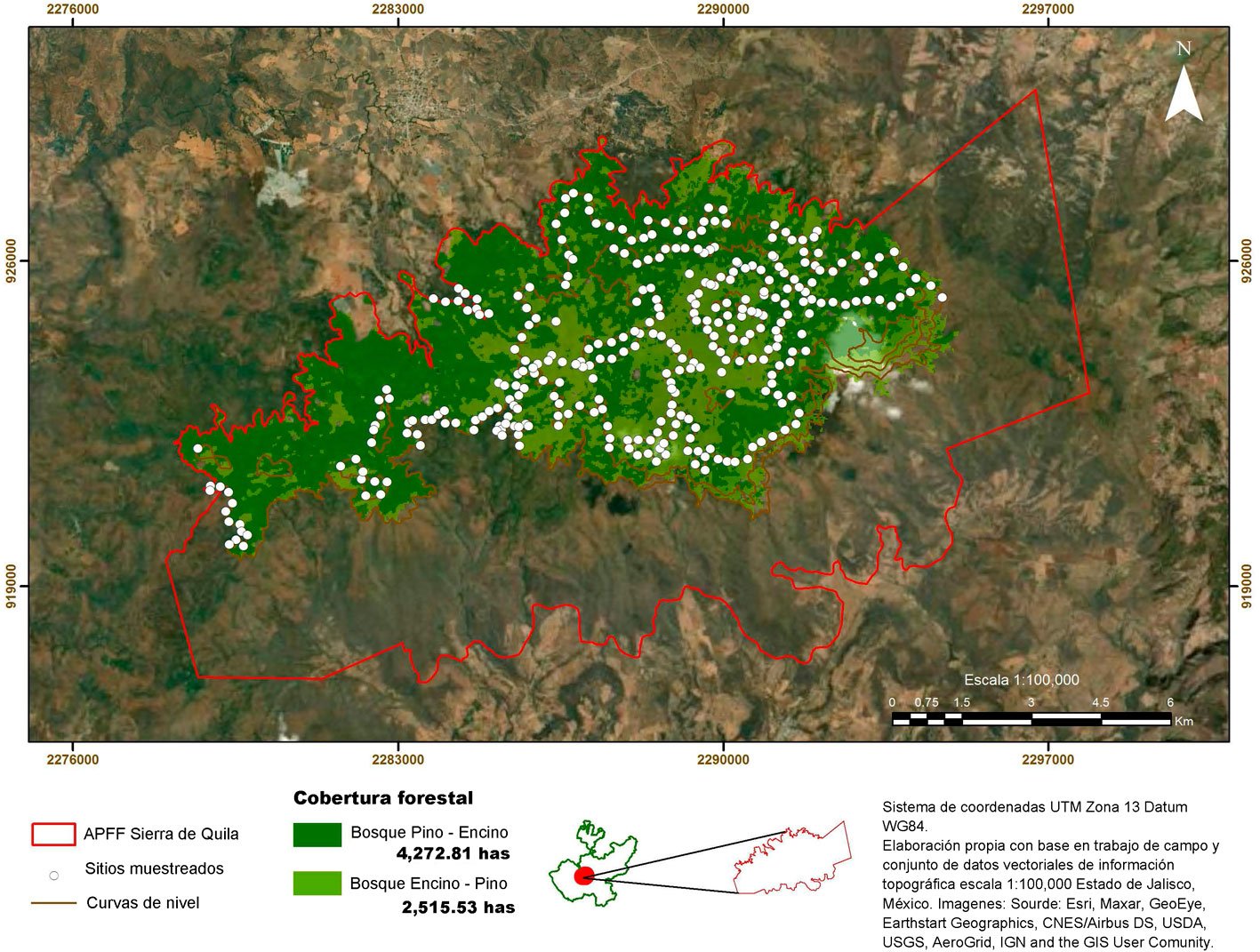
Este estudio se realizó en los bosques de pino-encino y encino- pino (6,788 ha); debido a que el mayor número de reportes de presencia de muérdago se encuentra en éstos (Cibrián-Tovar et al., 2007; Galván, 2016; Geils et al., 2002; Martínez-Ambriz, 2020; Ornelas, 2019; Rzedowski y Calderon de Rzedowski, 2011; Sandoval y Siqueiros, 2019). El muestreo se realizó de noviembre 2021 a noviembre 2022. Para ello, se establecieron 330 sitios de muestreo (SM) circulares de 0.1 ha (35.6 m de diámetro) establecidos sobre las curvas de nivel con una separación altitudinal de 100 m entre curvas (fig. 2); las cuales fueron ubicadas en 7 distintas altitudes fijadas entre 2,000 y 2,600 m snm. Lo anterior se hizo para considerar la variabilidad climática y los cambios de vegetación asociados a las diferencias de altitud (Mayer y Ott, 1991). Dentro de la misma cota altitudinal los sitios tuvieron una separación equidistante de 300 m, lo que permitió superar la intensidad mínima de muestreo recomendada de 0.28% (Dauber, 1995).
En cada SM se registraron las variables fisiográficas, las características de la vegetación (cobertura de pastos y arbustos) e información dasonómica de las especies arbóreas con un diámetro normal (DN) ≥ 2.5 (Endara-Agramont et al., 2022, 2023; Rojas-García et al., 2019) (tabla 1), así como, el nivel de infestación de los árboles con muérdago enano y verdadero (Alvarado-Rosales y Saavedra-Romero, 2005; Hawksworth, 1983) (tabla 2). Además, se recolectaron muestras botánicas de las plantas parásitas y del árbol hospedero, las cuales fueron identificadas mediante las claves taxonómicas de Alvarado-Cárdenas (2010), Martínez-Ambriz (2020) y Vázquez-Collazo et al. (2006), así como otros ejemplares en el Herbario CHAP, de la Universidad Autónoma Chapingo y el Herbario Nacional de México de la Universidad Nacional Autónoma de México, para poder rectificar que las muestras recolectadas coincidieran con otras previamente identificadas.
Se elaboró una base de datos que permitió generar los polígonos con la distribución espacial de muérdago a través del software ArGis ver. 10.8, en el cual se aplicó la metodología de interpolación de datos del vecino natural (Sibson, 1982), que consiste en generar polígonos con información específica (asignada a través de la generación de la base de datos) para formar superficies continuas (Childs, 2004; Pirotti y Tarolli, 2010).
Se realizó un Anova para determinar el efecto de la densidad del bosque sobre el porcentaje de cobertura y los individuos con muérdago con el programa Statgraphics Centurion XVI v.16.1.03 (StatPoint Technologies Inc., 2009). Finalmente, se realizó un gráfico descriptivo para analizar si existe relación entre la presencia de muérdago y la estructura arbórea.
Resultados
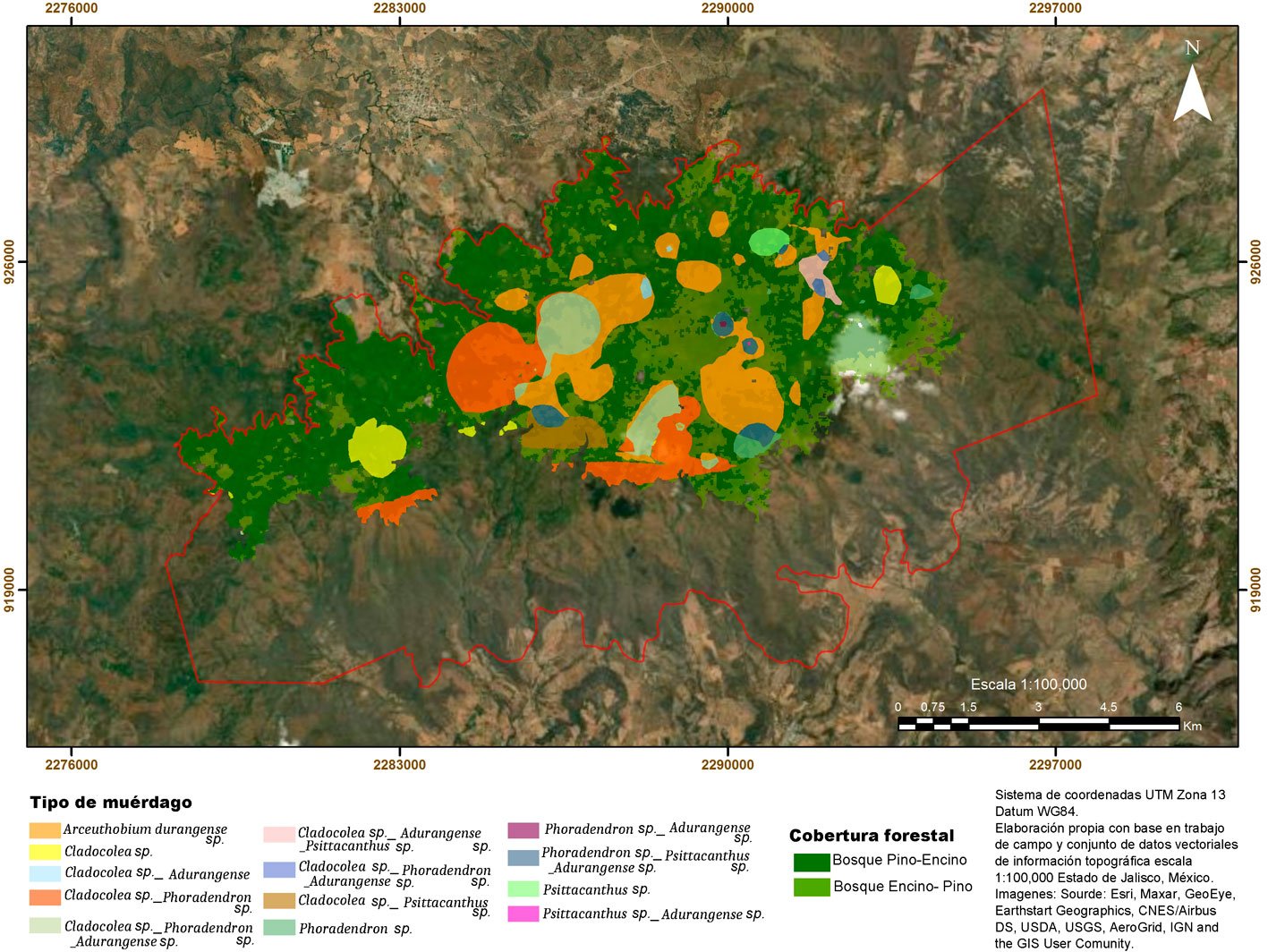
Se encontró que cerca de 40% de los SM tiene presencia de muérdagos, lo que corresponde a poco más de 2,050 ha afectadas. Las especies encontradas fueron: Arceuthobium durangense (A) (42%) sobre Pinus douglasiana y P. devoniana; Phoradendron bolleanum (B)(9%), P. brachystachyum (C)(2%), P. lanceolatum (D) (2%) y P. reichenbachianum (E) (2%) sobre Arbutus xalapensis, Quercus castanea, Q. laeta y Q. coccolobifolia; Cladocolea cupulata (F)(25%), C. grahamii (G) (5%), C. mcvaughii (H) (7%)y C. microphylla (I) (2%) sobre Pinus lumholtzii, P. oocarpa, Quercus coccolobifolia, Q. castanea, Q. resinosa, Q. rugosa y Q. crassipes; así como Psittacanthus calyculatus (J) (4%) sobre Quercus resinosa, Q. laeta y Q. rugosa (tabla 3), lo que muestra que estos bosques son dañados por una parte representativa de la diversidad de muérdagos presentes en Jalisco.
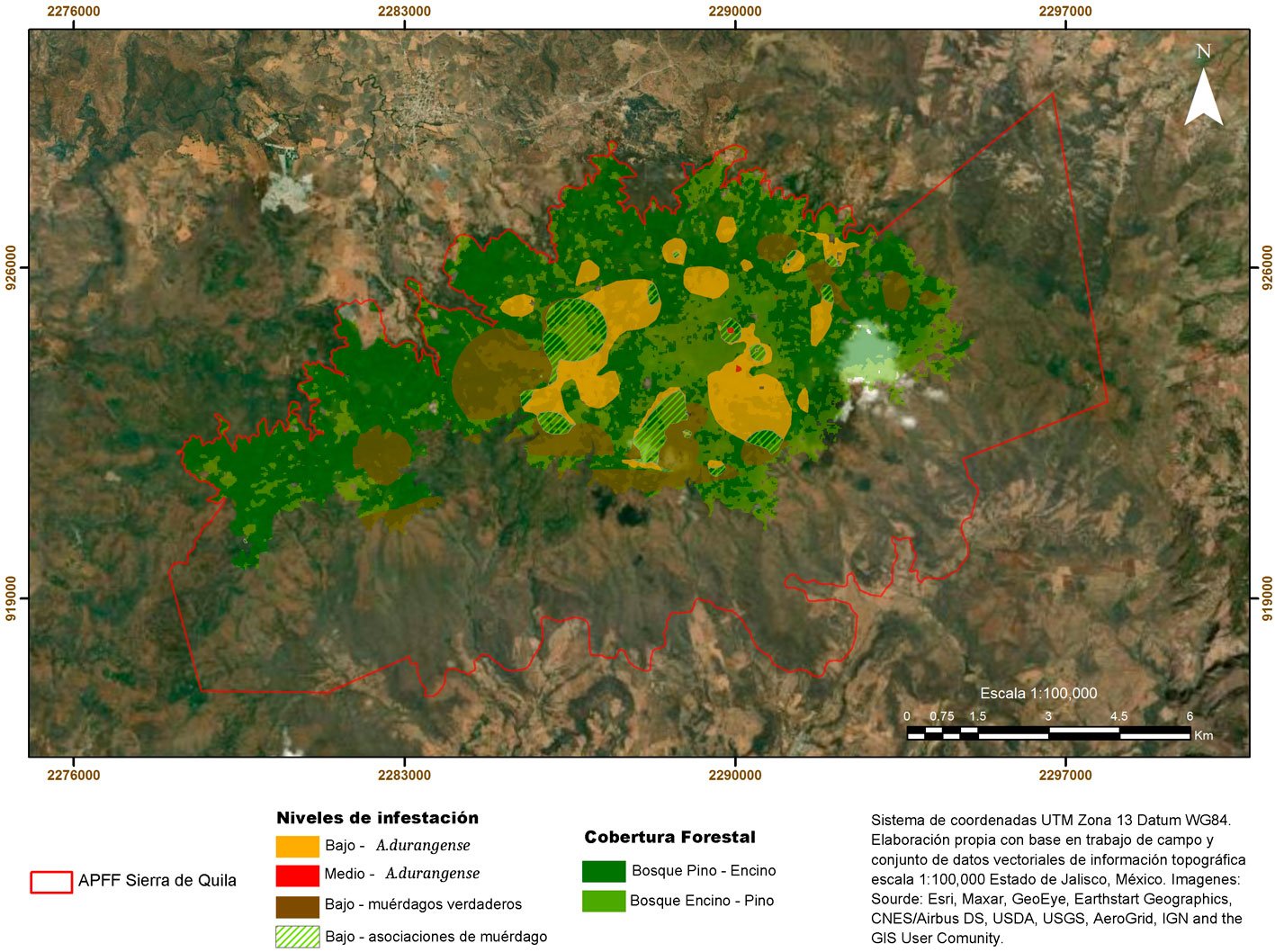
Asimismo, el muestreo señala que la distribución espacial de muérdagos enanos y verdaderos sobre los bosques de pino-encino y encino-pino de la sierra de Quila obedece patrones de distribución altitudinal para cada género de plantas parásitas: Cladocolea de 2,000 a 2,300 m, Arceuthobium de 2,100 a los 2,300 m, Phoradendron de 2,200 a 2,400 m y Psittacanthus únicamente en 2,400 m (fig. 3). Por otro lado, de acuerdo con la superficie afectada de la sierra de Quila, 35% presenta muérdago enano (A. durangense), 47% muérdago verdadero (Cladocolea, Phoradendron y Psitthacanthus) y 18% asociaciones de diferentes especies de muérdagos. Si bien, todas las especies de muérdago (A. durangense, Cladocolea, Phoradendron y Psitthacanthus) se encuentran en niveles bajos dentro de los hospederos (tabla 2), la distribución en términos de superficie es de 30% dentro de todo el bosque (fig. 4), pudiendo incrementar hacia otros hospederos y rodales.
Tabla 1
Variables de sitio e individuos; información recabada en los sitios de muestreo (SM).
| Fisiográficas | Vegetación | Características dasonómicas |
| Coordenadas (UTM) Ubicación (ladera, pie, cima) Pendiente (grados) Altitud (m snm) | Estructura (juvenil, senil, semillero, pastizal) | Diámetro a la altura del pecho (DAP) Altura total Adaptación de la escala del nivel de infestación para muérdagos enanos (Hawksworth, 1983) Nivel de infestación de muérdagos verdaderos (Alvarado-Rosales y Saavedra-Romero, 2005) Observaciones (daños por rayo, sin ápice o muertos) |
Tabla 2
Escalas de evaluación para muérdagos enanos y verdaderos.
| Escalas de evaluación de muérdagos enanos Adaptada de Hawksworth (1983) | Escalas de evaluación de muérdagos verdaderos Alvarado-Rosales y Saavedra-Romero (2005) |
| 0 Sano. No muestra rasgos de muérdago 1 Infestación ligera (50% o menos de las ramas infectadas) 2 Infestación severa (más de 50% de las ramas infectadas o en el fuste) | 0 Sano. Sin infección aparente 1 infección ligera (1-30%) 2 Infección media (31-60%) 3 Infección severa (61-90%) |
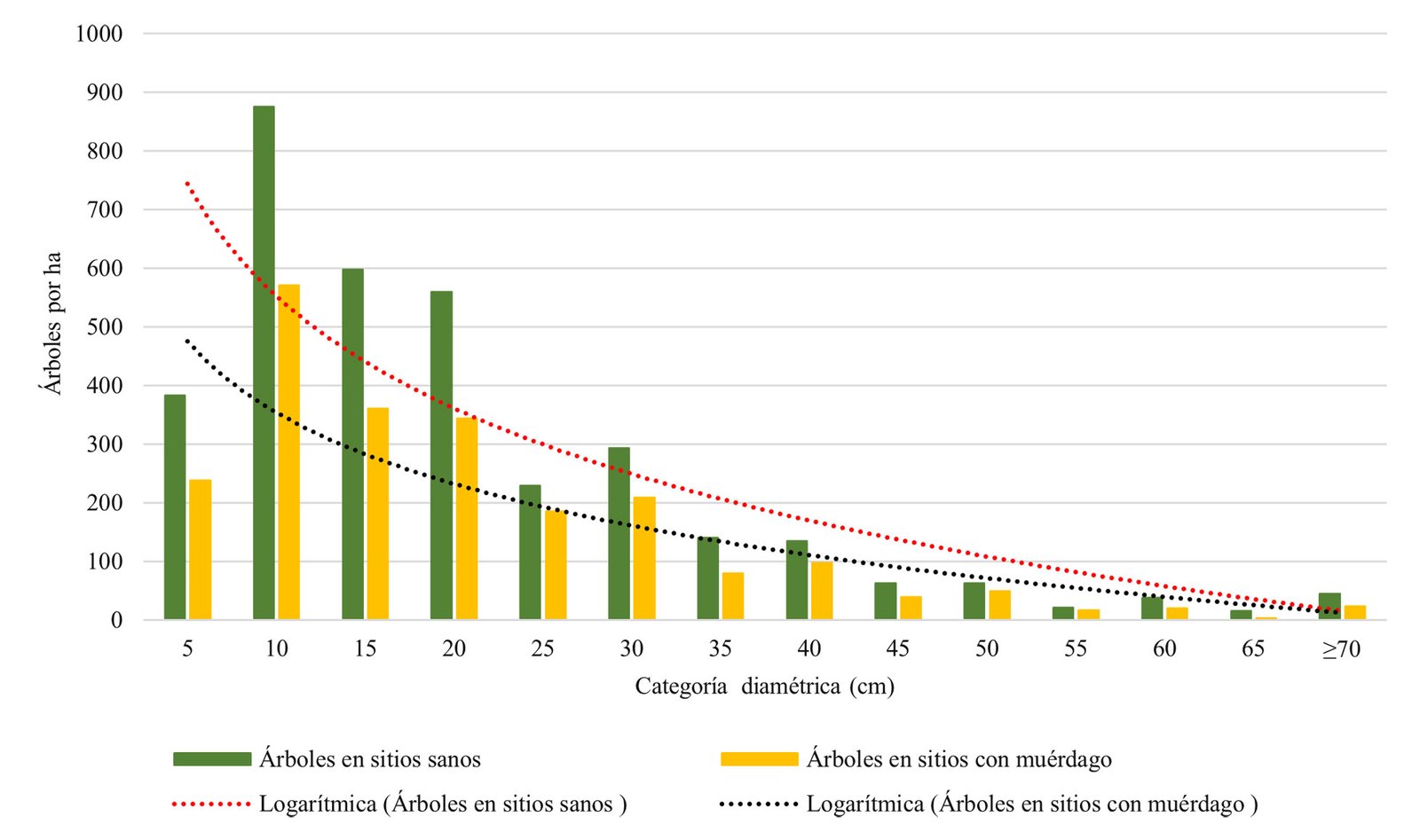
Otro factor que influye en la presencia de muérdagos es la densidad, lo que corrobora el Anova, donde se determinaron 3 categorías, denso, semidenso y fragmentado; el total de los árboles registrados fueron de 10,346 y se clasificaron de la siguiente manera: denso (≥ 361), semidenso (151-360) y fragmentado (30-150). (tabla 4). Estos resultados muestran que la presencia de muérdago se concentra en el bosque semidenso (13%) y fragmentado (34%) (fig. 5), observando estas afectaciones en el dosel dominante y codominante, donde se confirma que la reducción del número de árboles por unidad de superficie implica una mayor cantidad de luz al interior del bosque, relacionado con el incremento de las poblaciones de muérdago. Aun cuando el muérdago afecta generalmente a todas las categorías diamétricas, es más abundante entre los 5 y 40 cm de DAP (fig. 6). Del total de árboles registrados mencionados anteriormente, 10% está infestado por muérdago, lo que indica que los niveles se encuentran dentro de los límites del parasitismo (Mathiasen, 1998).
Discusión
El muestreo realizado indica un rango altitudinal que oscila entre 2,100 y 2,400 m snm, lo que coincide con Queijeiro-Bolaños et al. (2013) y Endara-Agramont et al. (2023), quienes mencionan que a mayor altitud existe menor número de las plantas parásitas; así mismo, la distribución de éstos obedece a la presencia de sus hospederos y a su capacidad de dispersión (Cibrián-Tovar et al., 2007). Estos registros muestran que Arceuthobium durangense es el único que presenta niveles de infestación medio, siendo más agresivo debido a que ataca en mayor medida al fuste en comparación con el muérdago verdadero (Hawksworth, 1961; Rey et al., 1991).
Cabe destacar que de los 122 SM con muérdago, 10% de los individuos se ve afectando principalmente a individuos juveniles de pinos y encinos (5 y 40 cm de DAP), lo que concuerda con Cibrián-Tovar et al. (2007), donde mencionan que el arbolado juvenil es el más susceptible al establecimiento de estas plantas parásitas. De acuerdo con la estructura y densidad del bosque, el dosel dominante y codominante compuesto por diversas especies de pinos y encinos y los rodales fragmentados son los más afectados, lo anterior coincide con lo reportado por Queijeiro-Bolaños y Cano-Santana (2015), quienes afirman que los muérdagos responden a presiones ambientales, donde la disminución de las densidad de los hospederos favorece la infestación debido a que estas condiciones son necesarias para la producción de estructuras vegetativas y reproductivas, teniendo menos competencia por recursos al tener una menor cantidad de árboles vecinos; por lo tanto, estos son directamente aprovechados por los muérdagos aumentando la probabilidad de infestación dentro de los individuos.
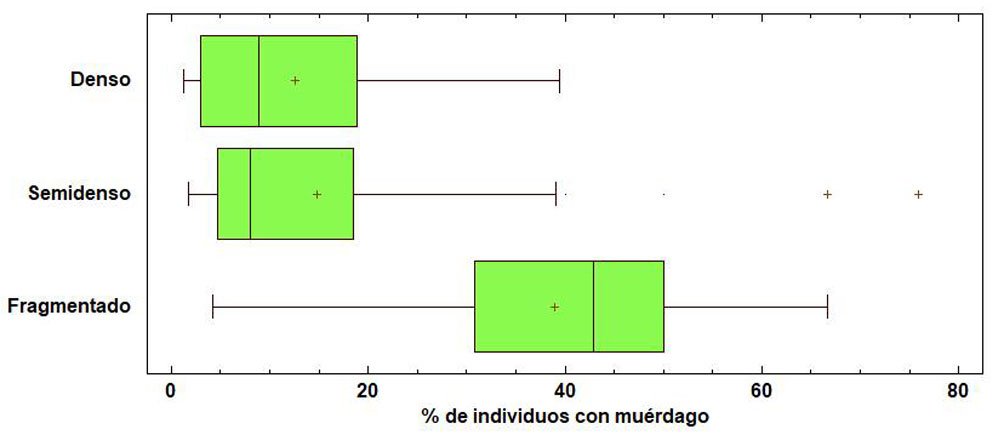

Tabla 4
Asociaciones arbóreas encontradas en el APFF Sierra de Quila, Jalisco.
| Asociaciones arbóreas | Densidad (núm./ha) | Especie | DAP Promedio (cm ± e.e.) | Altura promedio (m ± e.e.) | Dosel |
| Clethra sp. – Pinus sp. – Quercus sp. | 310 | Pinus sp. | 34 | 20 | Dominante |
| Quercus sp. | 24 | 16 | Codominante | ||
| Clethra sp. | 17 | 11 | Suprimido | ||
| Pinus sp. – Quercus sp. | 287 | Pinus sp. | 24 | 15 | Dominante |
| Quercus sp. | 18 | 11 | Codominante | ||
| Quercus sp. – Pinus sp. | 292 | Pinus sp. | 24 | 14 | Dominante |
| Quercus sp. | 17 | 11 | Codominante |
En este estudio se encontró que la altitud, la densidad de árboles, y el DN favorecen la prevalencia de muérdagos (Ferrenberg, 2020; Mathiasen, 2019), como consecuencia reducen el vigor de los árboles infestados, ésto coincide con lo encontrado por Cibrián-Tovar et al. (1995) y Endara-Agramont et al. (2023), quienes reportan que la pérdida del vigor y reducción en el crecimiento de árboles parasitados genera mayor probabilidad de muerte en los hospederos y facilita la infestación por descortezadores y/o enfermedades (Klutsch et al., 2014).
Hawksworth (1983) encontró que otro de los factores que afecta inversamente la prevalencia de los muérdagos es la densidad de árboles, a mayor densidad arbórea menor presencia de muérdago. En relación con ésto, se ha reportado que las semillas de estos organismos proliferan bajo un dosel forestal semidenso o fragmentado (Endara-Agramont et al., 2023). Más de 30% de la superficie de la sierra de Quila está infestada por muérdagos de 4 géneros (Arceuthobium, Cladocolea, Phoradendron y Psittacanthus), distribuidos entre 2,000 y 2,400 m snm, mismos que afectan a todas las categorías diamétricas, sobre todo los que están establecidos en bosques semidensos y fragmentados.
Agradecimientos
Agradecemos el apoyo financiero del proyecto Conafor-Conacyt A-S-130105 “Identificación, evaluación y manejo integrado de plantas parásitas en cuatro regiones de México: Noroeste (Durango), Centro-Occidente (Michoacán y Jalisco), Oriente (Puebla y Veracruz), Centro-Sur (Estado de México y Tlaxcala)”; a las brigadas contra incendios de las localidades de Quila, El Cobre, Mesa del Cobre y Lagunillas y al grupo de investigación de Bosques de Alta Montaña del Instituto de Ciencias Agropecuarias y Rurales.
Referencias
Alvarado-Cárdenas, L. (2010). Flora del valle de Tehuacán-Cuicatlán, fascículo 75, VISCACEAE Bastch. México D.F.: Universidad Nacional Autónoma de México.
Alvarado-Rosales, D. y Saavedra-Romero, L. (2005). El género Cladocolea (Loranthaceae) en México: muérdago verdadero o injerto. Revista Chapingo. Serie Ciencias Forestales y del Ambiente, 11, 5–9.
Alvarado-Rosales, D., Saavedra-Romero, L. y Almaraz-Sánchez, A. (2007). Agentes asociados y su papel en la declinación y muerte de encinos (Quercus, Fagaceae) en el centro-oeste de México. Polibotánica, 23, 1–21.
Bentz, B. J., Régniére, J., Fettig, C.J., Hansen, E. M., Hayes, J. L., Hicke, J. A. et al. (2010). Climate change and bark beetles of the Western United Satates and Canada: direct and indirect effects. BioScience, 60, 602–613. https://doi.org/10.1525/bio.2010.60.8.6
Cházaro, B., Huerta, M. M., Patiño, B. R., Sánchez, F. R., Lomelí, M. E. y Flores, M. A. (1992). Los muérdagos (Loranthaceae) de Jalisco, parásitas poco conocidas. Ciencia y Desarrollo, 17, 70–85.
Childs, C. (2004). Interpolating surfaces in ArcGIS spatial analyst. In ArcUser, ESRI, California, pp. 32-35.
Cibrián-Tovar, D., Alvarado-Rosales, D. y Gracia-Díaz, S. E. (2007). Enfermedades forestales en México. Chapingo, Estado de México: Universidad Autónoma de Chapingo.
Cibrián-Tovar, D., Méndez-Montiel, J. T., Campos-Bolaños, R., Tates III, H. O. y Flores-Lara, J. E. (1995). Insectos forestales de México. Chapingo, Estado de México: Universidad Autónoma de Chapingo.
Conafor (Comisión Nacional Forestal). (2007). Manual de sanidad vegetal. México D.F.: Conafor.
Conafor (Comisión Nacional Forestal). (2021). Estado que guarda el sector forestal en México. Bosques para el bienestar social y ambiental 202. México D.F.: Conafor.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2019). División política estatal, escala 1:250000. Catálogo de metadatos geográficos. México D.F.
Dauber, E. (1995). Guía práctica y teórica para el diseño de un inventario forestal de reconocimiento. Santa Cruz. BOLFOR.
Endara-Agramont, A, R., Heredia-Bobadilla, R. L., García-Almaraz, L. A., Luna-Gil, A. A y Aguirre-Zúñiga, J. J. (2023). Distribución espacial del descortezador Dendroctonus adjunctus Blandford, 1897 (Coleoptera: Curculionidae, Scolytinae) en dos bosques de alta montaña del centro de México. Acta Zoológica Mexicana, 39, 1–17. https://doi.org/10.21829/azm.2023.3912569
Endara-Agramont, A. R., Calderón-Contreras, R., Nava-Bernal, G. y Franco-Mass, S. (2013). Analysis of fragmentation processes in high-mountain forests of the Centre of México. American Journal of Plant Sciences, 4, 697–704. https://doi.org/10.4236/ajps.2013.43A088
Endara-Agramont, A. R., Heredia-Bobadilla, R. L., García-Almaraz, L. A., Luna-Gil, A. A., Franco-Mass, S. y Cibrián-Llanderal, V. D. (2022). Factores asociados con la distribución espacial de muérdagos enanos en dos poblaciones de Pinus hartwegii del centro de México. Revista Mexicana de Biodiversidad, 93, e935008. https://doi.org/10.22201/ib.20078706e.2022.93.5008
Espinoza-Zúñiga, P., Ramírez-Dávila, J. F., Cibrián-Tovar, D., Villanueva-Morales, A., Cibrián-Llanderal, V. D., Figueroa-Figueroa, D. K. y Rivera-Martínez, R. (2019). Modelación de la distribución espacial del muérdago (Santalales: Loranthaceae) en las áreas verdes de la delegación Tlalpan, México. Bosque,40, 17–28. https://doi.org/10.4067/S0717-92002019000100017
Ferrenberg, S. (2020). Dwarf mistletoe infection interacts with tree growth rate to produce opposing direct and indirect effects on resin duct defenses in lodgepole pine. Forests, 11, 222. https://doi.org/10.3390/f11020222
Galván, G. L. (2016). El género Cladocolea Tiegh. (Loranthaceae) en México (Tesis de maestría). Universidad Autónoma Metropolitana, Unidad Iztapalapa. México D.F.
Geils, B. W y Vázquez, C. (2002). Loranthaceae and Viscaceae in North America. En B. W. Geils, J. C. Tovar y B. Modoy (Coords.), Mistletoes of North America conifers. (pp. 1–5). Gen. Tech. Rep. RMRS-GTR-98. U. S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, Utah.
Guerrero, N. J. y Cházaro, B. (1995). Datos generales sobre la Sierra de Quila, Jalisco. En M. Cházaro Basáñez, E. Lomelí-Mijes, R. Acevedo-Rosas y S. Ellerbracke-Román (Compiladores) Antología botánica del estado de Jalisco (pp. 70–73). Guadalajara: Universidad de Guadalajara.
Hawksworth, F. (1961). Dwarf mistletoe of ponderosa pine in the Southwest. Technical Bulletin, 1246. USDA, Washington DC.
Hawksworth, F. (1983). Mistletoes as forest parasites. En M. Calder y P. Bernhardt (Eds.), The biology of mistletoes (pp. 320–329). Nueva York: Academic Press.
Klutsch, J. G., Beam, R. D., Jacobi, W. R. y Negrón, J. F. (2014). Bark beetles and dwarf mistletoe interact to alter downed woody material, canopy structure, and stand characteristics in northern Colorado ponderosa pine. Forest Ecology and Management, 315, 63–71. https://doi.org/10.1016/j.foreco.2013.12.024
Martínez-Ambriz, E. (2020). Familia Loranthaceae, fascículo 214. Flora del Bajío y Regiones adyacentes. Pátzcuaro, Mich.: Instituto de Ecología, A.C.
Mathiasen, R. L. (1998). Comparative susceptibility of conifers to larch dwarf mistletoe in the Pacific Northwest. Forest Science, 44, 559–568. https://doi.org/10.1093/forest
science/44.4.559
Mathiasen, R. L. (2019). Susceptibility of red fir and white fir to fir dwarf mistletoe (Arceuthobium abietiunum) in California. Forest Pathology, 2019, e12516.
Mathiasen, R. L., Nickrent, D., Shaw, D. y Watson, D. (2008). Mistletoes. Pathology, systematics, ecology, and management. The American Phytopathological Society, 92, 988–1006. https://doi.org/10.1094/pdis-92-7-0988
Matula, R., Svátek, M., Pálková, M., Volařík, D. y Vrška, T. (2015). Mistletoe infection in an oak forest is influenced by competition and host size. Plos One, 10, 1–11. https://doi.
org/10.1371/journal.pone.0127055
Mayer, H. y Ott, E. (1991) Gebirgswaldabau-Schutzwaldpflege: ein waldbauulicher Beitrag zur Landschaftssokologie und zum Umweltschutz (Silviculture in Mountain Forest-Management of protection forest: A silvicultural contribution to landscape ecology and enviromental protection), 2nd revised Edn. Stuttgart: Gustav Fischer.
Nickrent, D. L. y Musselman, L. J. (2004). Introduction to parasitic flowering plants. The plant health instructor. Heide-Jorgensen HS (Ed.) Brill. https://doi.org/10.1094/PHI-I-2004-0330-01
Ornelas, J. F. (2019). Los muérdagos Psittacanthus en México: ecología, evolución, manejo y conservación. Biodiversitas, 146, 12–16.
Pirotti, F. y Tarolli, P. (2010). Suitability of LiDAR point density and derived landform curvature maps for channel network extraction. Hydrological Processes, 24, 1187–1197. https://doi.org/10.1002/hyp.7582
Queijeiro-Bolaños, M. E. y Cano-Santana, Z. (2015). Dinámica temporal de la infestación por muérdago enano (Arceuthobium globosum y A.vaginatum) en Zoquiapan (Parque Nacional Iztaccíhuatl Popocatépetl), México. CienciaUAT, 9, 6–14.
Queijeiro-Bolaños, M. E., Cano-Santana, Z. y Castellanos-Vargas, I. (2013). Does disturbance determine the prevalence of dwarf mistletoe (Arceuthobium, Santalales: Viscaceae) in Central Mexico? Revista Chilena de Historia Natural, 2, 181–190. https://doi.org/10.4067/s0716-078×2013000200007
Rey, L., Sadik, A., Fer, A. y Renaudin, S. (1991). Trophic relations of the dwarf mistletoe Arceuthobium oxycedri with its host Juniperus oxycedrus. Journal of Plant Physiology, 138, 411–416. https://doi.org/10.1016/S0176-1617(11)80515-8
Rojas-García, F., Fredericksen, T. S., Vázquez, L. S. y Endara, A. A. R. (2019). Impact of timber harvesting on carbon storage in montane forests of central Mexico. New Forest, 50, 1043–1061. https://doi.org/10.1007/s11056-019-09714-z
Rzedowski, J. y Calderón-de Rzedowski, C. (2011). Dos especies notables de Phoradendron (Viscaceae) de la mixteca Oaxaqueña (México), una nueva y una complementada. Acta Botanica Mexicana, 1, 3–10. https://doi.org/10.21829/abm96.2011.254
Sandoval, O. M. y Siqueiros, D. M. (2019). Cladocolea loniceroides, un nuevo registro para la flora de Aguascalientes, México. Investigación y Ciencia de la Universidad Autónoma de Aguascalientes, 27, 51–54. https://doi.org/10.33064/iycuaa2019782232
Santiago, P. A. L., Domínguez, M. L., Rosas, V. E. y Rodríguez, J. M. (Coords.). (2012). Anfibios y reptiles de las montañas de Jalisco: Sierra de Quila. Orgánica Editores/ Conabio. Guadalajara, Jal., México.
SEMADET (Secretaría de Medio Ambiente y Desarrollo Territorial). (2021). Diagnóstico y plan de acción para la atención a plagas y enfermedades forestales en el Estado de Jalisco. Recuperado el 23 de noviembre, 2023 de: http://sivicoff.cnf.gob.mx/ContenidoPublico/02%20Informes%20de%20acciones%20operativas/DiagnosticosEstatales/2021/Jalisco.pdf
SEMADET (Secretaría de Medio Ambiente y Desarrollo Territorial). (2022). Programa operativo estatal de sanidad forestal para el estado de Jalisco. Recuperado el 23 de noviembre, 2023 de: http://sivicoff.cnf.gob.mx/ContenidoPublico/02%20Informes%20de%20acciones%20operativas/DiagnosticosEstatales/2022/Jalisco.pdf
Sibson, R. (1982). A brief description of natural neighbor interpolation. En John Wiley y Sons (Eds.), Interpolating multivariate data (pp. 21–36). Nueva York: John Wiley & Sons.
StatPoint Technologies, Inc. (2009). Version, 16, 17. Statgraphics Centurion, X. V. I.
USDA (United States Departament of Agriculture). (2013). National strategy framework for invasive species management. Forest Service. Washington: United States Department of Agriculture. Recuperado el 14 de diciembre, 2023 de: https://www.fs.usda.gov/foresthealth/publications/Framework_for_Invasive_Species_FS-1017.pdf
Vázquez-Collazo, I., Villa, R. y Madrigal, H. (2006). Los muérdagos (Loranthaceae) en Michoacán. Libro TécnicoNúm. 2. Mich. Uruapan, Michoacán: CIRPAC/ INIFAP.
Vázquez-García, J. A., Cházaro B., Nieves H., Vargas-Rodríguez, M., Vázquez-García, A. y Flores, M. (2004). Flora del norte de Jalisco y etnobotánica huichola. Guadalajara: Universidad de Guadalajara.
Vázquez-García, J. A., Cuevas, G. R., Cochrane, T. S., Litis, H. H., Santana, M. F y Guzmán, H. L. (1995). Flora de Manantlán: plantas vasculares de la Reserva de la Biósfera Sierra de Manantlán Jalisco-Colima, México. Sida, Botanical Miscellany, 13, 1−312.
Villavicencio, G. R., Ávila, C. R., Treviño, G. E. y Muñiz, C. M. (2015). Cartografía de la cobertura vegetal y uso actual del suelo de la Sierra de Quila, Jalisco, México. Memorias de resúmenes en extenso SELPER-XXI-México-UACJ-2015.
Villavicencio, G. R., Bauche, P., Gallegos, R., Santiago, P. y Huerta, M. (2005). Característica estructural y diversidad de comunidades arbóreas de La Sierra de Quila. Boletín del Instituto de Botánica de la Universidad de Guadalajara, 13, 67−76.


