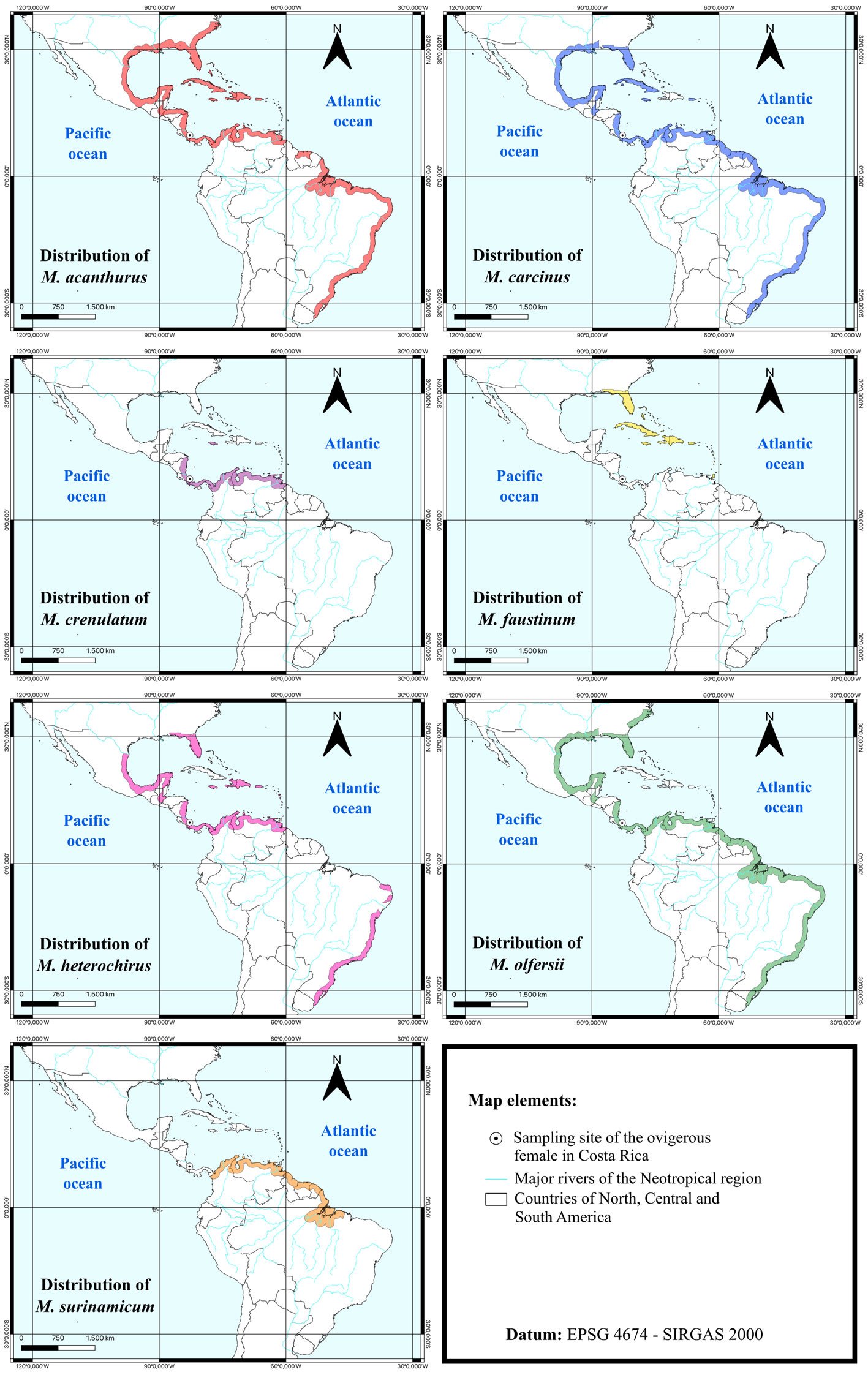
Redescripción de Stagmomantis hebardi (Mantodea: Mantidae), con la descripción de la hembra
Aarón Emilio Vásquez-Quintero a, Iker Cubillos-Macías b, Erick Omar Martínez-Luque c, Jovana M. Jasso-Martínez b y Fernando Varela-Hernández a, *
a Universidad Autónoma del Estado de Morelos, Escuela de Estudios Superiores del Jicarero, Laboratorio de Sistemática Molecular, Carretera Galeana-Tequesquitengo s/n, Col. El Jicarero, 62909 Jojutla, Morelos, Mexico b Universidad Nacional Autónoma de México, Instituto de Biología, Colección Nacional de Insectos, Tercer Circuito Exterior s/n, Cd. Universitaria, Copilco El Alto, Coyoacán, Apartado postal 70-233, 04510 Mexico City, Mexico c Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales, Av. de las Ciencias s/n, Juriquilla, 76230 Querétaro, Querétaro, Mexico
*Corresponding author: fernando.varela@uaem.mx (F. Varela-Hernández)
Received: 02 September 2025; accepted: 08 December 2025
Abstract
The genus Stagmomantis Saussure, 1869 comprises a group of New World mantids whose taxonomic delimitation and composition have been challenging due to complex morphological variation. Stagmomantis hebardi Rehn, 1935, a species distributed in the Neotropical region and the Mexican Transition Zone, was originally described based on both a female and a male specimen. However, the female was subsequently recognized as belonging to a different species within the subgenus Auromantis. In this study, we provide the redescription of the male and the description of the female of S. hebardi. Descriptions are based on external morphology and both sexes internal genitalia characters. The conspecificity of the male and female is supported by molecular evidence provided in previous studies and further validated by biological observations from captive rearing. The corkscrew-like pda (posterior process of the ventral phallomere) in the male genitalia and the lobe-like structure between the fifth and sixth tergites in the female abdomen are diagnostic characters that distinguish this species from others of the genus Stagmomantis.
El género Stagmomantis Saussure, 1869 es un grupo de mantis del Nuevo Mundo cuya delimitación y composición han sido desafiantes debido a su compleja variación morfológica. Stagmomantis hebardi Rehn, 1935, una especie distribuida en la región Neotropical y la Zona de Transición Mexicana, fue descrita originalmente con base en un macho y una hembra. Más tarde, se reconoció que la hembra pertenecía a una especie diferente del subgénero Auromantis. En este estudio, proporcionamos la redescripción del macho y la descripción de la hembra de S. hebardi. Ambas descripciones se basan en caracteres de morfología externa y de la genitalia de ambos sexos. La asociación específica entre el macho y la hembra está respaldada por análisis moleculares previos y validada por observación directa en cautiverio. El pda (proceso posterior del falómero ventral) en forma de sacacorchos en la genitalia del macho y la estructura en forma de lóbulo entre los terguitos quinto y sexto en el abdomen de la hembra son caracteres clave para separar a esta especie de otras del género Stagmomantis.
The order Mantodea (praying mantises) comprises about 2,500 species worldwide, distributed in 16 superfamilies, 29 families, 60 subfamilies, and 436 genera (Ehrmann, 2002; Otte & Spearman, 2005; Schwarz & Roy, 2019). Mantidae Latreille, 1802 is the most species-rich family, containing approximately half of the described species (Patel & Singh, 2016; Svenson & Whiting, 2004, 2009); however, the limits and phylogenetic relationships within Mantidae remain unclear. Several groups previously defined based on morphological data were later recovered as non-monophyletic by analyses using mitochondrial and nuclear genes (Svenson & Whiting, 2004, 2009), as well as mitochondrial genomes (Wang et al., 2022; Xu et al., 2021).
Stagmomantis Saussure, 1869 is a New World genus belonging to the subfamily Stagmomantinae Giglio-Tos, 1919. It is one of the most species-rich genera within Mantidae, with 25 recognized species (Anderson, 2025). Species delimitation within Stagmomantis has been challenging due to significant morphological variation among its members, rendering the number of valid species debatable (Agudelo-Rondón et al., 2007; Anderson, 2020a, b, 2021; Ehrmann, 2002; Maxwell, 2014; Otte & Spearman, 2005; Terra, 1995). For instance, Maxwell (2014) did not recognize S. conspurcata Serville, 1869 as a valid species, treating it as a synonym of S. carolina (Johansson, 1763). Later, Anderson (2020b) revalidated S. conspurcata, arguing that sufficient morphological differences —such as body size and male hindwing coloration— justified its recognition as a distinct species. This case exemplifies the taxonomic instability present within Stagmomantis, which has important implications, such as constant changes in the reported geographic distribution patterns of its species.
In 1935, Rehn described Stagmomantis hebardi —named after the American entomologist Morgan Hebard— based on a female and a male specimen from Sinaloa, Mexico. Hebard (1923) had previously identified these specimens as S. limbata Hahn, 1835 and S. tolteca Saussure, 1861, respectively. In his description, Rehn included images of the dorsal habitus of both the female and male, as well as an illustration of the male’s internal genitalia. The taxonomic status of S. hebardi, based on the male-female association established by Rehn, has remained unchallenged for decades.
Recently, a molecular analysis grouped together a female and a male specimen previously identified as S. carolina (Varela-Hernández et al., 2022). In addition to the molecular data, that study included high-resolution images of the dorsal habitus. Subsequent comparison of those specimens with the descriptions and illustrations in Rehn (1935) revealed that the male assigned to S. carolina corresponded to S. hebardi. Crucially, the morphological traits of the female described as S. hebardi by Rehn (1935) did not match those of the female specimen molecularly associated with the male in Varela-Hernández et al. (2022).
Based on this evidence, we consider that Rehn (1935) erroneously associated the sexes of S. hebardi, establishing a heterospecific type series. Consequently, the main objectives of this study are to correctly associate both sexes of S. hebardi, to redescribe the male, and to formally describe the female of S. hebardi (Rehn, 1935). This work is supported by the aforementioned molecular evidence and is based on external morphology as well as a comprehensive analysis of male and female genitalia.
Materials and methods
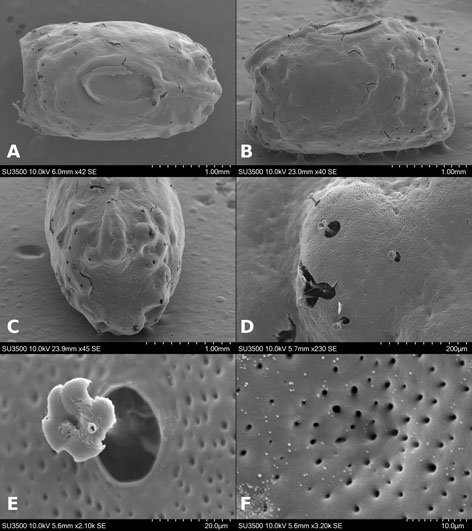
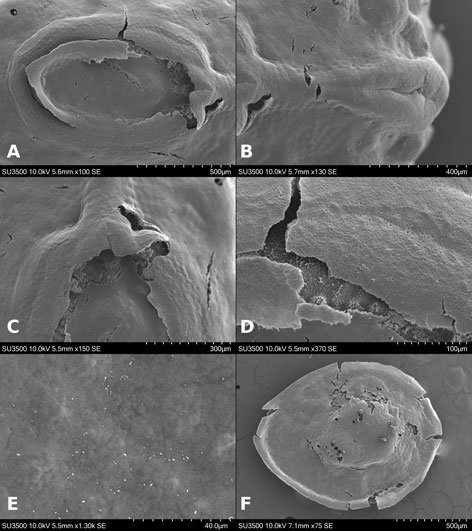
A total of 48 pinned specimens identified as Stagmomantis hebardi were examined (29 males and 15 females, see “Taxonomic summary”). These specimens are deposited at the Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México (CNIN-IBUNAM) and the Colección Entomológica de la Universidad Autónoma del Estado de Morelos, México (CEUAEM-MAN). Additionally, we examined high-resolution images of the holotype (Fig. 1) and allotype (Fig. 2) deposited at the Academy of Natural Sciences of Philadelphia (ANSP), USA.
Figure 1. Holotype of Stagmomantis hebardi Rehn, 1935, Academy of Natural Sciences of Philadelphia (ANSP). Images provided by Jason D. Weintraub (ANSP Collection Manager).Figure 2. Allotype of Stagmomantis hebardi Rehn, 1935, Academy of Natural Sciences of Philadelphia (ANSP). Images provided by Jason D. Weintraub (ANSP Collection Manager).
Observations and measurements were conducted using an Olympus SZ stereomicroscope. Specimens identification at genus and species level followed the keys and descriptions provided by Rehn (1935), Agudelo-Rondón and Chica (2002), Maxwell (2014), and Anderson (2021). Specimens of both sexes were photographed from multiple angles using a Canon EOS Rebel T5 digital camera equipped with an EF 100 mm f/2.8 Macro USM lens. Camera control was managed via EOS Utility software; images were subsequently stacked and edited using Zerene Stacker and Photoshop CC 2019.
Terminalia of both sexes were softened in hot water (80-100 °C) for 15-20 min, following the protocol of Ferraz et al. (2023). Genitalia were then cleared in a 10% KOH solution at 40 °C for 20-30 min (Brannoch et al., 2017). The extraction of female genitalia followed the method described by Rodrigues and Cancello (2016). Cleared genitalia were rinsed twice in distilled water and stored in 70% ethanol. Remaining soft tissue was removed, and structures were photographed suspended in antibacterial gel to maintain their position (Su, 2016). Abbreviations for all measurements and genitalic structures are provided in Table 1.
To visualize the geographical distribution of the examined specimens, a map was generated using ArcGIS software, utilizing the State Geostatistical Framework, scale 1:250,000 (INEGI, 2024).
Description
Stagmomantis hebardi Rehn, 1935
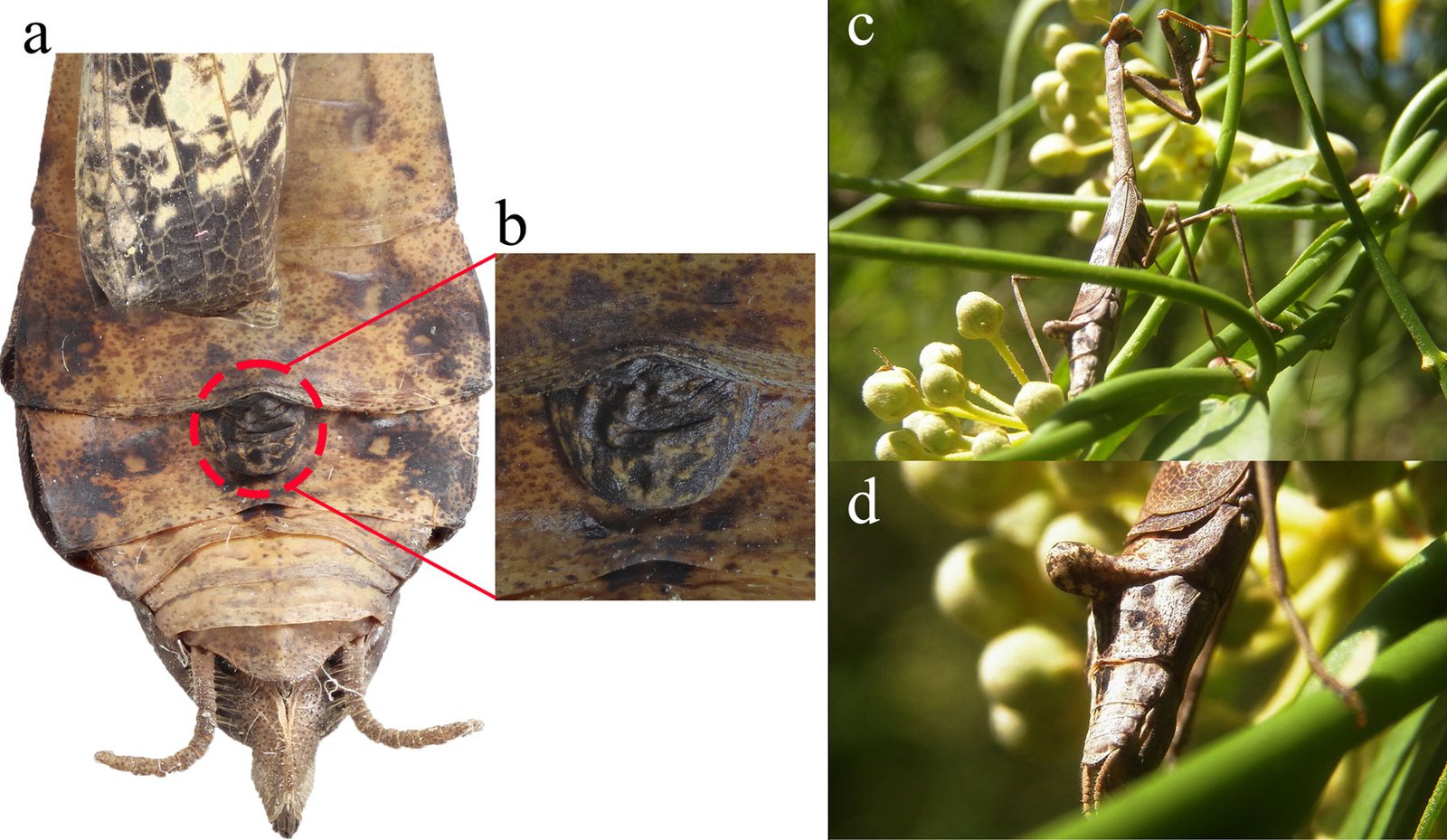
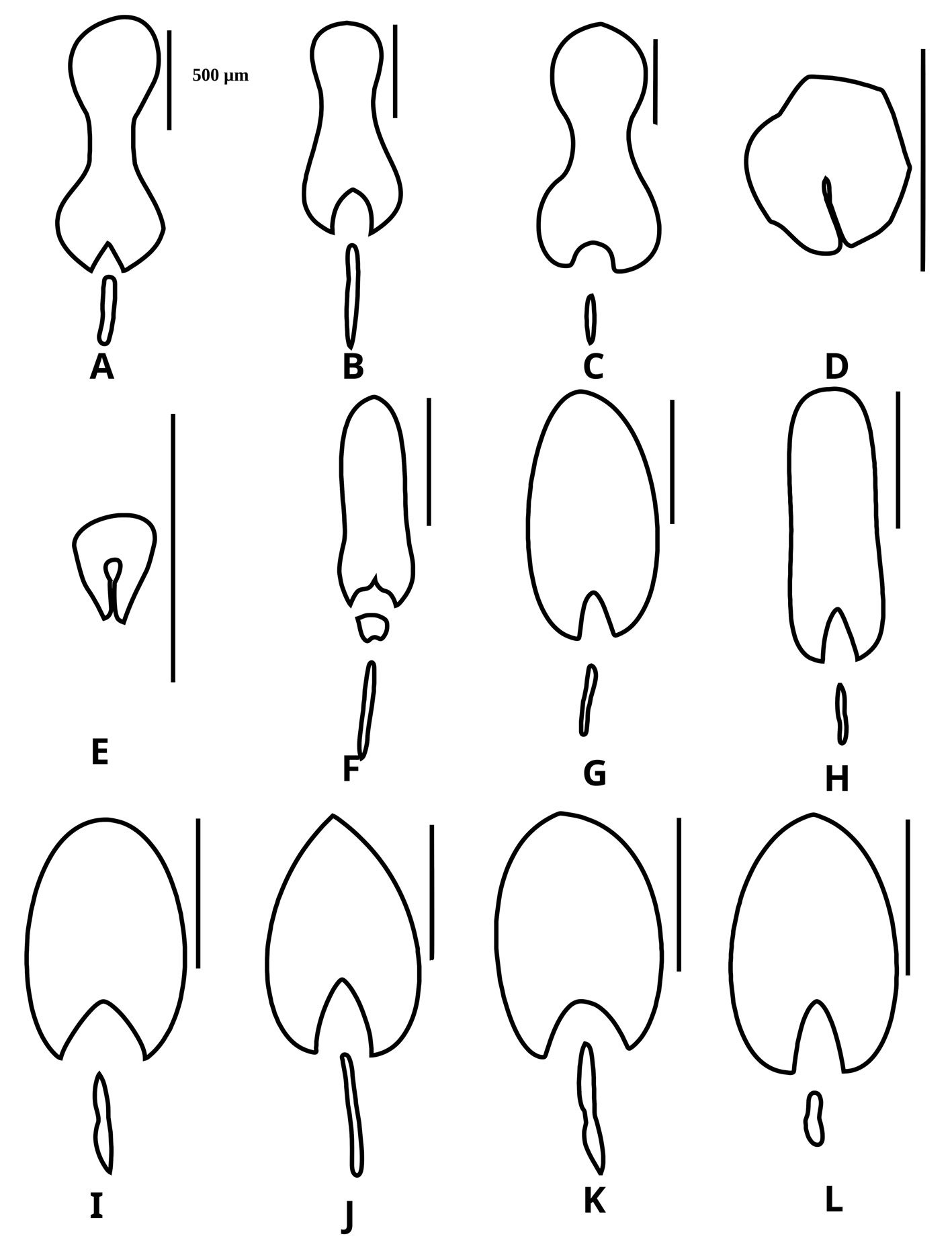
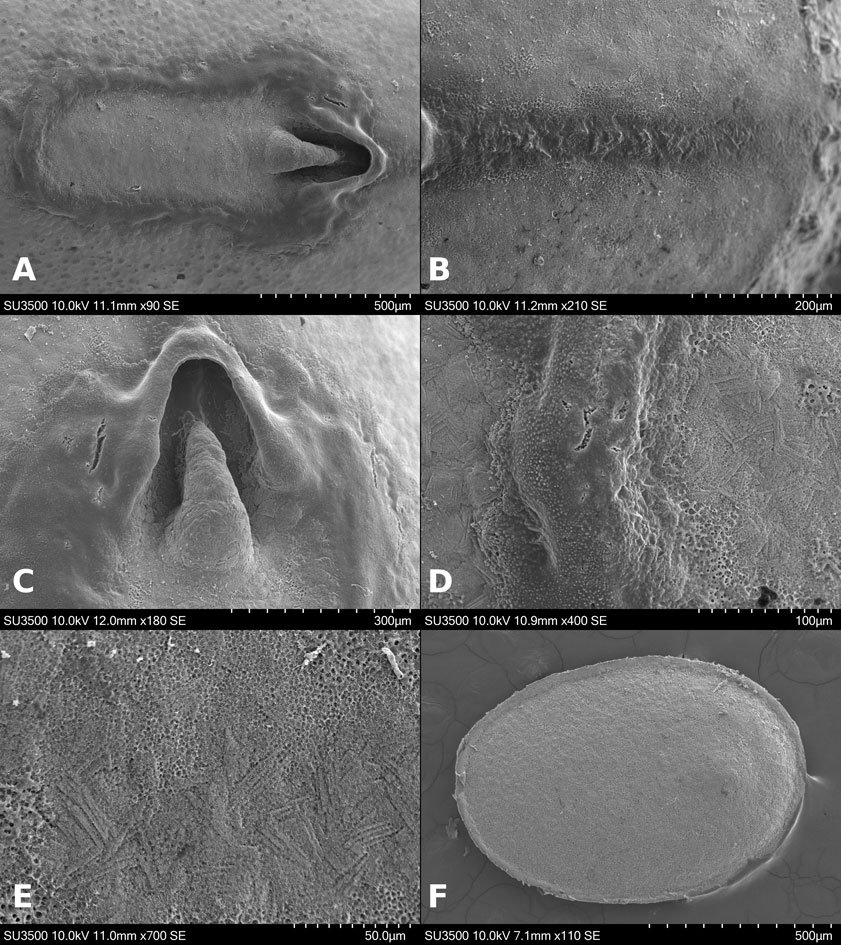
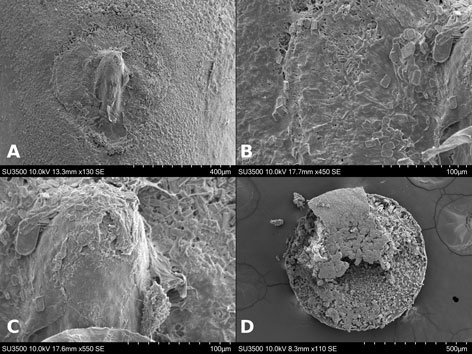
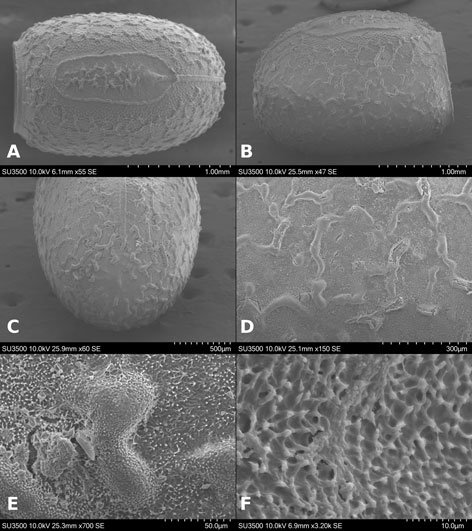
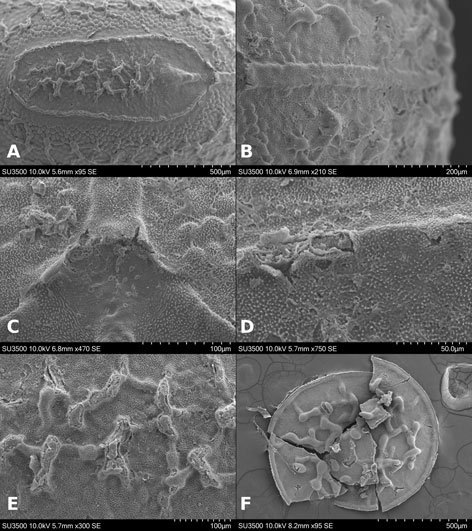
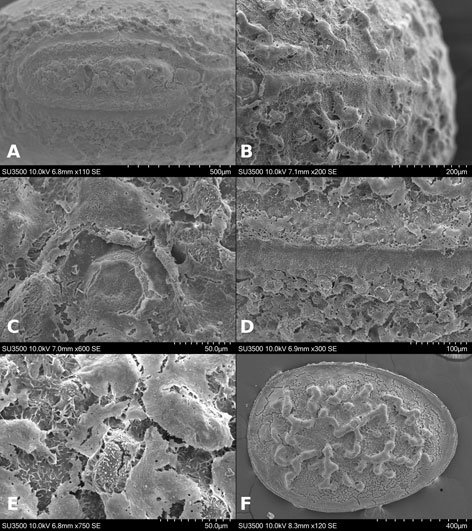
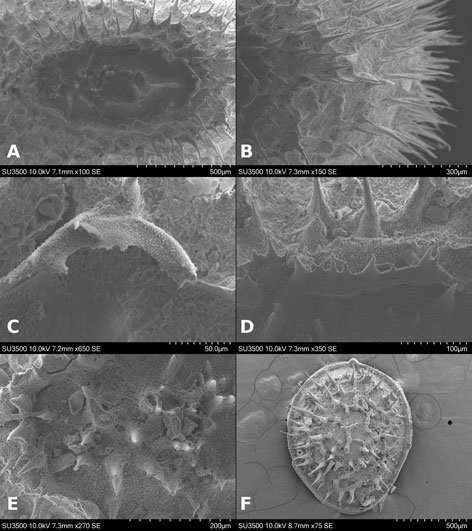
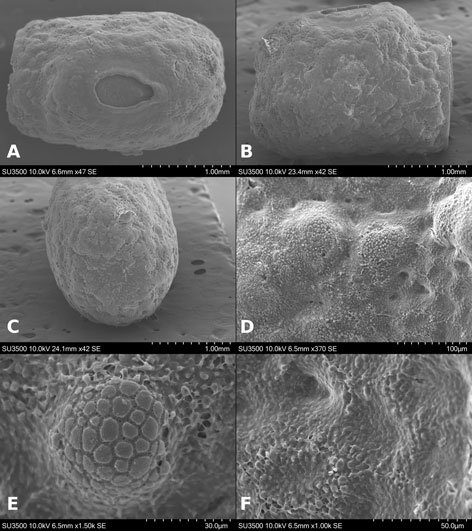
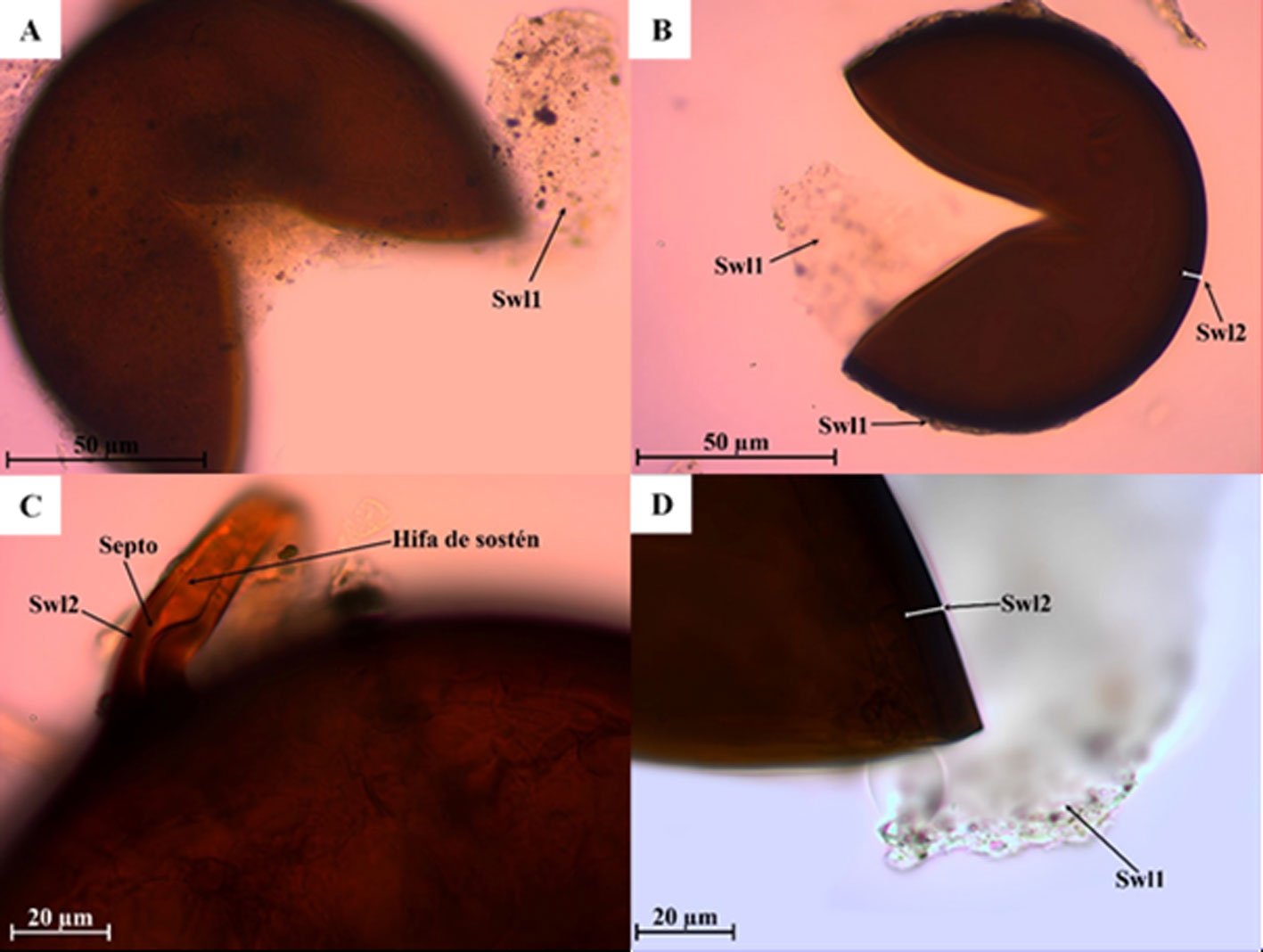
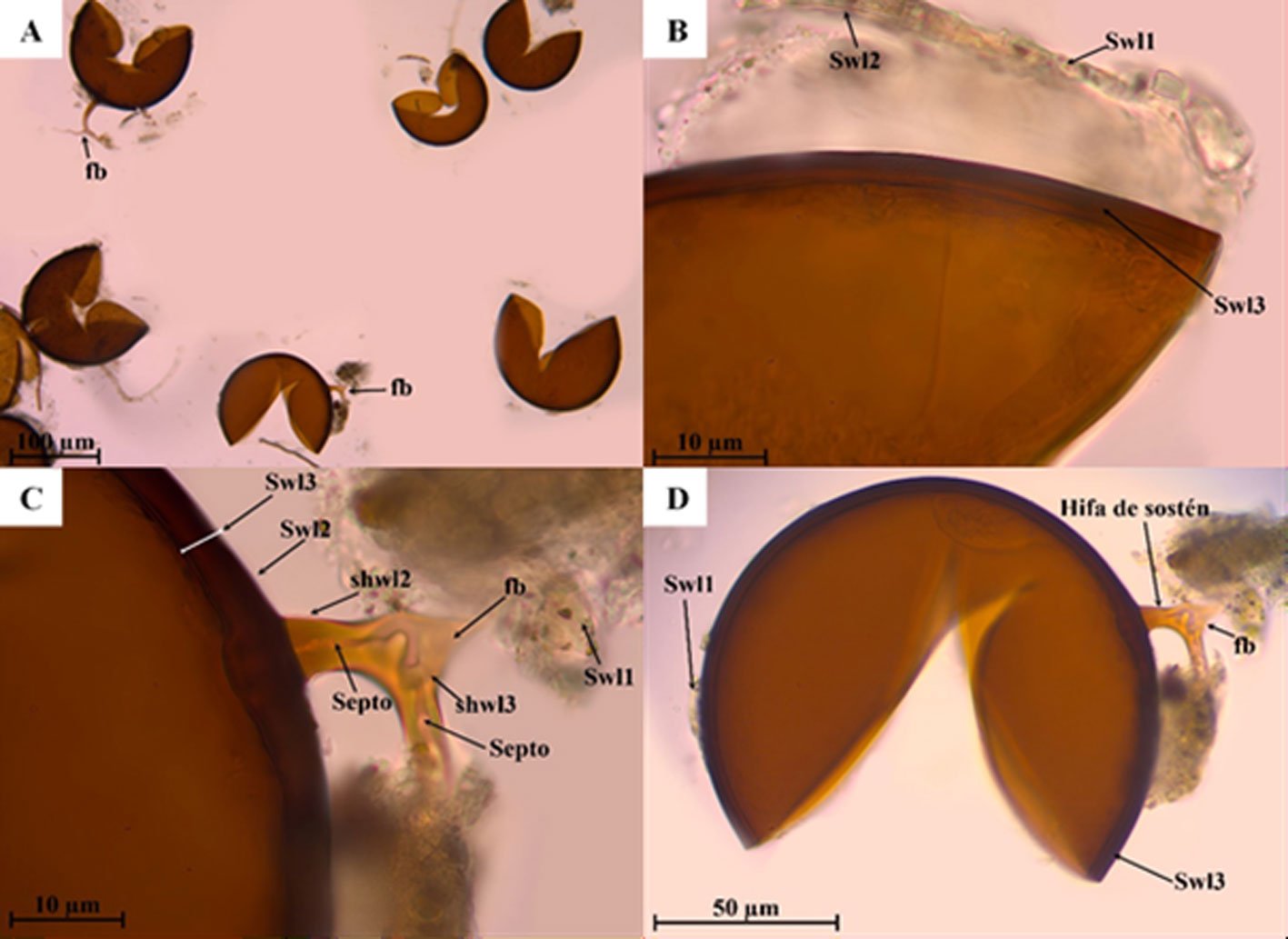
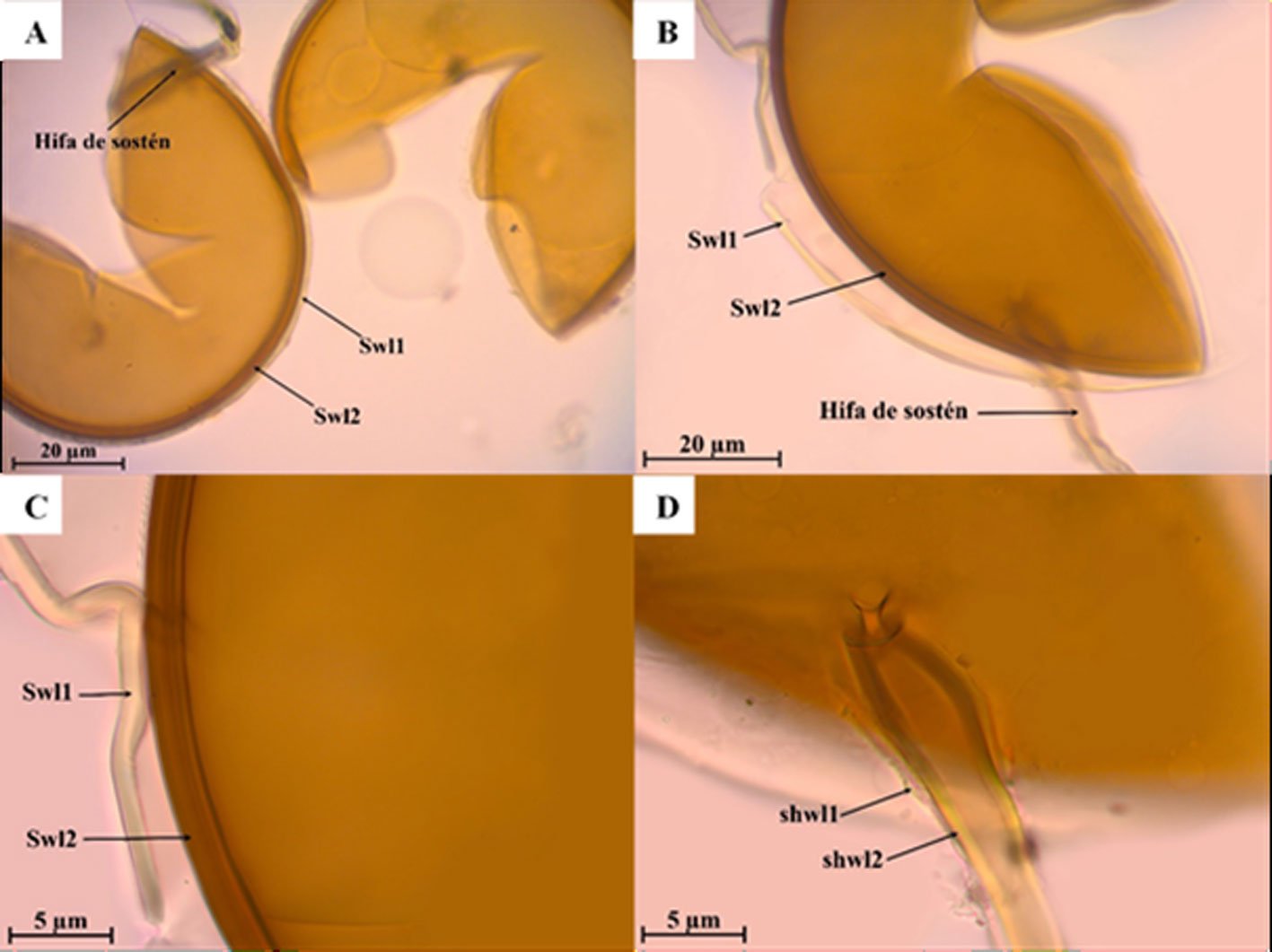
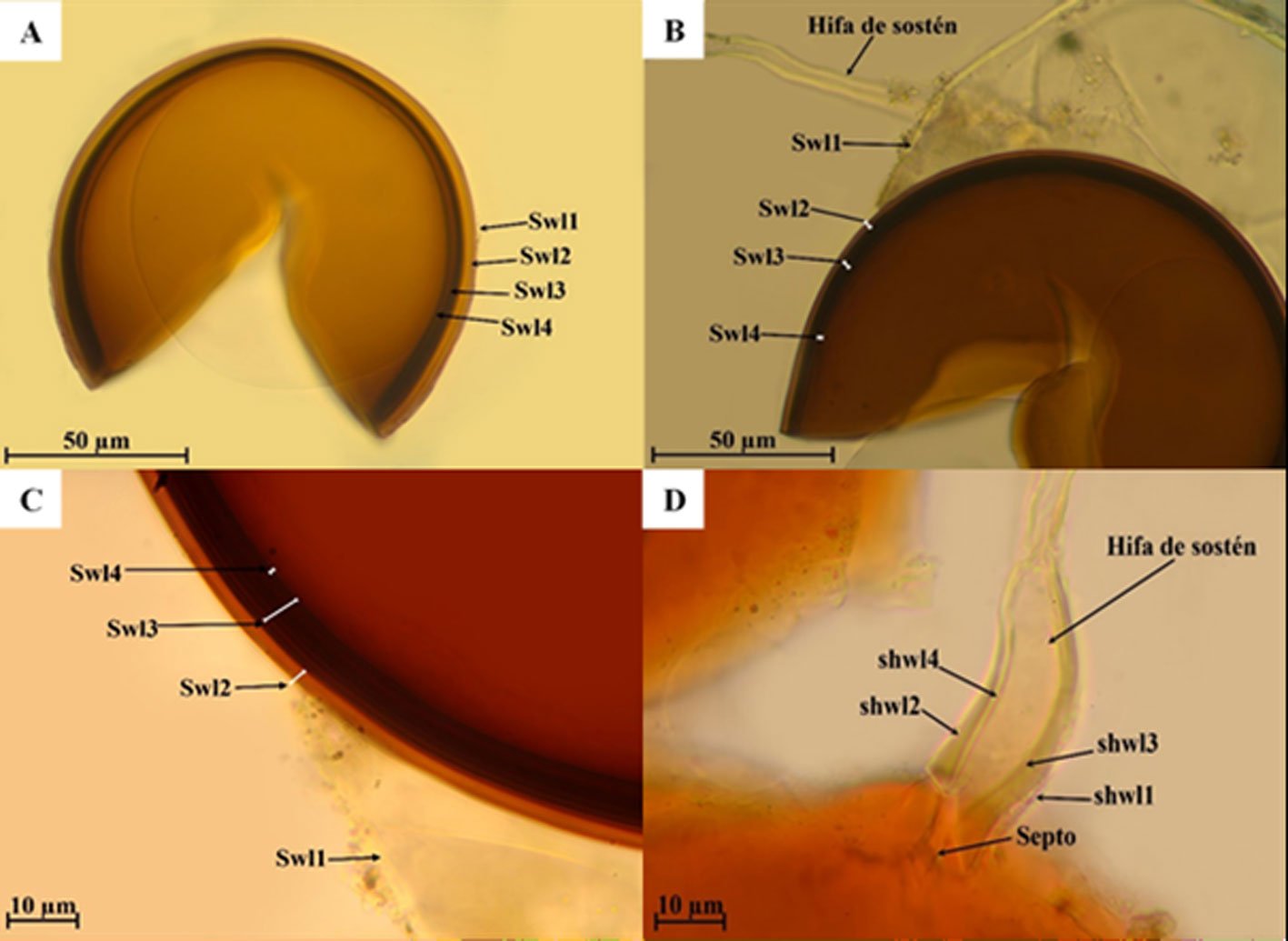
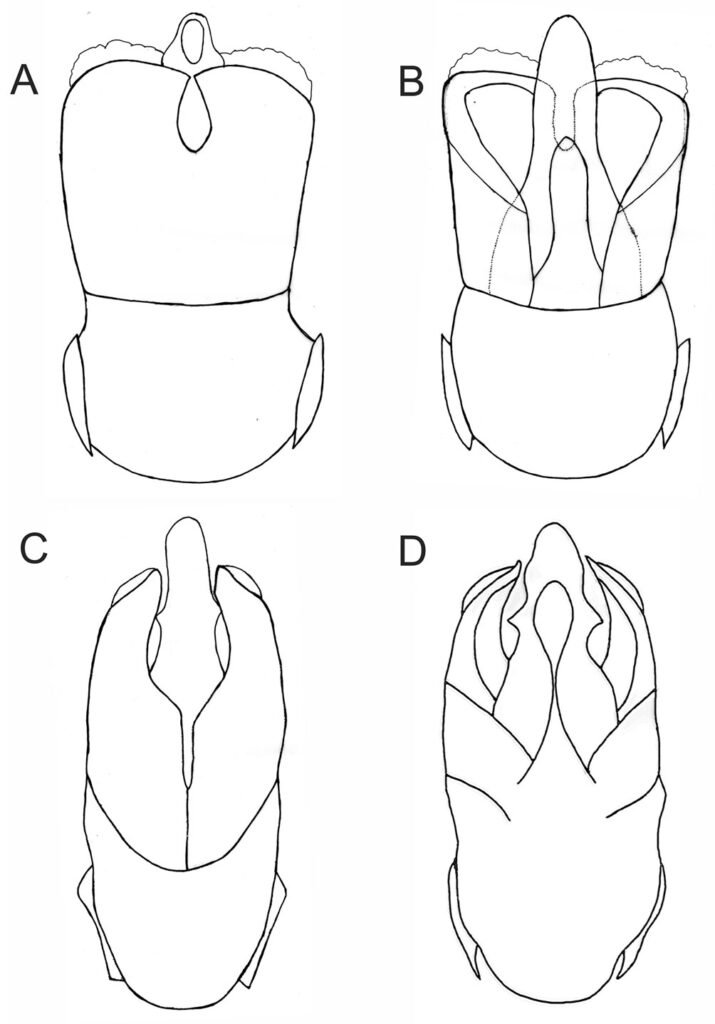
Diagnosis. Male: distinguished by the corkscrew-like posterior process of the ventral phallomere (pda) (Fig. 3). Female: prosternum (inter-coxal area) blackish; anterior coxae armed with 3-6 blackish spines; presence of a protrusible lobe-like structure located between abdominal tergites 5 and 6 (Fig. 4).
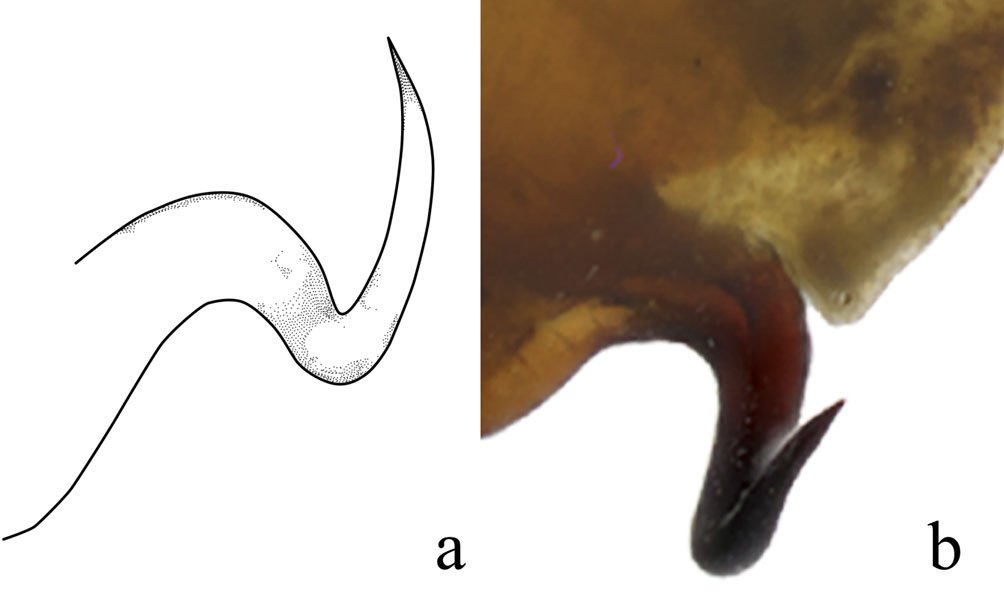
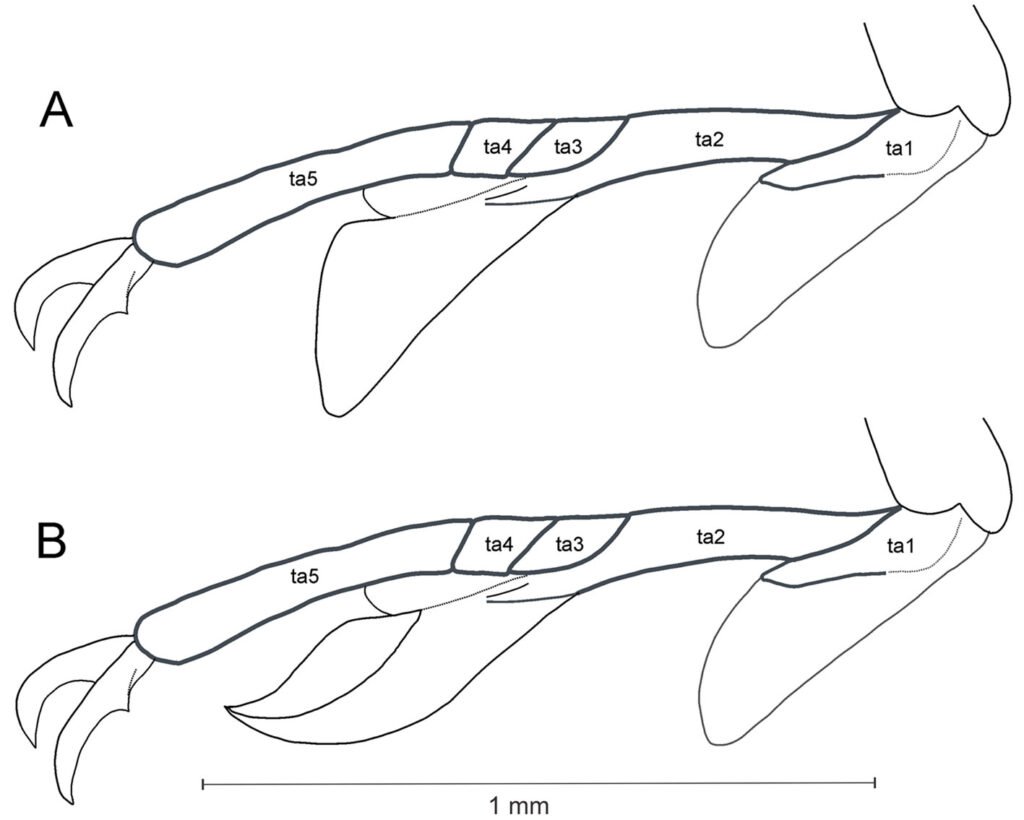
Figure 3. Comparison of the posterior process of ventral phallomere (pda) of the male genitalia of Stagmomantis hebardi. a) Illustration modified from Rehn (1935); b) specimen examined in this study.Figure 4. Protrusible lobe-like structure on the abdomen of female Stagmomantis hebardi. a-b) Dried specimen: a) dorsal view of the abdomen; b) close-up of the lobe-like structure (indicated in red). c-d) Live specimen: c) dorso-lateral view; d) close-up of the lobe-like structure. Specimens imaged: a-b (CEUAEM-MAN-139); c-d (iNaturalist observation 237796237, photo ID 423231786; photo by Edgar Salmerón Barrera).
Category
Abbreviation
Meaning
Morphological
ACL
Anterior coxae length
Morphological
AFL
Anterior femur length
Morphological
AFW
Anterior femur width
Morphological
ATL
Anterior tibia length
Morphological
AvS
Anteroventral spines
Morphological
DS
Discoidal spines
Morphological
FwL
Forewing length
Morphological
HH
Head height
Morphological
HW
Head width
Morphological
HwL
Hindwing length
Morphological
ML
Metazone length
Morphological
MFL
Metafemur length
Morphological
MTL
Metatibia length
Morphological
MsFL
Mesofemur length
Morphological
MsTL
Mesotibia length
Morphological
PL
Pronotum length
Morphological
PrL
Prozone length
Morphological
PvS
Posteroventral spines
Morphological
PW
Pronotum width
Morphological
TL
Total length
Male genitalia
aafa
Anterior lobe of phalloid apophysis (left phallomere)
Male genitalia
an
Anterior extension of sclerite R3 (right phallomere)
Male genitalia
bm
Dextral extension (right phallomere)
Male genitalia
fda
Main posterior lobe (right phallomere)
Male genitalia
L4A
Sclerite extending over the ventral wall (ventral phallomere)
Male genitalia
L4B
Sclerite extending over the dorsal wall (left phallomere)
Male genitalia
loa
Posteromesal (left phallomere)
Male genitalia
paa
Posterior process (left phallomere)
Male genitalia
pafa
Posterior lobe of phalloid apophysis (left phallomere)
Male genitalia
pda
Posterior process (ventral phallomere)
Male genitalia
pia
Process posterolateral to pva (right phallomere)
Male genitalia
pva
Process anteromesal to pia (right phallomere)
Male genitalia
R3
Anteriorly extending sclerite (right phallomere)
Male genitalia
ssp
Secondary spine
Female genitalia
agsl
Accessory gland supporting lobe
Female genitalia
CG8
Caudogyne
Female genitalia
CX8
Coxa 8
Female genitalia
cxdl
Dorsolateral coxal lobelet
Female genitalia
cxvl
Ventrolateral coxal lobelet
Female genitalia
gl9
Gonoplac 9
Female genitalia
gp8
Gonapophysis 8
Female genitalia
gp9
Gonapophysis 9
Female genitalia
gpal8
Apical lobe of gonapophysis 8
Female genitalia
gpmo8
Medial outgrowth of gonapophysis 8
Female genitalia
gptm9
Medial tine of gonapophysis 9
Female genitalia
rh
Rhachis
Female genitalia
spb
Spermathecal bulge
Female genitalia
vf7
Ventral segmental fold 7
Female genitalia
VS7
Vestibular sclerite 7
Table 1. Abbreviations of male and female genitalia characters. Morphological characters follow Garikipati (2024) and genitalia characters follow Brannoch et al. (2017).
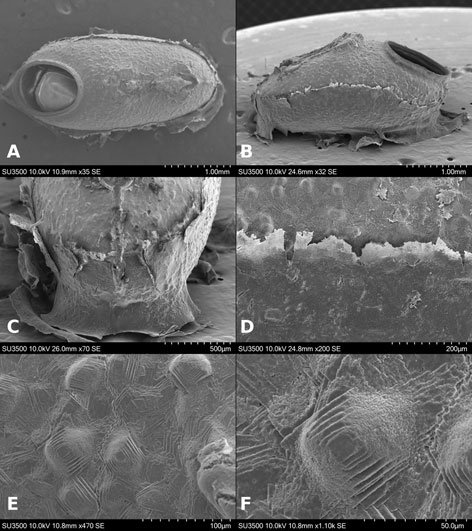
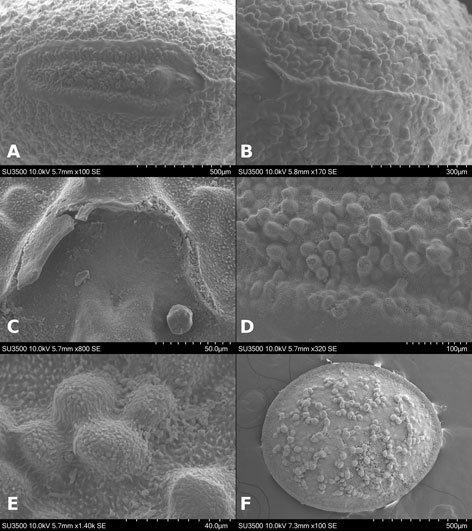
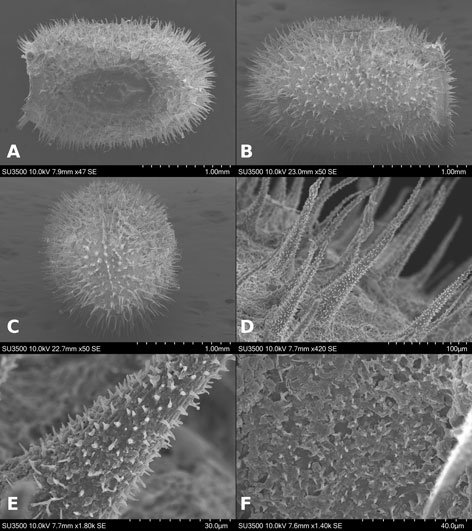
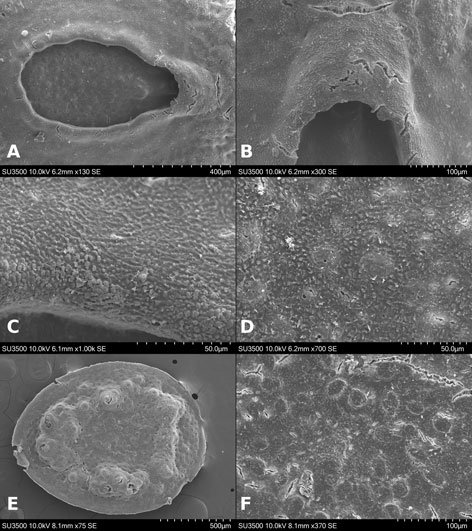
Identification. Males: antennae longer than pronotum. Tubercle present between antennifer and compound eyes. Wings longer than abdomen; fore- and hindwings hyaline; costal area of forewings opaque with brown spots. Genitalia with the posterior process of ventral phallomere (pda) corkscrew-shaped. Females: abdominal tergites 4-6 dorsoventrally flattened; protrusible lobe-like structure present between tergites 5 and 6. Supragenital plate wider than long with well-developed longitudinal medial keel. Forewings opaque, greenish or brownish, with brown spots; hindwings opaque, anteriorly yellowish with brown spots near posterior margin, or yellowish with colorless spots posteriorly. Both sexes: ocellar tubercles absent; ocelli flush with surface of frons. Lower frons smooth, wider than long. Pronotum long and slender; lateral margins denticulate; dorsal surface smooth. Forecoxae approximately 3/4 the length of pronotum; anterior and posterior margins denticulate; forecoxal lobes convergent. Hearing organ of DK type.
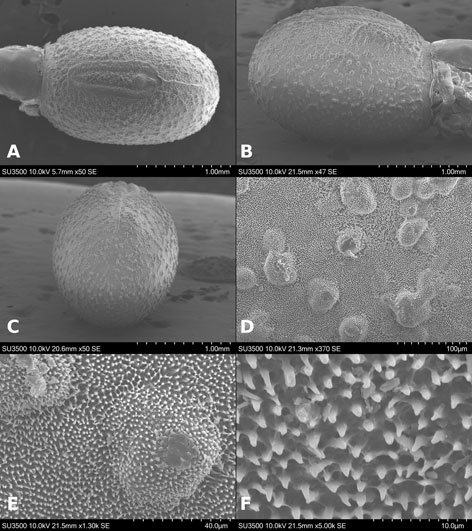
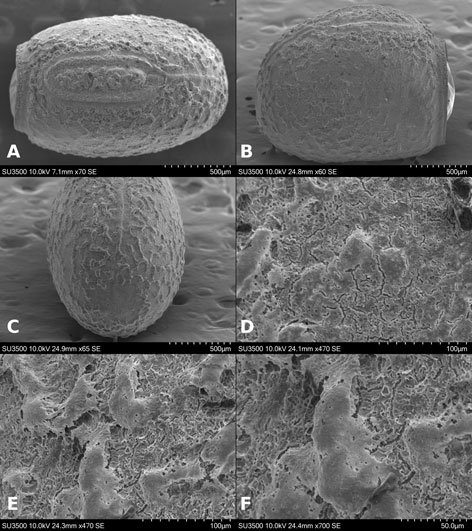
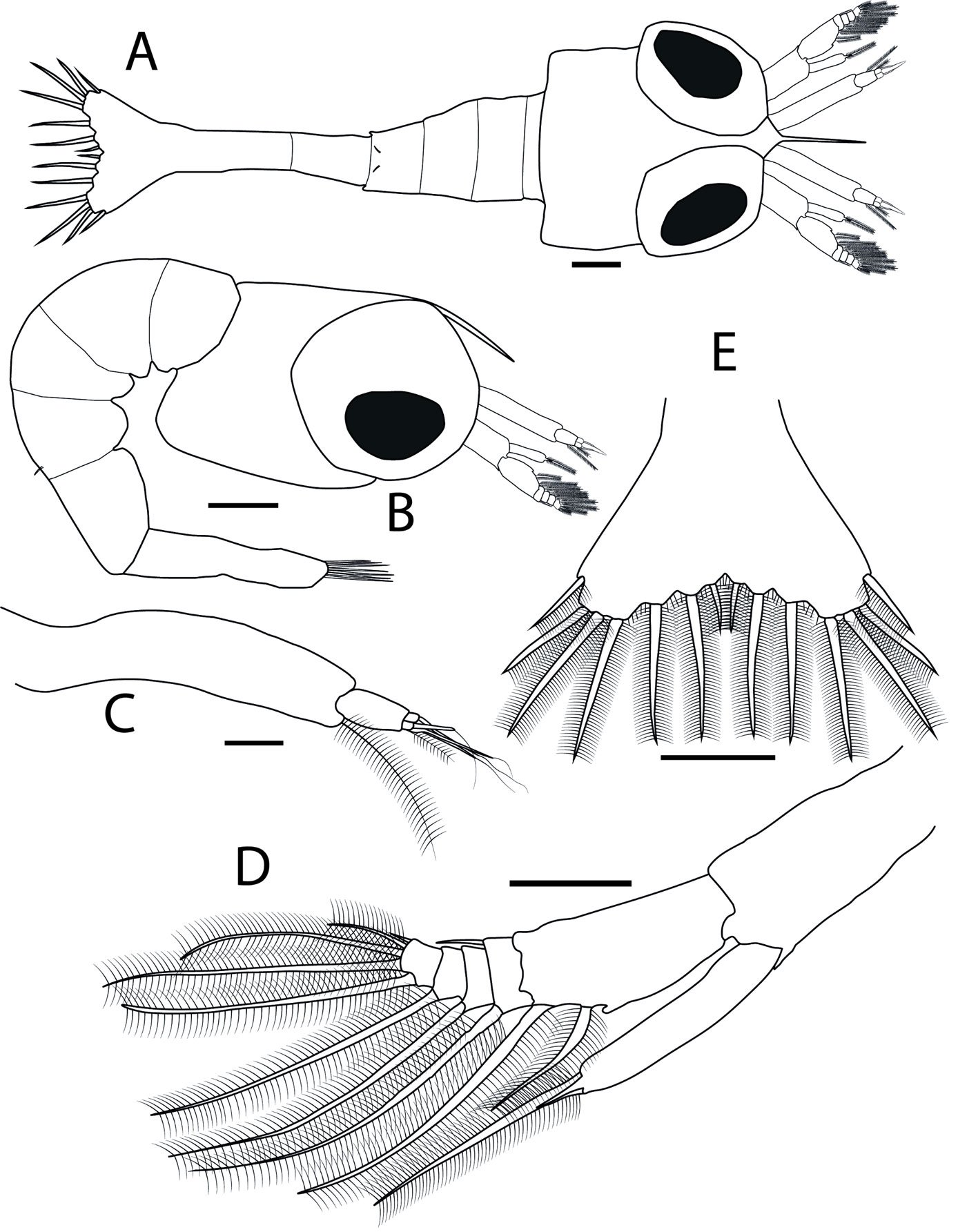
Male (Fig. 5). Measurements (Table 2). Head: head capsule (Fig. 6a) with oval eyes; juxtaocular bulges undeveloped; ocellar tubercles undeveloped. Anterior margin of vertex straight, depressed near parietal sulcus. Ocelli larger than in females. A small tubercle present between antennifer and compound eye. Antennae filiform, longer than pronotum, reaching anterior portion of the mesothorax; pedicel nearly as long as scape. Frons wider than long, with an M-shaped depression in the middle of the transverse carina. Thorax: pronotum, supracoxal dilation slightly pronounced; supracoxal groove well-marked. Lateral margins of prozone denticulate; lateral margins of metazone denticulate, denticles decreasing in size posteriorly. In ventral view, inter-coxal area dark-colored. Legs: forelegs, spination formula: F = 4DS/12-15AvS/4PvS; T = 9-14AvS/9-11PsV. Forecoxae armed with 3-5 blunt, often blackish spines, small denticles interspersed between them; coxal lobes convergent (Fig. 6b). Forefemur relatively long, slender; femoral brush located at the level of last 3 posteroventral spines; tibial spur groove located at approximately the middle of femur; first discoidal spine smaller than the others; third discoidal spine longer than the others; blackish spot present at the level of first anteroventral spine (AvS). Foretarsus: distal portion of each segment darkened (Fig. 6c, d). Wings: slightly surpassing tip of abdomen. Forewings: hyaline, with scattered brown spots in discoidal area; stigma covered by dark brown patch; anterior margin wide at base, narrowing posteriorly; apex rounded. Hindwings: hyaline with a dark patch at apex; main central longitudinal veins feature a series of spotted brownish lines running along posterior margin to about half the distance to anterior margin (Fig. 5). Abdomen: slender, cylindrical. Subgenital plate longer than wide. Supragenital plate wider than long; apex rounded, with marked longitudinal medial keel. Cerci elongate; cercomeres cylindrical, subequal in length; distal cercomere smaller, conical, rounded at apex.
Figure 5. Male of Stagmomantis hebardi (green/brown morphotype). a) Dorsal view; b) ventral view. Specimen imaged: CEUAEM-MAN-140. Scale bars = 10 mm.
Genitalia: left phallomere, sclerite L4B almost rectangular, longer than wide, with left margin projected anteriorly (Fig. 7a). Anterior lobe of phalloid apophysis (aafa) linguiform, sometimes oval, distally dentate. Posterior lobe of phalloid apophysis (pafa) elongated, apically rounded, dentate, bearing 3-6 ventral denticles. Posteromesal lobe (loa) oval, bearing short setae. Posterior process (paa) robust, sometimes slender. Right phallomere: triangular (Fig. 7b); posterior apex rounded with short setae. Dextral extension (bm) thick, slightly elongate; anterior extension of sclerite R3 (an) elongate, slender. Process anteromesal to pia (pva) short, thick, smooth with rounded apex; process posterolateral to pva (pia) short, smooth. Ventral phallomere: sclerite L4A oval (Fig. 7c, d). Secondary spine (ssp) well-sclerotized, triangular. Posterior process (pda) corkscrew-shaped, twisted left or right, with apex angled to slightly flattened.
Spec. ID
TL
PL
PrL
ML
PW
HH
HW
ACL
AFL
AFW
ATL
FwL
HwL
MsFL
MsTL
MFL
MTL
CEUAEM-MAN-140
51.0
16.5
3.5
13.0
2.5
4.0
5.0
8.5
9.5
1.0
6.0
33.0
30.5
10.0
9.0
13.0
13.0
CEUAEM-MAN-135
51.0
15.5
3.0
14.0
3.0
4.0
5.0
7.5
10.0
1.0
6.0
32.0
31.0
10.0
9.0
13.0
13.0
CEUAEM-MAN-013
55.0
18.0
4.0
14.5
3.0
4.0
5.5
9.0
11.0
1.5
7.0
35.0
34.0
11.0
10.0
13.0
13.0
CEUAEM-MAN-021
45.5
15.5
3.0
12.0
2.5
3.5
5.0
7.5
9.5
1.0
5.0
31.0
27.0
10.0
9.0
11.0
12.0
CEUAEM-MAN-015
47.0
16.5
4.0
12.5
2.5
4.0
5.0
8.0
10.0
1.5
6.0
34.0
31.5
10.0
9.0
13.0
12.0
CEUAEM-MAN-014
45.0
18.0
3.5
13.5
3.0
3.5
5.5
8.5
10.5
1.5
6.0
34.0
31.0
10.5
9.5
13.0
13.0
IBUNAM: CNIN: MA645
64.0
21.5
3.5
18.0
3.5
–
–
–
12.0
–
6.0
–
–
–
–
15.0
14.5
IBUNAM: CNIN: MA810
55.5
19.0
3.0
16.0
3.0
–
–
–
11.0
–
5.0
–
–
–
–
14.0
15.0
IBUNAM: CNIN: MA946
54.0
16.0
3.0
13.0
3.5
–
–
–
10.0
–
4.5
–
–
–
–
–
–
Table 2. Measurements (in mm) of males of Stagmomantis hebardi (n = 9).
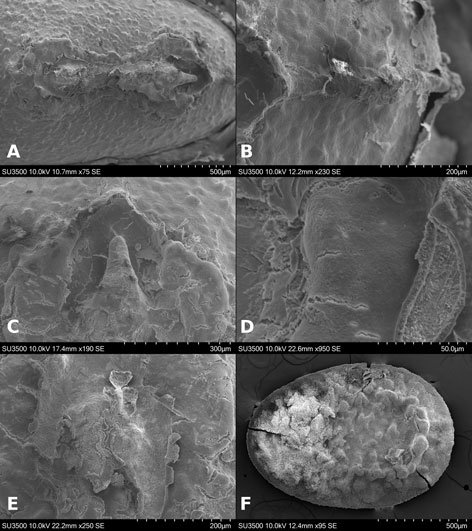
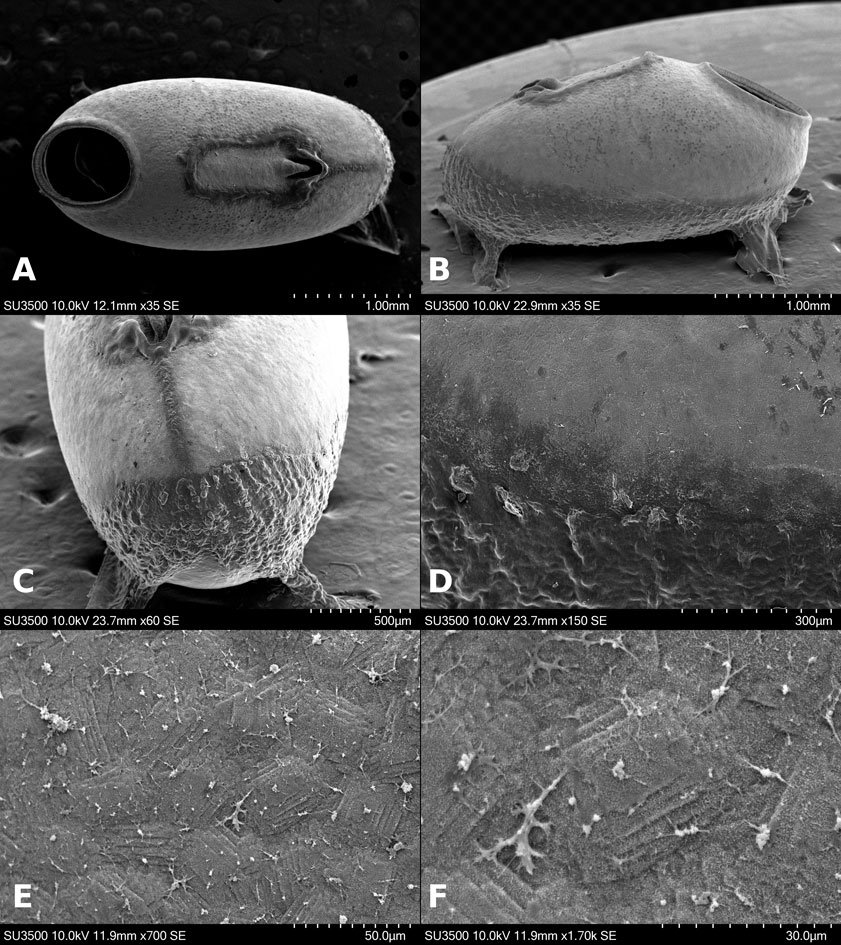
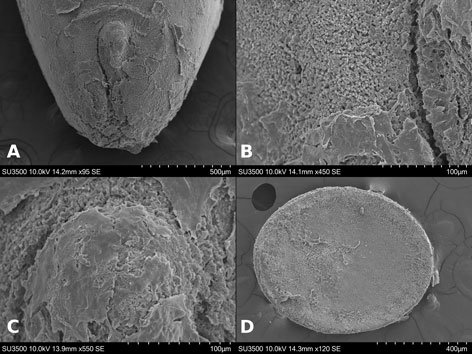
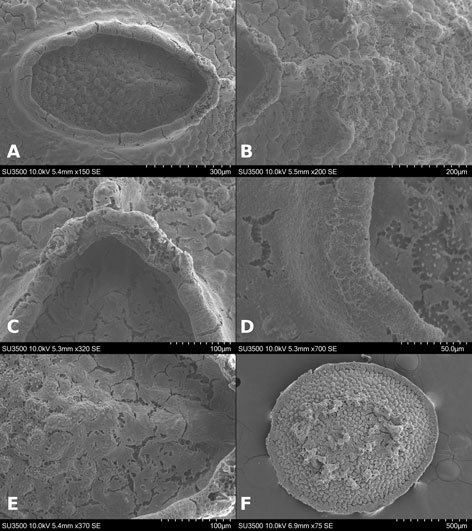
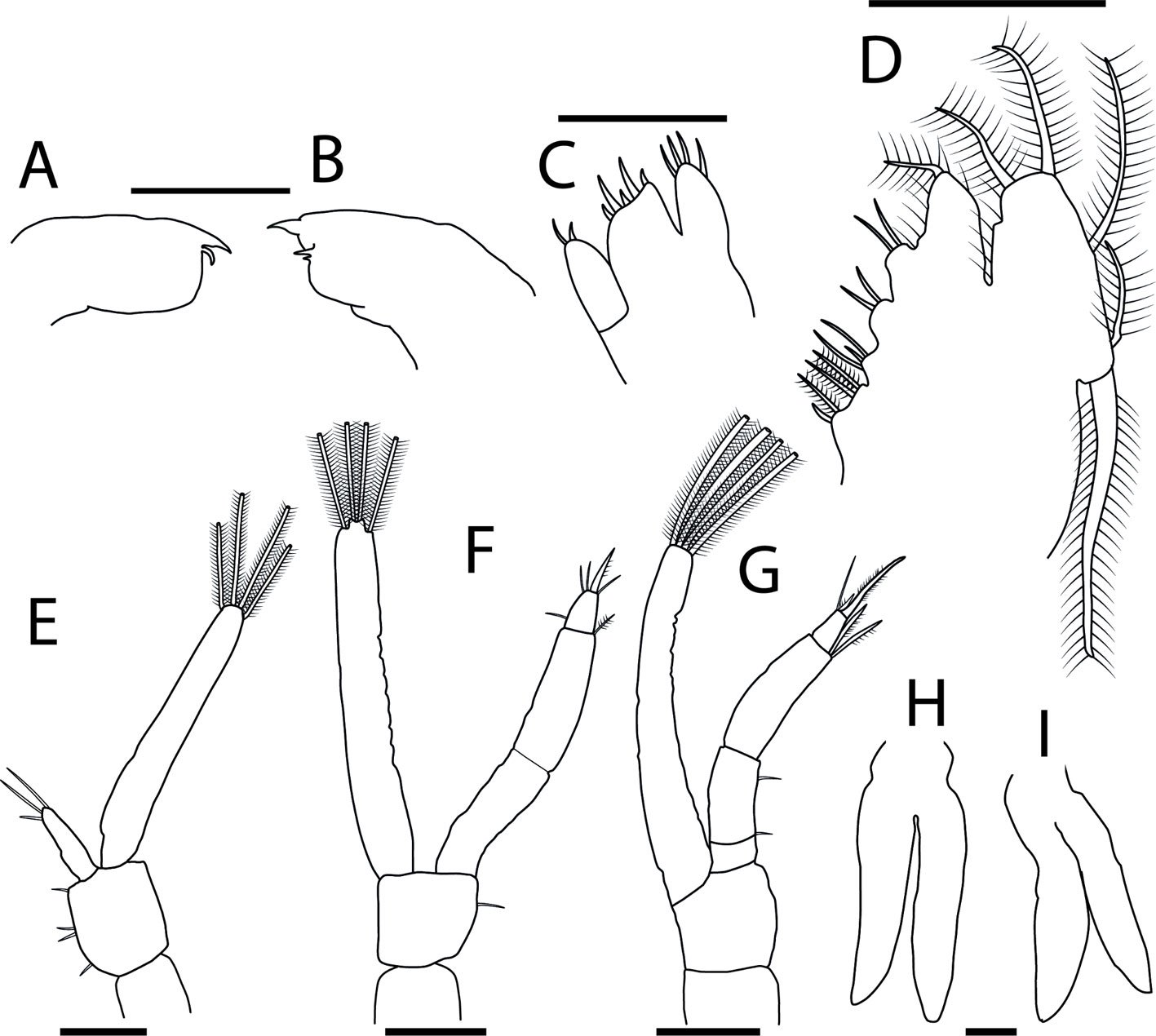
Female (Fig. 8). Measurements (Table 3). Head: head capsule with oval eyes; juxtaocular bulges absent; ocellar tubercles absent; ocelli smaller than in males. Anterior margin of vertex straight; vertex slightly depressed near parietal sulcus. Antennae filiform; scape almost twice as long as pedicel. Frons wider than long in frontal view, with M-shaped depression in the middle of transverse carina (Fig. 9a). Thorax: pronotum, supracoxal dilation slightly pronounced; supracoxal groove well-marked. Lateral margins of prozone denticulate; lateral margins of metazone denticulate, becoming smooth distally; medial keel of pronotum well-developed. In ventral view, inter-coxal area brownish. Legs: forelegs, spination formula: F = 4DS/14-16AvS/4PvS; T = 12-14AvS/8-10PvS. Forecoxae (Fig. 9b) armed with 3-6 apically blunt spines, often brownish, denticles interspersed among them; forecoxal lobes convergent. Forefemur relatively long, slender; femoral brush located at the level of last 3 posteroventral spines; tibial spur groove located at approximately half the length of femur; first discoidal spine smaller than the others; third discoidal spine longer than the others; with blackish spot at the level of first anteroventral spine (AvS). Foretarsus: distal portion of each segment blackish (Fig. 9c, d). Wings (fig. 10a-c): reaching approximately 3/4 the length of abdomen. Forewings: anterior margin wide at base, narrowing distally; color variable, ranging from brown to light hues; stigma darker than rest of wing. Hindwings fully colored; color variable, ranging from yellowish at base to middle area, with dark spots distally, to yellowish with hyaline spot; apex of hindwing yellowish with greenish portions. Abdomen: dorsoventrally flattened, reaching maximum width between 4th and 6th tergites, with prominent, protrusible lobe-like structure between 5th and 6th tergites. Supragenital plate wider than long, rounded at the apex, with a marked longitudinal medial keel. Cerci elongate; cercomeres cylindrical, subequal in length; distal cercomere smaller, conical, apically rounded.
Figure 8. Female of Stagmomantis hebardi (green morphotype). a) Dorsal view; b) ventral view. Specimen imaged: CEUAEM-MAN-138. Scale bars = 10 mm.Figure 10. Fore- and hindwing variation in female Stagmomantis hebardi. a) Green morphotype; b) light brown morphotype; c) dark brown morphotype. Specimens imaged: a (CEUAEM-MAN-138); b (CEUAEM-MAN-129); c (CEUAEM-MAN-139). Scale bars = 10 mm.
Genitalia (Fig. 11): gonoplacs (gl9), simple, elongate, apically rounded, bearing short setae throughout, with longer setae apically. Gonapophysis: gonapophysis 9 (gp9) membranous, elongate, shorter than gl9. Gonapophysis 8 (gp8) elongate, bearing short setae; in lateral view, distinct furrow near ventral margin; lateral margins sinuous depressed near base of apical lobe (gpal8). Apical lobe of gonapophysis 8 (gpal8) glove-shaped; dorsal portion laterally compressed, distally rounded; ventral portion short, elongate; surface covered with short setae, longer setae on inner surface. Medial outgrowth of gonapophysis 8 (gpmo8) well-developed. Coxa 8 (CX8): acuminate, broad, rounded at base, tapering distally; divided into dorsal and ventral regions by a marked depression, a prominent keel extending through middle of depression. Dorsal region of CX8 elongate lateral wider, more rounded than dorsal region. Caudogyne (CG8) weakly sclerotized; dorsal margin rounded, sinuous; ventral margin depressed at the middle (M-shaped). Spermathecal bulge (spb) well-sclerotized. Accessory gland supporting lobe (agsl) smooth, weakly sclerotized. Ventral segmental fold 7 (vf7) membranous, rounded, with sinuous margins. Vestibular sclerite (VS7) weakly sclerotized, rhomboidal in shape, with ventro-medial corner rounded.
Spec. ID
TL
PL
PrL
ML
PW
HH
HW
ACL
AFL
AFW
ATL
FwL
HwL
MsTL
MFL
MTL
CEUAEM-MAN-139
52.0
20.0
4.5
15.5
4.0
4.5
7.0
10.0
13.0
1.5
8.0
19.5
17.0
12.0
10.5
15.5
CEUAEM-MAN-129
48.0
18.0
3.5
15.0
3.0
4.2
6.0
9.0
11.5
1.5
7.0
19.0
17.0
11.0
10.0
15.0
CEUAEM-MAN-019
48.0
20.0
4.5
15.5
3.8
4.5
7.0
9.5
12.0
2.0
8.0
–
–
12
11
16.0
CEUAEM-MAN-018
49.0
21.0
4.5
17.0
4.0
4.5
6.5
10.0
13.0
2.0
8.0
21.0
–
12
11
15.5
CEUAEM-MAN-017
51.0
20.0
4.5
15.5
4.0
5.0
7.0
10.0
13.0
2.0
8.0
–
17.0
12.0
11.0
16.0
CEUAEM-MAN-138
53.0
20.0
4.5
16.0
4.0
5.0
7.0
10.5
13.0
2.0
8.0
22.0
18.0
12.0
11.5
16.0
CEUAEM-MAN-109
48.0
19.5
4.0
15.5
4.0
4.0
7.0
9.5
12.5
2.0
6.5
20.0
16.0
12.0
11.0
15.0
IBUNAM: CNIN: MA1560
55.0
23.0
5.0
18.0
4.0
–
–
–
13.0
–
6.5
–
–
–
–
17.0
IBUNAM: CNIN: MA946
54.0
16.0
3.0
13.0
3.5
–
–
–
10.0
–
7.0
–
–
–
–
17.5
Table 3. Measurements (in mm) of females of Stagmomantis hebardi (n = 9).
Taxonomic summary
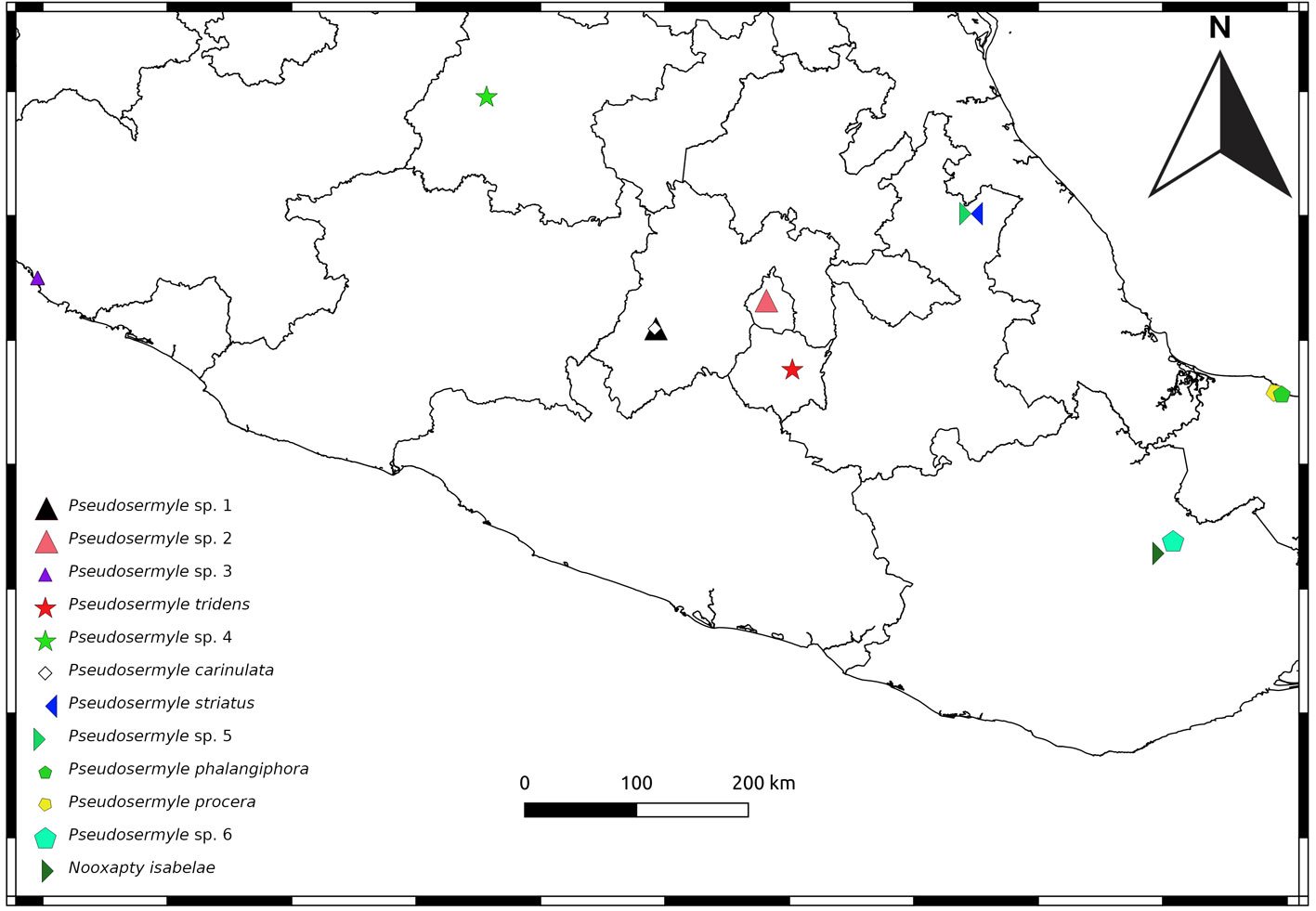
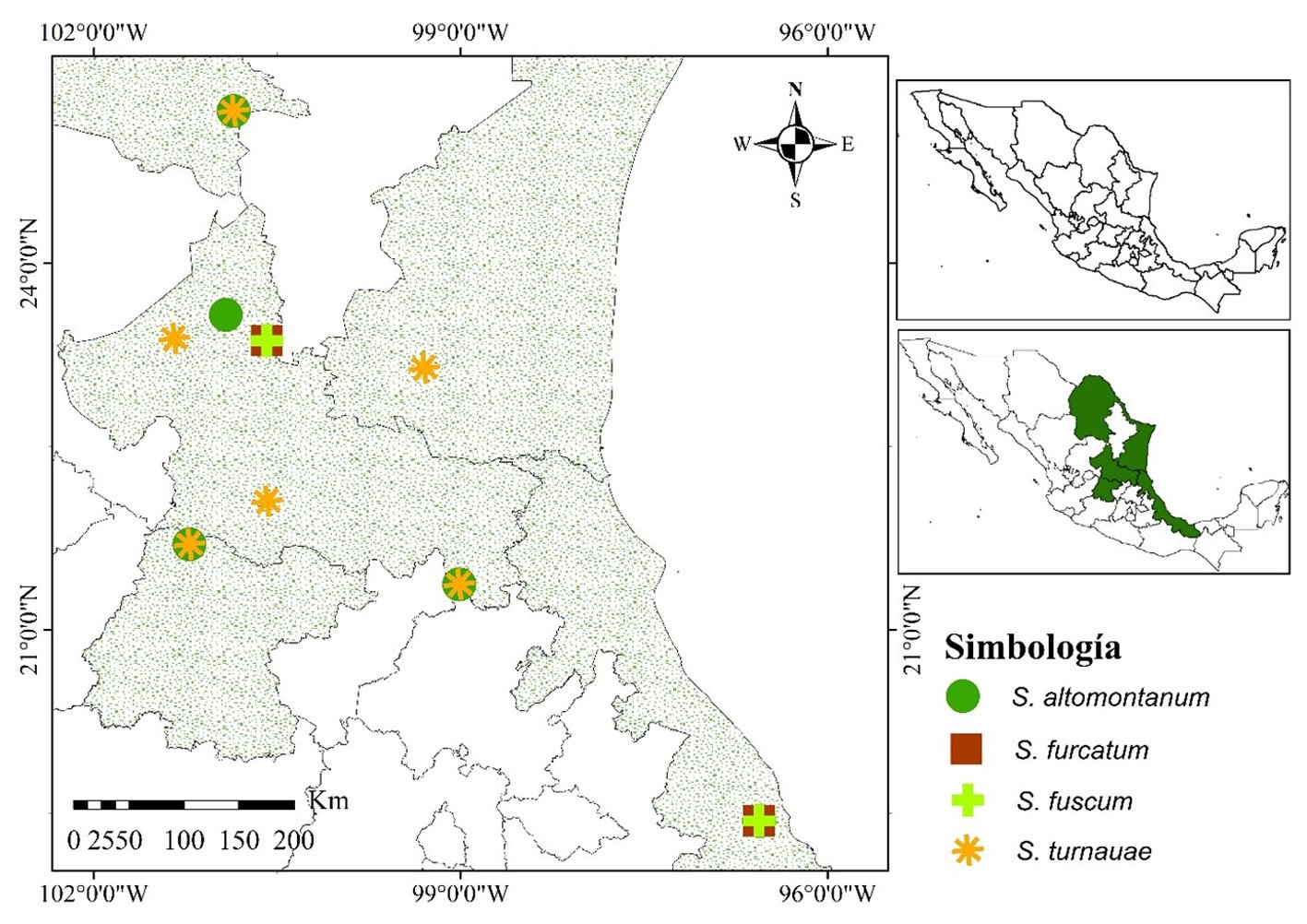
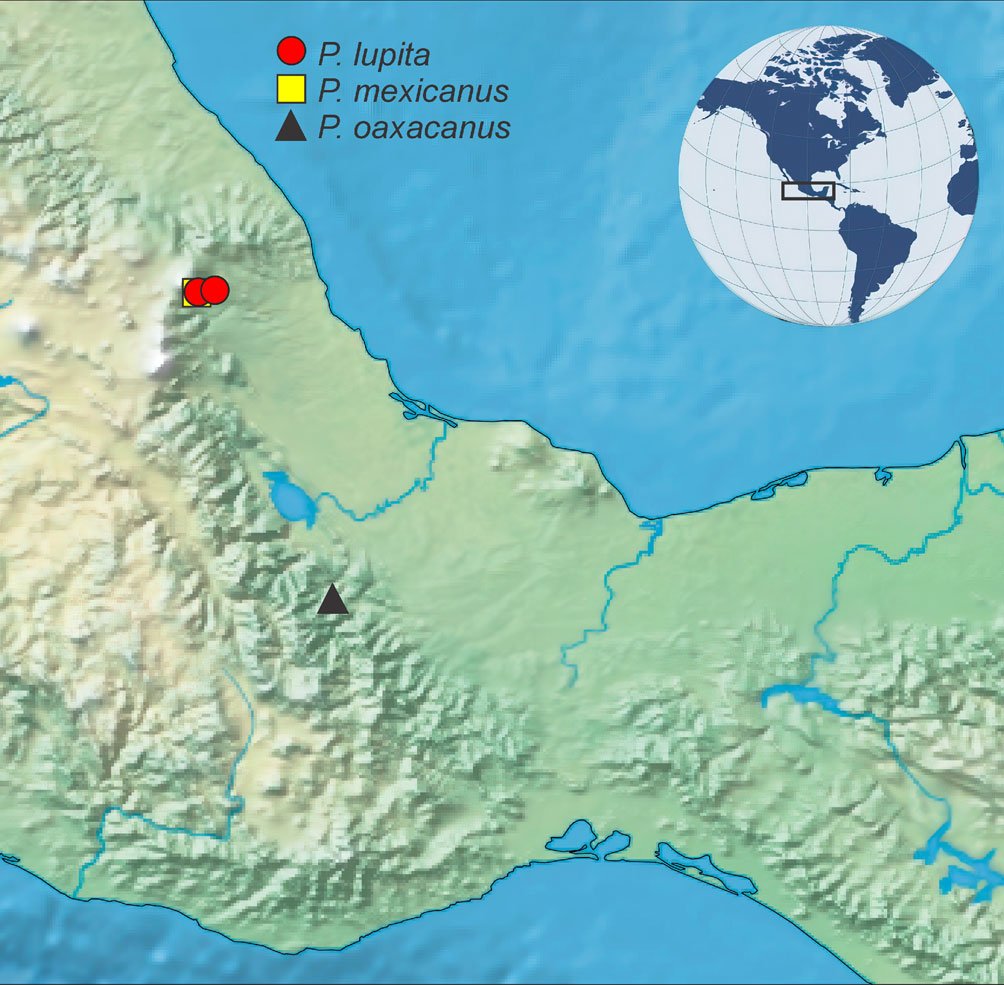
Holotype: 1 ♀, Mexico. Sinaloa: Venvidio, 2-6.VIII.1918, type # 8988 (ANSP). Allotype: 1 ♀, Sinaloa: Los Mochis, 25.XII.1918 (ANSP). Additional material examined: Mexico. Estado de México: 1 ♂, Nepantla, Finca Badoni, 18°59’0.8” N, 98°50’44.9” W, 2,020 m asl, 05.X.2023 (CEUAEM-MAN-135); 1 ♀, Malinalco, Chalma, 118°55’56.3” N, 99°26’11” W, 29.XI.1980 (IBUNAM: CNIN: MA909). Guerrero: 10 specimens (5 ♂, 5 ♀). Five ♂: Acahuizotla, 17°28’8.9” N, 99°25’10.3” W, 18.XI.1983 (IBUNAM: CNIN: MA644, MA645); Iguala, Km 3 desv. a Tepecoacuilco, 18°17’32” N, 99°28’58” W, 14.X.2006 (IBUNAM: CNIN: MA724); Iguala, Tuxpan, 2 km a Microondas, 18°23’34” N, 99°28’27” W, 14.X.2006 (IBUNAM: CNIN: MA813); Tecpan, La Laguna, Km 15 San Luis, 17°13’14.9” N, 100°37’9.3” W, 1,425 m asl, 20.VII.2005 (IBUNAM: CNIN: MA816). Five ♀: Atenango del Río, Km 36 Huitzuco-Atenango, 18°8’32” N, 99°7’49” W, 200 m asl, 14.X.2006 (IBUNAM: CNIN: MA823); Guerrero, 22.IX.2022 (CEUAEM-MAN-159, CEUAEM-MAN-160); Guerrero, 2024 (CEUAEM-MAN-161, CEUAEM-MAN-162). Jalisco: 13 specimens (11 ♂, 2 ♀). Eleven ♂: La Huerta, Estación de Biología Chamela (UNAM), 19°29’55.2” N, 105°2’39.5” W, 01.VIII.1981 (IBUNAM: CNIN: MA426); 10.V.1980 (IBUNAM: CNIN: MA802, MA804); 24.V.1980 (IBUNAM: CNIN: MA803); 10.VIII.1981 (IBUNAM: CNIN: MA805-MA810); 01.IV.1981 (IBUNAM: CNIN: MA811). Two ♀: La Huerta, Estación de Biología Chamela (UNAM), 19°29’55.2” N, 105°2’39.5” W, 14.X.1990 (IBUNAM: CNIN: MA723); 08.VIII.1975 (IBUNAM: CNIN: MA1560). Morelos: 23 specimens (12 ♂, 11 ♀). 12 ♂: Jojutla, EESJ, 18°36’49.8” N, 99°13’50.2” W, 957 m asl: 08.III.2022 (CEUAEM-MAN-013); 07.X.2018 (CEUAEM-MAN-014); 21.XI.2018 (CEUAEM-MAN-015); 20.XI.2019 (CEUAEM-MAN-021); Jojutla, 18°36’41.6” N, 99°10’44.7” W, 894 m asl, 21.X.2023 (CEUAEM-MAN-140); Morelos, 18°39’3.9” N, 99°2’20.5” W, 16.XI.1972 (IBUNAM: CNIN: MA519, MA520); Tepalcingo, Sierra de Huautla, 18°32’31.3” N, 98°56’9.4” W, 12.VI.2016 (IBUNAM: CNIN: MA943-MA946). Eleven ♀: Zacatepec, Galeana, 18°38’24.1” N, 99°12’39.6” W, 934 m asl, 22.IX.2019 (CEUAEM-MAN-017); Jojutla, EESJ, 18°36’49.8” N, 99°13’50.2” W, 957 m asl: 28.XI.2019 (CEUAEM-MAN-018); 22.XI.2019 (CEUAEM-MAN-019); 25.IX.2020 (CEUAEM-MAN-109); 13.XI.2020 (CEUAEM-MAN-110); 04.X.2023 (CEUAEM-MAN-129); Tehuixtla, 18°33’16.8” N, 99°16’12.8” W, 26.XII.2019 (CEUAEM-MAN-072); Yautepec, 18°53’12.8” N, 99°3’47.8” W, 1,216 m asl, 17.X.2023 (CEUAEM-MAN-138); Jojutla, 18°36’41.6” N, 99°10’44.7” W, 894 m asl, 27.X.2023 (CEUAEM-MAN-139); Cuautla, 18°48’59.4” N, 98°56’56” W, XI.1954 (IBUNAM: CNIN: MA905); Morelos, 18°39’3.9” N, 99°2’20.5” W, IV.1981 (IBUNAM: CNIN: MA906). Nayarit: 1 ♂, Bahía de Banderas, San Rafael, Arroyo de la Virgen, P. H. Aguamilpa, 20°50’19.5” N, 105°16’44.4” W, 734 m asl, 01.XI.1991 (IBUNAM: CNIN: MA824).
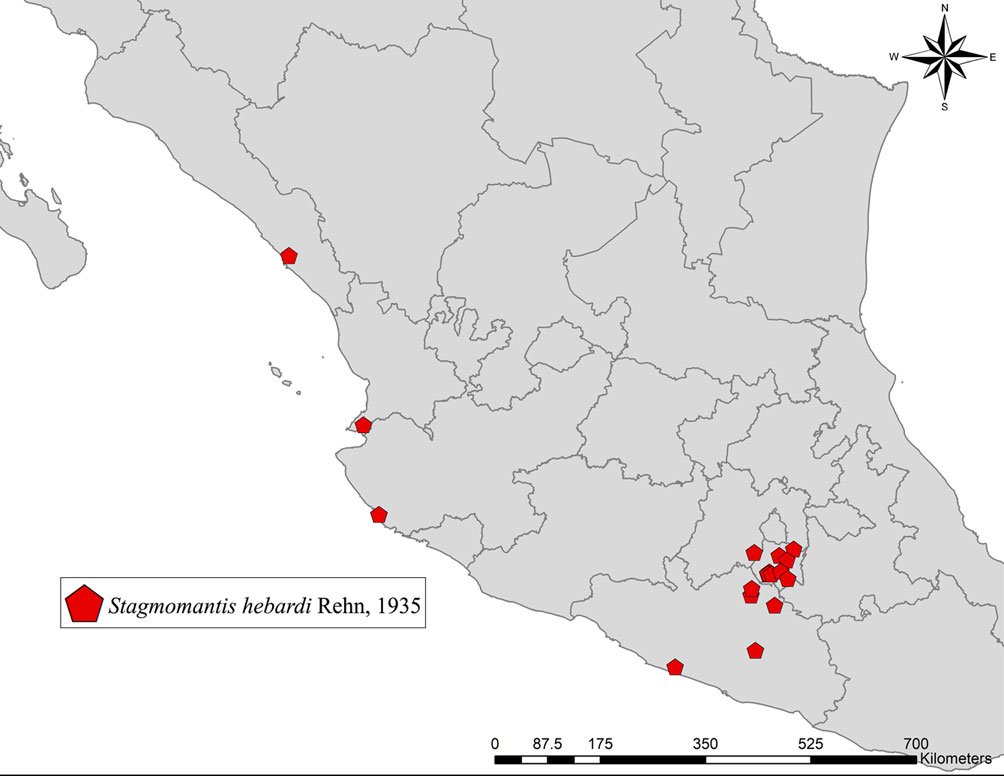
Geographic distribution: Stagmomantis hebardi is distributed in the states of Estado de México, Guerrero, Jalisco, Morelos, Nayarit, and Sinaloa (Fig. 12).
Remarks
Taxonomic delimitation within Stagmomantis based on male morphology has been a subject of study for decades. Notably, Saussure and Zehntner (1894) and Rehn (1935) considered the males of S. carolina and S. tolteca to be indistinguishable, grouping them within the “carolina group,” which currently corresponds to the subgenus Stagmomantis.
Morphological differences between the female originally described by Rehn (1935) and the females analyzed in this study are summarized in Table 4. Key distinctions include: a) the pronotum is proportionately wider in Rehn’s female compared to the slender pronotum in our specimens; b) wing shape and coloration differ significantly; Rehn’s specimen has wider forewings with a small, oval, light-colored stigma lacking a surrounding patch, whereas our specimens exhibit a stigma covered by a dark patch and surrounded by a white area covering nearly the proximal half of the wing; c) the abdomen in Rehn’s female is widely oval, whereas it is subfusiform (widening at the distal segments) in the females observed in this study; and d) the lobe-like structure between the 5th and 6th abdominal tergites, observed in all examined specimens, appears to be a diagnostic feature and a potential autapomorphy for this species.
Rehn (1935)
Present study
Pronotum wide
Pronotum slender
Forewings solid light colored
Forewings with different combinations of colors
Forewing’s stigma not covered by a colored patch
Forewing’s stigma covered by a white and dark patch
Forewings wider
Forewings slender
Abdomen wider almost oval
Abdomen subfusiform
Abdomen without lobe-like structure
Abdomen with lobe-like structure
Table 4. Comparison between the female of Stagmomantis hebardi in Rehn (1935) and the female specimens examined in this study.
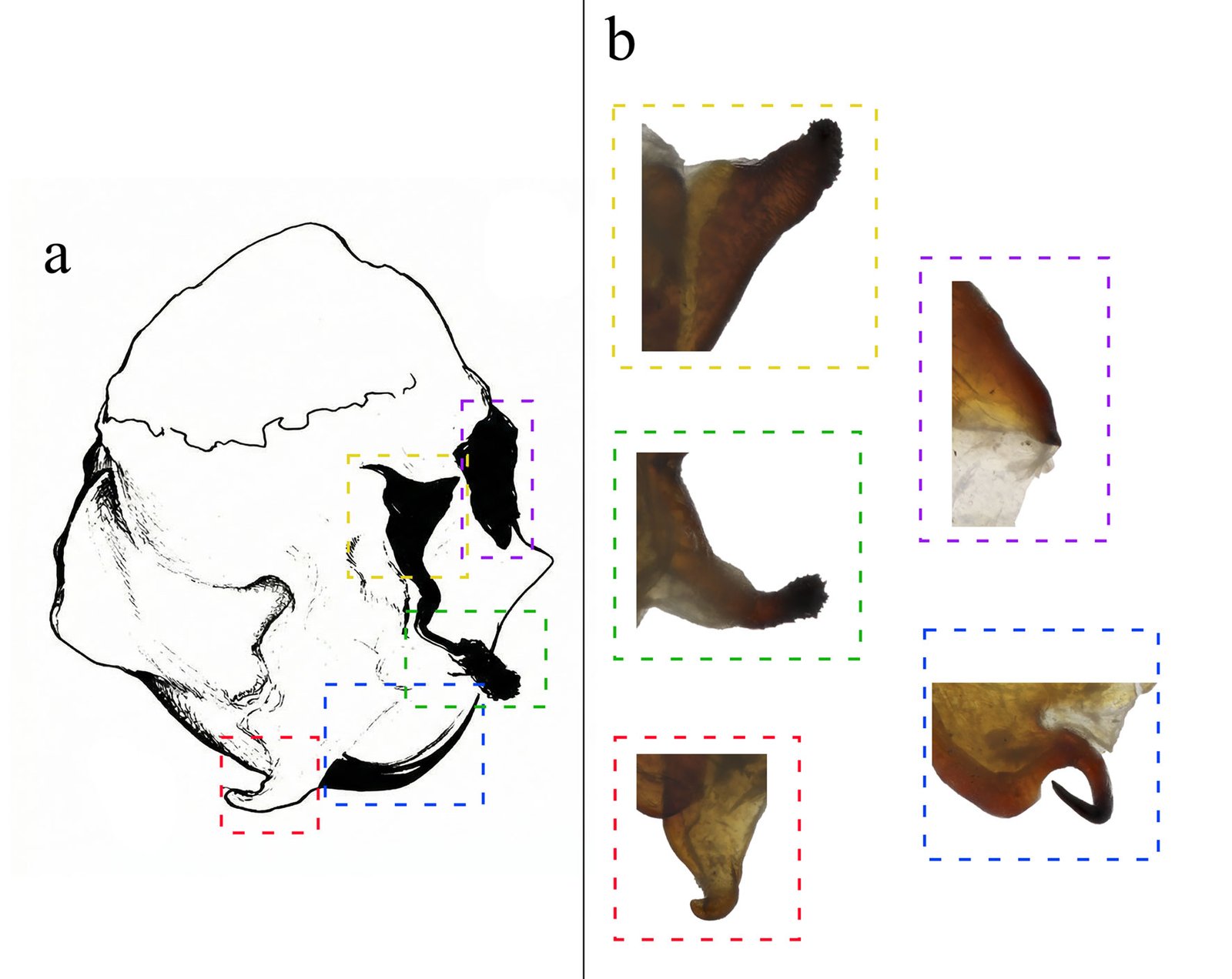
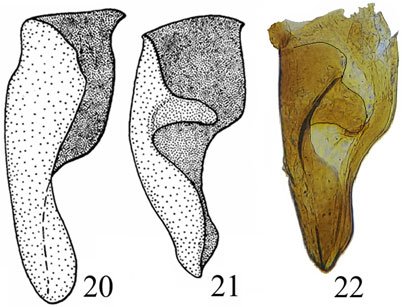
Although Stagmomantis tolteca and S. hebardi are morphologically similar, significant differences in male genitalia provide reliable diagnostic characters. As noted by Arteaga-Blanco et al. (2016), the posterior process of the ventral phallomere (pda) in S. tolteca is strongly sclerotized, wide at the base, tapering distally into a spine, and directed sinistrally (Fig. 13a, blue). Conversely, in S. hebardi, the pda is corkscrew-shaped, twisted either sinistrally or dextrally (Figs. 1, 5, 13b, blue). Further comparisons revealed differences in other genitalic structures: the aafa in S. tolteca is triangular with an angulate apex (Fig. 13a, yellow), whereas it is rounded in S. hebardi (Figs. 5, 13b, yellow); the ssp in S. tolteca is more developed with a slightly angulate inferior margin (Fig. 13a, purple), compared to the small and triangular ssp in S. hebardi (Figs. 5, 13b, purple); finally, the pafa in S. tolteca is elongate and curved near the middle, with the apex directed inward (Fig. 13a, green), while the pafa of S. hebardi projects laterally, is less curved, and has a more rounded apex (Figs. 5, 13b, green). The paa is morphologically similar in both species (Fig. 13, red). Rehn’s (1935) illustrations of the male genitalia correspond more closely to the pda morphology described in the present study than to that illustrated in Arteaga-Blanco et al. (2016). Thus, the sclerotized structure of the pda serves as a robust diagnostic character to separate both species and supports the identification of our male specimens as S. hebardi, in accordance with Rehn (1935).
Figure 12. Geographic distribution of examined specimens of Stagmomantis hebardi in Mexico.Figure 13. Comparison of male genitalia. a) Stagmomantis tolteca, modified from Arteaga-Blanco et al. (2016); b) Stagmomantis hebardi. Color coding: blue = pda; green = pafa; purple = ssp; red = paa; yellow = aafa. Specimen imaged: CEUAEM-MAN-021. See Table 1 for abbreviations.
A remarkable feature observed in this study is the presence of a lobe-like structure on the female abdomen. While comparable structures are rare in Mantodea, a few instances have been documented. For example, Robinson and Robinson (1979) described a terminal abdominal structure in females of Acanthops falcata Stål, 1877, identifying it as a pheromone-releasing gland. Similarly, Schwarz and Glaw (2021) described a Y-shaped protrusible structure near the abdominal apex of Stenophylla lobivertex Lombardo, 2000, hypothesizing that it represents a pheromone-releasing gland more efficient than simpler structures performing the same function.
The precise function of the lobe-like structure identified in this study remains unknown; however, several hypotheses can be proposed. According to Robinson and Robinson (1979) and Schwarz and Glaw (2021), such structures may function as sex pheromone glands, as reported for A. falcata and S. lobivertex. However, we observed this structure in both sexes during the early developmental stages of S. hebardi, which suggests it may possess an alternative or additional function. Another hypothesis involves the release of prey-attracting pheromones (kairomones), similar to the aggressive mimicry observed in bolas spiders (Mastophora Holmberg, 1876), which mimic lepidopteran sex pheromones to capture prey (Yeargan & Quate, 1997). Finally, this structure might enhance crypsis, similar to strategies observed in the subfamily Vatinae. However, unlike Vatinae, which are characterized by fixed cuticular lobes on the legs or abdominal margins, the structure in S. hebardi is distinctively protrusible.
Acknowledgements
We thank Cristina Mayorga-Martínez for her support in reviewing the CNIN-IBUNAM; Michael R. Maxwell, Henrique M. Rodrigues, and Lohitashwa Garikipati for their comments on an earlier version of this work; Jason D. Weintraub for providing us high resolution images of the holotype and allotype of Stagmomantis hebardi. Aarón Emilio Vásquez Quintero was supported by a scholarship from the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti; CVU 1315459).
References
Agudelo-Rondón, A. A., & Chica, E. L. M. (2002). Mántidos. Introducción al conocimiento del orden Mantodea. Universidad Distrital Francisco José de Caldas, Bogotá, Colombia.
Agudelo-Rondón, A. A., Lombardo, F., & Jantsch, L. J. (2007). Checklist of the Neotropical mantids (Insecta, Dictyoptera, Mantodea). Biota Colombiana, 8, 105–158.
Anderson, K. (2020a). Revision of Stagmomantis Saussure, 1869. Soothsayer, Journal of Mantodea Research, 1, 9–18.
Anderson, K. (2020b). Revalidation of Stagmomantis (Stagmomantis) conspurcata (Serville, 1839). Soothsayer, Journal of Mantodea Research, 1, 1–20.
Anderson, K. (2021). A new species of Stagmomantis Saussure, 1869 from North America. Soothsayer, Journal of Mantodea Research, 2, 86–96.
Anderson, K. (2025). Mantodea mundi: comprehensive nomenclatural catalogue of Mantodea (September 2025 edition). On line resource available at: file:///C:/Users/joelv/ Downloads/MantodeaMundi09.25.pdf
Arteaga-Blanco, L. A., de la Parra-Guerra, A. C., & Martínez- Hernández, N. J. (2016). Descripciones taxonómicas de mántidos (Insecta: Mantodea) del Departamento del Atlántico, Colombia, con apuntes sobre su distribución. Boletín Científico Centro de Museos. Museo de Historia Natural, 20, 211–236. https://doi.org/10.17151/bccm.2016.20.1.16
Brannoch, S. K., Wieland, F., Rivera, J., Klass, K. D., Olivier, B., & Svenson G. J. (2017). Manual of praying mantis morphology, nomenclature, and practices (Insecta, Mantodea). Zookeys, 696, 1–100. https://doi.org/10.3897/zookeys.696.12542
Ehrmann, R. (2002). Mantodea: Gottesanbeterinnen der Welt. Natur und Tier-Verlag. Münster, German.
Ferraz, B. R., Souza-Dias, P. G. B., & Rivera, J. (2023). A hidden gem from northeastern Brazil: a new species of Metaphotina Piza, 1964 (Mantodea, Acontistidae) ecologically linked to the Caatinga. Zootaxa, 5343, 31–54. https://doi.org/10.11646/zootaxa.5343.1.2
Garikipati, L. (2024). Description and total ontogeny of the Sonoran tiger mantis, Stagmomantis clauseni sp. nov., and key to the subgenus Nigralora (Mantodea: Mantidae). Zootaxa, 5501, 079–107. https://doi.org/10.11646/zootaxa.5501.1.4
Hebard, M. (1923). Dermaptera and Orthoptera from the state of Sinaloa, Mexico. Transactions of the American Entomological Society (1890-), 48, 157–196. INEGI (Instituto Nacional de Estadística y Geografía). (2024). Área geoestadística estatal, escala: 1:250000. Instituto Nacional de Estadística y Geografía. México, Ciudad de México. Retrieved March 23, 2025 from: https://www.inegi.org.mx/app/biblioteca/ficha.html?upc=794551132173
Otte, D., & Spearman, L. (2005). Mantida species file. Catalog of the mantids of the world. Philadelphia: Association of Insect Diversity.
Patel, S., & Singh, R. (2016). Updated checklist and distribution of Mantidae (Mantodea: Insecta) of the world. International Journal of Research Studies in Zoology, 2, 17–54. https://doi.org/10.20431/2454-941X.0204003
Rehn, J. A. G. (1935). On certain Mexican and Central American species of Melliera and Stagmomantis (Orthoptera, Mantidae). Transactions of the American Entomological Society, 61, 317–329.
Robinson, H. M., & Robinson, B. (1979). By dawn’s early light: matutinal mating and sex attractants in a Neotropical mantid. Science, 205, 825–827. https://doi.org/10.1126/science.205.4408.825
Rodrigues, H. M., & Cancello, E. M. (2016). Taxonomic revision of Stagmatoptera Burmeister, 1838 (Mantodea: Mantidae, Stagmatopterinae). Zootaxa, 4183, 1–78. https://doi.org/10.11646/zootaxa.4183.1.1
Saussure, H., & Zehntner, L. (1894). Fam. Mantidae. In H. Saussure, & L. Zehntner, A. (Eds.), Biologia Centrali- Americana, Insecta. Orthoptera, Vol. 1 (pp. 123–197). Paris: L‘Imprimerie Nationale.
Schwarz, C. J., & Glaw, F. (2021). The luring mantid: protrusible pheromone glands in Stenophylla lobivertex (Mantodea: Acanthopidae). Journal of Orthoptera Research, 30, 31–33. https://doi.org/10.3897/jor.30.55274
Schwarz, C. J., & Roy, R. (2019). The systematics of Mantodea revisited: an updated classification incorporating multiple data sources (Insecta: Dictyoptera). Annales de La Société Entomologique de France, 55, 101–196. https://doi.org/10.1080/00379271.2018.1556567
Su, Y. N. (2016). A simple and quick method of displaying liquid- preserved morphological structures for microphotography. Zootaxa, 4208, 592–593. https://doi.org/10.11646/zootaxa.4208.6.6
Svenson, G. J., & Whiting, M. F. (2004). Phylogeny of Mantodea based on molecular data: evolution of a charismatic predator. Systematic Entomology, 29, 359–370.
Svenson, G. J., & Whiting, M. F. (2009). Reconstructing the origins of praying mantises (Dictyoptera, Mantodea): the roles of Gondwanan vicariance and morphological convergence. Cladistics, 25, 468–514. https://doi.org/10.1111/j.1096-0031.2009.00263.x
Terra, P. S. (1995). Revisão sistemática dos gêneros de louva-a- deus da região Neotropical (Mantodea). Revista Brasileira de Entomologia, 39, 13−94.
Varela-Hernández, F., Martínez-Luque, E. O., Vázquez- Franco, C. M., & Pedraza-Lara, C. (2022). Contribution to the knowledge of mantids (Insecta, Mantodea) in Central Mexico: a morphological and molecular approach. Southwestern Entomologist, 47, 27–43.
Wang, W., Wang, H., Huang, H., Zhao, Y., & Zhou, Z. (2022). Mitochondrial genomes of 10 Mantidae species and their phylogenetic implications. Archives of Insect Biochemistry and Physiology, 111, e21874. https://doi.org/10.1002/arch.21874
Xu, X. D., Guan, J. Y., Zhang, Z. Y., Cao, Y. R., Storey, K. B., Yu, D. N. et al. (2021). Novel tRNA gene rearrangements in the mitochondrial genomes of praying mantises (Mantodea: Mantidae): translocation, duplication and pseudogenization. International Journal of Biological Macromolecules, 185, 403–411. https://doi.org/10.1016/j.ijbiomac.2021.06.096
Yeargan, K. V., & Quate, L. W. (1997). Adult male bolas spiders retain juvenile hunting tactics. Oecologia, 112, 572–576. https://doi.org/10.1007/s004420050347
Revisión de los Elmidae (Coleoptera: Dryopoidea) de Chile
Marcos Ferrú a, b, *, William Shepard c
a Universidad Católica de la Santísima Concepción, Alonso de Ribera 2850, Concepción, Chile b Centro de Muestreo y Análisis Biológico, Luis Acevedo 456 Chiguayante Bíobío-Concepción, Chile c University of California, Essig Museum of Entomology, 1101 Valley Life Sciences Bldg., Berkeley, California, 94720 USA
Received: 12 August 2025; accepted: 17 December 2025
Abstract
The family Elmidae is one of several families of aquatic beetles in the superfamily Dryopoidea. The Elmidae occur throughout the world (except Antarctica), with approximately 150 genera and 1,500 species. Elmidae have been studied as part of the taxa used to assess water quality. There are no keys to identify larvae of Chilean species, and to identify adults it is necessary to remove the male genitalia. Research carried out in Chile has included Elmidae in descriptions, lists and reviews of aquatic beetles in general. Until now, there has been no review dedicated entirely to Chilean Elmidae. In Chile, after this review there are 2 subfamilies, 7 genera and 18 species currently recognized. The species Austrelmis woytkowskii (Hinton), which has been cited in error for Chile, is eliminated from the Chilean list and the species Austrelmis sp. 1 is included, as is Austrolimnius elatus Hinton. Expanded distributions of some species are included.
Keywords: Elmidae; Elminae; Larainae; Diversity; Distribution
Resumen
La familia Elmidae es una de varias familias de escarabajos acuáticos perteneciente a la superfamilia Dryopoidea. Los Elmidae se encuentran en todo el mundo (excepto en la Antártida), con aproximadamente 150 géneros y 1,500 especies. Los Elmidae han sido estudiados como parte de los taxones utilizados para evaluar la calidad del agua. No existen claves para identificar larvas de las especies chilenas y para identificar adultos es necesario extraer los genitales de los machos. Investigaciones realizadas en Chile han incluido a los Elmidae en descripciones, listas y revisiones de escarabajos acuáticos en general. Hasta ahora, no había una revisión dedicada completamente a los Elmidae chilenos. En Chile, después de esta revisión hay 2 subfamilias, 7 géneros y 18 especies actualmente reconocidas. La especie Austrelmis woytkowskii (Hinton), que ha sido citada por error para Chile se elimina de la lista chilena y se incluye la especie Austrelmis sp. 1, al igual que Austrolimnius elatus Hinton. Se incluyen la ampliación de distribuciones de algunas especies.
The family Elmidae (Coleoptera: Dryopoidea) colloquially known as riffle beetles is a group of globally distributed beetles (except for Antarctica) found in clean, permanent waters. Elmidae is divided into 2 subfamilies (Elminae and Larainae), with approximately 150 genera and 1,500 species Jäch et al. (2016) and more taxa are described every year. In Chile, the Elmidae have 2 large faunal divisions. This is a fauna with continental characteristics that shares genera, such as Heterelmis Sharp and Microcylloepus Hinton, with other South American countries. The other faunal area is the Southern Cone, which contains genera shared with New Zealand and Australia, or has genera only shared with Argentina, as is the case of the genera Hydora Anon. [Broun], Luchoelmis Spangler and Staines and Stethelmis Hinton. The different species of Elmidae have similar life cycles. Eggs are laid on submerged substrates such as stones or wood, with hatching occurring in approximately 2 weeks. Pupation lasts about 2 weeks. After about 2 weeks of pupation, the adults emerge, some fly to find new habitats before re-entering the water, while others just re-enter the stream from which they came. Adults can live approximately 10 years (Brown, 1974). Larvae and adults of Elmidae often co-occur in appropriate microhabitats, such as sand, gravel, leaf bundles, aquatic macrophytes, wood, and rocks in clear water, etc. The main requirement for Elmidae is uncontaminated water with high levels of dissolved oxygen, up to moderately polluted environments González-Córdoba et al. (2020). Most Elmidae consume fine particulate organic matter that they browse from the surface or find in the interstitial spaces between gravel particles. This material is usually algae, fungal hyphae, and crushed leaves or wood. The few species that feed on decaying and submerged wood take several years to develop due to the low levels of nutrients wood presents. Reviews of elmid biology are in Brown (1987) and Elliott (2008). Elmidae breathe with tracheal gills (larvae) or plastrons (adults). Both systems require relatively high levels of dissolved oxygen (Brown, 1987; Kodada et al., 2016). Elmidae have been found to be very susceptible to contaminants such as surfactants in soap, petroleum products, accordiong to Brown (1984), Elliott (2008), Lafuente et al. (2019) and anything that removes oxygen from the water. Therefore, their presence in a stream is a sign of uncontaminated water during the lifespan of the Elmidae. Thus, Elmidae are often targeted during water quality assessment programs, along with Ephemeroptera, Plecoptera and Trichoptera, all of which are also susceptible to contaminants (Fierro et al., 2012). When these studies are carried out, it is very important that the sampling considers the microhabitat requirements of the different species. The new global catalog of Elmidae has helped to understand generic and species distributions (Jäch et al., 2016). An identification key for adult genera of South American Elmidae has been developed by Manzo (2005) and an identification key for most larvae of South American Elmidae genera has been developed by Manzo and Archangelsky (2008), Pasos et al. (2018). These have been important scientific advances since Elmidae are an integral part of the ecology of streams and rivers and are often collected during sampling (Fierro et al., 2012). None the less despite its importance there are no keys to identify the larvae to species, and generally to identify the adults it is necessary to extract the male genitalia. The first Elmidae described in Chile are found in 2 articles by Germain (1854, 1892). This was followed by articles published by Champion (1918), Hinton (1941, 1945), Janssens (1957), Spangler and Brown (1981) and Spangler and Staines (2002). Some of the species described in these papers are now found in different genera. Other papers dealing with Chilean dryopoid beetles include authors like Lataste (1897 a, b), Dajos (1974), Solervicens (1974), Shepard (2016) and Shepard and Barr (2023). Nonetheless, the information is scattered and Elmidae continues to be a relatively unknown group. This study will facilitate further efforts on Chilean Elmidae by bringing together information and providing new distribution data.
Materials and methods
The collection of elmid larvae and adults usually involves aquatic nets that are placed downstream of the sampling area where the substrates are disturbed. Elmidae then drifts in the current and into the net. The specimens are collected from the net and preserved in 70% or 95% ethyl alcohol. Some of the net contents collected by WDS were entirely preserved in 95% alcohol bottles and then processed in the laboratory. This makes it easier to collect smaller adults (such as Austrolimnius) and younger larvae. After collecting at each site, ecological characteristics were recorded on field data sheets. Collection locality maps were produced with QGIS and used all known localities, published and new. Locality data are presented primarily as they occur on the specimen labels. A forward slash “/” indicates separate lines in a label and a double forward slash “//” indicates separate labels. Information included by “( )” is below the locality labels. Additional data provided here, but not in the labels, is indicated by brackets “[ ]”. The “WDS-A- ____” numbers refer to WDS data sheets. Collection locality maps were made using ArcGIS and used all known localities, published and new. Additional records of the specimens were obtained from the collections: National Museum of History of Chile (Santiago) and the Museum of the Department of Zoology of the University of Concepción, as well as from published literature on the Elmidae. Abbreviations cited in the text include the following: National Museum of Natural History, Washington, DC, USA (USNM); Natural History Museum, London, England (NHML); Royal Belgian Institute of Natural Sciences, Brusseels, Belgium (RBINS); William D. Shepard (WDS).
Results
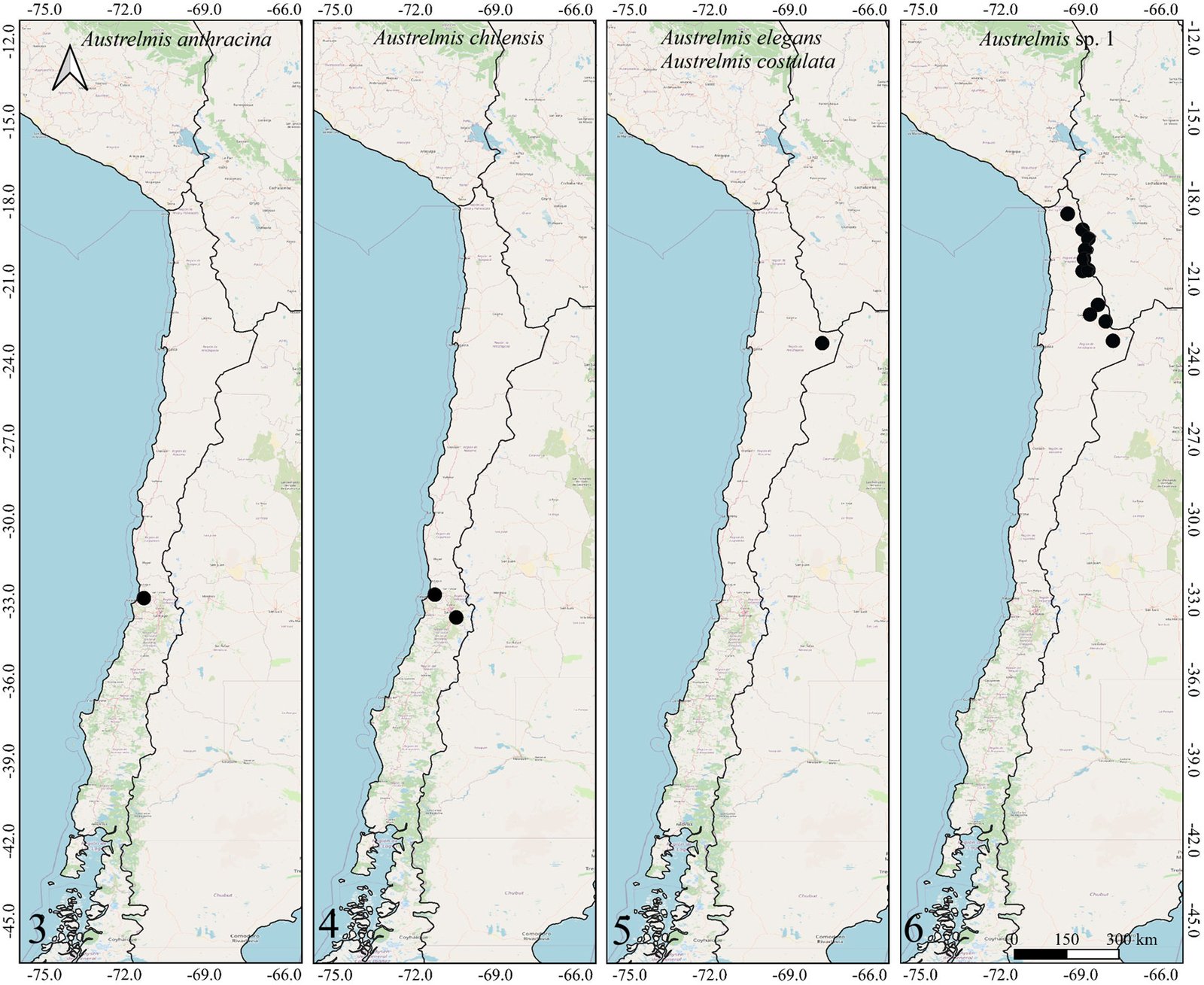
As a result of this work, we can consider the diversity of Elmidae to include 18 species, 7 genera and 2 subfamilies. The species Austrelmis woytkowskii (Hinton, 1937) is eliminated because the analysis of samples collected from various streams in northern Chile shows no evidence of its presence. The species Heterelmis trivialis (Germain, 1892) is also eliminated. One of the reasons for this decision is the loss of the type material. Another reason is the fragility of the stream where it was cited; this stream is small and urbanized and tends to have a considerable decrease in its water flow in summer. This species was searched for at the type locality on 2 occasions, thus corroborating its presence in Chile, but it has not been found or reported since its description. In these samplings, the other species described by Germain and cited for the area were found. In this work, the species Austrolimnius elatus, eliminated by Elgueta and Arriagada (1989), is reestablished; this is supported by the type material found. Finally, it is important to mention that despite the limited progress in species richness shown in this work, there are numerous new localities for the different species present in Chile. A list of the Elmidae known from Chile is provided (Table 1).
Key to the genera of adult Elmidae known from Chile extracted from Manzo and Archangelsky (2008) and Passos et al. (2018). 1 Body densely pubescent, without plastron; transverse procoxae, with exposed trochantins; adults fly easily from water or net — Larainae Hydora (Fig. 19). 1’ Body not pubescent, with plastron; procoxa globosa, with unexposed trochantins; adults do not fly from water or net — Elminae (2). 2 Thorax without sublateral carinae (Figs. 27, 30, 32) — 3. 2’ Thorax with sublateral carinae — 4. 3 Tarsal claws with basal teeth (Fig. 31) — Stethelmis. 3’ Tarsal claws without basal teeth — Luchoelmis. 4 Epipleura with 1 or 2 longitudinal lines of granules — Austrolimnius. 4’ Epipleura without longitudinal lines of granules — 5. 5 Pronotum with complete deep transverse groove (Fig. 29) — Neoelmis. 5’ Pronotum without deep transverse groove complete — 6. 6 Pronotum with circular anterior depression, with V-shaped carina in ½ basal, with sublateral elevation (Fig. 28) — Microcylloepus. 6’ Pronotum without depressions; sublateral carina restricted to anterior half (Figs. 25, 26) — Austrelmis.
#
Species
Original generic placement
Author
Endemic
Subfamily Elminae Curtis, 1830
1
Austrelmis anthracina
Elmis
(Germain, 1892)
Yes
2
Austrelmis chilensis
Elmis
(Germain, 1854)
Yes
3
Austrelmis costulata
Macrelmis
(Janssens, 1957)
Yes
4
Austrelmis elegans
Macrelmis
(Hinton, 1937)
No
5
Austrelmis sp.
Macrelmis
(Janssens, 1957)
Yes
6
Austrolimnius chiloensis
Elmis
(Champion, 1918)
Yes
7
Austrolimnius elatus
Austrolimnius
Hinton, 1941
No
8
Austrolimnius nycteloides
Elmis
(Germain, 1854)
No
9
Luchoelmis aequalis
Luchoelmis
Spangler & Staines 2002
Yes
10
Luchoelmis cekalovici
Luchoelmis
Spangler & Staines 2002
No
11
Luchoelmis magallanensis
Luchoelmis
Spangler & Staines 2002
Yes
12
Luchoelmis penai
Luchoelmis
Spangler & Staines 2002
Yes
13
Microcylloepus chilensis
Microcylloepus
Janssens, 1957
Yes
14
Neoelmis scissicollis
Elmis
(Germain, 1892)
Yes
15
Stethelmis chilensis
Stethelmis
Hinton 1945
Yes
16
Stethelmis kaszabi
Stethelmis
Hinton, 1970
No
Subfamily Larainae LeConte, 1861
17
Hydora annectens
Hydora
Spangler & Brown, 1981
No
18
Hydora lenta
Hydora
Spangler & Brown, 1981
Yes
Table 1. Elmidae of Chile and endemicity status.
Generic reviews
This section is reproduced from the article PDF; the genus-by-genus accounts (with new distributional records and material examined) are available in full in the linked PDF.
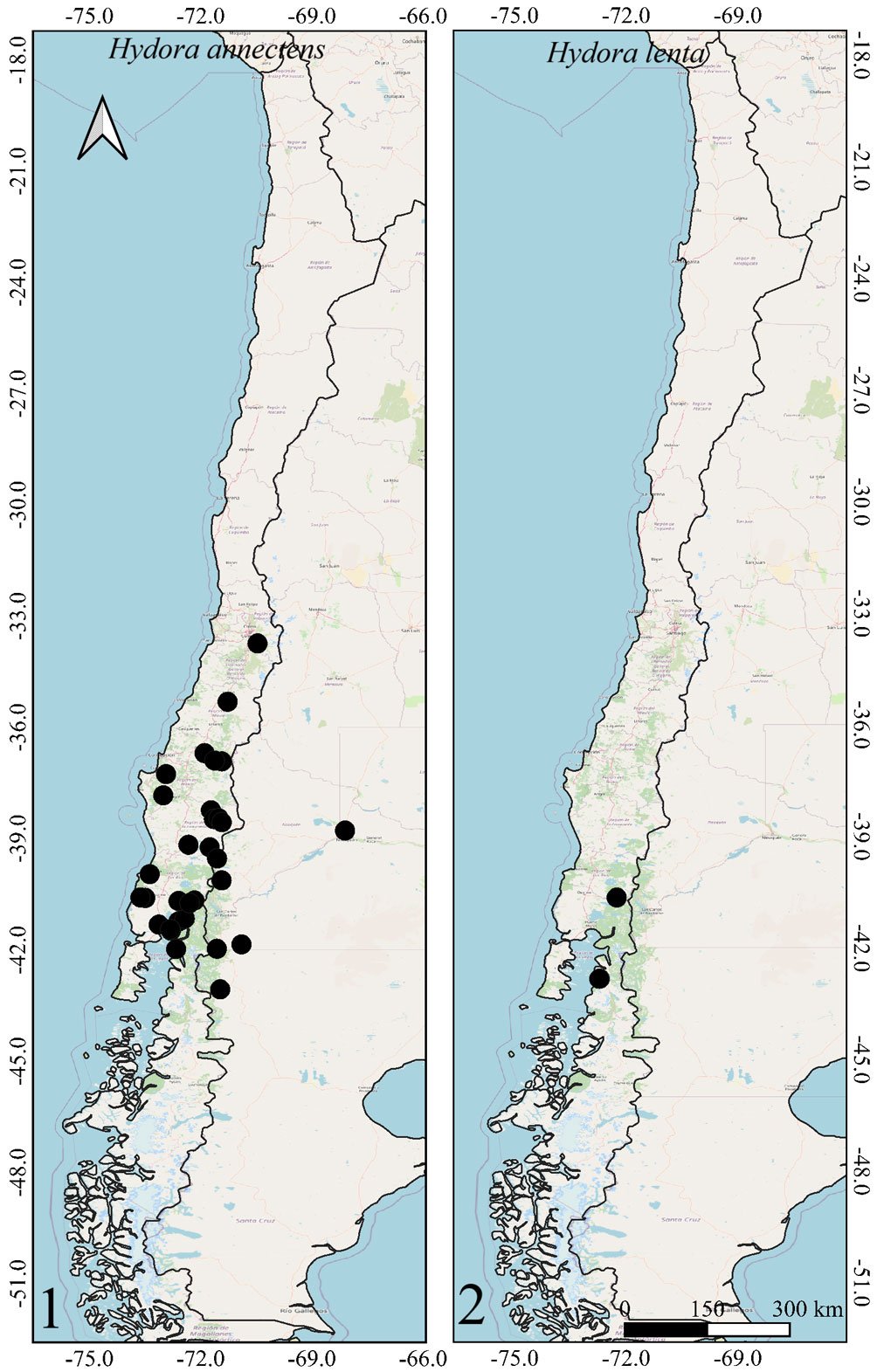
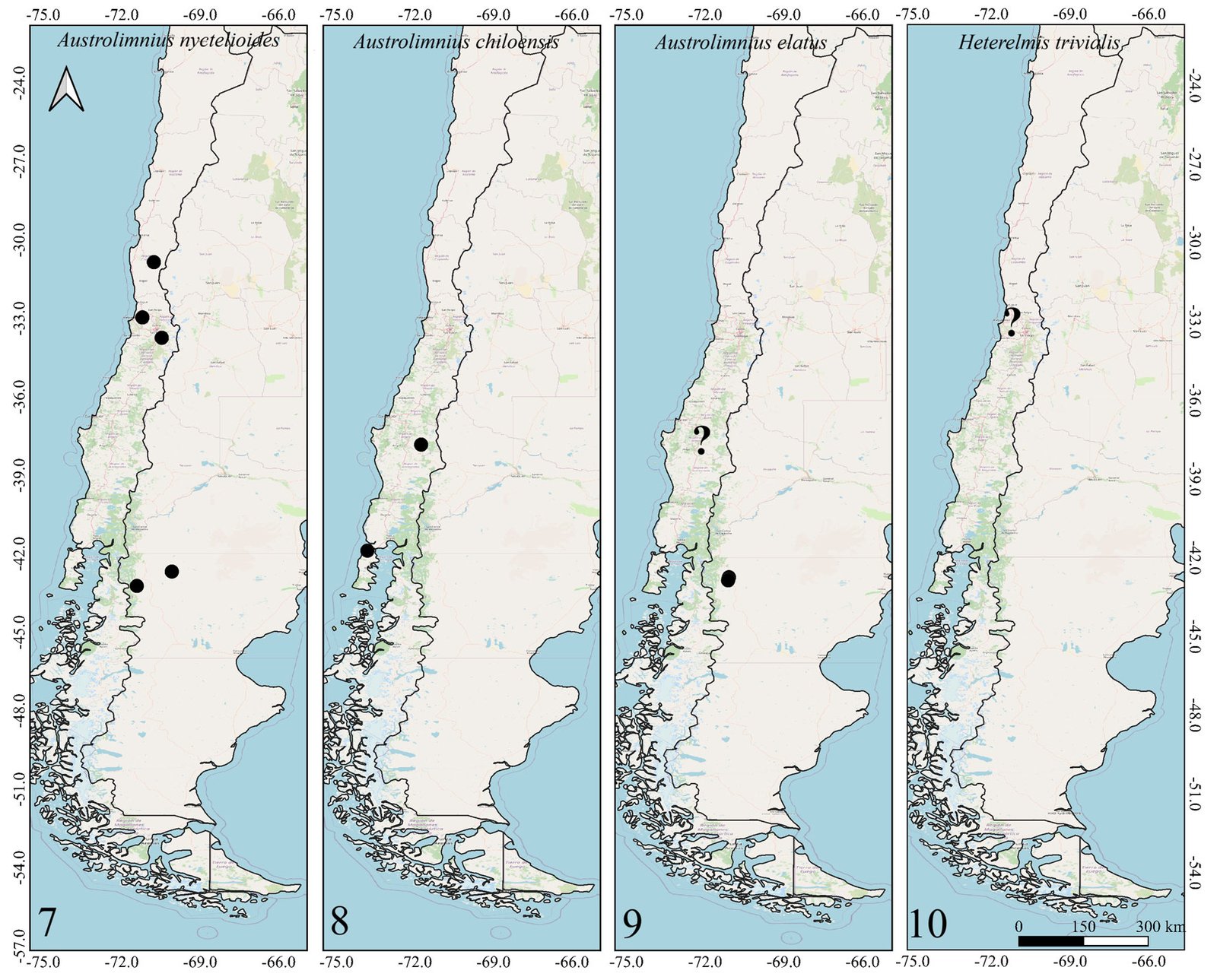
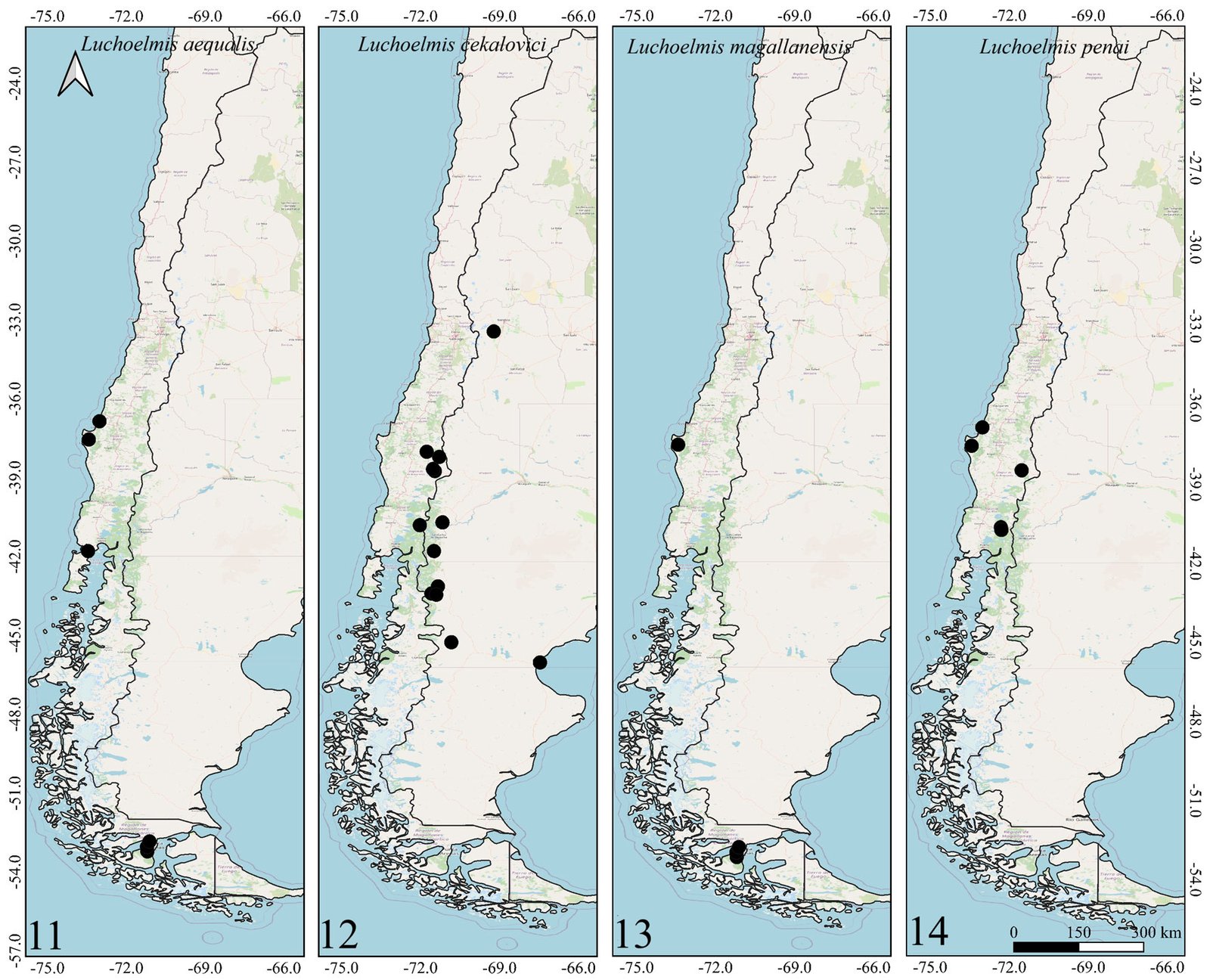
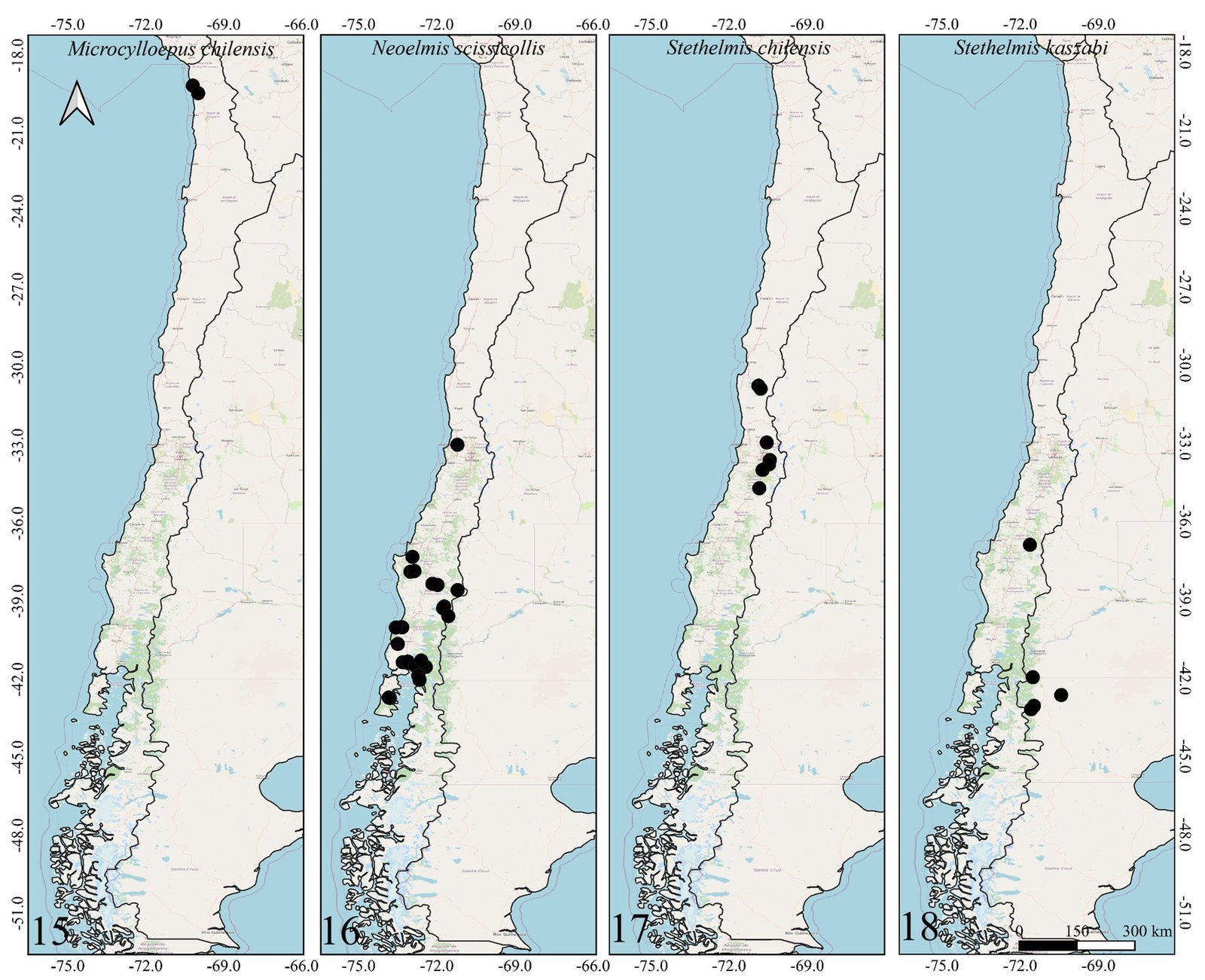
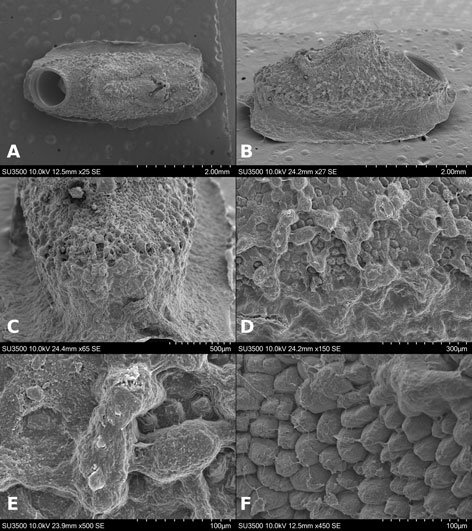
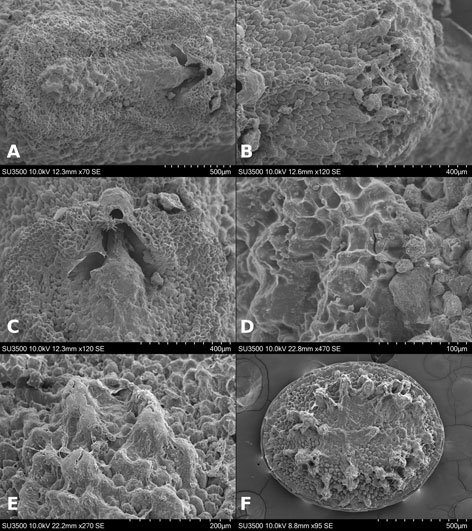
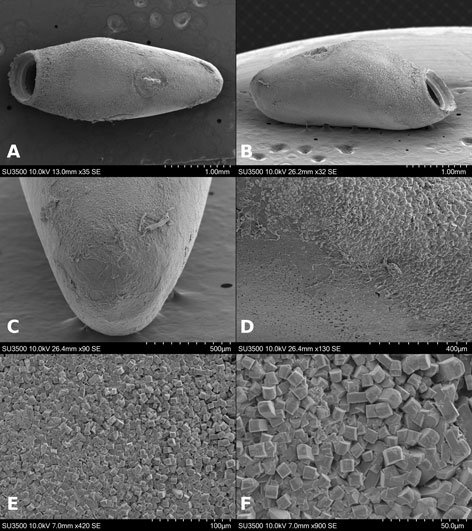
Figures 1-2. Distributions of the species belonging to the genus Hydora in Chile and neighboring countries.Figures 3-6. Distributions of Austrelmis species in Chile.Figures 7-10. 7-9, Distribution of the species of Austrolimnius; 10, Heterelmis in Chile and neighboring countries.Figures 11-14. Distributions of the species belonging to the genus Luchoelmis in Chile and neighboring countries.Figures 15-18. Distribution of the species belonging to the genera Microcylloepus, Neoelmis and Stethelmis in Chile and neighboring countries.Figure 19. Dorsal habitus of the type specimen of Hydora annectens and labels.Figures 20-22. Ventral view of paramero of Austrelmis. 2. A. woytkowskii Hinton; 3. A. consors Hinton; 4. A. sp. 1, where the paramere of the species collected in the highlands of Chile can be seen.Figure 23. Dorsal habitus of the type specimen of Austrelmis costulata and labels.Figure 24. Dorsal habitus of type specimen of Austrelmis elegans and labels.Figure 25. Dorsal habitus of the type specimen of Austrolimnius elatus and labels.Figure 26. Dorsal habitus of the type specimen of Austrolimnius chiloensis and labels.Figure 27. Dorsal habitus of Luchoelmis sp. (Photography: William Shepard).Figure 28. Dorsal habitus of the type specimen of Microcylloepus chilensis and labels (left). Dorsal thoracic habits delimiting the elevation in the posterior fraction of the pronotal disc depression of Microcylloepus chilensis (right).Figure 29. Thorax of Neoelmis scissicollis with carinae on pronotum (Photography: Marcos Ferrú).Figures 30-31. Thorax of Stethelmis sp., without carinae on pronotum; posterior leg of Stethelmis sp. (Photography: Marcos Ferrú).Figure 32. Dorsal habitus of the type specimen of Stethelmis chilensis and labels.
Discussion
The Chilean Elmidae include 2 subfamilies, 7 genera and 17 species. However, many additional species are known but not yet described. Many geographic areas remain unexplored, so additional species are likely to be found. Endemism is relatively high in the Chilean Elmidae, 82.3% (14 of 17 species) of the species are endemic to Chile. The only Chilean Elmidae not endemic to Chile are Austrolimnius elatus, A. nycteloides, and Hydora annectens.
The number of genera and species present in the South American countries on the Pacific slope decreases from north to south (Ecuador to Chile) (Table 2). Partly this is because the northernmost areas are closer to the equator, where temperatures are more favorable to elmid evolution. Also, partly this is due to Ecuador and Peru having areas located on the east front of the Andes near to the Amazon where the evolution of the Elmidae has gone in a different direction (Shepard unpublished data). Chile lacks an Amazonian component.
In Chile, the elmid fauna has been negatively impacted by coastal and Andean orogenies, especially in the north. This is mainly due to the isolation and desertification that characterizes this area (Shepard unpublished data). The only 2 genera present there are Austrelmis and Microcylloepus. Microcylloepus elsewhere has been able to maintain populations in desert environments where small springs or seeps occur, so it is not surprising that it would be found here, while Austrelmis is a very typical inhabitant of high mountain habitats throughout the Andes.
The Southern Cone area of Chile has genera that were thought to be restricted to this area, such as Austrolimnius, Luchoelmis, and Stethelmis. However, Austrolimnius and Stethelmis have now been collected somewhat farther north, Coquimbo in the case of Austrolimnius and Atacama in the case of Stethelmis, nonetheless their diversity remains in the Southern Cone area.
Two collection attempts in the type locality of Heterelmis trivialis failed to find extant populations. It is not known why this population might have disappeared. Likewise, attempts to find populations of Austrelmis woytkowskii were unsuccessful. These findings reinforce the necessity to amplify sampling focused on aquatic coleoptera and Elmidae specifically, in order to determine if these populations are truly extinct or only minimally abundant.
Country
Area (km²)
Number of genera and species
Reference
Colombia
916,445
31 genera and 65 especies
Hincapié-Montoya, 2023
Ecuador
276,841
19 genera and 59 especies
Monte and Mascani, 2012
Perú
1,285,216
17 genera and 51 especies
Shepard and Chaboo, 2015
Chile
756,950
7 genera and 18 especies
This review
Bolivia
1,098,581
12 genera and 33 especies
Manzo and Moya, 2010
Argentina
2,780,400
14 genera and 21 especies
Manzo and Archangelsky, 2014
Table 2. Number of genera and species of Elmidae and the area of the countries on the Pacific slope of South America (from north to south) and Bolivia and Argentina.
Acknowledgements
We thank all the people who made this work possible: Walter Sielfeld, for the support on the ground and collection of specimens; Mario Elgueta (National Museum of Natural History); Margarita Ruiz de Gamboa (CEMABIO); Pablo Fierro (Universidad del Valdivia) for his help, contribution of copies and ideas; the specialists Verónica Manzo (University of Tucumán, Argentina) and María Passos (Federal University of the State of Rio de Janeiro, Brazil) for their bibliography. And, finally, the Laboratory of Water Quality Bioindicators EULA-Chile, for their contribution in specimens and land. Additionally, we thank the following people for providing photographs of type specimens and labels under their care: Pol Limbourg (RBINS), Keita Matsumoto (NHML) and Charyn Micheli (USNM).
References
Archangelsky, M., & Brand, C. (2014). A new species of Luchoelmis Spangler & Staines (Coleoptera: Elmidae) from Argentina and its probable larva. Zootaxa, 3779, 563–572. https://doi.org/10.11646/zootaxa.3779.5.6
Archangelsky, M., & Manzo, V. (2006). The larva of Hydora annectens Spangler and Brown (Coleoptera: Elmidae, Larainae) and a key to New World Larainae larvae. Zootaxa, 1204, 41–52. https://doi.org/10.11646/zootaxa.1204.1.4
Archangelsky, M., & Manzo, V. (2007). Descripción de las larvas maduras de los géneros Stethelmis Hinton y Luchoelmis Spangler and Staines (Insecta: Coleoptera, Elmidae). Revista del Museo Argentino de Ciencias Naturales, 9, 79–87.
Archangelsky, M., Martínez-Román, N. R., & Manzo, V. (2016). Description of mature larvae of the genus Austrelmis Brown from Argentina (Insecta: Coleoptera, Elmidae). Annales de Limnologie, 52, 343–353. https://doi.org/10.1051/limn/2016024
Artigas, J. (1963). Descripción de la larva de un Psefenido chileno (Coleoptera-Psephenidae). Gayana Zoología, 8, 3–9.
Brown, H. P. (1974). Survival records for elmid beetles, with notes on laboratory rearing of various dryopoids (Coleoptera). Entomological News, 84, 278–284.
Brown, H. P. (1984). Neotropical Dryopoids, III. Major nomenclatural changes affecting Elsianus Sharp and Macrelmis motschulsky, with checklists of species (Coleoptera: Elmidae: Elminae). The Coleopterists Bulletin, 38, 121–129.
Champion, G. C. (1918). Notes on various South American Coleoptera collected by Charles Darwin during the voyage of the “Beagle” with descriptions of new genera and species. Entomologists’ Monthly Magazine, 54, 43–54.
Dajos, R. (1973). Description du Coléoptere Chiloea chilensis, n. g., n. sp., type d’une nouvelle familie: Chiloeidae. Annals de la Societe Etomologique de France (N. S.), 9, 173–179. https://doi.org/10.1080/21686351.1973.12278133
Elgueta, M., & Arriagada, G. (1989). Estado actual del conocimiento de los coleópteros de Chile (Insecta: Coleoptera). Revista Chilena de Entomología, 17, 5–60.
Elliott, J. M. (2008). The ecology of riffle beetles (Coleoptera: Elmidae). Freshwater Reviews, 1, 189–203. https://doi.org/10.1608/FRJ-1.2.4
Emmerich, D., Manzo, V., & Morelli, E. (2025). Lista comentada y actualizada de los escarabajos acuáticos de la familia Elmidae (Insecta: Coleoptera) de Uruguay. Revista de la Sociedad Entomológica Argentina, 84, e0210. https://doi.org/10.25085/rsea.840210
Ferrú, M., & Elgueta, M. (2011) Lista de coleópteros (Insecta: Coleoptera) de las regiones de Arica y Parinacota y de Tarapacá, Chile. Boletín del Museo Nacional de Historia Natural, Chile, 60, 9–61. https://doi.org/10.54830/bmnhn.v60.2011.178
Fierro, P., Bertrán, C., Mercado, M., Peña-Cortés, F., Tapia, J., Hauenstein, E. et al. (2012). Benthic macroinvertebrate assemblages as indicators of water quality applying a modified biotic index in a spatio-seasonal context in a coastal basin of southern Chile. Revista de Biología Marina y Oceanografia, 47, 23–33. https://doi.org/10.4067/S0718-19572012000100003
Fierro, P., Ferrú, M., & Lara, C. (2020). Effects of forest conversion on the aquatic Coleoptera assemblage in Mediterranean-climate streams. Ecological Indicators, 111, 106043. https://doi.org/10.1016/j.ecolind.2019.106043
Germain, P. (1854). Descripción de 21 especies nuevas de coleópteros de Chile. Anales de la Universidad de Chile, 1854, 324–336.
Germain, P. (1892). Notes sur les coléopteros du Chili. Actes de la Société Scientifique du Chile, 2, 241–261.
González-Córdoba, M., Zúñiga, M. C., Giraldo, L. P., Ramírez, Y. P., & Chará, J. (2020). Sensibilidad de Elmidae (Insecta: Coleoptera) a la perturbación del hábitat y la calidad fisicoquímica del agua en ambientes lóticos de los Andes colombianos. Revista de Biología Tropical, 68, 601–622. https://doi.org/10.15517/rbt.v68i2.36702
Hincapié-Montoya, D. M. (2023). Nuevos registros para Elmidae (Coleoptera: Byrrhoidea) en Colombia. Acta Biológica Colombiana, 28, 345–349. https://doi.org/10.15446/abc.v28n2.103984
Hinton, H. E. (1940). The Percy Sladen Trust Expedition to Lake Titicaca in 1937. VII. The Peruvian and Bolivian species of Macrelmis Motsch. (Coleoptera, Elmidae). The Transactions of the Linnean Society of London (Ser. 3), 1, 117–147.
Hinton, H. E. (1941). A synopsis of the American species of Austrolimnius Carter (Coleoptera, Elmidae). Entomologists’ Monthly Magazine, 77, 156–163.
Hinton, H. E. (1945). Stethelmis chilensis, new genus and species of Elmidae from Chile (Coleoptera). Proceedings of the Royal Entomological Society of London (B), 14, 73–76. https://doi.org/10.1111/j.1365-3113.1945.tb00022.x
Jäch, M. A., Kodada, J., Brojer, M., Shepard, W. D., & Čiampor, Jr. F. (2016). World catalogue of insects, Vol. 14. Coleoptera: Elmidae and Protelmidae. Leiden, The Netherlands: Koninklijke Brill.
Janssens, E. (1957). Contribution à l’étude des coléoptères torrenticoles. V. Elmidae de la cordillère des Andes. Bulletin de l’Institut royal des Sciences Naturelles de Belgique, 33, 1–8.
Lafuente, W., Soto L. M., López C., & Domínguez-Granda, L. (2019). Efectos de un derrame de petróleo crudo en la comunidad de macroinvertebrados bentónicos de un río amazónico ecuatoriano. Revista de Ciencias Ambientales, 53, 1–22. https://doi.org/10.15359/rca.53-1.1
Lambert, P., Maier, C. A., & Leschen, R. A. B. (2015). A new species and lectotype designations for New Zealand Hydora (Elmidae: Larainae) with a world checklist. New Zealand Entomologist, 38, 38–57. https://doi.org/10.1080/00779962.2014.963910
Lataste, F. (1897a). Présentation d’une larve de Pseudo-Névroptère d’aspect crustacéen. Actes de la Société Scientifique du Chili, 7, 103–106.
Lataste, F. (1897b). La larve crustacéiforme de Peñaflor est de Coléoptère et non de Pseudo-Névroptère. Actes de la Société Scientifique du Chili, 7, 107–108.
Manzo, V. (2005). Key to South America genera of Elmidae (Insecta: Coleoptera) with distributional data. Studies on Neotropical Fauna and Environment, 40, 201–208. https://doi.org/10.1080/01650520500140619
Manzo, V. (2007). Cinco citas nuevas de Elmidae (Coleoptera) para la Argentina, con la redescripción de Austrolimnius (Telmatelmis) nyctelioides. Revista de la Sociedad Entomológica Argentina, 66, 11–20.
Manzo, M. V., & Archangelsky, M. (2008). A key to the known larvae of South American Elmidae (Coleoptera: Byrrhoidea), with a description of the mature larva of Macrelmis saltensis Manzo. International Journal of Limnology, 44, 63–74. https://doi.org/10.1051/limn:2008023
Manzo, M. V., & Moya, N. (2010). Description of the first South American species of Neocylloepus Brown (Coleoptera: Elmidae): N. chaparensis sp. nov. from Bolivia. Studies on Neotropical Fauna and Environment, 45, 129–138. https://doi.org/10.1080/01650521.2010.520210
Martínez-Román, N. M., & Archangelsky, M. (2017). Description of the mature larva of Austrolimnius nyctelioides (Germain, 1892) and A. elatus Hinton, 1941 (Coleoptera: Elmidae). Zootaxa, 4216, 585. https://doi.org/10.11646/zootaxa.4216.6.5
Martínez-Román, N. R., Manzo, V., & Archangelsky, M. (2019). A new species of Stethelmis Hinton (Coleoptera: Elmidae) from Argentina and description of its larva. Anais da Academia Brasileira de Ciências, 91, 1–13. https://doi.org/10.1590/0001-3765201920180954
Martínez-Román, N., Manzo, V., & Archangelsky, M. (2025). Descriptions of the mature larvae of the genera Austrelmis and Macrelmis (Coleoptera: Elmidae) from Argentina. Zoosystematica Rossica, 34, 181–207. https://doi.org/10.31610/zsr/2025.34.1.181
Monte, C., & Mascagni, A. (2012). Review of the Elmidae of Ecuador with the description of ten new species (Coleoptera: Elmidae). Zootaxa, 3342, 1–38. https://doi.org/10.11646/zootaxa.3342.1.1
Passos, M. I. S., Manzo, V., & Maier, C. A. (2018). Families Dryopidae, Elmidae and Psephenidae. In J. Thorp, & A. Covich (Eds.), Thorp and Covich’s freshwater invertebrates. Fourth Edition, Volume 3 (pp. 583–598). Salt Lake City: Elsevier. https://doi.org/10.1016/B978-0-12-804223-6.00027-5
Polizei, T. T. S. (2018). A new species and a key for the Brazilian species of the genus Heterelmis Sharp, 1882 (Coleoptera: Elmidae, Elminae). Papéis Avulsos de Zoologia, 58, e20185820. https://doi.org/10.11606/1807-0205/2018.58.20
Polizei, T. T. S., Costa, L. S. M., & Bispo, P. C. (2020). Austrolimnius cleidecostae, a new species of riffle beetle (Coleoptera: Elmidae) from Brazil. Papéis Avulsos de Zoologia, 60, 1–20. https://doi.org/10.11606/1807-0205/2020.60.special-issue.12
Shepard, W. D. (2016). Notes on the endemic Chilean terrestrial dryopoid Sosteamorrphus verrucatus Hinton 1936 (Coleoptera: Dryopidae). Boletín del Museo Nacional de Historia Natural, Chile, 65, 175–183. https://doi.org/10.54830/bmnhn.v65.2016.93
Shepard, W. D., & Barr, C. B. (2023). A revision of the Chilean water penny beetle genus Tychepsephus Waterhouse, 1876 (Coleoptera, Psephenidae, Eubriinae). Zookeys, 1164, 23–61. https://doi.org/10.3897/zookeys.1164.103184
Shepard, W. D., & Chaboo, C. S. (2015). Beetles (Coleoptera) of Peru: a survey of the families Elmidae. Journal of the Kansas Entomological Society, 88, 167–168. https://doi.org/10.2317/kent-88-02-167-168.1
Solervicens, J. (1974). Observaciones biológicas y de distribución acerca de Chiloea chilensis Dajos 1973. Anales del Museo de Historia Natural, 7, 234–240.
Spangler, P. J., & Brown, H. P. (1981). The discovery of Hydora, a hitherto Australian-New Zealand genus of riffle beetles in Austral South America (Coleoptera; Elmidae). Proceedings of the Entomological Society of Washington, 83, 596–606.
Spangler, P. J., & Staines, C. L. (2002). Luchoelmis, a new genus of Elmidae (Coleoptera) from Chile and Argentina. Insecta Mundi, 16, 215–220.
Una especie incrustante nueva de Nymphon (Arthropoda: Pycnogonida) de la bahía de La Paz, golfo de California, México
Angel de León-Espinosa, Jesús Angel de León-González *
Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas, Laboratorio de Zoología de Invertebrados No Artrópodos, Av. Pedro de Alba s/n, Cd. Universitaria, 66451 San Nicolás de los Garza, Nuevo León, Mexico
*Corresponding author: jesus.deleongn@uanl.edu.mx (J.A. de León-González)
A new species of pycnogonid belonging to the genus Nymphon is described and illustrated from specimens collected at Bahía de La Paz (Gulf of California), where it is part of the dock fouling community. The new species is morphologically similar to Nymphon littus, both species can be separated based on the dentition of the oviger terminal claw, strigilis formula, the relative size of palp segments and chela ornamentation. An identification key for the species reported from the Eastern Pacific is included.
Se describe e ilustra una especie nueva de picnogónido perteneciente al género Nymphon a partir de especímenes recolectados en la bahía de La Paz (golfo de California), donde forma parte de la comunidad de incrustantes en andadores de muelles náuticos. La especie nueva es morfológicamente similar a Nymphon littus; ambas especies pueden distinguirse según la dentición de la uña terminal del ovígero, la fórmula de la strigilis, el tamaño relativo de los segmentos del palpo y la ornamentación de la quela. Se incluye una clave de identificación para las especies reportadas en el Pacífico oriental.
Palabras clave: Picnogónido; Nymphonidae; Nymphon childi; Mar de Cortés; Araña marina; Incrustantes
Introduction
Nymphon Fabricius, 1794 is the most diverse genus of the class Pycnogonida. Currently the genus includes 269 accepted species (Bamber et al., 2026) but a large number of species are poorly described, making difficult its study (Bamber, 1982; Takahashi et al., 2012). Four species of Nymphon have been recorded, so far, in Mexico (Munilla-León, 2002; de León-Espinosa et al., 2021; Ramírez-Tello et al., 2022): N. apheles Child, 1979, described from Panama City, reported also in Bahía de La Paz, Gulf of California (de León-Espinosa et al., 2021), N. floridanum Hedgpeth, 1948 described from Florida and reported in the Yucatán Channel (Stock, 1986); N. lituus Child, 1979 described from the northeastern Gulf of California, with records in the Pacific coast of Panama (Child, 1979), Ojo de Liebre Lagoon on the western coast of Baja California Peninsula (de León-Espinosa & de León-González, 2015), as well as Bahía Banderas, Nayarit (Villatoro-Ortiz & Salazar-Silva, 2022); and N. pixellae Scott, 1912 described from Vancouver, Canada and reported in the Gulf of California by Child (1979).
During a sampling effort carried out at public and private nautical marinas along the west coast of the Gulf of California, specimens of a new species of the genus Nymphon were collected inhabiting with the algae Padina caulescens Thivy, 1945, and the barnacles Tetraclita stalactifera (Lamarck, 1818) fixed to floating docks not deeper than 0.50 m. Type material was deposited in the Colección Carcinológica de la Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León (UANL-FCB-PYCNO).
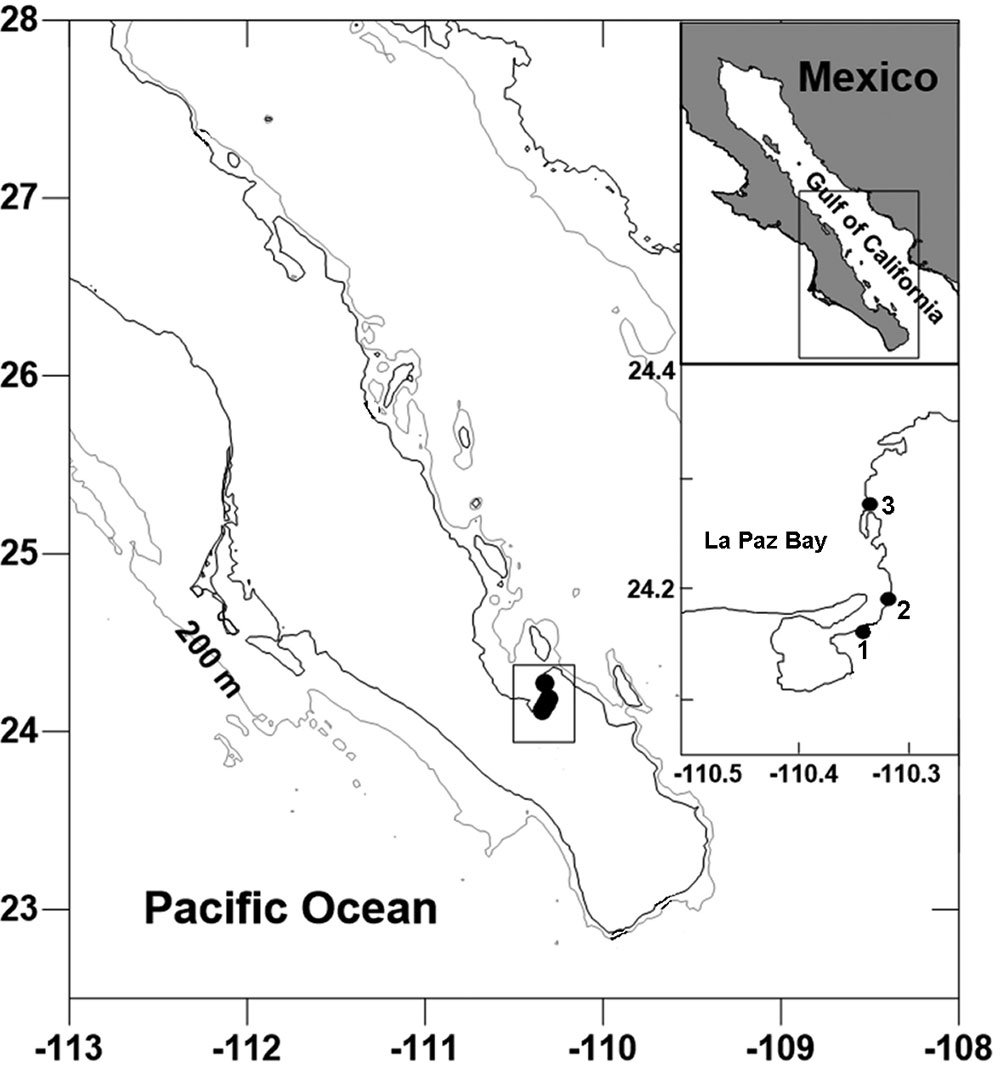
Materials and methods
Samples of Nymphon were collected from 3 marine docks located in Bahía de La Paz, Baja California Sur, Mexico (La Paz, Palmira and Cantamar) (Fig. 1). Each encrusting faunal sample was obtained by scraping an area of 400 cm² (20 × 20 cm) located immediately below the sea surface from the edges of the platform and floating docks. The scraping of encrusted fauna was done with a carpenter’s chisel or a spatula following a standard technique (Pech et al., 2023). The encrusted fauna detached from the dock were deposited in plastic bags (Ziploc®) labeled with the date, time and geographic location information for each collection site. The biological samples were later placed in a cooler with seawater obtained from the collection site to transfer to the processing site. At the processing site, specimens from each biological sample were identified and sorted in situ into large taxonomic groups.
Figure 1. Area of study showing the marine docks located [in] Bahía de La Paz, Gulf of California, Mexico: 1 = La Paz; 2 = Palmyra; 3 = Cantamar.
The pycnogonid specimens, sorted from the encrusted fauna, were fixed with a 10% formaldehyde solution in 2 mL cryovials. The pycnogonids were then washed a week later with distilled water to remove the excess formalin and salt and then preserved in 70% non-denatured ethanol.
Morphological observations were conducted using a stereomicroscope Olympus SZ, and an Olympus BX51 microscope with differential interference contrast (DIC). The figure backgrounds were cleaned and lightened as necessary, and final figures were assembled on a plate using Paint Shop Pro7. Description of the species is based on the morphology of the holotype.
Description
Class Pycnogonida Latreille, 1810 Order Pantopoda Gerstäcker, 1863 Family Nymphonidae Wilson, 1878 Genus Nymphon Fabricius, 1794 Type species: Nymphon grossipes (O. Fabricius, 1780)
Diagnosis. Trunk usually segmented and elongate. Lateral processes in general are well separated. Ocular tubercle is inserted at the posterior end of the cephalic segment. Abdomen is usually short. Proboscis variable in shape, cylindrical. Cheliphore robust, the scape formed by 1 segment, chelae complete and functional, fingers are long and have teeth of various shape and size. Palps 5-segmented, first one always short; the second one usually longest. The 10-segmented ovigers are present in both sexes, with a strigilis formed by the last 4 segments. They have denticulate spines and usually a toothed terminal claw. Propodus is heelless, main claw usually has auxiliary claws of various sizes. Cement glands, when present, are only present in males and open on the ventral side of the femur.
Remarks. Nymphon the most diverse genus within Pycnogonida, early descriptions are very brief, and type materials are unavailable, making its study very difficult. Traditionally, its species have been placed in artificial groups; however, most species have not been able to be placed in such groups for study.
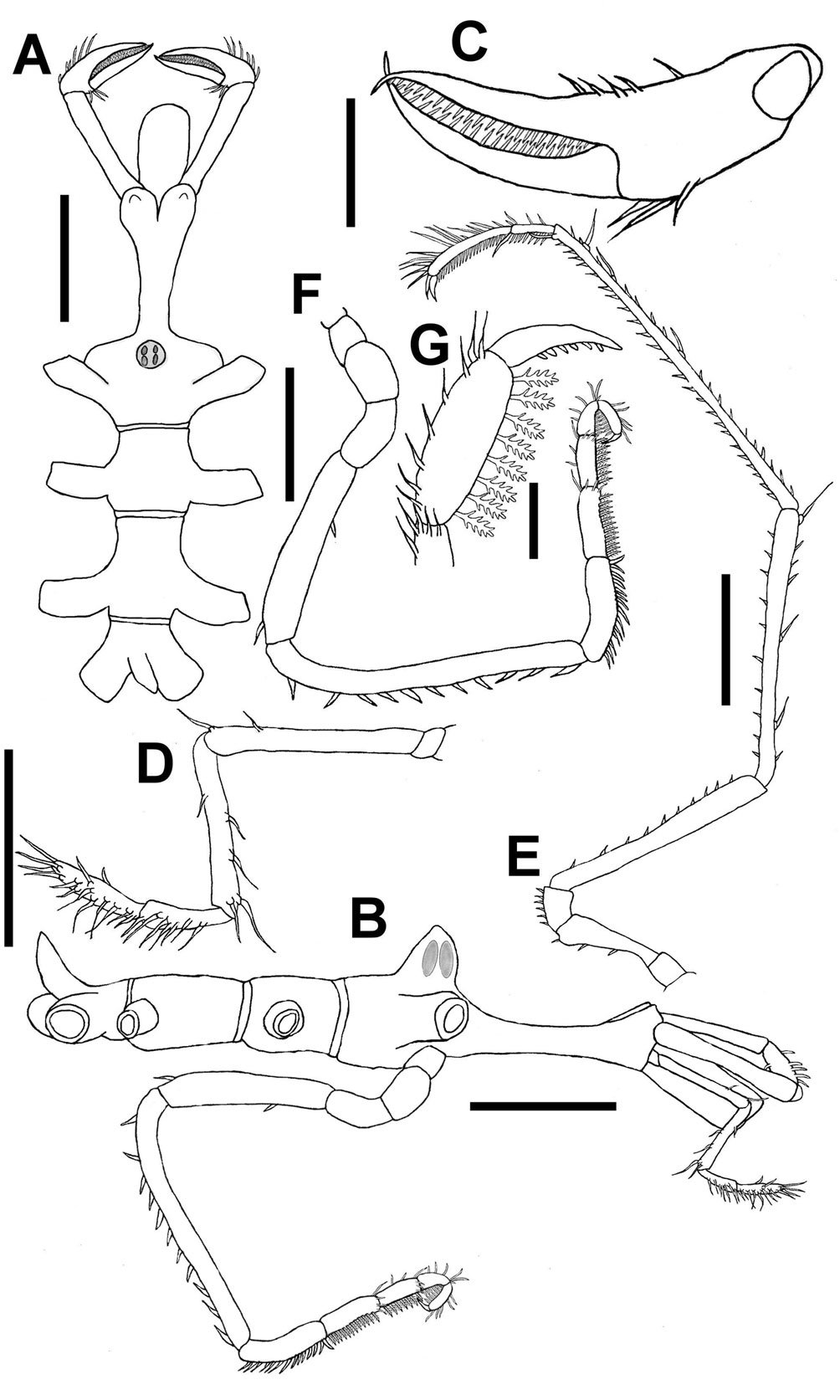
Nymphon childi sp. nov. (Fig. 2)
Description. Male holotype (UANL-FCB-PYCNO-0055), trunk slender, fully segmented, neck in front of oviger bases, half the width of trunk, 4.5 times longer than its width, cylindrical, widening in the form of a calyx, with pair of conical, short anterior projections (Fig. 2A, B); lateral processes between first and second pairs separated by 2 times their own diameters, second and third pairs separated by 2.5 times their own diameters, third and fourth pairs separated by their own diameters (Fig. 2A). Ocular tubercle as high as basal width, inserted between first pair of lateral processes and bases of ovigers, cone-shaped, with 2 small papilliform projections, with 2 pigmented pairs of eyes (Fig. 2A, B).
Proboscis cylindrical, longer than wide, horizontal to body (Fig. 2A, B), with 3 smooth lips.
Chelifore scape over-reaching proboscis, chela fingers curved inwards, crossing at tips, 20-21 needle-like teeth of uniform size on each finger, slender, longer than basal part, which is inflated and rectangular, with 5 median ventral spines and 3 large dorsal setae in a row at distal end, next to articulation with movable finger (Fig. 2C).
Abdomen narrowing distally, length over 2 times greatest width, as long as lateral processes of fourth pair of legs, elevated from body at an angle of about 30° (Fig. 2B).
Palps 5-segmented, first segment very short. Second segment longest, with 1 medio-dorsal seta and a large dorso-distal 1. Third segment about 2/3 length of segment 2 with line of 3 ventral setae and latero-distal one; with medio-dorsal small seta. Fourth segment 4.5 times as long as segment 1, with ventral row of 7 setae. Fifth segment 1.1 times as long as fourth segment, with 9 long ventral setae, medio-dorsal small seta, terminal end with cluster of 4 setae (Fig. 2D).
Oviger 10-segmented, inserted in distal half of first lateral processes (Fig. 2B). First 3 segments short, first segment half size of second one, second and third segments subequal. Fourth segment with dorsal seta and ventro-distal one. Fifth segment 1.6 times longer than fourth, with ventral row of 11 setae, dorsal surface bare. Sixth segment as long as first and second segments together, with ventral row of 14 setae. Last 4 segments compose the strigilis, armed with compound ovigerous leg spines, each with the following formula: 12: 7: 5: 8. Compound spines with 3-6 pairs of lateral teeth depending upon the size of the spine. Terminal claw long, curved, with 6 teeth (Fig. 2G). First segment of strigilis with transverse row of 4 distal setae, second one with 2 dorsal isolated setae and distal row with 4 setae, in both dorsal setae longest; third segment with 3 dorsal setae and group of 4 dorso-distal setae; fourth strigilis segment with row of 8 dorsal setae (Fig. 2F, G).
Third leg (Fig. 2E), coxa 1 short, without setae. Coxa 2, 3.3 times longer than coxa 1, with 3 ventral setae. Coxa 3 slightly longer than coxa 1, with row of 6 small ventral setae. Femur long, with 10 scattered ventral setae. Tibia 1 with row of 6 long median-dorsal chaetae and distal spine, ventral row of 10 smaller setae. Tibia 2, 1.3 longer than tibia 1, with row of 17 dorsal setae and 3 median and distal spines, 24 small ventral setae and 1 distal spine. Tarsus with dorsal row of 4 setae, and 7 ventral setae and distal spine. Propodus 2.2 times longer than tarsus, slightly curved, with dorsal row of 13 spines and setae, and row of 22 ventral setae. Main claw short, less than ¼ length of propodus, auxiliary claws nearly as long as main claw.
Female paratype (UANL-FCB-PYCNO-0056). Gonopores oval-shaped, present on ventrodistal end of coxa 2 in all leg pairs.
Taxonomic summary
Etymology. This species is named in honor of Dr. C. Allan Child (Department of Invertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington) in recognition of his life dedicated to the study of Pycnogonida. The species-group name is a noun in the genitive case (ICZN 1999, Art. 31.1.2).
Type material. Holotype (1 male), (UANL-FCB-PYCNO-0055), paratypes (1 male and 4 females), (UANL-FCB-PYCNO-0056), Marina La Paz, La Paz, Baja California Sur, Mexico, 24°07’29.3” N, 110°20’47.3” W, 3 September 2013.
Additional material. One specimen (female), (UANL-FCB-PYCNO-0059), Marina La Paz, La Paz, Baja California Sur, Mexico, 24°07’29.3” N, 110°20’47.3” W, 14 August 2011. One specimen (male), (UANL-FCB-PYCNO-0057), Marina Palmira, La Paz, Baja California Sur, Mexico, 24°11’05.3” N, 110°18’12.7” W, 3 April 2011; 10 specimens (male), (UANL-FCB-PYCNO-0058), same locality, 15 August 2011. Two specimens (1 male and 1 female), (UANL-FCB-PYCNO-0060), Marina Cantamar, La Paz, Baja California Sur, Mexico, 24°16’44” N, 110°19’49” W, 26 June 2017.
Figure 2. Nymphon childi sp. nov. Holotype. A) Trunk, dorsal view; B) trunk, lateral view; C) chela, lateral view; D) palp, lateral view; E) third leg, lateral view; F) oviger, lateral view; G) oviger, terminal end, lateral view. Scale bars: A, B, D, F = 0.5 mm; C, G = 0.1 mm; E = 1 mm.
Remarks
From the current Nymphon species known in the world, 21 have been reported from both coasts of America, 10 of these from the Eastern Pacific: N. aculeatum Child, 1994 (California, Oregon); N. apheles Child, 1979 (Panama); N. duospinum (Hilton, 1942) (Alaska); N. heterodenticulatum Hedgpeth, 1941 (Santa Catalina Island, California); N. hirsutum Child, 1995 (Bering Sea); N. lituus Child, 1979 (Gulf of California and Panama); N. longicollum Hoek, 1881 (Chile); N. pixellae Scott, 1912 (Vancouver, Oregon, Gulf of California); P. simulare Child, 1992 (Ecuador) and N. stipulum Child, 1990 (California).
Nymphon childi sp. nov. is morphologically similar to N. lituus Child, 1979 and N. simulare Child, 1992, all have a slender body, the third metamere of the body is longer than wide, the neck is 4 or more times longer than wide, and the tarsus is less than 50% of the length of the propodus. Nymphon childi sp. nov. differ from these species in the following characteristics: 1) the anterior tubercle over insertion of chelifores, conical in N. childi sp. nov., and as low anteriorly curved tubercle in N. lituus, N. simulare does not present anterior tubercles; 2) strigilis formula, of 12: 7: 5: 8 in N. childi sp. nov., 15: 10: 10: 11 in N. littus, and 11:10:10:10 in N. simulare; 3) second segment of the palp is shorter than third and fourth together in N. childi sp. nov. and N. simulare, and sligthly longer in N. littus; 4) third segment of the palp is 1.5 times longer than the fifth in N. childi sp. nov. and N. simulare and subequal to the fifth one in N. littus; 5) chela fingers, with 23 teeth in both chelae in N. childi sp. nov, immovable finger has 29 teeth, movable finger with 33 teeth in N. littus, and 27-28 teeth in N. simulare.
Another closer species to Nymphon childi sp. nov. is N. heterodenticulatum, both have the same structural body plan, proboscis horizontal to body, with a long cylindrical neck longer than wide ending in a structure with calyx shape, similar lateral processes separation, and abdomen shape. However, N. heterodenticulatum presents the ocular tubercle sharply pointed, ending in a cone shaped structure, chelifores with slender chela, fixed finger with 17 teeth, movable finger with 19 teeth; oviger with strigilis with a formula of 9: 9: 7: 7.
This species is currently known only from the type locality: Bahía de La Paz, Baja California Sur, Mexico, attached to dock fouling not deeper than 0.5 m.
Key to Nymphon species from Eastern Pacific
1. Tarsus longer than propodus … 2 — Tarsus shorter than propodus … 4 2. Ocular tubercle well developed; auxiliary claw present; fourth and fifth segment of palp equal in size … 3 — Ocular tubercle and eyes lacking; auxiliary claws absent; fourth segment of palp shorter than the fifth one … N. aculeatum Child, 1994 (California) 3. Auxiliary claws less than half the length of the main claw; strigilis of oviger as follow 20:9:14:12; terminal claw of oviger with 15-17 teeth; movable finger of chela with 35 thin teeth; immovable finger with 20 teeth … N. pixellae Scott, 1912 (Vancouver, Canada) — Auxiliary claws slightly longer than the main claw; strigilis of oviger as follow 10: 9: 8: 10; terminal claw of oviger with 8 teeth; movable finger of chela with 19 teeth; immovable finger with 17 teeth … N. heterodenticulatum Hedgepeth, 1941 (Santa Catalina Island, California) 4. Short neck, less than twice own width … 5 — Long neck, more than twice own width … 7 5. Auxiliary claws less than half the length of the main claw; cheliphore with long and thin scape … 6 — Auxiliary claws longer than half the length of the main claw; cheliphore with short and robust scape … N. apheles Child, 1979 (Panama) 6. Palp with third segment longer than second one, with a group of long distal chaetae; tarsus longer than half the size of the propodus … N. stipulum Child, 1990 (California) — Palp with third segment shorter than second one, with short chaetae along surface; tarsus shorter than half the size of the propodus … N. duospinum (Hilton, 1942) (Alaska) 7. Auxiliary claws thin and small, less than ¼ the size of the main claw; chelae with few teeth on both fingers, movable finger with 8 teeth, immovable finger with 5 teeth … N. longicollum Hoek, 1881 (Chile) — Auxiliary claws equal to, or longer than half the size of the terminal claw … 8 8. Fingers of the chelae shorter than palm, movable finger with 34 teeth, immovable finger with 26 teeth; oviger with strigilis as follow 11: 6: 6: 6; oviger claw with 20 teeth … N. hirsutum Child, 1995 (Bering Sea) — Fingers of the chelae longer than palm … 9 9. Oviger with strigilis as follow 12: 7: 5: 8; movable finger of chelae with 20 teeth, immovable finger with 20-21 teeth … N. childi sp. nov. (Bahía de La Paz, Gulf of California) — Last 3 segments of the strigilis with 10 or more compound spines; chelae fingers with more than 20 teeth each … 10 10. Third segment of the palp similar in size of the second one; distal end of neck without distal tubercules … N. simulare Child, 1992 (El Viejo Island, Ecuador) — Third segment of the palp ½ the size of the second one; distal end of neck with anteriorly curved low tubercle over insertion of chelifores … N. lituus Child, 1979 (Puerto Peñasco, Gulf of California)
Acknowledgements
This work was financially supported by the Universidad Autónoma de Nuevo León (grant number PAICYT 0235). Many thanks to Jesús Flores and Jesús Mendieta for their technical help with the collections and sorting out the specimens from the samples. This work could not be completed without the support of Claudia Arango, Lyudmila Slucheusky and Isabell Clasen who sent us some relevant scientific literature. We thank Mariana Tovar-Hernández for her review of the manuscript, and an anonymous reviewer.
References
Bamber, R. N. (1982). Variation in Nymphon brevirostre (Hodge), and the status of N. rubrum (Hodge) (Arthropoda: Pycnogonida). Biological Journal of the Linnean Society, 17, 275–288. https://doi.org/10.1111/j.1095-8312.1982.tb02022.x
Bamber, R. N., El Nagar, A., Arango, C. P., & Sabroux, R. (Eds.). (2026). Pycnobase: World Pycnogonida Database. Nymphon Fabricius, 1794. Accessed through: World Register of Marine Species at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=134591
Child, C. A. (1979). Shallow-water Pycnogonida of the Isthmus of Panama and the Coasts of Middle America. Smithsonian Contributions to Zoology, 293, 1–86. https://doi.org/10.5479/si.00810282.293
de León-Espinosa, A., & de León-González J. A. (2015). Pycnogonids associated with the giant lion´s-paw scallop Nodipecten subnodosus (Sowerby) in Ojo de Liebre Bay, Guerrero Negro, Baja California Sur, Mexico. Zookeys, 530, 129–149. https://doi.org/10.3897/zookeys.530.6064
de León-Espinosa, A., de León-González J. A., & Gómez-Gutiérrez J. (2021). Pycnogonids from marine docks located along the west coast of the Gulf of California, Mexico. Zootaxa, 4938, 151–195. https://doi.org/10.11646/zootaxa.4938.2.1
ICZN (International Code of Zoological Nomenclature). (1999). International Code of Zoological Nomenclature, 4th Edition. London: International Commission on Zoological Nomenclature.
Munilla-León, T. (2002). Pycnogonida. En J. Llorente-Bousquets, & J. J. Morrone (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia una síntesis de su conocimiento, Vol. III (pp. 215–222). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Pech, D., Tovar-Hernández, M. A., Bastida-Zavala, R., Carrera-Parra, L. F., de León-González, J. A. y Delgado-Blas, V. H. et al. (2023). Protocolo para el muestreo, análisis y estudio de la biota portuaria en México. Monterrey: Universidad Autónoma de Nuevo León.
Ramírez-Tello, L. M., Tello-Musi, J. L., & López-García, R. (2022). Pycnogonida of Mexican waters: a checklist with observations on diversity and ecology. Bulletin of Marine and Coastal Research, 51, 151–170. https://doi.org/10.25268/bimc.invemar.2022.51.1.1186
Stock, J. H. (1986). Pycnogonida from the Caribbean and the Straits of Florida. Bulletin of Marine Science, 38, 399–441.
Takahashi, Y., Kajihara, H., & Mawatari, S. F. (2012). Sea spiders of the genus Nymphon (Arthropoda: Pycnogonida) from waters around the Nansei Islands, Japan. Journal of Natural History, 46, 1337–1358. https://doi.org/10.1080/00222933.2012.655797
Villatoro-Ortiz, A., & Salazar-Silva, P. (2022). Picnogónidos (Arthropoda: Pycnogonida: Pantopoda) de la bahía de Banderas, Pacífico mexicano. Revista Ciencias Marinas y Costeras, 14, 71–90. https://doi.org/10.15359/revmar.14-2.5
Egg morphology and oviposition techniques in Pseudosermyle (Phasmatodea: Diapheromeridae)
Ulises López-Mora a, b, *, Enrique González-Soriano a, A. Celeste Martínez-Cervantes a, b, Jorge Llorente-Bousquets c
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Tercer Circuito Universitario s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
b Universidad Nacional Autónoma de México, Posgrado en Ciencias Biológicas, Instituto de Biología, Departamento de Zoología, Tercer Circuito Universitario s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
c Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Biología Evolutiva, Museo de Zoología Alfonso L. Herrera, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
* Autor para correspondencia: nxitl00@gmail.com (U. López-Mora)
Recibido: 30 julio 2025; aceptado: 04 octubre 2025
Resumen
Pseudosermyle es el género de insectos palo con mayor riqueza en América del Norte; muestra una distribución desde el sur de los EUA hasta América Central, desafortunadamente ha sido poco estudiado. En el presente trabajo se realiza la primera aproximación en extenso a la morfología del huevo en Pseudosermyle respecto a su microestructura, con el empleo de microscopía electrónica de barrido (MEB), así como el análisis de componentes principales para determinar la utilidad de las medidas estandarizadas que se utilizan en la descripción de huevos, además de su ornamentación. Se encontraron 4 patrones en el huevo de Pseudosermyle, entre los que se incluyen 3 tipos de oviposición: liberación, adheridos al sustrato y formación de ooteca. Esto sugiere que podría tratarse de géneros distintos bajo una posible subtribu. Además, la ornamentación del huevo es lo suficientemente distinta para reconocer diferentes especies. Por lo tanto, se debe analizar la importancia de estos caracteres en la descripción de especies de Pseudosermyle.
Pseudosermyle is the genus of stick insects with the greatest diversity in North America, with a range extending from the southern USA to Central America; unfortunately, it has been poorly studied. In this work, we present the first comprehensive analysis of the egg morphology in Pseudosermyle in relation to ultrastructure, using scanning electron microscopy, along with principal component analysis to evaluate the effectiveness of the standardized measurements used in the egg descriptions, as well as the ornamentation. We identified 4 general patterns in Pseudosermyle eggs corresponding to 3 different oviposition techniques: dropping, attaching to substrate and forming oothecae. These patterns suggest the possibility of recognizing different genera within a putative subtribe. Additionally, the ornamentation is sufficiently distinct to differentiate among species. Thus, we emphasize the importance of these characters in the description of species of Pseudosermyle.
El orden Phasmatodea comprende organismos denominados insectos palo, hoja o corteza. Por lo general se reconocen por su cuerpo alargado y cilíndrico o deprimido; es menos frecuente encontrar formas más robustas (Bradler y Buckley, 2018). Su coloración varía en distintos matices del castaño al verde, aunque existen algunos con coloraciones aposemáticas (rojo, amarillo o azul). En la etapa adulta ninguno de estos insectos mide menos de 2 cm de longitud (Seiler et al., 2006). Una característica notoria es su cripsis con la vegetación, que en conjunto con comportamientos especializados, como la catalepsia e imitación del balanceo de las hojas y el follaje durante el día (Bradler y Buckley, 2018), les permiten pasar inadvertidos frente a sus depredadores. Sus huevos se asemejan a semillas de las plantas donde habitan (Severin, 1910).
La ootaxonomía de Phasmatodea se estandarizó con los trabajos de Clark (1976a, b, 1979) y Sellick (1994, 1997a, b, 1998). A partir de entonces, el huevo se ha incluido en las descripciones de sus especies (Allred et al., 1986; Conle et al., 2007; López-Mora y Llorente-Bousquets, 2023) y también se han efectuado trabajos utilizando el microscopio electrónico de barrido (MEB) para el análisis de las microestructuras y la textura de la cápsula en especies pertenecientes a los géneros Agathemera Stål, 1875, Anisomorpha Gray, 1835, Bacillus Berthold, 1827, Burria Brunner von Wattenwyl, 1900, Carausius Stål, 1875, Clonopsis Pantel, 1915, Heteronemia Gray, 1835, Lonchodes Gray, 1835, Megaphasma Caudell, 1903, Orxines Stål, 1875, Paraprisopus Redtenbacher, 1906, Ramulus Saussure, 1862, Sipyloidea Brunner von Wattenwyl, 1893, Timema Scudder, 1895 y Xeropsis Redtenbacher, 1906 (Allred et al., 1986; Camousseight y Bustamante, 1991; Jintsuet al., 2010; Mazzini y Scali, 1977, 1980, 1983; Mazzini et al., 1982, 1984, 1992; Scali y Mazzini, 1977, 1981, 1982, 1983; Stark y Lentz, 1986). En particular, la textura de la cápsula y la ornamentación del opérculo han resultado ser muy útiles en la diferenciación de especies cercanamente relacionadas (Mazzini et al., 1982, 1992; Scali y Mazzini, 1981), incluso para diferenciar híbridos (Mazzini et al., 1984). En suma, los trabajos previos han concluido en la importancia de los estudios con MEB y microscopia óptica para la diferenciación entre especies cercanamente relacionadas en Phasmatodea y posibles híbridos.
Pseudosermyle Caudell, 1903 es el género más diverso en México y América del Norte, con 26 especies en total (de Luna, 2022; López-Mora y Llorente-Bousquets, 2018). Sin embargo, solo se conocen 4 ilustraciones, que no son de microestructura del huevo, para 3 especies: Pseudosermyle tridens (Burmeister, 1838), ilustrada por Kaup (1871) y Zompro (2001); P. truncata Caudell, 1903, por Caudell (1914) y P. phalangiphora Conle, Hennemann et Fontana, 2007, por ellos mismos. Al observar las imágenes de estas 3 especies se advierten diferencias significativas en las formas de la cápsula y la lámina micropilar, que no han sido evaluadas en su microestructura y son relevantes para distinguir especies.
Nuestro objetivo fue realizar una exploración más extensa sobre la morfología coriónica a nivel de microestructura utilizando MEB, incorporando 11 especies más de Pseudosermyle, así como analizar su empleo a nivel genérico y específico al reconocer jerarquías entre los caracteres que se utilizan para describir a las especies.
Materiales y métodos
Los acrónimos y abreviaturas de las colecciones son: CNIN = Colección Nacional de Insectos, Instituto de Biología, UNAM; MZFC = Museo de Zoología Alfonso L. Herrera, Facultad de Ciencias, UNAM. Estructuras coriónicas: H = altitud de la cápsula; L = longitud de la cápsula; mpi = lámina micropilar interna; mpl = longitud de la lámina micropilar; mpw = amplitud de la lámina micropilar, NM = lámina micropilar interna abierta con línea media; oph = altura del opérculo, opw = amplitud del opérculo; PO = lámina micropilar interna paralela sin línea media; W = amplitud de cápsula; %h/l = índice de elongación, %w/h = índice de compresión.
Se recolectaron 44 machos y 61 hembras en distintas localidades (fig. 1, tabla 1) y se depositaron en la CNIN y MZFC (tabla 2), Nooxapty isabelae se revisó de los ejemplares tipos depositados en las colecciones por López-Mora y Llorente-Bousquets (2023) (tabla 2). Los ejemplares que se obtuvieron de crianza aún no se depositan en colección alguna, pues se están utilizando para otro tipo de estudios. Los puntos de recolecta se muestran en un mapa (fig. 1) obtenido con QGIS 3.8.1-Zanzibar (QGIS, 2019). Para obtener los huevos, se seleccionaron 22 hembras de distintas especies (tabla 3), el número de hembra varió entre las especies pues algunas no sobrevivieron al traslado, se mantuvieron en cautiverio y aisladas entre sí (dentro de diferentes contenedores) con las siguientes condiciones: temperatura de 20-22 °C, humedad relativa 60-80%, alimentación con base en Galinsoga sp. (Asteraceae) que se colocó en frascos con agua. Estuvieron bajo ciclos de luz y oscuridad correspondientes a los presentados en la CDMX durante el lapso que estuvieron cautivas; se intentaron replicar las condiciones de temperatura y humedad tomadas en campo con un termohigrómetro digital PEAKMETER PM6508. Los organismos se identificaron comparando la morfología de adultos (machos y hembras) y el tipo de oviposición, en particular con las fotografías e ilustraciones de Brock et al. (2019), estos datos fueron cualitativos y no se incluyeron en el análisis de componentes principales (PCA). Para Nooxaptyisabelae López-Mora et Llorente-Bousquets, 2023 se examinaron los huevos de la serie tipo y se usó como grupo externo. Se observaron y midieron en total 255 huevos de 22 hembras de 12 especies (tabla 3) bajo un microscopio Olympus SZX12, con ocular de reglilla micrométrica y un objetivo Olympus DF PL 1.5 X-4. Se efectuaron las mediciones de los huevos acorde con Sellick (1997b); los datos se presentan en las tablas 4-9 y en las diagnosis se expresan las cifras bajo los siguientes parámetros estadísticos: n = tamaño de la muestra, promedio ± desviación estándar (valor mínimo-valor máximo) en las descripciones.
Tabla 1
Localidades de recolecta de las distintas especies de insectos palo, el orden es alfabético por estado.
Localidad
Longitud
Latitud
Especies
REPSA, Coyoacán, Ciudad de México, México
19.317634
-99.184854
Pseudosermyle sp. 2
Arriba cueva, Valle de Bravo, Estado de México
19.09248
-100.07218
Pseudosermyle sp. 1
Bajo mesa, Valle de Bravo, Estado de México, México
19.0861
-100.07665
Pseudosermyle carinulata
Villa Esmeralda, Silao, Guanajuato, México
20.95367
-101.43361
Pseudosermyle sp. 4
EBCH, La Huerta, Jalisco, México
19.49931
-105.04484
Pseudosermyle sp. 3
Cruz Verde, Ayala, Morelos, México
18.760174
-98.976098
Pseudosermyle tridens
Curva Choápam-Latani, Santiago Choápam, Oaxaca, México
17.377754
-95.916254
Pseudosermyle sp. 6
Curva 1 Amatepec-Chinantequilla, Totontepec Villa de Morelos, Oaxaca, México
17.28485
-95.988362
Nooxapty isabelae
Camino a Tzinacapan, Cuetzalan, Puebla, México
20.016558
-97.53983
Pseudosermyle striatusPsudosermyle sp. 5
EBTLT, San Andrés Tuxtla, Veracruz, México
18.5860833
-95.0755
Pseudosermyle phalangiphoraPseudosermyle procera
Tabla 2
Organismos revisados y depositados en las colecciones por especie, sexo y huevos indicando el voucher. Para Nooxapty isabelae se indican los ejemplares revisados. En Núm. de especímenes se separan los ejemplares depositados y revisados (primera cifra) de los revisados (segunda cifra) por un símbolo de más “+”.
Especie
CNIN
MZFC
Machos
Hembras
Huevos
Machos
Hembras
Huevos
Pseudosermyle sp. 1
PHMX351-354, 356-358
PHMX26
PHMX125
PHMX349-350, 355, 426
Tabla 2. Continúa
Especie
CNIN
MZFC
Machos
Hembras
Huevos
Machos
Hembras
Huevos
Pseudosermyle sp. 2
PHMX307-308, 311, 314-322
PHMX72-75, 78-79
PHMX76-77, 80
PHMX304-306, 309-310, 312-313
Pseudosermyle sp. 3
PHMX467-468
PHMX469-471
PHMX324, 327-328
PHMX323, 325-326
P. tridens
PHMX678
PHMX679-680
PHMX332-337, 341, 343-348, 616-618
PHMX329-331, 338-340, 342
Pseudosermyle sp. 4
PHMX91, 93
PHMX94-100
PHMX362-365, 367-373, 423-425
PHMX54-56, 90, 92
PHMX57-62
PHMX359-361, 366
P. carinulata
PHMX160-163
PHMX164-165, 168, 653
PHMX420-421
PHMX416-419, 422
P. striata
PHMX81-85
PHMX86-89
PHMX262-270, 272-273, 277-288, 292-300, 302-303
PHMX463
PHMX462, PHMX464-465
PHMX259-261, 271, 274-276, 289-291, 301
Pseudosermyle sp. 5
PHMX27
PHMX378-388
PHMX1-2, 11-12
PHMX4-6, 13
PHMX374-377
P. phalangiphora
PHMX101-102, 135, 148, 155
PHMX103-107, 146-147, 149
PHMX172-183, 188-198, 203-213
PHMX169-171, 184-187, 199-202
P. procera
PHMX108-110, 136-137, 151, 156-157
PHMX111-114, 150, 152-154,
PHMX217-228, 232-243, 248-258
PHMX214-216, 229-231, 244-247
Pseudosermyle sp. 6
PHMX681
PHMX405-415
PHMX401-404
Nooxapty isabelae
PHMX117, 119
PHMX115
PHMX389-393, 399-400
PHMX118
PHMX116
PHMX394-398
Núm. especímenes
27+2
38+1
179+7
17+1
23+1
70+5
Tabla 3
Especies de Phasmatodea revisadas, número de hembras por especie tentativamente bajo el género Pseudosermyle y número de huevos por hembra revisados.
Especies de Phasmatodea revisadas
Clave
Huevos por hembra
Nombre
Clave de ♀
Núm. de ♀
Pseudosermyle sp. 1
PtgVB
U
1
11
Pseudosermyle sp. 2
PtgDF
G1 G2
1 2
5 14
Pseudosermyle sp. 3
Pch
F1 F2
1 2
3 3
P. tridens
PtrMor
T1 T2
1 2
10 10
Tabla 3. Continúa
Especies de Phasmatodea revisadas
Clave
Huevos por hembra
Nombre
Clave de ♀
Núm. de ♀
Pseudosermyle sp. 4
PtrGto
U
1
15
P. carinulata
Pca
C1 C2
1 2
5 2
P. striata
Pst
S1 S2 S3
1 2 3
15 15 15
Pseudosermyle sp. 5
PspF
U
1
15
P. phalangiphora
Pph
H1 H2 H3
1 2 3
15 15 15
P. procera
Ppr
P1 P2 P3
1 2 3
15 15 15
Pseudosermyle sp. 6
Poax
U
1
15
N. isabelae
Nisa
U
1
12
Total: 12 spp.
—
—
22
255
Figura 1. Sitios de recolecta en México de las especies analizadas.
Para asociar machos con hembras que no se encontraban en cópula en el campo, se realizaron los siguientes pasos: a) se observaron los machos con los cuales copulaban las hembras en cautiverio; b) se separaron los huevos por morfología y hembra; c) éstos se incubaron por separado; d) cuando eclosionaron, las ninfas se mantuvieron en terrarios separados; e) se obtuvieron adultos y se comparó la morfología con la presentada por los parentales; f) cuando se obtienen machos y hembras de la misma hembra se corrobora si la morfología que presentan es similar a la de los parentales y se deduce qué machos parentales coinciden; g) se observa que los machos copulen con las hembras correspondientes con la morfología materna; y h) se compara la morfología coriónica de los huevos de las hembras hijas con las parentales para corroborar la correspondencia. Estos pasos permiten discernir cuando hay especies del mismo género que se encuentran en el mismo sitio, o cuando desconocemos la correspondencia entre sexos en los insectos palo de una localidad.
Se tomaron micrografías con una cámara de 8MP LEICA DFC490 adaptada a un microscopio LEICA Z16 AP0A y obtenidas con Leica Application Suite. Éstas se procesaron con GIMP 2.8.22 (GNU). Se llevaron a cabo observaciones bajo el MEB, los huevos se procesaron modificando el método de Nieves-Uribe et al. (2021) de la siguiente manera: serie descendente de etanol, 10 min en cada uno (96% → 70% → 50% → 30% → 10%) → H2O destilada (10 min) → jabón biológico (agitación manual constante) → Sonicar por 10 seg en 2 lapsos de 5 seg (con un sonicador modelo: Bronson 200) → enjuagar en agua destilada → agua destilada (10 min) → serie ascendente de etanol, 10 min en cada uno (10% → 30% → 50% → 70% → 96%) → dejar evaporar el alcohol (30 min. aprox.) → montar la muestra → dejar secar por 2 días en un recipiente hermético con una bolsita de silica gel (2 días). Las muestras se cubrieron en la ionizadora 19Quorum 150R ES en 2 ciclos de 20 min cada uno y se observaron bajo un microscopio Hitachi SU3500. Las abreviaturas en las medidas del huevo siguen la nomenclatura de Sellick (1997b, c, 1998). Las estructuras descritas y propuestas para el huevo se ilustran en la figura 2. Se siguió la nomenclatura de texturas de Harris (1979) para las descripciones de microestructura, en este caso solo se midieron las estructuras de 1 huevo por especie por el costo elevado de la técnica; éste corresponde con el huevo ilustrado en las micrografías obtenidas con el microscopio estereoscópico Leica. Es importante señalar que el nivel de detalle que obtuvimos permite documentar que las estructuras (v. gr. arrugas, espinas, gránulos) presentan textura, que se describe en ocasiones con frases que parecen contradictorias (v. gr. arrugas lisas, gránulos con espinas), pero se acompañan de imágenes.
Figura 2. Partes de un huevo sin capítulo de Phasmatodea adaptado de Clark (1978) a las especies de Pseudosermyle. A, P. striata vista dorsal; B, P. procera vista lateral. El relieve micropilar solo está presente en P. procera y se observa de una vista lateral. El tubérculo micropilar es apreciable en vista dorsal. La línea media solo la presenta P. striata; en ocasiones es apreciable en vista lateral.
Tabla 4
Medidas en micras (μm) de cada huevo de P. phalangiphora utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PphH1-1
2,774
1,752
1,971
876
474.5
1,423.5
1,131.5
PphH1-2
2,737.5
1,752
1,898
912.5
474.5
1,350.5
1,058.5
PphH1-3
2,664.5
1,533
1,788.5
912.5
511
1,350.5
1,095
PphH1-4
2,701
1,715.5
1,898
876
474.5
1,460
1,131.5
PphH1-5
2,628
1,679
1,861.5
949
511
1,460
1,095
PphH1-6
2,847
1,715.5
1,898
766.5
511
1,460
1,204.5
PphH1-7
2,737.5
1,679
1,861.5
766.5
511
1,496.5
1,095
PphH1-8
2,847
1,788.5
1,861.5
912.5
438
1,423.5
1,168
PphH1-9
2,774
1,642.5
1,861.5
803
511
1,460
1,204.5
PphH1-10
2,774
1,679
1,861.5
876
547.5
1,350.5
1,095
PphH1-11
2,810.5
1,679
1,934.5
839.5
438
1,460
1,168
PphH1-12
2,847
1,752
1,861.5
803
474.5
1,460
1,168
PphH1-13
2,737.5
1,715.5
1,861.5
766.5
474.5
1,460
1,131.5
PphH1-14
2,664.5
1,642.5
1,861.5
839.5
474.5
1,387
1,058.5
PphH1-15
2,920
1,788.5
1,934.5
912.5
511
1,496.5
1,168
PphH2-1
2,372.5
1,387
1,496.5
839.5
474.5
1,350.5
1,058.5
PphH2-2
2,555
1,679
1,752
803
438
1,715.5
1,387
PphH2-3
2,555
1,569.5
1,642.5
912.5
438
1,533
1,314
PphH2-4
2,591.5
1,898
1,861.5
766.5
438
1,752
1,387
PphH2-5
2,555
1,606
1,825
766.5
401.5
1,606
1,277.5
PphH2-6
2,591.5
1,606
1,825
766.5
438
1,642.5
1,387
PphH2-7
2,664.5
1,642.5
1,825
766.5
438
1,642.5
1,387
PphH2-8
2,555
1,679
1,861.5
803
438
1,642.5
1,277.5
PphH2-9
2,591.5
1,642.5
1,825
949
547.5
1,606
1,241
PphH2-10
2,628
1,642.5
1,788.5
693.5
438
1,533
1,314
PphH2-11
2,628
1,496.5
1,679
839.5
511
1,460
1,168
PphH2-12
2,591.5
1,606
1,752
839.5
438
1,606
1,314
PphH2-13
2,591.5
1,496.5
1,715.5
839.5
401.5
1,460
1,277.5
PphH2-14
2,591.5
1,679
1,788.5
730
438
1,423.5
1,350.5
PphH2-15
2,664.5
1,715.5
2,007.5
912.5
438
1,715.5
1,423.5
PphH3-1
2,774
1,642.5
1,898
803
474.5
1,387
1,095
PphH3-2
2,664.5
1,642.5
1,825
803
474.5
1,387
1,095
PphH3-3
2,664.5
1,642.5
1,788.5
839.5
511
1,314
1,058.5
PphH3-4
2,737.5
1,642.5
1,825
766.5
547.5
1,387
1,095
PphH3-5
2,628
1,679
1,788.5
803
511
1,350.5
1,058.5
PphH3-6
2,737.5
1,642.5
1,788.5
803
511
1,314
1,095
PphH3-7
2,774
1,642.5
1,825
803
511
1,387
1,095
Tabla 4. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PphH3-8
2,737.5
1,679
1,825
876
511
1,460
1,095
PphH3-9
2,701
1,679
1,861.5
803
547.5
1,350.5
1,022
PphH3-10
2,701
1,642.5
1,788.5
766.5
438
1,350.5
1,095
PphH3-11
2,701
1,606
1,898
839.5
547.5
1,496.5
1,131.5
PphH3-12
2,701
1,642.5
1,861.5
876
547.5
1,496.5
1,095
PphH3-13
2,664.5
1,606
1,825
839.5
438
1,350.5
1,204.5
PphH3-14
2,737.5
1,642.5
1,752
803
474.5
1,350.5
1,095
PphH3-15
2,774
1,679
1,788.5
876
438
1,168
1,058.5
Tabla 5
Medidas en μm de cada huevo de Pseudosermyleprocera utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PprP1-1
2,737.5
1,825
1,934.5
693.5
292
1,496.5
1,241
PprP1-2
2,774
1,788.5
1,934.5
730
292
1,460
1,241
PprP1-3
2,737.5
1,788.5
1,934.5
657
255.5
1,423.5
1,168
PprP1-4
2,737.5
1,679
1,861.5
693.5
292
1,423.5
1,168
PprP1-5
2,555
1,606
1,788.5
730
328.5
1,314
1,095
PprP1-6
2,664.5
1,642.5
1,861.5
657
292
1,350.5
1,204.5
PprP1-7
2,737.5
1,752
1,934.5
730
255.5
1,387
1,131.5
PprP1-8
2,737.5
1,788.5
1,934.5
730
292
1,314
1,131.5
PprP1-9
2,737.5
1,715.5
1,934.5
730
292
1,460
1,204.5
PprP1-10
2,737.5
1,752
1,971
693.5
255.5
1,387
1,131.5
PprP1-11
2,664.5
1,715.5
1,898
693.5
255.5
1,387
1,095
PprP1-12
2,810.5
1,788.5
1,898
693.5
292
1,387
1,095
PprP1-13
2,701
1,752
1,934.5
657
255.5
1,350.5
1,095
PprP1-14
2,810.5
1,752
1,934.5
766.5
292
1,460
1,168
PprP1-15
2,810.5
1,679
1,934.5
693.5
292
1,460
1,131.5
PprP2-1
2,774
1,715.5
1,861.5
693.5
365
1,350.5
1,204.5
PprP2-2
2,664.5
1,788.5
2,044
657
365
1,423.5
1,095
PprP2-3
2,737.5
1,679
1,971
584
328.5
1,569.5
1,131.5
PprP2-4
2,737.5
1,715.5
1,898
620.5
365
1,387
1,131.5
PprP2-5
2,737.5
1,752
1,898
693.5
365
1,496.5
1,131.5
PprP2-6
2,701
1,679
1,861.5
693.5
328.5
1,387
1,204.5
PprP2-7
2,774
1,715.5
1,898
620.5
365
1,460
1,095
PprP2-8
2,737.5
1,715.5
1,934.5
730
365
1,460
1,168
Tabla 5. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PprP2-9
2,701
1,715.5
1,934.5
657
328.5
1,460
1,168
PprP2-10
2,774
1,752
1,934.5
657
365
1,423.5
1,241
PprP2-11
2,737.5
1,715.5
1,898
730
401.5
1,460
1,168
PprP2-12
2,810.5
1,715.5
1,898
730
328.5
1,423.5
1,131.5
PprP2-13
2,664.5
1,679
1,861.5
620.5
292
1,460
1,131.5
PprP2-14
2,664.5
1,679
1,861.5
657
365
1,387
1,131.5
PprP2-15
2,810.5
1,679
1,898
693.5
365
1,387
1,168
PprP3-1
2,737.5
1,788.5
2,007.5
730
401.5
1,569.5
1,241
PprP3-2
2,628
1,788.5
1,898
730
401.5
1,460
1,204.5
PprP3-3
2,737.5
1,825
2,007.5
657
438
1,569.5
1,277.5
PprP3-4
2,701
1,788.5
1,971
730
401.5
1,533
1,241
PprP3-5
2,628
1,825
2,007.5
657
365
1,569.5
1,241
PprP3-6
2,701
1,788.5
1,934.5
657
401.5
1,496.5
1,204.5
PprP3-7
2,701
1,788.5
1,934.5
657
401.5
1,533
1,241
PprP3-8
2,591.5
1,788.5
1,971
620.5
365
1,496.5
1,168
PprP3-9
2,701
1,788.5
1,971
693.5
438
1,569.5
1,168
PprP3-10
2,591.5
1,788.5
1,971
620.5
401.5
1,642.5
1,241
PprP3-11
2,737.5
1,752
1,934.5
657
438
1,569.5
1,241
PprP3-12
2,774
1,825
2,007.5
730
401.5
1,569.5
1,277.5
PprP3-13
2,701
1,715.5
2,044
620.5
401.5
1,533
1,277.5
PprP3-14
2,628
1,825
2,007.5
693.5
401.5
1,533
1,204.5
PprP3-15
2,664.5
1,825
2,007.5
657
401.5
1,642.5
1,277.5
Tabla 6
Medidas en μm de cada huevo de Pseudosermylestriata utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PstS1-1
2,263.8
1,470
1,675.8
1,234.8
529.2
1,029
882
PstS1-2
2,352
1,587.6
1,734.6
1,234.8
529.2
1,146.6
1,029
PstS1-3
2,293.2
1,528.8
1,705.2
1,205.4
529.2
1,087.8
940.8
PstS1-4
2,352
1,558.2
1,646.4
1,205.4
529.2
1,087.8
1,029
PstS1-5
2,116.8
1,528.8
1,705.2
1,058.4
529.2
1,176
999.6
PstS1-6
2,175.6
1,499.4
1,705.2
1,087.8
588
1,117.2
970.2
PstS1-7
2,234.4
1,470
1,646.4
1,117.2
558.6
1,146.6
970.2
PstS1-8
2,263.8
1,528.8
1,646.4
1,058.4
558.6
1,176
1029
Tabla 6. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PstS1-9
2,175.6
1,470
1,675.8
1,087.8
588
1,146.6
999.6
PstS1-10
2,087.4
1,499.4
1,705.2
1,029
558.6
1,117.2
999.6
PstS1-11
2,175.6
1,440.6
1,646.4
1,058.4
529.2
1,146.6
970.2
PstS1-12
2,116.8
1,470
1,705.2
1,087.8
529.2
1,117.2
911.4
PstS1-13
2,116.8
1,499.4
1,675.8
1,176
529.2
1,117.2
940.8
PstS1-14
2,146.2
1,558.2
1,675.8
1,117.2
529.2
1,058.4
940.8
PstS1-15
2,205
1,499.4
1,646.4
1,087.8
588
1,087.8
911.4
PstS2-1
2,322.6
1,558.2
1,705.2
1,029
529.2
1,087.8
940.8
PstS2-2
2,293.2
1,558.2
1,705.2
1,117.2
588
1,058.4
970.2
PstS2-3
2,263.8
1,528.8
1,646.4
999.6
529.2
1,087.8
940.8
PstS2-4
2,263.8
1,352.4
1,528.8
1,058.4
558.6
999.6
911.4
PstS2-5
2,175.6
1,499.4
1,646.4
911.4
529.2
1,087.8
940.8
PstS2-6
2,322.6
1,558.2
1,734.6
1,058.4
529.2
1,087.8
940.8
PstS2-7
2,263.8
1,411.2
1,734.6
1,058.4
558.6
1,117.2
970.2
PstS2-8
2,205
1,558.2
1,675.8
1,029
558.6
1,087.8
940.8
PstS2-9
2,352
1,558.2
1,734.6
1,205.4
499.8
1,087.8
970.2
PstS2-10
2,381.4
1,558.2
1,675.8
1,087.8
558.6
1,029
940.8
PstS2-11
2,263.8
1,499.4
1,617
970.2
558.6
1,058.4
911.4
PstS2-12
2,234.4
1,499.4
1,675.8
1,087.8
558.6
1,029
911.4
PstS2-13
2,293.2
1,470
1,646.4
1,176
529.2
1,058.4
911.4
PstS2-14
2,175.6
1,411.2
1,558.2
970.2
529.2
1,029
882
PstS2-15
2,116.8
1,323
1,499.4
999.6
558.6
970.2
852.6
PstS3-1
2,146.2
1,440.6
1,587.6
999.6
470.4
999.6
911.4
PstS3-2
2,116.8
1,440.6
1,675.8
940.8
441
1,058.4
793.8
PstS3-3
2,175.6
1,352.4
1,528.8
940.8
558.6
940.8
852.6
PstS3-4
2,175.6
1,470
1,499.4
970.2
470.4
999.6
911.4
PstS3-5
2,146.2
1,440.6
1,587.6
999.6
529.2
999.6
940.8
PstS3-6
2,028.6
1,440.6
1,587.6
999.6
470.4
1,058.4
911.4
PstS3-7
2,146.2
1,440.6
1,617
940.8
441
1,029
911.4
PstS3-8
2,205
1,528.8
1,617
911.4
470.4
1,058.4
970.2
PstS3-9
2,116.8
1,470
1,558.2
970.2
470.4
1,029
911.4
PstS3-10
2,175.6
1,499.4
1,646.4
911.4
499.8
1,029
940.8
PstS3-11
2,087.4
1,411.2
1,470
852.6
529.2
999.6
882
PstS3-12
2,087.4
1,470
1,528.8
911.4
499.8
999.6
911.4
PstS3-13
2,175.6
1,470
1,528.8
940.8
529.2
1,029
911.4
PstS3-14
2,352
1,558.2
1,764
1,146.6
529.2
1,146.6
999.6
PstS3-15
2,410.8
1,528.8
1,705.2
1,264.2
529.2
1,117.2
1,058.4
Tabla 7
Medidas en μm de cada huevo de Pseudosermylecarinulata, Pseudosermyletridens, Pseudosermyle sp. 4, Pseudosermyle sp. 5 utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PtrGtoU-1
3,580.2
1,285.2
1,514.7
642.6
596.7
872.1
688.5
PtrGtoU-2
3,626.1
1,239.3
1,560.6
642.6
550.8
872.1
688.5
PtrGtoU-3
3,626.1
1,377
1,606.5
642.6
596.7
872.1
688.5
PtrGtoU-4
3,626.1
1,377
1,468.8
688.5
596.7
872.1
688.5
PtrGtoU-5
3,580.2
1,285.2
1,560.6
734.4
550.8
826.2
734.4
PtrGtoU-6
3,580.2
1,331.1
1,560.6
688.5
550.8
872.1
688.5
PtrGtoU-7
3,672
1,331.1
1,514.7
688.5
596.7
826.2
734.4
PtrGtoU-8
3,672
1,377
1,422.9
688.5
550.8
872.1
642.6
PtrGtoU-9
3,672
1,377
1,514.7
688.5
550.8
872.1
688.5
PtrGtoU-10
3,580.2
1,377
1,377
459
459
872.1
642.6
PtrGtoU-11
3,580.2
1,285.2
1,422.9
321.3
229.5
872.1
688.5
PtrGtoU-12
3,672
1,331.1
1,422.9
596.7
459
826.2
688.5
PtrGtoU-13
3,580.2
1,239.3
1,422.9
688.5
550.8
872.1
734.4
PtrGtoU-14
3,534.3
1,331.1
1,514.7
642.6
596.7
826.2
688.5
PtrGtoU-15
3,488.4
1,331.1
1,606.5
688.5
596.7
872.1
734.4
PtrMorT1-1
3,488.4
1,285.2
1,422.9
596.7
504.9
734.4
780.3
PtrMorT1-2
3,626.1
1,331.1
1,514.7
504.9
504.9
734.4
780.3
PtrMorT1-3
3,717.9
1,377
1,514.7
550.8
504.9
734.4
780.3
PtrMorT1-4
3,580.2
1,331.1
1,468.8
504.9
504.9
780.3
780.3
PtrMorT1-5
3,534.3
1,377
1,468.8
550.8
504.9
780.3
826.2
PtrMorT1-6
3,672
1,331.1
1,468.8
550.8
504.9
734.4
780.3
PtrMorT1-7
3,580.2
1,331.1
1,377
459
504.9
734.4
734.4
PtrMorT1-8
3,580.2
1,377
1,377
596.7
504.9
642.6
734.4
PtrMorT1-9
3,442.5
1,331.1
1,422.9
550.8
550.8
780.3
826.2
PtrMorT1-10
3,442.5
1,239.3
1,377
550.8
550.8
780.3
826.2
PtrMorT2-1
3,549.6
1,326
1,433.1
540.6
515.1
744.6
785.4
PtrMorT2-2
3,488.4
1,239.3
1,422.9
688.5
550.8
734.4
780.3
PtrMorT2-3
3,626.1
1,147.5
1,468.8
642.6
367.2
734.4
734.4
PtrMorT2-4
3,442.5
1,147.5
1,331.1
688.5
550.8
734.4
780.3
PtrMorT2-5
3,717.9
1,285.2
1,514.7
688.5
550.8
734.4
780.3
PtrMorT2-6
3,717.9
1,239.3
1,468.8
688.5
550.8
734.4
734.4
PtrMorT2-7
3,626.1
1,239.3
1,422.9
688.5
596.7
688.5
780.3
PtrMorT2-8
3,717.9
1,285.2
1,468.8
642.6
550.8
826.2
780.3
PtrMorT2-9
3,717.9
1,239.3
1,422.9
688.5
596.7
734.4
780.3
PtrMorT2-10
3,672
1,193.4
1,514.7
642.6
596.7
734.4
780.3
PcaC1-1
1,999.2
1,381.8
1,587.6
1,029
382.2
940.8
823.2
Tabla 7. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PcaC1-2
1,999.2
1,411.2
1,587.6
999.6
382.2
970.2
823.2
PcaC1-3
2,028.6
1,381.8
1,587.6
1,029
411.6
940.8
823.2
PcaC1-4
2,058
1,352.4
1,558.2
940.8
382.2
940.8
852.6
PcaC1-5
1,969.8
1,323
1,558.2
940.8
382.2
940.8
823.2
PcaC2-1
1,852.2
1,176
1,381.8
940.8
441
823.2
735
PcaC2-2
1,822.8
1,264.2
1,440.6
911.4
411.6
882
764.4
PspFU-1
2,058
1,323
1,558.2
1,117.2
411.6
999.6
823.2
PspFU-2
2,028.6
1,323
1,499.4
1,234.8
382.2
999.6
793.8
PspFU-3
2,087.4
1,352.4
1,587.6
1,146.6
382.2
911.4
735
Tabla 8
Medidas en μm de cada huevo de N. isabelae, Pseudosermyle sp. 1, Pseudosermyle sp. 2. Pseudosermyle sp. 5 utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PspFU-4
1,822.8
1,146.6
1,440.6
940.8
411.6
970.2
676.2
PspFU-5
1,940.4
1,264.2
1,499.4
970.2
382.2
911.4
676.2
PspFU-6
1,881.6
1,234.8
1,470
1,029
411.6
940.8
705.6
PspFU-7
1,940.4
1,293.6
1,470
940.8
411.6
911.4
705.6
PspFU-8
1,911
1,205.4
1,499.4
911.4
382.2
970.2
764.4
PspFU-9
1,852.2
1,205.4
1,470
940.8
382.2
970.2
764.4
PspFU-10
2,058
1,323
1,499.4
1,087.8
382.2
999.6
735
PspFU-11
2,087.4
1,323
1,499.4
1,058.4
382.2
999.6
793.8
PspFU-12
1,999.2
1,323
1,470
1,205.4
382.2
970.2
764.4
PspFU-13
1,881.6
1,234.8
1,440.6
1,058.4
411.6
911.4
793.8
PspFU-14
2,058
1,323
1,528.8
1,087.8
382.2
1,029
764.4
PspFU-15
1,940.4
1,293.6
1,499.4
999.6
382.2
999.6
705.6
NisaU-1
2,263.8
1,499.4
1,764
1,029
617.4
1,293.6
970.2
NisaU-2
2,293.2
1,470
1,705.2
999.6
588
1,146.6
882
NisaU-3
2,293.2
1,646.4
1,793.4
1,029
617.4
1,234.8
1,029
NisaU-4
2,175.6
1,558.2
1,705.2
999.6
558.6
1,205.4
1,029
NisaU-5
2,263.8
1,646.4
1,793.4
999.6
588
1,234.8
1,029
NisaU-6
2,146.2
1,470
1,558.2
1,087.8
558.6
1,117.2
940.8
NisaU-7
2,293.2
1,558.2
1,793.4
1,029
588
1,234.8
1,029
NisaU-8
2,146.2
1,440.6
1,617
999.6
558.6
1,087.8
940.8
NisaU-9
2,175.6
1,528.8
1,617
1,087.8
558.6
1,146.6
911.4
NisaU-10
2,293.2
1,587.6
1,793.4
970.2
588
1,176
911.4
Tabla 8. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
NisaU-11
2,116.8
1,558.2
1,793.4
970.2
617.4
1,234.8
1,058.4
NisaU-12
2,352
1,499.4
1,764
1,058.4
646.8
1,293.6
1,029
PtgVBU-1
3,394.5
1,569.5
1,715.5
1,533
657
1,277.5
912.5
PtgVBU-2
3,029.5
1,350.5
1,606
1,350.5
511
912.5
620.5
PtgVBU-3
3,467.5
1,533
1,752
1,496.5
730
1,204.5
803
PtgVBU-4
3,394.5
1,533
1,606
1,496.5
657
1,095
803
PtgVBU-5
3,321.5
1,496.5
1,642.5
1,533
620.5
1,204.5
912.5
PtgVBU-6
3,431
1,460
1,642.5
1,460
584
1,095
803
PtgVBU-7
3,431
1,460
1,642.5
1,460
657
1,241
839.5
PtgVBU-8
3,394.5
1,533
1,569.5
1,533
693.5
1,095
839.5
PtgVBU-9
3,467.5
1,533
1,715.5
1,460
620.5
1,095
766.5
PtgVBU-10
3,394.5
1,533
1,715.5
1,496.5
730
1,168
912.5
PtgVBU-11
3,285
1,606
1,679
1,460
620.5
1,204.5
876
PtgDFG1-1
3,321.5
1,533
1,715.5
1,095
730
985.5
766.5
PtgDFG1-2
3,102.5
1,533
1,606
1,131.5
693.5
839.5
693.5
PtgDFG1-3
3,285
1,533
1,642.5
1,204.5
730
912.5
766.5
PtgDFG1-4
3,102.5
1,460
1,533
1,204.5
693.5
949
766.5
PtgDFG1-5
3,248.5
1,569.5
1,569.5
1,168
766.5
803
657
PtgDFG2-1
3,248.5
1,533
1,642.5
1,241
657
949
620.5
PtgDFG2-2
3,285
1,496.5
1,606
1,095
620.5
949
730
PtgDFG2-3
3,248.5
1,496.5
1,569.5
1,350.5
730
949
803
PtgDFG2-4
3,285
1,606
1,715.5
1,314
876
912.5
730
PtgDFG2-5
3,102.5
1,606
1,752
1,241
876
949
730
Tabla 9
Medidas en μm de cada huevo de Pseudosermyle sp. 2. Pseudosermyle sp. 3, Pseudosermyle sp. 6 utilizadas para el PCA.
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PtgDFG2-6
3,285
1,496.5
1,606
1,314
620.5
912.5
657
PtgDFG2-7
3,212
1,460
1,569.5
1,277.5
620.5
912.5
657
PtgDFG2-8
3,321.5
1,533
1,679
1,277.5
657
985.5
693.5
PtgDFG2-9
3,212
1,569.5
1,679
1,314
876
912.5
766.5
PtgDFG2-10
3,175.5
1,606
1,752
1,387
657
876
657
PtgDFG2-11
3,467.5
1,606
1,715.5
1,241
657
949
657
PtgDFG2-12
3,248.5
1,606
1,642.5
1,277.5
693.5
985.5
839.5
PtgDFG2-13
3,248.5
1,606
1,642.5
1,241
693.5
985.5
657
Tabla 9. Continúa
Clave
Medidas de huevo
L
W
H
mpl
mpw
oph
opw
PtgDFG2-14
3,285
1,496.5
1,642.5
1,460
730
949
803
PchF1-1
3,626.1
1,147.5
1,422.9
1,239.3
688.5
918
780.3
PchF1-2
3,993.3
1,468.8
1,652.4
1,422.9
734.4
1,055.7
826.2
PchF1-3
3,947.4
1,514.7
1,606.5
1,468.8
826.2
1,101.6
872.1
PchF2-1
3,947.4
1,377
1,698.3
1,377
688.5
918
780.3
PchF2-2
3,672
1,377
1,560.6
1,377
780.3
872.1
688.5
PchF2-3
3,672
1,422.9
1,606.5
1,422.9
780.3
872.1
688.5
PoaxU-1
2,701
1,642.5
1,898
693.5
328.5
1,314
1,131.5
PoaxU-2
2,518.5
1,569.5
1,825
693.5
365
1,277.5
1,095
PoaxU-3
2,664.5
1,642.5
1,861.5
657
328.5
1,277.5
1,058.5
PoaxU-4
2,664.5
1,679
1,825
657
365
1,277.5
1,131.5
PoaxU-5
2,664.5
1,642.5
1,861.5
730
365
1,277.5
1,131.5
PoaxU-6
2,518.5
1,606
1,788.5
693.5
365
1,241
1,095
PoaxU-7
2,555
1,606
1,861.5
657
365
1,350.5
1,131.5
PoaxU-8
2,591.5
1,642.5
1,861.5
766.5
328.5
1,277.5
1,131.5
PoaxU-9
2,482
1,569.5
1,752
657
365
1,168
949
PoaxU-10
2,591.5
1,569.5
1,861.5
657
365
1,241
1,095
PoaxU-11
2,482
1,679
1,788.5
693.5
365
1,277.5
1,058.5
PoaxU-12
2,518.5
1,569.5
1,788.5
693.5
328.5
1,277.5
1,095
PoaxU-13
2,555
1,606
1,861.5
693.5
328.5
1,241
1,022
PoaxU-14
2,555
1,679
1,788.5
693.5
365
1,241
1,022
PoaxU-15
2,701
1,679
1,898
657
292
1,314
1,131.5
Se llevó a cabo un análisis de componentes principales (PCA) para examinar si las medidas —en micras— de los huevos eran relevantes al reconocer grupos. Las medidas utilizadas fueron: longitud de la cápsula (L), amplitud de la cápsula (W), altitud de la cápsula (H), longitud de la lámina micropilar (mpl), amplitud de la lámina micropilar (mpw), altitud del opérculo (oph) y amplitud del opérculo (opw) (tablas 4-9). Se incluyeron todos los huevos con el propósito de graficar estas mediciones y agruparlos según dichas medidas. Se empleó R (R Core Team, 2013) mediante la interfaz de Rstudio (Rstudio Team, 2015); las paqueterías utilizadas fueron: ggplot2 (Wickham, 2016), ggfortify (Horikoshi y Tang, 2016), rgl (Adler et al., 2018), pca3d (Weiner, 2017) y magick (Ooms, 2018). Cada huevo fue caracterizado de manera individual siguiendo un código que incluye los nombres de las columnas: nombre, clave de hembra de la tabla 3, más el número ascendente correspondiente (tablas 4-9). Para la descripción de los grupos, las medidas se convirtieron a milímetros (mm), mientras que para cada especie se mantuvieron en micras (μm).
Resultados
El análisis de componentes principales (PCA) muestra la formación de 4 grupos (fig. 3) explicados en detalle posteriormente, que coinciden con los patrones de morfología externa mostrada por los adultos (machos y hembras) y el tipo de oviposición. En el gráfico del PCA cada eje representa un componente; por lo tanto, se representan 2 gráficos del PCA, uno con rotación en el eje del primer componente (fig. 3A) y otro con rotación en el eje del segundo componente (fig. 3B), para una apreciación mejor de los 4 grupos. Los primeros 3 componentes explican 96.76% de la varianza (fig. 4A). El primer componente presenta mayor contribución de L (fig. 4B), mientras que el segundo presenta mayor homogeneidad en la contribución que recibe de las variables; sin embargo, la oph es la variable que más contribuye (fig. 4C). Finalmente, el tercer componente presenta mayor contribución de mpl (fig. 4D).
A continuación, se da una descripción breve de la microestructura coriónica general de cada grupo, así como para cada una de las especies incluidas en este trabajo. También se incluye una comparación de las distintas estructuras coriónicas entre los grupos y las especies, como se observa en las tablas 10-12. Muchas de estas especies no están descritas y pueden representar especies aún inéditas e innominadas, en consecuencia, se dejaron como morfoespecies y solo se enumeraron.
Tabla 10
Comparación entre estructuras coriónicas entre especies y grupos. Longitud de cápsula (L), amplitud de cápsula (W), altitud de cápsula (H).
Grupos
General
Cápsula
Tipo de huevo
Oviposición
Forma
Textura
Microestructura
L promedio (mm)
W promedio (mm)
H promedio (mm)
Grupo 1
adhesivo
Adheridos
Fusiforme
Gránulos
Basamentos piramidales
3.37 ±0.23
1.5 ±0.09
1.64 ±0.07
Pseudosermyle sp. 1
adhesivo
Adheridos
Fusiforme
Gránulos
Basamentos piramidales
3.36 ±0.12
1.5 ±0.06
1.66 ±0.05
Pseudosermyle sp. 2
adhesivo
Adheridos
Fusiforme
Gránulos
Basamentos piramidales
3.24 ±0.08
1.54 ±0.05
1.64 ±0.06
Pseudosermyle sp. 3
adhesivo
Adheridos
Fusiforme
Gránulos
Basamentos piramidales
3.8 ±0.16
1.38 ±0.12
1.59 ±0.09
Grupo 2
adhesivo
Ooteca
Fusiforme
Paralelepípedos
Liso
3.6 ±0.08
1.3 ±0.06
1.46 ±0.06
Pseudosermyle tridens
adhesivo
Ooteca
Fusiforme
Paralelepípedos
Liso
3.59 ±0.09
1.28 ±0.07
1.44 ±0.05
Pseudosermyle sp. 4
Adhesivo
Ooteca
Fusiforme
Paralelepípedos
Liso
3.6 ±0.05
1.32 ±0.04
1.49 ±0.07
Grupo 3
Alveolada
Por caída
Doliforme
Variable
Variable
2.14 ±0.14
1.44 ±0.11
1.61 ±0.1
Pseudosermyle carinulata
Alveolada
Por caída
Doliforme
Gránulos
Proyecciones digitiformes
1.96 ±0.08
1.32 ±0.08
1.52 ±0.08
Pseudosermyle striata
Alveolada
Por caída
Doliforme
Arrugas
Proyecciones digitiformes
2.21 ±0.09
1.48 ±0.06
1.64 ±0.07
Pseudosermyle sp. 5
Alveolada
Por caída
Doliforme
Arrugas
Rugulosa
1.96 ±0.08
1.27 ±0.05
1.49 ±0.03
Nooxapty isabelae
Alveolada
Por caída
Doliforme
Espinas
Proyecciones digitiformes
2.23 ±0.07
1.53 ±0.06
1.72 ±0.08
Grupo 4
Alveolada
Por caída
Doliforme
Variable
Variable
2.68 ±0.09
1.69 ±0.08
1.87 ±0.08
Pseudosermyle phalangiphora
Alveolada
Por caída
Doliforme
Gibas con espinas umbeliformes
Punteada
2.68 ±0.1
1.65 ±0.08
1.82 ±0.08
Pseudosermyle procera
Alveolada
Por caída
Doliforme
Verrugosa con fóveas
Verrugosa
2.71 ±0.06
1.74 ±0.05
1.93 ±0.05
Pseudosermyle sp. 6
Alveolar
Por caída
Doliforme
Coliculada con espinas umbeliformes
Verrugosa
2.58 ±0.07
1.62 ±0.04
1.83 ±0.04
Tabla 11
Comparación entre estructuras coriónicas entre especies y grupos. Porcentaje de cobertura de la longitud de lámina micropilar en la longitud de la cápsula (% de mpl/L).
Grupos
Opérculo
Lámina micropilar (mp)
Ornamentación
Ángulo opercular
Capas
Forma
Ornamentación
% de mpl/L
Relieve micropilar
Forma lámina micropilar interna
Grupo 1
Variable
+ 55°
1
Lemniscata
Similar a la cápsula
40
Variable
Abierta con línea media
Pseudosermyle sp. 1
Estructura solitaria
+ 55°
1
Lemniscata
Similar a la cápsula
43.98
Presen-te
Abierta con línea media
Pseudosermyle sp. 2
Similar a la cápsula
+ 55°
1
Lemniscata
Similar a la cápsula
38.63
Ausente
Abierta con línea media
Pseudosermyle sp. 3
Disposición radial
+ 55°
1
Lemniscata
Similar a la cápsula
36.34
Presen-te
Abierta con línea media
Grupo 2
Similar a la cápsula
+ 40°
2
Variable
Distinta a la cápsula
17
Ausente
Paralela sin línea media
Pseudosermyle tridens
Similar a la cápsula
+ 40°
2
Triangular
Distinta a la cápsula
16.7
Ausente
Paralela sin línea media
Pseudosermyle sp. 4
Similar a la cápsula
+ 40°
2
Circular
Distinta a la cápsula
17.57
Ausente
Paralela sin línea media
Grupo 3
Similar a la cápsula
≈ 0°
1
Oblonga
Similar a la cápsula
50
Ausente
Abierta con línea media
Pseudosermyle carinulata
Similar a la cápsula
≈ 0°
1
Oblonga
Similar a la cápsula
49.46
Ausente
Abierta con línea media
Pseudosermyle striata
Similar a la cápsula
≈ 0°
1
Oblonga
Similar a la cápsula
47.51
Ausente
Abierta con línea media
Pseudosermyle sp. 5
Similar a la cápsula
≈ 0°
1
Oblonga
Similar a la cápsula
53.23
Ausente
Abierta con línea media
Nooxapty isabelae
Similar a la cápsula
≈ 0°
1
Oblonga
Similar a la cápsula
45.72
Ausente
Abierta con línea media
Grupo 4
Acomodo de estructuras
≈ 0°
1
Variable
Variable
30
Varia-ble
Abierta con línea media
Pseudosermyle phalangiphora
Estructura solitaria
≈ 0°
1
Oblonga
Distinta a la cápsula
30.885
Ausente
Abierta con línea media
Pseudosermyle procera
Anillo
≈ 0°
1
Elíptica
Distinta a la cápsula
25.11
Presen-te
Abierta con línea media
Pseudosermyle sp. 6
Anillo
≈ 0°
1
Oblonga
Similar a la cápsula
26.55
Ausente
Abierta con línea media
Tabla 12
Comparación entre estructuras coriónicas entre especies y grupos. Lámina micropilar (mp).
Grupos
Borde
Tubérculo micropilar
Copa micropilar
Línea media externa
Forma
Posición
Forma
Posición
Presencia
Ornamentación
Longitud
Grupo 1
Carina que se ensancha hacia la parte posterior
Hacia la copa micropilar
Variable
Fusionada al borde de mp
Presente
Variable
Corta (no llega al área polar)
Pseudosermyle sp. 1
Carina que se ensancha hacia la parte posterior
Hacia la copa micropilar
Triángulo invertido
Fusionada al borde de mp
Presente
Carina
Corta (no llega al área polar)
Pseudosermyle sp. 2
Carina que se ensancha hacia la parte posterior
Hacia la copa micropilar
Triángulo invertido
Fusionada al borde de mp
Presente
Cambio de textura
Corta (no llega al área polar)
Pseudosermyle sp. 3
Carina que se ensancha hacia la parte posterior
Hacia la copa micropilar
Matraz invertido
Fusionada al borde de mp
Presente
Sucesión de estructuras
Corta (no llega al área polar)
Grupo 2
Variable
En la mitad de la copa micropilar
Carina
Ausente
Ausente
No aplica
No aplica
Pseudosermyle tridens
Cambio de textura
En la mitad de la copa micropilar
Carina
Ausente
Ausente
No aplica
No aplica
Pseudosermyle sp. 4
Ligera elevación
En la mitad de la copa micropilar
Carina
Ausente
Ausente
No aplica
No aplica
Grupo 3
Variable
Hacia la copa micropilar
Variable
Separada del borde de mp
Presente
Variable
Variable
Pseudosermyle carinulata
Surco
Hacia la copa micropilar
Rectangular
Separada del borde de mp
Presente
Carina
Larga (llega al área polar)
Pseudosermyle striata
Carina
Hacia la copa micropilar
Triángulo invertido
Separada del borde de mp
Presente
Carina
Larga (llega al centro del área polar)
Pseudosermyle sp. 5
Superficie lisa
Hacia la copa micropilar
Rectangular
Separada del borde de mp
Presente
Carina
Larga (llega al área polar)
Nooxapty isabelae
Carina
Hacia la copa micropilar
Forma de y
Separada del borde de mp
Presente
Sucesión de estructuras
Larga (llega al centro del área polar)
Grupo 4
Carina
Hacia la copa micropilar
Variable
Fusionada al borde de mp
Variable
Variable
Variable
Tabla 12. Continúa
Grupos
Borde
Tubérculo micropilar
Copa micropilar
Línea media externa
Forma
Posición
Forma
Posición
Presencia
Ornamentación
Longitud
Pseudosermyle phalangiphora
Carina
Hacia la copa micropilar
Semiesfera
Fusionada al borde de mp
Presente
Carina
Larga (llega al área polar)
Pseudosermyle procera
Carina
Hacia la copa micropilar
Semiesfera
Fusionada al borde de mp
Ausente
No aplica
No aplica
Pseudosermyle sp. 6
Carina
Hacia la copa micropilar
Triángulo invertido
Fusionada al borde de mp
Presente
Sucesión de estructuras
Larga (llega al área polar)
Figura 3. Gráfica de los 3 primeros componentes del PCA, que representan 96.76% de la varianza explicada. A, Vista rotando el eje del componente 1; B, vista rotando el eje del componente 2. Grupo 1: Pseudosermyle sp. 1 (rosa), Pseudosermyle sp. 2 (rojo) y Pseudosermyle sp. 3 (lila). Grupo 2: P. tridens (naranja oscuro) y Pseudosermyle sp. 4 (aguamarina). Grupo 3: P. carinulata (amarillo), P. striata (verde), Pseudosermyle sp. 5 (cian) y Nooxapty isabelae (naranja claro). Grupo 4: P. phalangiphora (azul), P. procera (morado)y Pseudosermyle sp. 6 (azul celeste).
Pseudosermyle grupo 1
Huevo: adhesivo, adherido individualmente sobre una superficie (fig. 5B). Medidas en mm (n = 36): L = 3.37 ± 0.23 (3.02-3.99), W = 1.5 ± 0.09 (1.14-1.6), H = 1.64 ± 0.07 (1.42-1.75), mpl = 1.34 ± 0.12 (1.09-1.53), mpw = 0.69 ± 0.08 (0.51-0.87), oph = 0.99 ± 0.12 (0.8-1.27) y opw = 0.76 ± 0.08 (0.62-0.91). Opérculo orientado en + 55° aproximadamente. Lámina micropilar en forma de lemniscata u oblonga con el centro constreñido; con una longitud aproximada entre más de un tercio y menos de la mitad de la longitud de la cápsula. Lámina micropilar interna abierta con línea media (fig. 6A-C).
Material examinado: 11 huevos. México. Edo. de México, Valle de Bravo, Arriba cueva, 19°5’32.928” N, 100°4’19.848” O, 2,260 m snm [voucher: PHMX 349-358 depositados en CNIN y MZFC].
Figura 4. Gráficas de PCA. A, Proporción de la varianza explicada en porcentaje por cada componente; B, contribución de las variables al componente 1; C, contribución de las variables al componente 2; D, contribución de las variables al componente 3. La línea punteada roja indica el valor promedio que cada variable aportaría al componente si estas contribuciones fueran equitativas.Figura 5. Tipos de oviposición encontrados en Pseudosermyle. A, Por caída; B, adheridos a una superficie; C, ooteca.Figura 6. Diversidad de formas en la lámina micropilar interna (mpi) de las distintas especies de Pseudosermyle. A-C, Grupo 1; D-E, grupo 2; F-I, grupo 3; J-L, grupo 4. A-B y F-L, mpi abierta con línea media (NM); D-E, mpi paralela sin línea media (PO). A, Pseudosermyle sp. 1; B, Pseudosermyle sp. 2; C, Pseudosermyle sp. 3; D, P. tridens; E, Pseudosermyle sp. 4; F, P. carinulata; G, P. striata; H, Pseudosermyle sp. 5; I, Nooxapty isabelae; J, P. phalangiphora; K, P. procera; L. Pseudosermyle sp. 6.
Figura 7. Diversidad de formas en los huevos de las distintas especies de Pseudosermyle, huevo en vista dorsal. A, Pseudosermyle sp. 1; B, Pseudosermyle sp. 2; C, Pseudosermyle sp. 3; D, P. tridens; E, Pseudosermyle sp. 4; F, P. carinulata; G, P. striata; H, Pseudosermyle sp. 5; I, Nooxapty isabelae; J, P. phalangiphora; K, P. procera; L, Pseudosermyle sp. 6.
Microestructura: (n = 1) (figs. 8, 9) cápsula de superficie granulosa (fig. 8A, B), con granos formados por basamentos piramidales de 6-8 niveles (fig. 8E, F); la longitud varía entre 37.18-59.04 μm. En toda la superficie se observa un patrón de terrazas (polígonos irregulares y escalonados, en su mayoría rectangulares), que pueden o no formar los gránulos (fig. 8E). El huevo suele estar cubierto (cuando es ovipuesto) por una sustancia adherente o pegajosa que presenta rugosidades irregulares y separadas, suele cubrir la mitad ventral (fig. 8C, D); las arrugas son lisas (fig. 8D). Lámina micropilar con ornamentación y textura similar a la cápsula (fig. 9A). Borde a manera de una carina con 66 μm de amplitud (fig. 9D), tiende a ensancharse hacia la copa micropilar (fig. 9C). Tubérculo micropilar como un triángulo invertido, con 286 μm de longitud y 198 μm de amplitud (fig. 9C). Copa micropilar con forma de ‘y’, fusionada al borde, con 362 μm de amplitud y 39 μm de grosor (fig. 9C). Línea media como una carina de 40 μm de amplitud (figs. 8C, 9B). Las estructuras antes mencionadas, salvo la superficie de la lámina micropilar (fig. 9E), son de textura rugosa. Opérculo de superficie rugosa, con arrugas que forman una elevación a manera de semicírculo en la región anterior; la textura de las arrugas es verrugosa (fig. 9F).
Pseudosermyle sp. 2 (figs. 1, 6B, 7B, 10, 11)
Material examinado:19 huevos. México, Ciudad de México, Coyoacán, REPSA, Espacio Escultórico-Caseta de vigilancia, 19°19’3.4824” N, 99°11’5.4744” O, 2,311 m snm [voucher: PHMX 304-322 depositados en CNIN y MZFC].
Figura 8. Pseudosermyle sp. 1 cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, límite entre superficie de la cápsula y pegamento; E, superficie de la cápsula; F, acercamiento a la textura de los gránulos.
Microestructura: (n = 1) (figs. 10, 11) cápsula de superficie coliculado-granulosa (fig. 10A, B), con granos formados por basamentos piramidales de 4-6 niveles; la longitud varía entre 24.66-34.39 μm (fig. 10E, F). En toda la superficie se presenta un patrón de polígonos irregulares y escalonados —en su mayoría rectangulares, que pueden o no formar los gránulos (fig. 10E); en ciertas regiones hay máculas irregulares de textura escabriculosa. El huevo suele estar cubierto (cuando es ovipuesto) por una sustancia pegajosa que muestra rugosidades irregulares y muy cercanas entre sí, suele cubrir la mitad ventral (fig. 10B, C); las arrugas son lisas (fig. 10D). Lámina micropilar con ornamentación similar al patrón presente en la superficie de la cápsula (fig. 11A), con secciones irregulares de textura escabriculosa (fig. 11E). Borde como una carina de 73 μm de amplitudl (fig. 11D) y tiende a ensancharse hacia la copa micropilar (fig. 11C), con textura papilada (fig. 11D). Tubérculo micropilar como un triángulo invertido, con 282 μm de longitud y 136 μm de amplitud (fig. 11C); textura similar a la presente en la superficie de la cápsula (fig. 11C). Copa micropilar con forma de ‘v’, fusionada al borde, con 321 μm de amplitud y 45 μm de grosor, de superficie rugosa (fig. 11A, C). Línea media con 533 μm de longitud y 61 μm de amplitud (figs. 10C, 11B); de textura tuberculosa al centro y escabriculosa en la unión con la cápsula (fig. 11B) – esta línea media exhibe un cambio de texturas en la cápsula. Opérculo de textura rugulosa que forma secciones poligonales de grupos de protuberancias (fig. 11F).
Pseudosermyle sp. 3 (figs. 1, 6C, 7C, 12, 13)
Material examinado: 6 huevos. México, Jalisco, La Huerta, alrededores EB Chamela, 19°29’57.516” N, 105°2’41.424” O, 87 m snm [voucher: PHMX 323-328 depositados en CNIN y MZFC].
Figura 9. Pseudosermyle sp. 1 huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, opérculo.
Microestructura: (n = 1) (figs. 12, 13) cápsula de superficie rugosa, con arrugas irregulares de longitud 54.04-148.04 μm y amplitud de 28.19-53.26 μm (fig. 12A, B); unidas por una especie de membrana (fig. 12D). Los espacios entre las arrugas están saturados de gránulos con diámetro 28.19-42.29 μm (fig. 12D, F), éstos suelen estar muy juntos hacia el dorso y más separados hacia los lados; en las superficies laterales se ven algunos niveles, como si fuesen basamentos piramidales (fig. 12E). El huevo suele estar cubierto (cuando ovipuesto) por una sustancia adherente que exhibe rugosidades irregulares y muy cercanas, suele cubrir la mitad ventral (fig. 12B, C); las arrugas son lisas (fig. 12D). Lámina micropilar con ornamentación y textura similar a la cápsula, salvo que las arrugas están hacia el centro (fig. 13E) y los gránulos hacia la periferia (fig. 13A).
Figura 10. Pseudosermyle sp. 2 cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, límite entre superficie de la cápsula y pegamento; E, superficie de la cápsula; F, acercamiento a la textura de los gránulos.
Borde de textura escabriculosa elevada a manera de carina, con 77 μm de amplitud (fig. 13D). Tubérculo micropilar en forma de matraz invertido con 381 μm de longitud y 237 μm de amplitud, de textura rugosa (fig. 13C). Copa micropilar con forma de ‘v’, fusionada al borde, con 437 μm de amplitud y 97 μm de grosor, de textura rugosa (fig. 13C). Línea media con 351 μm de longitud y 98 de amplitud; como una serie de arrugas elevadas con una textura escabriculosa en el centro (figs. 12C, 13B). Opérculo con espacio entre las arrugas radiales; hacia la periferia es granuloso como la cápsula y hacia el centro de las arrugas la textura es rugosa, pero sin formar arrugas del tamaño y forma a las que rodean esta zona (fig. 13F).
Pseudosermyle grupo 2
Huevo: adhesivo con oviposición en ooteca (fig. 5C). Medidas en mm (n = 35): L = 3.6 ± 0.08 (3.44-3.71), W = 1.3 ± 0.06 (1.14-1.37), H = 1.46 ± 0.06 (1.33-1.6), mpl = 0.61 ± 0.09 (0.32-0.73), mpw = 0.53 ± 0.07 (0.22-0.59), oph = 0.79 ± 0.06 (0.64-0.87) y opw = 0.74 ± 0.05 (0.64-0.82). Opérculo orientado hacia la superficie dorsal en + 40° aproximadamente. Lámina micropilar reducida, con una longitud cercana a un sexto de la longitud de la cápsula. Lámina micropilar interna paralela sin línea media (fig. 6D, E).
Especies incluidas:Pseudosermyle tridens (Burmeister, 1838) y Pseudosermyle sp. 4.
Material examinado: 20 huevos disecados y 3 ootecas. México, Morelos, Ayala, Venustiano Carranza esquina Iturbide, Cruz Verde, 18°45’36.6264” N, 98°58’33.9528” O, 1,295 m snm [voucher: PHMX 329-348, 616-618 depositados en CNIN y MZFC].
Figura 11. Pseudosermyle sp. 2 huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, opérculo.
Microestructura: (n = 1) (figs. 14, 15) cápsula con superficie dorsal cubierta por paralelepípedos (fig. 14A, B) con longitudes de arista entre 6-9.5 μm (fig. 14E, F). En las superficies laterales y ventrales es lisa (fig. 14C, D). Lámina micropilar en general con textura lisa hacia el centro (fig. 15A); con tendencia a formar fibras que se vuelven una retícula hacia la periferia (fig. 15B). Tubérculo micropilar lineal, liso, con 273 μm de longitud y 108 μm de amplitud aproximadamente (fig. 15C). El borde es el límite entra las texturas de la lámina micropilar y la cápsula. Sin copa micropilar, ni línea media. Opérculo de textura como en la cápsula (fig. 15D).
Pseudosermyle sp. 4 (figs. 1, 6E, 7E, 16, 17)
Material examinado: 15 huevos disecados y 3 ootecas. México, Guanajuato, Silao de la Victoria, Villa Esmeralda, 20°57’13.212” N, 101°26’0.996” O, 1,782 m snm [voucher: PHMX 359-373, 423-425 depositados en CNIN y MZFC].
Microestructura: (n = 1) (figs. 16, 17) cápsula de superficie dorsal cubierta por paralelepípedos (fig. 16A, B), con longitudes de arista entre 1-3 μm (fig. 16E, F). En las superficies laterales y ventral con una reducción gradual de los paralelepípedos (fig. 16C, D). Lámina micropilar en general de textura lisa hacia el tubérculo micropilar; hacia el borde es rugosa (fig. 17A). Tubérculo micropilar liso, con 321 μm de longitud y 136 μm de amplitud aproximadamente (fig. 17C).
Figura 12. Pseudosermyle sp. 3 cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, límite entre superficie de la cápsula y pegamento; E, superficie de la cápsula; F, acercamiento a la textura de los gránulos.
El borde es una ligera elevación que rodea la lámina micropilar y se encuentra abierto en el área polar (figs. 16C, 17A); su textura es rugosa (fig.17B), de aproximadamente 64 μm de amplitud. Sin copa micropilar, ni línea media (figs. 16C, 17A). Opérculo de textura como en la cápsula (fig. 17D).
Pseudosermyle grupo 3
Huevo: alveolar, de oviposición por caída (fig. 5A), doliforme. Medidas en mm (n = 79): L = 2.14 ± 0.14 (1.82-2.41), W = 1.44 ± 0.11 (1.14-1.64), H = 1.61 ± 0.1 (1.38-1.79), mpl = 1.03 ± 0.09 (0.85-1.26), mpw = 0.5 ± 0.07 (0.38-0.64), oph = 1.05 ± 0.09 (0.82-1.29) y opw = 0.89 ± 0.09 (0.67-1.05). Lámina micropilar en general oblonga y con una longitud cercana a la mitad de la longitud de la cápsula. Lámina micropilar interna abierta con línea media (fig. 6F-I).
Especies incluidas:Pseudosermyle carinulata (Brunner von Wattenwyl, 1907), P. striata (Burmeister, 1838), Pseudosermyle sp. 5 y se incluye a Nooxapty isabelae López-Mora et Llorente-Bousquets, 2023 como parte de este grupo debido a su similitud coriónica, aunque la microestructura es más especializada.
Material examinado: 7 huevos. México, Edo. de México, Valle de Bravo, bajo mesa, 19°5’9.96” N, 100°4’35.94” O, 2, 201 m snm [voucher: PHMX 416-422 depositados en CNIN y MZFC].
Figura 13. Pseudosermyle sp. 3 huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, opérculo.
Microestructura: (n = 1) (figs. 18, 19) cápsula de superficie granulosa (fig. 18A, B), con gránulos de diámetro entre 25.7-50.3 μm (fig. 18D), toda la superficie está cubierta por proyecciones digitiformes de altura entre 1.2-2.1 μm y una separación entre ellos de 0.7-3 μm (fig. 18E, F). Lámina micropilar de superficie como en la cápsula (fig. 19A), aunque los gránulos son de textura verrugosa (fig. 19E), el espacio entre los gránulos muestra la misma textura que aquellos de la cápsula (fig. 19E). Borde de una superficie con gránulos menos marcados y con un surco circumperiférico que lo separa del centro de la lámina micropilar (fig. 19A, D), con proyecciones digitiformes muy separadas, mide 76 μm de amplitud. Tubérculo micropilar como una giba más o menos rectangular de 97 μm de longitud y 69 μm de amplitud, su textura es verrugosa (figs. 18A, 19A). Copa micropilar separada del borde, con forma de ‘u’ abierta de 145 μm de amplitud y 22 μm de grosor (fig. 19C). Línea media como una carina de 31 μm de amplitud (figs. 18C, 19B). Las últimas 2 estructuras son de textura verrugosa. Opérculo de superficie granulosa (fig. 19F); la textura de los gránulos es similar a los que tiene la cápsula.
Figura 14. Pseudosermyle tridens cápsula del huevo en MEB. A, Dorsal; B, lateral; C, área polar; D, límite entre ambas texturas; E, textura dorsal; F, acercamiento a textura.
Material examinado: 45 huevos. México, Puebla, Cuetzalan del Progreso, camino a Tzinacapan, 20°0’59.6088” N, 97°32’23.388” O, 657 m snm [voucher: PHMX 259-303 depositados en CNIN y MZFC]
Microestructura: (n = 1) (figs. 20, 21) cápsula de superficie rugosa en general (fig. 20A, B), con arrugas que varían en tamaño entre 70-170 μm de longitud y 10-45 μm de amplitud (fig. 20D); estas estructuras están cubiertas por una retícula (fig. 20E) donde en cada intersección se produce una proyección cilíndrica o cónica de 1-3 μm de alto y una separación entre ellas de 1-3 μm (fig. 20F); con gránulos alrededor de la lámina micropilar cuyo diámetro mide entre 20-28 μm (figs. 20A, 21A), con textura similar a las arrugas. Lámina micropilar de textura rugosa (fig. 21E), con las arrugas hacia el centro y coliculada hacia el borde (fig. 21A). Borde a modo de carina periférica (fig. 21A); mide 13 μm de amplitud (fig. 21D).
Figura 15. Pseudosermyle tridens huevo en MEB. A-C, Lámina micropilar. A, Vista general; B, borde; C, protuberancia; D, opérculo.
Tubérculo micropilar en forma de triángulo invertido, mide 191 μm de longitud y 128 μm de amplitud (fig. 21A). Copa micropilar separada del borde, con forma de ‘u’ abierta; comienza donde termina el borde, mide 171 μm de amplitud y 24 μm de grosor (fig. 21C). Línea media es una carina con un surco en medio a lo largo, mide 56 μm de amplitud (figs. 20C, 21B). La textura de todas las estructuras anteriores es similar a la que exhiben las arrugas de la cápsula. Opérculo de composición similar a la cápsula, las arrugas suelen ser menos prominentes hacia el centro, el borde del opérculo es una carina periférica (fig. 21F). La textura es similar a la que poseen las arrugas de la cápsula.
Pseudosermyle sp. 5 (figs. 1, 6H, 7H, 22, 23)
Material examinado: 15 huevos. México, Puebla, Cuetzalan del Progreso, camino a Tzinacapan, 20°0’59.6088” N, 97°32’23.388” O, 657 m snm [voucher: PHMX 374-388 depositados en CNIN y MZFC].
Microestructura: (n = 1) (figs. 22, 23) cápsula de superficie rugosa en general (fig. 22A, B), con arrugas que varían en tamaño, mide entre 67-185 μm de longitud y 9-35 μm de amplitud (fig. 22D, E); estas estructuras están cubiertas por una textura rugulosa con orificios irregulares que varían en diámetro de 0.5-3.2 μm (fig. 22F). Lámina micropilar de superficie como en la cápsula (fig. 23A, E). Borde como un surco de 4.8 μm de amplitud (fig. 23D). Tubérculo micropilar como una giba más o menos rectangular, con 123 μm de longitud y 69 μm de amplitud (fig. 23A). Copa micropilar separada del borde, en forma de ‘u’ abierta, mide 187 μm de amplitud y 54 μm de grosor (fig. 23C). Línea media a modo de carina, mide 35 μm de amplitud (figs. 22C, 23B). La textura es similar a la que muestran las arrugas de la cápsula. Opérculo rugoso (fig. 23F), con arrugas similares a las que exhibe la cápsula y con la misma textura.
Figura 16. Pseudosermyle sp. 4 cápsula del huevo en MEB. A, Dorsal; B, lateral; C, área polar; D, límite entre ambas texturas; E, textura dorsal; F, acercamiento a textura.
Material examinado:12 huevos. México, Oaxaca, Totontepec Villa de Morelos, curva 1 Amatepec-Chinantequilla, 17°17’5.46” N, 95°59’18.1032” O, 1,562 m snm [voucher: PHMX 389-400 depositados en CNIN y MZFC].
Microestructura: (n = 1) (figs. 24, 25) cápsula de superficie espinosa (fig. 24A, B); espinas de diámetro entre 21-38 μm y altitud entre 81-211.5 μm (fig. 24A-D). Toda la superficie está cubierta por proyecciones digitiformes de altura entre 1.5-4.5 μm y una separación entre ellos de 1.7-4.4 μm (fig. 24D-F).
Figura 17. Pseudosermyle sp. 4 huevo en MEB. A-C, Lámina micropilar. A, Vista general; B, borde; C, protuberancia; D, opérculo.
Lámina micropilar cuya ornamentación y textura es similar a la cápsula (fig. 25A). Borde a modo de carina, con una sucesión de espinas unidas (similares a las de la cápsula) (fig. 25A, D). Tubérculo micropilar en forma de ‘y’, a manera de sucesión de espinas; mide 351 μm de longitud total, 199 μm de longitud hasta la bifurcación y 145 μm de longitud en cada brazo de la ‘y’, mide 27 μm de grosor (fig. 25A). Copa micropilar separada del borde, como una ‘u’ abierta, las espinas del borde acaban en los límites de la copa micropilar; mide 153 μm de amplitud y 24 μm de grosor (fig. 25C). Línea media a modo de sucesión de espinas solitarias (figs. 24C, 25B), de amplitud igual al diámetro de las mismas (21-38 μm). La textura en todas las estructuras anteriores es similar a la que posee la cápsula (fig. 25E). Opérculo espinoso con la misma composición, textura y medidas que las espinas de la cápsula; el borde carece de espinas y exhibe un surco que encierra a todas las espinas (fig. 25F).
Pseudosermyle grupo 4
Huevo: alveolar, de oviposición por caída (fig. 5A), doliforme.
Medidas en mm: (n = 105) L = 2.68 ± 0.09 (2.37-2.92), W = 1.69 ± 0.08 (1.38-1.89), H = 1.87 ± 0.08 (1.38-1.89), mpl = 0.74 ± 0.08 (0.58-0.94), mpw = 0.4 ± 0.08 (0.25-0.54), oph = 1.43 ± 0.11 (1.16-1.75) y opw = 1.16 ± 0.09 (0.94-1.42). Lámina micropilar en general oblonga y de longitud cercana a un tercio de la longitud de la cápsula, lámina micropilar interna abierta con línea media (fig. 6J-L).
Especies incluidas: Pseudosermyle phalangiphora (Rehn, 1907), P. procera Conle, Hennemann et Fontana, 2007 y Pseudosermyle sp. 6.
Material examinado: 45 huevos. México, Veracruz, San Andrés Tuxtla, camino viejo EBTLT, 18°35’9.89988” N, 95°4’31.8” O, 143 m snm [voucher: PHMX 169-213 depositados en CNIN y MZFC].
Figura 18. Pseudosermyle carinulata cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, textura de la cápsula; E, textura de gránulos; F, acercamiento a la textura de los gránulos.Figura 19. Pseudosermyle carinulata huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, Opérculo.
Microestructura: (n = 1) (figs. 26, 27) cápsula de superficie gibosa (fig. 26A, B), con gibas irregulares de diámetro entre 197.05-352.94 μm; suelen exhibir entre 1 y 4 poros de diámetro aproximado de 24 μm (fig. 26D), en el centro de este poro se produce una espina sombrilla (fig. 26D, E) de altura aproximada entre 44-48 μm, cuya amplitud es de 5 μm; la sombrilla de la espina formada por varios lóbulos es de diámetro aproximado de 18 μm (fig. 26E). La textura de la superficie dorsal es foveada (fig. 26D), con fóveas de 20 μm aproximadamente; éstas a su vez están punteadas con orificios de 0.5-1.8 μm de diámetro (fig. 26F); en el área polar en lugar de fóveas se forman gránulos (fig. 26C) de diámetro entre 12.5-17.18 μm.
Figura 20. Pseudosermyle striata cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, textura de la cápsula; E, textura de arrugas; F, acercamiento a la textura de las arrugas.
Lámina micropilar con superficie coliculada (fig. 27A) y textura rugulosa (fig. 27E). Borde como una carina de 207 μm de amplitud (fig. 27D); textura similar a la cápsula. Tubérculo micropilar similar a una media esfera de 114 μm de longitud y 153 μm de amplitud (fig. 27A); ornamentación similar a la lámina micropilar. Copa micropilar fusionada al borde con forma de ‘u’, mide 230 μm de amplitud y 129 μm de grosor (fig. 27C). Línea media a modo de carina de 179 μm de amplitud (figs. 26C, 27B). La textura de estas últimas 2 estructuras es similar a la cápsula. Opérculo muestra una giba al centro, mide 73-182 μm de longitud y entre 365-547 μm de amplitud, cuya altura es de 474-839 μm; está giba puede exhibir una muesca al centro, con esto adquiere una apariencia de pequeño cráter; en la cúspide presenta poros con espinas sombrilla del mismo tamaño y forma que los de la cápsula (fig. 27F). La textura es similar a la cápsula; en el borde se forma una carina circumperiférica (fig. 27F).
Material examinado: 45 huevos. México, Veracruz, San Andrés Tuxtla, camino viejo EBTLT, 18°35’9.89988” N, 95°4’31.8” O, 143 m snm [voucher: PHMX 214-258 depositados en CNIN y MZFC].
Microestructura: (n = 1) (figs. 28, 29) cápsula con superficie verrugosa (fig. 28A, B), cuyas verrugas miden de diámetro entre 40.8-289.32 μm, de textura rugulosa (fig. 28D, E), compuesta por protuberancias de diámetro de 2.42-8.53 μm (fig. 28E); entre las verrugas la superficie es foveada (fig. 28D), con fóveas de 17.68-28.06 μm de diámetro, e igual que las verrugas son de textura rugulosa (fig. 28F). Lámina micropilar de superficie escasamente coliculada (fig. 29A); textura rugulosa como la cápsula (fig. 29D). Borde a modo de carina (fig. 29A), mide 42 μm de amplitud, textura como en la cápsula (fig. 29C). Tubérculo micropilar como una media esfera de 61 μm de longitud y 80 μm de amplitud (fig. 29A); textura similar a la lámina micropilar. Copa micropilar fusionada al borde (fig. 29B), con forma de ‘u’, de 230 μm de amplitud y 159 μm de grosor, cuya textura es rugulosa. Sin línea media evidente. Opérculo con ornamentación similar a la cápsula, las verrugas forman un anillo periférico (fig. 29E); al centro la superficie es coliculada, pero mantiene la textura rugulosa (fig. 29F).
Pseudosermyle sp. 6 (figs. 1, 6L, 7L, 30, 31)
Material examinado: 15 huevos. México, Oaxaca, Santiago Choápam, curva Choápam-Latani, 17°22’39.9144” N, 95°54’58.5144” O, 701 m snm [voucher: PHMX 401-415 depositados en CNIN y MZFC].
Microestructura: (n = 1) (figs. 30, 31) cápsula coliculada (fig. 30A, B) con gránulos de 31.30-51.20 μm de diámetro, superficie rugulosa con verrugas de 1.76-4.12 de diámetro (fig. 30D, F). Las espinas sombrilla por lo general en grupos de al menos 3 (fig. 30E), aunque en ocasiones se encuentran solitarias; su altura es de 47-51 μm, su amplitud entre 5.9-10 μm; la sombrilla de la espina está formada por un círculo con diámetro entre 47.64-59.24 μm (fig. 30E). Lámina micropilar con ornamentación similar a la cápsula, pero carece de espinas sombrilla (fig. 31A). Borde a modo de carina de 49 μm de amplitud (fig. 31A, D), su textura rugulosa (fig. 31D). Tubérculo micropilar como un triángulo invertido de 252 μm de longitud y 155 μm de amplitud (fig. 31A); textura similar a la lámina micropilar (fig. 31E). Copa micropilar fusionada al borde, en forma de ‘u’, mide 139 μm de longitud y 48 μm de grosor (fig. 31C), de textura rugulosa como la cápsula. Línea media como una sucesión de gránulos (fig. 31B), puede exhibir espinas sombrilla; de 322 μm de longitud y 89 μm de amplitud, de textura rugulosa. Opérculo con espinas sombrilla que forman un anillo concéntrico rodeado de textura coliculada; al centro presenta la misma textura y algunas espinas sombrilla (fig. 31F).
Figura 21. Pseudosermyle striata huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, Opérculo.
Comportamiento de oviposición. Los grupos encontrados presentan 3 comportamientos distintos de oviposición: 1) por caída, para las especies: P. striata, P. carinulata, Pseudosermyle sp. 5 y Nooxapty isabelae del grupo 3 y P. phalangiphora, P. procera y Pseudosermyle sp. 6 del grupo 4 (fig. 5A); 2) pegados en sustrato, para las especies: Pseudosermyle sp. 1, Pseudosermyle sp. 2 y Pseudosermyle sp. 3 del grupo 1 (fig. 5B); y 3) agrupados en ooteca, para las especies: P. tridens y Pseudosermyle sp. 4 del grupo 2 (fig. 5C).
Variabilidad morfológica del huevo. Los caracteres constantes entre los grupos son: tipo de oviposición, tipo de huevo, forma de la cápsula, ángulo opercular, número de capas presentes en el opérculo, porcentaje de cobertura de la longitud de la lámina micropilar (mpl) con respecto a la longitud de la cápsula (L), forma de la lámina micropilar interna y posiciones del tubérculo y copa micropilar. Por lo tanto, suponemos que son caracteres relevantes para identificar los grupos obtenidos por PCA y nos permiten definirlos. En contraste, los caracteres como ornamentación y textura de la cápsula, opérculo, lámina micropilar y línea media son caracteres variables entre especies, pero constantes entre los huevos de distintas hembras de la misma especie. Podemos suponer que estos caracteres son específicos (tablas 10-12). A continuación, se discuten a nivel de grupo los caracteres mencionados como específicos y algunas de sus posibles tendencias.
Figura 22. Pseudosermyle sp. 4 cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, textura de la cápsula; E, arrugas; F, acercamiento a la textura de las arrugas.
Discusión
Las especies dePseudosermyle presentan distintos tipos de morfología del huevo, lo que nos permite reconocer 4 grupos, con Nooxapty isabelae próximo al grupo 3. Se reconocen 2 tipos de huevo, alveolar (grupos 3 y 4) y adhesivos (grupos 1 y 2); con 3 comportamientos de oviposición: por caída (grupos 3 y 4), adheridos al sustrato (grupo 1) e incluidos en 1 ooteca (grupo 2); se admiten 4 grupos acorde a las medidas obtenidas por PCA, que coinciden con aquellos reconocidos cualitativamente con otras estructuras (morfología del macho y la hembra). Esta diferenciación permite proponer el reconocimiento de 4 unidades ovocíticas distintas dentro de Pseudosermyle. Se observó que dentro de cada grupo existen ciertas características comunes en la morfología del huevo (tablas 10-12); pero a su vez existen diferencias suficientes entre las especies que los conforman (tablas 10-12). Por lo tanto, se puede reconocer y diferenciar de manera inequívoca a cada especie y el grupo al que pertenece. Las ilustraciones de P. truncata y Bacteria cacica Kaup, 1871 (sinónimo deP. tridens [Hebard, 1932]) corresponden conel patrón del huevo que encontramos en el grupo 1.
Figura 23. Pseudosermyle sp. 4 huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, Opérculo.
En cambio, la ilustración provista por Zompro (2001) de P. tridens sigue el patrón del grupo 2, lo que sugiere queB. cacica fue sinonimizada equivocadamente conP. tridens por Hebard (1932). Consideramos que es indispensable revisar más organismos de estas 2 especies con la finalidad de examinar esta sinonimia posiblemente errónea y reconocer a B. cacica como especie válida. Por último, P. phalangiphora corresponde con el grupo 4; no obstante, la morfología coriónica ilustrada por Conle et al. (2007) no corresponde con la encontrada en el presente trabajo. Conle et al.(2007) mencionan que esta especie es muy variable en sus genitales, por ende, una revisión detallada es necesaria para clarificar si se trata de variación intraespecífica o bien de 2 especies distintas con algunos caracteres homoplásticos.
Cabe destacar que en nuestro estudio se encontró un tipo de oviposición que previamente solo había sido registrado para la subfamilia Korinninae (hoy Necrosciinae) por Goldberg et al. (2015). Éste representa el segundo caso de formación de ooteca en Phasmatodea y a su vez una convergencia para ambos casos. Nuestros resultados representan el primer registro para América y el primero en la familia Diapheromeridae. Hay diferencia entre las ootecas de ambos taxones referente a la forma en la que los huevos están acomodados; mientras en Korinninae los huevos están acomodados de forma radial y se generan 4 cámaras, en nuestro caso, en el grupo 2de Pseudosermyle están acomodados en líneas sin formación de cámaras.
Figura 24. Nooxapty isabelae cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, superficie de la cápsula; E, textura de espinas; F, acercamiento a la textura de las espinas.
Debido a que las especies con ootecas, en este estudio, provienen de ambientes con temporada seca muy marcada como el matorral xerófilo y la selva baja subcaducifolia, deducimos que la formación de una ooteca con cápsula muy dura y ancha posiblemente evita que los huevos se desequen; tal como Salas-Araiza et al. (2013) reportan para algunos Acrididae hallados en matorral xerófilo, pues se trata de una adaptación a medios xéricos. Otro aspecto importante del fenómeno es que una condición previa a la formación de una ooteca es la presencia de huevos pegados a un sustrato, tal y como lo mencionan Robertson et al. (2018); en nuestro caso, también encontramos ese comportamiento de oviposición (grupo 1) en el mismo género donde se presentan ootecas. Para poner a prueba esta aseveración, es necesario realizar un análisis filogenético donde se incluyan más especies de Pseudosermyle y así examinar el proceso en términos de posibles tendencias evolutivas, de acuerdo con las secuencias reconocidas.
El huevo en Pseudosermyle es diferente en todas las especies respecto a su forma, texturas y la disposición de sus ornamentaciones, que son exclusivas de cada especie; además, estas características se agrupan en patrones más generales, que reconocemos como grupos. Asimismo, estos caracteres son constantes entre los huevos de distintas hembras de la misma especie. Dentro de Phasmatodea, las hembras de muchas especies son muy similares y el huevo ha sido un primer paso para reconocer si pertenecen a la misma o a distintas especies. A continuación, se listan los casos donde este carácter fue útil como primer paso en el reconocimiento de especies muy similares.
Figura 25. Nooxapty isabelae en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, Opérculo.
El primer caso corresponde con P. striata y Pseudosermyle sp. 5, donde son simpátridas e incluso los machos se parecen mucho. Al inicio, se pensó que pertenecían a la misma especie, hasta que se analizaron los huevos bajo el estereoscopio y el MEB. Se registró que los huevos eran diferentes en varios aspectos, como la ornamentación y la textura de la cápsula, la forma de la lámina micropilar y el opérculo. Entonces, para corroborar que se trataba de 2 especies distintas, se realizó la técnica descrita en materiales y método, con lo cual se confirmó nuestra hipótesis.
El segundo caso se dio entre P. phalangiphora y P. procera, que también son simpátridas. Se realizó el mismo procedimiento descrito en materiales y método. Las diferencias entre los huevos fueron la ornamentación de la cápsula, el opérculo y la presencia del relieve micropilar. En este caso, las diferencias entre machos eran conspicuas, así que la pregunta fue ¿cuál hembra y qué huevo correspondía a qué macho? Al igual que en el caso anterior, la respuesta se obtuvo al criar separadamente las ninfas eclosionadas de las 2 morfologías tal y como se describe en materiales y método. Después de obtener los adultos se pudo asignar a las hembras con sus respectivos machos y confirmar la continuidad morfológica con la reproducción.
Figura 26. Pseudosermyle phalangiphora cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, gibas; E, espina umbeliforme; F, textura de la cápsula.
El tercer caso es entre P. tridens y Pseudosermyle sp. 4, las diferencias entre machos suelen ser igual de sutiles que las mostradas entre hembras. En ese caso, las especies no son simpátridas y se pensaba que ambas eran una única especie, pues presentaban oviposición en ooteca y morfología genital similar. Por esta causa se recurrió a la disección de hembras para obtener los huevos y realizar la comparación; entonces se encontraron las primeras diferencias, pues la lámina micropilar de P. tridens es circular y está cerca del centro de la cápsula; la lámina de Pseudosermyle sp. 4 es un triángulo invertido desplazado al área polar. Para corroborarlo se obtuvo la primera generación de Pseudosermyle sp. 4, donde las hembras y los machos presentaban la misma morfología que los parentales; también se observó que los huevos tenían la misma morfología. El cultivo de P. tridens no se logró; cabe mencionar que fue hasta la observación de los huevos que se pudo determinar a P. tridens, pues el huevo es similar a la ilustración de Zompro (2001).
Con estos casos verificamos que la afirmación de Robertson et al.(2018) sobre la especificidad del huevo a nivel de especie es cierta para este clado de Phasmatodea. Al seguir esta aseveración pudimos notar que la imagen presentada porConle et al.(2007) del huevo de P. phalangiphora de un cultivo de Belice, puede ser una especie distinta a la que reconocemos aquí como P. phalangiphora.
Figura 27. Pseudosermyle phalangiphora huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, Opérculo.
Sin embargo, es complicado conocer la identidad con certeza, pues los autores anteriores mencionan que esta especie presenta gran variabilidad en los caracteres masculinos y femeninos; en consecuencia, aquí reconocimos a los ejemplares que entraban dentro del rango de variabilidad de P. phalangiphora como esta especie, aunque una revisión más exhaustiva de los caracteres de ésta y otras poblaciones es indispensable.
Figura 28. Pseudosermyle procera cápsula del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, superficie de la cápsula; E, textura de verrugas; F, Textura de la cápsula.
Los grupos 1 y 3 exhiben gránulos, pero la diferencia entre sus texturas es lo que los diferencia, pues el grupo 1 presenta gránulos como basamentos piramidales y el grupo 3 como semicírculos. También se observa una tendencia a aumentar o disminuir el número de niveles en el basamento piramidal y por lo tanto el tamaño del gránulo en el grupo 1; por el momento, hacen falta estudios comparativos que incluyan más especies, para después postular una posible tendencia (aumentar o disminuir el número de niveles en los basamentos piramidales) a este carácter, después de un ulterior análisis filogenético. El grupo 2 presenta especies que carecen de copa micropilar y línea media.
El grupo 3 se aproxima al género Nooxapty, por lo que se podría sugerir que al menos a nivel de medidas coriónicas son similares en forma. Este grupo exhibe gran variación en la microestructura coriónica de sus estructuras (tablas 10-12). La diferencia mayor entre Nooxapty y el grupo 3 es la presencia de espinas en el primero y de gránulos en el segundo. Se reconoce una tendencia de los gránulos a formar arrugas, pero se carece de hipótesis filogenética del género, que nos permita realizar una comparación precisa sobre tal tendencia. Además, el hecho que Nooxapty esté agrupado dentro de este grupo sugiere la necesidad de revisar estos caracteres bajo un enfoque filogenético, para aclarar si la similitud morfológica se debe a simplesiomorfías o a convergencia. Consideramos que un análisis filogenético nos permitiría contrastar y conocer los alcances de las medidas coriónicas, la ornamentación y la microestructura al momento de definir los géneros en Phasmatodea.
El grupo 4, al igual que el 3, exhibe gran variación en su microestructura coriónica (tablas 10-12). Bradler (2009) agrupa las 2 especies (P. phalangiphora y P. incongruens [Brunner von Wattenwyl, 1907]) dentro del género Pseudosermyle por presentar cercos asimétricos y con 2 ápices.Las demás, P. neptuna (Brunner von Wattenwyl, 1907), P. tridens (cercos con 3 ápices)y P. striata (con cercos simples), junto con este clado, forman un grupo monofilético a manera de peine, cuya sinapomorfía es la reducción del segundo esclerito del “poculum”. En particular, consideramos que tal variación en los cercos es muy grande y podría estar agrupando entidades distintas, por ejemplo, géneros distintos. En el presente trabajo mostramos evidencia que sustenta la formación de 4 grupos al usar la morfología coriónica y que cualitativamente observamos una correspondencia entre estos grupos y la morfología de los adultos. Sin embargo, se debe poner a prueba esta hipótesis bajo un enfoque filogenético que incluya más grupos de caracteres (v. gr. genitales internos) y especies.
Figura 29. Pseudosermyle procera huevo en MEB. A-D, Lámina micropilar. A, Vista general; B, copa micropilar; C, borde; D, textura, E-F, opérculo; E, vista general; F, textura.
A grandes rasgos, los caracteres como textura, forma de tubérculo micropilar, borde de la lámina micropilar, línea media externa y ornamentación (tablas 10-12) son de utilidad específica entre las especies. Para hacer más robusta esta hipótesis es necesario poner a prueba estos caracteres bajo un análisis filogenético, en el cual se deben considerar más especies de cada grupo. En los análisis previos, los grupos 1, 2 y 3 solo están representados por una especie y el grupo 4 por 2 especies (Bradler, 2009); mientras que el grupo 4 se ha incluido en análisis que consideran a todo Phasmatodea (Goldberg et al. 2015; Robertson et al. 2018) y Pseudosermyle no se incluyó en los análisis de Whiting et al. (2003) y Bradler et al. (2014), pese a ser el género con mayor diversidad en América del Norte (de Luna, 2022); esto evidencia la falta de información en un género tan ampliamente distribuido.
Figura 30. Pseudosermyle sp. 6 cápusla del huevo en MEB. A, Vista dorsal; B, vista lateral; C, área polar; D, superficie de la cápsula; E, espinas sombrilla; F, acercamiento a la textura.
Medidas coriónicas. Cuando aplicamos PCA a las medidas de huevo (L, W, H, mpl, mpw, oph y opw) obtuvimos 4 agrupaciones, que corresponden con los patrones generales del huevo explicados anteriormente. Al revisar la contribución de las variables a cada componente (fig. 4), nos percatamos de que la longitud de la cápsula (L), la altitud (oph) y amplitud (opw) del opérculo, así como la longitud micropilar (mpl) son las medidas coriónicas que mejor explican la variación de la forma del huevo. Podemos observar que L es una variable independiente de las restantes y la que más contribuye al componente 1 (fig. 4); mientras que oph y opw están más correlacionadas positivamente con la amplitud (W) y altitud (H) de la cápsula, las variables que más contribuyen al componente 2 (fig. 4). En cambio, mpl es la variable que más contribuye al componente 3 (fig. 4) y su correlación es inversamente proporcional a oph y opw; podemos destacar que cada estructura del huevo (cápsula, lámina micropilar y opérculo) está aportando una de las medidas que lo caracterizan. En consecuencia, recomendamos que al momento de realizar descripciones del huevo se incluyan todas las medidas mencionadas.
Las medidas del huevo nos permiten distinguir 4 grupos, que además son congruentes con otros caracteres. Esto podría indicar que el género Pseudosermyle pueda estar conformado por géneros distintos, como lo sugiere Conle et al. (2007), quienes llegan a esta conclusión debido a la variabilidad de formas en los genitales de los machos. Ellos admiten que el conocimiento del género es fragmentado, en consecuencia, no realizan una redescripción del mismo. Así también, ninguna de las especies que ellos describen sigue la descripción original de Pseudosermyle pues no cumplen con las características que menciona Caudell (1903); sin embargo, coinciden con la redescripción que realizó Zompro (2001). En contraste, resaltamos que las medidas por sí solas no son suficientes en la caracterización del huevo pues N. isabelae, por sus medidas, se aproxima al grupo 3, pero al revisar la microestructura notamos que la ornamentación es distinta entre esta especie y las restantes dentro del grupo, pues exhibe espinas, mientras que las demás presentan arrugas y gránulos. En conclusión, notamos que la microestructura, como sugieren Mazzini y Scali (1983), es bastante útil al reconocer especies cercanas dentro del mismo género.
Figura 31. Pseudosermyle sp. 6 huevo en MEB. A-E, Lámina micropilar. A, Vista general; B, línea media; C, copa micropilar; D, borde; E, textura; F, opérculo.
Por último, es importante destacar que Pseudosermyle exhibe gran variabilidad en los caracteres morfológicos del huevo, por lo cual consideramos que para aclarar si estos caracteres nos permiten inferir distintos géneros o representan variabilidad dentro del mismo, es indispensable revisar más caracteres, v. gr., genitales externos e internos para ambos sexos y observar una correspondencia entre los mismos. Así mismo, es necesario efectuar un análisis filogenético que nos permita postular hipótesis evolutivas sobre el género y sus caracteres o rasgos. Las especies restantes de Pseudosermyle no fueron incluidas en este trabajo debido a la carencia de tiempo y recursos para recolectar un área tan vasta como México, América Central y el sur de Estados Unidos. Por lo tanto, en el futuro será oportuno un trabajo más exhaustivo que incluya lo antes mencionado y amplíe el número de especies analizadas. Aún queda mucho trabajo por realizar con Pseudosermyle, pues al igual que muchos géneros de Phasmatodea americanos, se desconoce su bionomía.
Agradecimientos
Al posgrado en Ciencias Biológicas, UNAM, por el financiamiento a la beca de Secihti (cvu: 599426 y cvu: 1039296). A Susana Guzmán Gómez por la asistencia técnica en las micrografías en el Laboratorio de Microscopia y Fotografía de la Biodiversidad II, IB UNAM. A Arturo Arellano Covarrubias por la asesoría en la realización del mapa. A Isabel Vargas Fernández y María Berenit Mendoza Garfias por su asistencia técnica al montar las muestras y tomar las fotomicrografías con microscopio electrónico de barrido. Isabel Vargas ayudó críticamente en la elaboración de las figuras. JLB agradece al proyecto PAPIIT-UNAM IN 202415. ULM agradece a la Orthopterists Society por el Theodore J. Cohn Research Fund. Por último, agradecemos a 2 revisores anónimos en la revisión previa del presente manuscrito.
Referencias
Adler, D., Murdoch, D. Nenadic, O., Urbanek, S., Chen, M., Gebhardt, A. et al. (2018). rgl: 3D Visualization Using OpenGL. R package version 0.99.16. Recuperado el 10 junio 2019 de: https://cran.r-project.org/web/packages/shinyRGL/index.html
Allred, M. L., Stark, B. P. y Lentz; D, L. (1986). Egg capsule morphology of Anisomorpha buprestoides (Phasmatodea: Pseudophasmatidae). Entomological News, 97, 169–174.
Bradler, S. (2009). Die Phylogenie der Stab- und Gesepnstruchrecken (Insecta: Phasmatodea). Species, Phylogeny and Evolution, 2, 3–139. https://doi.org/10.17875/gup2009-710
Bradler, S. y Buckley, T. R. (2018). Biodiversity of Phasmatodea. En R. G. Foottit y P. H. Adler (Eds.), Insect biodiversity: science and society, Volumen II (pp. 281–313). Ottawa: Jonh Wiley & Sons Ltd. https://doi.org/10.1002/9781118945582.ch11
Bradler, S., Robertson, J. A. y Whiting, M. F. (2014). A molecular phylogeny of Phasmatodea with emphasis on Necrosciinae, the most species-rich subfamily of stick insects. Systematic Entomology, 39, 205–222. https://doi.org/10.1111/syen.12055
Brock, P. D., Büscher, T. y Baker; E, (2023). Phasmida Species File Online. Versión 5.0/5.0 Recuperado el 20 abril 2019 de: https://phasmida.speciesfile.org/
Camousseigth, A. y Bustamante, I. (1991). Descripción de los huevos de los fásmidos (Phasmatodea: Pseudophasma- tidae) de Chile. Revista Chilena de Entomología, 19, 39–43.
Caudell, A. N. (1903). The Phasmidae or walkingsticks, of the United States. Proceedings of the United States National Museum, 26, 863–885. https://doi.org/10.5479/si.00963801.26-1335.863
Caudell, A. N. (1914). The egg of Pseudosermyle truncata Caudell. Proceedings of the Entomological Society of Washington, 16, 96.
Clark, J. T. (1976a). The capitulum of phasmid eggs (Insecta: Phasmida). Zoological Journal of the Linnean Society, 59, 365–614. https://doi.org/10.1111/j.1096-3642.1976.tb01019.x
Clark, J. T. (1976b). The eggs of stick insects (Phasmida): a review with descriptions of the eggs of eleven species. Systematic Entomology, 1, 95–105. https://doi.org/10.1111/j.1365-3113.1976.tb00342.x
Clark, J. T. (1979). A key to the eggs of stick and leaf insects (Phasmida). Systematic Entomology, 4, 325–331. https://doi.org/10.1111/j.1365-3113.1979.tb00617.x
Conle, O. V., Hennemann, F, H. y Fontana, P. (2007). Studies on Neotropical Phasmatodea V: notes on certain species of Pseudosermyle Caudell, 1903, with the descriptions of three new species from Mexico (Phasmatodea: Diapheromeridae: Diapheromerinae: Diapheromerini). Zootaxa, 1496,31–51. https://doi.org/10.11646/zootaxa.1496.1.2
de Luna, M. (2022). Checklist of stick insects (Insecta: Phasmatodea) of North America, with three new records for Nuevo León, México. Phasmid Studies, 21, 90–105.
GNU Image Manipulation Program (GIMP 2.8.22). Recuperado el 10 junio 2019 de: https://www.gimp.org/
Goldberg, J., Bresseel, J., Constant, J., Kneubühler, B., Leubner, F., Michalik, P. et al. (2015) Extreme convergence in egg-laying strategy across insect orders. Scientific Reports, 5, 7825. https://doi.org/10.1038/srep07825
Harris, R. A. (1979). A glossary of surface sculpturing. Occasional Papers in Entomology, 28,1–31.
Hebard, M. (1932). New species and records of Mexican Orthoptera. Transactions of the American Entomological Society, 58,201–219.
Horikoshi, M. y Tang, Y. (2016). ggfortify: data visualization tools for statistical analysis results. Recuperado el 10 de junio de 2019 de: https://doi.org/10.32614/cran.package.ggfortify
Jintsu, Y., Uchifune, T. y Machida, R. (2010). Structural features of egg of the basal Phasmatodean Timema monikensis Vickery et Sandoval, 1988 (Insecta: Phasmatodea: Timematidae). Arthropod Systematics & Phylogeny, 68, 71–78. https://doi.org/10.3897/asp.68.e31716
Kaup, J. (1871). Ueber die Eier der Phasmiden. Berliner Entomologische Zeitschrift, 15, 17–24. https://doi.org/10. 1002/mmnd.18710150106
López-Mora, U. y Llorente-Bousquets, J. (2018). Lista de especies y clave ilustrada de los géneros de Phasmato- dea (Insecta) de México. Revista Mexicana de biodiversidad, 89, 46–78. https://doi.org/10.22201/ib.2007 8706e.2018.1.2175
López-Mora, U. y Llorente-Bousquets, J. (2023). Nooxapty gen. nov. con dos especies nuevas (Phasmatodea: Diapheromeridae) de Oaxaca, México. Dugesiana, 30, 11–33. https://doi.org/10.32870/dugesiana.v30i1.7272
Mazzinni, M., Carcupino, M. y Fausto, A. M. (1993). Egg chorion architecture in stick insects (Phasmatodea). International Journal of Insect Morphology and Embriology,22, 391–415. https://doi.org/10.1016/0020-7322(93)90021-r
Mazzini, M., Nascetti, G. y Bullini, L. (1982). The egg of the stick-insect Bacillus atticus caprai Nascetti et Bullini: a scanning electron microscope study (Cheleutoptera Bacillidae). Italian Journal of Zoology, 16, 93–103. https://doi.org/10.1080/01688170.1984.10510070
Mazzini, M., Nascetti, G. y Bullini, L. (1984). A scanning electron microscope study on the egg of a hybrid stick-insect (Cheleutoptera, Bacillidae). International Journal of Invertebrate Reproduction and Development, 7, 43–49. https://doi.org/10.1080/01688170.1984.10510070
Mazzini, M. y Scali, V. (1977). Fine structure of the insect micropyle. Scannin electron microscope investigations of the egg of the stick insect Bacillus rossius (Rossi) (Insecta Cheleutoptera). Monitore Zoologico Italiano-Italian Journal of Zoology, 11, 71–82. https://doi.org/10.1016/0020-7322(77)90020-4
Mazzini, M. y Scali, V. (1980). Eggs of the stick insects, Lonchodes pterodactylus Gray and Carausius morosus Br. (Phasmatodea: Heteronemiidae). International Journal of Insect Morphology & Embriology,9, 369–382. https://doi.org/10.1016/00207322(80)90010-0
Mazzini, M. y Scali, V. (1983). A scanning electron microscope study of the eggs of three stick insect species of Ramulus (Phasmatodea Heteronemiidae) from Somalia. Italian Journal of Zoology, 17, 89–103.
Nieves-Uribe. S., Llorente-Bousquets, J. y Flores-Gallardo, A. (2021). Toward standards in practices and techniques on ootaxonomy in the Pieridae (Lepidoptera: Papilionoidea). Zootaxa,4985, 301–344. https://doi.org/10.11646/zootaxa.4985.3.1
Ooms, J. (2018). magick: advanced graphics and image-processing in R. R package version 2.0. Recuperado el 10 junio 2019 de: https://CRAN.R-project.org/package=magick https://doi.org/10.32614/cran.package.magickgui
QGIS Dveelopment Team, (2019). QGIS Geographic Information System. Open Source Geospatial Foundation Project. Recuperado el 10 junio 2019 de: https://qgis.org.
R Core Team. (2013). R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna, Austria. Recuperado el 10 junio 2019 de: http://www.R- project.org/
Robertson, J. A., Bradler, S. y Whiting, M. F. (2018). Evolution of oviposition techniqus in stick and leaf insects (Phasmatodea). Frontiers in Ecology and Evolution, 6, 2016. https://doi.org/10.3389/fevo.2018.00216
RStudio Team (2015). RStudio: Integrated Development for R. RStudio, Inc., Boston, MA. Recuperado el 10 de junio de 2019 de: http://www.rstudio.com/. https://doi.org/10.4135/9781526472557
Salas-Araiza, M. D., Mackay, W. P., Valdez-Carrasco, J., Salazar-Solís; E, y Martínez-Jaime, O. A. (2013). Characterization and comparison of the eggs of seven species of Mexican grasshoppers. Southwestern Entomologist, 38,267–274. https://doi.org/10.3958/059.038.0210
Scali, V. y Mazzini, M. (1977). Fine morphology and amino acid analysis of the egg capsule of the stick insect, Clonopsis gallica (Charp.) (Cheleutoptera: Bacillinae). International Journal of Insect Morphology and Embryology, 6, 255–264. https://doi.org/10.1016/0020-7322(77)90020-4
Scali, V. y Mazzini, M. (1981). The eggs of stick insects, Sipyloidea sipylus (Westwood) and Orxines macklotti De Haan (Phasmatodea, Heteronemiidae). International Journal of Invertebrate Reproduction, 4, 25–38. https://doi.org/10.1080/01651269.1981.10553416
Scali, V. y Mazzini, M. (1982). Interpopulation differences in egg sculpturing of the stick insect, Clonopsis gallica (Charp.) (Phasmatodea: Bacillidae). International Journal of Insect Morphology and Embriology, 11, 189–195. https://doi.org/10.1016/s0020-7322(82)80004-4
Scali, V. y Mazzini, M. (1983). Egg capsule morphology of the stick insect Burria longixipha Brunner (Phasmatodea Heteronemiidae) from Somaliland. Italian Journal of Zoology, 17, 303–312.
Seiler, C., Bradler, S. y Koch, R. (2006). Fásmidos: insectos palo e insectos hoja. España: Hispano Europea.
Sellick, J. T. C. (1994). A redefinition of the orientation terminology of phasmid eggs. Phasmid Studies, 3, 1–2.
Sellick, J. T. C. (1997a). The range of egg capsule morphology within the Phasmatodea and its relevance to the taxonomy of the order. Italian Journal of Zoology,64, 97–104. https://doi.org/10.1080/11250009709356178
Sellick, J. T. C. (1997b). Descriptive terminology of the phasmid egg capsule, with an extended key to the phasmid genera based on egg structure. Systematic Entomology,22, 97–122. https://doi.org/10.1046/j.1365-3113.1997.d01-30.x
Sellick, J. T. C. (1997c). The “umbrella spines” and other surface projections of some phasmid eggs and some comments on phasmid taxonomy. Phasmid Studies, 6, 15–20. https://doi.org/10.1111/j.1096-3642.1988.tb01364.x
Sellick, J. T. C. (1998). The micropylar plate of the eggs of Phasmida, with a survey of the range of plate form within the order. Systematic Entomology, 23, 203–228. https://doi.org/10.1046/j.1365-3113.1998.00056.x
Severin, H. P. (1910). A study on the structure of the egg of the walking-stick Diapheromera femorata Say. Annals of The Entomological Society of America, 3, 83–92. https://doi.org/10.1093/aesa/3.2.83
Stark, B. P. y Lentz; D, L. (1986). Morphology of the egg capsule in Megaphasma dentricus (Phasmatodea: Heteronemiidae). Journal of the Kansas Entomological Society, 52, 398–401.
Whiting, M. F., Bradler, S. y Maxwell, T. (2003). Loss and recovery of wings in stick insects. Letter to Nature, 421, 264–267. https://doi.org/10.1038/nature01313
Wickham, H. (2016). ggplot2: elegant graphics for data analysis. Nueva York: Springer-Verlag.
Weiner, J. (2017). pca3d: Three dimensional PCA plots. R package version 0.10. Recuperado el 10 junio 2019 de: https://CRAN.R-project.org/package=pca3d
Zompro, O. (2001). A generic revision of the insect order Phasmatodea: the new world genera of the stick insect subfamily Diapheromeridae: Diapheromerinae = Heteronemiidae: Heteronemiinae sensu Gradley et Galil, 1977. Revue Suisse de Zoologie, 108, 189–255. https://doi.org/10.5962/bhl.part.79626
Four new records of the genus Septoglomus (Glomeromycota) in Mexico
Reyna P. Hipólito-Piedras a, Heriberto Méndez-Cortés b, *, Hugo M. Ramírez-Tobias b, Victor Olalde-Portugal c, Carlos Arturo Aguirre-Salado d, Ángel Natanael Rojas-Velázquez b y Aracely Mena-Echevarríae
a Universidad Autónoma de San Luis Potosí, Facultad de Agronomía y Veterinaria, Doctorado en Ciencias Agropecuarias, Carretera San Luis Potosí-Matehuala Km. 14.5, Ejido Palma de la Cruz, 78321 Soledad de Graciano Sánchez, San Luis Potosí, México
b Universidad Autónoma de San Luis Potosí, Facultad de Agronomía y Veterinaria, Carretera San Luis Potosí- Matehuala Km. 14.5, Ejido Palma de la Cruz, 78321 Soledad de Graciano Sánchez, San Luis Potosí, México
c Instituto Politécnico Nacional, Centro de Investigación y de Estudios Avanzados-Unidad Irapuato, Departamento de Biotecnología y Bioquímica, Libramiento Norte Carretera Irapuato-León Km 9.6, 36824 Irapuato, Guanajuato, México
d Universidad Autónoma de San Luis Potosí, Facultad de Ingeniería, Av. Dr. Manuel Nava 4, 78290 San Luis Potosí, San Luis Potosí, México
e Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Campo Experimental General Terán-INIFAP, Km 31 Carretera Montemorelos-China, 67400 General Terán, Nuevo León, México
*Autor para correspondencia: heriberto.mendez@uaslp.mx (H. Méndez-Cortés)
Recibido: 13 febrero 2024; aceptado: 23 julio 2025
Resumen
Los hongos micorrizógenos arbusculares (HMA) son organismos esenciales y de suma importancia para las plantas terrestres. Se seleccionaron distintas comunidades vegetales de México en los estados de Coahuila, Guanajuato, San Luis Potosí, Tamaulipas y Veracruz, en donde se recolectó suelo rizosférico con el objetivo de aislar esporas de HMA. La identificación taxonómica se llevó a cabo mediante el análisis de las características morfológicas de las esporas, comparándolas con los artículos originales de cada especie descrita. Se identificaron 4 especies de HMA pertenecientes al género Septoglomus (S. altomontanum, S. fuscum, S. furcatum y S. turnauae), que constituyen su primer registro en México. Con estas 4 especies, la riqueza total de HMA en México se eleva a 171, lo cual evidencia una diversidad significativa de HMA en las comunidades vegetales del país.
Arbuscular mycorrhizal fungi (AMF) are essential and extremely important organisms for terrestrial plants. Different plant communities of Mexico were selected in the states of Coahuila, Guanajuato, San Luis Potosí, Tamaulipas and Veracruz, where rhizospheric soil was collected with the objective of isolating AMF spores. Taxonomic identification was carried out by analyzing the morphological characteristics of the spores, comparing them with the original articles of each described species. Four species of AMF belonging to the genus Septoglomus (S. altomontanum, S. fuscum, S. furcatum, and S. turnauae) were identified, constituting their first record in Mexico. With the record of these 4 species, the total richness of AMF in Mexico rises to 171, evidencing a significant diversity of AMF in the plant communities of the country.
Los hongos micorrizógenos arbusculares (HMA) son organismos cruciales y de suma importancia en la ecología y fisiología de las plantas terrestres. Fueron considerados inicialmente en el filo Zygomycota; sin embargo, a través de sus características moleculares, morfológicas y ecológicas, fueron reorganizados en un nuevo filo monofilético denominado Glomeromycota (Schüßler et al., 2001). Actualmente, la clasificación taxonómica de los HMA incluye 3 clases, 4 órdenes y 43 géneros (Schüβler, 2025; Wijayawardene et al., 2020).
Uno de los géneros incluidos en Glomeromycota es Septoglomus. Originalmente, las especies que ahora se ubican en este género se agruparon dentro del género Glomus. Sin embargo, Schüßler y Walker (2010) las reubicaron en el género Funneliformis, basándose en criterios morfológicos. Posteriormente, Oehl et al. (2011) dividieron Glomus en 2 grupos taxonómicos, proponiendo los géneros Simiglomus y Septoglomus con base en análisis de secuencias ribosomales y en la morfología. Sin embargo, Redecker et al. (2013) validaron únicamente al género Septoglomus y recomendaron conservar su estatus en espera de mayor evidencia, particularmente a partir de estudios filogenéticos del DNAr.
La especie de referencia taxonómica para la clasificación del género Septoglomus fue Glomus constrictum (Trappe) Sieverd., GA Silva et Oehl (Glomeraceae) (actualmente, S. constrictum). A partir de ésta, se establecieron los criterios taxonómicos del género, que se caracterizan por la producción de esporas de manera individual o en pequeños grupos. Dichas esporas presentan una o múltiples capas y están conectadas a hifas que se extienden de forma continua hasta la pared de la espora, adoptan una forma cilíndrica o de embudo y presentan un poro cerrado por un septo ubicado en la base o debajo de la pared de la espora. Generalmente, estas esporas no reaccionan a la aplicación del reactivo de Melzer; sin embargo, sus estructuras adquieren una coloración azul oscuro al ser tratadas con azul de tripano (Oehl et al., 2011).
Actualmente, el género Septoglomus comprende 13 especies, lo que representa aproximadamente 4% de la diversidad total de HMA. Estas especies han sido documentadas en diversos estudios, como los de Błaszkowski et al. (2004, 2014, 2013, 2023), Goto et al. (2013), Symanczik et al. (2014), Chimal-Sánchez et al. (2020), Oehl et al. (2011, 2019), Palenzuela et al. (2013) y Guillén et al. (2020). Sin embargo, la clasificación de algunas de estas especies continúa en revisión. Por ejemplo, recientemente S. deserticola G.A. Silva, Oehl et Sieverd. y S. viscosum C. Walker, D. Redecker, D. Stille et A. Schüßler (Glomeraceae) fueron reclasificados en el nuevo género Blaszkowskia, con base en análisis filogenéticos y en su morfología distintiva (da Silva et al., 2023).
Las especies del género Septoglomus han sido registradas en diferentes continentes; en Europa, por Palenzuela et al. (2013), Blaszkowski et al. (2014), Oehl et al. (2019) y Guillén et al. (2020); en América, por Błaszkowski et al. (2013), Goto et al. (2013) y Chimal-Sánchez et al. (2020); en Asia, por Symanczik et al. (2014) y Oehl et al. (2019) y en África por Błaszkowski et al. (2010, 2013). El registro de estas especies fue en dunas marítimas, bosques lluviosos y tropicales, así como en matorrales xerófilos. No obstante, se ha observado que este género demuestra una destacada capacidad de adaptación y una dominancia notable en entornos áridos, en comparación con otros géneros de HMA. Además, muestra una eficiencia notable en la colonización de las raíces de cultivos agrícolas y forestales (Guardiola-Márquez et al., 2022).
En México se han identificado 4 especies del género Septoglomus. Sin embargo, es crucial llevar a cabo investigaciones a mayor escala, que abarquen tanto el ámbito ecológico como el morfológico para comprender mejor su distribución en el país. Asimismo, intensificar el muestreo para aumentar el conocimiento sobre la riqueza de especies de HMA en el territorio nacional (Polo-Marcial et al., 2021). El objetivo de esta investigación fue contribuir al conocimiento sobre la diversidad de Septoglomus mediante la documentación de 4 nuevos registros obtenidos en distintos tipos de vegetación de México.
Materiales y métodos
El muestreo del suelo rizosférico se realizó durante la época de lluvias (agosto, 2020-2021), en 6 sitios ubicados en 4 estados de la República Mexicana (fig. 1, tabla 1). En cada uno de los sitios se estableció un transecto de 1 km, donde cada 200 m se tomó una submuestra a una profundidad de 0 a 20 cm, la cual fue trasladada al laboratorio de Fitopatología, de la Facultad de Agronomía y Veterinaria, de la Universidad Autónoma de San Luis Potosí.
La extracción de esporas de HMA se realizó por el método de tamizado húmedo y decantación (Gerdemann y Nicolson, 1963), utilizando 500 g de suelo seco, seguido de una centrifugación en sacarosa al 70% a 2,000 rpm (Daniels y Skipper, 1982). Las esporas se montaron en preparaciones permanentes con alcohol polivinílico-lacto-glicerol (PVLG) y PVLG con reactivo de Melzer (INVAM, 2023).
La identificación se llevó a cabo de acuerdo con las características morfológicas de la espora, como lo sugieren da Silva et al. (2023). Así, se registraron el tamaño, las características de la pared de la espora y de la hifa de sostén, y la reacción al reactivo de Melzer. La observación de tales características fue en un microscopio óptico Zeiss Primo Star (Jena, Alemania) con iluminación fija Koehler.
Las mediciones de las características de las esporas de HMA se realizaron en fotografías tomadas con el software de microscopía ZEN blue (versión 3.4) (Jena, Alemania), con objetivos 40x y 100x. Tales características se contrastaron con lo descrito en las claves dicotómicas de Palenzuela et al. (2013) y Oehl et al. (2019); así como con los artículos de referencia (Blaszkowski et al., 2013, 2014; Palenzuela et al., 2013) y se elaboraron claves para la identificación de las especies. Los ejemplares de referencia fueron depositados en el herbario Isidro Palacios, perteneciente al Instituto de Investigación de Zonas Desérticas, de la Universidad Autónoma de San Luis Potosí (SLPM).
Descripciones
Se identificaron un total de 62 especies de HMA, en los 9 sitios de muestreo. De éstas, 21% pertenecen al género Acaulospora, 15% a Glomus, 11% a Septoglomus, 10% a Ambispora, 10% a Rhizophagus, 5% a Entrophospora, 5% a Funneliformis, 5% a Gigaspora, 5% a Scutellospora, 3% a Diversipora y 10% restante se distribuye en igual proporción entre los géneros Blazskowskia, Dominikia, Pacispora y Sieverdingia.
Figura 1. Distribución geográfica de sitios de muestreo y 4 especies del género Septoglomus en México.
Las especies registradas del género Septoglomus fueron: S. constrictum, S. mexicanum E. Chimal-Sánchez, C. Senés-Guerrero, NM Montaño, L. Varela, R. García-Sánchez, A. Pacheco, S. Montaño-Arias y SL Camargo-Ricalde, S. xanthium (Błaszk., Blanke, Renker y Buscot) GA Silva, Oehl y Sieverd., S. altomontanum Palenz., Oehl, Azcón-Aguilar y GA Silva, S. fuscum, S. furcatum Błaszk., Chwat, Kovács y Ryszka, y S. turnauae Błaszk., Chwat y Ryszka(Glomeraceae). Las últimas 4 son registro nacional, por lo cual se presenta la descripción morfológica y las ilustraciones correspondientes.
Tabla 1
Caracterización de los sitios de muestreo de Septoglomus en México.
Estado
Sitio
Coordenadas geográficas (Datum WGS84)
Vegetación
Suelo
Clima
San Luis Potosí
1. San José de Coronados
23°34’35.4” N 100°55’09.5” O
Bosque de pino
Xerosol
Árido templado
2. Xilitla
21°22’21.4” N 99°00’39.0” O
Bosque mesófilo de montaña
Litosol
Semicálido húmedo
3. Matehuala
23°21’51.1” N 100°35’01.7” O
Matorral xerófilo
Xerosol
Árido semicálido
4. Sierra de Álvarez
22°03’06.3” N 100°34’50.8” O
Bosque de encino
Litosol
Semicálido
5. Charcas
23°22’58.5” N 101°20’18.5” O
Pastizal
Xerosol
Semiárido
Guanajuato
6. San Felipe
21°41’56.4” N 101°12’57.6” O
Bosque de encino
Feozem
Semiárido templado
Veracruz
7. Plan de Higuera
19°26’17.0” N 96°33’35.6” O
Bosque tropical caducifolio
Vertisol
Cálido subhúmedo
Coahuila
8. Sierra Hermosa
25°14’42” N 100°51’18” O
Bosque de pino
Litosol
Semiárido templado
Tamaulipas
9. Jaumave
23°08’26.0” N 99°17’48.4” O
Bosque de encino
Litosol
Semiárido templado
Fuente: datos generados a partir de las capas disponibles en Geoportal y Enciclovida, México, Conabio (2018).
Septoglomus altomontanum Palenz., Oehl, Azcón-Aguilar y GA Silva, 2013.
Fig. 2a-d
Las esporas se forman individualmente en el suelo. Tienen forma ovalada y globosa, de 128-152 × 121-160 µm, de color marrón rojizo tornándose a negro rojizo. Las esporas están compuestas de 2 capas (Swl1-2). Swl1 es subhialina a amarillo oscuro, acompañada de partículas del suelo, de 2.1 a 3.2 µm de espesor, con la presión al oprimir el cubreobjetos esta capa se desprende. Swl2 es de marrón rojizo a negro rojizo, lisa, laminada, de 3.8 a 8.2 µm de espesor. Ninguna de las capas reacciona al reactivo de Melzer. La hifa de sostén es de forma curveada, continua a la pared de la espora, de un color marrón rojizo, es más ancha en la base de la espora y llega a medir 22 µm de largo. El poro de la espora está cerrado por un septo concoloro o una tonalidad más clara que la espora y surge de la capa Swl2.
Figura 2. Estructura subcelular de Septoglomus altomontanum. A) Espora con capa Swl1desprendida (40x); B) capas externas de la espora Swl1-2, en reactivo de Melzer (40x); C) hifa de sostén continua a la capa Swl2 con septo (100x); D) capas externas Swl1-2 en PVLG (100x).
Material examinado: México, Guanajuato, San Felipe (21°41’56.4” N, 101°12’57.6” O, 2,445 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-Piedras, 08-2021. México, San Luis Potosí, Catorce, San Antonio de Coronados (23°35’11.8” N, 100°54’01.2” O, 2,455 m snm) en un suelo rizosférico de un bosque de pino, R. P. Hipólito-Piedras, 08-2021. México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. (CH-SLPM-228).
Distribución y hábitat. Septoglomus altomontanum se registró por primera vez en el Parque Nacional Sierra Nevada en Andalucía, España, donde se encontró asociada a Pinguicola grandiflora Lam. y P. nevadensis H. Lindb(Lentibulariaceae), así como a otras especies vegetales presentes en el suelo circundante (Palenzuela et al., 2013). Posteriormente, se identificó en muestras de suelo rizosférico de eucaliptos en el noreste de Tailandia (Khaekhum et al., 2017), así como en un bosque enano de cerezo de montaña en el monte Fanjing, que forma parte de las montañas Wuling, ubicadas en la provincia de Guizhou, China (He et al., 2021). Adicionalmente, se ha documentado la presencia de S. altomontanum en un bosque de Cryptomeria japonica (Miq.) P.D.Sell, (Taxodiaceae) en estas mismas montañas (Ming et al., 2023). En México, se aisló de suelo rizosférico de un bosque de encino, en Guanajuato, bosque de pino en San Luis Potosí y bosque tropical caducifolio en Veracruz.
Septoglomus furcatum Błaszk., Chwat, Kovács y Ryszka, 2013.
Fig. 3a-d
Figura 3. Estructura subcelular de Septoglomus furcatum A) Grupo de esporas, bifurcación de hifa (fb), característica de la especie en PVLG (10x); B) capas externas Swl1-3 en reactivo de Melzer (100x); C) capas externas Swl1-3 de la pared de la espora, capas Shwl1-2, septo y bifurcación de la hifa de sostén en reactivo de Melzer (100x); D) espora e hifa de sostén en PVLG (40x).
Esporas de color marrón rojizo a marrón oscuro; globosas de (106-) 138 (-167) µm de diámetro. La pared de la espora consta de 3 capas (Swl1-3). Capa Swl1, formando la superficie de la espora, semipermanente, hialina con restos de suelo de 1.6 µm de espesor y se desprende con la presión al oprimir el cubreobjetos. Swl2 semipermanente, hialina de 1.3 a 3.1 µm de espesor. Swl3 es laminada, lisa, marrón rojizo de 5.3 a 10.9 µm de espesor. Ninguna de las capas de la pared de la espora se tiñe con el reactivo de Melzer. La hifa de sostén es de color marrón parduzco; de forma recta de 10.5 a 30.3 µm, frecuentemente presenta una bifurcación (fb) a distancia de la pared de la espora. La pared de la hifa está compuesta de 3 capas (shwl1-3), las cuales son continuas a las paredes de la espora (Swl1-3). El poro de la espora está cerrado. El septo se encuentra en la base de la espora y pueden encontrarse numerosos septos a lo largo de la hifa.
Material examinado: México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. México, San Luis Potosí, Matehuala (23°21’51.1” N, 100°35’01.7” O, 1,373 m snm) en un suelo rizosférico de matorral xerófilo, R. P. Hipólito-Piedras, 08-2021 (CH-SLPM-229).
Distribución y hábitat. Septoglomus furcatum, se registró por primera vez asociado a raíces de Cordia oncocalyx Allemann (Cordiaceae) en Brasil. Posteriormente se realizó un cultivo monoespecífico utilizando a Plantago lanceolata L. (Plantaginaceae) como planta huésped, en donde S. furcatum formó micorrizas con numerosos arbúsculos, vesículas e hifas poco frecuentes (Blaszkowski et al., 2013). El segundo reporte en Brasil, se evidenció a través de una investigación en torno a la retroalimentación de suelo-planta, donde la mencionan como una especie clave, propagada en Senna uniflora (Mill.) HSIrwin y Barneby (Caesalpiniaceae), con suelo de áreas degradadas que han sido restauradas en ese país (Medeiros et al., 2022). La presencia de S. furcatum en el continente asiático seregistró asociado a las raíces de Rosa acicularis Lindl. (Rosaceae) junto a una baja diversidad de 3 especies de HMA (Zu et al., 2019), mientras que Ming et al. (2023), la registraron en un bosque de Pinus damingshanensis W.C.Cheng y L.K.Fu (Pinaceae) , Cryptomeria japonica y en un bosque lanceolado de Cunninghamia R.Br. (Cupressaceae), junto con 27 especies más. En México, se aisló de suelo rizosférico de un bosque tropical caducifolio en Veracruz y matorral xerófilo en San Luis Potosí.
Septoglomus fuscum Błaszk., Chwat, Kovács y Ryszka, 2013.
Fig. 4a-d
Figura 4. Estructura subcelular de Septoglomus fuscum. A) Esporas solitarias en PVLG (40x); B) capas externas Swl1-2 de la pared de la espora e hifa de sostén en reactivo de Melzer (40x); C) capas externas Swl1-2 de la pared de la espora en reactivo de Melzer (100x); D) capas shwl1-2 de la pared de la hifa de sostén en PVLG (100x).
Las esporas se forman en el suelo en racimos sueltos. Grupos de 89-194 × 141-248 µm con 2 a 7 esporas. Esporas de color blanco amarillento en la juventud, anaranjado parduzco a marrón oscuro en la madurez; globosas a subglobosas; (32-) 51 (-102) µm de diámetro. La pared de la espora consta de 2 capas (Swl1-2). Swl1, es semipersistente, semiflexible, de color amarillo dorado, de 1.8 a 2.4 µm de grosor, al oprimir el cubreobjetos se separa de la capa Swl2. Swl2 es laminada, lisa, de amarillo a anaranjado parduzco de (1.8-) 3.5 (-6.7) µm de grosor. Ninguna de las capas de la pared de la espora se tiñe con el reactivo de Melzer. La hifa de sostén es de color anaranjado parduzco a marrón oscuro en esporas maduras; recta o recurvada, cilíndrica en forma de embudo, a veces ligeramente constreñida en la base de la espora; (5.8-) 7.4 (-12.3) µm de anchura en la base de la espora. Pared de la hifa de color amarillo a anaranjado parduzco de (0.8-) 2.7 (-5.8) µm de grosor. El poro es abierto de 1.5 a 2.7 µm de diámetro.
Material examinado: México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. México, San Luis Potosí, Matehuala (23°21’51.1” N, 100°35’01.7” O, 1,373 m snm) en un suelo rizosférico de matorral xerófilo, R.P. Hipólito-Piedras, 08-2021 (CH-SLP-230).
Distribución y hábitat.Septoglomus fuscum se registró por primera vez asociado con raíces de Arctotheca populifolia (P.J.Bergius) Norl. (Asteraceae) en Sudáfrica. Posteriormente, se realizaron cultivos monoespecíficos con Plantago lanceolata como cultivo huésped, en donde se observaron estructuras como arbúsculos, hifas intra y extrarradicales (Blaszkowski et al., 2013). En México, se aisló de suelo rizosférico de bosque tropical caducifolio en Veracruz y un matorral xerófilo en San Luis Potosí.
Septoglomus turnauae Błaszk., Chwat, Ryszka y Orfanoudakis, 2014.
Fig. 5a-d
Figura 5. Estructura subcelular de Septoglomus turnauae. A) Espora solitaria en reactivo de Melzer (40x); B) paredes externas de la pared de la espora Swl1-4 e hifa de sostén en PVLG (40x); C) capas externas de la pared de la espora en PVLG (100x); D) septo y paredes shwl1-4 de la hifa de sostén en reactivo de Melzer (100x).
Las esporas se forman de manera solitaria en el suelo. Esporas de color anaranjado amarronado a marrón oscuro, globosas de (45) 64 (-102) µm de diámetro. La pared de la espora está compuesta de 4 capas (Swl1-4). Swl1, es evanescente, hialina, de 3.1 µm de diámetro. Swl2 es permanente, laminada, lisa, de anaranjado claro de 1.8 a 9.2 µm de espesor. Swl3 es laminada, lisa, de color anaranjado claro a anaranjado parduzco, de 3.5 a 9.3 µm de espesor. Swl4 semiflexible, lisa, de color anaranjado parduzco de 1.0 a 1.7 µm de espesor. Ninguna de las capas se tiñe con el reactivo de Melzer. La hifa es de color anaranjado parduzco de (4.5-) 8.2 (-10.7) µm de espesor en la base de la espora, la hifa es continua con las capas Swl1-3. El poro es de (1.3-) 2.7 (-3.2) µm de diámetro, estrechándose con la edad.
Material examinado: Coahuila, Sierra Hermosa (25°14’42” N, 100°51’18” O, 2,255 m snm) en suelo rizosférico de un bosque de pino, H. Méndez-Cortés, 08-2020. Guanajuato, San Felipe (21°41’56.4” N, 101°12’57.6” O, 2,445 m snm), en un suelo rizosférico de un bosque de encino, R.P. Hipólito-Piedras. San Luis Potosí, Xilitla (21°22’21.4” N, 99°00’39.0” O, 857 m snm) en un suelo rizosférico de un bosque mesófilo de montaña, R. P. Hipólito-Piedras, 08-2021. México, San Luis Potosí, Charcas (23°22’58.5” N, 101°20’18.5” O, 2,471 m snm) en un suelo rizosférico de un pastizal, R. P. Hipólito-Piedras, 08-2021. San Luis Potosí, Sierra de Álvarez (22°03’06.3” N, 100°34’50.8” O, 2,700 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-Piedras, 08-2021. Tamaulipas, Jaumave (23°08’26.0” N, 99°17’48.4” O, 742 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-piedras, 08-2021. (CH-SLPM-230).
Distribución y hábitat. S. turnuauae se registró en cultivos trampa inoculados con el suelo de la rizósfera y raíces de Cistus sp. L. (Cistaceae), en Italia. Posteriormente, se realizaron cultivos monoespecíficos con Plantago lanceolata, donde se observaron arbúsculos e hifas (Blaszkowski et al., 2014). En México, se aisló de suelo rizosférico de bosques de encino en Guanajuato y Tamaulipas, bosques de pino en Coahuila, bosque mesófilo de montaña y pastizal en San Luis Potosí.
Clave de las especies del género Septoglomus presentes en México
1. Esporas mayores a 200 micras 4
1. Esporas menores a 200 micras 2
2. Esporas con 2 capas en la pared de la espora 3
2. Esporas con 3 capas en la pared de la espora 5
2. Esporas con 4 capas en la pared de la espora 6
3 Esporas amarillas a marrones 7
3. Esporas rojas a negras 8
4. Esporas solitarias; de color anaranjado rojizo a marrón rojizo; globosa; (154.5-)202.8 (-228.9) µm de diámetro. La pared de la espora costa de 4 capas (Swl1-4). Capa Swl1 es evanescente hialina, lisa, con la edad se torna rugosa o se desprende. Swl2 permanente, laminada de color anaranjado, Swl3 es permanente, lisa y laminada de color anaranjado a marrón. Swl4 semiflexible, hialina, lisa, raramente se separa de swl3. Hifa de sostén amarilla a marrón, recurvada de 4 capas (shwl1-4), con poro cerrado por un septo situado en la base de la espora Septoglomus mexicanum
5. Esporas solitarias; de color marrón rojizo a marrón oscuro; globosas a subglobosas; (106-) 138 (-167) µm de diámetro. La pared de la espora consta de 3 capas (Swl1-3). Capa Swl1, semipermanente, hialina a anaranjado claro, lisa con gránulos de suelo. Swl2 semipermanente, hialina a amarillo dorado. Swl3 es laminada, lisa, marrón rojizo a marrón oscuro. La hifa de sostén es de color marrón parduzco a marrón oscuro; rectas o curvadas, cilíndricas, bifurcada, compuestas por 3 capas (shwl1-3), poro cerrado por un septo en la base de la espora, además, a lo largo de la hifa se encontrarán septos transversales Septoglomus furcatum
6. Esporas solitarias; de color anaranjado parduzco a marrón oscuro; globosas a subglobosas; (110-) 133(-165) μm de diámetro. La pared de la espora consta de 4 capas (Swl1-4). Swl1, es evanescente, hialina y lisa. Swl2 permanente, laminada, lisa, de color anaranjado claro a anaranjado parduzco. Swl3 es laminada, lisa, de color anaranjado claro a anaranjado parduzco. Swl4 semiflexible, lisa, de color anaranjado claro a anaranjado parduzco. La hifa de sostén es de color anaranjado parduzco a marrón oscuro, constituida de 3 capas (shwl1-3) con un septo en la base de la espora que no invade el lumen de la hifa de sostén Septoglomus turnauae
7. Esporas en racimos o solitarias, de color amarillento en la juventud, anaranjado parduzco a marrón oscuro en la madurez; globosas a subglobosas; (20- ) 47 (-90) µm de diámetro. La pared de la espora consta de 2 capas (Swl1-2). Swl1, semi persistente, semiflexible, de color blanco anaranjado a amarillo dorado. Swl2 laminada, lisa, blanco amarillento en esporas juveniles, anaranjado parduzco a marrón oscuro en esporas maduras. La hifa de sostén de color anaranjado parduzco a marrón oscuro; recta o recurvada, cilíndrica a forma de embudo, con poro abierto, sin septo, estrechamiento gradual hasta llegar al centro del lumen de la hifa Septoglomus fuscum
8. Esporas solitarias; de color marrón rojizo oscuro a negro rojizo; ovoides, elípticas y globosas, de 137-175 (-208) × 125-170 (-204) µm de diámetro. La pared de la espora está compuesta por 2 capas (Swl1-2). Swl1 es subhialina, lisa, de color amarillo oscuro. Swl2 es de marrón rojizo oscuro a negro rojizo, lisa, laminada. La hifa de sostén de color amarillo oscuro a un marrón rojizo, cilíndrica, curveada, ancha en la base de la espora estrechándose debajo de esta, poro cerrado, septo concoloro a la base de la espora y tonalidad clara cuando se encuentra debajo de la pared de la espora Septoglomus altomontanum
Discusión
En México, el género Septoglomus está representado por 33% de las especies descritas en el mundo, lo que equivale a 5 de las 13 especies conocidas. Entre éstas, destaca S. constrictum debido a su amplia distribución y frecuentes registros en diversas comunidades vegetales. Por otro lado, S. xanthium ha sido principalmente registrada en agroecosistemas (Polo-Marcial et al., 2021). Septoglomus mexicanum se ha registrado en asociación con Prosopis laevigata (Humb. y Bonpl. ex Willd.) M.C.Johnst. (Mimosaceae) (Chimal-Sánchez et al., 2020), similar a S. titan BT Goto y GA Silva (Glomeraceae) que se ha registrado en zonas áridas del país (Guardiola-Márquez et al., 2022).
De acuerdo con los registros de las especies de HMA descritas en esta investigación, Septoglomus altomontanum fue la única especie cuya distribución coincidió con la descripción original al ser aislada en sitios con vegetación de coníferas y encinos, a altitudes superiores a 2,000 m, lo cual es similar a lo reportado por Palenzuela et al. (2013), quienes documentaron su presencia en zonas con vegetación similar, localizadas entre 1,800 y 3,100 m de altitud.
Septoglomus altomontanum se diferencia de las especies de HMA descritas hasta la fecha, incluso de las del mismo género, en términos de tamaño, color, estructura, forma e hifas de las esporas. La especie que podría generar algún debate conforme a su morfología es S. furcatum; no obstante, ésta presenta la particularidad de tener una hifa que se adelgaza a medida que se aleja de la base de la espora (Blaszkowski et al., 2013), mientras que S. altomontanum mantiene un grosor uniforme a lo largo de la pared de la espora y la hifa (Palenzuela et al., 2013).
Septoglomus furcatum se destaca por sus esporas grandes de tonalidad oscura que presentan una pared de 3 capas y, con frecuencia, hifas ramificadas, incluso llegando a tener 2 hifas cercanas entre sí, característica que no se encuentra en otras especies del género Septoglomus (Blaszkowski et al., 2013).A primera vista, las esporas de S. furcatum pueden parecer similares a las de S. constrictum (Trappe) Sieverd., GA Silva et Oehl; sin embargo, se diferencian por la estructura de la pared de la espora, ya que S. furcatum tiene 3 capas, mientras que S. constrictum tiene solo 2. Además, la espora de S. constrictum tiende a ser de un tono anaranjado parduzco, mientras que la de S. furcatum es de color marrón oscuro. La característica de S. furcatum de poseer una hifa ramificada, es similar a la de las esporas de G. brohultii Sieverd. y R.A. Herrera (Glomeraceae). Sin embargo, G. brohultii tiene una tonalidad más clara tanto en la pared de la espora como en la hifa, a diferencia de S. furcatum, que presenta una tonalidad oscura uniforme tanto en la pared como en la hifa (Blaszkowski et al., 2013; Oehl et al., 2011).
Septoglomus fuscum es similar a S. xanthium, tanto en características moleculares como en características morfológicas. No obstante, se ha observado que S. fuscum forma racimos sueltos de esporas, mientras que S. xanthium se compone de esporas solitarias que se adhieren y se desarrollan dentro de las raíces (Blaszkowski et al., 2004). Al analizar las capas, se nota que S. xanthium posee una capa adicional permanente que S. fuscum no presenta; sin embargo, S. fuscum tiene un grosor de capa laminada de 2 a 3 veces mayor que el de S. xanthium (Blaszkowski et al., 2013).
Desde una perspectiva filogenética, S. turnauae presenta una estrecha relación con S. fuscum, lo que sugiere un origen común entre ambas especies, a través de análisis ribosomales que las agrupan en un mismo clado. Sin embargo, se destaca que la mayoría de las esporas de S. fuscum se agrupan en racimos sueltos (Blaszkowski et al., 2013), en contraste con las esporas individuales de S. turnauae. Al comparar el número de capas, S. turnauae muestra similitudes con S. mexicanum, ambas compuestas por 4 capas en su estructura, aunque, difieren en el color de las esporas. Mientras que S. mexicanum presenta tonalidades que van desde el rojizo hasta el marrón (Chimal-Sánchez et al., 2020), las esporas de S. turnauae tienen un tono anaranjado parduzco y tonalidades diferentes, conforme empieza otra capa (Blaszkowski et al., 2014).
Previo a esta investigación, se consideraba que la distribución del género Septoglomus era limitada, sin embargo, donde se localiza expresa un comportamiento dominante respecto a los demás géneros de HMA (Guardiola-Márquez et al., 2022). No obstante, la presente investigación, que ha registrado 4 especies pertenecientes a este género, revela una riqueza de HMA previamente desconocida en las comunidades vegetales de México. Además, sugiere que este género no se encuentra restringido a una distribución específica, ya que se ha documentado en 5 comunidades vegetales distintas en el país. El reciente hallazgo de S. altomontanum, S. fuscum, S. furcatum y S. turnauae eleva el número de especies registradas en México a 171, ésto constituye aproximadamente 48% de la riqueza global de HMA y 62% de las especies se han reportado del género Septoglomus en el país.
Agradecimientos
A la Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti) por la beca otorgada (775694) a la primera autora para sus estudios de posgrado.
Referencias
Błaszkowski, J., Blanke, V., Renker, C., y Buscot, F. (2004). Glomus aurantium and G. anthium, new species in Glomeromycota. Mycotaxon, 90, 447–467. https://doi.org/10.5962/p.431247
Błaszkowski, J., Chwat, G., Góralska, A., Ryszka, P. y Orfanoudakis, M. (2014). Septoglomus jasnowskae and Septoglomus turnauae, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycology Progress, 13, 985. https://doi.org/10.1007/s11557-014-0985-z
Blaszkowski, J., Chwat, G., Kovacs, G. M., Gaspar, B. K., Ryszka, P., Orowska, E. et al. (2013). Septoglomus fuscum and S. furcatum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia, 105, 670–680. https://doi.org/10.3852/12-127
Błaszkowski, J., Kovács, G. M., Balazs, T. K., Orlowska, E., Sadravi, M., Wubet, T. et al. (2010). Glomus africanum and G. iranicum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia, 102, 1450–1462. https://doi.org/10.3852/09-302
Błaszkowski, J., Yamato, M., Niezgoda, P., Zubek, S., Milczarski, P., Malinowski, R. et al. (2023). A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycological Progress, 22, 1–15. https://doi.org/10.1007/s11557-023-01882-9
Chimal-Sánchez, E., Senés-Guerrero, C., Varela, L., Montaño, N. M., García-Sánchez, R., Pacheco, A. et al. L. (2020). Septoglomus mexicanum, a new species of arbuscular mycorrhizal fungi from semiarid regions in Mexico. Mycologia, 112, 121–132. https://doi.org/10.1080/00275514.2019.1671147
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2018). Sistema Nacional de Información sobre Biodiversidad (SNIB). Registros de ejemplares, versión 2017-12. Geoportal y Enciclovida, México. Recuperado el 28 julio, 2023 de: http://www.conabio.gob.mx/informacion/gis/
da Silva, G. A., Corazon-Guivin, M. A., de Assis, D. M. A. y Oehl, F. (2023). Blaszkowskia, a new genus in Glomeraceae. Mycological Progress, 22, 1–14. http://dx.doi.org/10.1007/s11557-023-01919-z
Daniels, H. B. A. y Skipper, H. D. (1982). Methods for the recovery and quantitative estimation of propagules from soil. En N. C. Schenck (Ed.), Methods and principles of mycorrhizal research (pp. 29–37). St. Paul, Minn.: American Society for Phytopathology,
Gerdemann, J. W. y Nicolson, T. J. (1963). Spores of mycorrhizal Endogonespecies extracted from soil by wet sieving and decanting. Transactions of the BritishMycological Society, 46, 235–244. http://dx.doi.org/10.1016/S0007-1536(63)80079-0
Goto, B. T., Araújo, A. F., Soares, A. C. F., de Almeida-Ferreira, A. C., Maia, L. C., da Silva-Sousa, C. et al. (2013). Septoglomus titan, a new fungus in the Glomeraceae (Glomeromycetes) from Bahia, Brazil. Mycotaxon, 124, 101–109. http://dx.doi.org/10.5248/124.101
Guardiola-Márquez, C. E., Pacheco, A., Mora-Godinez, S., Schüßler, A., Gradilla-Hernández, M. S. y Senés-Guerrero, C. (2022). Septoglomus species dominate the arbuscular mycorrhiza of five crop plants in an arid region of northern Mexico. Symbiosis, 87, 93–106. http://dx.doi.org/10.1007/s13199-022-00851-2
Guillén, A., Serrano-Tamay, F. J., Peris, J. B. y Arrillaga, I. (2020). Glomus ibericum, Septoglomus mediterraneum, and Funneliformis pilosus, three new species of arbuscular mycorrhizal fungi. Mycologia, 112, 819–828. https://doi.org/10.1080/00275514.2020.1771992
He, R. J., Ming, Y., Yao, L. M., Zhu, Q. Q. y Jiang, L. (2021). Diversity of arbuscular mycorrhizal fungi in mossy dwarf forest of Fanjing Mountain, Guizhou Province, Southwest China. Mycosystema, 40, 2843–2853. https://doi.org/10.13346/j.mycosystema.210188
INVAM (International Culture Collection of Vesicular Arbuscular Mycorrhizal Fungi). (2023). Recuperado el 28 agosto, 2023 de: http://invam.caf.wvu.edu/
Khaekhum, S., Lumyong, S., Kuyper, T. W. y Boonlue, S. (2017). Species richness and composition of arbuscular mycorrhizal fungi occurring on eucalypt trees (Eucalyptus camaldulensis Dehnh.) in rainy and dry season. Current Research in Environmental and Applied Mycology, 7, 282–292. http://dx.doi.org/10.5943/cream/7/4/5
Medeiros, A. S., Scaloppi, J. C., Damasceno, E. S., Goto, B. T., Vieira, D. C. M., Socolowski, F. et al. (2022). Arbuscular mycorrhizal fungi communities shaped by host-plant affect the outcome of plant-soil feedback in dryland restoration. Journal of Applied Ecology, 60, 507–518. https://doi.org/10.1111/1365-2664.14330
Ming, Y., He, R., Yao, L., Zhu, Q., Long, C. y Jiang, L. (2023). Diversity of soil arbuscular mycorrhizal fungi in different coniferous forests in Fanjing Mountain. Journal of Yunnan Agricultural University (Natural Science), 38, 149–157. https://doi.org.10.12101/j.issn.1004-390X(n).202108033
Niezgoda, P., Błaszkowski, J., Błaszkowski, T., Stanisławczyk, A., Zubek, S., Milczarski, P. et al. (2024). Three new species of arbuscular mycorrhizal fungi (Glomeromycota) and Acaulospora gedanensis revised. Frontiers in Microbiology, 15, 1320014. https://doi.org/10.3389/fmicb.2024.1320014
Oehl, F., Sánchez-Castro, I., da Silva, D. K. A., Santos, V. M., Palenzuela Jiménez, E. J. y da Silva, G. A. (2019). Septoglomus nigrum, a new arbuscular mycorrhizal fungus from France, Germany and Switzerland. Nova Hedwigia, 109, 121–134. http://dx.doi.org/10.1127/nova_hedwigia/2019/0523
Oehl, F., Silva, G. A. D., Goto, B. T. y Sieverding, E. (2011). Glomeromycota: three new genera and glomoid species reorganized. Mycotaxon, 116, 75–120. http://dx.doi.org/10.5248/116.75
Palenzuela, J., Azcón-Aguilar, C., Barea, J. M., da Silva, G. A. y Oehl, F. (2013). Septoglomus altomontanum, a new arbuscular mycorrhizal fungus from mountainous and alpine areas in Andalucía (southern Spain). IMA Fungus, 4, 243–249. https://doi.org/10.5598/imafungus.2013.04.02.09
Polo-Marcial, M. H., Lara-Pérez, L. A., Goto, B. T., Margarito-Vista, X. y Andrade-Torres A. (2021). Glomeromycota in Mexico, a country with very high richness. Sydowia, 74, 33–63. http://dx.doi.org/10.12905/0380.sydowia74-2021-0033
Redecker, D., Schüßler, A., Stockinger, H., Stürmer, S. L., Morton, J. B. y Walker, C. (2013). An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza, 23, 515–531. https://doi.org/10.1007/s00572-013-0486-y
Schüßler, A., Schwarzott, D. y Walker, C. (2001). A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycological Research, 105, 1413–1421. https://doi.org/10.1017/S0953756201005196
Schüßler, A. y Walker, C. (2010). The Glomeromycota. A species list with new families and new genera. Recuperado el 28 agosto, 2023 de: http://schuessler.userweb.mwn.de/amphylo/
Symanczik, S., Błaszkowski, J., Chwat, G., Boller, T., Wiemken, A. y Al-Yahya’ei, M. N. (2014). Three new species of arbuscular mycorrhizal fungi discovered at one location in a desert of Oman: Diversispora omaniana, Septoglomus nakheelum and Rhizophagus arabicus. Mycologia, 106, 243–259. https://doi.org/10.3852/106.2.243
Zu, Y., Ping, Y., Mu, L. y Yang, T. (2019). The diversity of arbuscular mycorrhizal fungi of Rosa acicularis ‘Luhe’in saline areas. Journal of Forestry Research, 30, 1507–1512. https://doi.org/10.1007/s11676-018-0748-9
Wijayawardene, N. N., Hyde, K. D., Al-Ani, L. K. T., Tedersoo, L., Haelewaters, D., Rajeshkumar, K. C. et al. (2020). Outline of fungi and fungus-like taxa. Mycosphere Online. Journal of Fungal Biology, 11, 1060–1456. https://doi.org/10.5943/mycosphere/13/1/2
Schüβler, A. 2025. Glomeromycota: species list. Recuperado en marzo, 2025 de: http://www.amf-phylogeny.com/
Descripción morfológica de la primera zoea del camarón de agua dulce Macrobrachium crenulatum (Decapoda: Palaemonidae) y una comparación con sus congéneres del Neotrópico
Lucas Oliveira-Rogeri a, *, Rodrigo Pantoni a, Ingo S. Wehrtmann b, c, d, Fernando L. Mantelatto a, João Alberto Farinelli Pantaleão e
a University of São Paulo, Faculty of Philosophy, Science and Letters at Ribeirão Preto, Department of Biology, Laboratory of Bioecology and Crustacean Systematics, Av. Bandeirantes, 3900, 14040-901, Ribeirão Preto, São Paulo, Brazil
b Universidad de Costa Rica, Unidad de Investigación Pesquera y Acuicultura, Centro de Investigación en Ciencias del Mar y Limnología, 11501-2060 San José, Costa Rica
c Universidad de Costa Rica, Centro de Investigación en Biodiversidad y Ecología Tropical, Museo de Zoología, 11501-2060 San José, Costa Rica
d Universidad del Valle de Guatemala, Editorial Universitaria, 18 Avenida 11-95, Zonas 15, Vista Hermosa III, Ciudad de Guatemala, Guatemala
e Federal University of Alagoas, Institute of Biological and Health Sciences, Laboratory of Bioecology and Development of Crustaceans, Av. Paulo Holanda, 143, 57072-970, Maceió, Alagoas, Brazil
Received: 08 April 2025; accepted: 13 November 2025
Abstract
We present the first description of Zoea I of Macrobrachium crenulatum and compare it with available data of other zoeae of Macrobrachium spp. of the Neotropics to identify morphological characteristics to differentiate the first zoea of M. crenulatum from zoeae of other species of the genus. A female with embryos was collected in the Atlantic drainage of Costa Rica, Central America. After hatching in the laboratory, larvae were preserved in glycerinate alcohol and later semi-permanent slides were prepared for morphological study and larval description. A table was prepared to compare the morphology of the first zoea of M. crenulatum to the first zoeae of other species of Macrobrachium. Among species of Macrobrachium phylogenetic and geographically closely related, the first zoea of M. crenulatum can be differentiated by the comparison of some structures as antenna (number of articles on scaphocerite), maxillipeds (number of plumose setae; setation pattern), maxilla (setation pattern), maxillule (number of spines), and telson (outer pair of setae). The larval morphology of M. crenulatum corroborated the phylogenetic position and may serve as a taxonomic tool for the distinction between the first zoea of closely related species.
Keywords: Atlantic drainage; Caridea; Costa Rica; Larval morphology
Resumen
Presentamos la primera descripción de la zoea I de Macrobrachium crenulatum y la comparamos con datos disponibles de otras primeras zoeas de especies de Macrobrachium del Neotrópico, con el objetivo de identificar características morfológicas que permitan diferenciar la primera zoea de M. crenulatum respecto de las de otras especies del género. Una hembra ovígera fue colectada en la cuenca atlántica de Costa Rica, Centroamérica. Tras la eclosión en laboratorio, las larvas fueron preservadas en alcohol glicerinado y, posteriormente, se prepararon láminas semipermanentes para su estudio morfológico y la descripción larval. Se elaboró una tabla comparativa con la morfología de la primera zoea de M. crenulatum respecto de las primeras zoeas de otras especies de Macrobrachium. Entre las especies filogenética y geográficamente relacionadas, la zoea I de M. crenulatum puede distinguirse mediante la comparación de estructuras como antenas (número de artejos del escafocerito), maxilípedos (número y patrón de setas plumosas), maxila (patrón de setas), maxílula (número de espinas) y telson (disposición del par lateral de setas marginales). La morfología larval de M. crenulatum respalda su posición filogenética y puede servir como herramienta taxonómica para diferenciar la primera zoea de especies estrechamente relacionadas.
Palabras clave: Vertiente Atlántica; Caridea; Costa Rica; Morfología larvaria
Introduction
The genus Macrobrachium Spence Bate, 1868 is one of the most speciose taxa among caridean shrimps, currently accounting for more than 270 species (De Grave & Fransen, 2011; Pantaleão et al., 2025; Poore & Ahyong, 2023). The genus is cosmopolite, with representatives inhabiting different environments with distinct climate conditions throughout the world. Macrobrachium is remarkably diverse in subtropical and tropical regions, such as Central and South America as well as in the Indo-Pacific region (Murphy & Austin, 2005; Pileggi & Mantelatto, 2012; Pileggi et al., 2014). Some species of Macrobrachium are marine, estuarine and anchialine, but the vast majority inhabit typically freshwater systems, such as rivers, lakes and floodplains (Chace & Bruce, 1993; Fujita et al., 2015; Poore & Ahyong, 2023).
Besides the elevated number of species, the genus is highly diverse in terms of morphological characteristics, ecological traits and reproductive strategies (Sastry, 1983; Wowor et al., 2009). Some species of Macrobrachium, for instance, complete their entire life cycle in freshwater. Others are amphidromous and depend on both freshwater and estuarine/marine environments to finish their cycle, which implies migration events from one water system to the other (Bauer, 2011a, b; Liu et al., 2007; Wowor et al., 2009). Another varying feature of the biology and ecology of species of Macrobrachium is the duration of larval development. While some species have extended larval development (ELD) with a long duration of the larval period and the number of larval stages varying between 9 and 13, others have an abbreviated larval development (ALD) with 1 or 2 free-swimming larval stages before reaching the juvenile stage (Anger, 2013; Jalihal et al., 1993; Shokita, 1985; Wowor et al., 2009).
One of the representatives of this shrimp genus is Macrobrachium crenulatum Holthuis, 1950, with its geographical distribution including the coastal waters of Nicaragua, Costa Rica, Panama, Colombia, Venezuela, Jamaica, Grenada, and Trinidad and Tobago (Holthuis, 1952; Hunte, 1979; Pileggi et al., 2014; Rossi, 2012). This species is characterized by an amphidromous life cycle with extended larval development (ELD) (Bauer, 2013; Pileggi et al., 2014). Adults have been the target of taxonomic, ecological and molecular studies (Pileggi et al., 2014; Rossi, 2012; Valencia & Campos, 2007); however, the larval morphology of M. crenulatum has never been described. This lack of information about the larval morphology hinders the global comprehension of species, since understanding larval features is not only useful for the larval recognition itself, but also for studies about the larval ecology, for resolving taxonomic problems and for providing phylogenetic insights (Lai et al., 2013; Pantaleão et al., 2025).
Pileggi et al. (2014) conducted a molecular study, which provided phylogenetic data about the “sibling species” of Macrobrachium from Central America. According to these authors, morphological features of adults are often not sufficient to distinguish the “sibling species” of the genus, including the distinction between transisthmian pairs of sibling species Macrobrachium hancocki Holthuis, 1950 and M. crenulatum. Recently, Pantaleão et al. (2025) conducted a novel study integrating larval morphology and molecular data to understand the phylogenetic context of relevant traits of species of Macrobrachium life history and larval morphology. Nonetheless, the authors highlighted that one of the limitations was the lack of larval descriptions for the vast majority of species of Macrobrachium as well as the accuracy of the available descriptions.
Considering the high diversity of Macrobrachium in Central America, the absence of larval descriptions hinders the identification of their larvae in plankton samples and thus a broader understanding of their larval ecology, especially in river systems where several species of Macrobrachium co-occur (see Lara & Wehrtmann, 2011). Therefore, here we describe for the first time the first zoea of M. crenulatum. The results include a comparison with other newly hatched larvae of species of Macrobrachium, which may serve as a taxonomic tool for the distinction between the first zoeae of these closely related species.
Materials and methods
An ovigerous female (carapace length = 16.6 mm) was collected on 23 May 2010 in the Suarez River (09º43’40” N, 22º50’25.5” W), at the district of Cahuita, in the province of Limón, in the Atlantic drainage of Costa Rica, Central America. The female was collected using a sieve through ascendant movements near the margins of the river; once obtained, it was moved to the laboratory where it was kept alive until the larvae hatched 1 day after the collection. The hatching larvae were collected using a Pasteur pipette and then transferred to a tube containing glycerinate alcohol 80% (1:1). The parental female was preserved in alcohol 80%. Both larvae and the female were deposited at the Coleção de Crustáceos do Departamento de Biologia (CCDB), Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto (FFCLRP), Universidade de São Paulo (USP). Genetic sequences of the parental female were generated by Rossi and Mantelatto (2013) and are available in GenBank under accession numbers JQ805902 for the cytochrome c oxidase subunit I (COI) and JQ805867 for the histone 3 (H3) markers (https://www.ncbi.nlm.nih.gov/nuccore/jq805902 and https://www.ncbi.nlm.nih.gov/nuccore/jq805867).
All the following methodological steps were performed at the Laboratório de Bioecologia e Sistemática de Crustáceos (LBSC/FFCLRP/USP). Firstly, semi-permanent slides were prepared to make possible the morphological study and the larval description (Pantaleão et al., 2011). Before mounting the slides, some larvae were colored with methylene blue 1% to facilitate the visualization of some structures. Slide preparation started with the dissection of larvae in glycerin mounted on a slide. Dissection was carried out under a stereomicroscope Leica™ M205 C equipped with a digital camera Leica™ DFC295. Before mounting the coverslip, larval body structures were organized to avoid over-position. Nail base was used to fix the coverslip. Slides were observed under a microscope Zeiss™ AxioScope A.1 equipped with a Zeiss™ AxioCam MRc 5. Photos of the slides were taken using AxioVision Rel. 4.8. These photos were used as a base for the drawings made posteriorly with the computational program Adobe Illustrator version 24.2.1.
The morphological study involved the analysis, the description and the drawing of the entire specimen with all its appendages, with special attention devoted to the number and shape of cuticular outgrowths (e.g., spines, setae). Larval description was based mainly on Clark et al. (1998) and updates by Clark and Cuesta (2015), but other referential studies on larval morphology were also used as guides for morphological analyses, the description and the drawings (Anger, 2001; Bueno & Rodrigues, 1995; Garm, 2004; Melo & Brossi-Garcia, 2005; Pantaleão et al., 2011; Pohle & Telford, 1981). Information about the larval size was obtained by measuring (see Pantaleão et al., 2021) the total length (TL) from the tip of the rostrum to the posterior margin of the telson, and carapace length (CL) from the posterior margin of the ocular orbit to the posterior margin of the carapace using a stereomicroscope equipped with a digital software (Leica Application Suite version 3.8).
An analysis was carried out to compare the morphology of the first zoea of M. crenulatum in relation to other species of Macrobrachium from the Neotropics that present ELD. We applied 2 criteria for species selection: phylogenetic proximity and similarity in geographic distribution. Considering the phylogenetic hypothesis presented by Pantaleão et al. (2025), our comparison includes M. faustinum (de Saussure, 1857), M. olfersii (Wiegmann, 1836), and M. surinamicum Holthuis, 1948 since all these species are closely related to M. crenulatum. Although M. acanthurus (Wiegmann, 1836), M. heterochirus (Wiegmann, 1836), and M. carcinus are also relatively close to M. crenulatum, their inclusion is particularly relevant because their geographic distributions overlap, at least partially, with that of M. crenulatum, M. olfersii and M. surinamicum. In some cases, this overlap includes freshwater systems in Costa Rica, the country of origin of the larvae examined here, and even cases of sympatry, such as in the Suarez River (Pileggi et al., 2014). Moreover, the inclusion of M. olfersii addresses a particular issue, as this species and M. crenulatum have been considered part of the same species complex (i.e., species complex of M. olfersii) (see Rossi & Mantelatto, 2013). In contrast, some species that are phylogenetically and/or geographically close to M. crenulatum were not included in the comparative analysis because, for several of them (e.g., M. digueti (Bouvier, 1895), M. hancocki Holthuis, 1950, M. hobbsi Nates & Villalobos in Villalobos-Hiriart & Nates-Rodríguez, 1990), no published description of their first zoea is yet available (Pantaleão et al., 2025).
The geographic distributions of all compared species, based on published reports, are presented in Figure 1. Morphological characteristics of Zoea I of all compared species with ELD life cycle were compiled and summarized in Table 1. Additionally, an identification key for the compared species was developed based on morphological traits of the first zoea.
Description
Order Decapoda Latreille, 1802
Suborder Pleocyemata Burkenroad, 1963
Family Palaemonidae Rafinesque, 1815
Genus Macrobrachium Spence Bate, 1868
Macrobrachium crenulatum Holthuis, 1950
Figures 2A-E, 3A-I
Locality: District of Cahuita, province of Limón, Costa Rica.
Material examined: a parental female and larvae mounted in 11 slides, each one containing one larva dissected (8 slides) or entire larvae (3 slides). Accession number: CCDB 3174.
Dimensions of newly hatched larvae: TL = 2.18 ± 0.12 mm; CL = 0.33 ± 0.02 mm (n = 10). Body (Fig. 2A, B): carapace smooth; rostrum long, curved downwards and with the shape of a spine; sessile eyes; pleon with 5 segments; fourth pleonite with 2 posterodorsal simple setae. Antennule (Fig. 2C): uniramous; peduncle long and unarticled; endopod (or primary flagellum) as a long terminal plumose seta; exopod (or accessory flagellum) with 4 terminal aesthetascs, and 1 short terminal plumose seta. Antenna (Fig. 2D): biramous; protopod unarticled with 1 terminal spine near insertion of endopod; endopod (or flagellum) unarticled with 1 short terminal spine and 1 long terminal plumose seta; exopod (or scaphocerite) 5-articled, first article with 2 plumose setae, 1 terminal and 1 subterminal and 1 medial protuberance, second with 2 setae, 1 inner plumose and 1 outer simple, third with 1 inner long plumose seta, fourth with 1 inner long plumose seta, and fifth with 5 setae, 1 long subterminal plumose, 2 long terminal plumose, 1 short terminal plumose, and 1 short terminal simple.
Table 1
Comparison of meristic and morphological characteristics of Zoea I among different species of Macrobrachium in the Neotropics with Extended Larval Development, adapted from Pantaleão et al. (2011) and Vieira et al. (2017). For multi-articled structures, cuticular outgrowths (plumose setae, simple setae or spines) on the same articles are separated by (+), while those on different articles are separated by comma. Other symbols and abbreviations used: aest. = aesthetascs; s. = setae; si. = simple setae; pl. = plumose setae; sp. = spine; (c.o.) = cuticular outgrowths; (*) = larval description and larval illustration are divergent; (**) = data exclusively based on larval illustration; description not available; (-) = information about the morphological trait unavailable; (?) = description of the appendage unavailable; (;) = data from 2 different larval descriptions.
7 + 7 plumose sp.; 1 + 1 si. (the innermost pair without setulae) and 6 + 6 pl.
7 + 7 pl., minute setae between 3 + 3 inner setae
7 + 7 pl., small sp. between 3 + 3 inner setae
Last pair of (c.o.) (most lateral)
With setulae only on inner margin
With setulae only on inner margin
With setulae only on inner margin**
With setulae only on inner margin
With setulae only on inner margin
With setulae on inner and outer margins
With setulae only on inner margin
Mandibles (Fig. 3A, B): both rudimentary, lacking palp; right mandible with molar process smooth, and incisor process with 2 subterminal teeth and 1 terminal sharp tooth of varying sizes; left mandible with molar and incisor process not clearly defined, bearing 2 terminal spines, 1 smaller and 1 larger. Maxillule (Fig. 3C): coxal and basial endites with 4 and 5 terminal simple setae, respectively; endopod with 2 terminal simple setae. Maxilla (Fig. 3D): biramous; coxal endite with 4 setae, 1 simple and 3 plumose; basial endite bilobed, with 3 and 2 simple setae on proximal and distal lobes, respectively; endopod (or palp) bilobed, with 2 proximal simple and 1 terminal plumose seta; exopod (or scaphognathite) margin with 4 proximal plumose setae and 1 long terminal plumose process; microtrichia on margins of endopod and exopod.
Figure 1. Geographic distribution of compared species of Macrobrachium with extended larval development from the Neotropical region (Almeida et al., 2008; Bowles et al., 2000; Ferreira et al., 2010; Magalhães & Pereira, 2007; Murienne et al., 2022; Pileggi & Mantelatto, 2010; Pileggi et al., 2014; Pompeu et al., 2006; Rossi & Mantelatto, 2013; Tejeda-Mazariegos et al., 2018; Valencia & Campos, 2007).Figure 2. First zoea of Macrobrachium crenulatum Holthuis, 1950. A) Dorsal view; B) lateral view; C) antennule; D) antenna; E) telson. Scale bars (mm) = A, B) 0.16; C) 0.06; D) 0.09; E) 0.12.
First maxilliped (Fig. 3E): biramous; coxa without setae; basis with 4 short simple setae arranged 1 + 2 + 1; endopod unarticled with 3 terminal simple setae, 1 short and 2 long; exopod unarticled, with 4 long terminal plumose natatory setae; endopod approximately 3 times shorter than exopod. Second maxilliped (Fig. 3F): biramous; coxa without setae; basis with 1 short simple seta; endopod 3-articled, first with 0, second with 1 terminal plumose, and third with 5, 1 short proximal simple, 3 short terminal simple and 1 short terminal sparsely plumose setae, respectively; exopod unarticled with 4 long terminal plumose natatory setae; endopod slightly shorter than exopod.
Figure 3. First zoea of Macrobrachium crenulatum Holthuis, 1950. A) Left mandible; B) right mandible; C) maxillule; D) maxilla; E) first maxilliped; F) second maxilliped; G) third maxilliped; H) first pereiopod; I) second pereiopod. Scale bars (mm) = A, B) 0.2; C) 0.08; D, E) 0.09; F, G) 0.1; H, I) 0.05.
Third maxilliped (Fig. 3G): biramous; coxa and basis without setae; endopod 4-articled, first article without setae, second with 1 short proximal simple, and 1 short subterminal simple, third with 2 long terminal plumose, and fourth with 3 terminal setae, 1 long simple, 1 short plumose and 1 long plumose; exopod unarticled with 4 long terminal plumose natatory setae; endopod and exopod of similar length. Pereiopods (Fig. 3H, I): first and second rudimentary and biramous, second slightly longer than the first; third, fourth and fifth absent. Telson (Fig. 2E): posterior bilobed region broader than the anterior, with 7 + 7 plumose setae, outer 2 plumose only in inner margin; minute spinules along the posterior margin and around the bases of 3 + 3 inner setae.
Remarks
The morphological description of Zoea I of Macrobrachium crenulatum enables the comparison among the first zoeae of closely related congeners as well as the definition of some morphological characteristics. All the species included in the comparison exhibit type I development, characterized by an extended larval development and an amphidromous life cycle (Bauer, 2013). This contrasts with type II development, which involves a partially abbreviated larval development and is common in freshwater species, and type III development, which is marked by a completely abbreviated larval development and is typical of species inhabiting mountain streams (Jalihal et al., 1993; Pileggi et al., 2014; Vieira et al., 2017). Adults of amphidromous species typically inhabit both freshwater and brackish environments, except for M. crenulatum, M. faustinum, and M. heterochirus, whose adults have been reported exclusively in freshwater (Holthuis, 1950; Hunte, 1980; Vega-Perez, 1984; Vieira et al., 2017).
Before comparing the morphology of the Zoea I among species, it is important to consider that most of the larval descriptions included in Table 1 were published at different times, each reflecting the research context on decapod larval morphology of that period. Thus, many of the current consensuses on terminology (e.g., Clark et al., 1998) postdate some of the descriptions examined. Moreover, recent studies employ more advanced technologies than those available in the 1970s, 80s and 90s, enabling, for example, more accurate and detailed descriptions of setal types and number of articles. Therefore, some inconsistencies were detected during the comparisons of the published descriptions. Thus, we aimed to distinguish true morphological differences of the larvae from inconsistencies likely resulting from the limited technological capabilities used in earlier studies. Although we kept the original terminology used by the authors in our table, in the following comparison we focus on taxonomic informative differences, highlighting instances where a variation may result from misinterpretation.
Macrobrachium crenulatum and M. olfersii have been considered part of the same species complex due to the high morphological similarity of adults (Rossi & Mantelatto, 2013; Villalobos-Figueroa, 1967). Their close phylogenetic relationship was later confirmed using molecular data by Pileggi et al. (2014) and, more recently, by Pantaleão et al. (2025). For M. olfersii, 2 larval descriptions of first zoea are available. Dugger and Dobkin (1975) provided the first, describing Zoea I of this species from females collected in Florida (USA). Later, Melo and Brossi-Garcia (2005) provided a description for hatched larvae of Macrobrachium birai Lobão, Melo & Fernandes, 1986, obtained from a female collected in São Paulo (Brazil). Subsequently, however, M. birai was regarded as a junior synonymous of M. olfersii by Pileggi and Mantelatto (2012). Thus, both studies are treated here as descriptions of the first zoeal stage of M. olfersii. Although generally similar, both descriptions show some differences in the number of cuticular outgrowths on the antennal flagellum, maxilla coxal endite, and maxilliped endopods. Considering that each study worked with larvae from distinct latitudes (USA and Brazil) with different temperature regimes and oceanographic dynamics, these differences may reflect plasticity in larval morphology, a phenomenon that has partially explored in the scientific literature (Sandifer & Smith, 1979; Thatje & Bacardit, 2000; Wehrtmann & Albornoz, 2003). For the comparison between M. crenulatum and M. olfersii, we focused only on features that are consistent across both descriptions of M. olfersii, avoiding traits that showed intraspecific variation. In Table 1, when differences are observed in the description of Zoea I of M. olfersii, data from both descriptions are presented (Dugger & Dobkin, 1975; Melo & Brossi-Garcia, 2005).
The maxillipeds are considerably distinct between M. crenulatum and M. olfersii. They differ by the number of plumose setae on the third maxilliped exopod: Macrobrachium crenulatum presents only 4 terminal plumose setae, while the first zoea of M. olfersii has 2 subterminal plumose setae in addition to the 4 terminal ones. Moreover, the basis of the first maxilliped bears one simple seta in M. olfersii, but 4 (1 + 2 + 1) in M. crenulatum. Finally, it is important to mention that the larval description published by Dugger and Dobkin (1975) reports a difference in the number of articles on the exopod of maxillipeds: 2 in M. olfersii versus only one in M. crenulatum. However, aside from being absent in the description from Melo and Brossi-Garcia (2005), this apparent difference may result from a misinterpretation of the number of articles in maxilliped exopods of M. olfersii, since the presence of only 1 article appears to be a consistent pattern among American species of Macrobrachium, regardless of whether they exhibit type I development (e.g., M. carcinus, M. heterochirus and M. surinamicum) or type II (e.g., Macrobrachium iheringi [Ortmann, 1897] and Macrobrachium jelskii [Miers, 1877]) (Bueno & Rodrigues, 1995; Choudhury, 1970, 1971; Magalhães, 2000; Pantaleão et al., 2025; Vega-Perez, 1984; Vieira et al., 2017).
Several morphological features can be used to distinguish the first zoeae of M. crenulatum and M. carcinus, 2 species whose geographic distributions partially overlap. Notably, these 2 species differ in the pattern of setae present on the endopod of the third maxilliped: M. crenulatum has setae on all 4 articles of the third maxilliped endopod (1, 1, 2, 1 + 2), while Zoea I of M. carcinus presents setae only on the last article (0, 0, 0, 1 + 2 or 3) (Choudhury, 1971). Analysis of the larval description of M. carcinus reveals possible inconsistencies in terminology. In several instances, the term “spine” is used to refer to a “seta” and vice-versa. Additionally, although illustrated, the second maxilliped of the first zoea of M. carcinus was not formally described, being mentioned only briefly within the description of the third maxilliped. These inconsistencies hinder a comprehensive morphological comparison and complicate phylogenetic inferences based on larval characters. Therefore, given the aforementioned issues related to technological advances and terminology standardization, we strongly recommend a redescription of the first zoeal stage of M. carcinus.
Macrobrachiumheterochirus is a species closely related to M. crenulatum both geographically and phylogenetically (Pantaleão et al., 2025; Pileggi et al., 2014), and the adults are morphologically distinct enough to be readily distinguished. Zoea I of both species, in turn, differ on the setal pattern of maxilla, both in the number and type of setae on coxal and basial endites: Macrobrachium crenulatum has 4 setae, one simple and 3 plumose, on the coxal endite and 5 simple setae on the basial endite, whereas the first zoea of M. heterochirus bears 5 setae, 4 plumose and one simple, on the coxal endite, and the basial endite carries only 4 plumose setae. According to the descriptions, these species also differ in the number of aesthetascs on the antennule exopod: 4 in M. crenulatum, and 3 in M. heterochirus (Vega-Perez, 1984). However, this difference may reflect a misinterpretation. Possibly, the simple seta described by the author as part of antennule exopod of M. heterochirus could in fact be an aesthetasc, bringing the total to 4, as observed in other species such as M. crenulatum and M. olfersii. Similar misunderstandings regarding the number of aesthetascs of the antennule may also applied to the descriptions of M. acanthurus, M. carcinus, M. faustinum, and M. surinamicum.
Several morphological characteristics allow the distinction between Zoea I of M. crenulatum and M. acanthurus, both species occurring in Costa Rica (Fig. 1). One of the most conspicuous features for identification is the number of aesthetascs on the antennule (4 on the first and only 3 on the latter), although this difference may reflect a misinterpretation in the earliest description, which dates back 56 years (Choudhury, 1970). Additionally, M. crenulatum has a spine on the protopod of the antenna, which is absent in M. acanthurus. Another notable difference lies in the number of endopod articles of the second maxilliped: Macrobrachium crenulatum has 3, whereas M. acanthurus presents 4, and the setal pattern on this structure also differs (Table 1). Finally, the telson of M. crenulatum bears small spines between the 3 innermost plumose setae, while in M. acanthurus, spines are present between all the setae of the telson.
As with M. acanthurus, several traits can be used to distinguish Zoea I of M. crenulatum and M. faustinum (Table 1), species that are relatively close phylogenetically (Pantaleão et al., 2025). Notably, the number of aesthetascs present in the antennule differs, with M. crenulatum having 4 and M. faustinum only 3. As previously noted, this feature should be interpreted cautiously, as the older description may not be fully accurate in this regard. Additionally, the scaphocerite of M. faustinum lacks a spine present in the first larval stage of M. crenulatum, and the molar process of the right mandible in M. crenulatum is unarmed, whereas in M. faustinum it bears 2 protuberances (Hunte, 1980).
Finally, several larval characteristics can also be used to identify the first zoeae of M. crenulatum and M. surinamicum, species relatively closely related according to Pantaleão et al. (2025). The differences between the newly hatched larvae refer to the number of articles on the scaphocerite (5 in M. crenulatum and 6 in M. surinamicum), the number of spines on basial endite of maxillule (5 in M. crenulatum and 2 in M. surinamicum), and the position of setulae on the outer pair of setae of telson margin. In M. crenulatum, setulae occur only on inner margin of the outer pair of setae of the distal margin of the telson, whereas in M. surinamicum, they are present on both the inner and outer margins (Table 1). Interestingly, none of the other species with described Zoea I included in Table 1 have setulae on the outer margin of this pair of setae. Therefore, the phylogenetic significance of this trait warrants to be analyzed in future studies.
The present study represents a first step toward improving our knowledge of larval morphology of M. crenulatum. Although it focuses only on the first larval stage of a single brood of Macrobrachium species, the results contribute to a broader effort of providing larval descriptions for Neotropical species of the genus. This information offers valuable insights for both theoretical studies about the evolution of the morphology of Macrobrachium and practical applications in larval ecology and aquaculture research focused on larval development. Additionally, the investigation of the larval morphology of M. crenulatum helps to fill a gap in the larval descriptions of a lineage of species of Macrobrachium for which no larval data were previously available (Pantaleão et al., 2025). This line of research should be continued by describing the larvae of other species phylogenetically close to M. crenulatum, such as M. digueti and M. hancocki, to broaden our understanding of phylogenetic relationships among these freshwater shrimps. Finally, our results highlight the need for redescriptions of the first zoea of several species of Macrobrachium, aimed at updating the larval terminology and clarifying ambiguous characters, which would in turn facilitate morphological comparisons and phylogenetic analysis.
Identification key for the compared species of Macrobrachium with ELD from Neotropics based on morphological traits of Zoea I
1. First article of third maxilliped endopod with setae 2
1’. First article of third maxilliped endopod without setae 3
2. Scaphocerite with 5 articles; basial endite of maxillule with 5 spines; most lateral pair of setae of telson margin with setulae only on inner margin M. crenulatum
2’. Scaphocerite with 6 articles; basial endite of maxillule with 2 spines; most lateral pair of setae of telson margin with setulae both on inner and outer margins M. surinamicum
3. Basial endite of first maxilliped with more than 1 seta; endopod of third maxilliped with 4 articles 4
3’. Basial endite of first maxilliped with only 1 seta; endopod of third maxilliped with 3 articles M. olfersii
4. Scaphocerite with 10 plumose setae; first maxilliped endopod with 3 cuticular outgrowths 5
4’. Scaphocerite with 11 plumose setae; first maxilliped endopod with 4 cuticular outgrowths M. heterochirus
5. Scaphocerite with 12 cuticular outgrowths; maxilla coxal endite with 4 plumose setae 6
5’. Scaphocerite with 11 cuticular outgrowths; maxilla coxal endite with 3 plumose setae M. faustinum
6. Lobes of maxilla basial endite with 2 plumose setae each M. acanthurus
6’. Lobes of maxilla basial endite with 2 plumose setae and 1 simple seta each M. carcinus
Acknowledgements
This study is result of the course “Special Topics in Comparative Biology: Larval morphology and phylogenetic relationships in decapod crustaceans” under the responsibility of FLM and JAFP and as part of Postgraduate Program in Comparative Biology at Faculty of Philosophy, Sciences and Letters at Ribeirão Preto. The authors are grateful to the LBSC team, in special to Gabriel L. Bochini, who helped with some dissections and slide preparation. We acknowledge the financial support provided by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (Projetos Temáticos BIOTA Proc. 2010/50188-8 and INTERCRUSTA Proc. 2018/13685-5) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (PPBio 07/2023 – Linha 8: Rede Costeira Marinha Proc. 442421/2023-0). FLM and ISW are thankful for the financial contribution supplied by CNPq (Proc. 491490/2004-6, 490353/2007-0, 490314/2011-2; 471011/2011-8) and CONICIT – Costa Rica (CII- 001-08, IQ-0001-11) during the development of the International Cooperative Project, which was crucial for the collection of the female with embryos and collaboration in the Brazil-Costa Rica research program. FLM thanks CNPq for an ongoing research grant (PQ 302706/2025-9). LOR and RP thank PhD grants provided by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (PROEX – Proc. 88887.804708/2023-00 and 88887.896987/2023-00, respectively). RP and LOR also thank the ongoing PhD fellowships by FAPESP (Proc. 2024/15232-9 and 2025/01018-8, respectively). The ovigerous female was collected under the collection and genetic access permit (Resolución No. 377 – Comisión Institucional de Biodiversidad de la Universidad de Costa Rica) to ISW. We also thank the anonymous reviewers and associate editor for their valuable comments, which improved the quality of the manuscript.
References
Almeida, A. O., Alves-Coelho, P., Rocha-Luz, J., Almeida-dos Santos, J. T., & Ribeiro-Ferraz, N. (2008). Decapod crustaceans in fresh waters of southeastern Bahia, Brazil. Revista de Biología Tropical, 56, 1225–1254. https://doi.org/10.15517/rbt.v56i3.5706
Anger, K. (2001). The biology of decapod crustacean larvae. Lisse, South Holland: AA Balkema Publishers.
Anger, K. (2013). Neotropical Macrobrachium (Caridea: Palaemonidae): on the biology, origin, and radiation of freshwater-invading shrimp. Journal of Crustacean Biology, 33, 151–183. https://doi.org/10.1163/1937240X-00002124
Bauer, R. T. (2011a). Amphidromy and migrations of freshwater shrimps. I. Costs, benefits, evolutionary origins, and an unusual case of amphidromy. New Frontiers in Crustacean Biology, 15, 145–156. https://doi.org/10.1163/ej.9789004174252.i-354.108
Bauer, R. T. (2011b). Amphidromy and migrations of freshwater shrimps. II. Delivery of hatching larvae to the sea, return juvenile upstream migration, and human impacts. New Frontiers in Crustacean Biology, 15, 157–168. https://doi.org/10.1163/EJ.9789004174252.I-354.115
Bauer, R. T. (2013). Amphidromy in shrimps: a life cycle between rivers and the sea. LatinAmerican Journal of Aquatic Research, 41, 633–650. https://doi.org/10.3856/vol41-issue4-fulltext-2
Bouvier, E. L., (1895). Sur les palémons receuillis dans les eaux douces de la Basse-Californie par M. Diguet. Bulletin du Muséum d’Histoire Naturelle, 1, 159–162.
Bowles, D. E., Aziz, K., & Knight, C. L. (2000). Macrobrachium (Decapoda: Caridea: Palaemonidae) in the contiguous United States: a review of the species and an assessment of threats to their survival. Journal of Crustacean Biology, 20, 158–171. https://doi.org/10.1163/20021975-99990025
Bueno, S. L. S., & Rodrigues, S. A. (1995). Abbreviated larval development of the freshwater prawn, Macrobrachium iheringi (Ortmann, 1897) (Decapoda, Palaemonidae), reared in the laboratory. Crustaceana, 68, 665–686. https://doi.org/10.1163/156854095X01880
Burkenroad, M. D. (1963). The evolution of the Eucarida (Crustacea, Eumalacostraca), in relation to the fossil record. Tulane Studies in Geology, 2, 3–17.
Clark, P. F., Calazans, D. K., & Pohle, G. W. (1998). Accuracy and standardization of brachyuran larval descriptions. Invertebrate Reproduction & Development, 33, 127–144. https://doi.org/10.1080/07924259.1998.9652627
Clark, P. F., & Cuesta, J. A. (2015). Larval systematics of Brachyura. In P. Castro, P. J. F. Davie, D. Guinot, F. R. Schram, & J. C. von Vaupel-Klein (Eds.), Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea 9C (pp. 981–1048). Leiden, South Holland: Brill. https://doi.org/10.1163/9789004190832_020
Chace, F. A., & Bruce, A. J. (1993). The caridean shrimps (Crustacea: Decapoda) of the Albatross Philippine expedition 1907-1910, part 6: superfamily Palaemonoidea. Smithsonian Contributions to Zoology, 543, 1–152. https://doi.org/10.5479/si.00810282.543
Choudhury, P. C. (1970). Complete larval development of the palaemonid shrimp Macrobrachium acanthurus (Wiegmann, 1836), reared in the laboratory. Crustaceana, 18, 113–132. https://doi.org/10.1163/156854070X00743
Choudhury, P. C. (1971). Complete larval development of the palaemonid shrimp Macrobrachium carcinus (L.), reared in the laboratory (Decapoda, Palaemonidae). Crustaceana, 20, 51–69. https://doi.org/10.1163/156854071X00526
De Grave, S., & Fransen, C. H. J. M. (2011). Carideorum catalogus: the recent species of the dendrobranchiate, stenopodidean, procarididean and caridean shrimps (Crustacea: Decapoda). Zoologische Mededelingen, 85, 195–589.
de Saussure, H. (1857). Diagnoses de quelques Crustacés nouveaux de l’Amérique tropicale. Revue et Magasin de Zoologie, Ser. 2, 9, 501–505.
Dugger, D. M., & Dobkin, S. (1975). A contribution to knowledge of the larval development of Macrobrachium olfersii (Wiegmann, 1836) (Decapoda, Palaemonidae). Crustaceana, 29, 1–30. https://doi.org/10.1163/156854075×00018
Ferreira, R. S., Vieira, R. R. R., & D’Incao, F. (2010). The marine and estuarine shrimps of the Palaemoninae (Crustacea: Decapoda: Caridea) from Brazil. Zootaxa, 2606, 1–24. https://doi.org/10.11646/zootaxa.2606.1.1
Fujita, Y., Davie, P. J., & Ng, P. K. (2015). A new stygobitic prawn of the genus Macrobrachium Spence Bate, 1864, from anchialine caves in Christmas Island, Indian Ocean; with a rediagnosis of M. miyakoense Komai & Fujita, 2005 (Crustacea: Decapoda: Caridea: Palaemonidae). Raffles Bulletin of Zoology, 63, 610–625.
Garm, A. (2004). Revising the definition of the crustacean seta and setal classification systems based on examination of the mouthpart setae of seven species of decapods. Zoological Journal of the Linnean Society, 142, 233–252. https://doi.org/10.1111/j.1096-3642.2004.00132.x
Holthuis, L. B. (1948). Note on some Crustacea Decapod Natantia from Surinam. Proceedings van de Koninklijke Nederlandsche Akademie van Wetenschappen, 51, 1104–1113.
Holthuis, L. B. (1950). Preliminary descriptions of twelve new species of palaemonid prawns from American waters (Crustacea Decapoda). Proceedings van de Koninklijke Nederlandsche Akademie van Wetenschappen, 53, 93–99.
Holthuis, L. B. (1952). A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. II. The subfamily Palaemonidae. Occasional Papers of the Allan Hancock Foundation, 12, 1–396.
Hunte, W. (1979). The rediscovery of the freshwater shrimp Macrobrachium crenulatum in Jamaica. Studies on the Fauna of Curaçao and other Caribbean Islands, 58, 69–74.
Hunte, W. (1980). The larval development of the shrimp Macrobrachium faustinum reared at the laboratory. Caribbean Journal of Science, 15, 49–68.
Jalihal, D. R., Sankolli, K. N., & Shenoy, S. (1993). Evolution of larval developmental patterns and the process of freshwaterization in the prawn genus Macrobrachium Bate, 1868 (Decapoda, Palaemonidae). Crustaceana, 65, 365–376. https://doi.org/10.1163/156854093X00793
Lai, J. C. Y., Thoma, B. P., Clark, P. F., Felder, D. L., & Ng, P. K. L. (2013). Phylogeny of eriphioid crabs (Brachyura, Eriphioidea) inferred from molecular and morphological studies. Zoologica Scripta, 43, 52–64. https://doi.org/10.1111/zsc.12030
Lara, R., & Wehrtmann, I. S. (2011). Diversity, abundance and distribution of river shrimps (Decapoda: Caridea) in the largest river basin of Costa Rica, Central America. In A. Asakura & C. Fransen (Eds.), New Frontiers in Crustacean Biology (pp. 197–211). Leiden, South Holland: Brill. http://dx.doi.org/10.1163/ej.9789004174252.i-354.140
Latreille, P. A. (1802). Histoire naturelle, générale et particulière des crustacés et des insectes. Familles naturelles des genre. Ouvrage faisant suite à l‘Histoire Naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieurs Sociétés savantes. de L‘imprimerie de F. Dufart, Paris.
Liu, M. Y., Cai, Y. X., & Tzeng, C. S. (2007). Molecular systematics of the freshwater prawn genus Macrobrachium Bate, 1868 (Crustacea: Decapoda: Palaemonidae) inferred from mtDNA sequences, with emphasis on East Asian species. Zoological Studies, 46, 272–289.
Lobão, V. L., Melo, G. A. S., & Fernandes, W. (1986). Descrição de uma nova espécie do gênero Macrobrachium (Crustacea: Decapoda: Palaemonidae) da região sul de São Paulo. In Resumos do Congresso Brasileiro de Zoologia, Volume 13. Cuiabá.
Magalhães, C. (2000). Abbreviated larval development of Macrobrachium jelskii (Miers, 1877) (Crustacea: Decapoda, Palaemonidae) from the Rio Solimões floodplain, Brazil, reared in the laboratory. Nauplius, 8, 1–14.
Magalhães, C., & Pereira, G. (2007). Assessment of the decapod crustacean diversity in the Guayana Shield region aiming at conservation decisions. Biota Neotropica, 7, 111–124. https://doi.org/10.1590/s1676-06032007000200013
Melo, S. G., & Brossi-Garcia, A. L. (2005). Desenvolvimento larval de Macrobrachium birai Lobão, Melo & Fernandes (Crustacea, Decapoda, Caridea, Palaemonidae) em laboratório. Revista Brasileira de Zoologia, 22, 131–152. https://doi.org/10.1590/S0101-81752005000100016
Miers, E. J. (1877). On a collection of Crustacea, Decapoda and Isopoda, chiefly from South America, with descriptions of new genera and species. Proceedings of the Zoological Society of London, 43, 653–679.
Murienne, J., Chevalier, J., & Clavier, S. (2022). On the presence of the giant freshwater prawn, Macrobrachium rosenbergii, in French Guiana confirmed by citizen science and genetic analyses. Water Biology and Security, 1, 100039. https://doi.org/10.1016/j.watbs.2022.100039
Murphy, N. P., & Austin, C. M. (2005). Phylogenetic relationships of the globally distributed freshwater prawn genus Macrobrachium (Crustacea: Decapoda: Palaemonidae): biogeography, taxonomy and the convergent evolution of abbreviated larval development. Zoologica Scripta, 34, 187–197. https://doi.org/10.1111/j.1463-6409.2005.00185.x
Ortmann, A. (1897). Os camarões da agua doce da America do Sul. Revista do Museu Paulista, 2, 173–216.
Pantaleão, J. A. F., Gregati, R. A., Taddei, F. G., & Costa, R. C. (2011). Morphology of the first larval stage of Macrobrachium brasiliense (Heller, 1868) (Caridea: Palaemonidae). Nauplius, 19, 79–85. https://doi.org/10.1590/S0104-64972011000100009
Pantaleão, J. A. F., Mantelatto, F. L., & Costa, R. C. (2021). The first larval stage (Zoea I) description of the caridean shrimp Ogyrides occidentalis (Ortmann, 1893) (Alpheoidea: Ogyrididae) reveals congruence with taxonomic status. Biota Neotropica, 21, e20201118. https://doi.org/10.1590/1676-0611-BN-2020-1118
Pantaleão, J. A. F., Peres, P. A., & Mantelatto, F. L. (2025). Mapping the congruence between larval morphology and molecular phylogeny: Convergent adaptations of fresh- water shrimps (Decapoda, Caridea). Zoologica Scripta, 54, 376–393. https://doi.org/10.1111/zsc.12709
Pileggi, L. G., & Mantelatto, F. L. (2010). Molecular phylogeny of the freshwater prawn genus Macrobrachium (Decapoda, Palaemonidae), with emphasis on the relationships among selected American species. Invertebrate Systematics, 24, 194–208. https://doi.org/10.1071/IS09043
Pileggi L. G., & Mantelatto, F. L. (2012). Taxonomic revision of doubtful Brazilian freshwater shrimp species of genus Macrobrachium (Decapoda, Palaemonidae). Iheringia, Série Zoologia, 102, 426–437. https://doi.org/10.1590/S0073-47212012005000012
Pileggi, L. G., Rossi, N., Wehrtmann, I. S., & Mantelatto, F. L. (2014). Molecular perspective on the American transisthmian species of Macrobrachium (Caridea, Palaemonidae). Zookeys, 457, 109–131. https://doi.org/10.3897/zookeys.457.6818
Pohle, G. W., & Telford, M. (1981). Morphology and classification of decapod crustacean larval cerdae: a scanning electron microscope study of Dissodactylus crinitichelis Moreira, 1901 (Brachyura: Pinnotheridae). Bulletin of Marine Science, 31, 736–752.
Pompeu, P. D. S., Vieira, F., & Martinez, C. B. (2006). Utilização do mecanismo de transposição de peixes da Usina Hidrelétrica Santa Clara por camarões (Palaemonidae), bacia do rio Mucuri, Minas Gerais, Brasil. Revista Brasileira de Zoologia, 23, 293–297. https://doi.org/10.1590/s0101-81752006000100024
Poore, G. C., & Ahyong, S. T. (2023). Marine decapod Crustacea: a guide to families and genera of the world. Clayton, Victoria: CSIRO Publishing. https://doi.org/10.1071/9781486311798
Rafinesque, C. S. (1815). Analyse de la nature ou Tableau de l’univers et des corps organisés. Palermo: Published by the author.
Rossi, N. (2012). Revisão das espécies de Macrobrachium Bate, 1868, pertencentes ao complexo M. olfersii (Crustacea, Palaemonidae): análises morfológicas e moleculares(M.Sc. Thesis). Universidade de São Paulo. Brazil, São Paulo.
Rossi, N., & Mantelatto, F. L. (2013). Molecular analysis of the freshwater prawn Macrobrachium olfersii (Decapoda, Palaemonidae) supports the existence of a single species throughout its distribution. Plos One, 8, e54698. https://doi.org/10.1371/journal.pone.0054698
Sandifer, P. A., & Smith, T. I. (1979). Possible significance of variation in the larval development of palaemonid shrimp. Journal of Experimental Marine Biology and Ecology, 39, 55–64. https://doi.org/10.1016/0022-0981(79)90004-2
Sastry, A. N. (1983). Ecological aspects of reproduction. In F. J. Vernberg, & W. B. Vernberg (Eds.), The biology of Crustacea: environmental adaptations, Volume 8 (pp. 179–270). New York: Academic Press.
Shokita, S. (1985). Larval development of the palaemonid prawn, Macrobrachium grandimanus (Randall), reared in the laboratory, with special reference to larval dispersal. Zoological Science, 2, 785–803.
Spence Bate, C. (1868). On a new genus, with four new species, of freshwater prawns. Proceedings of the Zoological Society of London, 363–368.
Tejeda-Mazariegos, J. C., Ortiz, L. M. M., López-Mejía, M., Crandall, K. A., Pérez-Losada, M., & Frausto-Martínez, O. (2018). Freshwater crustacean decapods: an important resource of Guatemala. In S. Ray (Ed.), Biological resources of water (pp. 169–180). London: IntechOpen. https://doi.org/10.5772/intechopen.73638
Thatje, S., & Bacardit, R. (2000). Morphological variability in larval stages of Nauticaris magellanica (A. Milne Edwards, 1891) (Decapoda: Caridea: Hippolytidae) from South American waters. Bulletin of Marine Science, 66, 375–398.
Valencia, D. M., & Campos, M. R. (2007). Freshwater prawns of the genus Macrobrachium Bate, 1868 (Crustacea: Decapoda: Palaemonidae) of Colombia. Zootaxa, 1456, 1–44. https://doi.org/10.11646/zootaxa.1456.1.1
Vega-Perez, L. A. (1984). Desenvolvimento larval de Macrobrachium heterochirus (Wiegmann, 1836), Macrobrachium amazonicum (Heller, 1862) e Macrobrachium brasiliense (Heller, 1868) (Crustacea, Decapoda, Palaemonidae), em laboratório(Ph.D. Thesis). Universidade de São Paulo. Brazil, São Paulo.
Vieira, R. R. R., Santos, C. R. M., & Anker, A. (2017). Morphology of the first zoeal stage of Macrobrachium surinamicum Holthuis, 1948 (Malacostraca, Decapoda, Palaemonidae) hatched in the laboratory. Crustaceana, 90, 1337–1350. https://doi.org/10.1163/15685403-00003723
Villalobos-Figueroa, A. (1967). Problemas de especiación en América de un grupo de Palaemonidae del género Macrobrachium. FAO Fisheries Report, 57, 1055–1066.
Villalobos-Hiriart, J. L., & Nates-Rodríguez, J. C. (1990). Dos especies nuevas de camarones de agua dulce del género Macrobrachium Bate, (Crustacea, Decapoda, Palaemonidae), de la vertiente occidental de México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 61, 1–11.
Wehrtmann, I. S., & Albornoz, L. (2003). Larvae of Nauticaris magellanica (Decapoda: Caridea: Hippolytidae) reared in the laboratory differ morphologically from those in nature. Journal of the Marine Biological Association of the United Kingdom, 83, 949–957. https://doi.org/10.1017/S0025315403008130h
Wiegmann, A. (1836). Beschreibung einiger neuen Crustaceen des Berliner Museums aus Mexiko und Brasilien. Archiv für Naturgeschichte, 2, 145–151.
Wowor, D., Muthu, V., Meier, R., Balke, M., Cai, Y., & Ng, P. K. L. (2009). Evolution of life history traits in Asian freshwater prawns of the genus Macrobrachium (Crustacea: Decapoda: Palaemonidae) based on multilocus molecular phylogenetic analysis. Molecular Phylogenetics and Evolution, 52, 340–350. https://doi.org/10.1016/j.ympev.2009.01.002
Received: 12 November 2024; accepted: 14 November 2025
Abstract
Non-native mollusk species can have significant ecological and economic impacts on invaded habitats. In recent decades, the number of non-native mollusks in Chile has steadily increased. This study reports the range expansion of Vallonia pulchella (Müller, 1774) and the first record of Vallonia cf. excentrica Sterki, 1893 in the country, using optical microscopy, scanning electron microscopy, and statistical analysis. The first species was found in Caldera (northern Chile) and in La Florida (Santiago, central Chile), while the latter was found only in the last locality. Vallonia pulchella is a potential intermediate host for parasitic worms that can affect local livestock, highlighting the need for sanitary control measures. The discovery of Vallonia cf. excentrica represents the first record of this species in South America.
Keywords:Alien species; Microgastropods; Morphological identification; New records; Terrestrial snails
Resumen
Las especies de moluscos no nativos pueden tener impactos ecológicos y económicos significativos en los hábitats invadidos. En las últimas décadas, el número de moluscos no nativos en Chile ha aumentado de manera constante. Este estudio documenta la expansión del rango de Vallonia pulchella (Müller, 1774) y el primer registro de Vallonia cf. excentrica Sterki, 1893 en el país, utilizando microscopía óptica, microscopía electrónica de barrido y análisis estadístico. La primera especie se encontró en Caldera (norte de Chile) y en La Florida (Santiago, centro de Chile), mientras que la segunda solo en esta última localidad. Vallonia pulchella es un huésped intermediario potencial para gusanos parásitos que pueden afectar al ganado local, lo que resalta la necesidad de medidas de control sanitario. El descubrimiento de Vallonia cf. excentrica representa el primer registro de esta especie en América del Sur.
Non-native mollusk species can cause significant ecological and economic impacts on invaded habitats (Darrigran et al., 2020; Lowe et al., 2000; Robinson, 1999; Sylvester & Sardiña, 2015). These species often disrupt local ecosystems by outcompeting native species for resources, altering food webs, and causing declines in biodiversity (Alonso et al., 2025; Collado et al., 2025; Vaughn, 2018). Their ability to rapidly colonize new areas can lead to changes in habitat structure and function, sometimes resulting in the loss of native species that are unable to compete with or adapt to the invaders. In addition, invasive mollusks can introduce new pathogens or parasites that harm local wildlife, livestock, and even humans (Cádiz et al., 2013; Collado et al., 2025).
During the last few decades, the introduction of non-native mollusks in Chile has steadily increased, with recent records of terrestrial and freshwater species on Easter Island and especially in the central region of the country (e.g., Cádiz & Gallardo, 2007; Cádiz et al., 2013; Collado, 2014, 2017; Darrigran et al., 2020; Kirch et al., 2009; Letelier et al., 2007). The number of non-native mollusks in Chile has grown from 13 species (Valdovinos-Zarges, 1999) to 34 species (Araya, 2015), including marine, freshwater, and terrestrial species. Of these, 20 are ground-dwelling terrestrial mollusks, making this the largest informal group of invasive species in any South American country, with numbers similar to those found in Argentina (Virgillito & Miquel, 2013). Most of these introduced species have a European origin, and some are known to negatively impact endemic biota, often by competing with or preying on native species, or acting as hosts for various parasites (Cádiz et al., 2013). Prevention and continuous monitoring are therefore essential to avoid the introduction and establishment of additional non-native species in new ecosystems.
In the present study, as part of an ongoing investigation on the diversity of terrestrial mollusks in Chile (Araya, 2016; Araya & Aliaga, 2015; Miquel & Araya, 2013, 2015), we report the range extension of Vallonia pulchella (Müller, 1774) and the first record of Vallonia cf. excentrica Sterki, 1893 in continental Chile. The Global Biodiversity Information Facility (GBIF, 2024) database had previously reported V. pulchella in Easter Island, as well as in Caldera and Coquimbo in continental Chile (unpublished data). Vallonia pulchella, the lovely Vallonia, is Holarctic in origin (Hotopp et al., 2013) and has been documented as an introduced species in South Africa, Madagascar, Australia, New Zealand and several other Asian countries (Gerber, 1996; Herbert, 2010; Mitra et al., 2005; Robinson, 1999; Roll et al., 2009; Stanisic, 1998), as well as in Argentina, Brazil, Peru, and Uruguay in South America (Rumi et al., 2010; Virgillito & Miquel, 2013). In Chile, the species was first reported as introduced by Philippi (1885) from the area surrounding Santiago. Later, Lataste (1896, fide Porter, 1926) found this species in Linderos, south of Santiago. More recently, it was reported by PNUD (2014), although the specific location was not detailed. On the other hand, V. cf. excentrica, another Holartic species (Metcalf, 1984; Nekola, 2002), is reported here for the first time as an introduced species in the country and in South America.
Materials and methods
Snail specimens were hand-collected from wet soil in 2017 in 2 urban areas of continental Chile: Caldera in the north and La Florida in Santiago City, in the central region of the country. Snails were sorted out from 2 soil samples of about 500 g in each location. Adult specimens were photographed using a Motic SMZ-168 stereomicroscope equipped with a Moticam 2000 digital camera and measured with a millimeter ruler. Specimens were also examined using a Hitachi SU3500 scanning electron microscope (SEM). Prior to observation, the snails were immersed in a dilute sodium hypochlorite solution for 3 minutes, rinsed in distilled water, and air-dried.
Species identification was based on original descriptions and additional literature (Burch & Jung, 1988; Herbert, 2010; Sterki, 1893). To assess morphometric variation among introduced populations, we measured 6 shell variables: shell length (SL), shell width (SW), shell height (SH), aperture height (AH), aperture width (AW), and the distance from the umbilicus to the last body whorl (UBW). The data were log-transformed to assess normality and homoscedasticity. Since some variables did not meet these assumptions, we applied the nonparametric Mann-Whitney U test (M-W U) using PAST software V. 4.13 (Hammer et al., 2001). Additionally, we performed a principal component analysis (PCA) in PAST to investigate whether shell shape variation could distinguish between the 2 morphologically similar species in morphometric space. This analysis included 14 individuals from Caldera and 13 from La Florida assigned to V. pulchella, and 3 individuals assigned to V. cf. excentrica. Specimens from Caldera and La Florida smaller than 2 mm were excluded from the analysis. Voucher specimens were deposited in the Museo de Ciencias Naturales Profesor Pedro Ramírez Fuentes (MCNPPRF), Chillán, Chile, and the Laboratorio de Malacología y Sistemática Molecular, Universidad del Bío-Bío, Chillán, Chile (LMSM) (see Results).
Results
Stereomicroscope and SEM observations revealed the presence of 2 non-native species of the genus Vallonia in continental Chile (Figs. 1-4).
Phylum Mollusca Linnaeus, 1758
Class Gastropoda Cuvier, 1797
Superfamily Pupilloidea W. Turton, 1831
Family Valloniidae E. S. Morse, 1864
Genus Vallonia Risso, 1826
Valloniapulchella (Müller, 1774)
Figs. 1A-K, 3A
Material examined. 17 specimens from Caldera (27°04’ S, 70°49’ W) (LMSM 1-17), 18 specimens from La Florida (33°27’ S, 70°40’ W) (MCNPPRF 139–24, MCNPPRF 139-25, LMSM 3-18).
Morphology. Shell with 3 1/8 whorls, fine axial striae (Fig. 1A, D, I), depressed, umbilicated, gray-white or corneous, translucent with a matte gloss. Adult shell diameter: 1.6-2.6 mm (Caldera), 1.3-2.4 mm (La Florida). Protoconch smooth (620 µm), with 1 1/8 whorls and a clear transition to the teleoconch (Fig. 3A). Round umbilicus (Fig. 1C, F, K), about 1/4 of the shell diameter. Near-circular aperture, slightly oblique (Fig. 1B, E, G, H, J), translucent peristome with a thickened lip in adults.
Remarks
Vallonia pulchella resembles Vallonia excentrica. Some authors consider V. excentrica to be a form of V. pulchella (Adams, 1906; Hubendick, 1950). Korte and Armbruster (2003) suggest that V. excentrica is paraphyletic, within the same clade as V. pulchella. However, V. pulchella can be distinguished by a higher spire, a less elongated umbilicus, and a translucent, thickened lip (in contrast, the lip of V. excentrica is white) (Sterki, 1893).
Vallonia cf. excentrica Sterki, 1893
Figs. 2A-F, 3B
Material examined. 3 specimens from La Florida (33°27’ S, 70°40’ W) (LMSM 1-3).
Morphology. Shell with about 3 1/8 whorls, fine axial striae (Fig. 2A-F), highly depressed, widely umbilicated, whitish or corneous, translucent. Adult size: 1.8-2.1 mm. Protoconch smooth (810 µm), 1 1/4 whorls (Fig. 3B), with clear transition to teleoconch. Round umbilicus (Fig. 2C, F), ~ 1/4 shell diameter. Subcircular to crescentic aperture, slightly oblique (Fig. 2B, E, J), thin lip.
Remarks
Valloniaexcentrica is distinguished from V. pulchella by its larger protoconch (Fig. 3), lower spire, and sharp, smooth aperture lip (thickened in V. pulchella). The crescentic aperture and the absence of radial sculpture of ribs distinguishes this species from all other Vallonia species.
No statistical differences were found between V. pulchella populations from Caldera and La Florida in any shell variable (M-W U test: p > 0.05) (Table 1). The first 2 principal components (PCAs) explained 90.02% of the variation (PC1: 84.7%, PC2: 5.3%). Populations of V. pulchella exhibited substantial overlap in morphometric space but were clearly separated from V. cf. excentrica specimens (Fig. 4).
Discussion
In the present study, we report a range extension of V. pulchella and the first documented occurrence of V. cf. excentrica in Chile and South America. The statistical analyses confirmed our typological assignment of snails from Caldera and La Florida to V. pulchella. There were no significant differences in any shell variables between the 2 samples, and in morphometric space, individuals from both localities clustered as a single group. Moreover, the analysis revealed that these specimens are clearly separated from those assigned to V. cf. excentrica, supporting their distinction as separate species.
Previous records of V. pulchella from Chile refer to preserved specimens from Bosque de Ceibos, Anakena, housed in the Field Museum of Natural History (FMNH 312411 and 312412), Arlington, Virginia (unpublished data). In addition, samples from Coquimbo were identified by Bernhard Hausdorf (iNaturalist 42089250), and those from Caldera are preserved in the Museu de Ciències Naturals de Barcelona (MCNB-Malac MZB 89-0910) (GBIF, 2024).
Although Vallonia species have low vagility, they can be dispersed by birds, fish, water and human activities (Altaba, 2015; Boycott, 1934), in the latter case through the unintentional transport of plants, soil, or leaf litter (Schembri & Lanfranco, 1996). However, the means of introduction of Vallonia species to Chile remains unknown, as does the origin of the samples from Easter Island, Caldera, and Coquimbo. Their introduction into the country is most likely associated with human activity, given that both Caldera and Coquimbo are very active ports and Easter Island has a high influx of tourism. Due to the species’ wide distribution in Chile, a single introduction event seems unlikely.
Figura 1. Vallonia pulchella collected in Caldera, northern Chile. A-C, Shells of different specimens imaged using SEM: A) apical view; B) apertural view; C) umbilical view. D-K, Shells photographed using stereomicroscope; D-G, specimen presented in A photographed in different views: D) apical view; E) apertural view; F) umbilical view; G) lateral view. H-K, Specimen presented in C photographed in different views: H) lateral view; I) apical view; J) apertural view; K) umbilical view. Scale bars: 1 mm.
Although we did not estimate the abundance of V. pulchella at the sampled sites, previous studies have reported densities ranging from fewer than 10 individuals/m² (Čejka & Hamerlik, 2009; Koralewska-Batura & Błoszyk, 2007) to up to 588 individuals/m² (Hermida et al., 1993). While we recorded only 3 individuals of V. cf. excentrica, this would represent the first record of the species as introduced in South America. However, this species has already been introduced to South Africa, Australia, and a great part of New Zealand (Barker, 1999).
Figura 2. Vallonia cf. excentrica collected in La Florida, Chile. A-C, Shell of a specimen imaged using SEM in different views: A) apical view; B) apertural view; C) umbilical view. D-F, Shell of a second individual photographed using stereomicroscope: D) apical view; E) apertural view; F) umbilical view. Scale bars: 1 mm.
Vallonia species are similar in size and shape (Gerber & Bössneck, 2009; Sterki, 1893). However, although V. pulchella and V. cf. excentrica are morphologically similar, they differ in the shape of the peristome lip, reflected and thickened in V. pulchella, but thin in the present V.excentrica specimens, and in protoconch size, which is larger in the latter species.
As an invasive species, V. pulchella could threaten livestock, particularly sheep and rabbits, as an intermediate host for nematodes such as Protostrongylus boughtoni (Goble & Dougherty, 1943), P. hobmaieri (Schulz et al., 1933), P. rufescens (Leuckart, 1865), and Varestrongylus pneumonicus (Bhalerao, 1932) (Grewal et al., 2003; Samson & Holmes, 1985). Continuous monitoring is essential to assess its spread. Due to its small size, adults, juveniles, and eggs, V. pulchella can be easily transported in soil and potted plants. Its iteroparous mode of reproduction and ability to self-fertilize could further contribute to its dispersal (Kuźnik-Kowalska & Proćków, 2016; Whitney, 1941). Given that most specimens were collected from plant litter and pots, the nursery trade could act as a dispersal vector, as seen with other small invasive snails (Forsyth, 2015).
Figura 3. Protoconch of Vallonia pulchella (A) and Vallonia cf. excentrica (B) imaged using SEM. Scale bar: 500 μm.
The small size of Vallonia species may explain the limited number of records, as small non-native snails are often recognized only long after they have arrived (Hutchinson et al., 2014). The new records presented in this study extend the known distribution of V. pulchella in South America and provide the first documented record of V. cf. excentrica in Chile and South America. However, these species are probably present elsewhere in the country, especially in the central region, where human population density is higher.
Table 1
Mann-Whitney U test results between Vallonia pulchella specimens collected in Caldera (northern Chile) and La Florida (central Chile). Shell length (SL), shell height (SH), shell width (SW), aperture height (AH), aperture width (AW), body whorl length (BWL), and the distance from the umbilicus to the last body whorl (UBW). Standard deviation: S.D.
Shell variable
Caldera (n)
Mean (±S.D.)
La Florida (n)
Mean (±S.D.)
U
p-value
SL
14
2.3 (± 0.2)
13
2.2 (± 0.1)
72.0
0.3549
SH
14
1.8 (± 0.1)
13
1.8 (± 0.1)
87.5
0.8736
SW
14
1.1 (± 0.1)
13
1.0 (± 0.1)
79.0
0.5483
AH
14
1.0 (± 0.1)
13
1.0 (± 0.1)
79.0
0.5358
AW
14
1.0 (± 0.1)
13
1.0 (± 0.1)
75
0.4097
UBW
14
1.3 (± 0.1)
13
1.2 (± 0.1)
73
0.3453
Figura 4. Principal component analysis (PCA) of 6 shell morphometric variables (SL, SH, SW, AH, AW, and UBW) for Vallonia species collected in Caldera (northern Chile) and La Florida (central Chile). Percent variance explained by PC1 and PC2 is shown on the axes.
It is also important to note that, although V. pulchella and V. cf.excentrica have been considered conspecific by some authors (e.g., Sterki, 1893), our study demonstrates that they are distinct species, based on clear morphological differences, particularly in the form of the lip and protoconch. Nevertheless, a comprehensive and integrative taxonomic revision of both species is still needed to fully resolve their relationship.
Acknowledgments
We thank Cristian Suárez for assistance with the SEM observations. We also thank FONDEQUIP Program (No. EQM-140088) and the anonymous reviewers. JFA also acknowledges the ANID doctoral fellowship 21180344, which supported the first 2 years of his doctoral studies.
References
Adams, L. E. (1906). Vallonia costata + excentrica. Journal of Conchology, 11, 364–365.
Alonso, Á., Collado, G. A., Gérard, C., Levri, E. P., Salvador, R. B., & Castro-Díez, P. (2025). Effects of the invasive aquatic snail Potamopyrgus antipodarum (Gray, 1853) on ecosystem properties and services. Hydrobiologia, 852, 1339–1357. https://doi.org/10.1007/s10750-022-05116-z
Altaba, C. R. (2015). Once a land of big wild rivers: specialism is context-dependent for riparian snails (Pulmonata: Valloniidae) in central Europe. Biological Journal of the Linnean Society, 115, 826–841. https://doi.org/10.1111/bij.12546
Araya, J. F. (2015). Current status of the non-non-native molluscs in Chile, with the first record of Otalapunctata (Müller, 1774) (Gastropoda, Helicidae) in the country and new records for Cornuaspersum (Müller, 1774) and Deroceraslaeve (Müller, 1774). Journal of Natural History, 49, 1731–1761. https://doi.org/10.1080/00222933.2015.1006703
Araya, J. F. (2016). On some land snails (Mollusca: Gastropoda) of Los Molles, central Chile. Revista Mexicana de Biodiversidad, 87, 1365–1368. https://doi.org/10.1016/j.rmb.2016.10.002
Araya, J. F., & Aliaga, J. A. (2015). A new species of Lilloiconcha Weyrauch,1965 (Pulmonata: Charopidae) from central Chile. Zootaxa, 4007, 295–297. https://doi.org/10.11646/zootaxa.4007.2.13
Barker, G. M. (1999). Naturalised terrestrial Stylommato-phora (Mollusca: Gastropoda). Fauna of New Zealand, 38, 1–253. https://doi.org/10.7931/J2/FNZ.38
Boycott, A. E. (1934). The habitats of land Mollusca in Britain. Journal of Ecology, 22, 1–38. https://doi.org/10.2307/2256094
Burch, J. B., & Jung, Y. (1988). Land snails of the University of Michigan Biological Station area. Walkerana, 3, 1–177.
Cádiz, F. J., & Gallardo, C. S. (2007). Arionintermedius (Gastropoda: Stylommatophora); first record of this introduced slug in Chile, with notes on its anatomy and natural history. Revista Chilena de Historia Natural, 80, 99–108.
Cádiz, F. J., Cádiz, D. G., & Grau, J. H. (2013). An invasive predatory snail Oxychilusalliarius (Miller, 1822) (Stylommatophora: Zonitidae) threatens the native malacofauna of continental Chile: a morphological and molecular confirmation. Studies on Neotropical Fauna and Environment,48, 119–124. https://doi.org/10.1080/01650521.2013.843910
Čejka, T., & Hamerlik, L. (2009). Land snails as indicators of soil humidity in Danubian woodland (SW Slovakia). Polish Journal of Ecology, 57, 741–747.
Collado, G. A. (2014). Out of New Zealand: molecular identification of the highly invasive freshwater mollusk Potamopyrgusantipodarum (Gray, 1843) in South America. Zoological Studies, 53, 70. https://doi.org/10.1186/s40555-014-0070-y
Collado, G. A. (2017). Unraveling cryptic invasion of a fresh-water snail in Chile based on molecular and morphological data. Biodiversity and Conservation,26, 567–578. https://doi.org/10.1007/s10531-016-1255-y
Collado, G. A., Cristina, D., Darrigran, G., & Alonso Á. (2025). Attributes and effects of the invasive air-breathing freshwater snail Physa acuta on ecosystem properties and services. Marine and Freshwater Research, 76, MF25023. https://doi.org/10.1071/MF25023
Darrigran, G., Agudo-Padrón, I., Baez, P., Belz, C., Cardoso, F., Carranza, A. et al. (2020). Non-native mollusks throughout South America: emergent patterns in an understudied continent. Biological Invasions, 22, 853–871. https://doi.org/10.1007/s10530-019-02178-4
Forsyth, R. G. (2015). First record of Carychiumminimum Müller, 1774 in New Brunswick, Canada (Mollusca: Gastropoda: Ellobioidea). Check List, 11, 1511. https://doi.org/10.15560/11.1.1511
Gerber, J. (1996). Revision der Gattung Vallonia Risso, 1826 (Mollusca: Gastropoda: Valloniidae). Schriften zur Malakozoologie aus dem Haus der Natur-Cismar, 8, 1–227.
Gerber, J., & Bössneck, U. (2009). The genus Vallonia in Nepal: (Gastropoda: Pulmonata: Valloniidae). Archiv für Molluskenkunde, 138, 43–52. https://doi.org/10.1127/arch.moll/0003-9284/138/043-052
GBIF (2024). Global Biodiversity Information Facility. Retrieved November 10, 2024 from: https://www.gbif.org/es/species/2296010
Grewal, P. S., Grewal, S. K., Tan, L., & Adams, B. J. (2003). Parasitism of molluscs by nematodes: types of associations and evolutionary trends. Journal of Nematology, 35, 146–156.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4, 9.
Herbert, D. G. (2010). The introduced terrestrial Mollusca of South Africa. SANBI Biodiversity Series 15. Pretoria: South African National Biodiversity Institute.
Hermida, J., Outeiro, A., & Rodríguez, T. (1993). Aspectos ecológicos de Vallonia pulchella (Muller, 1774) (Gastropoda, Pulmonata). Nova Acta Científica Compostelana (Bioloxía), 4, 129–135.
Hotopp, K. P., Pearce, T. A., Nekola, J. C., Slapcinsky, J., Dourson, D. C., Winslow, M. et al. (2013). Land snails and slugs of the Mid-Atlantic and Northeastern United States. Pittsburgh, PA: Carnegie Museum of Natural History. Retrieved November 10, 2024 from: https://www.carnegiemnh.org/science/mollusks/index.html
Hubendick, B. (1950). The validity of Vallonia excentrica Sterki. Proceedings of the Malacological Society of London, 28, 75–78. https://doi.org/10.1093/oxfordjournals.mollus.a064569
Hutchinson, J., Reise, H., & Robinson, D. (2014). A biography of an invasive terrestrial slug: the spread, distribution and habitat of Derocerasinvadens. NeoBiota, 23, 17–64. https://doi.org/10.3897/neobiota.23.7745
Kirch, P., Christensen, C., & Steadman, D. (2009). Subfossil land snails from Easter Island, including Hotumatuaanakenana, new genus and species (Pulmonata: Achatinellidae). Pacific Science,63, 105–122. https://doi.org/10.2984/15346188(2009)63[105:SLSFEI]2.0.CO;2
Koralewska-Batura, E., & Błoszyk, J. (2007). Stability of malacocoenoses in an ash-elm forest based on two-year observations. Folia Malacologica, 15, 175–180. https://doi.org/10.12657/folmal.015.015
Korte, A., & Armbruster, G. F. J. (2003). Apomorphic and plesiomorphic ITS-1 rDNA patterns in morphologically similar snails (Stylommatophora: Vallonia), with estimates of divergence time. Journal of Zoology, 260, 275–283. https://doi.org/10.1017/S0952836903003741
Kuźnik-Kowalska, E., & Proćków, M. (2016). Reproductive biology and growth of two Vallonia species in laboratory conditions (Gastropoda: Eupulmonata: Valloniidae). Folia Malacologica, 24, 265–273. https://doi.org/10.12657/folmal.024.022
Lataste, F. (1896). L’ Helix pulchella dans les vignes de Linderos. Santiago: Actes de la Societé Scientifique du Chili. Tome VI.
Letelier, S., Ramos, A., & Huaquín, L. (2007). Moluscos dulceacuícolas exóticos en Chile. Revista Mexicana de Biodiversidad, 78, 9–13. https://doi.org/10.22201/ib.20078706e.2007.002.301
Lowe, S., Browne, M. & Boudlejas, S. (2000). 100 of the world’s worst invasive alien species. A selection from the Global Invasive Species Database. The Invasive Species Specialist Group (ISSG), a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN), Auckland, New Zealand.
Metcalf, A. L. (1984). Distribution of land snails of the San Andres and Organ Mountains, Southern New Mexico. The Southwestern Naturalist, 29, 35–44. https://doi.org/10.2307/3670767
Miquel, S. E., & Araya, J. F. (2013). A new Charopidae from Chile and Argentina, Stephacharopacalderaensis n. gen. and n. sp., with remarks on the taxonomy of the genus Stephadiscus Hylton Scott 1981 (Mollusca: Gastropoda Pulmonata). Archiv für Molluskenkunde, 142, 227–235. https://doi.org/10.1127/arch.moll/1869-0963/142/227-235
Miquel, S. E., & Araya, J. F. (2015). New records of terrestrial molluscs of the Juan Fernández Archipelago (Chile), with the description of a new genus and species of Charopidae (Gastropoda Stylommatophora). Archiv für Molluskenkunde, 144, 155–167. https://doi.org/10.1127/arch.moll/1869-0963/144/155-167
Mitra, S. C., Dey, A., & Ramakrishna. (2005). Pictorial handbook. Indian land snails. Kolkata: Zoological Survey of India.
Nekola, J. C. (2002). Large-scale terrestrial gastropod community composition patterns in the Great Lakes region of North America. Diversity and Distributions, 9, 55–71. https://doi.org/10.1046/j.1472-4642.2003.00165.x
Philippi, R. A. (1885). Zoolojía. Sobre los animales introducidos en Chile desde su conquista por los españoles. Anales de la Universidad de Chile, 67, 319–335.
PNUD (Programa de las Naciones Unidas para el Desarrollo). (2014). Moluscos invasores de Chile. Documento del Taller Moluscos exóticos invasores en Chile: efectos en la biodiversidad. Santiago de Chile: Programa de las Naciones Unidas para el Desarrollo.
Porter, C. (1926). Los estudios malacológicos en Chile. Revista Chilena de Historia Natural, 30, 26–39.
Robinson, D. G. (1999). Alien invasions: the effects of the global economy on non-marine gastropod introductions into the United States. Malacologia, 41, 413–438.
Roll, U., Dayan, T., Simberloff, D., & Mienis, H. K. (2009). Non-indigenous land and freshwater gastropods in Israel. Biological Invasions, 11, 1963–1972. https://doi.org/10.1007/s10530-008-9373-4
Rumi, A., Sánchez, J., & Ferrando, N. S. (2010). Thebapisana (Müller, 1774) (Gastropoda, Helicidae) and other alien land molluscs species in Argentina. Biological Invasions, 12, 2985–2990. https://doi.org/10.1007/s10530-010-9715-x
Samson, J., & Holmes, J. C. (1985). The effect of temperature on rates of development of larval Protostrongylus spp. (Nematoda: Metastrongyloidea) from bighorn sheep, Oviscanadensiscanadensis, in the snail Valloniapulchella. Canadian Journal of Zoology, 63, 1445–1448. https://doi.org/10.1139/z85-215
Schembri, P. J., & Lanfranco, E. (1996). Introduced species in the Maltese Islands. In A. E. Baldacchino, & A. Pizzuto (Eds.), Introduction of alien species of flora and fauna. Proceedings of a seminar held at Qawra, Malta (pp. 29–54). Floriana, Malta: Environment Protection Department.
Stanisic, J. (1998). Family Valloniidae. In P. L. Beesley, G. J. B. Ross, & A. Wells (Eds.), Mollusca: the southern synthesis. Part B. Fauna of Australia, Vol. 5 (pp. 1085–1086). Melbourne: CSIRO Publishing.
Sterki, V. (1893). Observations on Vallonia. Proceedings of the Academy of Natural Sciences of Philadelphia, 45, 234–279.
Sylvester, F., & Sardiña, P. (2015). Relationships of Limnoperna fortunei with benthic animals. In D. Boltovskoy (Ed.), Limnoperna fortunei: the ecology, distribution and control of a swiftly spreading invasive fouling mussel (pp. 191–210). Cham: Springer International Publishing.
Valdovinos-Zarges, C. (1999). Biodiversidad de moluscos chilenos: base de datos taxonómica y distribucional. Gayana, 63, 111–164.
Vaughn, C. C. (2018). Ecosystem services provided by fresh-water mussels. Hydrobiologia, 810, 15–27. https://doi.org/10.1007/s10750-017-3139-x
Virgillito, M., & Miquel, S. E. (2013). New records of exotic land snails and slugs in Argentina. Revista del Museo Argentino de Ciencias Naturales, Nueva Serie, 15, 295–303.
Whitney, M. E. (1941). The hermaphrodite gland and germ cells of Valloniapulchella Müll. Michigan Academy of Science, 26, 311–338.
Epicoccum zapotae: una especie nueva aislada de frutos de Manilkara zapota en el centro de México
Limni Silday Ramírez-Gallegos a, Laura Navarro-de la Fuente b, Ángel Trigos b, Irene Lagunes b, *
a Universidad Veracruzana, Centro de Investigación en Micología Aplicada, Doctorado en Micología aplicada, Calle Médicos 5, U.H. Del Bosque, 91010 Xalapa, Veracruz, Mexico
b Universidad Veracruzana, Centro de Investigación en Micología Aplicada, Calle Médicos 5, U.H. Del Bosque, 91010 Xalapa, Veracruz, Mexico
Received: 07 October 2024; accepted: 03 November 2025
Abstract
The genus Epicoccum exhibits substantial intraspecific morphological and genetic diversity, which has made it difficult to correctly identify and delimit species on the basis of morphological characteristics alone. Against this background, this study was aimed to report the morphological and molecular identification of a new species belonging to the genus Epicoccum, isolated from sapodilla fruit (Manilkara zapota) in Veracruz, Mexico, named E. zapotae. The morphological characteristics and the phylogenetic analysis of the concatenated sequences of LSU, ITS, and β-TUB support the fact that the isolate is a new species of the genus Epicoccum. This discovery contributes to the knowledge of the diversity of Epicoccum species in tropical ecosystems.
Keywords: Didymellaceae; Epicoccum;Morphology; Phylogenetic; Sapodilla fruit
Resumen
El género Epicoccum presenta diversidad morfológica y genética intraespecífica considerable, lo cual ha dificultado la correcta identificación y delimitación de especies con base únicamente en características morfológicas. En este contexto, el objetivo de este estudio fue reportar la identificación morfológica y molecular de una especie nueva perteneciente al género Epicoccum, aislada del fruto chicozapote (Manilkara zapota) en Veracruz, México, denominada Epicoccum zapotae. Las características morfológicas y el análisis filogenético de las secuencias concatenadas de LSU, ITS y β-TUB respaldan el hecho de que el aislado es una nueva especie del género Epicoccum. Este descubrimiento contribuye al conocimiento de la diversidad de especies de Epicoccum en ecosistemas tropicales.
Palabras clave: Didymellaceae; Epicoccum;Morfología; Filogenética; Fruto del chicozapote
Introduction
The genus Epicoccum belongs to the Didymellaceae family. It exhibits substantial intraspecific morphological and genetic diversity. It can be found in the air, soil, water, and various parts of plants including flowers, branches, leaves, and bark (Chen et al., 2017; Fávaro et al., 2011; Lee et al., 2020; Li et al., 2022). More than 70 species of the genus Epicoccum have been described, of which 5 species (E. dendrobii, E. layuense, E. mezzettii, E. minitans, and E. nigrum) have demonstrated biocontrol activity against phytopathogens (Braga et al., 2018; Taguiam et al., 2021). Species of this genus may exhibit a saprophytic (Braga et al., 2018), phytopathogenic (E. nigrum and E. sorghinum) (Chen et al., 2017; Taguiam et al., 2021) or endophytic lifestyle, the latter having isolated compounds with antioxidant, anticancer and antimicrobial activities (Braga et al., 2018; Taguiam et al., 2021).
The genus Epicoccum was originally established by Link (1816) based on observations of sporodochia in dry plant stems, with the following diagnosis: compact globose stroma dotted with subglobose spores. Link (1816) described the type species E. nigrum, however, his diagnosis was unsatisfactory and generated confusion (Schol-Schwarz, 1959). Subsequently, many species belonging to this genus were described, based on the fungus characteristics in its natural habitat, and therefore, they were named of the plant species from which the specimen was isolated (Schol-Schwarz, 1959).
Schol-Schwarz (1959) conducted a thorough review of the Epicoccum genus, examining 70 specimens displaying a variety of hues, including red, yellow, or olive, alongside an additional 96 specimens from herbaria around the world. Despite this extensive examination, an incomplete diagnosis was noted with descriptions that were primarily based on the fungus’s original habitat characteristics and lacking the spore size measurements. For this reason, Schol-Schwarz (1959) considered the genus misclassified, concluding that color could not be used as a reliable taxonomic character, due to its susceptibility to various abiotic factors such as medium, light, pH, etc.; thus, no valid basis remained for splitting the genus into separate species, despite its wide range of conidial dimensions and cultural characters. Consequently, the entire genus was reduced to the single variable species E. nigrum (Kilpatrick & Chilvers, 1981).
Noting the lack of molecular evidence to support Schol-Schwarz´s classification, Wang and Guo (2004) performed the molecular identification of 45 E. nigrum isolates by amplifying the 5.8S gene and flanking internal transcribed spacer regions (ITS1 and ITS2) of the nuclear ribosomal DNA. They found that E. nigrum could comprise more than 1 species, ultimately concluding that conidial size and colony color should not be used as reliable taxonomic characteristics in the identification of E. nigrum. Later, Fávaro et al. (2011) acknowledged the need to reevaluate the classification of E. nigrum as a single variable species and many sequences deposited as E. nigrum in the GenBank database and fungal strains cultures many of which should be reclassified. Accurate species identification has long been challenging due to its heavy reliance on host plant morphology and association (Chen et al., 2015).
Therefore, taxonomic studies based on multilocus phylogeny using LSU, ITS, rpb2 and tub2 sequences in combination with morphological differences, allowed revising the classification within the Didymellaceae family, including the genus Epicoccum, providing a relatively robust phylogenetic basis for taxon delineation (Aveskamp et al., 2010; Braga et al.,2018; Chen et al., 2015; Chen et al., 2017). Currently, 18 Epicoccum species are accepted with their correct morphological and molecular identification (Braga et al., 2018). In this context, this study was aimed to report the morphological and molecular identification of a new species belonging to the genus Epicoccum.
Materials and methods
Sapodilla fruits were collected during May-June 2017 from 10-12 tall Manilkarazapota trees in one ~ 1 ha orchard close to the village of Apazapan, Veracruz, Mexico. Fruit samples were transported to the laboratory at the Centro de Investigación en Micología Aplicada (CIMA), Universidad Veracruzana. The samples were washed with running tap water to remove residual soil, afterwards the fruits were cut into small pieces (0.5 cm2), disinfected with 2% sodium hypochlorite for 30 seconds, and washed twice with sterile water. The disinfected samples were transferred onto Petri dishes containing Potato Dextrose Agar (PDA) medium (MCD LAB, San Jacinto Amilpas, Oax, MX) with 0.2 g/L chloramphenicol. The Petri dishes were incubated in dark conditions at 25 ± 2 °C for 2-3 days until mycelial growth was developed from the samples. Newly developed mycelia were immediately transferred to fresh PDA plates and incubated under the same conditions for fungal isolation. To ensure the purity of the fungal isolate, monosporic cultures were developed by cutting hyphal tips under a stereoscopic microscope (Leica EZ4), transferred to PDA plates and incubated under the same conditions established.
Culture characteristics were determined after 14 days of cultivation at 25 ± 2 °C on PDA. Colony diameters were measured after 7 days in incubation. The hyphae and reproductive structures produced on PDA were morphologically identified using an optical microscope at 100 × magnification. The mycelium was mounted on a microscope slide with a drop of lactophenol blue solution; the diameter of the conidia was measured (n = 100), means and standard errors (SE) were calculated, with extreme values shown in parentheses. The reproductive structures were identified using the taxonomic keys reported by Barnett and Hunter (1972). The holotype specimen (CM-CNRG 1003, dried agar plate culture) was deposited in the Microorganism Collection of the National Institute of Forestry, Agricultural and Livestock Research (INIFAP) in Mexico.
Genomic DNA was extracted from 14 days old mycelium grown on PDA according to Liu et al. (2000). The internal transcribed spacer region (ITS) of the nuclear ribosomal DNA (nrDNA) gene, the partial large subunit (LSU) nrDNA gene, and the β-tubulin (TUB) gene were amplified. PCR was performed using the primers listed in Table 1. DNA was amplified in a SureCycler 8800 thermal cycler with the same conditions for each gene, only the annealing temperature varied. The reaction mixture was incubated as follows: initial denaturation at 94 °C for 3 min, followed by 34 cycles at 94 °C for 1 min, the annealing temperatures are shown in Table 1, extension at 72 °C for 1 min, followed by a final extension at 72 °C for 10 min. The amplifications were purified using the Wizard® SV Gel and PCR System Clean-Up kit and sent to Labsergen Langebio (Cinvestav, Irapuato, Gto., Mexico) for sequencing on an AB3770 capillary sequencer.
Consensus sequences were created with BioEdit software (Hall, 1999) and were compared in the GenBank nucleotide sequence database (Benson et al., 2017) using BLAST search software (Zhang et al., 2000) to confirm the genus and percentage of identity. Closely related species sequences and our newly obtained sequences (Table 2) were incorporated into sequence datasets independently for each molecular marker using PhyDE v.0.9971 Phylogenetic Development (Müller et al.,2010). Each dataset was independently aligned using the MAFFT online service (Katoh et al., 2019). Inconsistencies were manually adjusted using the MESQUITE 3.61 phylogenetic data editor (Maddison & Maddison, 2019), the same program with which the concatenated ITS + LSU + TUB sequences dataset were integrated. The GTR + G nucleotide substitution model was selected using the jModelTest v.2.1.4 program (Darriba et al., 2012) according to the Akaike Information Criterion (AIC). The concatenated aligned dataset was analyzed using Maximum Likelihood (ML) and Bayesian Inference (BI). The ML analyses were conducted with RAxMLGUI2.0 (Edler et al., 2020) using 1,000 bootstrap replicates, the robustness of the analyses was evaluated by bootstrap support (BS) values. Bayesian Inference analyses were performed on MrBayes v.3.2.1 software, using the substitution model selected for each matrix by jModelTest, based on the Akaike Information Criterion (AIC) (Table 3), with 4 Markov Chains Monte Carlo (MCMC) and 1 million generations (Ronquist et al., 2012). The ITS, LSU and TUB sequences generated in this study were deposited in the GenBank database under the accession numbers provided in the taxonomic section.
The Genealogical Concordance Phylogenetic Species Recognition (GCPSR) model (Taylor et al., 2000) was used to delimit the fungal species and analyze phylogenetic relationships among closely related taxa. Recombination levels within these phylogenetically proximate species were assessed through the calculation of the pairwise homoplasy index (PHI) using the SplitsTree4 software (Huson, 1998; Huson & Bryant, 2006). A concatenated dataset including ITS, LSU, and TUB was used for the analyses. Splits graphs were generated in SplitsTree4 utilizing both the Log-Det transformation and splits decomposition options, facilitating the visualization of the phylogenetic relationships among the species. Interpretation of a pairwise homoplasy index value falling below the 0.05 significance threshold (Φw < 0.05) was taken to signify the presence of recombination within the analyzed dataset.
Table 1
Primers and annealing temperatures used in the PCR amplifications step.
Locus
Primer
Primer DNA sequences (5´–3´)
Annealing T (°C) / Time (s)
Reference
ITS
ITS1F
CTTGGTCATTTAGAGGAAGTAA
53/45
Gardes & Bruns, 1993; White et al., 1990
ITS4
TCCTCCGCTTATTGATATGC
LSU
LR0R
ACCCGCTGAACTTAAGC
55/60
Vilgalys & Hester, 1990
LR7
TACTACCACCAAGATCT
TUB
TUB1
AACATGCGTGAGATTGTAAGT
57/60
Woudenberg et al., 2009
TUB22
TCTGGATGTTGTTGGGAATCC
Table 2
Species and GenBank accession number of sequences used for the construction of the phylogenetic tree.
Species
Voucher
Country
GenBank accession number
ITS
LSU
TUB
E. brahmansense
CBS:990.95
Papua New Guinea
MN973514
MN943720
MT005614
E. brahmansense
CBS 985.95
Papua New Guinea
MN973513
MN943719
MT005613
E. camelliae
LC:4862
China
KY742092
KY742246
KY742334
E. dendrobii
LC:8145 T
China
KY742093
KY742247
KY742335
E. draconis
CBS 186.83
Rwanda
GU237795
GU238070
GU237607
E. duchesneae
CBS 218.81
India
MN972935
MN973322
MN983950
E. duchesneae
LC:5139 T
China
KY742095
KY742249
KY742337
E. hordei
LC:8149
Australia
KY742098
KY742252
KY742340
E. hordei
LC:8148 T
Australia
KY742097
KY742251
KY742339
E. huancayense
CBS:105.80
Peru
MH861244
MH873016
GU237615
E. huancayense
CBS 390.93
Peru
GU237857
GU238085
GU237616
E. layuense
LC:8155 T
China
KY742107
KY742261
KY742349
E. mezzettii
CBS 173.38
Italy
MN973496
MN943701
MT005596
E. mezzettii
CBS 238.59
No data
MN973494
MN943699
MT005594
E. multiceps
CBS:119734
Brazil
MN973512
MN943718
MT005612
E. nigrum
CBS 173.73
USA
FJ426996
GU237975
FJ427107
E. nigrum
CBS 125.82
Netherlands
FJ426995
GU237974
FJ427106
E. oryzae
CBS:174.34
Japan
MN973500
MN943705
MT005600
E. oryzae
CBS:173.34
Japan
MN973499
MN943704
MT005599
E. ovisporum
CBS 180.80 T
No data
NR158228
NG069492
FJ427174
E. plurivorum
CBS 558.81
New Zealand
GU237888
GU238132
GU237647
E. poaceicola
CBS:987.95
Papua New Guinea
MN972955
MN973343
MN983969
E. poae
LC:8161
USA
KY742114
KY742268
KY742356
E. poae
LC:8160 T
USA
KY742113
KY742267
KY742355
E. proteae
CBS:114179 T
South Africa
MH862956
MH874519
LT623230
E. tobaicum
JCK-CSHF10
South Korea
MW368670
MW368670
MW392085
E. variabile
CBS:119733
Brazil
MN973501
MN943706
MT005601
E. viticis
CGMCC 3.18344 T
China
NR158267
NG069447
KY742360
E. zapotae
CM-CNRG 1003
Mexico
OQ333010
OQ333009
OQ330858
D. americana
CBS 185.85
USA
FJ426972
GU237990
FJ427088
T Indicates type strains
Table 3
The nucleotide substitution model for each matrix obtained by jModelTest based on the Akaike Information Criterion.
Locus
Length
Substitution model
MrBayes parameters (Nst-rates)
ITS
455 pb
TIM2 + I + G
6-invgamma
LSU
765 pb
TrN + I + G
6-invgamma
TUB
330 pb
TIM3 + G
6-gamma
Description
Epicoccum zapotae L. Navarro & L. Ramírez sp. nov. (Fig. 1)
Diagnosis: dark sporodochia, conidia dark pigmented, globose or obovoid with radial separation and a verrucous outer surface. Compared to other species in the genus, colonies growth on PDA exhibits a distinctive pink pigmentation.
Sexual morph: not observed.
Asexual morph: Sporodochia aredark. Conidiophores are compact or loose, and light brown. Conidia dark pigmented, globose or obovoid with radial septation and a verrucous outer surface, with an average size 10.48 ± 1.64 µm (8.5-13.4 µm) (Fig. 2).
Culture characteristics: Colonies on PDA measured 33 ± 2 mm in diameter after 7 days, furrowed and circular with filamentous margin and flattened aerial mycelia. The central region of the colonies appears light pink, surrounded by concentric rings of intense pink coloration.
This strain was deposited at Colección de Microorganismos del Centro Nacional de Recursos Genéticos (CNRG), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Tepatitlán de Morelos, Jalisco, Mexico, under accession number CM-CNRG 1003.
Type: Mexico, Veracruz State: Apazapan (19°20’17.16” N, 96°43’54.12” W, 300 m), isolated from the fruit of M. zapota, June 2017, Holotype, CM-CNRG 1003.
Etymology: zapotae, referring to the host plant species (Manilkarazapota) from which it was isolated.
Remarks
Phylogenetic tree constructed from concatenated ITS + LSU + TUB sequences of 30 different species, with a final length of 1,550 characters including gaps. Sequences of Didymella americana CBS 185.85 were selected as outgroup (Table 2, Fig. 3). Branches of the phylogenetic tree are labeled with their respective bootstrap values (BS) and the Bayesian posterior probabilities (BPP). Our consensus phylogenetic tree generated from the ML analysis with BS/BPP values robustly supports (BS = 98 / BPP = 1) the assignment of our isolate as an independent species within the Epicoccum genus, positioned as a sister taxon to E.dendrobii (Fig. 3).
Application of the GCPSR analysis to our dataset yielded a PHI value of 0.5502 (Fig. 4). This outcome suggests that there is no substantial evidence of recombination occurring between E. zapotae and its related taxa. These results indicate that the E. zapotae isolate is distinctly differentiated from E.dendrobii and there is no indication of recombination events between them.
The robustness of BS/BPP values 98/1 supports the placement of E. zapotae as a sister species to E. dendrobii. This relationship forms a distinct clade, positioned adjacent to E. layuense, E. mezzettii, E. nigrum, E. oryzae, E. poae, and E. tobaicum.
Figure 1. E. zapotae (CM-CNRG 1003). A) Colony on PDA with the presence of sporodochia, B) conidiophore 100 ×, C) conidia 100 ×. Figure 2. Schematic representation of the microscopic characteristics of Epicoccum zapotae. A) Conidiophore, B) conidia.Figure 3. Maximum likelihood phylogenetic tree generated from the analysis of the concatenated ITS + LSU + TUB sequences of E. zapotae (holotype, CM-CNRG 1003) and species of the genus Epicoccum reported in the GenBank. The numbers at the nodes indicated Bootstrap values for 1,000 replicates (BS ≥ 70 %)/Bayesian Posterior Probabilities (BPP ≥ 0.9).Figure 4. Results of the pairwise homoplasy index (PHI) test for E. zapotae and closely related species. PHI test results (φw) > 0.05 indicate no significant evidence of recombination within the analyzed dataset.
Epicoccumdendrobii exhibits the closest genetic proximity to E. zapotae. Although E. zapotae shares general morphological traits with other Epicoccum species, distinct features support the classification of E. zapotae as a separate species. Conidia of E. dendrobii are multicellular-phragmosporous, verrucose, subglobose-pyriform, brown, with a basal cell, 11-19 µm diam (Chen et al., 2017). In contrast, conidia of E. zapotae are smaller, ranging from 8.84-12.12 µm diam. Additionally, the radial growth rate of E.dendrobii on PDA at 25 °C reaches 34-38 mm diam after 7 days. There is also a significant difference in culture characteristics on PDA between the 2 species, colonies of E.dendrobii display a regular margin, with flat aerial mycelia felty to floccose in texture, white to buff in color, becoming olivaceous near the center (Chen et al., 2017). In contrast, colonies of E. zapotae show slower growth on PDA (31-35 mm) for 7 days and their colonies are distinguished by their intense pink color (Table 4).
Table 4
Comparative morphological characteristics between E. dendrobii and E. zapotae. Data for E. dendrobii adapted from Chen et al. (2017).
Characteristic
E. dendrobii
E. zapotae
Conidia
Multicellular-phragmosporous, verrucose, subglobose-pyriform, brown
Dark pigmented, globose to obovoid with radial separation and a verrucous outer surface
Conidial diameter
11-19 µm
8.84-12.12 µm
Colony margin (PDA)
Regular
Furrowed and circular
Aerial mycelium (PDA)
Felty to floccose
Flattened
Colony color (PDA)
Beige
Pink
Colony diameter (7 days)
34-38 mm
31-35 mm
Epicoccum dendrobii has been reported to cause black leaf spots on Dendrobium fimbriatum (Chen et al., 2017). However, E. zapotae was isolated from sapodilla fruits without visible disease symptoms; therefore, it was probably plant-associated as an endophytic fungus, that can grow asymptomatically within plant tissues (Wen et al., 2022). The ability of endophytic fungi to produce bioactive compounds with biomedical applications has been widely explored (Hashem et al., 2023). In particular, from Epicoccum sp. known as an endophyte of Theobroma cacao, the bioactive compounds epicolactone, epicoccolide A and epicoccolide B have been isolated (Talontsi et al., 2013). Therefore, E. zapotae represents an opportunity for mycochemical studies aimed at exploring the biomedical potential of the bioactive compounds it produced.
Our phylogenetic and morphological analysis, as well as the results from the GCPSR analysis confirm that our isolate, despite its greater genetic proximity to E. dendrobii, presents distinct morphological differences ranging from the size and shape of the conidia to their growth on PDA. This demonstrates that our isolate represents a new species of the genus Epicoccum.
Our comprehensive morphological and molecular analyses have led us to conclude that fungal isolate recovered from the M. zapota fruit, is a new species within the genus Epicoccum which we report as E. zapotae. This discovery represents the first report of E. zapotae isolated from the sapodilla fruits in Veracruz, Mexico, and provides an opportunity for bioprospecting studies aimed at exploring its pharmaceutical potential.
Acknowledgements
This research was supported by Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti), formerly Conahcyt (FORDECYT PRONACES CF/304020 project) and the Universidad Veracruzana (UV-CA-354). L.S.R-G thanks Secihti for the predoctoral grant 802758.
References
Aveskamp, M. M., de Gruyter, J., Woudenberg, J. H. C., Verkley, G. J. M., & Crous, P. W. (2010). Highlights of the Didymellaceae: a polyphasic approach to characterize Phoma and related pleosporalean genera. Studies in Mycology, 65, 1–60. https://doi.org/10.3114/sim.2010.65.01
Barnett, H. L., & Hunter, B. B. (1972). Illustrated genera of imperfect fungi. 3rd Ed. Minneapolis: Burgess Publishing Company.
Benson, D. A., Cavanaugh, M., Clark, K., Karsch-Mizrachi, I., Lipman, D. J., Ostell, J. et al. (2017). GenBank. Nucleic Acids Research, 45, D37–D42. https://doi.org/10.1093/nar/gkw1070
Braga, R., Padilla, G., & Araújo, W. L. (2018). The biotechnological potential of Epicoccum spp.: diversity of secondary metabolites. Critical Reviews in Microbiology, 44, 759–778. https://doi.org/10.1080/1040841X.2018.1514364
Chen, Q., Jiang, J. R., Zhang, G. Z., Cai, L., & Crous, P. W. (2015). Resolving the Phoma enigma. Studies in Mycology, 82, 137–217. https://doi.org/10.1016/j.simyco.2015.10.003
Chen, Q., Hou, L. W., Duan, W. J., Crous, P. W., & Cai, L. (2017). Didymellaceae revisited. Studies in Mycology, 87, 105–159. https://doi.org/10.1016/j.simyco.2017.06.002
Darriba, D., Taboada, G. L., Doallo, R., & Posada, D. (2012). JModelTest 2: More models, new heuristics and parallel computing. Nature Methods, 9, 772. https://doi.org/10.1038/nmeth.2109
Edler, D., Klein, J., Antonelli, A., & Silvestro, D. (2020). raxmlGUI 2.0: a graphical interface and toolkit for phylogenetic analyses using RAxML. Methods in Ecology and Evolution, 12, 373–377. https://doi.org/10.1111/2041-210X.13512
Fávaro, L. C., de Melo, F. L., Aguilar-Vildoso, C. I., & Araújo, W. L. (2011). Polyphasic analysis of intraspecific diversity in Epicoccum nigrum warrants reclassification into separate species. Plos One, 6, e14828. https://doi.org/10.1371/journal.pone.0014828
Gardes, M., & Bruns, T. D. (1993). ITS primers with enhanced specificity for basidiomycetes-application to the identifi- cation of mycorrhizae and rusts. Molecular Ecology, 2,113–118. https://doi.org/10.1111/j.1365-294X.1993.tb00005.x
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symposium Series, 41, 95-98.
Hashem, A. H., Attia, M. S., Kandil, E. K., Fwazi, M. M., Abdelrahman, A. S., Khader, M. S. et al. (2023). Bioactive compounds and biomedical application of endophytic fungi: a recent review. Microbial Cell Factories, 22, 107. https://doi.org/10.1186/s12934-023-02118-x
Huson, D. H. (1998). SplitsTree: analyzing and visualizing evolutionary data. Bioinformatics, 14, 68–73. https://doi.org/10.1093/bioinformatics/14.1.68
Huson, D. H., & Bryant, D. (2006). Application of phylogenetic networks in evolutionary studies. Molecular Biology and Evolution, 23, 254–267. https://doi.org/10.1093/molbev/msj030
Katoh, K., Rozewicki, J., & Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Kilpatrick, J. A., & Chilvers, G. A. (1981). Variation in a natural population of Epicoccum purpurascens. Transactions of the British Mycological Society, 77, 497–508. https://doi.org/10.1016/S0007-1536(81)80097-6
Lee, A. J., Cadelis, M. M., Kim, S. H., Swift, S., Copp, B. R., & Villas-Boas, S. G. (2020). Epipyrone A, a broad-spectrum antifungal compound produced by Epicoccum nigrum ICMP 19927. Molecules, 25, 5997. https://doi.org/10.3390/molecules25245997
Li, T., Im, J., & Lee, J. (2022). Genetic diversity of Epicoccum nigrum and its effects on Fusarium graminearum. Mycobiology, 50, 457–466. https://doi:10.1080/12298093.2022.2148394
Link, H. F. (1816). Observationes in ordines plantarum naturales III. Magazin der Gesellschaft Naturforschender Freunde Berlin, 7, 37–38.
Liu, D., Coloe, S., Baird, R., & Pedersen, J. (2000). Rapid mini-preparation of fungal DNA for PCR. Journal of Clinical Microbiology, 38, 471. https://doi:10.1128/jcm.38.1.471-471.2000
Maddison, W., & Maddison, D. (2019). Mesquite: a modular system for evolutionary analysis. Version 3.61. https://www.mesquiteproject.org
Müller, J., Müller, K., Neinhuis, C., & Quandt, D. (2010). PhyDE Phylogenetic data editor, version 0.9971. Program distributed by the authors. http://www.phyde.de
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S. et al. (2012). Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61, 539–542. https://doi.org/10.1093/sysbio/sys029
Schol-Schwarz, M. B. (1959). The genus Epicoccum link. Transactions of the British Mycological Society, 42, 149–173. https://doi.org/10.1016/S0007-1536(59)80024-3
Taguiam, J. D., Evallo, E., & Balendres, M. A. (2021). Epicoccum species: ubiquitous plant pathogens and effective biological control agents. European Journal of Plant Pathology, 159, 713–725. https://doi.org/10.1007/s10658-021-02207-w
Talontsi, F. M., Dittrich, B., Schüffler, A., Sun, H., & Laatsch, H. (2013) Epicoccolides: antimicrobial and antifungal polyketides from an endophytic fungus Epicoccum sp. associated with Theobroma cacao. European Journal of Organic Chemistry, 2013, 3174–3180. https://doi.org/10.1002/ejoc.201300146
Taylor, J. W., Jacobson, D. J., Kroken, S., Kasuga, T., Geiser, D. M., Hibbett, D. S. et al. (2000). Phylogenetic species recognition and species concepts in fungi. Fungal Genetics and Biology, 31, 21–32. https://doi.org/10.1006/fgbi.2000.1228
Vilgalys, R., & Hester, M. (1990). Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology, 172, 4238–4246. https://doi.org/10.1128/jb.172.8.4238-4246.1990
Wang, Y., & Guo, L. D. (2004). Morphological and molecular identification of an endophytic fungus Epicoccum nigrum. Mycosystema, 23, 474–479. https://doi:10.3329/bipt.v29i2.63534
Wen, J., Okyere, S. K., Wang, S., Wang, J., Xie, L., Ran, Y. et al. (2022). Endophytic fungi: an effective alternative source of plant-derived bioactive compounds for pharmacological studies. Journal of Fungi, 8, 205. https://doi.org/10.3390/jof8020205
White, T. J., Bruns, T. D., Lee, S. B., & Taylor, J. W. (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In M. A. Innis, D. H. Gelfand, J. J. Sninsky, & T. J. White (Eds.), PCR Protocols: a guide to methods and applications (pp. 315–322). New York: Academic Press. http://dx.doi.org/10.1016/B978-0-12-372180-8.50042-1
Woudenberg, J. H. C., Aveskamp, M. M., de Gruyter, J., Spiers, A. G., & Crous, P. W. (2009). Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia: Molecular Phylogeny and Evolution of Fungi, 22, 56–62. https://doi.org/10.3767/003158509X427808
Zhang, Z., Schwartz, S., Wagner, L., & Miller, W. (2000). A greedy algorithm for aligning DNA sequences. Journal of Computational Biology, 7, 203–214. https://doi.org/10.1089/10665270050081478
Filogenia basada en COI y caracterización morfológica de una población brasileña de Lambornella trichoglossa (Ciliophora: Tetrahymenidae)
Vítor Ribeiro-Halfeld *
Universidade Federal de Juiz de Fora, Laboratório de Protozoologia, Rua José Lourenço Kelmer, s/n, São Pedro, Juiz de Fora, 36033-900 Minas Gerais, Brazil
In this work, the morphological characterization and molecular phylogeny (SSU rDNA and COI) of Lambornella trichoglossa Foissner, 2003, found in phytotelmata environments in southeastern Brazil is presented. The morphological study performed with silver impregnation techniques demonstrated parameters similar to those recorded in the type population, and additional new information was also obtained, including the conjugation process and the occurrence of specimens with an increased number of caudal cilia. The phylogenetic reconstructions based on both markers suggest the inclusion of L. trichoglossa in the genus Tetrahymena Furgason, 1940. The COI sequence obtained in this work is the first mitochondrial sequence of the genus Lambornella deposited in GenBank and published in a scientific article.
En este trabajo se realizó la caracterización morfométrica y la filogenia molecular (SSU rDNA y COI) de Lambornella trichoglossa Foissner, 2003, encontrada en ambientes de fitotelmata en el sureste de Brasil. El estudio morfológico realizado con técnicas de impregnación de plata demostró parámetros similares a los registrados en la población tipo y también se obtuvo nueva información adicional, incluyendo el proceso de conjugación y la presencia de especímenes con un mayor número de cilios caudales. Las reconstrucciones filogenéticas basadas en ambos marcadores sugieren la inclusión de L. trichoglossa en el género Tetrahymena Furgason, 1940. La secuencia COI obtenida en este trabajo es la primera de tipo mitocondrial del género Lambornella depositada en GenBank y publicada en un artículo científico.
The genus Lambornella has its systematic history marked by alterations. The type species, Lambornellastegomyiae Keilin, 1921, is a mosquito (Diptera, Culicidae) parasite. It was reclassified in the genus Tetrahymena Furgason, 1940 by Corliss (1960). However, the description of Lambornellaclarki Corliss, 1976, another parasitic species, revalidated the genus, indicating the formation of cuticular cysts and the number of post-oral kineties as diagnostic features. Nevertheless, Strüder-Kypke et al. (2001) suggested the non-validity of the genus Lambornella once again, due to its position within the Tetrahymena clade, based on phylogenetic analyses inferred from small subunit ribosomal DNA (SSU). Results presented by Bourland and Stüder-Kypke (2010), and Dunthorn et al. (2012), also based on SSU sequences, demonstrated a similar grouping.
Lambornella trichoglossa Foissner, 2003 is the only free-living species of the genus. It occurs endemically in phytotelmata environments (Foissner, 2003). Despite the recurrent records in the Neotropical region (Buosi et al., 2014, 2015; Durán-Ramírez et al., 2015; Foissner, 2003; Foissner et al., 2003), no studies have investigated Lambornella’s phylogenetic position based on mitochondrial markers. According to Chantangsi and Lynn (2008), the mitochondrial gene cytochrome c oxidase-subunit I (COI) is effective in elucidating recent phylogenetic events in the genus Tetrahymena. In this context, the present work aimed to investigate the molecular phylogeny of L. trichoglossa based on the mitochondrial marker COI. Morphological analysis of the specimens was also performed in order to confirm specific identification and investigate possible morphological variation.
Materials and methods
One hundred milliliters of phytotelmata content were collected from 12 bromeliads belonging to Portea petropolitana at the Botanical Garden of the Federal University of Juiz de Fora (21°43’74” S, 43°22’06” W), in September, 2019. On the same day of collections, the samples were analyzed under a stereoscopic microscope with transmitted light. Active ciliates were picked with glass micropipettes and processed, according to Foissner (2014), to perform silver carbonate and dry silver nitrate impregnation techniques. From the collection day, over a period of 7 days, 20 specimens of L. trichoglossa were screened from the samples and measured in vivo to check for possible alterations in body length. Obtained data were analyzed using the Shapiro-Wilk normality test, and the means recorded on the first and seventh days were compared using the Student’s t-test. All statistical analyses related to the morphological data were conducted using PAST software, version 4.03 (Hammer et al., 2001).
Thirty specimens of L. trichoglossa were picked from the samples and fixed in absolute ethanol for molecular analysis. Total DNA extraction was performed using the Blood and Tissue kit (Qiagen®), following the manufacturer’s guidelines. Primers F388dT and R1184dT (Strüder-Kypke & Lynn, 2010) were used to amplify the COI gene in 25-microliter reactions. The SSU marker was also sequenced for comparison with the type population. Therefore, primers 18S F9Euk and 18S R1513 (Schrallhammer et al., 2013) were used.
PCR products were visualized in a 1% agarose gel and purified using the QIAquick PCR Purification Kit (Qiagen®), following the manufacturer’s guidelines. Subsequently, the material was sent for sequencing according to the Sanger method in 7 µl reactions, using the M13 forward and M13 reverse primers (Messing, 1983) for COI, and 18S R536, 18S F783, 18S F919, and 18S R1052 for SSU (Modeo et al., 2006). Sequencing reactions were performed on an ABI PRISM® 3100 sequencer (Applied Biosystems).
Obtained sequences were added to the respective datasets, jointly with COI and SSU sequences of approximately 20 species of the genus Tetrahymena, as well as outgroups obtained from GenBank accessed in May 2025. Sequences were aligned using MAFFT software, version 7 (Katoh et al., 2017). The resulting alignment was edited in the GBlocks platform, version 0.91b (Talavera & Catresana, 2007). The determination of the best nucleotide substitution model (GTR + G + I to COI, and TN93 + G to SSU) was performed with the aid of MegaX software (Kumar et al., 2018), using the maximum likelihood method, considering all sites. The maximum likelihood phylogenetic analysis was inferred using RAxML software, version 8 (Stamatakis, 2014), at its default settings, with 500 bootstrap replicates for COI, and 1,200 for SSU. The evolutionary distances of COI and SSU sequences were computed in Mega X, using the Kimura 2-parameter method (Kimura, 1980). A third dataset with 74 COI sequences of Tetrahymena species was prepared, in addition to outgroups, with the aim of performing the pairwise distance calculations with a greater number of species (Supplementary material). For this analysis, the Maximum Composite Likelihood model (Tamura et al., 2004) was used, with the aid of MegaX (Kumar et al., 2018).
Results
Table 1 presents the morphological data obtained. Morphological characterization of the L. trichoglossa population analyzed in this study resembles the type population described by Foissner (2003), presenting slightly higher values for body size and the number of somatic kineties. However, postoral kineties appear in lower numbers. Another important morphological characteristic distinct from the type population detected in the present study was the occurrence of specimens with a greater number of caudal cilia (Fig. 1h). These organisms have a complex of caudal cilia, as shown in Figure 1h and Supplementary material. However, they were not included in the morphological statistical analysis due to the unsatisfactory results of the impregnation techniques in their oral and some somatic structures. Opportunely, conjugating forms of L. trichoglossa were recorded, which is unprecedented for the species (Fig. 1a). The temporal analyses on body length performed demonstrated a significant decrease in the average size of the organisms over 7 days (Fig. 2, Table 2).
The obtained SSU sequence was deposited in GenBank with accession number MN567691. It matches 99.97% of the type population’s sequence (AJ810078) (Table 3). The phylogenetic reconstruction inferred by the maximum likelihood method revealed that the new sequence is grouped with L. trichoglossa (AJ810078), T. corlissi (U17356), and T. berger (AF364039) (Fig. 3).
Lambornella trichoglossa’s COI sequence was deposited in GenBank with accession number MN477017. It has 774 nucleotides, with the following percentage of bases: A = 33%, C = 12%, G = 13.4%, and T = 41.6%. Phylogenetic reconstruction using the maximum likelihood method demonstrated L. trichoglossa clustering in a clade with the same topology revealed by the SSU phylogeny; it also grouped with T. corlissi (EF070279) and T. bergeri (EF070270) with a support value of 86%, besides T. scolopax (KJ028669) (Fig. 4). The average paired difference of the 74 Tetraymena sequences used in the phylogenetic analyses was 12.99%. The L.trichoglossa sequence differs by 13.04% from the T.corlissi sequence (Table 4). Although the value is slightly above the average for the Tetrahymena species analyzed, this percentage is lower than the difference observed between T. corlissi and T. caudata (14.29%), T.paravorax (14%), and T. glochidiophila (16.39%), for example.
Table 1
Morphological data on silver carbonate-impregnated specimens of Lambornella trichoglossa.
x
M
SD
SE
CV
Min
Max
n
Body, lenght
269.75
269.43
26.72
4.88
9.906
226.65
327.91
30
Body, width in ventral view
73.105
73.295
16.05
2.93
21.96
47.44
107.77
30
Body lenght: Lateral width, ratio
3.82
3.906
0.81
0.14
21.19
2.606
5.36
30
Anterior body end to membranelle 1, distance
47.91
48.73
7.407
1.35
15.46
34.14
63.87
30
Oral opening, width
32.45
31.42
5.11
0.93
15.77
21.28
44.12
30
Anterior body end to macronucleus, distance
94.55
92.88
13.86
2.53
14.66
74.16
126.35
30
Macronucleus, length
49.72
43.96
11.48
2.09
23.08
36.92
77.75
30
Macronucleus, width
51.27
48.71
9.42
1.72
18.38
33.74
66.86
30
Macronucleus, number
1
1
0
0
0
1
1
30
Micronucleus, width
5.67
5.5
1.15
0.21
20.408
4
9.56
27
Micronucleus, number
1.03
1
0.19
0.035
18.68
1
2
27
Somatic kineties, total number
52.33
52.5
3.79
0.69
7.24
45
62
30
Somatic kineties, postoral number
6.1
6
0.84
0.15
13.85
5
8
30
Caudal cilia, number
1
1
0
0
0
1
1
30
Measurements in μm. x, arithmetic mean. M, median. SD, standard deviation. SE, standard error of arithmetic mean. CV, coefficient of variation (%). Min, minimum. Max, maximum. n, number of individuals investigated.
Figure 1. Morfological characterization of Lambornella trichoglossa. A, B, E. In vivo. C, D, F. Dry silver impregnation. G, H. Silver Carbonate impregnation. A, First recorded conjugating specimens; B, lateral view; C, oral apparatus; D, specimen with normal number of caudal cilium (dorsal view); E, nuclear apparatus; F, ventral view, showing oral apparatus and postoral kineties, below; G, specimen recorded during divisional morphogenesis, with multiple food vacuoles scattered throughout the body; H, detail of caudal cilia complex recorded in larger specimens. OA (oral apparatus), FV (food vacuoles), S (preoral suture), PM (paroral membrane), M1 and M2 (adoral membranelles), MA (macronucleus), MI (micronucleus), CC (caudal cilia). Asterisks indicate macronucleus during divisional morphogenesis process. Scale bars in A, B, D, F, G = 100 μm. In C, E H = 50μm. Figure 2. Body length reduction of L. trichoglossa over a 7-day period. Figure 3. Maximum likelihood (ML) tree inferred from small subunit ribosomal DNA (SSU rDNA). Bold: sequence obtained in the present work. Numbers at nodes represent the bootstrap values of ML out of 1,200 replicates.
Table 2
Statistical data on the difference in body length of L. trichoglossa 7 days after collection.
Number of measured specimens
1st day
20
7th day
20
Body length mean (μm)
1st day
205.66
7th day
148.255
Maximum length (μm)
1st day
222.79
7th day
164.84
Minimum length (μm)
1st day
190.18
7th day
132.43
Standart deviation
1st day
10.97027077
7th day
10.06698748
Variance
1st day
120.3468408
7th day
101.3442368
Shapiro-Wilk test (α = 0.05)
1st day
W: 0.9273
p-value: 0.1371
7th day
W: 0.9556
p-value: 0.4607
Student’s t-test
t: 17.241
p: 1.47.10-19
Critical t-value (p = 0.05): 2.0244
Discussion
The similarity of the morphological parameters between the specimens analyzed in the present study and the data presented in the description of L. trichoglossa ensures the correct taxonomic identification of the organisms. It should be noted that the characterization performed by Foissner (2003) occurred a few days after collection, whereas in the present work, the morphological study techniques were performed a few hours after collection. The temporal analyses on body length performed in the present work demonstrated a decrease in the average size of the organisms over time (Fig. 2), which would justify the slightly higher average values compared to the type population.
Regarding the occurrence of specimens with a greater number of caudal cilia, polymorphic life cycles are a common characteristic of the genus Tetrahymena. Lynn and Doerder (2012) mentioned the occurrence of changes in size, body shape, organization, morphology, and cilia of the oral apparatus of Tetrahymena species, due to environmental changes. The aforementioned authors highlighted the great genotypic and phenotypic plasticity found in the clade.
Phylogenetic reconstructions performed with both markers demonstrate the internal position of L. trichoglossa in the genus Tetrahymena, corroborating analyses based on the SSU marker presented by Strüder-Kypke et al. (2001), Bourland and Strüder-Kypke (2010), and Dunthorn et al. (2012). Several authors demonstrated the division of the genus Tetrahymena into 2 strongly molecularly supported clades, Australis and Borealis (Chantangsi, 2007; Chantangsi & Lynn, 2008; Lynn et al., 2018; Nanney, 1998; Strüder-Kypke et al., 2001). This division was observed in the inferred reconstructions by the SSU marker, performed in the present study, demonstrating the inclusion of L. trichoglossa into the Borealis clade. Doerder (2018) states that this clade has greater molecular diversity and more species than the Australis clade, thus being an important source of diversity for the genus. Lambornella trichoglossa, included in this clade, is the greatest example of phenotypic diversification, given its morphological and ontogenetic particularities demonstrated by Foissner (2003).
The phylogenetic analyses inferred by the COI gene demonstrated the inclusion of L. trichoglossa in a clade called coxset A4 by Chantangsi and Lynn (2008). The species T. corlissi and T. bergeri, members of this clade, share the parasitic lifestyle and have particular morphological characteristics (Hoffman et al., 1975; Imai et al., 2000; Strüder-Kypke et al., 2001). Foissner (2003) performed infection tests with L. trichoglossa and different mosquito species, noting that the ciliate does not develop a parasitic association. However, congeners L. stegomyiae and L. clarki are parasites of culicid dipterans, demonstrating that this way of life is predominant in coxiset A4, corroborating Chantangsi and Lynn (2008). Probably, parasitism also represents the ancestral condition of this clade, since Strüder-Kypke et al. (2001) demonstrated that histophagy is a recurrent phenomenon in the evolution of Tetrahymena.
Figure 4. Maximum likelihood (ML) tree inferred from cytochrome oxidase subunit I (COI). Bold: sequence obtained in the present work. Numbers at nodes represent the bootstrap values of ML out of 500 replicates.
Table 3
Pairwise distance between SSU sequences of Lambornella trichoglossa and Tetrahymena spp.
MN567691
L. trichoglossa
AJ810078
L. trichoglossa*
0.0029
JQ723973
Lambornella sp.**
0.0053
0.0047
U17356
T. corlissi
0.0059
0.0063
0.0077
AF364039
T. bergeri
0.0071
0.0092
0.0089
0.0046
EF070247
T. furgasoni
0.0130
0.0149
0.0149
0.0099
0.0092
AF364040
T. mobilis
0.0130
0.0151
0.0149
0.0104
0.0092
0.0006
KJ028502
T. colerunensis
0.0132
0.0139
0.0139
0.0087
0.0080
0.0051
0.0058
KJ028504
T. scolopax
0.0135
0.0156
0.0156
0.0094
0.0067
0.0135
0.0142
0.0125
Bold font: sequence obtained in the present study. * Sequence obtained from the type population (Dunthorn et al., 2012). ** Sequences obtained from samples collected in Jamaica (Dunthorn et al., 2012).
Table 4
Pairwise distance between COI sequences of Lambornella trichoglossa and Tetrahymena spp.
MN477017
L. trichoglossa
EF070279
T. corlissi
0.1294
EF070270
T. bergeri
0.1450
0.0921
EF070291
T. malaccensis
0.1536
0.1187
0.1320
EF070268
T. asiatica
0.1557
0.1026
0.1113
0.1062
KY218158
T. alphathermophila
0.1713
0.1152
0.1403
0.1175
0.1281
EF070296
T. paravorax
0.1961
0.1400
0.1491
0.1413
0.1185
0.1407
KJ028669
T. scolopax
0.1982
0.1364
0.1376
0.1461
0.1184
0.1534
0.1491
EF070278
T. caudata
0.1857
0.1429
0.1492
0.1622
0.1331
0.1553
0.1735
0.1615
MF693881
T. glochidiophila
0.2187
0.1639
0.1867
0.1626
0.1631
0.1703
0.1500
0.1926
0.2051
Bold font: sequence obtained in the present study.
Traditionally recognized Tetrahymena species, described based on morphological or biochemical characteristics, and molecularly supported, may have high evolutionary difference values when comparing COI sequences. For example, T. caudata differs by over 15% from T. corlissi, T. termophila, and T. glochidiophila. Some species described by Doerder (2018) based mainly on molecular diversity differ by values close to 20%. Therefore, the 12.94% distance observed between the COI sequence of L. trichoglossa and T. corlissi does not represent a discrepancy concerning the molecular variability observed in Tetrahymena.
According to Chantangsi and Lynn (2008), the mitochondrial COI gene is effective in elucidating Tetrahymena’s recent phylogeny, while the SSU marker is more suited to assessing the deep phylogeny of the genus. The maintenance of topology in the reconstructions performed with both markers demonstrates that L. trichoglossa, T. corlissi, and T. bergeri share a long evolutionary history, and the differentiation of the species belonging to the genus Lambornella could be recent within the phylogeny of Tetrahymena. The phylogenetic position of L. trichoglossa also demonstrated by the COI gene, reinforces the proposals for synonymization of the genera Lambornella and Tetrahymena. Although Lambornella’s description precedes that of Tetrahymena, the principle of stability is appropriate to maintain the name of the latter taxon. In any case, it is necessary to establish an extended diagnosis for Tetrahymena, so that the species involved in new combinations are covered.
Finally, the molecular analysis of L. trichoglossa based on the COI marker supports the proposal for synonymization of the genera Lambornella and Tetrahymena. This systematic readjustment also points to the great molecular, morphological, evolutionary and ecological complexity of the genus Tetrahymena, which represents one of the greatest examples of evolutionary adaptation among eukaryotic organisms.
References
Bourland, W. A., & Strüder-Kypke, M. (2010). Agolohymena aspidocauda nov. gen., nov.spec., a histophagous freshwater tetrahymenid ciliate in the family Deltopylidae (Ciliophora, Hymenostomatia), from Idaho (northwest U.S.A.): morphology, ontogenesis and molecular phylogeny. European Journal of Protistology, 46, 221–242. https://doi.org/10.1016/j.ejop.2010.04.003
Buosi, P. R. B., Utz, L. R. P., Meira, B. R., Silva, B. T. S., Lansac-Tôha, F. M., Lansac-Tôha, F. A. et al. (2014). Rainfall influence on species composition of the ciliate community inhabiting bromeliad phytotelmata. Zoological Studies, 53, 32–44. https://doi.org/10.1186/s40555-014-0032-4
Buosi, P. R. B., Cabral, A. F., Utz, L. R. P., Vieira, L. C. G., & Velho, L. F. M. (2015). Effects of seasonality and dispersal on the ciliate community inhabiting bromeliad phytotelmata in riparian vegetation of a large tropical river. Journal of Eukaryotic Microbiology, 62, 737–749. https://doi.org/10.1111/jeu.12232
Chantangsi, C., & Lynn, D. H. (2008). Phylogenetic relationships within the genus Tetrahymena inferred from the cytochrome c oxidase subunit 1 and the small subunit ribosomal RNA genes. Molecular Phylogenetics and Evolution, 49, 979–87. https://doi.org/10.1016/j.ympev.2008.09.017
Chantangsi, C., Lynn, D. H., Brandl, M. T., Cole, J. C., Hetrick, N., & Ikonomi, P. (2007). Barcoding ciliates: a comprehensive study of 75 isolates of the genus Tetrahymena. International Journal of Systematic and Evolutionary Microbiology, 57, 2412–2423. https://doi.org/10.1099/ijs.0.64865-0
Corliss, J. O., & Coats, D. W. (1976). A new cuticular cyst-producing tetrahymenid ciliate, Lambornella clarki n. sp., and the current status of ciliatosis in culicine mosquitoes. Transactions of the American Microscopical Society, 95, 725–739. https://doi.org/10.2307/3225400
Corliss, J. O. (1960). Tetrahymena chironomi sp. nov., a ciliate from midge larvae, and the current status of facultative parasitism in the genus Tetrahymena. Parasitology, 50, 111–153. https://doi.org/10.1017/S0031182000025245
Doerder, F. P. (2018). Barcodes reveal 48 new species of Tetrahymena, Dexiostoma, and Glaucoma: phylogeny, ecology, and biogeography of new and established species. Journal of Eukaryotic Microbiology, 66, 182–208. https://doi.org/10.1111/jeu.12642
Dunthorn, M., Stoeck, T., Wolf, K., Breiner, H., & Foissner, W. (2012). Diversity and endemism of ciliates inhabiting Neotropical phytotelmata. Systematics and Biodiversity, 10, 195–205. https://doi.org/10.1080/14772000.2012.685195
Durán-Ramírez, C., García-Franco, J. G., Foissner, W., & Mayén-Estrada, R. (2015). Free-living ciliates from epiphytic tank bromeliads in Mexico. European Journal of Protistology, 51, 15–33. https://doi.org/10.1016/j.ejop.2014.09.002
Foissner, W. (2003). Morphology and ontogenesis of Lambornella trichoglossa nov. spec., a new tetrahymenid ciliate (Protozoa,Ciliophora) from Brazilian tank bromeliads (Bromeliaceae). European Journal of Protistology, 39, 63–82. https://doi.org/10.1078/0932-4739-00887
Foissner, W. (2014). An update of basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. International Journal of Systematic and Evolutionary Microbiology, 64, 271–292. https://doi.org/10.1099/ijs.0.057893-0
Foissner, W., Strüder-Kypke, M., van der Staay, G. W. M., van der Staay, S. M., & Hackstein, J. H. P. (2003). Endemic ciliates (Protozoa,Ciliophora) from tank bromeliads (Bromeliaceae): a combined morphological, molecular, and ecological study. European Journal of Protistology, 39, 365–372. https://doi.org/0.1078/0932-4739-00005
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4, 1–9.
Hoffman, G. L., Lando, M., Camper, J. E., Coats, D. W., Stookey, J. L., & Burek, J. D. (1975). A disease of freshwater fishes caused by Tetrahymena corlissi Thompson, 1955, and a key for identification of holotrich ciliates of freshwater fishes. The Journal of Parasitolgy, 61, 217–223. https://doi.org/10.1111/j.1365-2761.2009.01062.x
Imai, S., Tsurimaki, S., Goto, E., Wakita, K., & Hatai, K. (2000). Tetrahymena infection in Guppy, Poecilia reticulata. Fish Pathology, 35, 67–72. https://doi.org/10.3147/jsfp.35.67
Katoh, K., Rozewicki, J., & Yamada, K. D. (2017). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Keilin, D. (1921). On a new ciliate: Lambornella stegomyiae n. g., n. sp., parasitic in the body-cavity of the larvae of Stegomyia scutellaris Walker (Diptera, Nematocera, Culicidae). Parasitology, 13, 216–224. https://doi.org/10.1017/s0031182000012488
Kimura, M. (1980). A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 16, 111–120. https://doi.org/10.1007/BF01731581
Kumar, S., Stecher, G., Li, M., Knyaz, C., & Tamura, K. (2018). MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Molecular Biology and Evolution, 35, 1547–1549. https://doi.org/10.1093/molbev/msy096
Lynn, D. H., & Doerder, F. P. (2012). The life and times of Tetrahymena. In Collins, K. Methods in cell biology. Elsevier, 109, 9–27. https://doi.org/10.1016/B978-0-12-385 967-9.00002-5
Lynn, D. H., Doerder, F. P., Gillis, P. L., & Prosser, R. S. (2018). Tetrahymena glochidiophila n. sp., a new species of Tetrahymena (Ciliophora) that causes mortality to glochidia larvae of freshwater mussels (Bivalvia). Diseases of Aquatic Organisms, 127, 125–136. https://doi.org/10.3354/dao03188
Lynn, D. H. (2008). The ciliated taxa including families and genera. In D. H. Lynn (Ed.), The ciliated protozoa (pp. 339–440). Guelph, Canada: Springer.
Messing, J. (1983). New M13 vectors for cloning. Methods in Enzymology, 101, 20–78. https://doi.org/10.1016/0076-6879(83)01005-8
Modeo, L., Rosati, L., Andreoli, I., Simoni, G., Verni, F., & Petroni, G. (2006). Molecular systematics and ultrastructural characterization of a forgotten species: Chattonidium setense (Ciliophora, Heterotrichea). Proceedings of the Japan Academy, 82, 359–374. https://doi.org/10.2183/pjab.82.359
Nanney, D. L., Park, C., Preparata, R., & Simon, E. M. (1998). Comparison of sequence differences in a variable 23S rRNA domain among sets of cryptic species of ciliated protozoa. Journal of Eukaryotic Microbiology, 45, 91–100. https://doi.org/10.1111/j.1550-7408.1998.tb05075.x
Schrallhammer, M., Ferrantini, F., Vannini, C., Galati, S., Schweikert, M., Görtz, H. et al. (2013). Candidatus Megaira polyxenophila gen. nov., sp. nov.: Considerations on evolutionary history, host range and shift of early divergent Rickettsiae. Plos One, 8, e72581. https://doi.org/10.1371/journal.pone.0072581
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics, 30, 1312–3. https://doi.org/10.1093/bioinformatics/btu033
Strüder-Kypke, M., Wright, H. D., Jerome, C. A, & Lynn, D. H. (2001). Parallel evolution of histophagy in ciliates of the genus Tetrahymena. Evolutionary Biology, 1, 1–5. https://doi.org/10.1186/1471-2148-1-5
Strüder-Kypke, M., & Lynn, D. H. (2010). Comparative analysis of the mitochondrial cytochrome c oxidase subunit I (COI) gene in ciliates (Alveolata, Ciliophora) and evaluation of its suitability as a biodiversity marker. Systematics and Biodiversity, 8, 131–148. https://doi.org/10.1080/14772000903507744
Tamura, K., Nei, M., & Kumar, S. (2004). Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences (USA), 101, 11030–11035. https://doi.org/10.1073/pnas.0404206101
Talavera, G., & Castresana, J. (2007). Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Systematic Biology, 56, 564–577. https://doi.org/10.1080/10635150701472164
The genus Psephenops (Coleoptera: Dryopoidea: Psephenidae) in Mexico, with the description of a new species and notes on the species from Central America and the Antilles
Roberto Arce-Pérez, J. Antonio Gómez-Anaya y Rodolfo Novelo-Gutiérrez *
Instituto de Ecología, A. C., Red de Biodiversidad y Sistemática, Carretera Antigua a Coatepec 351, Col. El Haya, 91073 Xalapa, Veracruz, México
*Autor para correspondencia: rodolfo.novelo@inecol.mx (R. Novelo-Gutiérrez)
Recibido: 05 junio 2025; aceptado: 26 septiembre 2025
El género neotropical Psephenops Grouvelle incluye actualmente 15 especies descritas, de las cuales 13 se distribuyen en México, América Central y las Antillas, más una de Argentina y otra de Perú. Se reporta por primera vez a la subfamilia Psepheninae con Psephenops oaxacanus sp. nov. para el estado de Oaxaca, México, elevando a 3 el número de especies conocidas para el país, 14 en México, América Central y las Antillas y 16 a nivel continental. Se proporciona un mapa de distribución y una clave ilustrada para las especies presentes en México.
Palabras clave: Psepheninae; Moneditas de agua; Oaxaca; México; Taxonomía
Abstract
The Neotropical genus Psephenops Grouvelle currently includes 15 described species, 13 of which are distributed in Mexico, Central America, and the Antilles, plus one from Argentina and another from Peru. The subfamily Psepheninae is reported for the first time with Psephenops oaxacanus sp. nov. from the state of Oaxaca, Mexico, increasing the number of known species in the country to 3, 14 in Mexico, Central America and the Antilles, and 16 at the continental level. A distribution map and an illustrated key for the species present in Mexico are provided.
Keywords: Psepheninae; Water pennies; Oaxaca; Mexico; Taxonomy
Introducción
El género neotropical Psephenops Grouvelle, 1898 (= Xexanchorinus) está representado por 15 especies descritas hasta la fecha: P. argentinensis Deléve, 1967 (Argentina); P. bifidus Arce-Pérez y Novelo-Gutiérrez, 2017 (Panamá); P. grouvellei Champion, 1913 (Guatemala); P. haitianus Darlington, 1936 (Haití); P. lupita Arce-Pérez, 2002 (México); P. maculicollis Darlington, 1936 (Costa Rica y Colombia); P. mexicanus Arce-Pérez y Novelo-Gutiérrez, 2000 (México); P. panamaensis Arce-Pérez y Novelo-Gutiérrez, 2015 (Panamá); P. prestonae Spangler 1990 (Costa Rica); P. robacki (Spangler), 1966 (Perú); P. shepardi Arce-Pérez y Novelo-Gutiérrez, 2013 (Belice); P. smithi Grouvelle, 1898 (Antillas menores); P. spiniparameri Arce-Pérez y Novelo-Gutiérrez, 2013 (Belice); P. triangularis Arce-Pérez y Novelo-Gutiérrez, 2017 (Panamá); y P. trini Barr y Shepard, 2024 (Antillas menores) (Arce-Pérez 2002; Arce-Pérez y Novelo-Gutiérrez, 2000, 2013, 2015, 2017; Bameul, 2001; Barr y Shepard, 2024; Deléve, 1967). En este trabajo se reporta por primera vez al género Psephenops para Oaxaca, México, con base en ejemplares recolectados en el municipio de Santiago Comaltepec y se describe e ilustra a Psephenops oaxacanus sp. nov.
Materiales y métodos
Los ejemplares de la especie nueva se recolectaron mediante una red acuática con marco tipo D y se colocaron en frascos con etanol al 96%, en un tributario del río Soyolapam que corre por la selva alta perennifolia (fig. 3), en el poblado San Martín Soyolapam, municipio Santiago Comaltepec, en Oaxaca, México en febrero del 2016 (fig. 4). Los especímenes fueron identificados utilizando las descripciones y claves originales de Arce-Pérez y Novelo-Gutiérrez (2000, 2013, 2015, 2017). El examen morfológico se realizó utilizando un microscopio estereoscópico Nikon SMZ 745. Las imágenes y mediciones de estructuras específicas se obtuvieron mediante el uso de un microscopio estereoscópico Nikon SMZ25 acoplado al software Nikon NIS-Elements Imaging seguido de la preparación de imágenes con Photoshop CS5 (Adobe System Inc. San José, California). Las micrografías electrónicas se obtuvieron utilizando un microscopio electrónico de barrido Jeol JSM-5600LV. Las mediciones están en mm. El mapa de distribución geográfica se realizó con Simplemappr (Shorthouse, 2010).
Descripción
Con base en 6 machos y 6 hembras se describe Psephenops oaxacanus sp. nov., representando el primer registro de la subfamilia para el estado de Oaxaca, elevando a 3 el número de especies a nivel nacional, 14 en México, América Central y las Antillas y 16 a nivel continental. Se proporciona una lista anotada, claves dicotómicas y mapa de distribución para las especies presentes en México.
Familia Psephenidae Lacordaire, 1854
Subfamilia Psepheninae Lacordaire, 1854
Genero Psephenops Grouvelle, 1898
Psephenops oaxacanus Arce-Pérez, Gómez-Anaya y Novelo-Gutiérrez sp. nov. (figs. 1-6)
Diagnosis. Habitus (fig. 1): longitud total 3.2 mm; ancho humeral 1.4 mm. Cuerpo ovalado, deprimido; cabeza, pronoto y escutelo negro rojizo, élitros pardo rojizos, ventralmente pardo oscuros, al igual que patas, antenas y palpos. Dorsalmente cubierto completamente por vestidura de sedas diminutas de color amarillo con reflejos dorados, además cabeza y pronoto con sedas más largas y gruesas de color rojizo oscuro; pronoto con cresta levantada en el tercio distal, antenas moniliformes cortas que no rebasan región distal del pronoto. Ventralmente la vestidura presenta sedas diminutas blanquecinas amarillentas con reflejos dorados. Tarsos con los lóbulos de los tarsómeros 1 y 2 cortos, con vestidura ventral amarilla (fig. 1a, b). Se distingue de las otras especies mexicanas del género por la combinación de los caracteres de la tabla 1.
Holotipo macho. Dorsalmente (fig. 1a): cabeza corta, más ancha que larga; clípeo subrectangular, en declive menor a 90 grados desde plano frontal, margen distal ampliamente emarginado; vértex con surco longitudinal medio; superficie fronto-clipeal y vértex micropunteado, con sedas diminutas de color amarillo y sedas más largas y gruesas de color rojo oscuro. Antenas cortas, moniliformes, de 11 antenómeros, sin alcanzar margen posterior del pronoto; escapo más largo y robusto; pedicelo subgloboso, mitad de largo que escapo, ambos ligeramente más claros que antenómeros del flagelo; flagelo de 8 antenómeros, subtriangulares anchos, antenómero apical acuminado. Ojos laterales, redondeados, muy prominentes, de color rojo oscuro, con zona postocular amarilla y sedas largas amarillentas. Labro subrectangular similar al clípeo, pero más corto, ampliamente emarginado. Palpos maxilares de 4 artejos, cubiertos con sedas cortas, palpómero basal más pequeño y redondo, palpómero apical más grande, acuminado y con área apical sensorial amplia, longitud de los palpómeros: 0.04, 0.08, 0.06 y 0.14 mm. Palpos labiales muy cortos, de 3 artejos, palpómeros 1-2 redondeados y anchos, 3 subcónico, con ápice redondeado y ligeramente más pequeño que anteriores. Ventralmente (fig. 1b): negra rojiza, partes bucales pardo oscuro. Tórax: pronoto subtrapezoidal, casi tan ancho como base de élitros, 0.75 mm de largo por 1.2 mm de ancho basal; margen anterior arqueado, margen posterior bisinuado; márgenes laterales casi rectos a lo largo de mitad anterior y expandido lateralmente en mitad posterior. Disco pronotal con una cresta longitudinal bien desarrollada en tercio distal y ligera depresión ovalada a cada lado de ésta, además de otra depresión en margen laterodistal (fig. 1a); superficie pronotal diminutamente punteada, cubierta por vestidura de sedas diminutas amarillas con reflejos dorados y sedas largas de color rojo oscuro. Escutelo corto, más ancho en la base que largo, ápice redondeado. Élitros largos, exponiendo solo último esternito abdominal (pigidio), con márgenes laterales paralelos en los 0.75 basales, luego convergiendo gradualmente hacia atrás, con ancho humeral de 1.4 mm y cubiertos por una vestidura de sedas diminutas amarillas con reflejos dorados. Prosternón estrecho, con proceso postcoxal largo y lanceolado, 0.50 mm de largo por 0.13 mm de ancho en región lanceolada, levantado en toda su longitud y carenado en mitad distal, alcanzando mitad anterior del mesoventrito. Mesoventrito corto, con surco amplio para recepción del proceso postcoxal. Metaventrito grande y voluminoso, con un discrimen evidente que se observa como surco. Patas con coxas anteriores y medias cortas globosas, posteriores planas y transversas; fémures robustos; tibias largas y delgadas, ligeramente más anchas en ápice; protibia con pequeño dentículo apicolateral; metatibias más largas que metafémures; tarsómeros 1 y 2 ventralmente lobulados, con vestidura gruesa parecida a esponja; lóbulo ventral del tarsómero 1 se extiende distalmente sobre 0.30 basales del tarsómero 2; tarsómero 2 el más largo, casi 2 veces más largo que el 1, extendiéndose distalmente y cubriendo completamente a tarsómeros 3, 4 y poco más de 0.50 basales del 5; longitud de tarsómeros: 0.53, 0.94, 0.19, 0.17 y 0.54 mm (sumergidos en alcohol); garras apicales largas, curvas y delgadas, cada una con pequeño diente basal. Abdomenconvestidura corta, fina y densa, blanquecino amarillenta, con reflejos dorados; ventrito VI poco visible, oculto debajo de ventrito V; ventritos I y II anchos, con márgenes anterior y posterior sinuados; ventritos III y IV estrechos, con margen posterior ligeramente arqueado; ventrito V con margen posterior ampliamente arqueado; ventrito VI poco visible, corto, subtriangular, con margen posterior ampliamente arqueado; ventrito VII o placa anal, subtriangular, cubierta de sedas largas y amarillentas; último tergito abdominal o pigidio subtriangular, cubierto de sedas largas rojizo oscuras. Genitales en vista ventral subrectangulares, anchos, longitud de 0.55 mm de falobase al ápice del lóbulo medio, ancho de 0.25 mm entre parámeros y la falobase; lóbulo medio digitiforme con ápice en punta, longitud 0.29 mm, con esclerito medio esbelto, de 0.25 mm de largo, triangular en base, esbelto y claviforme hacia ápice (fig. 2a). En vista dorsal parámeros grandes y robustos de 0.37 mm, con ¼ apical subtriangular y delgada membrana lateral; falobase reducida, 0.18 mm y cóncava (fig. 2b). En vista ventrolateral falobase convexa, parámeros curvados y esclerito del lóbulo medio largo y agudo (fig. 2d). En vista frontal membrana de lóbulo medio fuertemente esclerotizada y escamosa formando cubierta dura (fig. 2c).
Variación en machos. Escasa, excepto por la longitud corporal (n = 5) de 3 a 3.5 mm por 1.2 a 1.5 mm de ancho humeral. La cresta pronotal se puede prolongar como ligera banda longitudinal semidesnuda hacia la región anterior; los élitros pueden estar ligeramente oscurecidos en la región posterior.
Hembra. Más grande y robusta que el macho, longitud 3.6 mm por 1.8 de anchura humeral (fig. 1c, d). Cabeza con ojos muy protuberantes y amplia zona postocular amarilla, cubierta por sedas largas rojizo oscuro; pronoto subtrapezoidal ancho y expandido posteriormente, con 0.90 mm de largo, por 1.7 mm de ancho distal (fig. 1c); proceso postcoxal muy ancho y lanceolado, 0.60 mm de largo por 0.22 mm de ancho en región lanceolada; mesoventrito ampliamente acanalado para recepción del proceso postcoxal; ventritos II y III con corta hendidura transversal a cada lado de línea media (fig. 1d); las patas son más claras, coxas, trocánteres y fémures mayormente rojizo amarillentos, ápice de fémures oscuros, tibias rojizas en margen interno y oscuras en el externo; tarsos totalmente amarillos con uñas y ápice del tarsómero apical oscuros; tarsómeros subcónicos, sin lóbulos ventrales, primeros 4 cortos y subcónicos, 5 esbelto, tan largo como primeros 4 combinados. Genitales con ovipositor ligeramente esclerotizado, hialino, longitud 0.40 mm; valvífera con 2 laterotergitos ligeramente esclerotizados, longitud 0.16 mm; gonocoxitos longitud 0.21 mm, juntos forman placa ancha y plana sin sedas ni puntos aparentes; estilos oblongos, longitud 0.03 mm (fig. 2e).
Variación en hembras. No se observa mucha variación en coloración y vestidura, únicamente algunas con élitros un poco más oscuros; también varían en la longitud corporal (n = 5) de 3.2 a 3.9 mm por 1.5 a 1.9 mm de ancho humeral.
Tabla 1
Caracteres diferenciales de las especies de Psephenops Grouvelle en México, América Central y las Antillas.
Psephenops spp/estructuras
smithi guadeloupensi
grouvellei
haitianus
maculicollis
prestone
mexicanus
lupita
spiniparameri
shepardi
panamaensis
bifidus
triangularis
oaxacanus
Longitud total (mm)
2.51
3.4
2.5
2.5
2.46
3.3
3.8
2.75
2.65
2.4
2.95
2.5
3.2
Tubérculos pronotales
0
3
0
0
0
0
0
0
0
0
0
0
0
Superficie elitral
con estrías
con bordes longitudi- nales
estriado
estriado
liso
liso
liso
liso
liso
liso
liso
liso
liso
Tipo de antenas
monili- formes
monili- formes?
filifor- mes?
monili- formes?
?
monili- formes
filifor- mes
monili- formes
monili- formes
monili- formes
monili- formes
monili- formes
monili- formes
Longitud antenal*
largas
largas
largas
cortas
cortas
cortas
largas
cortas
largas
largas
cortas
cortas
cortas
Proceso prosternal
subcilin- drico no bifido?
?
?
?
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado no bífido
lanceo- lado y bífido
lanceo- lado no bífido
lanceo- lado no bífido
Protibias
?
?
?
?
sin diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
con diente- cillo
Patas con lóbulos en los tarsómeros
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 en frontales y medias, 1 posteriores
1 y 2 de todas las patas
1 a 3 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
1 y 2 de todas las patas
Longitud de lóbulos tarsales**
largos
cortos?
?
largos?
largos
largos
cortos
largos
cortos
largos
largos
largos
cortos
Ápice de parameros
truncados
?
?
no truncado,
no truncado
no truncado
truncado
no truncado, divergente
no truncado,
truncado
no truncado
no truncado
no truncado
Parameros con dientecillo
ausente
ausente
ausente
ausente
ausente
ausente
ausente
ausente
presente
ausente
presente
presente
ausente
mitad basal de los parameros
fusionados
?
?
fusionados
fusionados
con una profunda hendidura
fusionados
con una profunda hendidura
fusionados
fusionados
con una profunda hendidura
fusionados
fusionados con ligera hendidura
Pene con proyección lateral
ausentes
?
?
ausentes
ausentes
pequeños dentículos
ausentes
ausentes
ausentes
ausentes
ausentes
proyección subtrian- gular
ausentes
Forma del esclerito del pene
?
?
?
?
?
lanceolado
digiti- forme
subtrian- gular esbelto
subtrian- gular, ápice bífido
subtrian- gular ancho
recto, ancho en la base
como un reloj de arena
esbelto lanceolado
Distribución
Antillas
Guatemala
Haití
Costa Rica, Colombia
Costa rica
México
México
Belice
Belice
Panamá
Panamá
Panamá
México
*Cortas = Cuando está completamente extendido hacia atrás, no alcanza el margen posterior del pronoto; Largas = que alcanza o sobrepasa el margen posterior del pronoto. ** Corto = Cuando el lóbulo del tarsómero 2 cubre ventralmente toda la longitud del tarsómero 3-4 y la mitad basal o menos del tarsómero 5; Largo = el menor pero que alcanza al menos 80% de la longitud del 5.
Figura 1. Vista dorsal y ventral del macho y hembra de Psephenops oaxacanus. A) Hábito dorsal; B) hábito ventral; C) hábito dorsal; D) hábito ventral. Figura 2. Vista del genital masculino y femenino de Psephenops oaxacanus. A) Vista dorsal; B) vista ventral; C) vista frontal; D) vista ventrolateral; E) vista ventral del genital femenino mostrando los estilos, gonocoxitos y laterotergitos.
Resumen taxonómico
Material tipo. Holotipo macho. México: localidad tipo, estado de Oaxaca, Municipio Santiago Comaltepec, San Martín Soyolapam, río Soyolapam, 136 m snm (17°41’58” N, -96°16’59” O), 5.II.2016. Cols. J.A. Gómez et R. Novelo [escrito a máquina, etiqueta blanca], “Holotype ♂/ Psephenops oaxacanus nov. sp. / Arce-Pérez, Gómez-Anaya & Novelo-Gutiérrez 2025” [escrito a máquina, etiqueta roja], Paratipos: 5 ♂, 6 ♀ mismos datos del holotipo [escrito a máquina, etiqueta amarilla] (IEXA).
Etimología. Esta especie recibe el nombre oaxacanus por ser la primera especie de la subfamilia Psepheninae y del género Psephenops para el estado de Oaxaca, México.
El holotipo y paratipos se encuentran depositados en la Colección Entomológica del Instituto de Ecología, A.C., Xalapa, México (IEXA), con el número de catálogo IEXA-2025-IM20.
Comentarios taxonómicos
Se considera más estrechamente relacionada con P. mexicanus por presentar ambas el mismo tipo de antenas cortas y moniliformes, parámeros en vista dorsal subtriangulares, separados por un profundo surco y fusionados en su base (figs. 2a, 6c), además por el tamaño corporal de 3 a 3.5 mm. Con base en la literatura disponible, la nueva especie puede distinguirse de las de México, América Central y las Antillas por las características anotadas en la tabla 1.
Hábitat. El río Soyolapam se localiza en la localidad del mismo nombre en la sierra de Juárez, Oaxaca, en la subcuenca del río Valle Nacional (fig. 4), el tipo de vegetación es selva alta perennifolia a una altitud de 136 m snm. El río presenta en su cauce cantos y rocas emergentes cubiertas parcialmente por algas y vegetación abundante en sus márgenes, los cuales en algunas partes son playones arenosos (fig. 3).
Figura 3. Localidad tipo de Psephenops oaxacanus en el río Sayolapam (San Martín), Santiago Comaltepec, Oaxaca, México (febrero de 2016).
Hasta el momento, esta especie solo está presente en Oaxaca, al sur de México, en el río Soyolapan, a 136 m snm (fig. 4). La localidad tipo se encuentra en la parte norte del estado, en la provincia sierra Madre del Sur, en la subprovincia de los Altos de Oaxaca. Esta especie se recolectó sintópicamente con ejemplares de los géneros Cylloepus y Macrelmis de la familia Elmidae.
Lista actualizada de las especies de Psephenops Grouvelle en México.
Psephenops lupita (Arce-Pérez)
Psephenops lupita (Arce-Pérez, 2002: 964)
Registros publicados. México: Veracruz: Municipio Coatepec, Poblado Coatepec, río Huehueyapan (Sección La Marina), 19º26’55” N, 96º58’52” O, 1, 200 m, bosque mesófilo de montaña (fig. 4); municipio Xico, Poblado Xico Viejo, río Malseseca, 19º26’44.89” N, 97º03’22.22” O, 1,800 m, bosque mesófilo de montaña (fig. 4).
Psephenops mexicanus (Arce-Pérez y Novelo-Gutiérrez)
Psephenops mexicanus (Arce-Pérez y Novelo-Gutiérrez, 2000: 196)
Registros publicados. México: Veracruz: Municipio Coatepec, Poblado Coatepec, río Huehueyapan (Sección La Marina), 19º26’55” N, 96º58’52” O, 1,200 m, bosque mesófilo de montaña (fig. 4); municipio Xico, Poblado Xico Viejo, río Malseseca, 19º26’44.89” N, 97º03’22.22” O, 1,800 m, bosque mesófilo de montaña (fig. 4).
Figura 4. Distribución de las especies de Psephenops en México. Figura 5. Vista lateral de las patas mostrando lóbulos tarsales en Psephenops spp. A) Cortos; B) largos, ta = tarsómeros. Figura. 6 A-D. Vista del genital masculino de P. lupita y P. mexicanus. A) Dorsal; B) ventral; C) dorsal; D) ventral. Figuras A, B tomadas de Arce-Pérez (2002); C, D tomadas de Arce-Pérez y Novelo-Gutierrez (2000).
Clave para la identificación de las especies mexicanas de Psephenops Grouvelle (modificada de Arce-Pérez y Novelo-Gutiérrez, 2013, 2017)
1 Antenas filiformes, largas, tocando base de pronoto; parámeros en vista dorsal anchos, subcuadrados y fusionados en base en ligera hendidura en forma de gota (fig. 6a), en vista ventral lóbulo medio y esclerito interno digitiformes (fig. 6b); lóbulos tarsales cortos, sin alcanzar más de 0.5 basales del tarsómero 5 (fig. 5a); longitud total 3.75 – 3.85 mm P. lupita
1’ Antenas cortas, moniliformes, no tocan base del pronoto; parámeros en vista dorsal subtriangulares, separados por hendidura que va de ligera a profunda (figs. 2a, 6c); en vista ventral lóbulo medio y esclerito interno no digitiformes; lóbulos tarsales cortos o largos (figs. 5a, b) 2
2 (1’) Lóbulos tarsales largos, cubriendo 0.8 basales del tarsómero 5 (fig. 5b); parámeros en vista dorsal con profunda hendidura longitudinal media (fig. 6c); en vista ventral lóbulo medio subtriangular, con dientecillo a cada lado; esclerito interno del lóbulo medio lanceolado (fig. 6d); longitud 3.15 – 3.55 mm P. mexicanus
2’ Lóbulos tarsales cortos, sin alcanzar más de 0.5 basales del tarsómero 5 (fig. 5a); parámeros en vista dorsal separados por profunda hendidura ancha en forma de lanza invertida y continuada por ligera hendidura que no los separa (fig. 2a); en vista ventral lóbulo medio digitiforme sin dientecillos laterales, terminando en punta ancha, con esclerito interno esbelto, claviforme hacia el ápice, en vista lateral agudo (figs. 2b-d); longitud 3.0 – 3.5 mm P. oaxacanus sp. nov.
Agradecimientos
A Emmanuel Arriaga Varela (INECOL) y a un revisor anónimo por sus atinadas correcciones y sugerencias que mejoraron el manuscrito final.
Referencias
Arce-Pérez, R. (2002). A new species of Psephenops Grouvelle (Coleoptera: Psephenidae) from Mexico. Proceedings of the Entomological Society of Washington, 104, 964–967.
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2000). First record of the genus Psephenops (Coleoptera: Psephenidae) from Mexico, with a description of a new species. Entomological News, 111, 196–200.
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2013). Two new species of Psephenops Grouvelle from Belize (Coleoptera: Byrrhoidea: Psephenidae), with a key to the known species from Mexico and Central America. Zootaxa, 3670, 63–70. http://dx.doi.org/10.11646/zootaxa.3670.1.5
Arce-Pérez, R. y Novelo-Gutiérrez, R. (2017). Two new species of Psephenops Grouvelle (Coleoptera: Byrrhoidea: Psephenidae) from Panama. Zootaxa, 4323, 109–118. https://doi.org/10.11646/zootaxa.4323.1.8
Arce-Pérez, R., Novelo-Gutiérrez, R. y Cornejo, A. (2015). Psephenops panamaensis sp. nov. (Coleoptera: Byrrhoidea: Psephenidae) from Panamá. Zootaxa, 4052, 233–236. https://doi.org/10.11646/zootaxa.4052.2.8
Bameul, F. (2001). Un nouveau Psephenops Grouvelle de la Guadeloupe, avec la description de sa larve (Coleoptera, Psephenidae). Nouvelle Revue d’Entomologie, 18, 161–172.
Barr, C. B. y Shepard, W. D. (2024). A taxonomic review of the genus Psephenops Grouvelle of the Lesser Antilles with description of new species Psephenops trini, and reassignment of Peruvian species Psephenus robacki Spangler (Coleoptera: Psephenidae: Psepheninae). Insecta Mundi, 1045, 1–22. https://doi.org/10.5281/zenodo.11450152
Deléve, J. (1967). Contribution á l’ étude des Dryopoidea (Coleoptera). XIX. Notes diverses et descriptions d’ spéces nouvelles. Bulletin et Annales de la Société Entomologique de Belgique, 103, 414–446.