
Descripción morfológica de la primera zoea del camarón de agua dulce Macrobrachium crenulatum (Decapoda: Palaemonidae) y una comparación con sus congéneres del Neotrópico
Lucas Oliveira-Rogeri a, *, Rodrigo Pantoni a, Ingo S. Wehrtmann b, c, d, Fernando L. Mantelatto a, João Alberto Farinelli Pantaleão e
a University of São Paulo, Faculty of Philosophy, Science and Letters at Ribeirão Preto, Department of Biology, Laboratory of Bioecology and Crustacean Systematics, Av. Bandeirantes, 3900, 14040-901, Ribeirão Preto, São Paulo, Brazil
b Universidad de Costa Rica, Unidad de Investigación Pesquera y Acuicultura, Centro de Investigación en Ciencias del Mar y Limnología, 11501-2060 San José, Costa Rica
c Universidad de Costa Rica, Centro de Investigación en Biodiversidad y Ecología Tropical, Museo de Zoología, 11501-2060 San José, Costa Rica
d Universidad del Valle de Guatemala, Editorial Universitaria, 18 Avenida 11-95, Zonas 15, Vista Hermosa III, Ciudad de Guatemala, Guatemala
e Federal University of Alagoas, Institute of Biological and Health Sciences, Laboratory of Bioecology and Development of Crustaceans, Av. Paulo Holanda, 143, 57072-970, Maceió, Alagoas, Brazil
*Corresponding author: luquinhaor.bio55@usp.br (L. Oliveira-Rogeri)
Received: 08 April 2025; accepted: 13 November 2025
Abstract
We present the first description of Zoea I of Macrobrachium crenulatum and compare it with available data of other zoeae of Macrobrachium spp. of the Neotropics to identify morphological characteristics to differentiate the first zoea of M. crenulatum from zoeae of other species of the genus. A female with embryos was collected in the Atlantic drainage of Costa Rica, Central America. After hatching in the laboratory, larvae were preserved in glycerinate alcohol and later semi-permanent slides were prepared for morphological study and larval description. A table was prepared to compare the morphology of the first zoea of M. crenulatum to the first zoeae of other species of Macrobrachium. Among species of Macrobrachium phylogenetic and geographically closely related, the first zoea of M. crenulatum can be differentiated by the comparison of some structures as antenna (number of articles on scaphocerite), maxillipeds (number of plumose setae; setation pattern), maxilla (setation pattern), maxillule (number of spines), and telson (outer pair of setae). The larval morphology of M. crenulatum corroborated the phylogenetic position and may serve as a taxonomic tool for the distinction between the first zoea of closely related species.
Keywords: Atlantic drainage; Caridea; Costa Rica; Larval morphology
Resumen
Presentamos la primera descripción de la zoea I de Macrobrachium crenulatum y la comparamos con datos disponibles de otras primeras zoeas de especies de Macrobrachium del Neotrópico, con el objetivo de identificar características morfológicas que permitan diferenciar la primera zoea de M. crenulatum respecto de las de otras especies del género. Una hembra ovígera fue colectada en la cuenca atlántica de Costa Rica, Centroamérica. Tras la eclosión en laboratorio, las larvas fueron preservadas en alcohol glicerinado y, posteriormente, se prepararon láminas semipermanentes para su estudio morfológico y la descripción larval. Se elaboró una tabla comparativa con la morfología de la primera zoea de M. crenulatum respecto de las primeras zoeas de otras especies de Macrobrachium. Entre las especies filogenética y geográficamente relacionadas, la zoea I de M. crenulatum puede distinguirse mediante la comparación de estructuras como antenas (número de artejos del escafocerito), maxilípedos (número y patrón de setas plumosas), maxila (patrón de setas), maxílula (número de espinas) y telson (disposición del par lateral de setas marginales). La morfología larval de M. crenulatum respalda su posición filogenética y puede servir como herramienta taxonómica para diferenciar la primera zoea de especies estrechamente relacionadas.
Palabras clave: Vertiente Atlántica; Caridea; Costa Rica; Morfología larvaria
Introduction
The genus Macrobrachium Spence Bate, 1868 is one of the most speciose taxa among caridean shrimps, currently accounting for more than 270 species (De Grave & Fransen, 2011; Pantaleão et al., 2025; Poore & Ahyong, 2023). The genus is cosmopolite, with representatives inhabiting different environments with distinct climate conditions throughout the world. Macrobrachium is remarkably diverse in subtropical and tropical regions, such as Central and South America as well as in the Indo-Pacific region (Murphy & Austin, 2005; Pileggi & Mantelatto, 2012; Pileggi et al., 2014). Some species of Macrobrachium are marine, estuarine and anchialine, but the vast majority inhabit typically freshwater systems, such as rivers, lakes and floodplains (Chace & Bruce, 1993; Fujita et al., 2015; Poore & Ahyong, 2023).
Besides the elevated number of species, the genus is highly diverse in terms of morphological characteristics, ecological traits and reproductive strategies (Sastry, 1983; Wowor et al., 2009). Some species of Macrobrachium, for instance, complete their entire life cycle in freshwater. Others are amphidromous and depend on both freshwater and estuarine/marine environments to finish their cycle, which implies migration events from one water system to the other (Bauer, 2011a, b; Liu et al., 2007; Wowor et al., 2009). Another varying feature of the biology and ecology of species of Macrobrachium is the duration of larval development. While some species have extended larval development (ELD) with a long duration of the larval period and the number of larval stages varying between 9 and 13, others have an abbreviated larval development (ALD) with 1 or 2 free-swimming larval stages before reaching the juvenile stage (Anger, 2013; Jalihal et al., 1993; Shokita, 1985; Wowor et al., 2009).
One of the representatives of this shrimp genus is Macrobrachium crenulatum Holthuis, 1950, with its geographical distribution including the coastal waters of Nicaragua, Costa Rica, Panama, Colombia, Venezuela, Jamaica, Grenada, and Trinidad and Tobago (Holthuis, 1952; Hunte, 1979; Pileggi et al., 2014; Rossi, 2012). This species is characterized by an amphidromous life cycle with extended larval development (ELD) (Bauer, 2013; Pileggi et al., 2014). Adults have been the target of taxonomic, ecological and molecular studies (Pileggi et al., 2014; Rossi, 2012; Valencia & Campos, 2007); however, the larval morphology of M. crenulatum has never been described. This lack of information about the larval morphology hinders the global comprehension of species, since understanding larval features is not only useful for the larval recognition itself, but also for studies about the larval ecology, for resolving taxonomic problems and for providing phylogenetic insights (Lai et al., 2013; Pantaleão et al., 2025).
Pileggi et al. (2014) conducted a molecular study, which provided phylogenetic data about the “sibling species” of Macrobrachium from Central America. According to these authors, morphological features of adults are often not sufficient to distinguish the “sibling species” of the genus, including the distinction between transisthmian pairs of sibling species Macrobrachium hancocki Holthuis, 1950 and M. crenulatum. Recently, Pantaleão et al. (2025) conducted a novel study integrating larval morphology and molecular data to understand the phylogenetic context of relevant traits of species of Macrobrachium life history and larval morphology. Nonetheless, the authors highlighted that one of the limitations was the lack of larval descriptions for the vast majority of species of Macrobrachium as well as the accuracy of the available descriptions.
Considering the high diversity of Macrobrachium in Central America, the absence of larval descriptions hinders the identification of their larvae in plankton samples and thus a broader understanding of their larval ecology, especially in river systems where several species of Macrobrachium co-occur (see Lara & Wehrtmann, 2011). Therefore, here we describe for the first time the first zoea of M. crenulatum. The results include a comparison with other newly hatched larvae of species of Macrobrachium, which may serve as a taxonomic tool for the distinction between the first zoeae of these closely related species.
Materials and methods
An ovigerous female (carapace length = 16.6 mm) was collected on 23 May 2010 in the Suarez River (09º43’40” N, 22º50’25.5” W), at the district of Cahuita, in the province of Limón, in the Atlantic drainage of Costa Rica, Central America. The female was collected using a sieve through ascendant movements near the margins of the river; once obtained, it was moved to the laboratory where it was kept alive until the larvae hatched 1 day after the collection. The hatching larvae were collected using a Pasteur pipette and then transferred to a tube containing glycerinate alcohol 80% (1:1). The parental female was preserved in alcohol 80%. Both larvae and the female were deposited at the Coleção de Crustáceos do Departamento de Biologia (CCDB), Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto (FFCLRP), Universidade de São Paulo (USP). Genetic sequences of the parental female were generated by Rossi and Mantelatto (2013) and are available in GenBank under accession numbers JQ805902 for the cytochrome c oxidase subunit I (COI) and JQ805867 for the histone 3 (H3) markers (https://www.ncbi.nlm.nih.gov/nuccore/jq805902 and https://www.ncbi.nlm.nih.gov/nuccore/jq805867).
All the following methodological steps were performed at the Laboratório de Bioecologia e Sistemática de Crustáceos (LBSC/FFCLRP/USP). Firstly, semi-permanent slides were prepared to make possible the morphological study and the larval description (Pantaleão et al., 2011). Before mounting the slides, some larvae were colored with methylene blue 1% to facilitate the visualization of some structures. Slide preparation started with the dissection of larvae in glycerin mounted on a slide. Dissection was carried out under a stereomicroscope Leica™ M205 C equipped with a digital camera Leica™ DFC295. Before mounting the coverslip, larval body structures were organized to avoid over-position. Nail base was used to fix the coverslip. Slides were observed under a microscope Zeiss™ AxioScope A.1 equipped with a Zeiss™ AxioCam MRc 5. Photos of the slides were taken using AxioVision Rel. 4.8. These photos were used as a base for the drawings made posteriorly with the computational program Adobe Illustrator version 24.2.1.
The morphological study involved the analysis, the description and the drawing of the entire specimen with all its appendages, with special attention devoted to the number and shape of cuticular outgrowths (e.g., spines, setae). Larval description was based mainly on Clark et al. (1998) and updates by Clark and Cuesta (2015), but other referential studies on larval morphology were also used as guides for morphological analyses, the description and the drawings (Anger, 2001; Bueno & Rodrigues, 1995; Garm, 2004; Melo & Brossi-Garcia, 2005; Pantaleão et al., 2011; Pohle & Telford, 1981). Information about the larval size was obtained by measuring (see Pantaleão et al., 2021) the total length (TL) from the tip of the rostrum to the posterior margin of the telson, and carapace length (CL) from the posterior margin of the ocular orbit to the posterior margin of the carapace using a stereomicroscope equipped with a digital software (Leica Application Suite version 3.8).
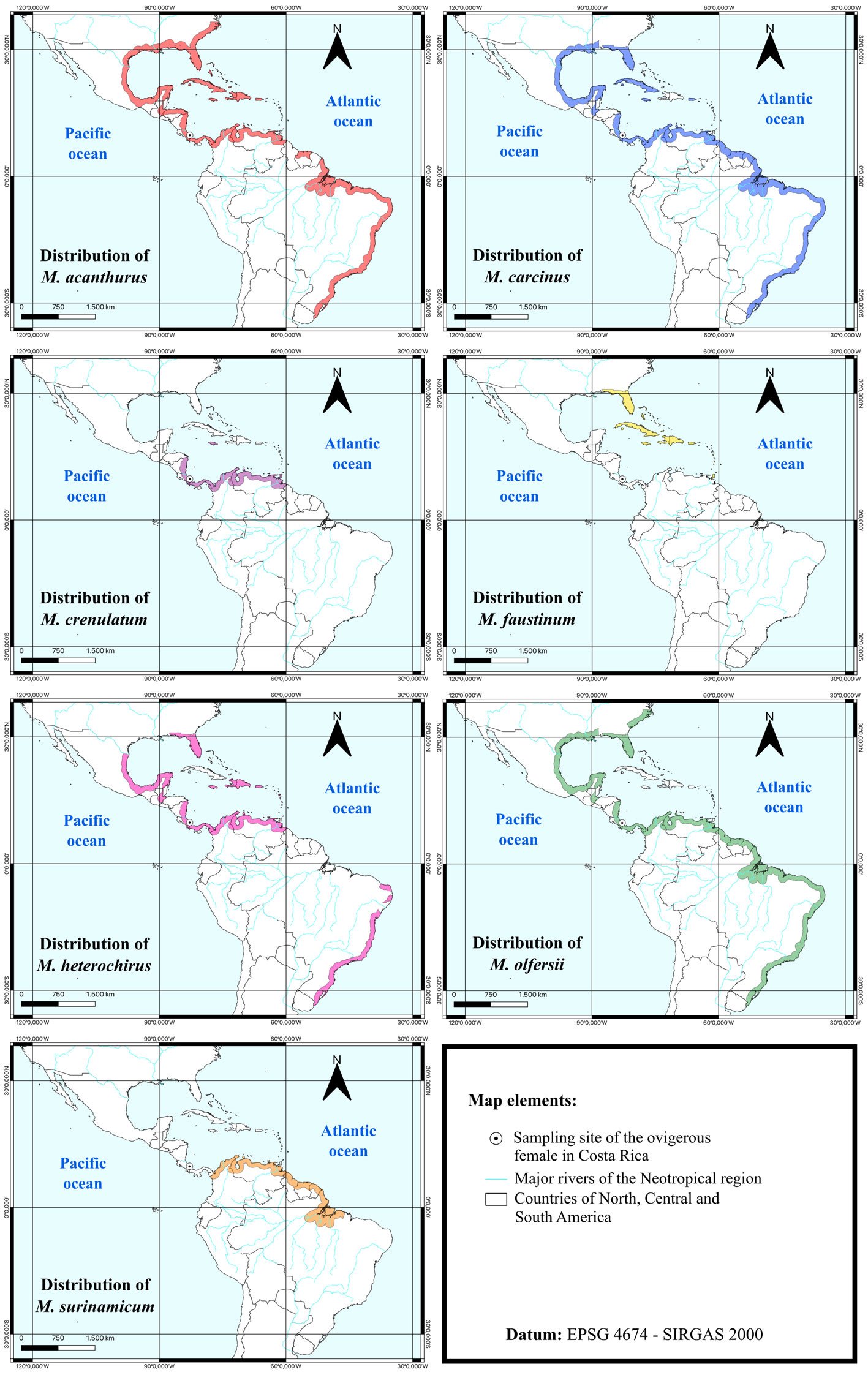
An analysis was carried out to compare the morphology of the first zoea of M. crenulatum in relation to other species of Macrobrachium from the Neotropics that present ELD. We applied 2 criteria for species selection: phylogenetic proximity and similarity in geographic distribution. Considering the phylogenetic hypothesis presented by Pantaleão et al. (2025), our comparison includes M. faustinum (de Saussure, 1857), M. olfersii (Wiegmann, 1836), and M. surinamicum Holthuis, 1948 since all these species are closely related to M. crenulatum. Although M. acanthurus (Wiegmann, 1836), M. heterochirus (Wiegmann, 1836), and M. carcinus are also relatively close to M. crenulatum, their inclusion is particularly relevant because their geographic distributions overlap, at least partially, with that of M. crenulatum, M. olfersii and M. surinamicum. In some cases, this overlap includes freshwater systems in Costa Rica, the country of origin of the larvae examined here, and even cases of sympatry, such as in the Suarez River (Pileggi et al., 2014). Moreover, the inclusion of M. olfersii addresses a particular issue, as this species and M. crenulatum have been considered part of the same species complex (i.e., species complex of M. olfersii) (see Rossi & Mantelatto, 2013). In contrast, some species that are phylogenetically and/or geographically close to M. crenulatum were not included in the comparative analysis because, for several of them (e.g., M. digueti (Bouvier, 1895), M. hancocki Holthuis, 1950, M. hobbsi Nates & Villalobos in Villalobos-Hiriart & Nates-Rodríguez, 1990), no published description of their first zoea is yet available (Pantaleão et al., 2025).
The geographic distributions of all compared species, based on published reports, are presented in Figure 1. Morphological characteristics of Zoea I of all compared species with ELD life cycle were compiled and summarized in Table 1. Additionally, an identification key for the compared species was developed based on morphological traits of the first zoea.
Description
Order Decapoda Latreille, 1802
Suborder Pleocyemata Burkenroad, 1963
Family Palaemonidae Rafinesque, 1815
Genus Macrobrachium Spence Bate, 1868
Macrobrachium crenulatum Holthuis, 1950
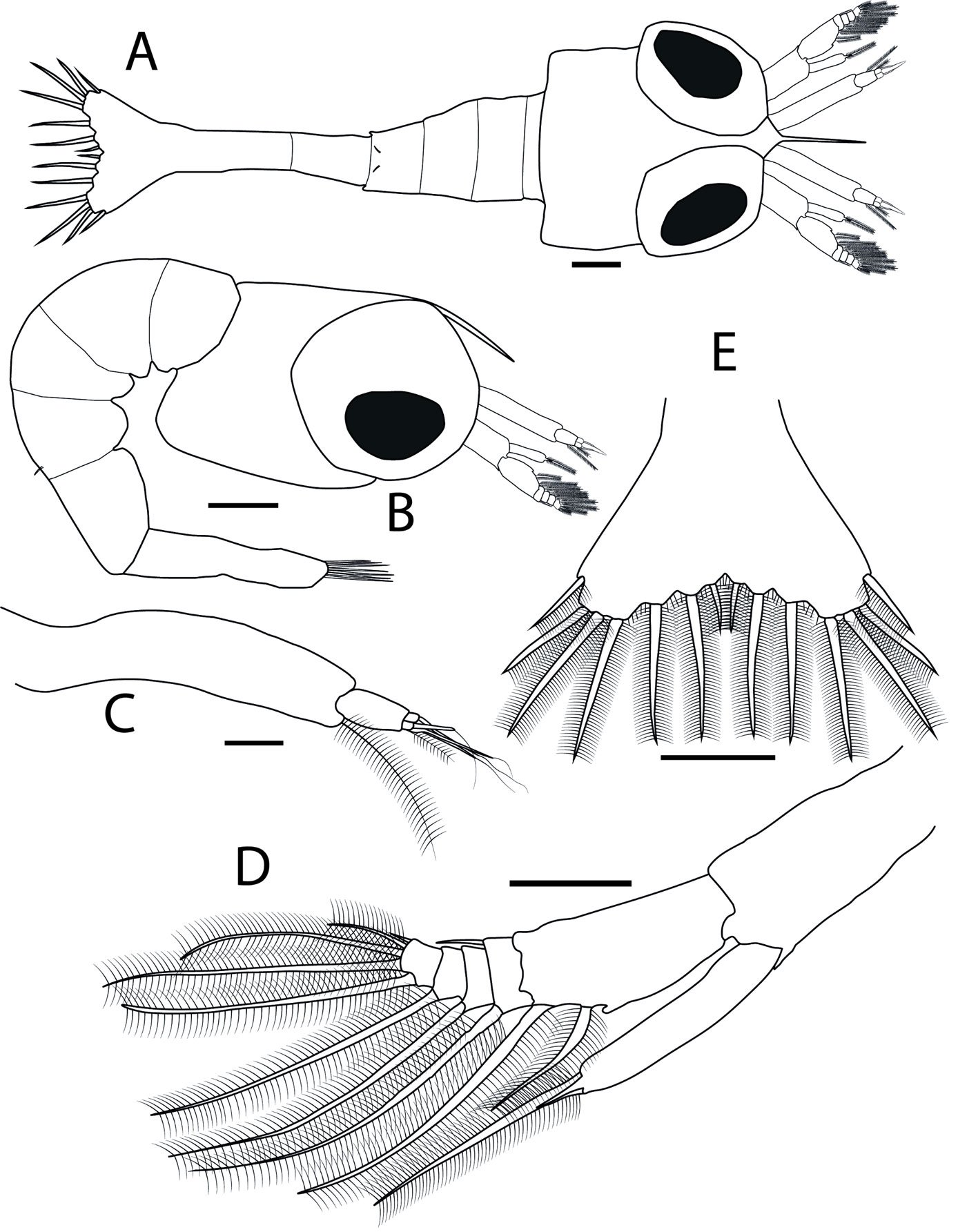
Figures 2A-E, 3A-I
Locality: District of Cahuita, province of Limón, Costa Rica.
Material examined: a parental female and larvae mounted in 11 slides, each one containing one larva dissected (8 slides) or entire larvae (3 slides). Accession number: CCDB 3174.
Dimensions of newly hatched larvae: TL = 2.18 ± 0.12 mm; CL = 0.33 ± 0.02 mm (n = 10). Body (Fig. 2A, B): carapace smooth; rostrum long, curved downwards and with the shape of a spine; sessile eyes; pleon with 5 segments; fourth pleonite with 2 posterodorsal simple setae. Antennule (Fig. 2C): uniramous; peduncle long and unarticled; endopod (or primary flagellum) as a long terminal plumose seta; exopod (or accessory flagellum) with 4 terminal aesthetascs, and 1 short terminal plumose seta. Antenna (Fig. 2D): biramous; protopod unarticled with 1 terminal spine near insertion of endopod; endopod (or flagellum) unarticled with 1 short terminal spine and 1 long terminal plumose seta; exopod (or scaphocerite) 5-articled, first article with 2 plumose setae, 1 terminal and 1 subterminal and 1 medial protuberance, second with 2 setae, 1 inner plumose and 1 outer simple, third with 1 inner long plumose seta, fourth with 1 inner long plumose seta, and fifth with 5 setae, 1 long subterminal plumose, 2 long terminal plumose, 1 short terminal plumose, and 1 short terminal simple.
Table 1
Comparison of meristic and morphological characteristics of Zoea I among different species of Macrobrachium in the Neotropics with Extended Larval Development, adapted from Pantaleão et al. (2011) and Vieira et al. (2017). For multi-articled structures, cuticular outgrowths (plumose setae, simple setae or spines) on the same articles are separated by (+), while those on different articles are separated by comma. Other symbols and abbreviations used: aest. = aesthetascs; s. = setae; si. = simple setae; pl. = plumose setae; sp. = spine; (c.o.) = cuticular outgrowths; (*) = larval description and larval illustration are divergent; (**) = data exclusively based on larval illustration; description not available; (-) = information about the morphological trait unavailable; (?) = description of the appendage unavailable; (;) = data from 2 different larval descriptions.
| Authors | Choudhury (1970) | Choudhury (1971) | Hunte (1980) | Vega-Perez (1984) | Dugger & Dobkin (1975); Melo & Brossi-Garcia (2005) | Vieira et al. (2017) | Present study | |
| Species | M. acanthurus | M. carcinus | M. faustinum | M. heterochirus | M. olfersii | M. surinamicum | M. crenulatum | |
| Locality | Jamaica, Westmoreland | Jamaica, St. Elizabeth | – | Brazil, SP | USA, Florida; Brazil, SP | Brazil, PA | Costa Rica, Limón | |
| Size (mm) | Total length | 2.25-2.35 | 2.0-2.1 | 1.85-1.95 | 2.208 | 1.72-1.77; 1.8-2.05 | 1.55-1.84 | 2.18 |
| Structures | Trait | |||||||
| Antennule | Exopod (c.o.) | 3 aest. + 1 seta + 1 pl. | 3 aest. + 1 si. + 1 pl. | 3 aest. + 1 si. + 1 pl. | 3 aest. + 1 si. + 1 pl. | 4 aest. + 1 pl.; 3 aest. + 1 pl.* | 3 aest. + 1 si. + 1 pl. | 4 aest. + 1 pl. |
| Antenna | Protopod spines | Absent** | Absent | – | 1 | Absent; 1 | Absent | 1 |
| Flagellum (c.o.) | 1 si. + 1 pl. | 1 si. + 1 pl. | 1 si. + 1 pl. | 1 si. + 1 pl. | 2 si. + 1 pl.; 1 si. + 1 pl. | 1 si. + 1 pl. | 1 sp. + 1 pl. | |
| Scaphocerite articles | 4 or 5 | 5 | 4 or 5 | 5* | 5**; 6 | 6 | 5 | |
| Scaphocerite (c.o.) | 10 pl. + 1 si. + 1 seta | 1 sp. + 10 pl. + 1 si. | 10 pl. + 1 seta | 11 pl. + 1 sp. | 1 sp. + 10 pl. + 1 si. | 10 pl. | 1 sp. + 9 pl. + 2 si. | |
| Table 1. Continued | ||||||||
| Authors | Choudhury (1970) | Choudhury (1971) | Hunte (1980) | Vega-Perez (1984) | Dugger & Dobkin (1975); Melo & Brossi-Garcia (2005) | Vieira et al. (2017) | Present study | |
| Mandible | Left mandible (c.o.) | Molar process with 4 or 5 small teeth, incisor process with 3 teeth | Molar process unarmed, incisor process with 4 teeth | Molar process with 2 protuberances, incisor process with 4 teeth | Molar process with 2 teeth, incisor process with 3 teeth | Molar process unarmed, incisor process with 3 teeth; Molar process unarmed, incisor process with 1 tooth and 1 pl. | Incisor process with 2 teeth | 2 sp. |
| Right mandible (c.o.) | Molar process with 2 protuberances, incisor process with 2 teeth | Molar process unarmed, incisor process with 2-3 teeth | Molar process with 2 protuberances, incisor process with 2 teeth | Molar process with 4 teeth, incisor process with 3 teeth | – ; Molar process unarmed, incisor process with 2 teeth and 1 si. | Incisor process with 1 larger tooth and several smaller teeth | Molar process unarmed, incisor process with 3 teeth | |
| Maxilulle | Coxal endite (c.o.) | 4 s. | 4 s. | 4 s. | 4 or 5 si. | 4 sp.; 0 | 4 si. | 4 si. |
| Basial endite (c.o.) | 2 small teeth + 2 large teeth | 2 small teeth + 2 large teeth + 1 seta | 2 small teeth + 2 large teeth | 5 sp. | 4 sp.; 2 si.* | 2 si. | 5 si. | |
| Endopod (c.o.) | 2 sp. | 2 sp. | 2 sp. | 2 si. | 2 sp.; 0 | 1 si. | 2 si. | |
| Maxilla | Coxal endite s. | 4 pl. | 4 pl. | 3 pl. | 4 pl. + 1 si. | 2 si. + 2 pl.; 4 si. | 4 pl. | 1 si. + 3 pl. |
| Basial endite proximal / distal lobe s. | 2 pl. / 2 pl. | 2 pl. + 1 si. / 2 pl. + 1 si. | 2 pl.* / 2 pl. | 2 pl. / 2 pl. | 2 si. / 2 si. | 3 pl. / 2 pl. | 3 si. / 2 si. | |
| Palp proximal / distal lobe s. | 2 s. / 1 pl. | 2 pl. / 1 pl. | 2 s.* / 1 si. | 2 pl. / 1 pl. | 1 si. + 1 pl. / 1 pl.; 2 si. / 1 si. | 2 pl. / 1 pl. | 2 si. / 1 pl. | |
| First maxilliped | Basial s. | 1 seta + 1 seta + 1 seta | 3 si. | 1 seta + 1 seta + 1 seta | 4 or 5 si. | 1 si. | 0 | 1 si. + 2 si. + 1 si. |
| Endopod (c.o.) | 3 s. | 3 sp. | 3 s. | 4 si. | 3 si. + 1 si.; 3 si. | 2 si. + 2 pl. | 3 si. | |
| Exopod articles | Unarticled** | – | Unarticled** | Unarticled | 2 | Unarticled | Unarticled | |
| Exopod s. | 4 pl. | 4 pl. | 4 pl. | 4 pl. | 2 pl. + 4 pl.; 4 pl. | 4 pl. | 4 pl. | |
| Table 1. Continued | ||||||||
| Authors | Choudhury (1970) | Choudhury (1971) | Hunte (1980) | Vega-Perez (1984) | Dugger & Dobkin (1975); Melo & Brossi-Garcia (2005) | Vieira et al. (2017) | Present study | |
| Second maxilliped | Basial s. | 0** | ? | – | 1 si. | 1 si. | 0 | 1 si. |
| Endopod articles | 4 | ? | 4 | 4 | 3 | 3 | 3 | |
| Endopod s. | 0, 0, 1 pl. + 1 seta**, 1 claw + 2 s. | ? | 0, 0, 0, 1 sp. + 2 s. | 0, 0, 2 pl., 2 or 3 si. + 1 pl. | 0, 0, 2 si + 1 si.; 0, 1 pl., 2 si. + 1 pl. | 0, 1 pl., 4 pl. | 0, 1 pl., 4 si + 1 pl. | |
| Exopod articles | Unarticled** | ? | – | Unarticled | 2 | Unarticled | Unarticled | |
| Exopod s. | 4 pl. | ? | – | 4 pl. | 2 pl. + 4 pl.; 4 pl. | 4 pl. | 4 pl. | |
| Third maxilliped | Basial s. | 0** | – | – | 1 si. | 0 | 0 | 0 |
| Endopod articles | 4 | 4 | 4 | 4 | 3 | 3 | 4 | |
| Endopod (c.o.) | 0, 0, 1 seta**, 1 claw + 3 s.* | 0, 0, 0, 1 sp. + 2 or 3 s. | 0, 0, 0, 1 sp. + 2 s.* | 0, 0, 2 or 3 pl., 2 si. + 1 pl. | 0, 0, 3 si. + 2 si. + 1 pl.; 0, 1 si. + 1 pl.; 1 si. + 1 pl. | 2 pl., 2 pl., 3 pl. | 1 si., 1 si., 2 pl., 1 si + 2 pl. | |
| Exopod articles | Unarticled** | Unarticled | – | Unarticled** | 2 | Unarticled | Unarticled | |
| Exopod s. | 4 pl. | 4 pl. | – | 4 pl.** | 2 pl. + 4 pl. | 4 pl. | 4 pl. | |
| Telson | Distal margin (c.o.) | 7 + 7 pl., many small sp. between s. | 7 + 7 pl., small sp. between 6 + 6 inner s. | 7 + 7 pl., many small sp. between s.* | 7 + 7 pl., small si. between pl. | 7 + 7 plumose sp.; 1 + 1 si. (the innermost pair without setulae) and 6 + 6 pl. | 7 + 7 pl., minute setae between 3 + 3 inner setae | 7 + 7 pl., small sp. between 3 + 3 inner setae |
| Last pair of (c.o.) (most lateral) | With setulae only on inner margin | With setulae only on inner margin | With setulae only on inner margin** | With setulae only on inner margin | With setulae only on inner margin | With setulae on inner and outer margins | With setulae only on inner margin |
Mandibles (Fig. 3A, B): both rudimentary, lacking palp; right mandible with molar process smooth, and incisor process with 2 subterminal teeth and 1 terminal sharp tooth of varying sizes; left mandible with molar and incisor process not clearly defined, bearing 2 terminal spines, 1 smaller and 1 larger. Maxillule (Fig. 3C): coxal and basial endites with 4 and 5 terminal simple setae, respectively; endopod with 2 terminal simple setae. Maxilla (Fig. 3D): biramous; coxal endite with 4 setae, 1 simple and 3 plumose; basial endite bilobed, with 3 and 2 simple setae on proximal and distal lobes, respectively; endopod (or palp) bilobed, with 2 proximal simple and 1 terminal plumose seta; exopod (or scaphognathite) margin with 4 proximal plumose setae and 1 long terminal plumose process; microtrichia on margins of endopod and exopod.
First maxilliped (Fig. 3E): biramous; coxa without setae; basis with 4 short simple setae arranged 1 + 2 + 1; endopod unarticled with 3 terminal simple setae, 1 short and 2 long; exopod unarticled, with 4 long terminal plumose natatory setae; endopod approximately 3 times shorter than exopod. Second maxilliped (Fig. 3F): biramous; coxa without setae; basis with 1 short simple seta; endopod 3-articled, first with 0, second with 1 terminal plumose, and third with 5, 1 short proximal simple, 3 short terminal simple and 1 short terminal sparsely plumose setae, respectively; exopod unarticled with 4 long terminal plumose natatory setae; endopod slightly shorter than exopod.
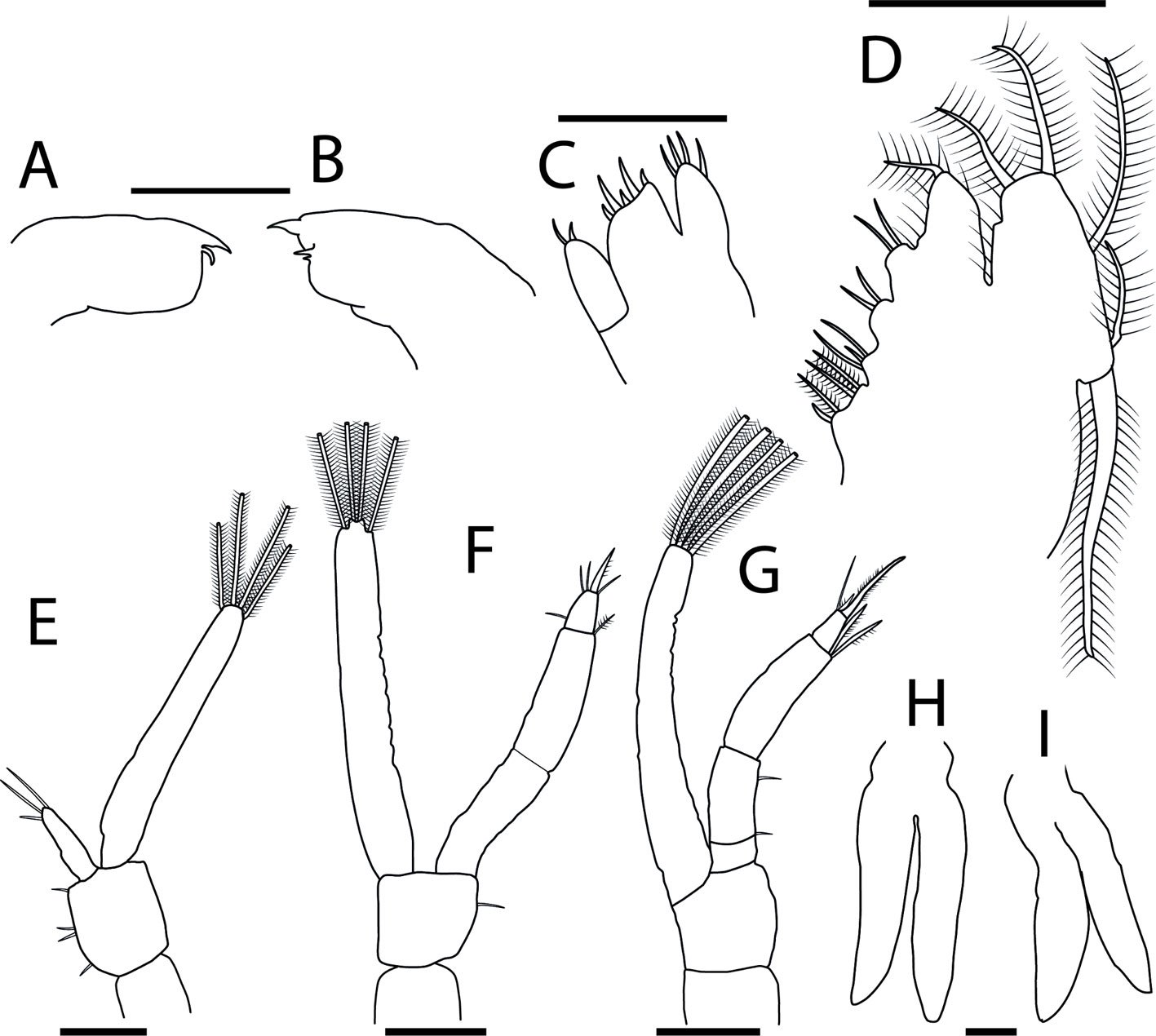
Third maxilliped (Fig. 3G): biramous; coxa and basis without setae; endopod 4-articled, first article without setae, second with 1 short proximal simple, and 1 short subterminal simple, third with 2 long terminal plumose, and fourth with 3 terminal setae, 1 long simple, 1 short plumose and 1 long plumose; exopod unarticled with 4 long terminal plumose natatory setae; endopod and exopod of similar length. Pereiopods (Fig. 3H, I): first and second rudimentary and biramous, second slightly longer than the first; third, fourth and fifth absent. Telson (Fig. 2E): posterior bilobed region broader than the anterior, with 7 + 7 plumose setae, outer 2 plumose only in inner margin; minute spinules along the posterior margin and around the bases of 3 + 3 inner setae.
Remarks
The morphological description of Zoea I of Macrobrachium crenulatum enables the comparison among the first zoeae of closely related congeners as well as the definition of some morphological characteristics. All the species included in the comparison exhibit type I development, characterized by an extended larval development and an amphidromous life cycle (Bauer, 2013). This contrasts with type II development, which involves a partially abbreviated larval development and is common in freshwater species, and type III development, which is marked by a completely abbreviated larval development and is typical of species inhabiting mountain streams (Jalihal et al., 1993; Pileggi et al., 2014; Vieira et al., 2017). Adults of amphidromous species typically inhabit both freshwater and brackish environments, except for M. crenulatum, M. faustinum, and M. heterochirus, whose adults have been reported exclusively in freshwater (Holthuis, 1950; Hunte, 1980; Vega-Perez, 1984; Vieira et al., 2017).
Before comparing the morphology of the Zoea I among species, it is important to consider that most of the larval descriptions included in Table 1 were published at different times, each reflecting the research context on decapod larval morphology of that period. Thus, many of the current consensuses on terminology (e.g., Clark et al., 1998) postdate some of the descriptions examined. Moreover, recent studies employ more advanced technologies than those available in the 1970s, 80s and 90s, enabling, for example, more accurate and detailed descriptions of setal types and number of articles. Therefore, some inconsistencies were detected during the comparisons of the published descriptions. Thus, we aimed to distinguish true morphological differences of the larvae from inconsistencies likely resulting from the limited technological capabilities used in earlier studies. Although we kept the original terminology used by the authors in our table, in the following comparison we focus on taxonomic informative differences, highlighting instances where a variation may result from misinterpretation.
Macrobrachium crenulatum and M. olfersii have been considered part of the same species complex due to the high morphological similarity of adults (Rossi & Mantelatto, 2013; Villalobos-Figueroa, 1967). Their close phylogenetic relationship was later confirmed using molecular data by Pileggi et al. (2014) and, more recently, by Pantaleão et al. (2025). For M. olfersii, 2 larval descriptions of first zoea are available. Dugger and Dobkin (1975) provided the first, describing Zoea I of this species from females collected in Florida (USA). Later, Melo and Brossi-Garcia (2005) provided a description for hatched larvae of Macrobrachium birai Lobão, Melo & Fernandes, 1986, obtained from a female collected in São Paulo (Brazil). Subsequently, however, M. birai was regarded as a junior synonymous of M. olfersii by Pileggi and Mantelatto (2012). Thus, both studies are treated here as descriptions of the first zoeal stage of M. olfersii. Although generally similar, both descriptions show some differences in the number of cuticular outgrowths on the antennal flagellum, maxilla coxal endite, and maxilliped endopods. Considering that each study worked with larvae from distinct latitudes (USA and Brazil) with different temperature regimes and oceanographic dynamics, these differences may reflect plasticity in larval morphology, a phenomenon that has partially explored in the scientific literature (Sandifer & Smith, 1979; Thatje & Bacardit, 2000; Wehrtmann & Albornoz, 2003). For the comparison between M. crenulatum and M. olfersii, we focused only on features that are consistent across both descriptions of M. olfersii, avoiding traits that showed intraspecific variation. In Table 1, when differences are observed in the description of Zoea I of M. olfersii, data from both descriptions are presented (Dugger & Dobkin, 1975; Melo & Brossi-Garcia, 2005).
The maxillipeds are considerably distinct between M. crenulatum and M. olfersii. They differ by the number of plumose setae on the third maxilliped exopod: Macrobrachium crenulatum presents only 4 terminal plumose setae, while the first zoea of M. olfersii has 2 subterminal plumose setae in addition to the 4 terminal ones. Moreover, the basis of the first maxilliped bears one simple seta in M. olfersii, but 4 (1 + 2 + 1) in M. crenulatum. Finally, it is important to mention that the larval description published by Dugger and Dobkin (1975) reports a difference in the number of articles on the exopod of maxillipeds: 2 in M. olfersii versus only one in M. crenulatum. However, aside from being absent in the description from Melo and Brossi-Garcia (2005), this apparent difference may result from a misinterpretation of the number of articles in maxilliped exopods of M. olfersii, since the presence of only 1 article appears to be a consistent pattern among American species of Macrobrachium, regardless of whether they exhibit type I development (e.g., M. carcinus, M. heterochirus and M. surinamicum) or type II (e.g., Macrobrachium iheringi [Ortmann, 1897] and Macrobrachium jelskii [Miers, 1877]) (Bueno & Rodrigues, 1995; Choudhury, 1970, 1971; Magalhães, 2000; Pantaleão et al., 2025; Vega-Perez, 1984; Vieira et al., 2017).
Several morphological features can be used to distinguish the first zoeae of M. crenulatum and M. carcinus, 2 species whose geographic distributions partially overlap. Notably, these 2 species differ in the pattern of setae present on the endopod of the third maxilliped: M. crenulatum has setae on all 4 articles of the third maxilliped endopod (1, 1, 2, 1 + 2), while Zoea I of M. carcinus presents setae only on the last article (0, 0, 0, 1 + 2 or 3) (Choudhury, 1971). Analysis of the larval description of M. carcinus reveals possible inconsistencies in terminology. In several instances, the term “spine” is used to refer to a “seta” and vice-versa. Additionally, although illustrated, the second maxilliped of the first zoea of M. carcinus was not formally described, being mentioned only briefly within the description of the third maxilliped. These inconsistencies hinder a comprehensive morphological comparison and complicate phylogenetic inferences based on larval characters. Therefore, given the aforementioned issues related to technological advances and terminology standardization, we strongly recommend a redescription of the first zoeal stage of M. carcinus.
Macrobrachium heterochirus is a species closely related to M. crenulatum both geographically and phylogenetically (Pantaleão et al., 2025; Pileggi et al., 2014), and the adults are morphologically distinct enough to be readily distinguished. Zoea I of both species, in turn, differ on the setal pattern of maxilla, both in the number and type of setae on coxal and basial endites: Macrobrachium crenulatum has 4 setae, one simple and 3 plumose, on the coxal endite and 5 simple setae on the basial endite, whereas the first zoea of M. heterochirus bears 5 setae, 4 plumose and one simple, on the coxal endite, and the basial endite carries only 4 plumose setae. According to the descriptions, these species also differ in the number of aesthetascs on the antennule exopod: 4 in M. crenulatum, and 3 in M. heterochirus (Vega-Perez, 1984). However, this difference may reflect a misinterpretation. Possibly, the simple seta described by the author as part of antennule exopod of M. heterochirus could in fact be an aesthetasc, bringing the total to 4, as observed in other species such as M. crenulatum and M. olfersii. Similar misunderstandings regarding the number of aesthetascs of the antennule may also applied to the descriptions of M. acanthurus, M. carcinus, M. faustinum, and M. surinamicum.
Several morphological characteristics allow the distinction between Zoea I of M. crenulatum and M. acanthurus, both species occurring in Costa Rica (Fig. 1). One of the most conspicuous features for identification is the number of aesthetascs on the antennule (4 on the first and only 3 on the latter), although this difference may reflect a misinterpretation in the earliest description, which dates back 56 years (Choudhury, 1970). Additionally, M. crenulatum has a spine on the protopod of the antenna, which is absent in M. acanthurus. Another notable difference lies in the number of endopod articles of the second maxilliped: Macrobrachium crenulatum has 3, whereas M. acanthurus presents 4, and the setal pattern on this structure also differs (Table 1). Finally, the telson of M. crenulatum bears small spines between the 3 innermost plumose setae, while in M. acanthurus, spines are present between all the setae of the telson.
As with M. acanthurus, several traits can be used to distinguish Zoea I of M. crenulatum and M. faustinum (Table 1), species that are relatively close phylogenetically (Pantaleão et al., 2025). Notably, the number of aesthetascs present in the antennule differs, with M. crenulatum having 4 and M. faustinum only 3. As previously noted, this feature should be interpreted cautiously, as the older description may not be fully accurate in this regard. Additionally, the scaphocerite of M. faustinum lacks a spine present in the first larval stage of M. crenulatum, and the molar process of the right mandible in M. crenulatum is unarmed, whereas in M. faustinum it bears 2 protuberances (Hunte, 1980).
Finally, several larval characteristics can also be used to identify the first zoeae of M. crenulatum and M. surinamicum, species relatively closely related according to Pantaleão et al. (2025). The differences between the newly hatched larvae refer to the number of articles on the scaphocerite (5 in M. crenulatum and 6 in M. surinamicum), the number of spines on basial endite of maxillule (5 in M. crenulatum and 2 in M. surinamicum), and the position of setulae on the outer pair of setae of telson margin. In M. crenulatum, setulae occur only on inner margin of the outer pair of setae of the distal margin of the telson, whereas in M. surinamicum, they are present on both the inner and outer margins (Table 1). Interestingly, none of the other species with described Zoea I included in Table 1 have setulae on the outer margin of this pair of setae. Therefore, the phylogenetic significance of this trait warrants to be analyzed in future studies.
The present study represents a first step toward improving our knowledge of larval morphology of M. crenulatum. Although it focuses only on the first larval stage of a single brood of Macrobrachium species, the results contribute to a broader effort of providing larval descriptions for Neotropical species of the genus. This information offers valuable insights for both theoretical studies about the evolution of the morphology of Macrobrachium and practical applications in larval ecology and aquaculture research focused on larval development. Additionally, the investigation of the larval morphology of M. crenulatum helps to fill a gap in the larval descriptions of a lineage of species of Macrobrachium for which no larval data were previously available (Pantaleão et al., 2025). This line of research should be continued by describing the larvae of other species phylogenetically close to M. crenulatum, such as M. digueti and M. hancocki, to broaden our understanding of phylogenetic relationships among these freshwater shrimps. Finally, our results highlight the need for redescriptions of the first zoea of several species of Macrobrachium, aimed at updating the larval terminology and clarifying ambiguous characters, which would in turn facilitate morphological comparisons and phylogenetic analysis.
Identification key for the compared species of Macrobrachium with ELD from Neotropics based on morphological traits of Zoea I
1. First article of third maxilliped endopod with setae 2
1’. First article of third maxilliped endopod without setae 3
2. Scaphocerite with 5 articles; basial endite of maxillule with 5 spines; most lateral pair of setae of telson margin with setulae only on inner margin M. crenulatum
2’. Scaphocerite with 6 articles; basial endite of maxillule with 2 spines; most lateral pair of setae of telson margin with setulae both on inner and outer margins M. surinamicum
3. Basial endite of first maxilliped with more than 1 seta; endopod of third maxilliped with 4 articles 4
3’. Basial endite of first maxilliped with only 1 seta; endopod of third maxilliped with 3 articles M. olfersii
4. Scaphocerite with 10 plumose setae; first maxilliped endopod with 3 cuticular outgrowths 5
4’. Scaphocerite with 11 plumose setae; first maxilliped endopod with 4 cuticular outgrowths M. heterochirus
5. Scaphocerite with 12 cuticular outgrowths; maxilla coxal endite with 4 plumose setae 6
5’. Scaphocerite with 11 cuticular outgrowths; maxilla coxal endite with 3 plumose setae M. faustinum
6. Lobes of maxilla basial endite with 2 plumose setae each M. acanthurus
6’. Lobes of maxilla basial endite with 2 plumose setae and 1 simple seta each M. carcinus
Acknowledgements
This study is result of the course “Special Topics in Comparative Biology: Larval morphology and phylogenetic relationships in decapod crustaceans” under the responsibility of FLM and JAFP and as part of Postgraduate Program in Comparative Biology at Faculty of Philosophy, Sciences and Letters at Ribeirão Preto. The authors are grateful to the LBSC team, in special to Gabriel L. Bochini, who helped with some dissections and slide preparation. We acknowledge the financial support provided by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (Projetos Temáticos BIOTA Proc. 2010/50188-8 and INTERCRUSTA Proc. 2018/13685-5) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (PPBio 07/2023 – Linha 8: Rede Costeira Marinha Proc. 442421/2023-0). FLM and ISW are thankful for the financial contribution supplied by CNPq (Proc. 491490/2004-6, 490353/2007-0, 490314/2011-2; 471011/2011-8) and CONICIT – Costa Rica (CII- 001-08, IQ-0001-11) during the development of the International Cooperative Project, which was crucial for the collection of the female with embryos and collaboration in the Brazil-Costa Rica research program. FLM thanks CNPq for an ongoing research grant (PQ 302706/2025-9). LOR and RP thank PhD grants provided by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (PROEX – Proc. 88887.804708/2023-00 and 88887.896987/2023-00, respectively). RP and LOR also thank the ongoing PhD fellowships by FAPESP (Proc. 2024/15232-9 and 2025/01018-8, respectively). The ovigerous female was collected under the collection and genetic access permit (Resolución No. 377 – Comisión Institucional de Biodiversidad de la Universidad de Costa Rica) to ISW. We also thank the anonymous reviewers and associate editor for their valuable comments, which improved the quality of the manuscript.
References
Almeida, A. O., Alves-Coelho, P., Rocha-Luz, J., Almeida-dos Santos, J. T., & Ribeiro-Ferraz, N. (2008). Decapod crustaceans in fresh waters of southeastern Bahia, Brazil. Revista de Biología Tropical, 56, 1225–1254. https://doi.org/10.15517/rbt.v56i3.5706
Anger, K. (2001). The biology of decapod crustacean larvae. Lisse, South Holland: AA Balkema Publishers.
Anger, K. (2013). Neotropical Macrobrachium (Caridea: Palaemonidae): on the biology, origin, and radiation of freshwater-invading shrimp. Journal of Crustacean Biology, 33, 151–183. https://doi.org/10.1163/1937240X-00002124
Bauer, R. T. (2011a). Amphidromy and migrations of freshwater shrimps. I. Costs, benefits, evolutionary origins, and an unusual case of amphidromy. New Frontiers in Crustacean Biology, 15, 145–156. https://doi.org/10.1163/ej.9789004174252.i-354.108
Bauer, R. T. (2011b). Amphidromy and migrations of freshwater shrimps. II. Delivery of hatching larvae to the sea, return juvenile upstream migration, and human impacts. New Frontiers in Crustacean Biology, 15, 157–168. https://doi.org/10.1163/EJ.9789004174252.I-354.115
Bauer, R. T. (2013). Amphidromy in shrimps: a life cycle between rivers and the sea. Latin American Journal of Aquatic Research, 41, 633–650. https://doi.org/10.3856/vol41-issue4-fulltext-2
Bouvier, E. L., (1895). Sur les palémons receuillis dans les eaux douces de la Basse-Californie par M. Diguet. Bulletin du Muséum d’Histoire Naturelle, 1, 159–162.
Bowles, D. E., Aziz, K., & Knight, C. L. (2000). Macrobrachium (Decapoda: Caridea: Palaemonidae) in the contiguous United States: a review of the species and an assessment of threats to their survival. Journal of Crustacean Biology, 20, 158–171. https://doi.org/10.1163/20021975-99990025
Bueno, S. L. S., & Rodrigues, S. A. (1995). Abbreviated larval development of the freshwater prawn, Macrobrachium iheringi (Ortmann, 1897) (Decapoda, Palaemonidae), reared in the laboratory. Crustaceana, 68, 665–686. https://doi.org/10.1163/156854095X01880
Burkenroad, M. D. (1963). The evolution of the Eucarida (Crustacea, Eumalacostraca), in relation to the fossil record. Tulane Studies in Geology, 2, 3–17.
Clark, P. F., Calazans, D. K., & Pohle, G. W. (1998). Accuracy and standardization of brachyuran larval descriptions. Invertebrate Reproduction & Development, 33, 127–144. https://doi.org/10.1080/07924259.1998.9652627
Clark, P. F., & Cuesta, J. A. (2015). Larval systematics of Brachyura. In P. Castro, P. J. F. Davie, D. Guinot, F. R. Schram, & J. C. von Vaupel-Klein (Eds.), Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea 9C (pp. 981–1048). Leiden, South Holland: Brill. https://doi.org/10.1163/9789004190832_020
Chace, F. A., & Bruce, A. J. (1993). The caridean shrimps (Crustacea: Decapoda) of the Albatross Philippine expedition 1907-1910, part 6: superfamily Palaemonoidea. Smithsonian Contributions to Zoology, 543, 1–152. https://doi.org/10.5479/si.00810282.543
Choudhury, P. C. (1970). Complete larval development of the palaemonid shrimp Macrobrachium acanthurus (Wiegmann, 1836), reared in the laboratory. Crustaceana, 18, 113–132. https://doi.org/10.1163/156854070X00743
Choudhury, P. C. (1971). Complete larval development of the palaemonid shrimp Macrobrachium carcinus (L.), reared in the laboratory (Decapoda, Palaemonidae). Crustaceana, 20, 51–69. https://doi.org/10.1163/156854071X00526
De Grave, S., & Fransen, C. H. J. M. (2011). Carideorum catalogus: the recent species of the dendrobranchiate, stenopodidean, procarididean and caridean shrimps (Crustacea: Decapoda). Zoologische Mededelingen, 85, 195–589.
de Saussure, H. (1857). Diagnoses de quelques Crustacés nouveaux de l’Amérique tropicale. Revue et Magasin de Zoologie, Ser. 2, 9, 501–505.
Dugger, D. M., & Dobkin, S. (1975). A contribution to knowledge of the larval development of Macrobrachium olfersii (Wiegmann, 1836) (Decapoda, Palaemonidae). Crustaceana, 29, 1–30. https://doi.org/10.1163/156854075×00018
Ferreira, R. S., Vieira, R. R. R., & D’Incao, F. (2010). The marine and estuarine shrimps of the Palaemoninae (Crustacea: Decapoda: Caridea) from Brazil. Zootaxa, 2606, 1–24. https://doi.org/10.11646/zootaxa.2606.1.1
Fujita, Y., Davie, P. J., & Ng, P. K. (2015). A new stygobitic prawn of the genus Macrobrachium Spence Bate, 1864, from anchialine caves in Christmas Island, Indian Ocean; with a rediagnosis of M. miyakoense Komai & Fujita, 2005 (Crustacea: Decapoda: Caridea: Palaemonidae). Raffles Bulletin of Zoology, 63, 610–625.
Garm, A. (2004). Revising the definition of the crustacean seta and setal classification systems based on examination of the mouthpart setae of seven species of decapods. Zoological Journal of the Linnean Society, 142, 233–252. https://doi.org/10.1111/j.1096-3642.2004.00132.x
Holthuis, L. B. (1948). Note on some Crustacea Decapod Natantia from Surinam. Proceedings van de Koninklijke Nederlandsche Akademie van Wetenschappen, 51, 1104–1113.
Holthuis, L. B. (1950). Preliminary descriptions of twelve new species of palaemonid prawns from American waters (Crustacea Decapoda). Proceedings van de Koninklijke Nederlandsche Akademie van Wetenschappen, 53, 93–99.
Holthuis, L. B. (1952). A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. II. The subfamily Palaemonidae. Occasional Papers of the Allan Hancock Foundation, 12, 1–396.
Hunte, W. (1979). The rediscovery of the freshwater shrimp Macrobrachium crenulatum in Jamaica. Studies on the Fauna of Curaçao and other Caribbean Islands, 58, 69–74.
Hunte, W. (1980). The larval development of the shrimp Macrobrachium faustinum reared at the laboratory. Caribbean Journal of Science, 15, 49–68.
Jalihal, D. R., Sankolli, K. N., & Shenoy, S. (1993). Evolution of larval developmental patterns and the process of freshwaterization in the prawn genus Macrobrachium Bate, 1868 (Decapoda, Palaemonidae). Crustaceana, 65, 365–376. https://doi.org/10.1163/156854093X00793
Lai, J. C. Y., Thoma, B. P., Clark, P. F., Felder, D. L., & Ng, P. K. L. (2013). Phylogeny of eriphioid crabs (Brachyura, Eriphioidea) inferred from molecular and morphological studies. Zoologica Scripta, 43, 52–64. https://doi.org/10.1111/zsc.12030
Lara, R., & Wehrtmann, I. S. (2011). Diversity, abundance and distribution of river shrimps (Decapoda: Caridea) in the largest river basin of Costa Rica, Central America. In A. Asakura & C. Fransen (Eds.), New Frontiers in Crustacean Biology (pp. 197–211). Leiden, South Holland: Brill. http://dx.doi.org/10.1163/ej.9789004174252.i-354.140
Latreille, P. A. (1802). Histoire naturelle, générale et particulière des crustacés et des insectes. Familles naturelles des genre. Ouvrage faisant suite à l‘Histoire Naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieurs Sociétés savantes. de L‘imprimerie de F. Dufart, Paris.
Liu, M. Y., Cai, Y. X., & Tzeng, C. S. (2007). Molecular systematics of the freshwater prawn genus Macrobrachium Bate, 1868 (Crustacea: Decapoda: Palaemonidae) inferred from mtDNA sequences, with emphasis on East Asian species. Zoological Studies, 46, 272–289.
Lobão, V. L., Melo, G. A. S., & Fernandes, W. (1986). Descrição de uma nova espécie do gênero Macrobrachium (Crustacea: Decapoda: Palaemonidae) da região sul de São Paulo. In Resumos do Congresso Brasileiro de Zoologia, Volume 13. Cuiabá.
Magalhães, C. (2000). Abbreviated larval development of Macrobrachium jelskii (Miers, 1877) (Crustacea: Decapoda, Palaemonidae) from the Rio Solimões floodplain, Brazil, reared in the laboratory. Nauplius, 8, 1–14.
Magalhães, C., & Pereira, G. (2007). Assessment of the decapod crustacean diversity in the Guayana Shield region aiming at conservation decisions. Biota Neotropica, 7, 111–124. https://doi.org/10.1590/s1676-06032007000200013
Melo, S. G., & Brossi-Garcia, A. L. (2005). Desenvolvimento larval de Macrobrachium birai Lobão, Melo & Fernandes (Crustacea, Decapoda, Caridea, Palaemonidae) em laboratório. Revista Brasileira de Zoologia, 22, 131–152. https://doi.org/10.1590/S0101-81752005000100016
Miers, E. J. (1877). On a collection of Crustacea, Decapoda and Isopoda, chiefly from South America, with descriptions of new genera and species. Proceedings of the Zoological Society of London, 43, 653–679.
Murienne, J., Chevalier, J., & Clavier, S. (2022). On the presence of the giant freshwater prawn, Macrobrachium rosenbergii, in French Guiana confirmed by citizen science and genetic analyses. Water Biology and Security, 1, 100039. https://doi.org/10.1016/j.watbs.2022.100039
Murphy, N. P., & Austin, C. M. (2005). Phylogenetic relationships of the globally distributed freshwater prawn genus Macrobrachium (Crustacea: Decapoda: Palaemonidae): biogeography, taxonomy and the convergent evolution of abbreviated larval development. Zoologica Scripta, 34, 187–197. https://doi.org/10.1111/j.1463-6409.2005.00185.x
Ortmann, A. (1897). Os camarões da agua doce da America do Sul. Revista do Museu Paulista, 2, 173–216.
Pantaleão, J. A. F., Gregati, R. A., Taddei, F. G., & Costa, R. C. (2011). Morphology of the first larval stage of Macrobrachium brasiliense (Heller, 1868) (Caridea: Palaemonidae). Nauplius, 19, 79–85. https://doi.org/10.1590/S0104-64972011000100009
Pantaleão, J. A. F., Mantelatto, F. L., & Costa, R. C. (2021). The first larval stage (Zoea I) description of the caridean shrimp Ogyrides occidentalis (Ortmann, 1893) (Alpheoidea: Ogyrididae) reveals congruence with taxonomic status. Biota Neotropica, 21, e20201118. https://doi.org/10.1590/1676-0611-BN-2020-1118
Pantaleão, J. A. F., Peres, P. A., & Mantelatto, F. L. (2025). Mapping the congruence between larval morphology and molecular phylogeny: Convergent adaptations of fresh-
water shrimps (Decapoda, Caridea). Zoologica Scripta, 54, 376–393. https://doi.org/10.1111/zsc.12709
Pileggi, L. G., & Mantelatto, F. L. (2010). Molecular phylogeny of the freshwater prawn genus Macrobrachium (Decapoda, Palaemonidae), with emphasis on the relationships among selected American species. Invertebrate Systematics, 24, 194–208. https://doi.org/10.1071/IS09043
Pileggi L. G., & Mantelatto, F. L. (2012). Taxonomic revision of doubtful Brazilian freshwater shrimp species of genus Macrobrachium (Decapoda, Palaemonidae). Iheringia, Série Zoologia, 102, 426–437. https://doi.org/10.1590/S0073-47212012005000012
Pileggi, L. G., Rossi, N., Wehrtmann, I. S., & Mantelatto, F. L. (2014). Molecular perspective on the American transisthmian species of Macrobrachium (Caridea, Palaemonidae). Zookeys, 457, 109–131. https://doi.org/10.3897/zookeys.457.6818
Pohle, G. W., & Telford, M. (1981). Morphology and classification of decapod crustacean larval cerdae: a scanning electron microscope study of Dissodactylus crinitichelis Moreira, 1901 (Brachyura: Pinnotheridae). Bulletin of Marine Science, 31, 736–752.
Pompeu, P. D. S., Vieira, F., & Martinez, C. B. (2006). Utilização do mecanismo de transposição de peixes da Usina Hidrelétrica Santa Clara por camarões (Palaemonidae), bacia do rio Mucuri, Minas Gerais, Brasil. Revista Brasileira de Zoologia, 23, 293–297. https://doi.org/10.1590/s0101-81752006000100024
Poore, G. C., & Ahyong, S. T. (2023). Marine decapod Crustacea: a guide to families and genera of the world. Clayton, Victoria: CSIRO Publishing. https://doi.org/10.1071/9781486311798
Rafinesque, C. S. (1815). Analyse de la nature ou Tableau de l’univers et des corps organisés. Palermo: Published by the author.
Rossi, N. (2012). Revisão das espécies de Macrobrachium Bate, 1868, pertencentes ao complexo M. olfersii (Crustacea, Palaemonidae): análises morfológicas e moleculares (M.Sc. Thesis). Universidade de São Paulo. Brazil, São Paulo.
Rossi, N., & Mantelatto, F. L. (2013). Molecular analysis of the freshwater prawn Macrobrachium olfersii (Decapoda, Palaemonidae) supports the existence of a single species throughout its distribution. Plos One, 8, e54698. https://doi.org/10.1371/journal.pone.0054698
Sandifer, P. A., & Smith, T. I. (1979). Possible significance of variation in the larval development of palaemonid shrimp. Journal of Experimental Marine Biology and Ecology, 39, 55–64. https://doi.org/10.1016/0022-0981(79)90004-2
Sastry, A. N. (1983). Ecological aspects of reproduction. In F. J. Vernberg, & W. B. Vernberg (Eds.), The biology of Crustacea: environmental adaptations, Volume 8 (pp. 179–270). New York: Academic Press.
Shokita, S. (1985). Larval development of the palaemonid prawn, Macrobrachium grandimanus (Randall), reared in the laboratory, with special reference to larval dispersal. Zoological Science, 2, 785–803.
Spence Bate, C. (1868). On a new genus, with four new species, of freshwater prawns. Proceedings of the Zoological Society of London, 363–368.
Tejeda-Mazariegos, J. C., Ortiz, L. M. M., López-Mejía, M., Crandall, K. A., Pérez-Losada, M., & Frausto-Martínez, O. (2018). Freshwater crustacean decapods: an important resource of Guatemala. In S. Ray (Ed.), Biological resources of water (pp. 169–180). London: IntechOpen. https://doi.org/10.5772/intechopen.73638
Thatje, S., & Bacardit, R. (2000). Morphological variability in larval stages of Nauticaris magellanica (A. Milne Edwards, 1891) (Decapoda: Caridea: Hippolytidae) from South American waters. Bulletin of Marine Science, 66, 375–398.
Valencia, D. M., & Campos, M. R. (2007). Freshwater prawns of the genus Macrobrachium Bate, 1868 (Crustacea: Decapoda: Palaemonidae) of Colombia. Zootaxa, 1456, 1–44. https://doi.org/10.11646/zootaxa.1456.1.1
Vega-Perez, L. A. (1984). Desenvolvimento larval de Macrobrachium heterochirus (Wiegmann, 1836), Macrobrachium amazonicum (Heller, 1862) e Macrobrachium brasiliense (Heller, 1868) (Crustacea, Decapoda, Palaemonidae), em laboratório (Ph.D. Thesis). Universidade de São Paulo. Brazil, São Paulo.
Vieira, R. R. R., Santos, C. R. M., & Anker, A. (2017). Morphology of the first zoeal stage of Macrobrachium surinamicum Holthuis, 1948 (Malacostraca, Decapoda, Palaemonidae) hatched in the laboratory. Crustaceana, 90, 1337–1350. https://doi.org/10.1163/15685403-00003723
Villalobos-Figueroa, A. (1967). Problemas de especiación en América de un grupo de Palaemonidae del género Macrobrachium. FAO Fisheries Report, 57, 1055–1066.
Villalobos-Hiriart, J. L., & Nates-Rodríguez, J. C. (1990). Dos especies nuevas de camarones de agua dulce del género Macrobrachium Bate, (Crustacea, Decapoda, Palaemonidae), de la vertiente occidental de México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 61, 1–11.
Wehrtmann, I. S., & Albornoz, L. (2003). Larvae of Nauticaris magellanica (Decapoda: Caridea: Hippolytidae) reared in the laboratory differ morphologically from those in nature. Journal of the Marine Biological Association of the United Kingdom, 83, 949–957. https://doi.org/10.1017/S0025315403008130h
Wiegmann, A. (1836). Beschreibung einiger neuen Crustaceen des Berliner Museums aus Mexiko und Brasilien. Archiv für Naturgeschichte, 2, 145–151.
Wowor, D., Muthu, V., Meier, R., Balke, M., Cai, Y., & Ng, P. K. L. (2009). Evolution of life history traits in Asian freshwater prawns of the genus Macrobrachium (Crustacea: Decapoda: Palaemonidae) based on multilocus molecular phylogenetic analysis. Molecular Phylogenetics and Evolution, 52, 340–350. https://doi.org/10.1016/j.ympev.2009.01.002


