Filogenia basada en COI y caracterización morfológica de una población brasileña de Lambornella trichoglossa (Ciliophora: Tetrahymenidae)
Vítor Ribeiro-Halfeld *
Universidade Federal de Juiz de Fora, Laboratório de Protozoologia, Rua José Lourenço Kelmer, s/n, São Pedro, Juiz de Fora, 36033-900 Minas Gerais, Brazil
In this work, the morphological characterization and molecular phylogeny (SSU rDNA and COI) of Lambornella trichoglossa Foissner, 2003, found in phytotelmata environments in southeastern Brazil is presented. The morphological study performed with silver impregnation techniques demonstrated parameters similar to those recorded in the type population, and additional new information was also obtained, including the conjugation process and the occurrence of specimens with an increased number of caudal cilia. The phylogenetic reconstructions based on both markers suggest the inclusion of L. trichoglossa in the genus Tetrahymena Furgason, 1940. The COI sequence obtained in this work is the first mitochondrial sequence of the genus Lambornella deposited in GenBank and published in a scientific article.
En este trabajo se realizó la caracterización morfométrica y la filogenia molecular (SSU rDNA y COI) de Lambornella trichoglossa Foissner, 2003, encontrada en ambientes de fitotelmata en el sureste de Brasil. El estudio morfológico realizado con técnicas de impregnación de plata demostró parámetros similares a los registrados en la población tipo y también se obtuvo nueva información adicional, incluyendo el proceso de conjugación y la presencia de especímenes con un mayor número de cilios caudales. Las reconstrucciones filogenéticas basadas en ambos marcadores sugieren la inclusión de L. trichoglossa en el género Tetrahymena Furgason, 1940. La secuencia COI obtenida en este trabajo es la primera de tipo mitocondrial del género Lambornella depositada en GenBank y publicada en un artículo científico.
The genus Lambornella has its systematic history marked by alterations. The type species, Lambornellastegomyiae Keilin, 1921, is a mosquito (Diptera, Culicidae) parasite. It was reclassified in the genus Tetrahymena Furgason, 1940 by Corliss (1960). However, the description of Lambornellaclarki Corliss, 1976, another parasitic species, revalidated the genus, indicating the formation of cuticular cysts and the number of post-oral kineties as diagnostic features. Nevertheless, Strüder-Kypke et al. (2001) suggested the non-validity of the genus Lambornella once again, due to its position within the Tetrahymena clade, based on phylogenetic analyses inferred from small subunit ribosomal DNA (SSU). Results presented by Bourland and Stüder-Kypke (2010), and Dunthorn et al. (2012), also based on SSU sequences, demonstrated a similar grouping.
Lambornella trichoglossa Foissner, 2003 is the only free-living species of the genus. It occurs endemically in phytotelmata environments (Foissner, 2003). Despite the recurrent records in the Neotropical region (Buosi et al., 2014, 2015; Durán-Ramírez et al., 2015; Foissner, 2003; Foissner et al., 2003), no studies have investigated Lambornella’s phylogenetic position based on mitochondrial markers. According to Chantangsi and Lynn (2008), the mitochondrial gene cytochrome c oxidase-subunit I (COI) is effective in elucidating recent phylogenetic events in the genus Tetrahymena. In this context, the present work aimed to investigate the molecular phylogeny of L. trichoglossa based on the mitochondrial marker COI. Morphological analysis of the specimens was also performed in order to confirm specific identification and investigate possible morphological variation.
Materials and methods
One hundred milliliters of phytotelmata content were collected from 12 bromeliads belonging to Portea petropolitana at the Botanical Garden of the Federal University of Juiz de Fora (21°43’74” S, 43°22’06” W), in September, 2019. On the same day of collections, the samples were analyzed under a stereoscopic microscope with transmitted light. Active ciliates were picked with glass micropipettes and processed, according to Foissner (2014), to perform silver carbonate and dry silver nitrate impregnation techniques. From the collection day, over a period of 7 days, 20 specimens of L. trichoglossa were screened from the samples and measured in vivo to check for possible alterations in body length. Obtained data were analyzed using the Shapiro-Wilk normality test, and the means recorded on the first and seventh days were compared using the Student’s t-test. All statistical analyses related to the morphological data were conducted using PAST software, version 4.03 (Hammer et al., 2001).
Thirty specimens of L. trichoglossa were picked from the samples and fixed in absolute ethanol for molecular analysis. Total DNA extraction was performed using the Blood and Tissue kit (Qiagen®), following the manufacturer’s guidelines. Primers F388dT and R1184dT (Strüder-Kypke & Lynn, 2010) were used to amplify the COI gene in 25-microliter reactions. The SSU marker was also sequenced for comparison with the type population. Therefore, primers 18S F9Euk and 18S R1513 (Schrallhammer et al., 2013) were used.
PCR products were visualized in a 1% agarose gel and purified using the QIAquick PCR Purification Kit (Qiagen®), following the manufacturer’s guidelines. Subsequently, the material was sent for sequencing according to the Sanger method in 7 µl reactions, using the M13 forward and M13 reverse primers (Messing, 1983) for COI, and 18S R536, 18S F783, 18S F919, and 18S R1052 for SSU (Modeo et al., 2006). Sequencing reactions were performed on an ABI PRISM® 3100 sequencer (Applied Biosystems).
Obtained sequences were added to the respective datasets, jointly with COI and SSU sequences of approximately 20 species of the genus Tetrahymena, as well as outgroups obtained from GenBank accessed in May 2025. Sequences were aligned using MAFFT software, version 7 (Katoh et al., 2017). The resulting alignment was edited in the GBlocks platform, version 0.91b (Talavera & Catresana, 2007). The determination of the best nucleotide substitution model (GTR + G + I to COI, and TN93 + G to SSU) was performed with the aid of MegaX software (Kumar et al., 2018), using the maximum likelihood method, considering all sites. The maximum likelihood phylogenetic analysis was inferred using RAxML software, version 8 (Stamatakis, 2014), at its default settings, with 500 bootstrap replicates for COI, and 1,200 for SSU. The evolutionary distances of COI and SSU sequences were computed in Mega X, using the Kimura 2-parameter method (Kimura, 1980). A third dataset with 74 COI sequences of Tetrahymena species was prepared, in addition to outgroups, with the aim of performing the pairwise distance calculations with a greater number of species (Supplementary material). For this analysis, the Maximum Composite Likelihood model (Tamura et al., 2004) was used, with the aid of MegaX (Kumar et al., 2018).
Results
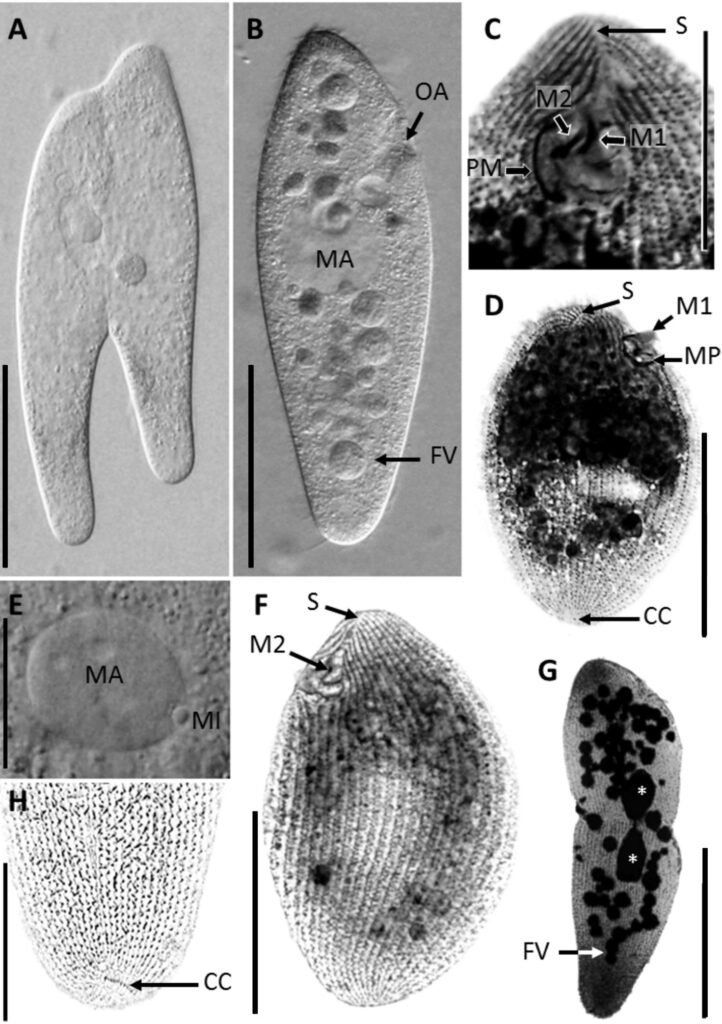
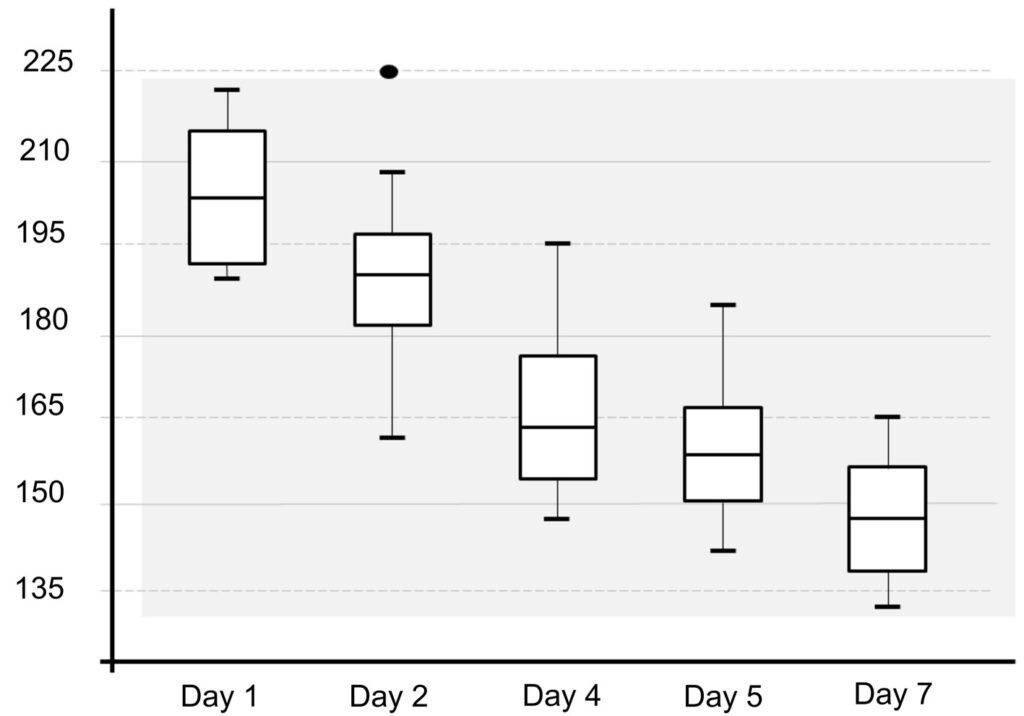
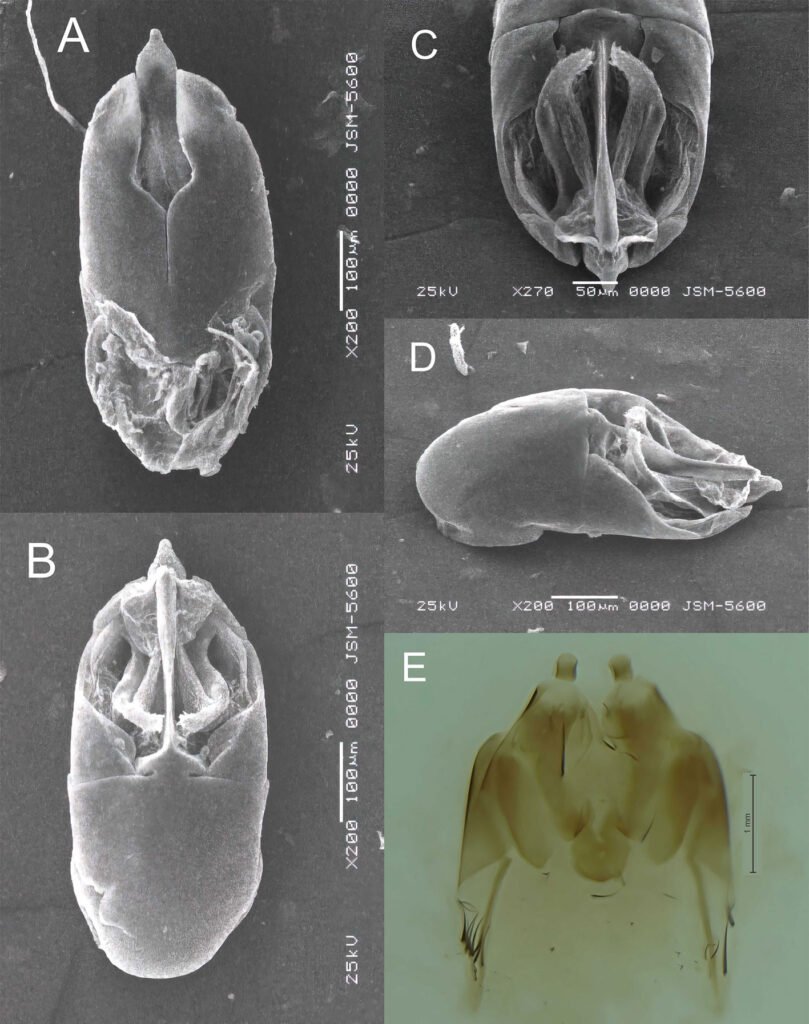
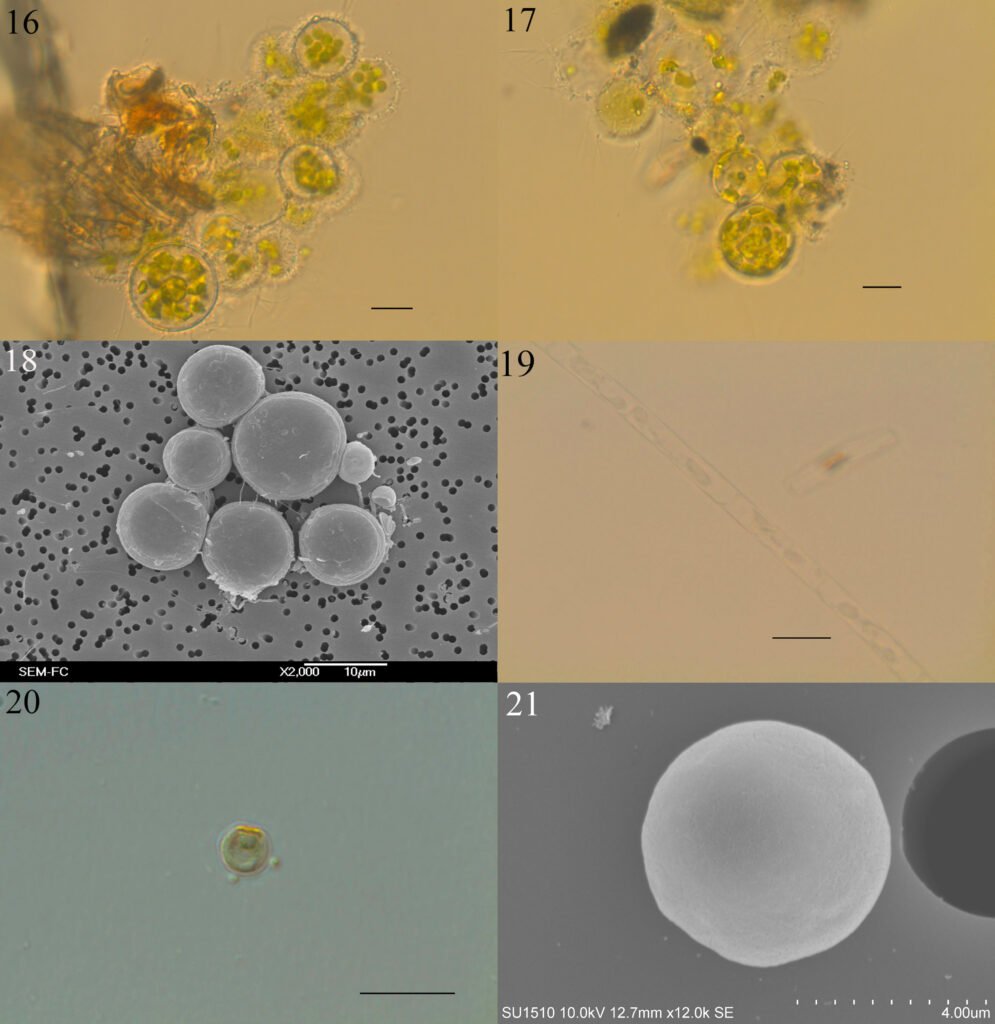
Table 1 presents the morphological data obtained. Morphological characterization of the L. trichoglossa population analyzed in this study resembles the type population described by Foissner (2003), presenting slightly higher values for body size and the number of somatic kineties. However, postoral kineties appear in lower numbers. Another important morphological characteristic distinct from the type population detected in the present study was the occurrence of specimens with a greater number of caudal cilia (Fig. 1h). These organisms have a complex of caudal cilia, as shown in Figure 1h and Supplementary material. However, they were not included in the morphological statistical analysis due to the unsatisfactory results of the impregnation techniques in their oral and some somatic structures. Opportunely, conjugating forms of L. trichoglossa were recorded, which is unprecedented for the species (Fig. 1a). The temporal analyses on body length performed demonstrated a significant decrease in the average size of the organisms over 7 days (Fig. 2, Table 2).
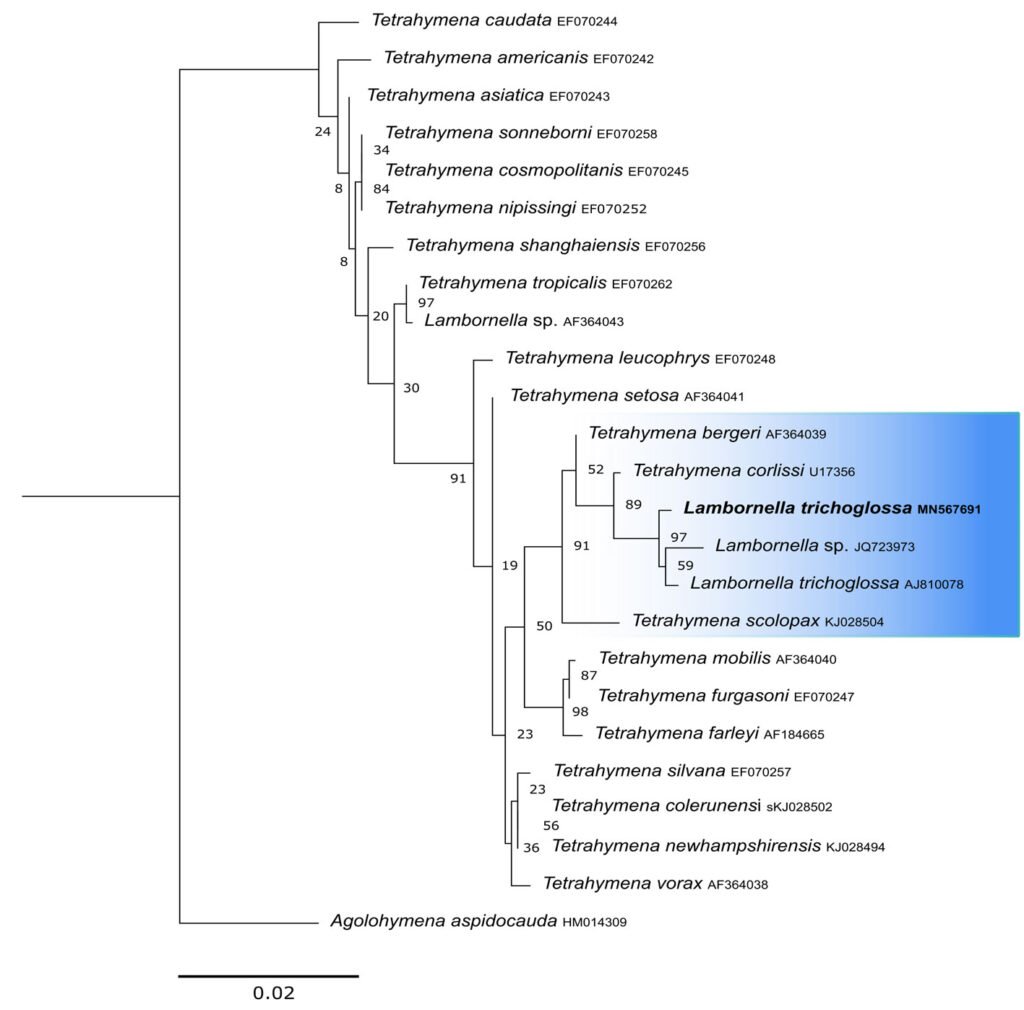
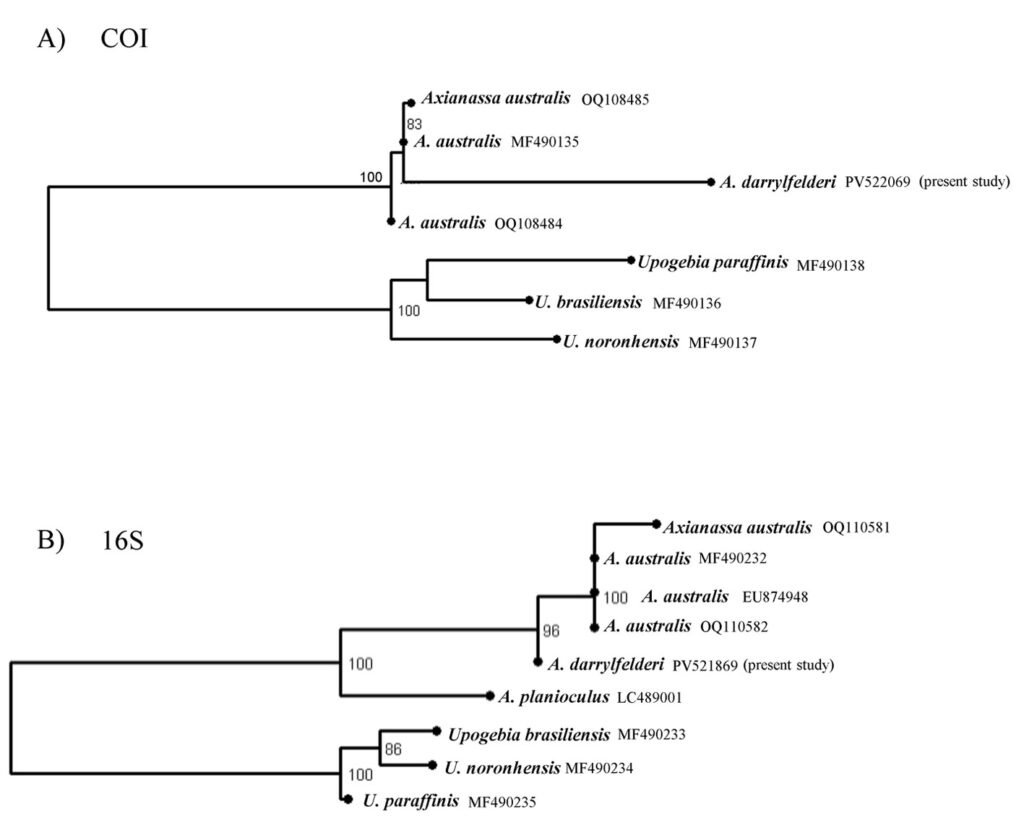
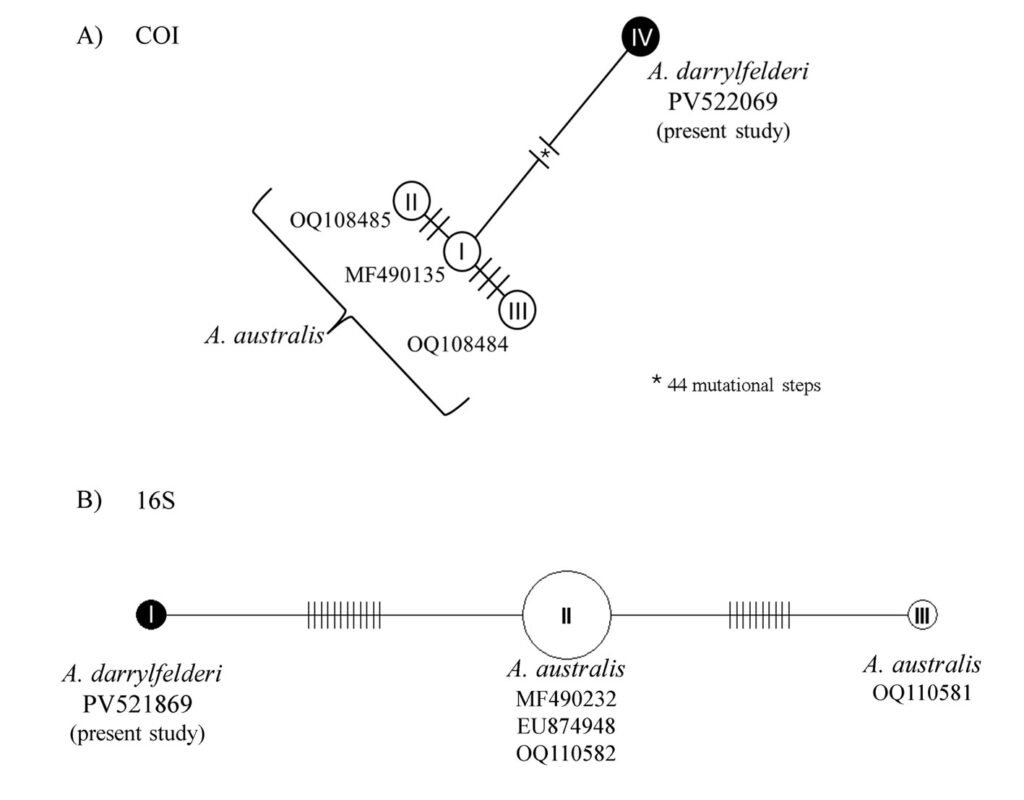
The obtained SSU sequence was deposited in GenBank with accession number MN567691. It matches 99.97% of the type population’s sequence (AJ810078) (Table 3). The phylogenetic reconstruction inferred by the maximum likelihood method revealed that the new sequence is grouped with L. trichoglossa (AJ810078), T. corlissi (U17356), and T. berger (AF364039) (Fig. 3).
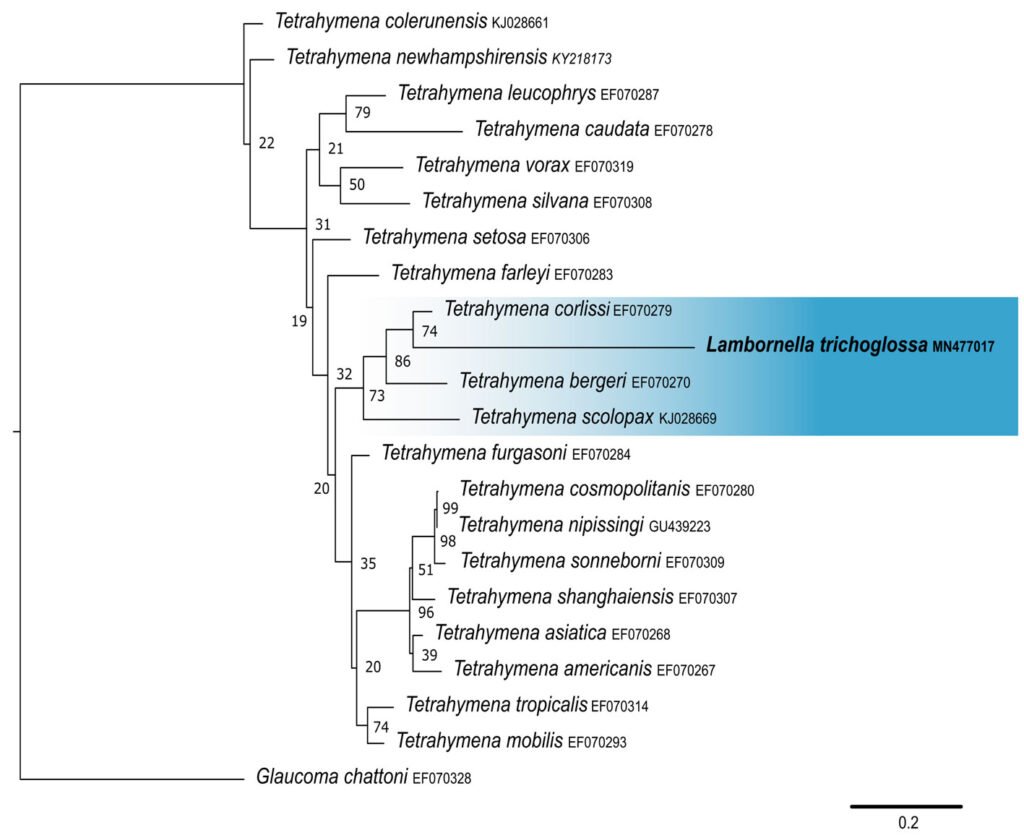
Lambornella trichoglossa’s COI sequence was deposited in GenBank with accession number MN477017. It has 774 nucleotides, with the following percentage of bases: A = 33%, C = 12%, G = 13.4%, and T = 41.6%. Phylogenetic reconstruction using the maximum likelihood method demonstrated L. trichoglossa clustering in a clade with the same topology revealed by the SSU phylogeny; it also grouped with T. corlissi (EF070279) and T. bergeri (EF070270) with a support value of 86%, besides T. scolopax (KJ028669) (Fig. 4). The average paired difference of the 74 Tetraymena sequences used in the phylogenetic analyses was 12.99%. The L.trichoglossa sequence differs by 13.04% from the T.corlissi sequence (Table 4). Although the value is slightly above the average for the Tetrahymena species analyzed, this percentage is lower than the difference observed between T. corlissi and T. caudata (14.29%), T.paravorax (14%), and T. glochidiophila (16.39%), for example.
Table 1
Morphological data on silver carbonate-impregnated specimens of Lambornella trichoglossa.
x
M
SD
SE
CV
Min
Max
n
Body, lenght
269.75
269.43
26.72
4.88
9.906
226.65
327.91
30
Body, width in ventral view
73.105
73.295
16.05
2.93
21.96
47.44
107.77
30
Body lenght: Lateral width, ratio
3.82
3.906
0.81
0.14
21.19
2.606
5.36
30
Anterior body end to membranelle 1, distance
47.91
48.73
7.407
1.35
15.46
34.14
63.87
30
Oral opening, width
32.45
31.42
5.11
0.93
15.77
21.28
44.12
30
Anterior body end to macronucleus, distance
94.55
92.88
13.86
2.53
14.66
74.16
126.35
30
Macronucleus, length
49.72
43.96
11.48
2.09
23.08
36.92
77.75
30
Macronucleus, width
51.27
48.71
9.42
1.72
18.38
33.74
66.86
30
Macronucleus, number
1
1
0
0
0
1
1
30
Micronucleus, width
5.67
5.5
1.15
0.21
20.408
4
9.56
27
Micronucleus, number
1.03
1
0.19
0.035
18.68
1
2
27
Somatic kineties, total number
52.33
52.5
3.79
0.69
7.24
45
62
30
Somatic kineties, postoral number
6.1
6
0.84
0.15
13.85
5
8
30
Caudal cilia, number
1
1
0
0
0
1
1
30
Measurements in μm. x, arithmetic mean. M, median. SD, standard deviation. SE, standard error of arithmetic mean. CV, coefficient of variation (%). Min, minimum. Max, maximum. n, number of individuals investigated.
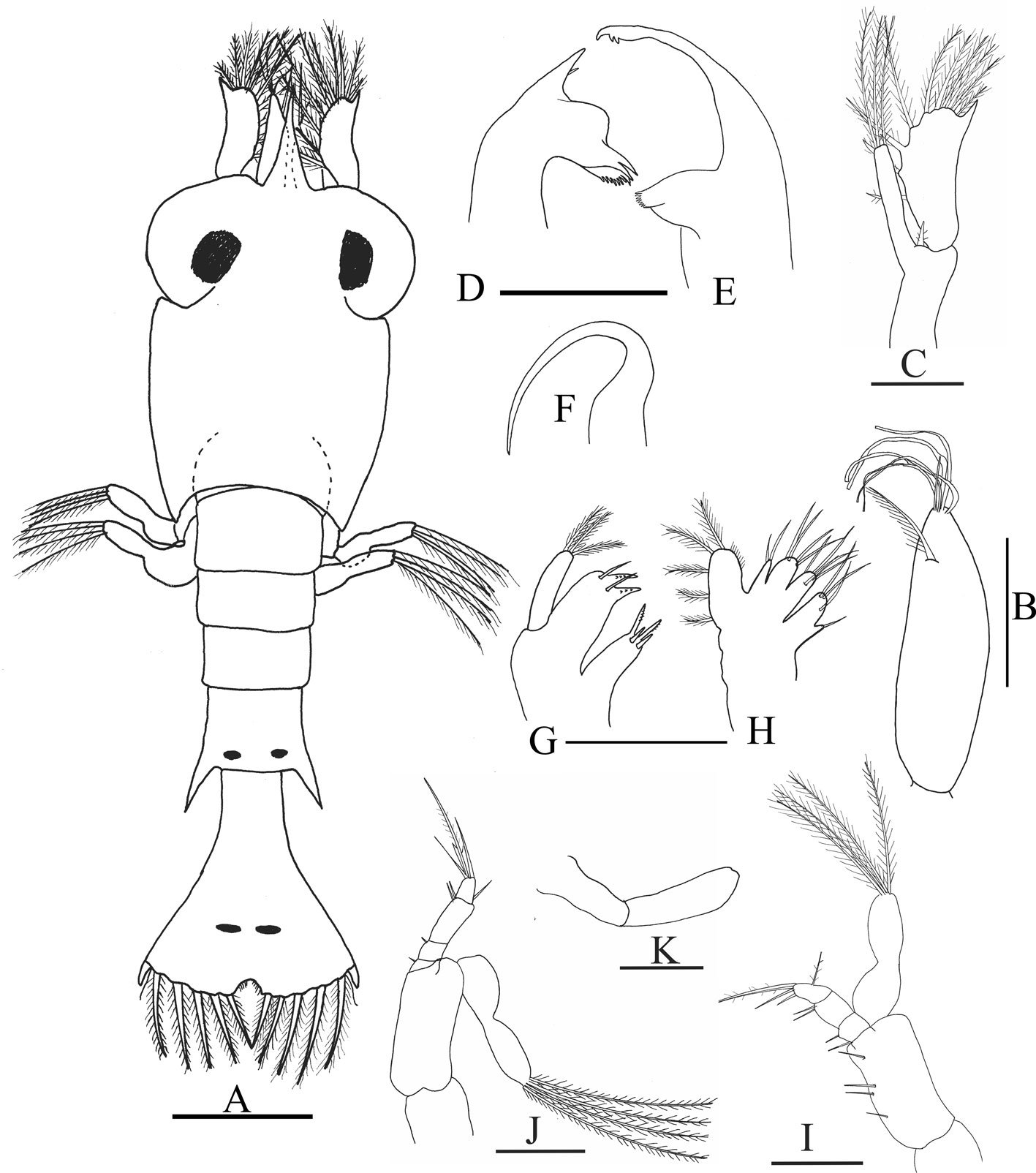
Figure 1. Morfological characterization of Lambornella trichoglossa. A, B, E. In vivo. C, D, F. Dry silver impregnation. G, H. Silver Carbonate impregnation. A, First recorded conjugating specimens; B, lateral view; C, oral apparatus; D, specimen with normal number of caudal cilium (dorsal view); E, nuclear apparatus; F, ventral view, showing oral apparatus and postoral kineties, below; G, specimen recorded during divisional morphogenesis, with multiple food vacuoles scattered throughout the body; H, detail of caudal cilia complex recorded in larger specimens. OA (oral apparatus), FV (food vacuoles), S (preoral suture), PM (paroral membrane), M1 and M2 (adoral membranelles), MA (macronucleus), MI (micronucleus), CC (caudal cilia). Asterisks indicate macronucleus during divisional morphogenesis process. Scale bars in A, B, D, F, G = 100 μm. In C, E H = 50μm. Figure 2. Body length reduction of L. trichoglossa over a 7-day period. Figure 3. Maximum likelihood (ML) tree inferred from small subunit ribosomal DNA (SSU rDNA). Bold: sequence obtained in the present work. Numbers at nodes represent the bootstrap values of ML out of 1,200 replicates.
Table 2
Statistical data on the difference in body length of L. trichoglossa 7 days after collection.
Number of measured specimens
1st day
20
7th day
20
Body length mean (μm)
1st day
205.66
7th day
148.255
Maximum length (μm)
1st day
222.79
7th day
164.84
Minimum length (μm)
1st day
190.18
7th day
132.43
Standart deviation
1st day
10.97027077
7th day
10.06698748
Variance
1st day
120.3468408
7th day
101.3442368
Shapiro-Wilk test (α = 0.05)
1st day
W: 0.9273
p-value: 0.1371
7th day
W: 0.9556
p-value: 0.4607
Student’s t-test
t: 17.241
p: 1.47.10-19
Critical t-value (p = 0.05): 2.0244
Discussion
The similarity of the morphological parameters between the specimens analyzed in the present study and the data presented in the description of L. trichoglossa ensures the correct taxonomic identification of the organisms. It should be noted that the characterization performed by Foissner (2003) occurred a few days after collection, whereas in the present work, the morphological study techniques were performed a few hours after collection. The temporal analyses on body length performed in the present work demonstrated a decrease in the average size of the organisms over time (Fig. 2), which would justify the slightly higher average values compared to the type population.
Regarding the occurrence of specimens with a greater number of caudal cilia, polymorphic life cycles are a common characteristic of the genus Tetrahymena. Lynn and Doerder (2012) mentioned the occurrence of changes in size, body shape, organization, morphology, and cilia of the oral apparatus of Tetrahymena species, due to environmental changes. The aforementioned authors highlighted the great genotypic and phenotypic plasticity found in the clade.
Phylogenetic reconstructions performed with both markers demonstrate the internal position of L. trichoglossa in the genus Tetrahymena, corroborating analyses based on the SSU marker presented by Strüder-Kypke et al. (2001), Bourland and Strüder-Kypke (2010), and Dunthorn et al. (2012). Several authors demonstrated the division of the genus Tetrahymena into 2 strongly molecularly supported clades, Australis and Borealis (Chantangsi, 2007; Chantangsi & Lynn, 2008; Lynn et al., 2018; Nanney, 1998; Strüder-Kypke et al., 2001). This division was observed in the inferred reconstructions by the SSU marker, performed in the present study, demonstrating the inclusion of L. trichoglossa into the Borealis clade. Doerder (2018) states that this clade has greater molecular diversity and more species than the Australis clade, thus being an important source of diversity for the genus. Lambornella trichoglossa, included in this clade, is the greatest example of phenotypic diversification, given its morphological and ontogenetic particularities demonstrated by Foissner (2003).
The phylogenetic analyses inferred by the COI gene demonstrated the inclusion of L. trichoglossa in a clade called coxset A4 by Chantangsi and Lynn (2008). The species T. corlissi and T. bergeri, members of this clade, share the parasitic lifestyle and have particular morphological characteristics (Hoffman et al., 1975; Imai et al., 2000; Strüder-Kypke et al., 2001). Foissner (2003) performed infection tests with L. trichoglossa and different mosquito species, noting that the ciliate does not develop a parasitic association. However, congeners L. stegomyiae and L. clarki are parasites of culicid dipterans, demonstrating that this way of life is predominant in coxiset A4, corroborating Chantangsi and Lynn (2008). Probably, parasitism also represents the ancestral condition of this clade, since Strüder-Kypke et al. (2001) demonstrated that histophagy is a recurrent phenomenon in the evolution of Tetrahymena.
Figure 4. Maximum likelihood (ML) tree inferred from cytochrome oxidase subunit I (COI). Bold: sequence obtained in the present work. Numbers at nodes represent the bootstrap values of ML out of 500 replicates.
Table 3
Pairwise distance between SSU sequences of Lambornella trichoglossa and Tetrahymena spp.
MN567691
L. trichoglossa
AJ810078
L. trichoglossa*
0.0029
JQ723973
Lambornella sp.**
0.0053
0.0047
U17356
T. corlissi
0.0059
0.0063
0.0077
AF364039
T. bergeri
0.0071
0.0092
0.0089
0.0046
EF070247
T. furgasoni
0.0130
0.0149
0.0149
0.0099
0.0092
AF364040
T. mobilis
0.0130
0.0151
0.0149
0.0104
0.0092
0.0006
KJ028502
T. colerunensis
0.0132
0.0139
0.0139
0.0087
0.0080
0.0051
0.0058
KJ028504
T. scolopax
0.0135
0.0156
0.0156
0.0094
0.0067
0.0135
0.0142
0.0125
Bold font: sequence obtained in the present study. * Sequence obtained from the type population (Dunthorn et al., 2012). ** Sequences obtained from samples collected in Jamaica (Dunthorn et al., 2012).
Table 4
Pairwise distance between COI sequences of Lambornella trichoglossa and Tetrahymena spp.
MN477017
L. trichoglossa
EF070279
T. corlissi
0.1294
EF070270
T. bergeri
0.1450
0.0921
EF070291
T. malaccensis
0.1536
0.1187
0.1320
EF070268
T. asiatica
0.1557
0.1026
0.1113
0.1062
KY218158
T. alphathermophila
0.1713
0.1152
0.1403
0.1175
0.1281
EF070296
T. paravorax
0.1961
0.1400
0.1491
0.1413
0.1185
0.1407
KJ028669
T. scolopax
0.1982
0.1364
0.1376
0.1461
0.1184
0.1534
0.1491
EF070278
T. caudata
0.1857
0.1429
0.1492
0.1622
0.1331
0.1553
0.1735
0.1615
MF693881
T. glochidiophila
0.2187
0.1639
0.1867
0.1626
0.1631
0.1703
0.1500
0.1926
0.2051
Bold font: sequence obtained in the present study.
Traditionally recognized Tetrahymena species, described based on morphological or biochemical characteristics, and molecularly supported, may have high evolutionary difference values when comparing COI sequences. For example, T. caudata differs by over 15% from T. corlissi, T. termophila, and T. glochidiophila. Some species described by Doerder (2018) based mainly on molecular diversity differ by values close to 20%. Therefore, the 12.94% distance observed between the COI sequence of L. trichoglossa and T. corlissi does not represent a discrepancy concerning the molecular variability observed in Tetrahymena.
According to Chantangsi and Lynn (2008), the mitochondrial COI gene is effective in elucidating Tetrahymena’s recent phylogeny, while the SSU marker is more suited to assessing the deep phylogeny of the genus. The maintenance of topology in the reconstructions performed with both markers demonstrates that L. trichoglossa, T. corlissi, and T. bergeri share a long evolutionary history, and the differentiation of the species belonging to the genus Lambornella could be recent within the phylogeny of Tetrahymena. The phylogenetic position of L. trichoglossa also demonstrated by the COI gene, reinforces the proposals for synonymization of the genera Lambornella and Tetrahymena. Although Lambornella’s description precedes that of Tetrahymena, the principle of stability is appropriate to maintain the name of the latter taxon. In any case, it is necessary to establish an extended diagnosis for Tetrahymena, so that the species involved in new combinations are covered.
Finally, the molecular analysis of L. trichoglossa based on the COI marker supports the proposal for synonymization of the genera Lambornella and Tetrahymena. This systematic readjustment also points to the great molecular, morphological, evolutionary and ecological complexity of the genus Tetrahymena, which represents one of the greatest examples of evolutionary adaptation among eukaryotic organisms.
References
Bourland, W. A., & Strüder-Kypke, M. (2010). Agolohymena aspidocauda nov. gen., nov.spec., a histophagous freshwater tetrahymenid ciliate in the family Deltopylidae (Ciliophora, Hymenostomatia), from Idaho (northwest U.S.A.): morphology, ontogenesis and molecular phylogeny. European Journal of Protistology, 46, 221–242. https://doi.org/10.1016/j.ejop.2010.04.003
Buosi, P. R. B., Utz, L. R. P., Meira, B. R., Silva, B. T. S., Lansac-Tôha, F. M., Lansac-Tôha, F. A. et al. (2014). Rainfall influence on species composition of the ciliate community inhabiting bromeliad phytotelmata. Zoological Studies, 53, 32–44. https://doi.org/10.1186/s40555-014-0032-4
Buosi, P. R. B., Cabral, A. F., Utz, L. R. P., Vieira, L. C. G., & Velho, L. F. M. (2015). Effects of seasonality and dispersal on the ciliate community inhabiting bromeliad phytotelmata in riparian vegetation of a large tropical river. Journal of Eukaryotic Microbiology, 62, 737–749. https://doi.org/10.1111/jeu.12232
Chantangsi, C., & Lynn, D. H. (2008). Phylogenetic relationships within the genus Tetrahymena inferred from the cytochrome c oxidase subunit 1 and the small subunit ribosomal RNA genes. Molecular Phylogenetics and Evolution, 49, 979–87. https://doi.org/10.1016/j.ympev.2008.09.017
Chantangsi, C., Lynn, D. H., Brandl, M. T., Cole, J. C., Hetrick, N., & Ikonomi, P. (2007). Barcoding ciliates: a comprehensive study of 75 isolates of the genus Tetrahymena. International Journal of Systematic and Evolutionary Microbiology, 57, 2412–2423. https://doi.org/10.1099/ijs.0.64865-0
Corliss, J. O., & Coats, D. W. (1976). A new cuticular cyst-producing tetrahymenid ciliate, Lambornella clarki n. sp., and the current status of ciliatosis in culicine mosquitoes. Transactions of the American Microscopical Society, 95, 725–739. https://doi.org/10.2307/3225400
Corliss, J. O. (1960). Tetrahymena chironomi sp. nov., a ciliate from midge larvae, and the current status of facultative parasitism in the genus Tetrahymena. Parasitology, 50, 111–153. https://doi.org/10.1017/S0031182000025245
Doerder, F. P. (2018). Barcodes reveal 48 new species of Tetrahymena, Dexiostoma, and Glaucoma: phylogeny, ecology, and biogeography of new and established species. Journal of Eukaryotic Microbiology, 66, 182–208. https://doi.org/10.1111/jeu.12642
Dunthorn, M., Stoeck, T., Wolf, K., Breiner, H., & Foissner, W. (2012). Diversity and endemism of ciliates inhabiting Neotropical phytotelmata. Systematics and Biodiversity, 10, 195–205. https://doi.org/10.1080/14772000.2012.685195
Durán-Ramírez, C., García-Franco, J. G., Foissner, W., & Mayén-Estrada, R. (2015). Free-living ciliates from epiphytic tank bromeliads in Mexico. European Journal of Protistology, 51, 15–33. https://doi.org/10.1016/j.ejop.2014.09.002
Foissner, W. (2003). Morphology and ontogenesis of Lambornella trichoglossa nov. spec., a new tetrahymenid ciliate (Protozoa,Ciliophora) from Brazilian tank bromeliads (Bromeliaceae). European Journal of Protistology, 39, 63–82. https://doi.org/10.1078/0932-4739-00887
Foissner, W. (2014). An update of basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. International Journal of Systematic and Evolutionary Microbiology, 64, 271–292. https://doi.org/10.1099/ijs.0.057893-0
Foissner, W., Strüder-Kypke, M., van der Staay, G. W. M., van der Staay, S. M., & Hackstein, J. H. P. (2003). Endemic ciliates (Protozoa,Ciliophora) from tank bromeliads (Bromeliaceae): a combined morphological, molecular, and ecological study. European Journal of Protistology, 39, 365–372. https://doi.org/0.1078/0932-4739-00005
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4, 1–9.
Hoffman, G. L., Lando, M., Camper, J. E., Coats, D. W., Stookey, J. L., & Burek, J. D. (1975). A disease of freshwater fishes caused by Tetrahymena corlissi Thompson, 1955, and a key for identification of holotrich ciliates of freshwater fishes. The Journal of Parasitolgy, 61, 217–223. https://doi.org/10.1111/j.1365-2761.2009.01062.x
Imai, S., Tsurimaki, S., Goto, E., Wakita, K., & Hatai, K. (2000). Tetrahymena infection in Guppy, Poecilia reticulata. Fish Pathology, 35, 67–72. https://doi.org/10.3147/jsfp.35.67
Katoh, K., Rozewicki, J., & Yamada, K. D. (2017). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Keilin, D. (1921). On a new ciliate: Lambornella stegomyiae n. g., n. sp., parasitic in the body-cavity of the larvae of Stegomyia scutellaris Walker (Diptera, Nematocera, Culicidae). Parasitology, 13, 216–224. https://doi.org/10.1017/s0031182000012488
Kimura, M. (1980). A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 16, 111–120. https://doi.org/10.1007/BF01731581
Kumar, S., Stecher, G., Li, M., Knyaz, C., & Tamura, K. (2018). MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Molecular Biology and Evolution, 35, 1547–1549. https://doi.org/10.1093/molbev/msy096
Lynn, D. H., & Doerder, F. P. (2012). The life and times of Tetrahymena. In Collins, K. Methods in cell biology. Elsevier, 109, 9–27. https://doi.org/10.1016/B978-0-12-385 967-9.00002-5
Lynn, D. H., Doerder, F. P., Gillis, P. L., & Prosser, R. S. (2018). Tetrahymena glochidiophila n. sp., a new species of Tetrahymena (Ciliophora) that causes mortality to glochidia larvae of freshwater mussels (Bivalvia). Diseases of Aquatic Organisms, 127, 125–136. https://doi.org/10.3354/dao03188
Lynn, D. H. (2008). The ciliated taxa including families and genera. In D. H. Lynn (Ed.), The ciliated protozoa (pp. 339–440). Guelph, Canada: Springer.
Messing, J. (1983). New M13 vectors for cloning. Methods in Enzymology, 101, 20–78. https://doi.org/10.1016/0076-6879(83)01005-8
Modeo, L., Rosati, L., Andreoli, I., Simoni, G., Verni, F., & Petroni, G. (2006). Molecular systematics and ultrastructural characterization of a forgotten species: Chattonidium setense (Ciliophora, Heterotrichea). Proceedings of the Japan Academy, 82, 359–374. https://doi.org/10.2183/pjab.82.359
Nanney, D. L., Park, C., Preparata, R., & Simon, E. M. (1998). Comparison of sequence differences in a variable 23S rRNA domain among sets of cryptic species of ciliated protozoa. Journal of Eukaryotic Microbiology, 45, 91–100. https://doi.org/10.1111/j.1550-7408.1998.tb05075.x
Schrallhammer, M., Ferrantini, F., Vannini, C., Galati, S., Schweikert, M., Görtz, H. et al. (2013). Candidatus Megaira polyxenophila gen. nov., sp. nov.: Considerations on evolutionary history, host range and shift of early divergent Rickettsiae. Plos One, 8, e72581. https://doi.org/10.1371/journal.pone.0072581
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics, 30, 1312–3. https://doi.org/10.1093/bioinformatics/btu033
Strüder-Kypke, M., Wright, H. D., Jerome, C. A, & Lynn, D. H. (2001). Parallel evolution of histophagy in ciliates of the genus Tetrahymena. Evolutionary Biology, 1, 1–5. https://doi.org/10.1186/1471-2148-1-5
Strüder-Kypke, M., & Lynn, D. H. (2010). Comparative analysis of the mitochondrial cytochrome c oxidase subunit I (COI) gene in ciliates (Alveolata, Ciliophora) and evaluation of its suitability as a biodiversity marker. Systematics and Biodiversity, 8, 131–148. https://doi.org/10.1080/14772000903507744
Tamura, K., Nei, M., & Kumar, S. (2004). Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences (USA), 101, 11030–11035. https://doi.org/10.1073/pnas.0404206101
Talavera, G., & Castresana, J. (2007). Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Systematic Biology, 56, 564–577. https://doi.org/10.1080/10635150701472164
The genus Psephenops (Coleoptera: Dryopoidea: Psephenidae) in Mexico, with the description of a new species and notes on the species from Central America and the Antilles
Roberto Arce-Pérez, J. Antonio Gómez-Anaya y Rodolfo Novelo-Gutiérrez *
Instituto de Ecología, A. C., Red de Biodiversidad y Sistemática, Carretera Antigua a Coatepec 351, Col. El Haya, 91073 Xalapa, Veracruz, México
*Autor para correspondencia: rodolfo.novelo@inecol.mx (R. Novelo-Gutiérrez)
Recibido: 05 junio 2025; aceptado: 26 septiembre 2025
El género neotropical Psephenops Grouvelle incluye actualmente 15 especies descritas, de las cuales 13 se distribuyen en México, América Central y las Antillas, más una de Argentina y otra de Perú. Se reporta por primera vez a la subfamilia Psepheninae con Psephenops oaxacanus sp. nov. para el estado de Oaxaca, México, elevando a 3 el número de especies conocidas para el país, 14 en México, América Central y las Antillas y 16 a nivel continental. Se proporciona un mapa de distribución y una clave ilustrada para las especies presentes en México.
Palabras clave: Psepheninae; Moneditas de agua; Oaxaca; México; Taxonomía
Abstract
The Neotropical genus Psephenops Grouvelle currently includes 15 described species, 13 of which are distributed in Mexico, Central America, and the Antilles, plus one from Argentina and another from Peru. The subfamily Psepheninae is reported for the first time with Psephenops oaxacanus sp. nov. from the state of Oaxaca, Mexico, increasing the number of known species in the country to 3, 14 in Mexico, Central America and the Antilles, and 16 at the continental level. A distribution map and an illustrated key for the species present in Mexico are provided.
Keywords: Psepheninae; Water pennies; Oaxaca; Mexico; Taxonomy
Introducción
El género neotropical Psephenops Grouvelle, 1898 (= Xexanchorinus) está representado por 15 especies descritas hasta la fecha: P. argentinensis Deléve, 1967 (Argentina); P. bifidus Arce-Pérez y Novelo-Gutiérrez, 2017 (Panamá); P. grouvellei Champion, 1913 (Guatemala); P. haitianus Darlington, 1936 (Haití); P. lupita Arce-Pérez, 2002 (México); P. maculicollis Darlington, 1936 (Costa Rica y Colombia); P. mexicanus Arce-Pérez y Novelo-Gutiérrez, 2000 (México); P. panamaensis Arce-Pérez y Novelo-Gutiérrez, 2015 (Panamá); P. prestonae Spangler 1990 (Costa Rica); P. robacki (Spangler), 1966 (Perú); P. shepardi Arce-Pérez y Novelo-Gutiérrez, 2013 (Belice); P. smithi Grouvelle, 1898 (Antillas menores); P. spiniparameri Arce-Pérez y Novelo-Gutiérrez, 2013 (Belice); P. triangularis Arce-Pérez y Novelo-Gutiérrez, 2017 (Panamá); y P. trini Barr y Shepard, 2024 (Antillas menores) (Arce-Pérez 2002; Arce-Pérez y Novelo-Gutiérrez, 2000, 2013, 2015, 2017; Bameul, 2001; Barr y Shepard, 2024; Deléve, 1967). En este trabajo se reporta por primera vez al género Psephenops para Oaxaca, México, con base en ejemplares recolectados en el municipio de Santiago Comaltepec y se describe e ilustra a Psephenops oaxacanus sp. nov.
Materiales y métodos
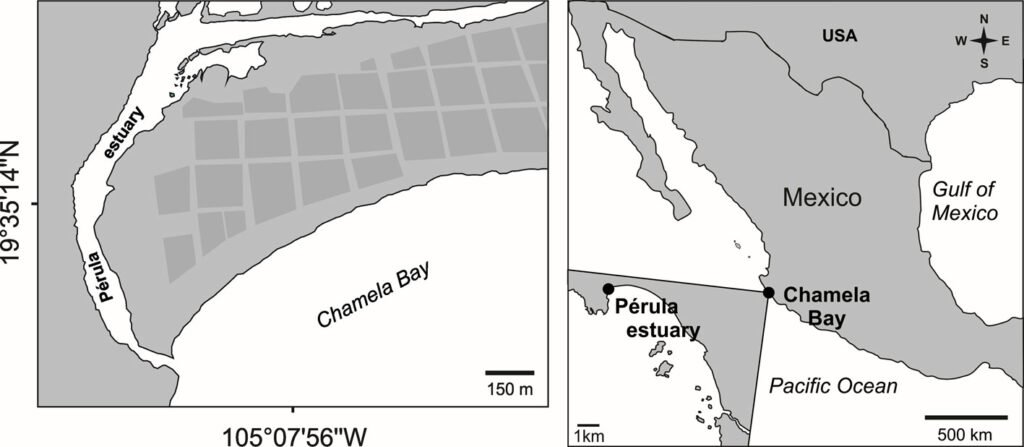
Los ejemplares de la especie nueva se recolectaron mediante una red acuática con marco tipo D y se colocaron en frascos con etanol al 96%, en un tributario del río Soyolapam que corre por la selva alta perennifolia (fig. 3), en el poblado San Martín Soyolapam, municipio Santiago Comaltepec, en Oaxaca, México en febrero del 2016 (fig. 4). Los especímenes fueron identificados utilizando las descripciones y claves originales de Arce-Pérez y Novelo-Gutiérrez (2000, 2013, 2015, 2017). El examen morfológico se realizó utilizando un microscopio estereoscópico Nikon SMZ 745. Las imágenes y mediciones de estructuras específicas se obtuvieron mediante el uso de un microscopio estereoscópico Nikon SMZ25 acoplado al software Nikon NIS-Elements Imaging seguido de la preparación de imágenes con Photoshop CS5 (Adobe System Inc. San José, California). Las micrografías electrónicas se obtuvieron utilizando un microscopio electrónico de barrido Jeol JSM-5600LV. Las mediciones están en mm. El mapa de distribución geográfica se realizó con Simplemappr (Shorthouse, 2010).
Descripción
Con base en 6 machos y 6 hembras se describe Psephenops oaxacanus sp. nov., representando el primer registro de la subfamilia para el estado de Oaxaca, elevando a 3 el número de especies a nivel nacional, 14 en México, América Central y las Antillas y 16 a nivel continental. Se proporciona una lista anotada, claves dicotómicas y mapa de distribución para las especies presentes en México.
Familia Psephenidae Lacordaire, 1854
Subfamilia Psepheninae Lacordaire, 1854
Genero Psephenops Grouvelle, 1898
Psephenops oaxacanus Arce-Pérez, Gómez-Anaya y Novelo-Gutiérrez sp. nov. (figs. 1-6)
Diagnosis. Habitus (fig. 1): longitud total 3.2 mm; ancho humeral 1.4 mm. Cuerpo ovalado, deprimido; cabeza, pronoto y escutelo negro rojizo, élitros pardo rojizos, ventralmente pardo oscuros, al igual que patas, antenas y palpos. Dorsalmente cubierto completamente por vestidura de sedas diminutas de color amarillo con reflejos dorados, además cabeza y pronoto con sedas más largas y gruesas de color rojizo oscuro; pronoto con cresta levantada en el tercio distal, antenas moniliformes cortas que no rebasan región distal del pronoto. Ventralmente la vestidura presenta sedas diminutas blanquecinas amarillentas con reflejos dorados. Tarsos con los lóbulos de los tarsómeros 1 y 2 cortos, con vestidura ventral amarilla (fig. 1a, b). Se distingue de las otras especies mexicanas del género por la combinación de los caracteres de la tabla 1.
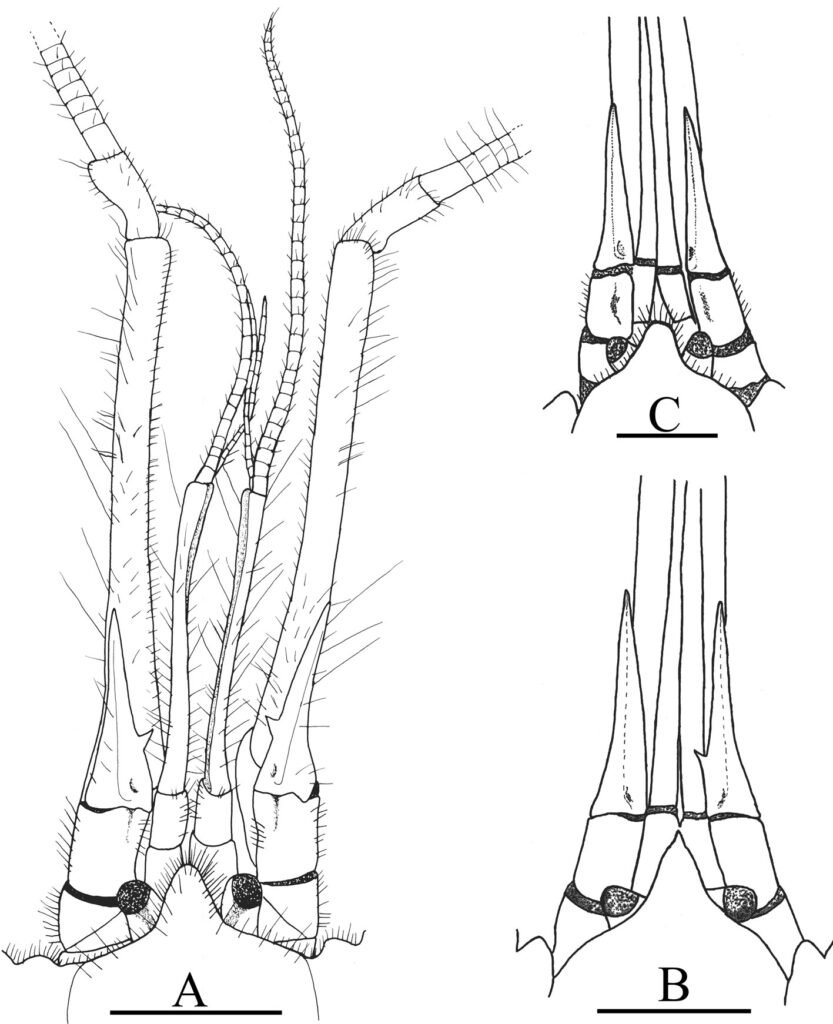
Holotipo macho. Dorsalmente (fig. 1a): cabeza corta, más ancha que larga; clípeo subrectangular, en declive menor a 90 grados desde plano frontal, margen distal ampliamente emarginado; vértex con surco longitudinal medio; superficie fronto-clipeal y vértex micropunteado, con sedas diminutas de color amarillo y sedas más largas y gruesas de color rojo oscuro. Antenas cortas, moniliformes, de 11 antenómeros, sin alcanzar margen posterior del pronoto; escapo más largo y robusto; pedicelo subgloboso, mitad de largo que escapo, ambos ligeramente más claros que antenómeros del flagelo; flagelo de 8 antenómeros, subtriangulares anchos, antenómero apical acuminado. Ojos laterales, redondeados, muy prominentes, de color rojo oscuro, con zona postocular amarilla y sedas largas amarillentas. Labro subrectangular similar al clípeo, pero más corto, ampliamente emarginado. Palpos maxilares de 4 artejos, cubiertos con sedas cortas, palpómero basal más pequeño y redondo, palpómero apical más grande, acuminado y con área apical sensorial amplia, longitud de los palpómeros: 0.04, 0.08, 0.06 y 0.14 mm. Palpos labiales muy cortos, de 3 artejos, palpómeros 1-2 redondeados y anchos, 3 subcónico, con ápice redondeado y ligeramente más pequeño que anteriores. Ventralmente (fig. 1b): negra rojiza, partes bucales pardo oscuro. Tórax: pronoto subtrapezoidal, casi tan ancho como base de élitros, 0.75 mm de largo por 1.2 mm de ancho basal; margen anterior arqueado, margen posterior bisinuado; márgenes laterales casi rectos a lo largo de mitad anterior y expandido lateralmente en mitad posterior. Disco pronotal con una cresta longitudinal bien desarrollada en tercio distal y ligera depresión ovalada a cada lado de ésta, además de otra depresión en margen laterodistal (fig. 1a); superficie pronotal diminutamente punteada, cubierta por vestidura de sedas diminutas amarillas con reflejos dorados y sedas largas de color rojo oscuro. Escutelo corto, más ancho en la base que largo, ápice redondeado. Élitros largos, exponiendo solo último esternito abdominal (pigidio), con márgenes laterales paralelos en los 0.75 basales, luego convergiendo gradualmente hacia atrás, con ancho humeral de 1.4 mm y cubiertos por una vestidura de sedas diminutas amarillas con reflejos dorados. Prosternón estrecho, con proceso postcoxal largo y lanceolado, 0.50 mm de largo por 0.13 mm de ancho en región lanceolada, levantado en toda su longitud y carenado en mitad distal, alcanzando mitad anterior del mesoventrito. Mesoventrito corto, con surco amplio para recepción del proceso postcoxal. Metaventrito grande y voluminoso, con un discrimen evidente que se observa como surco. Patas con coxas anteriores y medias cortas globosas, posteriores planas y transversas; fémures robustos; tibias largas y delgadas, ligeramente más anchas en ápice; protibia con pequeño dentículo apicolateral; metatibias más largas que metafémures; tarsómeros 1 y 2 ventralmente lobulados, con vestidura gruesa parecida a esponja; lóbulo ventral del tarsómero 1 se extiende distalmente sobre 0.30 basales del tarsómero 2; tarsómero 2 el más largo, casi 2 veces más largo que el 1, extendiéndose distalmente y cubriendo completamente a tarsómeros 3, 4 y poco más de 0.50 basales del 5; longitud de tarsómeros: 0.53, 0.94, 0.19, 0.17 y 0.54 mm (sumergidos en alcohol); garras apicales largas, curvas y delgadas, cada una con pequeño diente basal. Abdomenconvestidura corta, fina y densa, blanquecino amarillenta, con reflejos dorados; ventrito VI poco visible, oculto debajo de ventrito V; ventritos I y II anchos, con márgenes anterior y posterior sinuados; ventritos III y IV estrechos, con margen posterior ligeramente arqueado; ventrito V con margen posterior ampliamente arqueado; ventrito VI poco visible, corto, subtriangular, con margen posterior ampliamente arqueado; ventrito VII o placa anal, subtriangular, cubierta de sedas largas y amarillentas; último tergito abdominal o pigidio subtriangular, cubierto de sedas largas rojizo oscuras. Genitales en vista ventral subrectangulares, anchos, longitud de 0.55 mm de falobase al ápice del lóbulo medio, ancho de 0.25 mm entre parámeros y la falobase; lóbulo medio digitiforme con ápice en punta, longitud 0.29 mm, con esclerito medio esbelto, de 0.25 mm de largo, triangular en base, esbelto y claviforme hacia ápice (fig. 2a). En vista dorsal parámeros grandes y robustos de 0.37 mm, con ¼ apical subtriangular y delgada membrana lateral; falobase reducida, 0.18 mm y cóncava (fig. 2b). En vista ventrolateral falobase convexa, parámeros curvados y esclerito del lóbulo medio largo y agudo (fig. 2d). En vista frontal membrana de lóbulo medio fuertemente esclerotizada y escamosa formando cubierta dura (fig. 2c).
Variación en machos. Escasa, excepto por la longitud corporal (n = 5) de 3 a 3.5 mm por 1.2 a 1.5 mm de ancho humeral. La cresta pronotal se puede prolongar como ligera banda longitudinal semidesnuda hacia la región anterior; los élitros pueden estar ligeramente oscurecidos en la región posterior.
Hembra. Más grande y robusta que el macho, longitud 3.6 mm por 1.8 de anchura humeral (fig. 1c, d). Cabeza con ojos muy protuberantes y amplia zona postocular amarilla, cubierta por sedas largas rojizo oscuro; pronoto subtrapezoidal ancho y expandido posteriormente, con 0.90 mm de largo, por 1.7 mm de ancho distal (fig. 1c); proceso postcoxal muy ancho y lanceolado, 0.60 mm de largo por 0.22 mm de ancho en región lanceolada; mesoventrito ampliamente acanalado para recepción del proceso postcoxal; ventritos II y III con corta hendidura transversal a cada lado de línea media (fig. 1d); las patas son más claras, coxas, trocánteres y fémures mayormente rojizo amarillentos, ápice de fémures oscuros, tibias rojizas en margen interno y oscuras en el externo; tarsos totalmente amarillos con uñas y ápice del tarsómero apical oscuros; tarsómeros subcónicos, sin lóbulos ventrales, primeros 4 cortos y subcónicos, 5 esbelto, tan largo como primeros 4 combinados. Genitales con ovipositor ligeramente esclerotizado, hialino, longitud 0.40 mm; valvífera con 2 laterotergitos ligeramente esclerotizados, longitud 0.16 mm; gonocoxitos longitud 0.21 mm, juntos forman placa ancha y plana sin sedas ni puntos aparentes; estilos oblongos, longitud 0.03 mm (fig. 2e).
Variación en hembras. No se observa mucha variación en coloración y vestidura, únicamente algunas con élitros un poco más oscuros; también varían en la longitud corporal (n = 5) de 3.2 a 3.9 mm por 1.5 a 1.9 mm de ancho humeral.
Tabla 1
Caracteres diferenciales de las especies de Psephenops Grouvelle en México, América Central y las Antillas.
Psephenops spp/estructuras
smithi guadeloupensi
grouvellei
haitianus
maculicollis
prestone
mexicanus
lupita
spiniparameri
shepardi
panamaensis
bifidus
triangularis
oaxacanus
Longitud total (mm)
2.51
3.4
2.5
2.5
2.46
3.3
3.8
2.75
2.65
2.4
2.95
2.5
3.2
Tubérculos pronotales
0
3
0
0
0
0
0
0
0
0
0
0
0
Superficie elitral
con estrías
con bordes longitudi- nales
estriado
estriado
liso
liso
liso
liso
liso
liso
liso
liso
liso
Tipo de antenas
monili- formes
monili- formes?
filifor- mes?
monili- formes?
?
monili- formes
filifor- mes
monili- formes
monili- formes
monili- formes
monili- formes
monili- formes
monili- formes
Longitud antenal*
largas
largas
largas
cortas
cortas
cortas
largas
cortas
largas
largas
cortas
cortas
cortas
Proceso prosternal
subcilin- drico no bifido?
?
?
?
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado y bífido
lanceo- lado no bífido
lanceo- lado no bífido
Protibias
?
?
?
?
sin diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
Patas con lóbulos en los tarsómeros
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 en frontales y medias, 1 posteriores
1 y 2 de todas las patas
1 a 3 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
Longitud de lóbulos tarsales**
largos
cortos?
?
largos?
largos
largos
cortos
largos
cortos
largos
largos
largos
cortos
Ápice de parameros
truncados
?
?
no truncado,
no truncado
no truncado
truncado
no truncado, divergente
no truncado,
truncado
no truncado
no truncado
no truncado
Parameros con dientecillo
ausente
ausente
ausente
ausente
ausente
ausente
ausente
ausente
presente
ausente
presente
presente
ausente
mitad basal de los parameros
fusionados
?
?
fusionados
fusionados
con una profunda hendidura
fusionados
con una profunda hendidura
fusionados
fusionados
con una profunda hendidura
fusionados
fusionados con ligera hendidura
Pene con proyección lateral
ausentes
?
?
ausentes
ausentes
pequeños dentículos
ausentes
ausentes
ausentes
ausentes
ausentes
proyección subtrian- gular
ausentes
Forma del esclerito del pene
?
?
?
?
?
lanceolado
digiti- forme
subtrian- gular esbelto
subtrian- gular, ápice bífido
subtrian- gular ancho
recto, ancho en la base
como un reloj de arena
esbelto lanceolado
Distribución
Antillas
Guatemala
Haití
Costa Rica, Colombia
Costa rica
México
México
Belice
Belice
Panamá
Panamá
Panamá
México
*Cortas = Cuando está completamente extendido hacia atrás, no alcanza el margen posterior del pronoto; Largas = que alcanza o sobrepasa el margen posterior del pronoto. ** Corto = Cuando el lóbulo del tarsómero 2 cubre ventralmente toda la longitud del tarsómero 3-4 y la mitad basal o menos del tarsómero 5; Largo = el menor pero que alcanza al menos 80% de la longitud del 5.
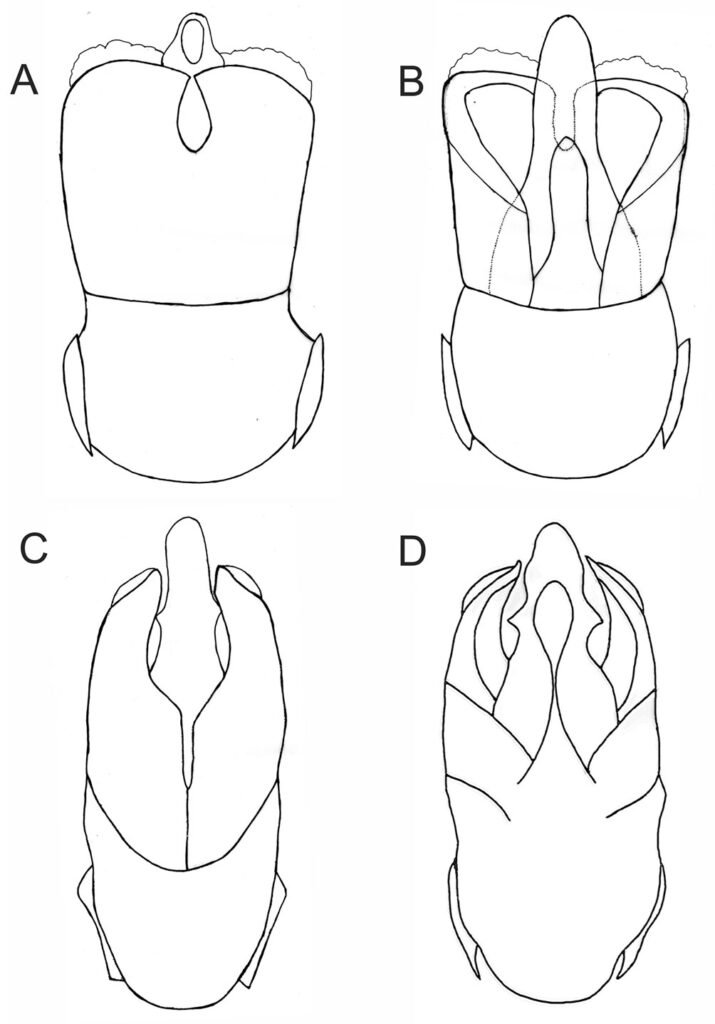
Figura 1. Vista dorsal y ventral del macho y hembra de Psephenops oaxacanus. A) Hábito dorsal; B) hábito ventral; C) hábito dorsal; D) hábito ventral. Figura 2. Vista del genital masculino y femenino de Psephenops oaxacanus. A) Vista dorsal; B) vista ventral; C) vista frontal; D) vista ventrolateral; E) vista ventral del genital femenino mostrando los estilos, gonocoxitos y laterotergitos.
Resumen taxonómico
Material tipo. Holotipo macho. México: localidad tipo, estado de Oaxaca, Municipio Santiago Comaltepec, San Martín Soyolapam, río Soyolapam, 136 m snm (17°41’58” N, -96°16’59” O), 5.II.2016. Cols. J.A. Gómez et R. Novelo [escrito a máquina, etiqueta blanca], “Holotype ♂/ Psephenops oaxacanus nov. sp. / Arce-Pérez, Gómez-Anaya & Novelo-Gutiérrez 2025” [escrito a máquina, etiqueta roja], Paratipos: 5 ♂, 6 ♀ mismos datos del holotipo [escrito a máquina, etiqueta amarilla] (IEXA).
Etimología. Esta especie recibe el nombre oaxacanus por ser la primera especie de la subfamilia Psepheninae y del género Psephenops para el estado de Oaxaca, México.
El holotipo y paratipos se encuentran depositados en la Colección Entomológica del Instituto de Ecología, A.C., Xalapa, México (IEXA), con el número de catálogo IEXA-2025-IM20.
Comentarios taxonómicos
Se considera más estrechamente relacionada con P. mexicanus por presentar ambas el mismo tipo de antenas cortas y moniliformes, parámeros en vista dorsal subtriangulares, separados por un profundo surco y fusionados en su base (figs. 2a, 6c), además por el tamaño corporal de 3 a 3.5 mm. Con base en la literatura disponible, la nueva especie puede distinguirse de las de México, América Central y las Antillas por las características anotadas en la tabla 1.
Hábitat. El río Soyolapam se localiza en la localidad del mismo nombre en la sierra de Juárez, Oaxaca, en la subcuenca del río Valle Nacional (fig. 4), el tipo de vegetación es selva alta perennifolia a una altitud de 136 m snm. El río presenta en su cauce cantos y rocas emergentes cubiertas parcialmente por algas y vegetación abundante en sus márgenes, los cuales en algunas partes son playones arenosos (fig. 3).
Figura 3. Localidad tipo de Psephenops oaxacanus en el río Sayolapam (San Martín), Santiago Comaltepec, Oaxaca, México (febrero de 2016).
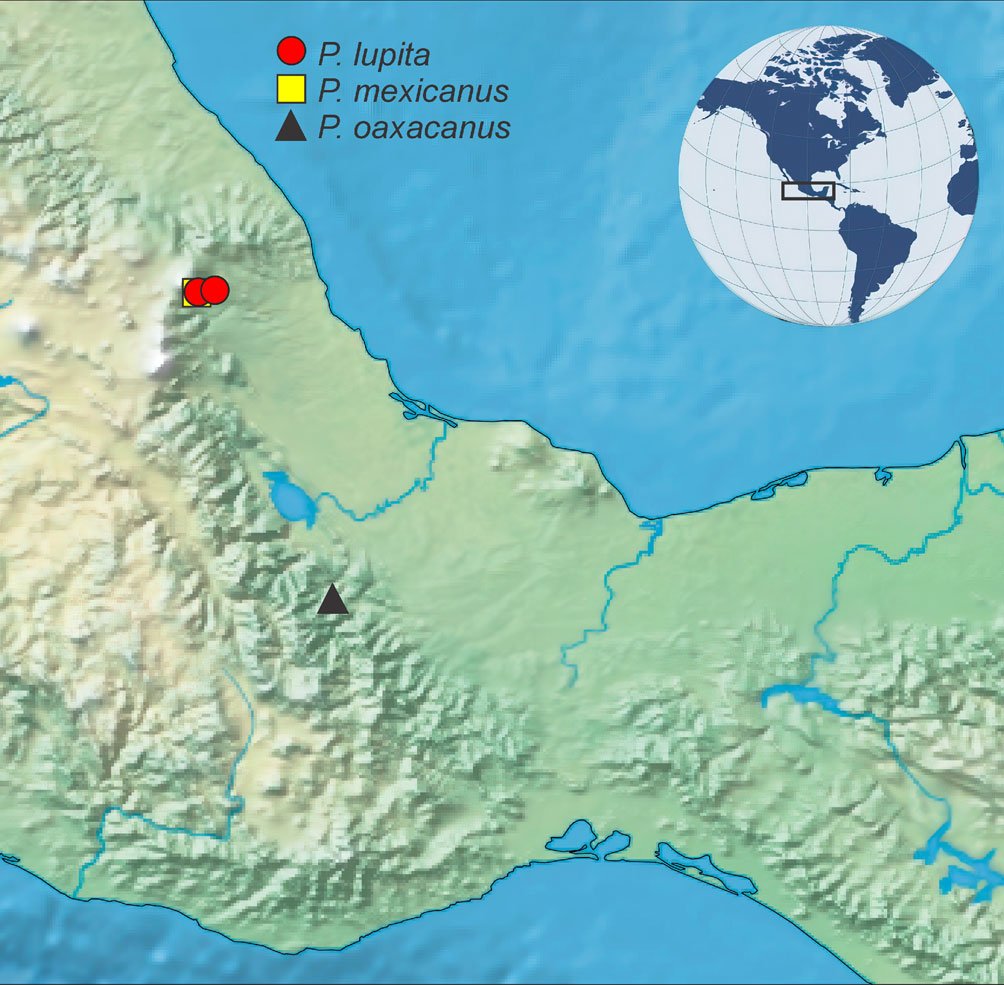
Hasta el momento, esta especie solo está presente en Oaxaca, al sur de México, en el río Soyolapan, a 136 m snm (fig. 4). La localidad tipo se encuentra en la parte norte del estado, en la provincia sierra Madre del Sur, en la subprovincia de los Altos de Oaxaca. Esta especie se recolectó sintópicamente con ejemplares de los géneros Cylloepus y Macrelmis de la familia Elmidae.
Lista actualizada de las especies de Psephenops Grouvelle en México.
Psephenops lupita (Arce-Pérez)
Psephenops lupita (Arce-Pérez, 2002: 964)
Registros publicados. México: Veracruz: Municipio Coatepec, Poblado Coatepec, río Huehueyapan (Sección La Marina), 19º26’55” N, 96º58’52” O, 1, 200 m, bosque mesófilo de montaña (fig. 4); municipio Xico, Poblado Xico Viejo, río Malseseca, 19º26’44.89” N, 97º03’22.22” O, 1,800 m, bosque mesófilo de montaña (fig. 4).
Psephenops mexicanus (Arce-Pérez y Novelo-Gutiérrez)
Psephenops mexicanus (Arce-Pérez y Novelo-Gutiérrez, 2000: 196)
Registros publicados. México: Veracruz: Municipio Coatepec, Poblado Coatepec, río Huehueyapan (Sección La Marina), 19º26’55” N, 96º58’52” O, 1,200 m, bosque mesófilo de montaña (fig. 4); municipio Xico, Poblado Xico Viejo, río Malseseca, 19º26’44.89” N, 97º03’22.22” O, 1,800 m, bosque mesófilo de montaña (fig. 4).
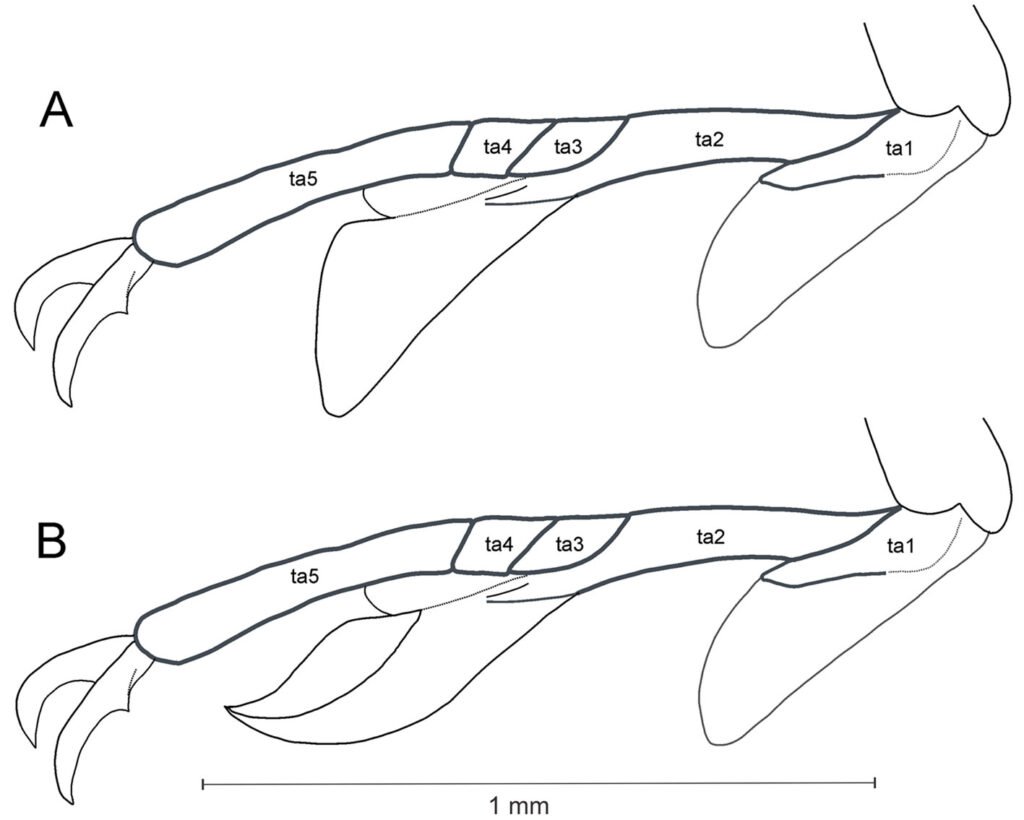
Figura 4. Distribución de las especies de Psephenops en México. Figura 5. Vista lateral de las patas mostrando lóbulos tarsales en Psephenops spp. A) Cortos; B) largos, ta = tarsómeros. Figura. 6 A-D. Vista del genital masculino de P. lupita y P. mexicanus. A) Dorsal; B) ventral; C) dorsal; D) ventral. Figuras A, B tomadas de Arce-Pérez (2002); C, D tomadas de Arce-Pérez y Novelo-Gutierrez (2000).
Clave para la identificación de las especies mexicanas de Psephenops Grouvelle (modificada de Arce-Pérez y Novelo-Gutiérrez, 2013, 2017)
1 Antenas filiformes, largas, tocando base de pronoto; parámeros en vista dorsal anchos, subcuadrados y fusionados en base en ligera hendidura en forma de gota (fig. 6a), en vista ventral lóbulo medio y esclerito interno digitiformes (fig. 6b); lóbulos tarsales cortos, sin alcanzar más de 0.5 basales del tarsómero 5 (fig. 5a); longitud total 3.75 – 3.85 mm P. lupita
1’ Antenas cortas, moniliformes, no tocan base del pronoto; parámeros en vista dorsal subtriangulares, separados por hendidura que va de ligera a profunda (figs. 2a, 6c); en vista ventral lóbulo medio y esclerito interno no digitiformes; lóbulos tarsales cortos o largos (figs. 5a, b) 2
2 (1’) Lóbulos tarsales largos, cubriendo 0.8 basales del tarsómero 5 (fig. 5b); parámeros en vista dorsal con profunda hendidura longitudinal media (fig. 6c); en vista ventral lóbulo medio subtriangular, con dientecillo a cada lado; esclerito interno del lóbulo medio lanceolado (fig. 6d); longitud 3.15 – 3.55 mm P. mexicanus
2’ Lóbulos tarsales cortos, sin alcanzar más de 0.5 basales del tarsómero 5 (fig. 5a); parámeros en vista dorsal separados por profunda hendidura ancha en forma de lanza invertida y continuada por ligera hendidura que no los separa (fig. 2a); en vista ventral lóbulo medio digitiforme sin dientecillos laterales, terminando en punta ancha, con esclerito interno esbelto, claviforme hacia el ápice, en vista lateral agudo (figs. 2b-d); longitud 3.0 – 3.5 mm P. oaxacanus sp. nov.
Agradecimientos
A Emmanuel Arriaga Varela (INECOL) y a un revisor anónimo por sus atinadas correcciones y sugerencias que mejoraron el manuscrito final.
Referencias
Arce-Pérez, R. (2002). A new species of Psephenops Grouvelle (Coleoptera: Psephenidae) from Mexico. Proceedings of the Entomological Society of Washington, 104, 964–967.
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2000). First record of the genus Psephenops (Coleoptera: Psephenidae) from Mexico, with a description of a new species. Entomological News, 111, 196–200.
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2013). Two new species of Psephenops Grouvelle from Belize (Coleoptera: Byrrhoidea: Psephenidae), with a key to the known species from Mexico and Central America. Zootaxa, 3670, 63–70. http://dx.doi.org/10.11646/zootaxa.3670.1.5
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2017). Two new species of Psephenops Grouvelle (Coleoptera: Byrrhoidea: Psephenidae) from Panama. Zootaxa, 4323, 109–118. https://doi.org/10.11646/zootaxa.4323.1.8
Arce-Pérez, R., Novelo-Gutiérrez, R. y Cornejo, A. (2015). Psephenops panamaensis sp. nov. (Coleoptera: Byrrhoidea: Psephenidae) from Panamá. Zootaxa, 4052, 233–236. https://doi.org/10.11646/zootaxa.4052.2.8
Bameul, F. (2001). Un nouveau Psephenops Grouvelle de la Guadeloupe, avec la description de sa larve (Coleoptera, Psephenidae). Nouvelle Revue d’Entomologie, 18, 161–172.
Barr, C. B. y Shepard, W. D. (2024). A taxonomic review of the genus Psephenops Grouvelle of the Lesser Antilles with description of new species Psephenops trini, and reassignment of Peruvian species Psephenus robacki Spangler (Coleoptera: Psephenidae: Psepheninae). Insecta Mundi, 1045, 1–22. https://doi.org/10.5281/zenodo.11450152
Deléve, J. (1967). Contribution á l’ étude des Dryopoidea (Coleoptera). XIX. Notes diverses et descriptions d’ spéces nouvelles. Bulletin et Annales de la Société Entomologique de Belgique, 103, 414–446.
Algae diversity and new records of Xanthophyceae and Eustigmatophyceae (Heterokontophyta) in the central region of Mexico
Alejandra Mireles-Vázquez a, b, * y Eberto Novelo b
a Universidad Nacional Autónoma de México, Posgrado en Ciencias del Mar y Limnología, Av. Universidad 3000, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
b Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Biología Comparada, Laboratorio Algas Continentales, Ecología y Taxonomía, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
*Autor para correspondencia: alemirelesv@ciencias.unam.mx (A. Mireles-Vázquez)
Recibido: 24 marzo 2025; aceptado: 25 agosto 2025
Resumen
Heterokontophyta incluye una gran variedad de algas con distintos niveles de organización, ambientes, presencia de pigmentos e incluso líneas evolutivas. En México se han registrado 72 especies de aguas continentales que pertenecen a las clases Xanthophyceae y Eustigmatophyceae desde distintos enfoques: florístico, ecológico, limnológico, morfométrico, químico y filogenético. Con el objetivo de conocer la diversidad de algas doradas en el centro de México y actualizar los registros en bases de datos especializadas nacionales, se recolectó material en 8 cuerpos de agua de la región central; en cada sitio se filtraron 10 L con una red de fitoplancton. De algunas muestras se desarrollaron cultivos que permitieron obtener imágenes en microscopio electrónico de barrido. Se realizó un análisis de similitud con el índice de Jaccard y un agrupamiento de los sitios usando variables ambientales. Se identificaron y caracterizaron 18 especies de algas de las clases Xanthophyceae (10) y Eustigmatophyceae (8), 6 de ellas son nuevos registros para el país. Los análisis muestran una caracterización de los sitios y una distribución de las especies muy particular, pues los valores de similitud son de 0.125 a 0.33. Con estos resultados actualizamos el panorama de las algas doradas en la región central de México.
Palabras clave: Algas continentales; Florística; Heterokontophyta
Abstract
Heterokontophyta includes a wide variety of algae with different levels of organization, environments, presence of pigments and even evolutionary lines. In Mexico, 72 species of inland waters belonging to the classes Xanthophyceae and Eustigmatophyceae have been recorded from different approaches: floristic, ecological, limnological, morphometric, chemical and phylogenetic. To understand the diversity of golden algae in the central region of Mexico and update the records in specialized national databases, material was collected in eight bodies of water in the central region, at each site 10 L were filtered with a phytoplankton net. Cultures were developed from some samples that allowed obtaining images with a scanning electron microscope. A similarity analysis was performed with the Jaccard index and a grouping of the sites using environmental variables. Eighteen species of algae from the classes Xanthophyceae (10) and Eustigmatophyceae (8) were identified and characterized, 6 of which are new records for the country. The analyses show a very particular characterization of the sites and distribution of the species, since the similarity values are from 0.125 to 0.33. With these results we update the panorama of golden algae in the central region and their distribution.
La división Heterokontophyta está constituida por algas con una gran heterogeneidad de niveles de organización, ambientes, distribución geográfica, pigmentos principales y accesorios, ultraestructura celular y líneas evolutivas. Dos de las clases de esta división son representantes muy similares: Xanthophyceae y Eustigmatophyceae (Yang et al., 2012), que por más de 100 años estuvieron clasificadas en un solo grupo, Heterokontae (Rybalka, 2015).
En México se han registrado 72 especies continentales de Xanthophyceae y Eustigmatophyceae. Estos taxones están distribuidos en 19 entidades federativas de México registradas en 45 publicaciones, 17 corresponden a tesis (8 de licenciatura, 7 de maestría y 2 de doctorado), 18 en revistas periódicas y 10 son capítulos o libros (fig. 1). Las fuentes consultadas describen generalmente 1 o 2 especies con algún enfoque: 1a) listado florístico (Alcocer y Escobar, 1992; Cameron, 1960; Esqueda-Lara et al., 2016; Figueroa, 2009; Figueroa et al., 2015; Garduño-Solórzano et al., 2009; Godínez-Ortega et al., 2001; Hernández-Morales et al., 2011; López-Adrián y Barrientos-Medina, 2005; López-Adrián y Catzim, 2010; Mendoza-González, 1985; Mendoza-González et al., 1985; Montejano-Zurita et al., 2000, 2004; Moreno-Ruiz, 2005; Moreno-Ruiz et al., 2008; Novelo et al., 2023; Núñez-Márquez y Reyes González, 1995; Ortega-Murillo et al., 2014; Quiroz-Castelán et al., 2007; Vázquez y Blanco-Pérez, 2011); 1b) listado con ilustraciones o datos relacionados a otras especies (Aguirre-Cavazos et al., 2018; Carmona-Jiménez et al., 2016; Godínez-Ortega et al., 2017; Hernández-Morales et al., 2008; Moreno-Ruiz, 1985; Novelo et al., 2009; Ortega, 1984; Valadez-Cruz, 1998); 1c) con descripciones y datos de las especies (Adrián-Serrano, 2014; Agredano, 2019; Amateco-Rivero, 2011; Carmona-Jiménez, 1990; Flechtner et al., 1998; Flores-Granados, 1980; García-Rodríguez y Tavera, 1998; García-Rodríguez, 2004; González-Barrera, 1991; Martínez, 2016; Mireles, 2019; Moreno y Palacios, 1987; Novelo, 1998, 2014; Ochoa, 2023; Osorio y López, 2005; Pedraza, 2020; Ponce et al., 2019; Rodríguez-Flores, 2014; Sámano-Bishop, 1934; Valadez-Cruz et al., 1996; Zariñana-Leguízamo, 1997); 2) ecológico (Alvarado-Villanueva, 2003; Bojorge et al., 2010; Cantoral et al., 1999; Carmona-Jiménez y Montejano-Zurita, 1993; Carmona-Jiménez et al., 2022; Flórez, 2011; Jiménez, 2010; López-Adrián et al., 2017; Mora-Navarro, 2004; Mora-Navarro et al., 2004, 2006; Rodríguez-Flores y Carmona, 2018; Tavera y Díez, 2009; Valadez-Cruz et al., 2015); 3) limnológicos (Banderas-Tarabay, 1994, 1997; Ramírez, 2010; Sánchez-Gómez y Lara-Villa, 1986; Schmitter-Soto et al., 2002); 4) morfométrico (Chávez et al., 2005); 5) químico (Favari et al., 2002; Pérez-Gutiérrez et al., 2008); 6) filogenia molecular (Bonilla-Rodríguez et al., 2013).
La caracterización morfológica de las clases Xanthophyceae (Allorge ex F.E. Fritsch, 1935) y Eustigmatophyceae (D.J. Hibberd et Leedale, 1971) es complicada por varias causas: las poblaciones son pequeñas en las algas microscópicas y las macroscópicas, con frecuencia, no forman estructuras reproductoras. Las células de las algas microscópicas son generalmente pequeñas (menores de 15 micrómetros) y no se observan fácilmente en aumentos bajos. Hay muchas similitudes morfológicas con géneros de Chrysophyceae y Chlorophyceae, y además frecuentemente son confundidas entre ellas.
Analizar y trabajar los cuerpos de agua con énfasis en algas doradas es un desafío, sin embargo, dará información necesaria para avanzar en el conocimiento de la distribución geográfica y ambiental en la que viven estas algas. La agrupación (clustering) como herramienta de clasificación de los sitios de muestreo, permite identificar conjuntos de datos similares, las propiedades que se atribuyen (datos ambientales) se analizan y a mayor diferencia entre ellos, más alejados se muestran en un dendrograma (Rodríguez et al., 2019). El índice de Jaccard ha sido una medida fundamental en estudios de diversidad como éste, pues evalúa la coincidencia de un conjunto de datos (distribución de especies) y revela valores de menor o mayor similitud (Jaccard, 1901), es muy común en la comparación de diversidad de especies entre sitios o ecosistemas (Magurran, 2004). El objetivo de este trabajo es documentar la diversidad de algas doradas en la región central de México, su distribución a través de la similitud de presencia y ausencia en los ambientes sitios y actualizar las bases de datos especializadas nacionales.
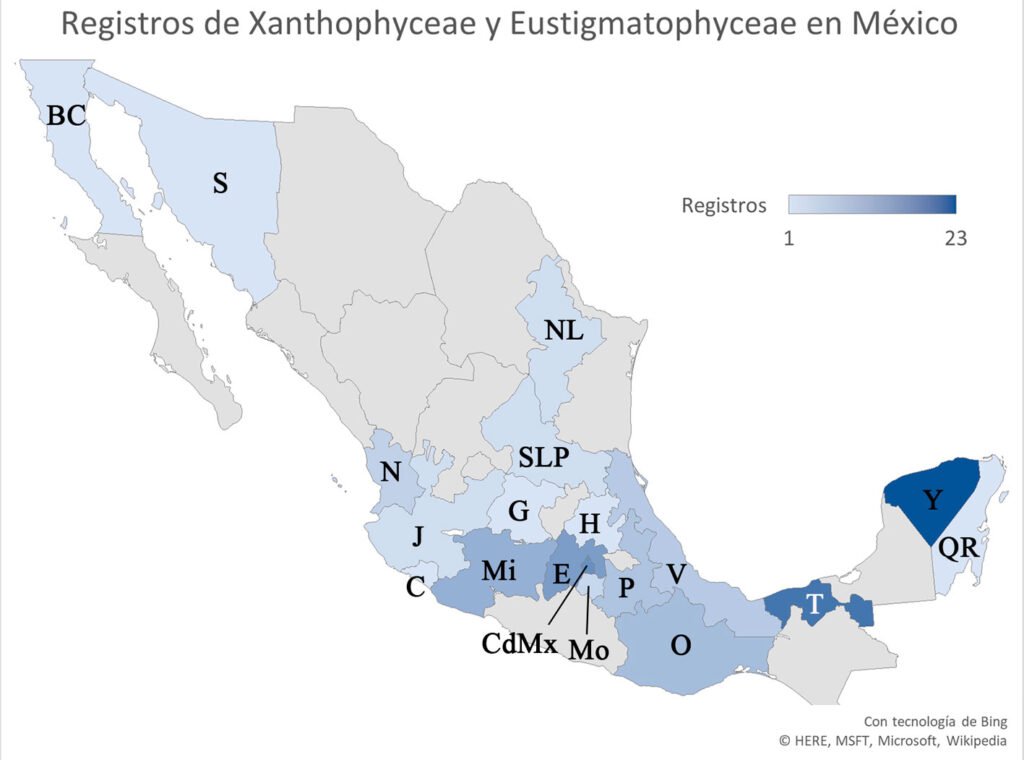
Figura 1. Registros de especies en México. BC: Baja California, C: Colima, CDMX: Ciudad de México, E: Estado de México, G: Guanajuato, H: Hidalgo, J: Jalisco, Mi: Michoacán, Mo: Morelos, N: Nayarit, NL: Nuevo León, O: Oaxaca, P: Puebla, Q: Quintana Roo, SLP: San Luis Potosí, S: Sonora, T: Tabasco, V: Veracruz, Y: Yucatán.
Materiales y métodos
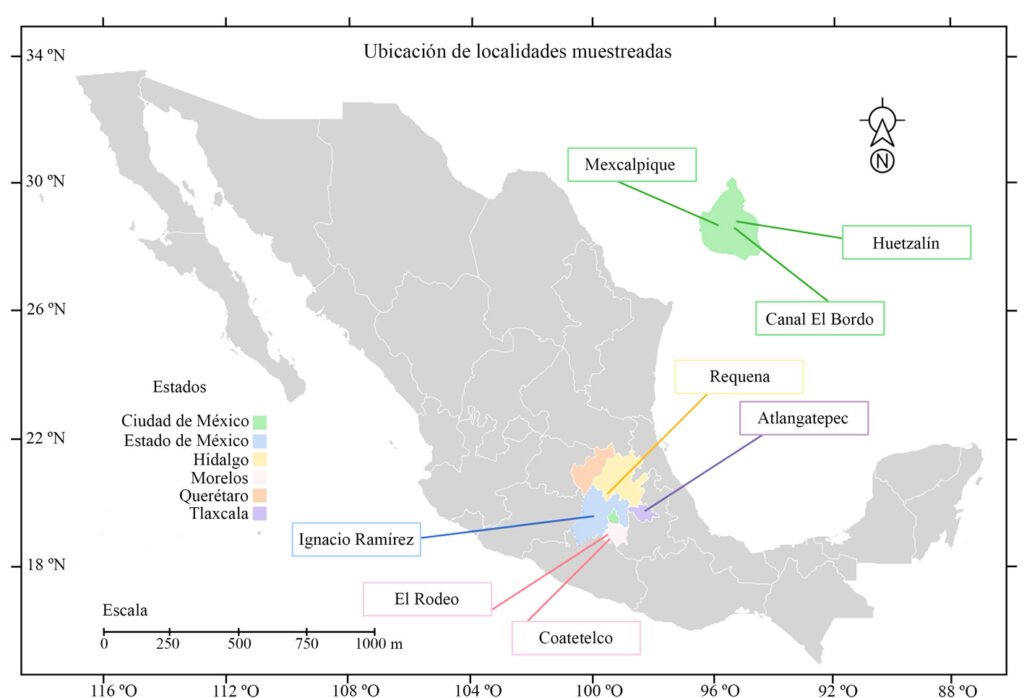
Los cuerpos de agua muestreados en este trabajo corresponden a sitios con registros previos de especies de estas clases (Cantera Oriente, canal El Bordo, lago Huetzalín, presa Ignacio Ramírez) y otras localidades que se agregaron como posibles ambientes en donde podríamos encontrar las especies de interés (presa Requena, lagos Coatetelco, El Rodeo y Atlangatepec). En total, se muestrearon 8 lagos en la región central de México contemplando la Ciudad de México, Estado de México, Hidalgo, Morelos y Tlaxcala (fig. 2, tabla 1).
Tabla 1
Información sobre las localidades y fechas de colecta.
Estado/ municipio
Cuerpo de agua
Número de localidad
Fecha de colecta
Muestra de herbario
Ciudad de México Xochimilco
Huetzalín
1
13 de octubre de 2021
FCME_CM971
Coyoacán
Mexcalpique
2
19 de octubre de 2021
FCME_CM972-FCME_CM973
Xochimilco
Canal El Bordo
3
26 de octubre de 2021
FCME_CM974-FCME_CM977
Estado de México Mayorazgo
Presa Ignacio Ramírez
4
16 de marzo de 2022
FCME_EDM64-FCME_EDM65
Hidalgo Entre Tepeji del Río de Ocampo y Tula de Allende
Presa Requena
5
20 de marzo de 2023
FCME_HGO090-FCME_HGO091
Morelos Jojutla
Coatetelco
6
26 de mayo de 2023
FCME_MOR002
Miacatlán
El Rodeo
7
26 de mayo de 2023
FCME_MOR003
Tlaxcala Atlangatepec
Atlangatepec
8
23 de junio de 2023
FCME_TLX001-FCME_TLX002
Localidad 1: lago Huetzalín pertenece al Parque Ecológico Xochimilco (PEX), una zona de recuperación ubicada en Xochimilco con una superficie de 1.9 km2 (Tavera et al., 2018). Localidad 2: la Cantera Oriente (CO) área rodeada por una pared de basalto (con una altura de hasta 40 m) que alberga varios cuerpos de agua con una superficie total de 11,906.45 m2 (Lot, 2007), perteneciente a la Reserva Ecológica del Pedregal de San Ángel. Localidad 3: el canal El Bordo es uno de los componentes de la zona chinampera de Xochimilco establecida como área natural protegida desde 1992, inscrita en la lista de humedales de importancia internacional por la Convención Internacional sobre Humedales Ramsar en 2004 (Figueroa et al., 2015). Localidad 4: presa Ignacio Ramírez (municipio Mayorazgo, Estado de México) es el acuífero más importante de la zona pues cuenta con disponibilidad de agua subterránea. Localidad 5: presa Requena (Hidalgo) es indispensable para la regulación de los escurrimientos del río Tula (Ríos, 2009), además de que está contemplada en el programa de ordenamiento ecológico local del municipio de Tepeji del Río de Ocampo desde 2004 (Valdez, 2012). Localidad 6: lago Coatetelco recibe aportes de arroyos del municipio Miacatlán, es un espacio de gran importancia social y económica (Macedonio, 2024). Los lagos Coatetelco y El Rodeo (localidad 7) representan los principales recursos hídricos del estado de Morelos. Ambos están dentro del programa de ordenamiento ecológico con el objetivo de restaurar las funciones ecológicas del sitio y disminuir acciones antropogénicas (Conabio, 2020). Localidad 8: lago Atlangatepec, representa un recurso relevante dentro de su territorio, en Tlaxcala, pues es el más grande y es alimentado por el río Zahuapan, el cual representa un riesgo por la contaminación que presenta por la descarga de material urbano e industrial (López-Segovia et al., 2024).
En cada sitio se filtraron 10 L de agua con una red de fitoplancton (10 µm), este material se concentró en 250 mL y una alícuota de 20 mL se guardó como muestra de herbario y en otro frasco de 200 mL como material de cultivo y observación. El frasco de cultivo se mantuvo en hielo hasta llegar al laboratorio y observar el material vivo. El frasco que se incorporó a las muestras de herbario (FCME) se preservó usando formaldehído al 2.5%. Se tomaron datos de las variables fisicoquímicas: radiación fotosintéticamente activa (PAR, sensores HOBO Penclant Couple MR), temperatura, pH, conductividad, oxígeno disuelto y clorofila a con una sonda multiparamétrica (Hydrolab DS 5 MR).
Figura 2. Ubicación de los lagos que se muestrearon en este estudio (Google Earth, 2025).
El cultivo de las especies se realizó en 2 medios de cultivo líquido: bold basal y bristol. El material filtrado se revisó y al identificar especies de interés se continuó con el aislamiento de ejemplares usando un capilar y material de succión. Los cultivos se ubicaron en condiciones de luz y oscuridad (16:8), con un promedio de 3,150 luxes y un intervalo de temperatura de 17-32 °C. Una parte del material de cultivo se procesó para fotografiarlo con microscopía electrónica de barrido. El procedimiento que se realizó usando glutaraldehído fue siguiendo a Takano y Hiroguchi (2005) para su observación. Los equipos utilizados fueron: Jeol JSM-5310LV y Hitachi SU1510.
Se realizaron preparaciones semipermanentes usando el material fijado (2.5%) y gelatina glicerinada (González-González y Novelo, 1986). La observación del material se realizó en un microscopio con contraste interdiferencial (Nikon 80i), para confirmar algunos caracteres morfológicos. Las fotografías se tomaron con una cámara Jenoptik adecuada al microscopio. Las fotografías y láminas se editaron para mejorar el contraste con el programa Photoshop (Adobe Photoshop [versión 26.8.1, 2025]).
En la identificación de las especies, las principales fuentes de consulta fueron: Bourrelly (1952, 1981), Ettl (1978), Ettl y Gärtner (1995); John et al. (2002), Pascher et al. (1925) y Starmach (1968). Y de manera particular se usaron: Bicudo et al. (2006: Characiopsis anabaenae, Chloridella cystiformis, Goniochloris mutica, Isthmochloron lobulatum, Pseudopolyedriopsis skujae, Tribonema minus); Hollerbach (1962: Pseudopolydriopsis skujae); Islam e Irfanullah (2001: Tetraplektron torsum); Krienitz et al. (1993: Pseudogoniochloris tripus); Krienitz et al. (2000: Nannochloropsis limnetica); Novelo et al. (2009: Botrydiopsis arhiza y Chloridella cystiformis).
Los datos de distribución geográfica y ambiental se obtuvieron de bdLACET (Novelo y Tavera, 2025), AlgaeBase (Guiry y Guiry 2025) y GBIF (Global Biodiversity Information Facility, 2025). El análisis de similitud con el índice de Jaccard se realizó con el programa PAST versión 4.17 (Hammer et al., 2001), construyendo una matriz de presencia y ausencia de las especies en los 8 sitios (con los parámetros establecidos en el programa). El análisis de agrupamiento (con distancia euclidiana, con el algoritmo UPGMA) se realizó utilizando los datos ambientales medidos en cada sitio de muestreo: temperatura (°C), pH, conductividad (µS), oxígeno disuelto (mg/L) y concentración de clorofila a (µg/L) (se realizó con los parámetros establecidos en el programa), los datos se transformaron con logaritmo base 10.
Resultados
Los datos de variables ambientales obtenidos en campo de los 8 lagos se presentan en la tabla 2, así como también la distribución de las especies por sitio y registro previos en México. El análisis de similitud con el índice de Jaccard se muestra en la tabla 3. Los valores obtenidos nos indican valores bajos de similitud en relación con las especies distribuidas en los sitios. Los valores obtenidos están en un intervalo de 0 a 0.33.
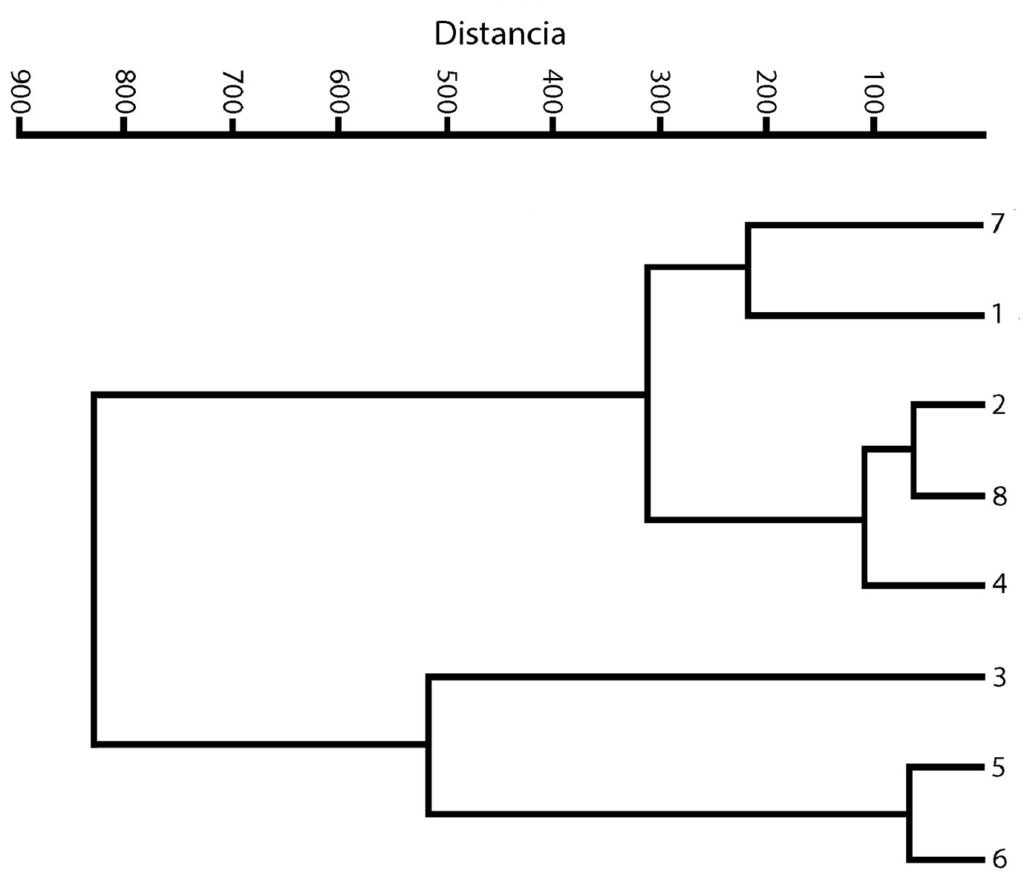
El agrupamiento de los sitios se representa en la figura 3. Los grupos que se formaron en el análisis son 3. El primer grupo (con los sitios más cercanos) es el que se representa con los números 2, 4 y 8, que corresponden a los sitios Mexcalpique (2), presa Ignacio Ramírez (4) y Atlangatepec (8); el siguiente grupo, con los sitios 1 (Huetzalín) y 7 (El Rodeo), y el último grupo con los sitios 5 (presa Requena), 6 (Coatetelco) y 3 (Bordo). En las muestras se identificaron en total 18 especies en la región central de México y corresponden a las 2 clases, de acuerdo con bases de datos actualizadas (bdLACET y AlgaeBase), 10 de ellas se clasifican en la clase Xanthophyceae y 8 en la clase Eustigmatophyceae.
Xanthophyceae P. Allorge ex F.E. Fritsch
Rhizochloridales Pascher
Stipitococcaceae Ettl
Stipitococcus W. et G. S. West
Stipitococcus capensis Prescott, 1937 (fig. 4)
Células solitarias, lorica con forma elipsoidal-ovoide ligeramente alargada, en la base presenta un pedúnculo corto que se adhiere a otras algas (encontradas sobre células de Stephanocyclus meneghinianus). Célula elipsoidal con un plastidio lobulado. Las células miden 7.8-9.7 µm de largo y 4.0-6.3 µm de ancho, con un pedúnculo corto de hasta 2.0 µm.
Distribución en México: Oaxaca, Ciudad de México (Xochimilco). Mundial: España, EUA e India.
Ambientes: estanques, lagos.
Formas de vida: planctónica, epífita y metafítica.
Valores de variables fisicoquímicas obtenidos en campo en los 8 sitios de muestreo: 1) Huetzalín – PEX; 2) Mexcalpique – Cantera Oriente; 3) canal El Bordo – Xochimilco; 4) presa Ignacio Ramírez; 5) presa Requena; 6) Coatetelco; 7) El Rodeo; 8) Atlangatepec.
Variable y especie/ sitio
1
2
3
4
5
6
7
8
Distribución en México
Temperatura (°C)
24
16.4
15.4
24
19.4
32.8
29.9
26.1
pH
8.7
8.1
6.9
9.1
9.5
8
8.3
9.2
Conductividad (µS)
0
364
932
291
826
878
168
401
DO (mg/L)
6.19
7.12
0.22
10.6
5.26
5.06
5.78
8.25
Clorofila a (µg/L)
0.96
59.05
3.75
29.94
6.94
6.31
41.1
26.97
Stipitococcus capensis
x
CdMx, Oaxaca
Chloridella cystiformis
x
x
CdMx, Tabasco
Ellipsoidion oocystoides
x
Nuevo Registro
Monodus guttula
x
Nuevo Registro
Tetraplektron torsum
x
Michoacán, Yucatán.
Pseudogoniochloris tripus
x
x
Tabasco, Nayarit.
Isthmochloron lobulatum
x
Tabasco, Nayarit.
Pseudopolydriopsis skujae
x
Nuevo registro
Botrydiopsis arhiza
x
Jalisco
Tribonema minus
x
x
CdMx
Nannochloropsis limnetica
x
Nuevo registro
Characiopsis anabaenae
x
Nuevo registro
Goniochloris mutica
x
x
x
CdMx, Tabasco, Nayarit.
G. iyengarii
x
x
x
Tabasco
Pseudostaurastrum limneticum
x
Estado de México
Tetraedriella acuta
x
Tabasco
Trachydiscus minutus
x
Tabasco
T. lenticularis
x
Nuevo registro
Tabla 3
Valores de similitud (índice de Jaccard).
1
2
3
4
5
6
7
8
1
1
0.16666667
0.14285714
0
0
0
0
0.14285714
2
0.16666667
1
0
0
0
0.14285714
0
0
3
0.14285714
0
1
0
0
0.125
0
0.33333333
4
0
0
0
1
0
0
0
0
5
0
0
0
0
1
0
0
0
6
0
0.14285714
0.125
0
0
1
0
0.125
7
0
0
0
0
0
0
1
0
8
0.14285714
0
0.33333333
0
0
0.125
0
1
Figura 3. Dendrograma. Agrupamiento de los 8 sitios recolectados.
Células solitarias, generalmente esféricas con pared lisa a veces amarillenta y gruesa, 2-4 plastidios discoidales, parietales. Las células miden 4.8-9.3 µm de diámetro.
Distribución en México: Ciudad de México (Cantera Oriente, PEX), Tabasco. Mundial: Alemania, Brasil, España, Eslovaquia, EUA, Polonia, República Checa, Rusia y Ucrania.
Ambientes: estanques y lagos.
Forma de vida: metafítica.
Referencias: Bicudo et al., 2006; Bourrelly, 1981; Ettl, 1978; Novelo et al., 2009; Pascher et al., 1925.
Nota: se encuentra en grandes cantidades en cuerpos de agua con un alto contenido de hierro y valores ácidos de pH (Ettl, 1978).
Células elipsoidales con extremos redondeados, generalmente 1.5 veces más largas que anchas, pared celular lisa, un plastidio laminar parietal. Las células miden 5.8-6.5 µm de largo y 2.9-3.8 µm de ancho.
Distribución en México: primer registro en Estado de México (presa Ignacio Ramírez) y México. Mundial: Alemania, Australia, Austria, Brasil, Corea del Norte, Eslovaquia, España, Islandia, Japón, República Checa, Rumania, Rusia y Tayikistán.
Ambientes: lagos y suelos.
Formas de vida: metafítica, edáfica.
Nota: común en aguas y suelos ácidos (Ettl, 1978).
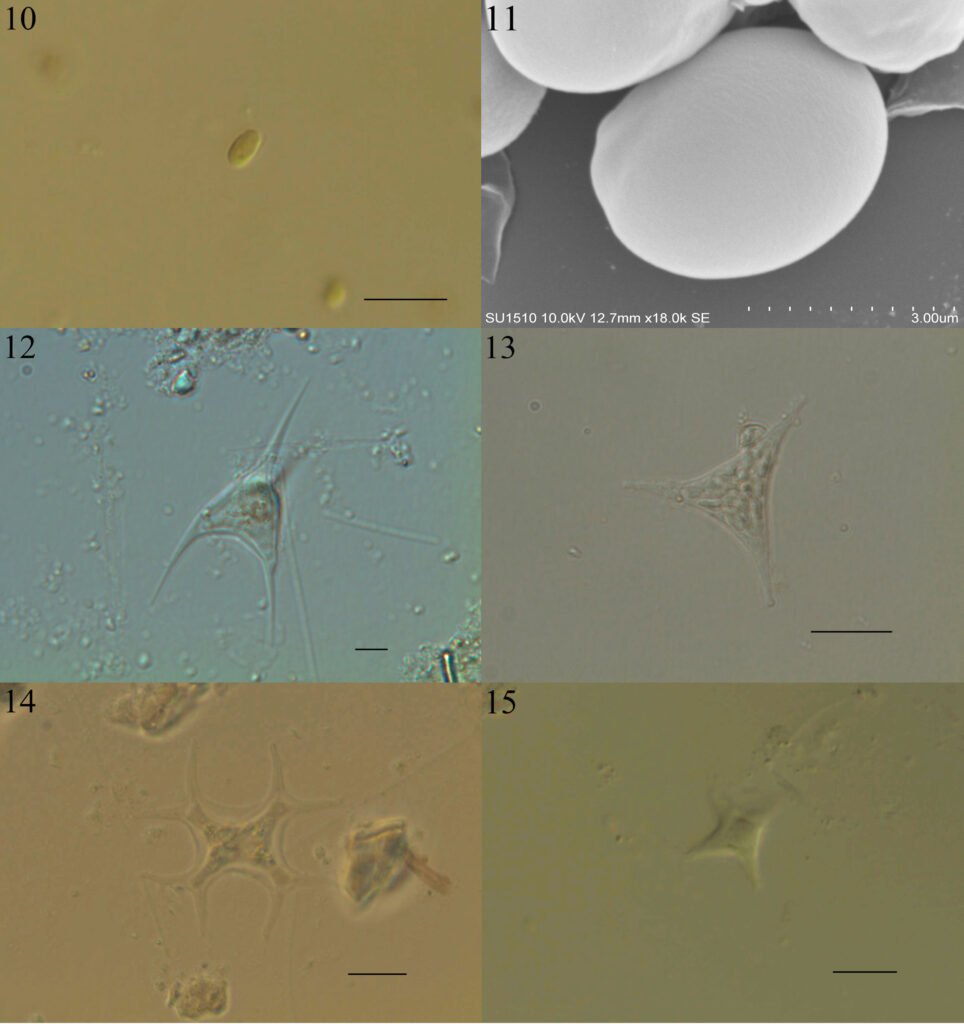
Figuras 4-9. 4, Stipitococcus capensis; 5, Chloridella cystiformis; 6, C. cystiformis (MEB); 7, Ellipsoidion oocystoides; 8, E. oocystoides (MEB); 9, Monodus guttula (cultivo). Barra = 10 µm.
Monodus Chodat
Monodus guttula Pascher, 1938 (figs. 9-11)
Células solitarias globosas u ovadas, puntiagudas en un extremo, ligeramente curvas, pared lisa y delgada, un plastidio laminar parietal. Las células miden 3.8-4.9 µm de largo y 2.2-2.9 µm de ancho.
Distribución en México: primer registro en Ciudad de México (Cantera Oriente). Mundial: Alemania, Bulgaria, Eslovaquia, España, Islas Shetland del Sur, Moldavia, República Checa, Rumania y Rusia.
Ambientes: lagos.
Formas de vida: metafítica.
Nota: se encuentran en lugares húmedos y zanjas en las cordilleras entre Alemania y la República Checa (Ettl, 1978).
Células solitarias, tetraédricas aplanadas, cada lado termina en una espina, un lado enmascara otro, pared celular lisa y gruesa, varios plastidios discoidales. Las células miden 44.2- 46.6 µm de largo y 22.1-22.9 µm de ancho.
Distribución en México: Morelos (El Rodeo), Michoacán y Yucatán. Mundial: Alemania, Australia, Bangladesh, Brasil, Costa de Marfil, EUA, Italia, Myanmar, Países Bajos, Paraguay, Suecia y Ucrania.
Ambientes: lagos.
Formas de vida: Planctónica, perifítica y metafítica.
Referencias: Bourrelly, 1981; Ettl, 1978; Islam e Irfanullah, 2001; John et al., 2002; Starmach, 1968.
Pseudogoniochloris (Pascher) Krienitz, Hegewald, Reymond et Peschke
Células triangulares irregulares (ligeramente torcidas) con bordes agudos, lados cóncavos, pared celular lisa o ligeramente ornamentada, 3-5 plastidios (puede haber más) discoidales, parietales. Las células miden 19.5-28.3 µm por lado.
Distribución en México: Ciudad de México (Xochimilco), Tlaxcala (Atlangatepec), Tabasco y Nayarit. Mundial: Alemania, Argentina, Australia, Austria, Bangladesh, Bélgica, Bielorrusia, Botsuana, Brasil, China, Eslovaquia, EUA, Estonia, Francia, India, Irlanda, Israel, Japón, Países Bajos, Paraguay, Reino Unido, República Checa, Rumania, Rusia, Suecia, Sudáfrica, Turquía y Ucrania.
Células solitarias, cuadrangulares con lados cóncavos que se dividen dicotómicamente al final, pared celular lisa, varios plastidios discoidales, parietales. Las células miden 7.8-11.7 en la parte central, sin brazos y miden 28.2-34.7 µm de cada lado con brazos.
Distribución en México: Morelos (Coatetelco), Tabasco y Nayarit. Mundial: en los 5 continentes.
Ambientes: lagos.
Formas de vida: metafítica y planctónica.
Referencias: Bicudo et al., 2006; Bourrelly 1952, 1981; Ettl, 1978; John et al., 2002; Starmach, 1968.
Células solitarias, aplanadas con 5 proyecciones angulosas, pared celular gruesa, ornamentación muy fina, varios plastidios discoidales parietales. Las células miden 9.3-17.1 µm de cada lado. Presentan 2-4 espinas de 13.2-18.6 µm en cada proyección.
Distribución en México: primer registro en México, Ciudad de México (Xochimilco). Mundial: Alemania, Australia, Bélgica, Bielorrusia, Brasil, EUA, Estonia, Francia, Países Bajos y Suecia.
Ambientes: lagos.
Formas de vida: metafítica.
Nota: los ejemplares observados sólo presentan espinas en campo, no en los cultivos.
Células esféricas, generalmente en grupos unidos por mucílago incoloro o pardo, pared celular gruesa a muy gruesa, varios plastidios discoidales o rectangulares. Las células miden 13.6-23.5 µm de diámetro.
Distribución en México: Ciudad de México (Cantera Oriente), Jalisco. Mundial: EUA, Japón, India, Países Bajos, Portugal, Reino Unido, Rusia, Suecia y Ucrania.
Ambientes: suelos, lagos, arenas y charcos.
Formas de vida: edáfica, planctónica y metafítica.
Referencias: Bourrelly, 1981; Ettl, 1978; John et al., 2002; Novelo et al., 2009; Pascher et al., 1925; Starmach, 1968.
Tribonematales Pascher
Tribonemataceae Pascher
Tribonema Derbès et Solier
Tribonema minus Wille (Hazen), 1902 (fig. 19)
Filamentos uniseriados, cortos, rectos o curvos. Células cilíndricas con septos definidos y sin constricción, pared celular lisa, 2 a 3 plastidios laminares, parietales. Las células miden 4.4-5.9 µm de ancho y 17.5-26.3 µm de largo.
Distribución en México: Ciudad de México (Cantera Oriente). Mundial: Alemania, Arabia Saudita, Austria, Brasil, Canadá, España, EUA, Estonia, Grecia, Hungría, Nigeria, Países Bajos, Polonia, Reino Unido, Suecia, Ucrania y Uzbekistán.
Ambientes: ríos, arroyos, lagos, humedales, charcos, estanques, embalses y turberas.
Formas de vida: bentónica, metafítica, planctónica.
Referencias: Bicudo et al., 2006; Bourrelly, 1981; Ettl, 1978; Ettl y Gärtner, 1995; John et al., 2002; Starmach, 1968.
Células esféricas, ovaladas o en forma de gota, pared celular lisa, 1-2 plastidios discoidales, parietales. Las células miden 3.1-3.4 µm de diámetro.
Distribución en México: Ciudad de México (Cantera Oriente). Mundial: Alemania, Canadá, China, Curazao, Dinamarca, Eslovenia, EUA, Finlandia, Francia, Grecia, Israel, Noruega, Polonia, Reino Unido, Rusia y Suecia.
Células solitarias, pequeñas, fusiformes, con un lado más convexo que el otro, terminación aguda de un extremo en donde se encuentra el disco basal, pared celular lisa, 2 plastidios laminares. Las células miden 10.7-18.6 µm de largo y 2.0-5.9 µm de ancho.
Distribución en México: primer registro en México, Hidalgo (Presa Requena). Mundial: Afganistán, Alemania, Brasil, Países Bajos, Portugal, Rumania, Rusia y Ucrania.
Ambientes: lagos.
Forma de vida: metafítica.
Referencias: Bicudo et al., 2006; Bourrelly, 1981; Ettl, 1978; Ettl y Gärtner, 1995; Starmach, 1968.
Goniochloridales K.P. Fawley, M. Eliás et M.W. Fawley
Células solitarias, triangulares regulares con bordes redondeados, lados cóncavos, pared celular ornamentada con pequeñas verrugas, 2-3 plastidios discoidales. Las células miden 5.8-9.8 µm de cada lado.
Distribución en México: Ciudad de México (Xochimilco), Tlaxcala (Atlangatepec), Tabasco y Nayarit. Mundial: Alemania, Australia, Argentina, Bangladesh, Bélgica, Bielorrusia, Botswana, Brasil, Burindi, Canadá, China, Costa de Marfil, Eslovaquia, España, EUA, Estonia, Francia, India, Irak, Irán, Israel, Italia, Niger, Nueva Zelanda, Países Bajos, Portugal, Reino Unido, República Checa, Rumania, Rusia, Sudán, Suecia, Taiwán, Tayjikistán, Turquía y Ucrania.
Ambientes: humedales y lagos.
Formas de vida: planctónica y metafítica.
Referencias: Bicudo et al., 2006; Bourrelly 1952, 1981; Ettl, 1978; John et al., 2002; Starmach, 1968.
Células solitarias, triangulares con terminaciones agudas, lados ligeramente cóncavos, pared celular gruesa con ornamentación escrobiculada, varios plastidios discoidales, parietales. Las células miden 22.3-26.4 µm de cada lado.
Distribución en México: Ciudad de México (Xochimilco), Morelos (Coatetelco), Tlaxcala (Atlangatepec), Tabasco. Mundial: Alemania, Brasil, Francia (Guadaloupe), India y Países Bajos.
Ambientes: charcos y lagos.
Formas de vida: metafítica.
Referencias: Bourrelly 1981; Ettl, 1978; John et al., 2002; Starmach, 1968.
Figuras 16-21. 16, Botrydiopsis arhiza (campo); 17, B. arhiza (cultivo); 18, B. arhiza (MEB); 19, Tribonema minus; 20, Nannochloropsis limnetica; 21, N. limnetica (MEB). Barra = 10 µm.
Células solitarias, tetraédricas con cuerpo central ligeramente amplio que termina en 4 brazos radiados, bifurcados en los extremos, pared celular lisa y gruesa, varios plastidios discoidales y parietales. Las células miden 29.3-34.9 µm de cada lado.
Distribución en México: Tlaxcala (Atlangatepec) y Estado de México. Mundial: Alemania, Argentina, Australia, Bangladesh, Bioelorrusia, Brasil, Canadá, China, Cuba, España, EUA, Estonia, India, Irak, Irlanda, Israel, Japón, Latvia, Mianmar, Nueva Zelanda, Países Bajos, Paquistán, Portugal, Reino Unido, República Checa, Rumania, Rusia, Sudán, Suecia, Taiwán, Turquía, Ucrania y Venezuela.
Ambientes: lagos.
Formas de vida: planctónica y metafítica.
Referencias: Bourrelly, 1981; Ettl, 1978; John et al., 2002; Starmach, 1968.
Tetraedriella Pascher
Tetraedriella acuta Pascher, 1930 (fig. 27)
Células solitarias, tetraédricas, con lados convexos y bordes ligeramente afilados, pared celular gruesa ornamentada en forma de panal con bordes aún más engrosados, 4 plastidios discoidales. Las células miden 10.3-13.7 µm de cada lado.
Distribución en México: Tabasco y Ciudad de México (Xochimilco). Mundial: Alemania, Brasil, Bulgaria, Costa de Marfil, España, Estonia, Francia, Países Bajos y República Checa.
Figuras 22-27. 22, Characiopsis anabaenae; 23, C. anabaenae (MEB); 24, Goniochloris mutica, 25; G. iyengarii, 26; Pseudostaurastrum limneticum, 27; Tetraedriella acuta. Barra = 10 µm.
Células solitarias, pentagonales redondeadas en los ángulos, pared celular delicadamente ornamentada, 4 a 5 plastidios discoidales. Las células miden 6.8-9.8 µm de diámetro.
Distribución en México: Tlaxcala (Atlangatepec) y Tabasco. Mundial: Argentina, Australia, Bélgica, Bulgaria, Canadá, China, Colombia, Corea del Sur, EUA, Finlandia, Francia, Grecia, Polonia, Reino Unido, República Checa, Suecia, Turquía y Ucrania.
Células solitarias, lenticulares, biconvexas, pared celular ornamentada con verrugas pequeñas, 3 plastos discoidales. Las células miden 8.6-9.8 µm de diámetro.
Distribución en México: primer registro en México, Tlaxcala (Atlangatepec). Mundial: Alemania, Bélgica, España, Francia, Irlanda, Países Bajos, República Checa y Reino Unido.
Figuras 28-29. 28, Trachydiscus minutus; 29, T. lenticularis. Barra = 10 µm.
Discusión
Los registros previos de especies de las Xanthophyceae y Eustigmatophyceae en los sitios seleccionados de la región central de México se muestran en la tabla 4. En la Ciudad de México, de las 8 especies registradas, confirmamos la presencia actual de 3 de ellas, Botrydiopsis arhiza, Chloridella cystiformis y Tribonema minus. B. arhiza ha sido reportada en 3 trabajos, el primero, en el catálogo de algas de Ortega (1984); el segundo, como un taxón a comparar que se encuentra por periodos en distintos cuerpos de agua siendo parte de distintas comunidades: fitoplancton, metafiton y perifiton (Novelo et al., 2009); nosotros la encontramos en un cuerpo de agua estancado dentro de la Cantera Oriente, en octubre, formando un crecimiento evidente en la superficie casi monoespecífico de fitoplancton; el tercer registro, menciona la especie como parte de las algas de la cuenca de la ciudad (Figueroa, 2009). Previamente registrada en EUA, varios países de Europa, Rusia, Japón e India.
C. cystiformis fue registrada por primera vez en la Cantera Oriente como una especie de ambientes templados y ácidos (Novelo et al., 2009); en este trabajo se registra en el mismo sitio, lago Mexcalpique, en el metafiton, corroborando su presencia y crecimiento en la misma comunidad, sin embargo, las condiciones no son similares pues los valores de pH no son ácidos (como se ha registrado previamente en localidades de Europa), sino básicos en el sitio donde la observamos (tabla 2), además también la encontramos en el lago Huetzalín (PEX) y en sitios con valores de pH básico.
En el caso de T. minus, se ha reportado en más ocasiones, ejemplo de la desigualdad de trabajos que registran especies de mayor talla, en este caso filamentosas que especies unicelulares, en canales de Xochimilco (Figueroa, 2009; Figueroa et al., 2015; Flores, 1980; González-Barrera, 1991) y en Cantera Oriente (Ponce et al., 2019). Esta especie ha sido registrada previamente en localidades frías de Europa, pero también en templadas o cálidas como EUA y Brasil.
Como nuevos registros de la Ciudad de México se tienen a Monodus guttula y Pseudopolydriopsis skujae. La primera descrita para el lago Mexcalpique en la Cantera Oriente, con registros previos en Europa y la segunda para el canal El Bordo en Xochimilco, presente en Europa y en Brasil. Por último, algunos registros que no estaban reportados en las zonas seleccionadas, pero sí en el país son: Stipitococcus capensis, en el Bordo, en Xochimilco, con registros en Europa, Norteamérica y Asia, en México se había descrito en Oaxaca. Pseudogoniochloris tripus, previamente registrado en Tabasco (Agredano, 2019; Mireles, 2019; Pedraza, 2020) y Nayarit (Ochoa, 2023), de amplia distribución mundial, se encontró en el canal Bordo; Nannochloropsis limnetica, se registra en el lago Mexcalpique, es una especie que ha proliferado en muchos ambientes en el mundo, ahora como primer registro en Cantera Oriente. En el lago Huetzalín, se observaron en el metafiton a Goniochloris mutica, con registros previos en los canales de Xochimilco (Tavera y Diez, 2009), Tabasco (Mireles, 2019) y Nayarit (Ochoa, 2023) y amplia distribución mundial; así como a G. iyengarii con registro anterior en Tabasco (Mireles, 2019) y con poca distribución mundial; y a Tetraedriella acuta, con registro previo en Tabasco (Mireles, 2019), en Europa y Brasil.
Tabla 4
Especies registradas previamente en las zonas de la región central de México.
Estado (número de especies registradas)
Especie
Referencias
CdMx (8)
Botrydiopsis arhiza Borzì
Ortega, 1984 Novelo et al., 2009 Figueroa, 2009
Characiopsis naegelii (A. Braun) Lemmermann
Ortega, 1984, Figueroa, 2009
Tabla 4. Continúa
Estado (número de especies registradas)
Especie
Referencias
Chloridella cystiformis Pascher
Novelo et al., 2009
Dichotomosiphon tuberosus (A. Braun) Ernest
Figueroa, 2009 Cantoral et al., 1999
Ducelleria chodatii (Ducellier) Teiling
Godínez-Ortega et al., 2017
Gloeobotrys limneticus (G.M. Smith) Pascher
Ortega, 1984 Figueroa, 2009
Ophiocytium capitatum Wolle
Figueroa et al., 2015 Figueroa, 2009 Flores-Granados, 1980
Tribonema minus (Wille) Hazen
Figueroa, 2009 Flores-Granados, 1980 González-Barrera, 1991 Figueroa et al., 2015 Ponce et al., 2019
Estado de México (10)
Bumilleriopsis brevis (Gerenck) Printz
Moreno y Palacios, 1987
Characiopsis longipes (Rabenhorst) Borzì
Banderas-Tarabay, 1994 Banderas-Tarabay, 1997
Chlorocloster pyreniger Pascher
Favari et al., 2002
Ophiocytium capitatum Wolle
Chávez et al., 2005
O. cochleare (Eichwald) A. Braun
Mendoza, 1985 Ortega, 1984 Chávez et al., 2005 Figueroa, 2009
O. lagerheimii Lemmermann
Chávez et al., 2005
O. majus Nägeli
Chávez et al., 2005
O. parvulum (Perty) A. Braun
Zariñana-Leguízamo, 1997 Chávez et al., 2005
Vaucheria bursata (O.F. Müller) C. Agardh
Bojorge et al., 2010 Ramírez, 2010 Rodríguez-Flores, 2014 Rodríguez-Flores y Carmona, 2018
V. geminata (Vaucher) de Candolle
Ortega, 1984 Mendoza, 1985 Figueroa, 2009 Garduño-Solórzano et al., 2009 Ceballos et al., 2009
Morelos (3)
Ophiocytium capitatum Wolle
Flórez, 2011
Tribonema bombycinum (C.A. Agardh) Derbés et Solier
García-Rodríguez y Tavera, 1998 Quiroz-Castelán et al., 2007 García-Rodríguez, 2004
Vaucheria sessilis f. clavata (De Candolle) Heering
Valadez-Cruz et al., 1996 Valadez-Cruz, 1998
Ducelleria chodatii (Ducellier) Teiling
Godínez-Ortega et al., 2017
De los registros en el Estado de México (10) no corroboramos ninguna de las especies de la tabla 4. Favari et al. (2002) reportan para la presa Ignacio Ramírez a Chlorocloster pyreniger, en este estudio con la observación de material de campo tanto vivo como fijado, no se corroboró su presencia. En el caso de Ellipsoidion oocystoides, se registra por primera vez en el país, con reportes previos en zona templada de Europa, Japón y zonas tropicales como Brasil, Australia y Nueva Zelanda.
Para la presa Requena, Hidalgo, no existían registros publicados sobre este grupo de algas, en la revisión de las muestras de este estudio encontramos a Characiopsis anabaenae, primer registro para México, previamente descrito en Europa, Asia y Brasil.
En Morelos, los 4 registros previos de algas doradas no se corroboraron, incluso en el sitio que visitamos, El Rodeo. En el lago Coatetelco identificamos Goniochloris mutica (mencionada para Huetzalín) e Isthmochloron lobulatum que ya tenía registros previos en Tabasco (Mireles, 2019) y Nayarit (Ochoa, 2023) y con amplia distribución mundial. En el lago El Rodeo, también identificamos a I. lobulatum y Tetraplektron torsum, esta última se había registrado en Michoacán (Martínez, 2016) y Yucatán (López-Adrián y Catzim, 2010), en parte de Europa, Sudamérica y Asia. Pseudostaurastrum limneticum, previamente registrada en el Estado de México (Jiménez, 2010) y con amplia distribución mundial.
En Tlaxcala, en el lago Atlangatepec, identificamos 6 especies: Pseudogoniochloris tripus, Goniochloris mutica, G. iyengarii, Pseudostaurastrum limneticum, Trachydiscus minutus y T. lenticularis. T. minutus, con registro previo en Tabasco (Pedraza, 2020), Europa y Argentina, muy escasos son los trabajos con esta especie. Mientras, que T. lenticularis, es primer registro en México y solo se ha reportado en ambientes fríos de Europa, Alemania, Bélgica, España, Francia, Irlanda, Países Bajos, República Checa y Reino Unido (Ettl, 1978; Starmach, 1968).
En relación con los sitios de muestreo y las variables ambientales, el dendrograma obtenido muestra que los sitios más parecidos son: Mexcalpique, presa Ignacio Ramírez y Atlangatepec (localidades 2, 4 y 8, respectivamente), esto resulta así pues los valores ambientales son cercanos en 3 variables, conductividad, oxígeno disuelto y concentración de clorofila a. Los sitios: Huetzalín y El Rodeo (1 y 7, respectivamente) presentan valores cercanos de pH y de oxígeno disuelto. Finalmente, el último grupo, Bordo, presa Requena y Coatetelco (3, 5 y 6 respectivamente), presentan valores cercanos de conductividad y de concentración de clorofila a.
La distribución de las especies utilizando una matriz de presencia/ausencia y aplicando el índice de Jaccard, dio como resultado valores entre 0 y 0.33, estos valores indican una similitud de especies nula o baja entre los sitios (Magurran, 2004). Huetzalín (1) muestra valores 0 con la presa Ignacio Ramírez (4), Presa Requena (5), Coatetelco (6) y El Rodeo (7), un valor de 0.14286 con los lagos El Bordo (3) y Atlangatepec (8) y un valor de 0.167 con Mexcalpique, esto indica una similitud bastante baja, pues solo comparten una especie (Goniochloris mutica) y la mayoría de las especies son diferentes entre sitios. Los valores 0, nos indican que los elementos son 100% diferentes, mientras que en los otros sitios son casi 90% diferentes. Los sitios que muestran los valores más altos (0.334) son El Bordo (3) y Atlangatepec (8), esto es debido a que comparten la presencia de 3 especies (Pseudogoniochloris tripus, Goniochloris mutica y Goniochloris iyengarii). La presencia de especies en cada sitio prácticamente es única, esto se explica por la variación en las condiciones ambientales que pueden limitar el crecimiento de algunas especies y la distancia entre los sitios es lo suficiente para no compartir especies (Magurran, 2004).
Se esperaba que los grupos que se formaron en el dendrograma coincidieran con los sitios con más similitud con la presencia de especies, sin embargo, no ocurrió, lo que nos indica que son conjuntos casi únicos (Real y Vargas, 1996).
Todas las algas son un reto en su identificación, las algas doradas presentadas en este trabajo son un ejemplo de la diversidad que aún desconocemos en distintos sitios del país, seguir estudiando la flora dorada es indispensable en el avance de una solución taxonómica, en un contexto integral.
Agradecimientos
Al Posgrado en Ciencias del Mar y Limnología, UNAM, por los estudios dirigidos a la obtención del grado de Doctor. Al apoyo recibido de Conahcyt (beca con clave 2020-000026-02NACF). A Guadalupe Vidal Gaona por su apoyo en el laboratorio y en campo. A Beatriz Lira Hernández por su apoyo en campo. A Silvia Espinosa Matías de la Facultad de Ciencias por el procesamiento de muestras y obtención de imágenes con el microscopio electrónico de barrido Jeol JSM-5310LV. A María Berenit Mendoza Garfias del Laboratorio Nacional de Biodiversidad del Instituto de Biología por su servicio en el procesamiento de muestra y obtención de imágenes con el microscopio electrónico de barrido Hitachi SU1510. A los revisores, que nos dieron sugerencias para mejorar el manuscrito.
Referencias
Adrián-Serrano, J. (2024). Estudio de la biodiversidad fitoplanctónica del Canal Japón, Xochimilco, México D. F. (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Agredano, Z. O. (2019). Ficoflora de la laguna El Camarón en Villahermosa, Tabasco, México(Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Aguirre-Cavazos, D. E., Moreno-Limón, S. y Salcedo-Martínez, S. M. (2018). Especies de algas de ríos de Nuevo León, México. Nuevos registros para el estado. Polibotánica, 46, 1–25. https://doi.org/10.18387/polibotanica.46.1
Alcocer, J. y Escobar-Briones, E. (1992). The aquatic biota of the now extinct lacustrine complex of Mexico basin. Freshwater forum, 2, 171–183.
Alvarado-Villanueva, R. (2003). Dinámica estacional y circadiana del fitoplancton en el Lago Zirahuén,Michoacán, México (Tesis de maestría). Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo. Morelia.
Amateco-Rivero, F. S. (2011). Composición del fitoplancton del lago de Catemaco, Veracruz, México (Tesis). Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México. Estado de México.
Banderas-Tarabay, A. G. (1994). Limnología del lago El Sol, Nevado de Toluca, México(Tesis doctoral). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Banderas-Tarabay, A. G. (1997). Phycoflora of the tropical high-mountain lake El Sol, Central Mexico, and some biogeographical relationships. Hydrobiologia, 354, 17–40. https://doi.org/10.1023/A:1003082500767
Bicudo, D. C., Bicudo, C. E. M., Oliveira, A. M., Ferragut, C., Fonseca, B. M., Lopes, M. R. M. et al. (2006). Criptógamos do Parque Estadual das Fontes de Ipiranga, São Paulo, S.P. Algas, 21: Xanthophyceae. Hoehnea, 33, 291–316.
Bojorge, G. M. G., Carmona, J. J., Beltrán, M. Y. y Cartajena, A. M. G. (2010). Temporal and spatial distribution of macroalgal communities of mountain streams in Valle de Bravo Basin, central Mexico. Hydrobiologia, 641, 159–169. https://doi.org/10.1007/s10750-009-0074-5
Bonilla-Rodríguez, M., Garduño Solórzano, G., Martínez-García, M., Campos, J. E., Monsalvo Reyes, A. C. y Quintanar-Zúñiga, R. E. (2013). Vaucheria zapotecana (Xanthophyceae), a new species from Oaxaca, Mexico. Phycologia, 52, 550–556. https://doi.org/10.2216/13-162.1
Bourrelly, P. (1952). Xanthophycées rares ou nouvelles. Bulletin du Museum National d´Histoire Naturelle, Série 2, 23, 666–672.
Bourrelly, P. (1981). Les algues d’eau douce. Initiation à la Systématique. II. Les algues jaunes et brunes. Chrysophycées, Phéophycées, Xanthophycées et Diatomées. París: Société Nouvelle des Éditions Boubée.
Cameron, R. E. (1960). Communities of soil algae ocurring in the Sonoran Desert in Arizona. Journal of the Arizona Academy of Sciences, 1, 85–88.
Cantoral, U. E. A., Carmona, J. J., González-González, J. y Montejano, Z. G. (1999). Algas indicadoras de la calidad del agua en el río la Magdalena, Delegación Magdalena Contreras, Distrito Federal, México. En D. Ponce-N. y M. V. Alegre (Eds.), Consejo de estudios para la restauración y valoración ambiental (aire-agua) (pp. 299–332). Ciudad de México: Gobierno del Distrito Federal.
Carmona-Jiménez, J. (1990). Ficoflora de manantiales de la Huasteca Postosina (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México, México.
Carmona-Jiménez, J. y Montejano-Zurita, G. (1993). Carac- terización ficológica en manantiales de la cuenca baja del sistema hidrológico de Pánuco, México. Boletín de la Sociedad Botánica de México, 53, 21–41. https://doi.org/10.17129/botsci.1414
Carmona-Jiménez, J., Ramírez-Rodríguez, R., Bojorge-García, M., González-Hidalgo, B. Y. y Cantoral-Uriza, E. A. (2016). Estudio del valor indicativo de las comunidades de algas bentónicas: una propuesta de evaluación y aplicación en el río Magdalena, Ciudad de México. Revista Internacional de Contaminación Ambiental, 32, 139–152. https://doi.org/10.20937/RICA.2016.32.02.01
Carmona-Jiménez, J. Salinas-Camarillo, V. H. y Caro-Borrego, A. (2022). The Macroalgae Ecological Quality Index (MEQI) in the Basin of Mexico: a proposal of aquatic bioindicators for peri-urban rivers. Revista Mexicana de Biodiversidad, 93, 1–17. https://doi.org/10.22201/ib.20078706e.2022.93.3899
Ceballos, G., List, K., Garduño-Solórzano, G., López-Cano, R., Muñozcano-Quintanar, M. J., Collado, E. & San Román, J. E. (Eds.). (2009). La diversidad biológica del Estado de México. Estudio de estado. Toluca de Lerdo: Gobierno del Estado de México.
Chávez, V., Zariñana, L., Novelo, E. y Tavera, R. (2005). Variación morfológica de algunas especies de Ophiocytium Nägeli (Xanthophyceae) de cuerpos de agua temporales del Estado de México. Hidrobiológica, 15, 311–320.
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio). (2020). Sexto informe nacional al convenio sobre la diversidad biológica. Conabio. https://www.biodiversidad.gob.mx/media/1/planeta/internacional/files/6IN_WEB_CLOSE.pdf
Esqueda-Lara, K., Sánchez, A. D. J., Valdés-Lagunes, G., Salcedo, M. Á., Franco-Torres, E. Y. y Florido, R. (2016). Fitoplancton en el humedal tropical Chaschoc en la cuenca baja del río Usumacinta. Revista Mexicana de Biodiversidad, 87, 1177–1188. https://doi.org/10.1016/j.rmb.2016.10.015
Ettl, H. (1978). Xanthophyceae. Teil 1. Stuttgart: Gustav Fischer Verlag.
Ettl, H. et Gärtner, G. (1995). Syllabus der Boden, Luft- und Flechtenalgen. Stuttgart: Gustav Fischer Verlag.
Favari, L., López, E., Martínez-Tabché, L. y Diaz-Pardo, E. (2002). Effect of insecticides on plankton and fish of Ignacio Ramírez Reservoir (Mexico): a biochemical and biomagnification study. Ecotoxicology and Environmental Safety, 51, 177–186. https://doi.org/10.1006/eesa.2002.2142
Figueroa, T. M. G. (2009). Algas de la cuenca de México. (Serie Académicos, CBS, UAM-Xochimilco). México D.F.: Universidad Autónoma Metropolitana. Unidad Xochimilco.
Figueroa, T. M. G., Arana-Magallón, F., Almanza-Encarnación, S., Ramos-Espinosa, M. G. y Ferrara-Guerrero, M. J. (2015). Microalgas del área natural protegida ejidos de Xochimilco y San Gregorio Atlapulco, México. Ciencia UAT, 9, 15–29.
Flechtner, V. R., Johansen, J. R. et Clark, W. H. (1998). Algal composition of microbiotic crusts from the central desert of Baja California, México. Great Basin Naturalist, 58, 295–311. https://doi.org/10.2307/41713069
Flores-Granados, C. (1980). Variaciones estacionales en la composición florística del fitoplancton de dos canales de Xochimilco, D.F. México (Tesis). Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. México D.F.
Flórez, M. A. E. (2011). Composición y variación temporal de los grupos funcionales fitoplanctónicos en un embalse tropical(Tesis de maestría). Posgrado en Ciencias del Mar y Limnología (Limnología), Universidad Nacional Autónoma de México. México D.F.
García-Rodríguez, J. (2004). Distribución espacio tiempo del fitoplancton del lago Zempoala, Morelos, México durante un ciclo anual (Tesis de maestría). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
García-Rodríguez, J. y Tavera, R. (1998). Fitoplancton del Lago Zempoala. Boletín de la Sociedad Botánica de México, 63, 85–100. https://doi.org/10.17129/botsci.1570
Garduño-Solórzano, G., Oliva-Martínez, M. G. y Ortega, M. (2009). Algas. En G. Ceballos, R. List, G. Garduño-Solórzano, R. López-Cano, M. J. Muñozcano-Quintanar, E. Collado et al. (Eds.), La diversidad biológica del Estado de México. Estudio de estado (pp. 153–161). Toluca de Lerdo: Gobierno del Estado de México.
Godínez-Ortega, J. L., Ortega, M. M., Garduño, G., Oliva, M. G. y Vilaclara, G. (2001). Traditional knowledge of Mexican continental algae. Journal of Ethnobiology, 21, 57–88.
Godínez-Ortega, J. L., Oliva-Martínez, M. G., Escobar-Oliva, M. A. y Mendoza-Garfias, B. (2017). Diversidad algal del Parque Nacional Lagunas de Zempoala, México, excepto diatomeas. Hidrobiológica, 27, 45–58. https://doi.org/10.24275/uam/izt/dcbs/hidro/2017v27n1/Godinez
GBIF: The Global Biodiversity Information Facility. (2024). What is GBIF? Recuperado el 10 de diciembre, 2024 de: https://www.gbif.org/what-is-gbif
González-Barrera, A. (1991). Contribución al estudio ficológico estacional de la laguna de Tlila y canales adyacentes, Xochimilco, México, D.F. (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
González-González, J. y Novelo, E. (1986). Algas. En A. Lot y F. Chiang (Eds.). Manual de herbario. Administración y manejo de colecciones, técnicas de recolección y preparación de ejemplares botánicos (pp. 47–54). Consejo Nacional de Flora de México, México D.F.
Guiry, M. D. y Guiry, G. M. (2025). AlgaeBase. World-wide electronic publication, University of Galway. Recuperado el 03 de marzo, 2025 de: https://www.algaebase.org
Hammer, Ø., Harper, D. A. T. y Ryan, P. D. (2001). PAST: Paleontological Statictics software package for education and data análisis. Paleontologia Electronica, 4, 9.
Hernández-Morales, R., Ortega-Murillo, M. R., Alvarado-Villanueva, R., Sánchez-Heredia, J. D. y Medrano-Zarco, F. (2008). Variación anual del fitoplancton en el lago cráter la Alberca de Tacámbaro, Michoacán, México. Biológicas, 10, 5–17.
Hernández-Morales, R., Ortega-Murillo, R., Sánchez, J. D., Alvarado-Villanueva, R. y Aguilera, M. S. (2011). Distribución estacional del fitoplancton en un lago cálido monomíctico en Michoacán, México. Biológicas, 13, 21–28.
Hollerbach, M. M. (1962). De vi taxonomica Polyedriopsis spinulosae Schmidle et genere novo Xanthophytorum pseudopolyedriopsis Hollerb. notula. Botanicheskij Mater- ialy, 15, 62–67.
Islam, A. K. M. N. e Irfanullah, H. M. (2001). Some new records of algae for Bangladesh: Cyanarcus, Chloremys, Myrmecia, Selenodictyum, Tetraplektron, and Pseudostaurastrum. Bangladesh. Journal of Plant Taxonomy, 8, 1–7.
Jaccard, P. (1901). Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bulletin of the New York Botanical Garden, 3, 1–46. https://doi.org/10.5169/SEALS-266450
Jiménez, A. G. (2010). Variaciones en la dominancia de grupos fitoplanctónicos en la capa de mezcla/zona trofogénica de un embalse tropical eutrófico con cambios significativos de nivel (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. México D.F.
John, D. M., Whitton, B. A. y Brook, A. J. (2002). The freshwater algal flora of the British Isles: an identification guide to freshwater and terrestrial algae. Cambridge University Press. U.K.
Krienitz, L., Hegewald, E., Reymond, O. L. y Peschke, T. (1993). Variability of LM, TEM and SEM characteristics of Pseudogoniochloris tripus gen. et comb. nov. (Xanthophyceae). Archiv für Hydrobiologie Supplement, 97, 67–82.
Krienitz, L., Hepperle, D., Stich, H. B. y Weiler, W. (2000). Nannochloropsis limnetica (Eustigmatophyceae), a new species of picoplankton from freshwater. Phycologia, 39, 219–227. https://doi.org/10.2216/i0031-8884-39-3-219.1
López-Adrián, S. J. y Barrientos-Medina, R. C. (2005). Diversidad y distribución del fitoplancton de agua dulce en la península de Yucatán, México. Ibugana.Boletín IBUG, 13, 3–13.
López-Adrián, S. J. y Catzim, C. L. A. (2010). Microalgas dulceacuícolas. En R. Durán García y M. E. Méndez González (Eds.), Biodiversidad y desarrollo humano en Yucatán (pp. 185–186). Mérida: CICY/ PPD-FMAM/ Conabio/ SEDUMA.
López-Adrián, S. J., Barrientos-Medina, R. C., Tavera, R. y Novelo, E. (2017). Ficoflora de la zona urbana y conurbada de Yucatán. En S. J. López-Adrián y E. Novelo-Maldonado (Eds.), Microalgas de la península de Yucatán (pp. 28–38). Mérida: Edición privada.
López-Segovia, E., Pérez-Díaz, J., Del Moral-Flores, L. F. y Hernández-Arellano, T. (2024). New records on the distribution of the Mexclapique, Girardinichthys viviparus (Bustamante, 1837) (Cyprinodontiformes, Goodeidae), an endangered species in Mexico. Neotropical Biology and Conservation, 19, 347–359. https://doi.org/10.3897/neotropical.19.e126767
Lot, A. (2007). Guía ilustrada de la Cantera Oriente. Caracterización ambiental e inventario biológico. México D.F.: Secretaría Ejecutiva de la Reserva Ecológica del Pedregal de San Ángel, Coordinación de la Investigación Científica, UNAM.
Macedonio, F. J. (2024). El lago de Coatetelco como patrimonio biocultural. En A. García-Flores, H. Colín-Bahena y J. M. Rivas-González (Coord.), Conocer para conservar: estudios de la diversidad biocultural en Coatetelco, Morelos (pp. 47–66). Cuernavaca: Universidad Autónoma del Estado de Morelos.
Magurran, A. E. (2004). Measuring biological diversity. London: Blackwell Publishing.
Martínez, M. M. (2016). Estructura de la microflora planctónica y perifítica en el lago de Cuitzeo (Tesis de maestría). Universidad Michoacana de San Nicolás de Hidalgo, Morelia. México.
Mendoza-González, A. C. (1985). Estudio florístico ficológico estacional de la laguna Victoria o de Santiago Tilapa, México. Phytologia, 58, 479–487.
Mendoza-González, A. C., Huerta-Múzquiz, L. y Flores-Granados, C. (1985). Estudio florístico del fitoplancton del lago Ziruahuén, Michoacán, México. Phytologia, 59, 1–8.
Mireles, V. A. (2019). Aspectos ecológicos de algas metafíticas en dos lagos de Tabasco, México(Tesis de maestría). Posgrado en Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. México D.F.
Montejano-Zurita, G., Carmona-Jiménez, J. y Cantoral-Uriza, E. A. (2000). Algal communities from calcareous springs in La Huasteca, central Mexico: a synthesis. En M. Munawar, S. G. Lawrence, I. F. Munawar y D. F. Malley (Eds.), Aquatic ecosystems of Mexico. Status and scope (pp. 135–149). Leiden: Backhuys Pub.
Montejano-Zurita, G., Cantoral-Uriza, E. A. y Carmona-Jiménez, J. (2004). Algas de ambientes lóticos en la cuenca baja del río Pánuco. En I. Luna, J. J. Morrone y D. Espinosa (Eds.), Biodiversidad de la Sierra Madre Oriental (pp. 111–126). Las Prensas de Ciencias, México D.F.: Universidad Nacional Autónoma de México.
Mora-Navarro, M. R. (2004). Fitoplancton del lago de Chapala, Jalisco-Michoacán, México (Tesis doctoral). Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara.
Mora-Navarro, M. d. R., Vázquez-García, J. A. y Vargas, Y. L. (2004). Ordenación de comunidades de fitoplancton en el lago de Chapala, Jalisco-Michoacán, México. Hidrobiológica (Iztapalapa), 14, 91–103.
Mora-Navarro, M. R., Vázquez-García, J. A., Guzmán-Arroyo, M., Mora-Navarro, M. A., Núñez-Márquez, I. G. y Reyes-González, M. E. (2006). Fitoplancton del lago de Chapala, Jalisco-Michoacán, México. En M. R. Mora-Navarro, J. A. Vázquez-García, Y. L. Vargas-Rodríguez y R. M. Hernández-Herrera (Eds.), Algas del occidente de México. Florística y ecología (pp. 8–29). Guadalajara: Universidad de Guadalajara (CUCBA)/ Fundación Gonzalo Río Arronte/ Sociedad Ficológica de México.
Moreno, C. R. y Palacios, D. C. E. (1987). Estudio ficoflorísticopreliminar en la presa de La Concepción, Tepotzotlán, Estado de México (Tesis). Facultad de Estudios Superiores-Iztacala, Universidad Nacional Autónoma de México. México.
Moreno-Ruiz, J. L. (1985). Contribución al estudio básico (análisis inicial de la diversidad alimenticia) de Ictiobus merinionalis Günter (Cypriniformes: Catastomidae), en algunas localidades de la cuenca baja del río Papaloapan (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Moreno-Ruiz, J. L. (2005). Fitoplancton. En J. Bueno, F. Álvarez y S. Santiago (Eds.), Biodiversidad del estado de Tabasco (pp. 33–63). México D.F.: Conabio/ Instituto de Biología, UNAM.
Moreno-Ruiz, J. L., Tapia-García, M., González-Macías, M. C. y Figueroa-Torres, M. G. (2008). Fitoplancton del río Tehuantepec, Oaxaca, México y algunas relaciones biogeográficas. Revista de Biología Tropical, 56, 27–54.
Novelo, E. (1998). Floras ficológicas del Valle de Tehuacán, Puebla(Tesis doctoral). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Novelo, E. (2014). Heterokontophyta Hoek. Flora del Valle de Tehuacán-Cuicatlán. Instituto de Biología, Universidad Nacional Autónoma de México. México D.F.
Novelo, E. y Tavera, R. (2025). BdLACET Base de datos de algas continentales. Facultad de Ciencias. Universidad Nacional Autónoma de México. México D.F. Consultado el 19 de octubre, 2024 en: https://bdlacet.mx/
Novelo, E., Tavera, R. y Lira, B. (2023). Un estudio a largo plazo de la flora algal en lagos urbanos. Revista Mexicana de Biodiversidad, 94, e945149. https://doi.org/10.22201/ib.20078706e.2023.94.5149
Novelo, E., Ponce, M. E. y Ramírez, R. (2009). Las microalgas de la Cantera Oriente. En A. Lot y Z. Cano-Santana (Eds.), Biodiversidad del ecosistema del Pedregal de San Ángel (pp. 71–79). Libro conmemorativo del 25 aniversario de la Reserva Ecológica de Ciudad Universitaria (1983-2008). México D.F: Universidad Nacional Autónoma de México.
Núñez-Márquez, I. G. y Reyes-González, M. E. (1995). Contribución al conocimiento del fitoplancton del lago de Chapala, Jalisco, México, durante el período de febrero a mayo de 1989, 90 y 91(Tesis). Universidad de Guadalajara, CUCBA, División de Ciencias Biológicas y Ambientales.
Ochoa, Z. G. G. (2023). Ficoflora de sistemas lóticos y lénticos de Nayarit(Tesis de maestría). Universidad Autónoma de Nayarit. Tepic.
Ortega, M. M. (1984). Catálogo de algas continentales recientes de México. México D.F: Universidad Nacional Autónoma de México.
Ortega-Murillo, M. R., Aguilar-Gómez, M. E., Alvarado-Villanueva, R., Hernández-Morales, R. y Díaz-Martínez, N. T. (2014). Riqueza específica de las microalgas de la subcuenca del río Cupatitzio, Michoacán, México. Boletín de la Sociedad Mexicana de Ficología, 4, 10–20.
Osorio, S. J. J. y López, P. R. (2005). Diversidad y distribución del fitoplancton de la laguna El Balsón, Tabasco, México (Tesis). División Académica de Ciencias Biológicas. Universidad Juárez Autónoma de Tabasco. Villahermosa.
Pascher, A., Schiller, J. y Migula, W. (1925). Heterokontae, Phaeophyta, Rhodophyta, Charophyta. Die Süsswasserflora Deutschlands, Österreichs und der Schweiz, Heft 11. Jena. https://doi.org/10.5962/bhl.title.21607
Pedraza, A. C. I. (2020). Relación de la comunidad fitoplanctónica y los factores abióticos del lago Cantemual en Tabasco (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. México D.F.
Pérez-Gutiérrez, R. M., Martínez-Flores, A., Vargas-Solís, R. y Jiménez, J. (2008). Two new antibacterial norabietane diterpenoids from cyanobacteria, Microcoleus lacustris. Journal of Natural Medicines, 62, 328–331. https://doi.org/10.1007/s11418-008-0238-z
Ponce, M. M. E., Ramírez, R. R. y Ramírez, V. M. (2019). Algas de la Cantera Oriente, Reserva Ecológica del Pedregal de San Ángel. Guía de campo y laboratorio. Las Prensas de Ciencias, Universidad Nacional Autónoma de México, México.
Quiroz-Castelán, H., Molina-Astudillo, F. I., García-Rodríguez, J. y Díaz-Vargas, M. (2007). Los lagos Zempoala y Tonatiahua del Parque Nacional Lagunas de Zempoala, Morelos. En G. De la Lanza-Espino y S. Hernández-Pulido (Eds.), Las aguas interiores de México: conceptos y casos (pp. 141–167). México D.F.: AGT Editor, S.A.
Ramírez, L. J. E. (2010). Caracterización del ecosistema de ribera y su valor indicador del estado ecológico en la subcuenca Valle de Bravo-Amanalco, Estado de México(Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. México D.F.
Real, R. y Vargas, J. M. (1996). The probabilistic basis of jaccard’s index of similarity. Systematic Biology, 45, 380–385. https://doi.org/10.1093/sysbio/45.3.380
Ríos, T. R. M. (2009). Composición química de las aguas del cauce Tula-Moctezuma-Claro, estado de Hidalgo (Tesis de maestría). Colegio de Postgraduados, Campus Montecillo. Texcoco, Estado de México.
Rodríguez, M. Z., Comin, C. H., Casanova, D., Bruno, O. M., Amancio, D. R., Costa, L. et al. (2019). Clustering algorithms: a comparative approach. Plos One, 14, e0210236. https://doi.org/10.1371/journal.pone.0210236
Rodríguez-Flores, R. (2014). Diversidad y distribución de algas macroscópicas en ríos de alta montaña de la cuenca de México(Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Rodríguez-Flores, R. y Carmona, J. J. (2018). Ecology and distribution of macroscopic algae communities in streams from the Basin of Mexico. Botanical Sciences, 97, 63–75. https://doi.org/10.17129/botsci.1237
Rybalka, N. (2015). Assessing phylogenetic delineations of taxa of filamentous Xanthophyceae (Stramenopiles) using DNA sequence analyses and morphology. (Ph.D. dissertation). Kiel University. Kiel.
Sámano-Bishop, A. (1934). Contribución al conocimiento de las algas verdes de los lagos del Valle de México. Anales del Instituto de Biología. Universidad Nacional Autónoma de México, 5, 149–173.
Sánchez-Gómez, S. P. y Lara-Villa, M. A. (1986). Análisis fitoplanctónico e hidrológico de un cuerpo de agua dulce tropical: laguna de Amela, Colima, México(Tesis). Escuela Nacional de Estudios Profesionales-Zaragoza. Universidad Nacional Autónoma de México. México D.F.
Schmitter-Soto, J. J., Comín, F. A., Escobar-Briones, E., Herrera-Silveira, J., Alcocer, J., Suárez-Morales, E. et al. (2002). Hydrogeochemical and biological characteristics of cenotes in the Yucatán Peninsula (SE Mexico). Hydrobiologia, 467, 215–228. https://doi.org/10.1023/A:1014923217206
Starmach, K. (1968). Flora slodkowodna Polski [Freshwater flora of Poland]. Volume 7. Chrysophyta III. Xanthophyceae Ròżnowiciowe. Varsovia: Państwowe Wydawn.
Takano, Y. y Horiguchi, T. (2005). Acquiring scanning electron microscopical, light microscopical and multiple gene sequence data from a single dinoflagellate cell. Journal of Phycology, 42, 251–256. https://doi.org/10.1111/j.1529-8817.2006.00177.x
Tavera, R. y Diez, B. (2009). Multifaceted approach for the analysis of the phototrophic microbial community in a freshwater recreational area of Xochimilco, México. Hydrobiologia, 636, 353–368. https://doi.org/10.1007/s10750-009-9965-8
Tavera, R., Novelo, E. y Orozco, C. (2018). Las diatomeas del Parque Ecológico de Xochimilco, México ¿Qué tan importante es la flora de un grupo de un pequeño lugar? Cymbella, 4, 3–46. https://cymbella.fciencias.unam.mx/articulos/V4/01/Las_diatomeas_del_Parque_Ecologico_de_Xochimilco_Mexico.html
Valadez-Cruz, F. (1998). Empleo de crecimientos algales para la caracterización de ambientes lóticos en la cuenca baja del río Amacuzac, Morelos (Tesis de maestría). Facultad de Ciencias, Universidad Nacional Autónoma de México.
Valadez-Cruz, F., Carmona, J. J. y Cantoral, U. E. (1996). Algas de ambientes lóticos en el Estado de Morelos, México. Anales del Instituto de Biología, Serie Botánica, 67, 227–282. https://repositorio.unam.mx/contenidos/28749
Valadez-Cruz, F., Rosiles-González, G. y Ortega-Rubio, A. (2015). Potenciales bioindicadores del elenco ficológico de la Reserva de la Biosfera Sian Ka’an. En A. Ortega-Rubio, A. M. J. Pinkus-Rendón e I. C. Espitia-Moreno (Eds.). Las áreas naturales protegidas y la investigación científica en México (pp. 285–314). La Paz: Centro de Investigaciones Biológicas del Noroeste/ Universidad Autónoma de Yucatán/ Universidad Michoacana de San Nicolás de Hidalgo.
Valdez, S. Z. (2012). Diagnóstico ambiental en las inmediaciones del embalse Requena, municipio de Tepeji del Río, Hidalgo(Tesis). Facultad de Estudios Superiores – Iztacala, Universidad Nacional Autónoma de México. Estado de México.
Vázquez, G. y Blanco-Pérez, R. (2011). Microalgas dulceacuícolas. En Comisión Nacional para el Conocimiento y uso de la Biodiversidad (Ed.), La biodiversidad en Veracruz. Estudio de estado (pp. 77–88). México D.F.: Comisión Nacional para el Conocimiento y uso de la Biodiversidad/ Gobierno del Estado de Veracruz/ Universidad Veracruzana/ Instituto de Ecología, A.C.
Yang, E. C., Boo, G. H., Kim, H. J., Cho, S. M., Boo, S. M., Andersen, R. A. et al. (2012). Supermatrix data highlight the phylogenetic relationships of photosynthetic Stramen- opiles. Protist, 163, 217–231. https://doi.org/10.1016/j.protist.2011.08.001
Zariñana-Leguízamo, M. L. 1997. Variación estacional de Ophiocytium parvulum (Perty) A. Braun 1855 en un sistema acuático inestable en el municipio de El Oro, Estado de México(Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Primer registro de Axianassa darrylfelderi (Crustacea: Decapoda: Axianassidae) en México y descripción de su primer estadio de zoea
Ignacio Cáceres a, b, José Salgado-Barragán c, Brenda Díaz-Cárdenas b, Eduardo Ríos-Jara a, Manuel Ayón-Parente a, *
a Universidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias, Departamento de Ecología, Laboratorio de Ecología Molecular, Microbiología y Taxonomía, Camino Ramón Padilla Sánchez No. 2100, 45110 Nextipac, Zapopan, Jalisco, Mexico
b Universidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias, Departamento de Biología Celular y Molecular, Laboratorio de Marcadores Moleculares en Biomedicina y Ecología, Camino Ramón Padilla Sánchez No. 2100, 45110 Nextipac, Zapopan, Jalisco, Mexico
c Universidad Nacional Autónoma de México, Unidad Académica Mazatlán, Instituto de Ciencias del Mar y Limnología, Laboratorio de Invertebrados Bentónicos, PO Box 811, 82000 Mazatlán, Sinaloa, Mexico
Received: 21 May 2024; accepted: 11 September 2025
Abstract
Axianassa darrylfelderi, previously known only from the Pacific coast of Colombia and Panama, is reported for the first time in Mexico and its first larval stage is described. Morphological characters of adults and the first zoea, in addition to a phylogenetic analysis were used to review the taxonomic status of the species. Our results show that A. darrylfelderi and A. australis are indeed related, but both species can be considered as different.
Keywords: Mud shrimps; Decapod; New records; Central Mexican Pacific
Resumen
Axianassa darrylfelderi, anteriormente conocida solo de la costa pacífica de Colombia y Panamá, se reporta por primera vez para México y también se describe su primer estadio larvario. Se utilizaron caracteres morfológicos de adultos y de la primera zoea, además de análisis filogenéticos, para revisar el estatus taxonómico de ésta. Los resultados muestran que A. darrylfelderi y A. australis efectivamente están relacionadas, pero ambas especies pueden considerarse diferentes.
Palabras clave: Camarones de lodo; Decápodo; Nuevo registro; Pacífico central mexicano
Introduction
As part of a research project on the inventory of decapod crustaceans from the bay of Chamela, Jalisco, Mexico, in the Mexican Pacific coast, 65 specimens of the mud shrimp Axianassa darrylfelderi Anker & Lazarus, 2015 were collected in mud bottoms of the Pérula Estuary. According to the World Register of Marine Species (WoRMS) consulted in February 2024, the genus Axianassa is composed of 15 species distributed in tropical and subtropical waters of the western Atlantic (A. intermedia Schmitt, 1924, A. arenaria Kensley & Heard, 1990, A. jamaicensis Kensley & Heard, 1990, A. australis Rodrigues & Shimizu, 1992, A. ferrazeae Blanco-Rambla & Kensley, 1998, and A. ovis Blanco-Rambla & Kensley, 1998), the eastern Pacific (A. mineri Boone, 1931, A. canalis Kensley & Heard, 1990, A. darrylfelderi Anker & Lazarus, 2015, A. christi Anker & Pachele, 2016, and A. linda Anker & Pachele, 2016), and the Indo-Pacific (A. ngochoae Anker, 2010, A. sinica Liu & Liu, 2010, A. planioculus Komai & Fujita, 2019, and A. microlepis Komai, Watanabe, Matsui & Tamego, 2020). Of the species recorded in the eastern Pacific, only A. mineri has been recorded in Mexico, Hernández-Aguilera (1998) reported 2 specimens from the intertidal of Isla Isabel and the Islas Marías.
Axianassa darrylfelderi was described from material collected in Bahía Málaga, Pacific coast of Colombia. The authors mentioned that it is very similar to A. australis from the coasts of Brazil and indicated that both species could be differentiated by the presence of several teeth on the lateral margin of the uropodal exopodite of A. darrylfelderi and absent in A. australis, the relative length of the antennal acicle, in relation to the 4th antennal segment is longer in A. darrylfelderi than in A. australis, and the armature of the cutting edge of the polex of the minor cheliped is armed with 3 strong (bicuspid) and numerous small teeth in A. darrylfelderi versus 1 proximal bi or tricuspid tooth and 2 simple distal teeth in A. australis. On the other hand, Anker and Pachelle (2016) reported A. darrylfelderi on the Pacific coast of Panama and observed that the species have more morphological variations. Due to the above, this species can be differentiated from A. australis solely by differences in the proportion of the relative length of the antennal acicle in relation to the 4th antennal peduncle (0.35 in A. darrylfelderi vs. 0.25 in A. australis) and other minor differences, e.g., 3rd to 5th abdominal pleurae with moderate pubescence in A. darrylfelderi vs. strongly pubescent in A. australis. Therefore, the authors considered A. darrylfelderi as a valid species, although with reservations (see the species identification key in Anker and Pachelle [2016]: 122) and suggested that a molecular analysis would be useful to confirm A. darrylfelderi as a distinct species.
It is known that the larval development of invertebrates provides important information about the biology of the species and their phylogenetic relationships; however, information on the larval stages of many invertebrates, including decapod crustaceans, is still scarce. Within the genus Axianassa, the larval development of A. australis is the only one known. Rodrigues and Shimizu (1992) described A. australis as a new species and included the description of its first zoeal stage (ZI). On the other hand, Ngoc-Ho (1981) included the complete larval development of Axianassa sp. (presumably A. australis) and Strasser and Felder (2005) presented the complete larval development of this same species, including a comparative analysis of its ZI, and those described in previous works.
Molecular analyses have become a useful tool to solve problems about the status of species (Mantelatto et al., 2006, 2009; Matzen da Silva et al., 2011) or to determine phylogenetic relationships among species or groups of species (Felder & Robles, 2009; Henmi et al., 2022; Morrison et al., 2002; Robles et al., 2009, 2020). Given the close morphological resemblance between A. darrylfelderi and A. australis and the uncertainty of the validity of A. darrylfelderi as a distinct species, the importance of combining morphological traits with molecular evidence to elucidate this problem is evident. The objective of this work is to present a new distribution record for A. darrylfelderi in the Pacific coast of Mexico, as well as to contribute information on the validity of the species through a molecular analysis and the description of its ZI.
Materials and methods
The Pérula Estuary is in the northwest portion of the town of Pérula, in Chamela Bay, Jalisco, Mexico (Fig. 1). The estuary is 2.5 km long with a width ranging between 16 and 65 m and an average depth of 0.8 m. The surrounding vegetation is classified as low deciduous forest type and the estuary is partially surrounded by mangroves (Avicennia germinans, Conocarpus erectus, Laguncularia racemosa, and Rhizophora mangle), those species are within the category of non-endemic threatened species in the Mexican Official Norm (NOM-059-SEMARNAT-2010).
The organisms were collected using a “yabby” suction pump in muddy substrates, at depths of 0-0.5 m. Once collected, specimens of A. darrylfelderi were kept alive for several hours for observation. One of the ovigerous females collected in April 2018 was separated and kept until it released the zoea larvae. Unfortunately, the larvae only survived for a few hours and were collected and fixed in 70% ethanol. Several larvae were selected for dissection and description of the larval stage. Carapace length (CL) was measured along the dorsal midline from the tip of the rostral projection to the posterior margin of the carapace and total length (TL) was measured from the tip of the rostrum to the posterior midpoint of the telson. All measurements are in millimeters. In the description of larval appendages, the arrangement of the setae was listed sequentially from the proximal to the distal ones, as suggested by Konishi (1989), Nates et al. (1997), and Strasser and Felder (2005). In the listings of the groups of setae, those on the different segments were separated by a comma and those on the same segment or on different lobes of the same segment, article or endite, were separated with a plus sign. Roman numerals were used to describe the pattern of processes on the posterior margin of the telson. The dissection of the larvae was performed with the aid of a Leica M125 stereoscopic microscope. Ten larvae were dissected for their description and both appendages (right and left) were used for counting the setae. The illustrations were made using a Leica DM1000 microscope equipped with a camera lucida. Adults, juveniles, and larvae of Axianassa were preserved in 70% ethanol and deposited, with an accession number, in the Crustacean Collection of the Laboratory of Marine Ecosystems and Aquaculture (LEMA-CR) of the University Center for Biological and Agricultural Sciences (CUCBA) in the University of Guadalajara (U de G).
Figure 1. Geographical location of the study area.
Phylogenetic analysis, genetic distances and haplotype networks
Total genomic DNA of an individual of A. darrylfelderi was extracted from muscle tissue of the walking legs. Partial DNA sequences were obtained from 2 genes, 16S and COI. DNA was extracted using the Quick-DNA TM Tissue/Insect Microprep kit, Catalog No. D6015 of ZYMO RESEARCH. The 16S region was amplified with the primer “CCT GTT TAN CAA AA CAT and AGA AAA CCA ACC TGG” (Crandal & Fitzpatrick Jr., 1996), and the following thermal cycler conditions: 5 min at 95 °C; annealing for 35 cycles: 1 min at 95 °C, 1 min at 54 °C, 1 min at 72 °C; final extension 10 min at 72 °C. For the COI region we used the primer “TAA ACT TCA GGG TGA CCA AAA AAT CA and GGT CCA CAA ATC ATA AAG ATA TTG G” (Folmer et al., 1994), and the following thermal cycler program: 5 min at 95 °C; annealing for 30 cycles: 1 min at 95 °C, 1 min at 40 °C, 1 min at 72 °C; final extension 10 min at 72 °C. The PCR took place in a final volume of 25 μL consisting of 12.1 μL of H2O, 2 μL of reaction buffer, 0.2 μL of dNTPs, 0.08 μL of each primer, 1.5 μL of MgCl2 0.1 μL of Taq Platinum (Invitrogen), and 20 ng of DNA. Amplification of the regions was confirmed by agarose gel electrophoresis at 1%. The amplified products were sent to the National Laboratory for Plant Identification and Characterization (LANIVEG) at CUCBA, UdeG, for sequencing. The generated sequences were deposited in GenBank under the accession numbers PV52069 for the COI region and PV521869 for 16S region.
The COI and 16S sequences obtained were edited with Sequencher v 4.1.4 (Gene Codes Corp., Ann Arbor, Michigan). The BLAST tool from Genbank (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to determine the percentage of similarity between A. darrylfelderi and A. australis. To test the divergence between the collected individual and individuals of A. australis, we analyzed the genetic distances under the Tamura-Nei 93 (TN93) model in the ape package (Paradis & Schliep, 2019) in R (R Core Team, 2021). To perform a phylogenetic analysis, we obtained the following sequences of A. australis from GenBank: MF490135, OQ108484 and OQ108485 for the COI region and MF490232, EU874948, OQ110581 and OQ110582 for the 16S region. Following Teles & Mantelatto (2023) we used Upogebia brasiliensis (COI: MF490136, 16S: MF490236), U. noronhensis (COI: MF490138, 16S: MF490234) and U. paraffinis (COI: MF490137, 16S: MF490235) to root the tree. Regarding the 16S tree, we also add A. planioculus (LC489001) as outgroup. The sequences were visually aligned in the PhyDe v 0.9971 program (Müller et al., 2010).
A phylogenetic analysis using maximum likelihood was performed in RAxML GUI 2.0 (Edler et al., 2020) for each region with the HKY + I + G evolutionary model for the COI region and the HKY + G model for the 16S region, both models were inferred in RAxML based on the Bayesian information criterion. To assess nodal support a resampling of 10,000 rapid bootstrap inferences was done. To corroborate the distance between A. australis and A. darrylfelderi sequences,we generated a haplotype network in the R package ape (Paradis & Schliep, 2019; R Core Team, 2021).
The discovery of A. darrylfelderi on the coast of Jalisco, represents the third record of the species in the Eastern Tropical Pacific. The 2 previous records correspond to the type locality, Bahía Málaga, Colombia, and Playa El Agallito, Panama (Anker & Lazarus, 2015; Anker & Pachele, 2016). The specimens collected in the Pérula Estuary represent the first record of the species for Mexico and thus extends its geographical distribution about 3,000 km to the north.
According to Anker and Pachele (2016), specimens of A. darrylfelderi from Colombia and Panama have morphological variations between individuals from the same populations and between males and females. We were also able to observe morphological variations among the specimens from the Pérula Estuary. The most evident variations were: the number of teeth on the ischium of the third maxillipeds varied from 13 to 18, with an average of 14; the number of teeth on the ischium of the largest cheliped varied from 1 to 6 in males and from 3 to 5 in females, while the number of teeth on the ischium of the minor cheliped ranged from 3 to 5 in males and from 3 to 4 in females. Uropods in all the specimens always carried a spiniform seta and from 0 to 5 fixed teeth on the external margin. Most of the individuals presented a rounded and unarmed rostrum (Fig. 2A, C), and only 2 specimens had a rostrum ending in a spine (Fig. 2B). The majority of the specimens had both antennal acicles armed with a mesial tooth (Fig. 2A) but 3 specimens had just 1 armed acicle (Fig. 2B) and 1 specimen had unarmed antennal acicles (Fig. 2C). The dorsomesial surface of the coxa of the third maxilliped bears only 1 large tooth in most of the specimens and only 2 specimens had 2 strong adjacent teeth.
Description of zoea I
(Fig. 3A-K)
Size. CL = 0.63 ± 0.038 mm, range 0.57-0.67 mm; TL = 1.56 ± 0.075 mm, range 1.48-1.63 mm. Carapace (Fig. 3A) about the same length as abdomen; rostrum elongate, rounded in cross-section, smooth; eyes fused to carapace. Abdomen (Fig. 3A) somite 5 with posterolateral projections about length of somite; lacking dorsal spines. Pleopods not developed.
Telson (Fig. 3A) narrow, anteriorly not differentiated from abdominal segment 6, triangular posterior with medial cleft, anal spine absent; 7 + 7 processes on posterior margin arranged as (I. II, III, II, I) where I is outermost naked seta, II is very thin (anomuran hair), III is 5 plumodenticulate setae.
Antennule (Fig. 3B) elongate, exopodal and endopodal lobes not distinct; 5 terminal aesthetascs and 1 seta; 1 long plumose seta on future endopodal lobe. Antenna (Fig. 3C) protopod with 1 distal spine between rami; endopod with 3 long, plumose setae; scaphocerite (exopod) armed with 1 strong distolateral spine, 10 plumose setae on inner margin. Mandibles (Fig. 3D, E) asymmetrical; right mandible with 2 prominent teeth at base, lower plate with numerous small teeth, distal end pointed but not as strongly produced as on left mandible; left mandible sickle-shaped, distal end pointed with 2 teeth, inner surface of base with several teeth. Left lobe of paragnath (Fig. 3F) sickle-shaped. Maxillule (Fig. 3G) coxal endite with 4 marginal setae; basal endite with 2 setae and 2 large dentate spines; endopodal lobe disctinct from protopod, with 3 setea on distal margin; propotopod without setae. Maxilla (Fig. 3H) coxal endite bilobed, 1 seta on proximal lobe, and 3-4 setae on distal lobe; basal endite bilobed with 4 setae on proximal lobe, 3-5 setae on distal lobe; endopod small, unsegmented with 2 terminal setae; scaphognathite with 5 plumose setae. Maxilliped 1 (Fig. 3I) coxa without setae; basis with 1 + 2 + 1 setae; endopod 4-segmented, with 1, 1, 2, 5 setae; exopod unsegmented, 4 plumose setae on distal margin. Maxilliped 2 (Fig. 3J) coxa and basis without setae; endopod 4-segmented, 1, 1, 2, 5 setae, exopod unsegmented, 4 plumose setae on distal margin. Maxilliped 3 (Fig. 3K) 2-segmented, without setae.
Figure 2. Axianassa darryfelderi from Pérula Estuary, Jalisco. A, Frontal region in dorsal view of a male (CL = 10.2 mm); B, rostrum, eyestalks and antennal acicles in dorsal view of a female (CL = 9.6 mm); C, rostrum, eyestalks and antennal acicles in dorsal view of a male (CL = 11.4 mm). Scale = 2 mm.
Table 1
Percentage identity (similarity) and genetic distances between Axianassa darrylfelderi and A. australis according to COI and 16S mitochondrial regions.
Percentage identity
Genetic distance
COI
89
0.124
16S
97
0.047
Figure 3. Axianassadarrylfelderi, zoea I. A, Dorsal view; B, antennule; C, antenna; D, right mandible; E, left mandible; F, left lobe of paragnath; G, maxillule; H, maxilla; I, maxilliped I; J, maxilliped 2; K, maxilliped 3. Scale bars = O.1 mm.
Pereopods. Not developed.
Sequences of 644 bp and 492 bp were obtained for COI and 16S, respectively. BLAST analyses showed 89% similarity to the A. australis COI region and 97% similarity to the A.australis 16S region (Table 1). According to the genetic distances, A. darrylfelderi was 12.4% (COI) and 4.7% different from A. australis (Table 1). The ML phylogenetic hypothesis of the COI region, groups A. darrylfelderi and A. australis with low support (49) and the 16S hypothesis indicates that A. darrylfelderi and A. australis are sister groups witha boostrap support of 96 (Fig. 4). The haplotype network showed a distance of 44 mutational steps in the COI network and 12 mutational steps in the 16S network between A. darrylfelderi and A. australis (Fig. 5). The genetic analyses presented in this study are a partial contribution to establishing that, based on the specimens collected in the Pérula Estuary, the genetic sequences of A. darrylfelderi are close, but different to those of A. australis compiled from GenBank, which would allow us to establish that both species can be considered as valid.
Discussion
The use of complementary tools for taxonomic analysis has allowed us to clarify the phylogenetic status of several species of decapod crustaceans. The great morphological similarity between A. darrylfelderi and A. australis led Anker and Pachelle (2016) to question whether the former is a valid species or to consider if it is a synonym of the latter. We believe that the application of molecular analyses (partial 16S rDNA and COI sequences) and a morphological review of adults and the Z1 allowed us to solve this problem, and to consider A. darrylfelderi as a valid species.
Anker and Pachelle (2016) modified the criteria used by Anker and Lazarus (2015) to differentiate specimens of A. darrylfelderi and A. australis (presence or absence of teeth on the distolateral margin of the uropodal exopodite, the proportional length of the antennal acicle and the armature of the cutting edges of the pollex in the male´s minor cheliped) because they observed that the specimens of A. darrylfelderi collected in Panama showed a wide morphological variation that overlapped with the variations observed in A. australis, so they suggested that only the relative length of the antennal acicle could be considered as a valid difference between both species (0.35 of the length of the 4th article of the antennal peduncle in A. darrylfelderi vs. 0.25 of the length of the 4th article of the antennal peduncle in A. australis). Furthermore, they added that abdominal pleurae 3-5 of A. australis are densely pubescent, obscuring their margin, while those of A. darrylfelderi are moderately pubescent and that the dorsomesial surface of the coxa of the third maxilliped of A. darrylfelteri has 2 small teeth (occasionally 1) vs. 1 tooth in A. australis.
Figure 4. Phylogenetic hypotheses of Axianassa darrylfelderi obtained by the maximum likelihood method with the COI (A) and 16S (B) regions. Node numbers indicate bootstrap support values above 70. GenBank accession numbers are shown in the tips of the tree. Figure 5. Haplotype networks of Axianassa darrylfelderi and A. australis. The lines between haplotypes represent mutational steps. The numbers indicate GenBank accession numbers.
The specimens collected in Pérula Estuary could be identified as A. darrylfelderi because their antennal acicles are 0.35 of the length of the 4th article of the antennal peduncle and the abdominal pleurae 3-5 are scarcely pubescent. However, unlike what was established by Anker and Pachelle (2016), the majority of our specimens had only 1 tooth on the dorsomesial surface of the coxa of the third maxilliped, and only 2 specimens had 2 teeth; so, we consider that this characteristic cannot be used to separate the 2 species. In addition, we observed in A. darrylfelderi other morphological variations, for example, most of our specimens have an unarmed rostrum (as mentioned in the original description of the species) but 2 of them have a terminal small spine or papilla and other specimens have antennal acicles without a ventromesial spine. This is important to highlight because those characters are used to distinguish other species within the genus Axianassa. Kensley and Heard (1990) pointed out that A. arenaria has a narrow to rounded rostrum ending in a small papilla (Fig. 5B, p. 565) similar to that observed in our specimens. On the other hand, A. arenaria, A. christyi, A. ngochoae, and A. sinica have an unarmed antennal acicle as pointed out by Anker and Pachelle (2016).
The morphology of the Z1 of A. darrylfelderi from the the Pérula Estuary has a close resemblance to the ZI of A. australis analyzed by Ngoc-Ho (1981), Rodrigues and Shimizu (1992) and Strasser and Felder (2005) (Table 2). Perhaps the greatest differences can be observed with those described by Rodrigues and Shimizu (1992), however, this work should be considered with caution since there are some inconsistencies in its illustrations. For example, figures 25 and 26 correspond to the left mandible and left lobe paragnath, respectively, and not to right and left mandibles as they mentioned, so it is possible that the larvae observed by Rodrigues and Shimizu (1992) were not so different from those described in the other works, and they did not illustrate other structures (e.g., subterminal plumose seta on the antennula or a spine on the antennal peduncle), thus we could conclude that the Z1 of species of Axianassa are relatively similar and the differentiation among species can be made only in advanced stages, as seen in other mud shrimp (e.g., De Oliveira et al., 2014; Miyabe et al., 1998; Pohle et al., 2011; Strasser & Felder, 1999) or even in other decapods as hermit or panopeid crabs (Salgado-Barragán & Ruiz-Guerero, 2005; Siddiqui et al., 1993). Concerning the differences in the ZI shown in table 2, the only reliable character that allowed us to clearly distinguish the ZI of A. darrylfelderi from that of A. australis is the presence of first pereopods as small bilobed buds in A. australis that are absent in the ZI of A. darrylfelderi. Miyabe et al. (1998) pointed out that the Zl of Callianassa japonica (= Neotrypaea japonica) and C. petalura (= N. petalura) could be differentiated solely by the presence (in the former) or the absence (in the latter) of a coxal seta present in the second maxilliped. We observed that the morphological characters used to separate A. darrylfelderi and A. australis are very variable and significantly hinder the differentiation between these species.
COI and 16S regions have been widely used to delimit species in crustaceans (Costa et al., 2007; Lefébure et al., 2006; Magalhães et al., 2016). According to our results, the divergence between A. darrylfelderi and A. australis is above the lower interspecific distances reported in the literature (Costa et al., 2007; Lefébure et al., 2006; Magalhães et al., 2016), which supports our hypothesis. Phylogenetic analysis showed confusing results. While phylogenetic inference with 16S region recovers both species as sister groups, the COI hypothesis groups both species in the same clade but with a low support. This could be due either to the lack of genetic information regarding Axianassa genus or to a close relationship between A. darrylfelderi and A. australis. Nonetheless, the haplotype networks revealed a great genetic distance between both species, supporting our hypothesis.
Table 2
Comparison between larva zoea I of A. australis and A. darrylfelderi. Rodrigues and Shimizu, 1992 (1); Ngoc-Ho, 1981 (2); Strasser and Felder, 2005 (3).
Axianassa australis (1)
Axianassa australis (2)
Axianassa australis (3)
Axianassa darrylfelderi (Present study)
Rostrum
As long as antennae
As long as antennae
As long as antennae
As long as antennae
Eyes
Round, sessile
Round, sessile
Round, sessile
Round, sessile
Abdomen
Somite 5 with lateral spines Somite 6 fused with telson,
Somite 5 with lateral spines Somite 6 fused with telson,
Somite 5 with lateral spines Somite 6 fused with telson
Somite 5 with lateral spines Somite 6 fused with telson
2-segmented, 4 apical setae 4-segmented, 0,0,2,4 setae No setae
Unsegmented, 4 apical setae 4-segmented, 0-1,0-1,0-2,4-5 setae No setae
Unsegmented, 4 apical setae 4-segmented. 1,1,2,5 setae No setae
Maxilliped 3: Exopod Endopod Pereopods
2-segmented, no setae Absent Absent
2-segmented, no setae Absent Pereopods 1 and 2 present as small buds
2-segmented, no setae Absent Pereopod 1 as small bilobed bud
2-segmented, no setae Absent Absent
Telson processes Medial spine Anomuran hair
7 +7 Absent Present
7 +7 Absent Present
7 + 7 Absent Present
7 + 7 Absent Present
Acknowledgments
Special thanks to the biology students from Universidad de Guadalajara who supported the sampling. Thanks also to the field station of the Instituto de Biología (Universidad Nacional Autónoma de México) that offered important support in its facilities in Chamela. We are grateful to two anonymous reviewers whose comments improved the manuscript significantly.
References
Anker, A. (2010). The mud-shrimp genus Axianassa Schmitt, 1924 (Crustacea, Decapoda, Axianassidae) in the Indo-West Pacific, with description of a new species from French Polynesia. Zootaxa, 2557, 49–59. https://doi.org/10.11646/zootaxa.2557.1.5
Anker, A., & Lazarus, J. F. (2015). Description of two new associated infaunal decapod crustaceans (Axianassidae and Alpheidae) from the tropical eastern Pacific. Papéis Avulsos de Zoologia, 55, 115–129. https://doi.org/10.1590/0031-1049.2015.55.08
Anker, A., & Pachelle, P. P. G. (2016). Mud-shrimps of the genus Axianassa Schmitt, 1924 from Panama, with description of two new species (Decapoda: Gebiidea: Laomediidae). Zootaxa, 4111, 101–125. https://doi.org/10.11646/zootaxa.4111.2.1
Blanco-Rambla, J. P., & Kensley, B. (1998). Dos nuevas especies de camarones laomedidos del género Axianassa Schmitt, 1924 (Crustacea: Decapoda: Thalassinidea) para el Caribe sur. In XLVIII Convención Anual AsoVAC, Maracaibo, 9-13 de noviembre de 1998. Acta Científica Venezolana, 49, 349.
Boone, L. (1931). Anomuran, macruran Crustacea from the Bay of Panama and the freshwaters of the Canal zone. Bulletin of the American Museum of Natural History, 61, 137–189.
Costa, F. O., deWaard, J. R., Boutillier, J., Ratnasingham, S., Dooh, R. T., Hajibabaei, M. et al. (2007). Biological identifications through DNA barcodes: the case of the Crustacea. Canadian Journal of Fisheries and Aquatic Sciences, 64, 272–295. https://doi.org/10.1139/F07-008
Crandal, K. A., & Fitspatrick Jr., J. F. (1996). Crayfish molecular systematics: Using a combination of procedures to estimate phylogeny. Systematic Biology, 45, 1–26. https://doi.org/10.1093/sysbio/45.1.1
De Oliveira, D. B., Martinelle-Lemos, J. M., & Araújo-Abrunhosa, F. (2014). The complete larval development of the mud shrimp Upogebia vasquezi (Gebiidea: Upogebiidae) reared in the laboratory. Zootaxa, 3826, 517–543. https://doi.org/10.11646/zootaxa.3826.3.5
Edler, D., Klein, J., Antonelli, A., & Silvestro, D. (2020). raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods in Ecology and Evolution, 12, 373–377. https://doi.org/10.1111/2041-210X.13512
Felder, D. L., & Robles, R. (2009). Molecular phylogeny of the family Callianassidae based on preliminary analyses of two mitochondrial genes. In J. W. Martin, K. A. Crandall, & D. L. Felder (Eds.), Decapod crustacean phylogenetics. Crustacean Issues. Koenemann, S. (Series ed.) Vol. 18 (pp. 327–342). Boca Raton, London, New York: CRC Press, Taylor & Francis Group.
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
Henmi, Y., Itani, G., Osawa, M., & Komai, T. (2022). A new species of the ghost shrimp genus Callianassa Leach, 1814 (Decapoda: Axiidea: Callianassidae) from Wakasa Bay, western Japan: the first representative of the genus from the Pacific region. Zootaxa, 5182, 465–478. https://doi.org/10.11646/zootaxa.5182.5.4
Hernández-Aguilera, J. L. (1998) Sobre una colección de talasínidos (Crustacea: Decapoda) de la costa del Pacífico de México, con la descripción de una especie nueva del género Biffarius. Ciencias Marinas, 24, 303–312. http://dx.doi.org/10.7773/cm.v24i3.754
Kensley, B., & Heard, R. (1990). The genus Axianassa (Crustacea: Decapoda: Thalassinidea) in the Americas. Proceedings of the Biological Society of Washington, 103, 558–572.
Komai, T., & Fujita, Y. (2019). A new species of the mud shrimp genus Axianassa Schmitt, 1924 (Decapoda: Gebiidea: Laomediidae) from the Ryukyu Islands, southern Japan. Zootaxa, 4658, 452–460. https://doi.org/10.11646/zootaxa.4658.3.2
Komai, T., Watanabe, T., Matsui, S., & Tamego, T. (2020). A new species of the mud shrimp genus Axianassa Schmitt, 1924 (Decapoda: Gebiidea: Laomeiidae) from Japon. Zootaxa, 4852, 461–474. https://doi.org/10.11646/zootaxa.4852.4.4
Konishi, K. (1989). Larval development of the mud shrimp Upogebia (Upogebia) major (De Hann) (Crustacea: Thalassinidea: Upogebiidae) under laboratory conditions, with comments on larval characters of thalassinid families. Bulletin of the National Research Institute of Aquaculture, 15, 1–17.
Lefébure, T., Douady, C. J., Gouy, M., & Gilbert, J. (2006). Relationship between morphological taxonomy and molecular divergence within Crustacea: proposal of a molecular threshold to help species delimitation. Molecular Phylogenetics and Evolution, 40, 435–447. https://doi.org/10.1016/j.ympev.2006.03.014
Liu, W., & Liu, R. (2010). First record of the gebiidean genus Axianassa Schmitt, 1924 (Crustacea: Decapoda: Gebiidea: Axianassidae) in the West Pacific, with description of a new species from the South China Sea. Raffles Bulletin of Zoology, 58, 193–197. http://dx.doi.org/10.5281/zenodo.5342684
Magalhães, T., Robles, R., Felder, D. L., & Mantelatto, F. L. (2016). Integrative taxonomic study of the purse crab genus Persephona Leach, 1817 (Brachyura: Leucosiidae): combining morphology and molecular data. Plos One, 11, e0152627. https://doi.org/10.1371/journal.pone.0152627
Mantelatto, F. L., Robles, R., Biagi, R., & Felder, D. L. (2006). Molecular analysis of the taxonomic and distributional status for the hermit crab genera Loxopagurus Forest, 1964, and Isocheles Stimpson, 1858 (Decapoda, Anomura, Diogenidae). Zoosystema, 28, 495–506.
Mantelatto F. L., Robles, R., Schubart, C. D., & Felder, D. L. (2009). Molecular phylogeny of the genus Cronius Stimpson, 1860, with reassignment of C. tumidulus and several American species of Portunus to the genus Achelous De Haan, 1833 (Brachyura: Portunidae). In J. W. Martin, K. A. Crandall, & D. L. Felder (Eds.), Crustacean issues: Decapod Crustacean Phylogenetics (pp. 567–579). Boca Raton: Taylor and Francis/ CRC Press.
Matzen da Silva, J., Creer, S., Dos Santos, A., Costa, A. C., Cunha, M. R., Costa, F. O. et al. (2011). Systematic and evolutionary insights derived from mtDNA COI barcode diversity in the decapoda (Crustacea: Malacostraca). Plos One, 6, e19449. http://dx.doi.org/10.1371/journal.pone.0019449
Miyabe, S., Konishi, K., Fukuda, Y., & Tamaki, A. (1998). The complete larval development of the ghost shrimp, Callianassa japonica Ortmann, 1891 (Decapoda: Thalassinidea: Callianassidae), reared in the laboratory. Crustacean Research, 27, 101–121. https://doi.org/10.18353/crustacea.27.0_101
Morrison, C. L., Harvey, A. W., Lavery, S., Tieu, K., Huang, Y., & Cunningham, C. W. (2002). Mitochondrial gene rearrangements confirm the parallel evolution of the crab-like form. Proceedings of the Royal Society of London,Series B, 269, 345–350. https://doi.org/10.1098/rspb.2001.1886
Müller, J., Müller, K., Neinhuis, C., & Quandt, D. (2010). PhyDE-Phylogenetic Data Editor, versión 0.9971. Program distributed by the autor: http://www.phyde.de
Nates, S. F., Felder, D. L., & Lemaitre, R. (1997). Comparative larval development in two species of the burrowing ghost shrimp genus Lepidophthalmus (Decapoda: Callianassidae). Journal of Crustacean Biology, 17, 497–519. https://doi.org/10.2307/1549444
Ngoc-Ho, N. (1981). A taxonomic study of the larvae of four thalassinid species (Decapoda, Thalassinidea) from the Gulf of Mexico. Bulletin of the British Museum (Natural History) Zoology, 40, 237–273.
Paradis, E., & Schliep, K. (2019). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics, 35, 526–528. https://doi.org/10.1093/bioinformatics/bty633
Pohle, G., Santana, W., Jansen, G., & Greenlaw, M. (2011). Plankton-caught zoeal stages and megalopa of the lobster shrimp Axius serratus (Decapoda: Axiidae) from the Bay of Fundy, Canada, with a summary of axiidean and gebiidean literature on larval descriptions. Journal of Crustacean Biology, 31, 82–99. http://dx.doi.org/10.1651/10-3321.1
R Core Team. (2021). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Robles, R., Dworschak, P. C., Felder, D. L., Poore, G. C. B., & Mantelatto, F. L. (2020). A molecular phylogeny of Callianassidae and related families (Crustacea: Decapoda: Axiidea) with morphological support. Invertebrate Systematics, 34, 113–132. https://doi.org.10.1071/IS9021
Robles, R., Tudge, C. C., Dworschak, P. C., Poore, G. C. B., & Felder, D. L. (2009). Molecular phylogeny of the Thalassinidea based on nuclear and mitochondrial genes. In J. W. Martin, K. A. Crandall, & D. L. Felder (Eds.), Crustacean Issues 18: Crustacean issues: Decapod Crustacean Phylogenetics (pp. 309–326). Boca Raton: Taylor and Francis/ CRC Press.
Rodrigues, S. A., & Shimizu, R. M. (1992). Description of a new Axianassa (Crustacea: Decapoda: Thalassinidea) from Brazil, and its first larval stage. Biological Society of Washington, 105, 317–323.
Salgado-Barragán, J., & Ruiz-Guerrero, M. (2005). Larval development of the eastern Pacific mud crab Acantholobulus mirafloresensis (Abele and Kim, 1989) (Decapoda: Brachyura: Panopeidae) described from laboratory-reared material. Invertebrate Reproduction & Development, 47, 133–145. https://doi.org/10.1080/07924259.2005.9652154
Schmitt, W. L. (1924). The macruran, anomuran and stomato- pod Crustacea. Bijdragen tot der kennis der fauna van Curaçao. Resultaden eener reis von Dr. C. J. van der Horst in 1920. Bijdragen tot de Dierkunde, 23, 9–82. https://doi.org/10.1163/26660644-02301005
Semarnat (Secretaría del Medio Ambiente y Recursos Naturales). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental – Especies nativas de México de flora y fauna silvestres – Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio – Lista de especies en riesgo. Diario Oficial de la Federación. 30 de diciembre de 2010, Segunda Sección, México.
Siddiqui, E. A., McLaughlin, P. A., & Crain, J. A. (1993). Larval development of the hermit crab Clibanarius albidigitus (Crustacea: Anomura: Diogenidae) reared under laboratory conditions. Marine Biology, 116, 603–613. https://doi.org/10.1007/BF00355479
Strasser, K. M., & Felder, D. L. (1999). Larval development in two populations of the ghost shrimp Callichirus major (Decapoda: Thalassinidea) under laboratory conditions. Journal of Crustacean Biology, 19, 844–878. http://www.jstor.org/stable/1549168
Strasser, M. K., & Felder, L. D. (2005). Larval development of the mud shrimp Axianassa australis (Decapoda: Thalassinidea) under laboratory conditions. Journal of Natural History, 39, 2289–2306. https://doi.org/10.1080/00222930500061320
Teles, J. N., & Mantelatto, F. L. (2023). New records of the shrimp-like decapods Alpheus angulosus, Alpheus buckupi, and Axianassa australis from Maranhão, Brazil. Nauplius, 31, e2023022. https://doi.org/10.1590/2358-2936e2023022
WoRMS Editorial Board. (2024). World register of marine species. https://doi.org/10.14284/170