José G. Palacios-Vargas a, Erika Rivero-Sánchez a, Yan Gao b, Margarita Ojeda a, *
a Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Ecología y Recursos Naturales, Laboratorio de Ecología y Sistemática de Microartrópodos, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
b China Shanghai Natural History Museum, Shanghai Science & Technology Museum, Shanhaiguan Road, Jing’an district, Shanghai 200041, China
As a result of 40 years of work and many projects on soil fauna and especially springtails, a collection of edaphic microarthropods has been established at the Facultad de Ciencias, UNAM, and has the name of “Colección de ácaros y colémbolos del Laboratorio de Ecología y Sistemática de Microartrópodos”. During the revision of material belonging to the subfamily Pseudachorutinae kept in the Collembola collection, we obtained 581 records of 28 species of Pseudachorutes, from 20 states and 62 localities from Mexico. Three are new records: P. ca. algidensis, P. ca. crassus, and P. reductus, and 5 new species from soil, litter and epiphytic plants in Mexico are described and illustrated herein: P.tabasquensis sp. nov., P. mexicanus sp. nov., P. chichinautzin sp. nov., P. tillandsiodes sp. nov., and P.veracruzensis sp. nov.
Cinco especies nuevas de Pseudachorutes (Collembola: Neanuridae) de México
Resumen
Como resultado del trabajo de más de 40 años y diversos proyectos sobre la fauna del suelo, especialmente colémbolos, se estableció en la Facultad de Ciencias, UNAM, una colección de microartrópodos edáficos, registrada con el nombre de “Colección de ácaros y colémbolos del Laboratorio de Ecología y Sistemática de Microartrópodos”. Durante la revisión del material depositado en ella, especialmente de la subfamilia Pseudachorutinae, se obtuvo información de 581 registros de 28 especies de Pseudachorutes distribuidos en 20 estados y 62 localidades del país. De éstas, 3 son nuevos registros (P. ca. algidensis, P. ca. crasus y P. reductus) y 5 son especies nuevas provenientes de suelo, hojarasca y plantas epífitas en México, que se describen e ilustran aquí: P.tabasquensis sp. nov., P. mexicanus sp. nov., P. chichinautzin sp. nov., P. tillandsiodes sp. nov. y P.veracruzensis sp. nov.
The genus Pseudachorutes Tullberg, 1871 (Neanuridae: Pseudachorutinae) was based on the type species Pseudachorutessubcrassus Tullberg, 1871 characterized by: 1) ocelli 8+8; 2) postantennal organ in one circle or ellipse; 3) Ant. III and IV dorsally fused, Ant. IV generally with 6 sensilla and apical bulb, Ant. III organ with 2 microsensilla in a cuticular fold, 2 guard sensilla and one microsensillum; 4) bucal cone sharp, mandible with 2 or more teeth, maxilla styliform; 5) unguiculus absent; 6) furcula usually well developed, mucro present; 7) sixth abdominal segment always visible in dorsal view, anal spines absent (Christiansen & Bellinger, 1998; Fjellberg, 1998; Palacios-Vargas, 1990).
Currently there are 119 species of Pseudachorutes in the world (Bellinger et al., 2023), and for Mexico records of 20 species from 18 states are known (Arango-Galván et al., 2007; Cutz-Pool et al., 2003, 2007a, b, 2008; Palacios-Vargas, 1997, 2005; Palacios-Vargas & Castaño-Meneses, 2003; Palacios-Vargas et al., 2000, 2007; Vázquez, 1988; Vázquez & Palacios-Vargas, 1990, 2004). The genus is found in great abundance in many ecosystems and biotopes, including soil, litter, mosses, epiphytic plants and even the forest canopy.
During a recent project to study a collection of the subfamily Pseudachorutinae housed in the Collembola collection of the Laboratorio de Ecología y Sistemática de Microartrópodos (LESM), we found 5 new species of Pseudachorutes and their descriptions and illustrations are given herein.
Materials and methods
Examination of the material deposited in the LESM scientific collection “Colección de ácaros y colémbolos” (register number D.F.-ENT-229-09-09, issued by Subsecretaría de Gestión para la Protección Ambiental, Dirección de Vida Silvestre) was made to separate all the specimens of the genus Pseudachorutes.The specimens are permanently mounted in Hoyer’s medium slides. Measurements are presented as the range with means in parentheses and expressed in micrometers (µm). Drawings were made with the aid of a phase contrast microscope Carl Zeiss Standard 3 K7, equipped with a drawing tube. The full body scales correspond to 500 µm, and the rest of the structures to 100 µm.
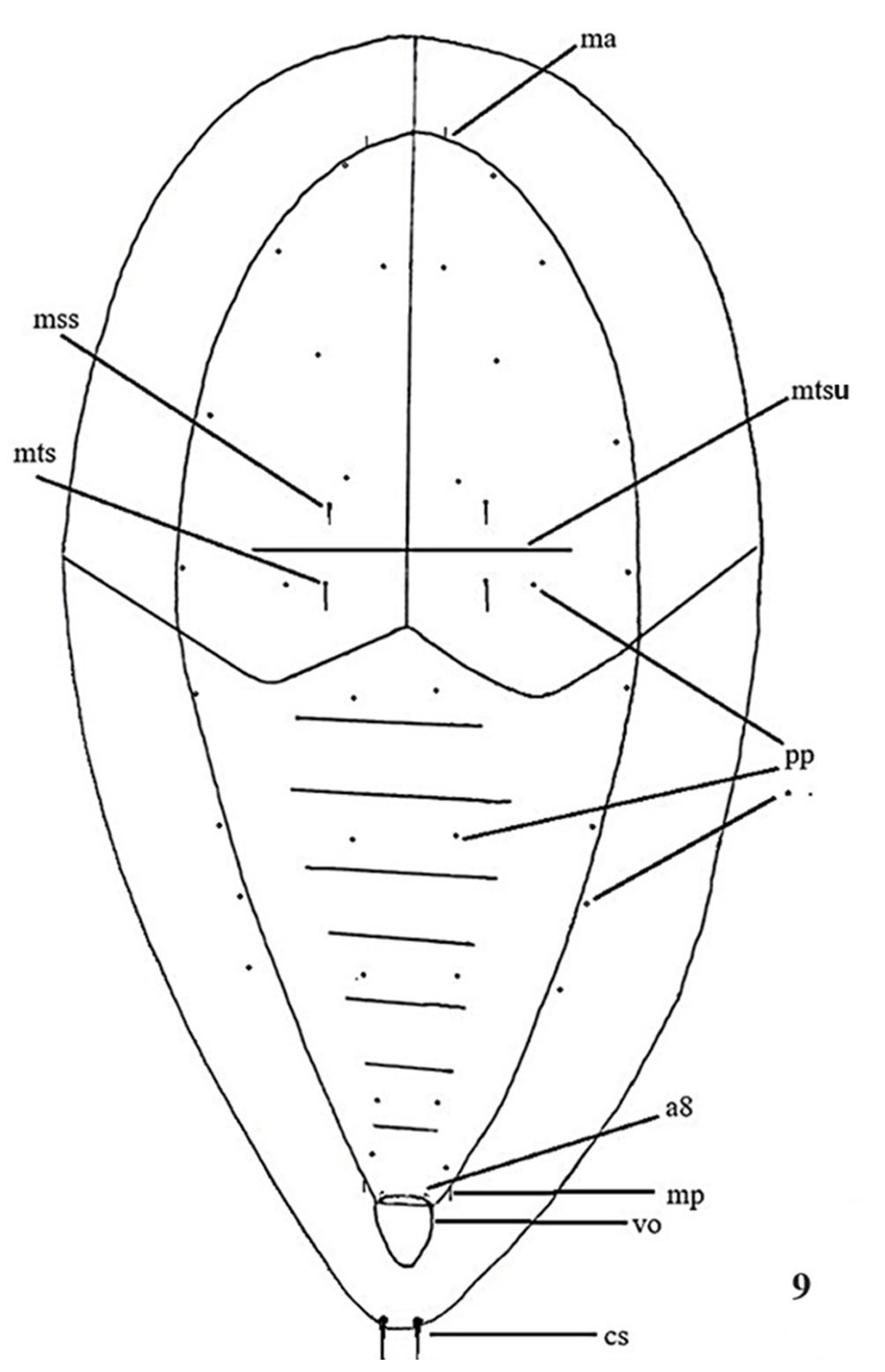
Type specimens are deposited in the LESM. The chaetotaxy system follows that of Jordana et al. (1997). Abbreviations used in this paper are: Ant. = antennal segment (s), Abd. = abdominal segment (s), PAO = postantennal organ, sgd = dorsal guard sensillum, sgv = ventral guard sensillum, Th. = thoracic segment (s).
Descriptions
Class Collembola Lubbock, 1870
Order Poduromorpha Börner, 1913
Family Neanuridae Börner, 1901
Subfamily Pseudachorutinae Börner, 1906
Genus Pseudachorutes Tullberg, 1871
New records
Pseudachorutes ca. algidensis Carpenter, 1925. Mexico: Hidalgo: Mineral El Chico, 2,900 m asl, 1 specimen, ex Tillandsia violacea, 10-IX-1998, J.A. Monterrubio, col. Pseudachorutes ca. crassus da Gama, 1964. Mexico: Popocatépetl, 3,800 m asl, 1 specimen, ex litter, 29-I-1983, J.G. Palacios, col. Pseudachorutes reductus Thibaud & Massoud, 1983. Mexico: Veracruz: Estación de Biología Tropical, Los Tuxtlas, 8 specimens, 20-X-1997, J. Álvarez, col. Tamaulipas: Rancho El Cielo, 975 m asl, 1 specimen, ex soil, XI-1987, F. J. Villalobos, col.
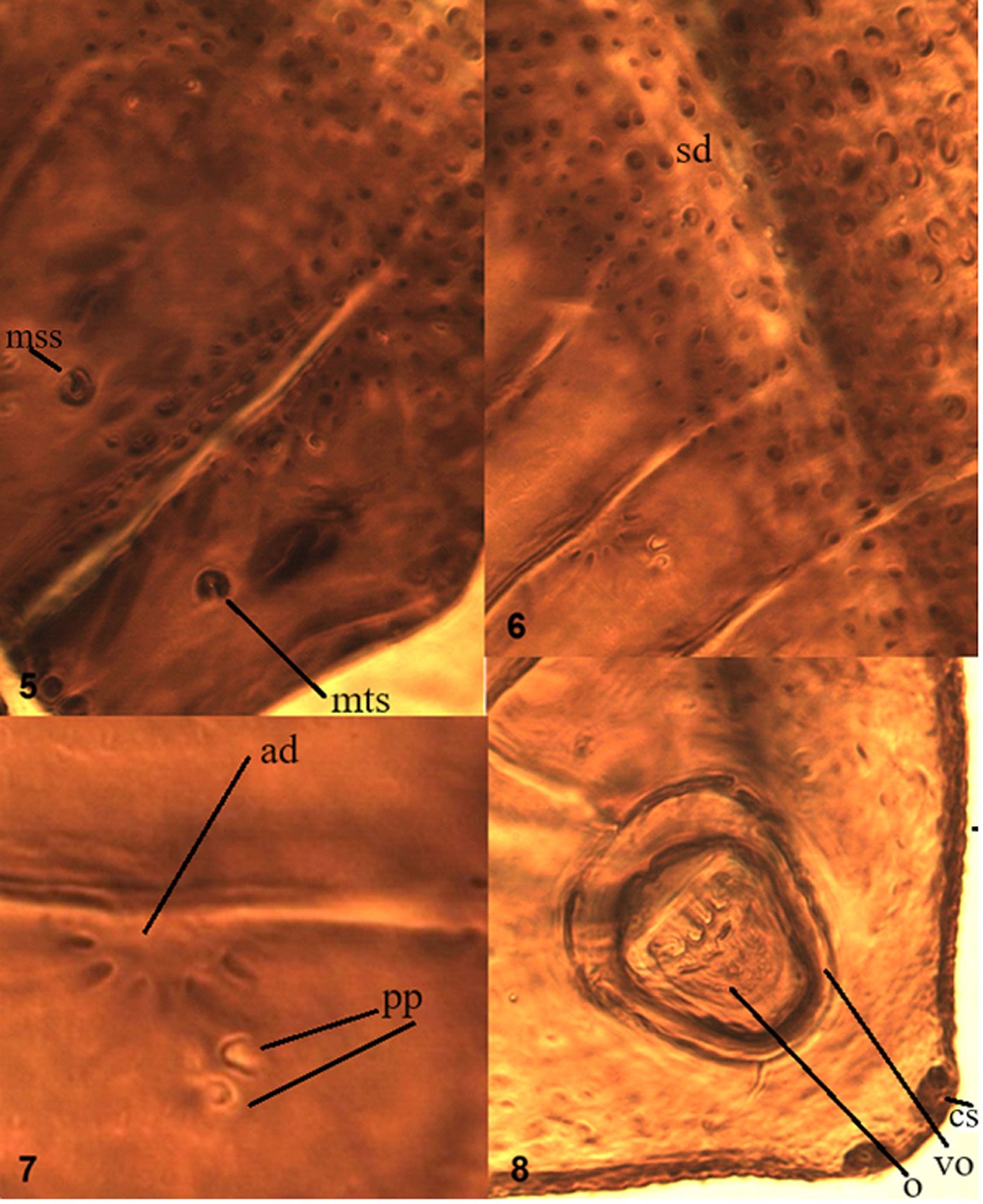
Description. Body length (n = 7): 2,625 µm (range: 1,250-4,000 µm). Body color gray-violet, with dark eyes patches. Granulations are fine and homogenous. Body setae simple and smooth, but with 2 kinds of setae, long macrosetae (M 22-24 µm) and short microsetae (m 10-12 µm), the sensorial setae relatively long (54-56 µm) (Fig. 1 A, B).
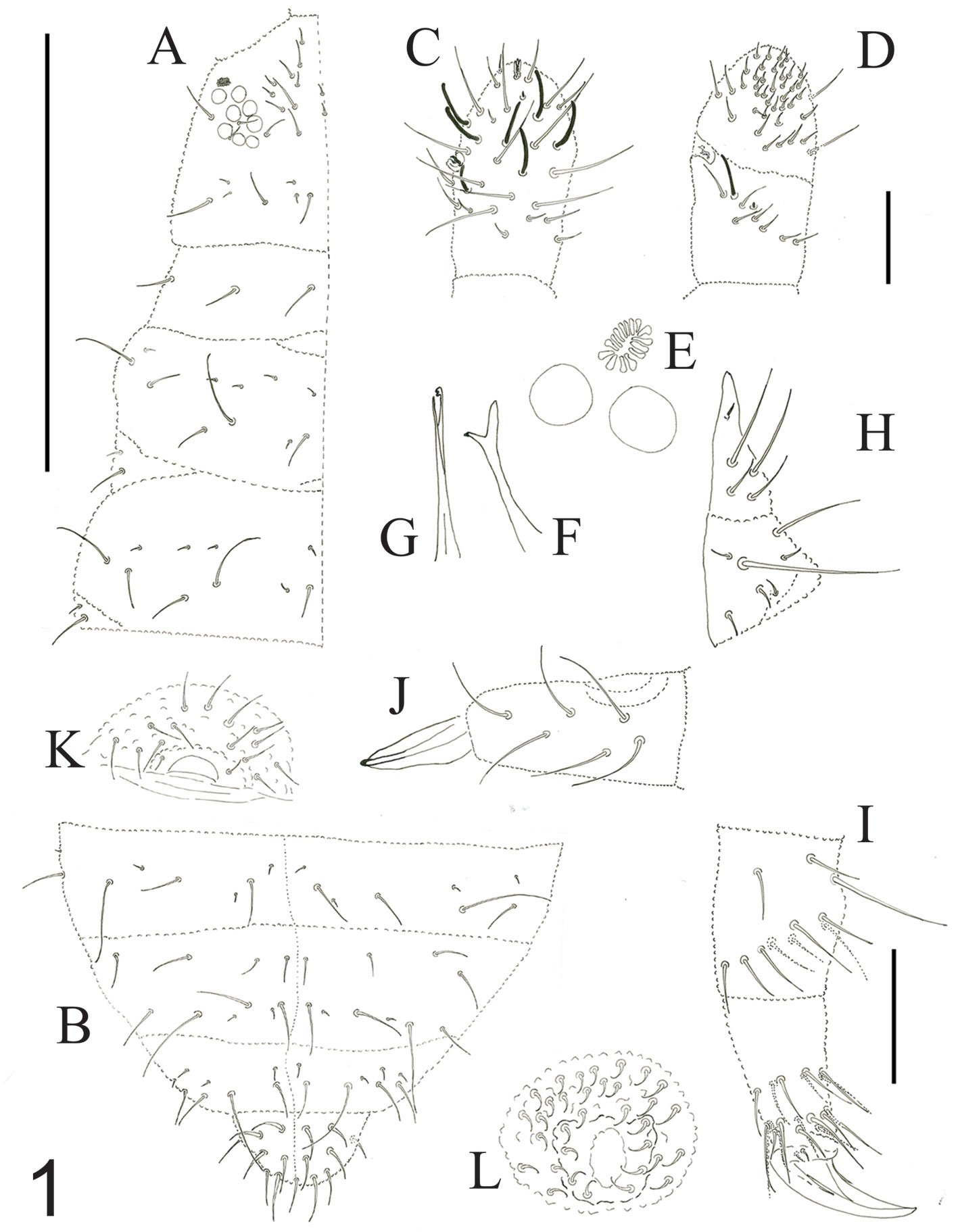
Figure 1. A-L. Pseudachorutestabasquensis sp. nov. A) Dorsal chaetotaxy from head to Th. III; B) dorsal chaetotaxy from Abd. III. to Abd. VI; C) Ant. III-IV right antenna, dorsal view; D) Ant. III-IV right antenna, ventral view; E) PAO and nearby eyes; F) mandible; G) maxilla; H) labium; I) femur, tibiotarsus, and unguis III; J) dens and mucron; K) female genital plate; L) male genital plate.
Table 1
Pseudachorutes tabasquensis sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. orghidani, P. conicus, and P. tabasquensis sp. nov.
1a.
sd
d
oc
c
p
Number of setae
5
4+1
3
1
4
Setae absent
c1, c3, c4
1b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
2
5
a5
m5
Th. III
3
2
5
a2, a5
m5
Abd. I-III
4
–
5
a2
Abd. IV
4
–
6
Abd. V
4
–
4
Abd. VI
2
2
2+1
1c.
Characters
P. conicus
P. tabasquensis sp. nov.
P. orghidani
Labium setae L
–
+
?
Ant. IV sensilla
5
6
5
Ventral tube setae
3+3
4+4
?
Mandible teeth
3
2
3
Ventral file on Ant. IV
–
25-30 short setae
25-30 cuniform
Vesicles of PAO
13-15
13-17
17
Antennae as long as head. Ant. I with 7 setae, Ant. II with 11 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II: III+IV as 1: 1; 1.8. Ant. III-organ with 2 small straight internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.4 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 cylindrical sensilla, seta “i”, and one subapical organite (Fig. 1C), ventral file with 25-30 short and strongly spine-like setae (Fig. 1D). PAO elliptical composed of 13-17 simple vesicles, 0.9 times smaller than the nearest eyes (Fig. 1E). 8+8 eyes, F, G are 0.7 times smaller than others. Buccal cone elongated. Mandible with 2 slender teeth (Fig. 1F). Maxilla styliform, with 2 blades, one has an apical tooth, another has 2 apical teeth (Fig. 1G). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae. Setae L spine-shape (Fig. 1H).
Dorsal chaetotaxy as in figure 1A-B, table 1a, b. Seta a0 on the head absent, unpaired seta d1 present, sometimes, one additional seta d3’ between the seta d3 present. Th. I with 3+3 setae, plus 1+1 lateral. Seta a2 present on Th. II, but absent from Th. III to Abd. V. Sensorial setae s on the body in position of p4 and m6 on the thoracic segments II and III, and p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 2 times as long as the macrosetae. The ratio of the largest Abd. V setae and inner unguis length is 1.0. Thoracic sterna without setae.
Legs setation from I to III is, tibiotarsi 19, 19, 18, without tenent hairs; Femora 10, 10, 11, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 7; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis wide with one inner tooth near 1/3 part from the basal, and a weakly subbasal lateral tooth. Ratio of tibiotarsus III and unguis about 1.6. Unguiculus absent (Fig. 1I).
Furcula is well developed. Dens dorsally with 6 setae, ventral with a smooth area. Mucro straight, 1.8 times shorter than dens, with granulations and 2 small lamellae (Fig. 1J). Tenaculum with 3+3 teeth. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 6-14 circumgenital setae and 1+1 eugenital setae (Fig. 1K). Male genital plate with 3+3 pregenital setae, 25 circumgenital setae and 4+4 eugenital setae (Fig. 1L).
Taxonomic summary
Type material. Holotype: male mounted on a slide (FC-UNAM: LESM-AC: 23013). 5 paratypes females and 1 juvenile mounted on slides (FC-UNAM: LESM-AC: 23014-23019), same data as holotype.
Type locality. Mexico, Tabasco, Tapijulapa, outside cave “Las Sardinas”, ex litter, 14-III-2002, D.A. Estrada col.
Etymology. The name is locative for the state of Tabasco where the type locality is.
Remarks
Pseudachorutes tabasquensis sp. nov. shares with P. orghidani Massoud & Gruia, 1973 the presence of 1 internal and 1 lateral tooth on unguis. The new species also resembles P. conicus Lee & Kim, 1994 from Korea due to the presence of 2 types of body setae. They all share a similar number of PAO vesicles (Table 1c), dens with 6 setae and tenaculum with 3+3 teeth. Main differences between them are shown in Table 1c. Additionally, P. conicus has a very long and thin unguis, but in P. tabasquensis it is short and thick.
Description. Body length (n = 17): 814 µm (range: 470-1,350 µm). Color of the body gray-violet, with a dark eyes patch. Granulations are fine and homogenous. Posterior setae of body long and capitated (Fig. 2A).
Antennae as long as head. Ant. I with 7 setae, Ant. II with 11 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1.4; 3.2. Ant. III-organ with 2 small straight internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.2 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with simple apical bulb, 6 cylindrical sensilla, seta “i”, 1 microsensillum and 1 subapical organite (Fig. 2B), ventral file poorly developed, with 20-35 short setae (Fig. 2C). PAO elliptical composed of 5-6 simple vesicles, 1.1 times as long as the nearest ocelli (Fig. 2D). 8+8 small ocelli, F, G are 0.9 times smaller than others. Buccal cone short. Mandible with 2 slender teeth (Fig. 2E). Maxilla with 2 blades, each has 2 apical teeth (Fig. 2F). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae, setae L spine-shape (Fig. 2G).
Dorsal chaetotaxy as in figure 2A, table 2b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. V. Sensory setae s on the body in position of p4 and m6 on Th. II and III, and p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensory setae longer than 1.1-1.5 times as long as body setae. Ratio of largest Abd. V setae and inner unguis length is 1.4. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 2+2 pregenital setae, 4-9 circumgenital setae and 1+1 eugenital setae (Fig. 2H). Male genital plate with 3+3 pregenital setae, 10 circumgenital setae and 4+4 eugenital setae (Fig. 2I).
Tibiotarsi I, II, III with 18, 18, 17 setae respectively, with 1 tenent hair long and capitate (Fig. B8). Femora I, II, III with 9, 9, 10 setae respectively. Trochanters with 5 setae each. Coxae I, II, III with 3, 7, 7 setae respectively. Subcoxae 2 I, II, III with 0, 2, 2 setae respectively. Subcoxae 1. I, II, III with 1, 2-3, 2-3 setae respectively. Unguis wide with 1 weakly apical inner tooth. Ratio of tibiotarsus III and unguis about 1.0. Unguiculus absent (Fig. 2J).
Furcula well developed. Dens dorsally with 5 setae. Mucro straight, 3 times shorter than dens, with two larger bladder-like swelling visible and a hook-like end (Fig. 2K). Tenaculum with 3+3 teeth.
Figure 2. A-K. Pseudachorutesmexicanus sp. nov. A) Dorsal chaetotaxy from head to abdomen VI; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO and nearby eye; E) mandible; F) maxilla; G) labium; H) female genital plate; I) male genital plate; J) femur, tibiotarsus, and unguis III; K) furcula and tenaculum.
Taxonomic summary
Type material. Holotype: female mounted on a slide (FC-UNAM: LESM-AC: 22996). 16 paratypes: 4 females, 1 male and 11 juveniles under slides (FC-UNAM: LESM-AC: 22997-23012), same data as holotype.
Type locality. Mexico, Hidalgo, Mineral El Chico, 10-IX-98, J.A. Monterrubio, col. Ex Tillandsia violacea.
Etymology. The name of the new species is after the country of the type locality: Mexico.
Remarks
This species resembles P. americanus Stach, 1949 with a similar number of vesicles in PAO (P. americanus with 5-8 vesicles), unguis with 1 minute tooth near apex, one large, clavate tenent hair, ventral tube with 4+4 setae, tenaculum with 3+3 teeth and posterior abdominal setae clearly capitate. However, dens of the new species with 5 setae, maxilla with 2 blades and each has 2 apical teeth, 6 cylindrical sensilla on Ant. IV and a poorly developed ventral file with 20-35 short setae is different from P. americanus.
Description.Body length (n = 8): 1,904 µm (range: 850-2,010 µm). Color of the body gray-violet, with a dark eyes patch. Granulations homogenous. Lateral and posterior body setae are longer and truncate (Fig. 3A).
Table 2
Pseudachorutes mexicanus sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy.
2a.
sd
d
oc
c
p
Number of setae
5
4+1
3
4
4
2b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
3
5
a4
Th. III
3
3
5
Abd. I-III
3
2
5
Abd. IV
4
–
5
Abd. V
3
–
4
Abd. VI
2
2
2+1
Antennae as long as head. Ant. I with 7 setae, Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1.2: 2.0. Ant. III-organ with 2 small internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.1 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 cylindrical sensilla, seta “i”, and one subapical organite (Fig. 3B), ventral file with about 10 strong, spine-like setae and several slender, normal setae (Fig. 3C). Some setae on Ant. IV are blunt. PAO elliptical composed of 10-12 vesicles, sometimes, 1 or 2 of them inside in the others, subequal to nearest eyes (Fig. 3D). 8+8 eyes, F, G are 0.7 times smaller than others. Buccal cone elongated. Mandible is not clearly detected, about 2 or 3 slender teeth. Maxilla with 2 blades and one has an apical tooth, and the other styliform (Fig. 3E). Labium with normal chaetotaxy of the genus from setae A to G and 5 lateral setae, setae L spine-shape (Fig. 3F).
Dorsal chaetotaxy as in Fig. 3A, table 3a, b. Seta a0 on the head absent, unpaired seta d1 present. Th. I with 3+3 setae, plus 1+1 lateral. Setae a2 present on Th. II, but absent from Th. III to Abd. V. Sensorial setae s on the body in position of p4 and m6 on the thoracic segments II and III, and p5 from Abd. I to IV and p3 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 1.2-1.7 times as long as the normal setae. The lateral setae of the body longer and blunt. Ratio of largest Abd. V setae and inner unguis length is 1.0. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 9 circumgenital setae and 1+1 eugenital setae (Fig. 3G). Male genital plate with 3+3 pregenital setae, 8 circumgenital setae and 4+4 eugenital setae (Fig. 3H).
Leg setation from I to III, is tibiotarsi 19, 19, 18, with 1 acuminate tenent hair a little longer than others; femora 9, 9,10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 7; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis wide, with one inner tooth near 1/3 part from the basal. Sometimes, a pair of weakly subbasal lateral tooth is present. Ratio of tibiotarsus III and unguis about 1.6. Unguiculus absent (Fig. 3I).
Figure 3. A-J. Pseudachoruteschichinautzin sp. nov. A) Dorsal chaetotaxy; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO, eye patch and ocular setae; E) maxilla; F) labium; G) female genital plate; H) male genital plate; I) femur, tibiotarsus, and unguis III; J) furcula and tenaculum.
Furcula well developed. Dens dorsally with 6 setae, ventral with fine granulate. Mucro straight, 1.7 times shorter than dens, with 2 big lamella and a clearly hook-like end. Tenaculum with 3+3 teeth (Fig. 3J).
Taxonomic summary
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 22974). 7 paratypes: 4 females, 2 males and 1 juvenile under slides, same data as holotype (FC-UNAM: LESM-AC: 22975-22981).
Type locality. Mexico; Morelos, Derrame Chichinautzin. 12-IX-1976. J. Palacios, col. Ex. Tillandsia prodigiosa.
Etymology. The name is that of the Chichinautzin lava flow (state of Morelos), as a noun of type locality.
Remarks
Pseudachorutes chichinautzin sp. nov. shares with P. orghidani and P. subcrassus Tullberg, 1871 the presence of 1 internal and 1 pair of lateral teeth on unguis. The new species also resembles P. tabasquensis sp. nov. with 2 types of body setae, Ant. IV with 6 sensilla, trilobed apical bulb. Dens with 6 setae and tenaculum with 3+3 teeth. Main differences between them can be seen in table 3c.
Table 3
Pseudachorutes chichinautzin sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. orghidani, P. subcrassus, P. chichinautzin sp. nov. and P. tabasquensis sp. nov.
Description. Body length (n = 6): 2,260 µm (range: 1,750-2,860 µm). Color of the body violet, with a white strip from Th. I to Th. II and a dark eyes patch. Granulations fine and homogenous. Body with simple and spine-like setae, p row longer on last abdominal segments (24-40 µm), long sensorial setae (75-100 µm) (Fig. 4A, B).
Antennae shorter than head, 155 µm and 180 µm respectively. Ant. I with 7 setae Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1; 1.9. Ant. III-organ with 2 small curving sensilla under a cuticular fold, 2 guard sensilla (sgv is about 1.4 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 thin and cylindrical sensilla, seta “i”, one microsensillum and one subapical organite (Fig. 4C), ventral side with about 20 setae with some thick and spine-like (Fig. 4D). PAO elliptical composed of 14 vesicles, 1.2 times as long as the nearest eyes. 8+8 eyes, F, G 0.9 times as big as others (Fig. 4E). Buccal cone elongated. Mandible has 2 big teeth, the apical with clearly 3 small teeth (Fig. 4F). Maxilla with 2 blades, one with 2 apical teeth (Fig. 4G). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae, setae L reduced to a minus spine, difficult to see (Fig. 4H).
Dorsal chaetotaxy as in figure 4A, B, table 4a, b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. V. m5 present on Th. II to Th. III. Sensorial setae on body in position of p4 and m6 on thoracic segments II and III, p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 2.5-4.0 times as long as the normal setae. Ratio of largest Abd. V setae and inner unguis length is 0.6. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 9 circumgenital setae and 1+1 eugenital setae (Fig. 4I). No males were found.
Figure 4. A-K. Pseudachorutes tillandsiodes sp. nov. A) Dorsal chaetotaxy from head to Th. III; B) dorsal chaetotaxy from Abd. III. to Abd. VI; C) Ant. III-IV right antenna, dorsal view; D) Ant. III-IV right antenna, ventral view; E) PAO, eye patch and ocular setae; F) mandible; G) maxilla; H) labium; I) female genital plate; J) femur, tibiotarsus, and unguis III; K) dens and mucron.
Leg setation from I to III, is tibiotarsi 19, 19, 18, no tenent hair; femora 9, 10, 10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 8; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis with one clearly inner tooth at the basal side. Ratio of tibiotarsus III and unguis about 1.5. Unguiculus absent (Fig. 4J).
Furcula well developed. Dens dorsally with 6 setae, ventral with fine granulations. Mucro straight, 2.2 times shorter than dens, with 2 thin but long lamella, without hook-like end (Fig. 4K). Tenaculum with 3+3 teeth.
Table 4
Pseudachorutes tillandsiodes sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. gilvusi, and P.tillandsiodes sp. nov.
4a.
sd
d
oc
c
p
Number of setae
5
4+1
3
4
4
4b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
5
2
5
m4
Th. III
4
2
5
a2
m4
Abd. I-III
4
–
5
a2
Abd. IV
4
–
5
a2
Abd. V
4
–
3
P4
Abd. VI
2
2
2+1
4c.
Characters
P. gilvus
P. tillandsiodes sp. nov.
Setae d1 on head
1+1
1
Setae number on Th. I
2+2
3+3
Setae a2 on Th. II
–
+
Ant. IV sensilla
7
6
Ventral tuve setae
3+3
4+4
Taxonomic summary
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 23172). 5 Paratypes: 4 female and 1 juvenile (FC-UNAM: LESM-AC: 23173-23177).
Type locality. Mexico; Hidalgo, Mineral El Chico. 10-IX-98. J. A. Monterrubio, col. Ex Tillandsia violacea.
Etymology. The name is taken from the epiphytic genus Tillandsia (Bromeliacea) habitat where the species was found.
Remarks
Pseudachorutes tillandsiodes sp. nov. resembles P. gilvus Oliveira & Deharveng, 1995 with white strips and long sensilla on the body, similar number of postantennal organ vesicles (P. gilvus with 11-15 vesicles), unguis with 1 inner tooth, tibiotarsi I-III with 19, 19, 18 setae and dens with 6 setae. The main differences between them are shown in table 4c. However, P. gilvus has 3 white stripes on the body: the first on the posterior part of head and the middle of Th. I, the second on the mesothorax and the third on Abd. I-II; P. tillandsiodes sp. nov. only with 1 white stripe from Th. I to Th. II.
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 2133a). 10 paratypes: 7 females and 3 males under slides, same data as holotype (FC-UNAM: LESM-AC: 2131a-2132c, 2132a-2132e, 2133b, 2133c)
Type locality. Mexico; Veracruz, Xalapa, La Herradura. 26-IX-26-10/1998, ex Bosque Mesófilo de Montaña, J. Márquez, col.
Description. Body length (n = 11): 1,900 (range: 1,050-3,400 µm). Color body violet, with a dark eyes patch. Granulations fine and homogenous. Body with short and simple setae (10-12), some of them longer in abdomen segments, especially on 4th segment (22-26 µm), long sensorial setae (67-75 µm) (Fig. 5A).
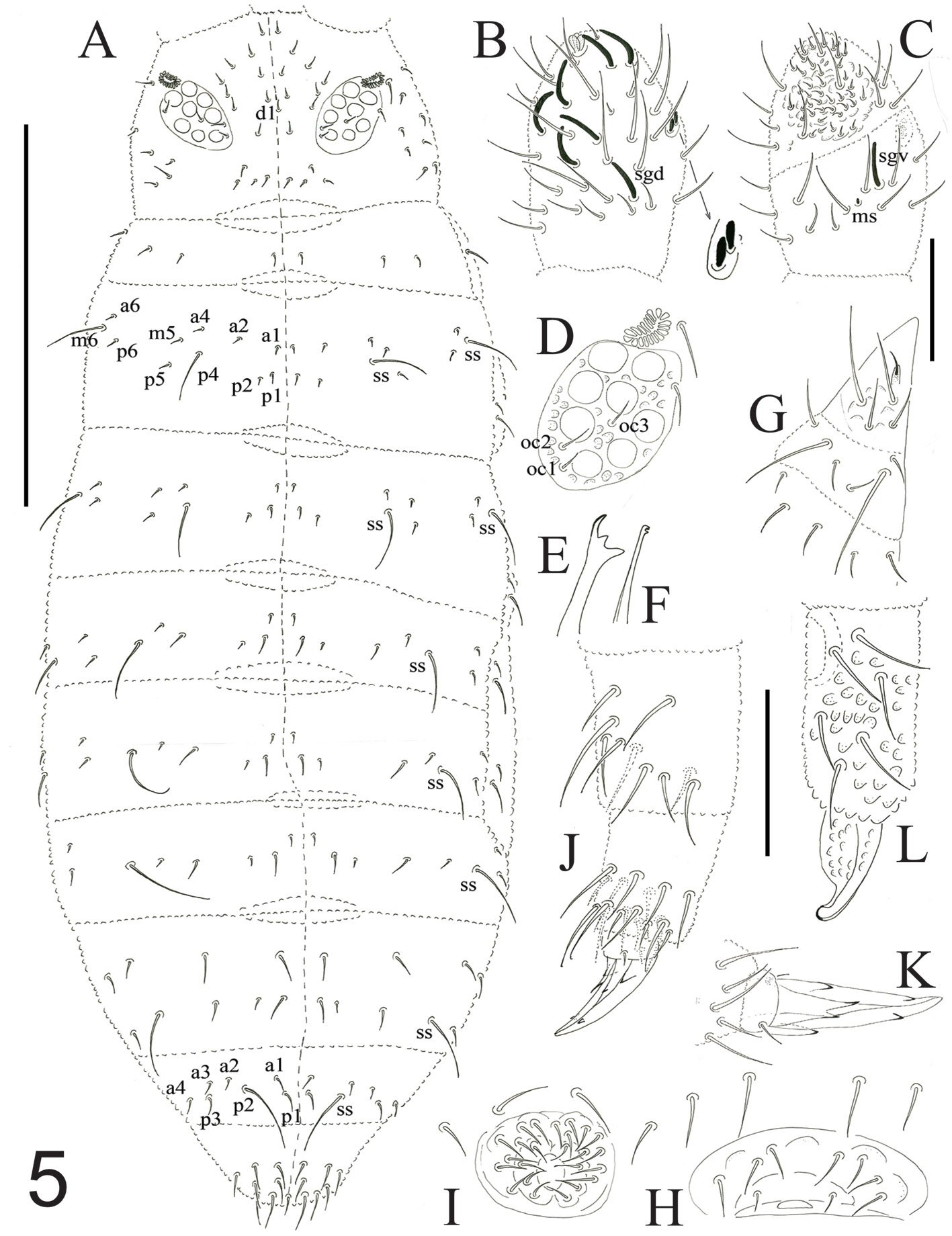
Antennae little shorter than head, 100 µm and 105 µm, respectively. Ant. I with 7 setae, Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1; 2. Ant. III-organ with 2 small straight sensilla under a cuticular fold, 2 guard sensilla (the sgv is about 1.1 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV dorsally with trilobed apical bulb, 6 thin and cylindrical sensilla, seta “i”, 1 microsensillum and 1 subapical organite (Fig. 5B), ventral side has a distinct ventral file with about 40 short setae (Fig. 5C). PAO elliptical composed of 17-20 vesicles, 1.5 times as long as the nearest eyes. 8+8 eyes, F, G 0.8 times as big as others (Fig. 5D). Buccal cone elongated. Mandible has 2-3 big teeth (Fig. 5E). Maxilla with 1 blade and 2 apical teeth (Fig. 5F). Labium with normal chaetotaxy of the genus from setae A to G and 3 lateral setae, one longer spine-like setae present at the place of setae L (Fig. 5G).
Dorsal chaetotaxy as in figure 5A and table 5a,b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. IV. m5 present on Th. II to Th. III. Sensorial setae s on the body in position of p4 and m6 on Th. II and III, p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 6.5-7.0 times as long as the shorter setae. The ratio of the largest Abd. V setae and inner unguis length is 0.5. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 7 circumgenital setae and 1+1 eugenital setae (Fig. 5H). Male genital plate with 2+2 pregenital setae, 16 circumgenital setae and 4+4 eugenital setae (Fig. 5I).
Leg setation from I to III, is tibiotarsi 19, 19, 18, no tenent hairs; femora 13, 11, 10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 6,6,5; coxae 3, 7, 8; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis with 1 big and 1 small inner tooth together with 2 pairs of lateral teeth (Fig. 5J). Ratio of tibiotarsus III and unguis about 1.4. Unguiculus absent (Fig. 5K).
Furcula well developed. Dens dorsally with 6 setae, ventral granulate. Mucron granulated with broad and long lamella, 1 slightly hook-like end, 2.0 times shorter than dens (Fig. 5L). Tenaculum with 3+3 teeth.
Etymology. The name is a locative for the State of Veracruz where the type locality is found.
Remarks
Pseudachorutes veracruzensis sp. nov. resembles P. orghidani with granules on mucron and dens with 6 setae, mandible with 3 teeth, unguis with lateral teeth, similar number of PAO (P. orghidani with 17 vesicles) and ventral file on Ant. IV. The main difference between the species is the shape of short setae in the ventral file (P. orghidani has small setae with apex truncate) and number of sensilla on Ant. IV (P. orghidani with 5) and the teeth on unguis (P. orghidani only with 1+1 lateral teeth).
Discussion
After revision and analysis of the material deposited in the LESM collection, we were able to update the knowledge of genus Pseudachorutes in Mexico, describing 5 new species: P. tabasquensis sp. nov., P. veracrucensis sp. nov., P. tillandsiodes sp. nov., P. chichinautzin sp. nov., and P. mexicanus sp. nov. Three new records for the country are added, for the following species: P. ca. algidensis from Hidalgo, P. ca. crassus from Estado de México, these must be confirmed with the collection of more specimens that will allow the species to be fully determined. For now, this information is presented as an element to show the richness of species of the genus in the country. Pseudachorutes reductus has a distribution in the Antilles and southern Florida, the new records in Mexico extend its distribution area and confirm the affinity of the species to the Atlantic area. Total records for Pseudachorutes is increased to 28 species, from 20 states and 62 localities within the country. Quintana Roo and Hidalgo were the states with the highest number of species present (11 and 10, respectively), followed by Morelos, Estado de México, Puebla, Veracruz, and Guerrero (8, 7, 6, 6, and 4 species recorded in each one), 6 states have 3 species, 5 have 2 and Querétaro and San Luis Potosí only have 1 species recorded. Three species are widely distributed in Mexico: P. corticolus, P. simplex, and P. subcrassoides. Diversity of the genus in Mexico is around 21% of the total known worldwide, and therefore the country becomes one with the greatest number of species present.
Figure 5. A-L. Pseudachorutes veracruzensis sp. nov. A) Dorsal chaetotaxy; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO, eye patch and ocular setae; E) mandible; F) maxilla; G) labium; H) female genital plate; I) male genital plate; J) femur, tibiotarsus, and unguis III; K) unguis III, ventral view; L) dens and mucron.
Regarding biotopes where the species preferably live, we found that litter, epiphytic plants, soil, mosses and decaying wood are the most suitable habitats for this springtail. However, sand, canopy, wood, caves and anthills are also microhabitats used by some species.
Table 5
Pseudachorutes veracruzensis sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy.
5a.
sd
d
oc
c
p
Number of setae
5
4+1
3
2
4
Setae absent
c1, c3 c4
5b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
2
5
a5
m4
Th. III
3
2
5
a2, a5
m4
Abd. I-III
3
–
5
a2, a5
Abd. IV
4
–
5
a2, a3
Abd. V
3
–
4
a4
Abd. VI
3
2
2+1
The analysis and revision of the morphology of the 5 species described, allowed us to come to the following conclusions about the morphology of the genus that needs to be taken in consideration. Size of body setae should be a characteristic to be taken into consideration for the description of the species, since we observed that it is a variable character. Antennae are a uniform structure and characters associated with the sensory organ of Ant. III have little relevance at a specific level. Sensory file of Ant. IV, present a varied shape and number, from 8 to 40 setae. Apical vesicle of Ant. lV is trilobed in most species and others have a simple shape. Number of ocelli is stable within the genus, presenting 8+8. The maxillae and mandibles vary in all species. Maxillae can be needle-shaped, crocheted or styliform, they may or may not be lamellar and the number of lamellae is variable, and in most cases, there are 1 to 2 apical teeth. The number of teeth in the mandibles in almost all species ranges from 1 to 5. The number of teeth in the retinaculum is a constant character among the species of the genus, with 3+3 teeth. The ventral tube in almost all species has 4+4 setae. The number of setae in the dens varies from three to seven, but most have six setae. The mucron has 2 lamellae in all species, except in P. reductus, where it is greatly reduced. The shape of the mucron is variable, from elliptical, triangular, elongated, short, curved, widened, or with a bladder-shaped base.
Acknowledgments
This study had the support from the International Relationships of Universidad Nacional Autónoma de México issued to the first author and partially by the National Natural Sciences Foundation of China (No. 3217047 and 32370491). Blanca E. Mejía Recamier helped remount specimens. Kenneth A. Christiansen gave criticism, suggestions, and literature. We thank Erick García and Eduardo Pacheco who assigned the catalogue numbers for the slides. Daniel A. Estrada, Jesús A. Monterrubio, Juan Márquez donated the specimens.
References
Arango-Galván, A., Cutz-Pool, L. Q., & Cano-Santana, Z. (2007). Estructura de la comunidad de colémbolos del mantillo de la Reserva Ecológica del Pedregal de San Ángel: Riqueza, composición y abundancia relativa. Entomología Mexicana, 6, 397–400.
Bellinger, P. F., Christiansen, K. A., & Janssens, F. (2023). Checklist of the Collembola of the World. Retrieved on November 11th, 2023 from: http://www.collembola.org
Christiansen, K. A., & Bellinger, P. F. (1998). The Collembola of North America, North of the Río Grande: a taxonomic analysis. Part I. Grinnell, Iowa Grinnell College.
Cutz-Pool, L. Q., Palacios-Vargas, J. G., & Vázquez, M. M. (2003). Comparación de algunos aspectos ecológicos de Collembola en cuatro asociaciones vegetales de Noh-Bec, Quintana Roo, México. Folia Entomológica Mexicana, 42, 91–101.
Cutz-Pool, L. Q., García-Gómez, A., & Palacios-Vargas, J. G. (2007a). Primer estudio de colémbolos (Hexapoda: Collembola) de la Hoya de las Guaguas en el estado de San Luis Potosí, México. Dugesiana, 14, 47–51.
Cutz-Pool, L. Q., Palacios-Vargas, J. G., Castaño-Meneses, G., & García-Calderón, N. E. (2007b). Edaphic Collembola from two agroecosystems with contrasting irrigation type in Hidalgo State, Mexico. Applied Soil Ecology, 36, 46–52. https://doi.org/10.1016/j.apsoil.2006.11.009
Cutz-Pool, L. Q., Palacios-Vargas, J. G., & Castaño-Meneses, G. (2008). Estructura de la comunidad de colémbolos (Hexapoda: Collembola) en musgos corticícolas en un gradiente altitudinal de un bosque templado subhúmedo. Revista de Biología Tropical, 56, 1–10. https://doi.org/10.15517/rbt.v56i2.5620
Fjellberg, A. (1998). Fauna entomologica scandinavica. The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. Vol. 35. Leiden: Brill Publishers.
Jordana, R., Arbea, J. I., Simón, C., & Luciáñez, M. J. (1997). Fauna ibérica. Vol. 8. Collembola, Poduromorpha. Madrid: Museo Nacional de Ciencias Naturales.
Palacios-Vargas, J. G. (1990). Manuales y guías para el estudio de microartrópodos I. Diagnosis y clave para determinar las familias de los Collembola de la Región Neotropical. México D.F.: Facultad de Ciencias, Universidad Nacional Autónoma de México.
Palacios-Vargas, J. G. (1997). Catálogo de los Collembola de México. Las Prensas de Ciencias, UNAM, México D.F.
Palacios-Vargas, J. G. (2005). Collembola. In G. Sánchez-Ramos, P. Reyes-Castillo y R. Dirzo (Eds.), Historia natural de la Reserva de la Biosfera El Cielo Tamaulipas, México (pp. 354–366). Ciudad Victoria, Tamaulipas: Universidad Autónoma de Tamaulipas.
Palacios-Vargas, J. G., & Castaño-Meneses, G. (2003). Seasonality and community composition of springtails in Mexican forests. In Y. Basset, V. Novotny, S. E. Millar, & R. L. Kitching (Eds.), Arthropods of tropical forests. Spatio-temporal dynamics and resource use in the canopy (pp. 159–169). Cambridge, UK: Cambridge University Press.
Palacios-Vargas, J. G., Castaño-Meneses, G., & Mejía-Recamier, B. E. (2000). Collembola. In J. Llorente-Bousquets, E. González-Soriano, & N. Papayero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México. Hacia una síntesis de su conocimiento II (pp. 249–273).México D.F.: Las Prensas de Ciencias, UNAM.
Palacios-Vargas, J. G., Cutz-Pool, L. Q., & Estrada-Barcenas, D. A. (2007). Collembola. In I. Luna, J. J. Morrone, & D. Espinosa (Eds.). Biodiversidad de la Faja Volcánica Transmexicana (pp. 331–344).México D.F.: Las Prensas de Ciencias, UNAM.
Tullberg, T. (1871). Förteckning öfver Svenska Podukrider. Öfversigt af Kongliga Vetenskaaps -Akademiens Förhandlingar, 28, 143–155.
Vázquez, M. M. (1988). Fauna colembológica de hojarasca y suelo. In L. Arriaga, & A. Ortega (Eds.), La sierra de la Laguna de Baja California Sur (pp. 133–148). La Paz, Baja California Sur: Centro de Investigaciones Biológicas de Baja California Sur, A.C.
Vázquez, M. M., & Palacios-Vargas, J. G. (1990). Nuevos registros y aspectos biogeográficos de los colémbolos de la sierra de la Laguna, B. C. S., México. Folia Entomológica Mexicana, 78, 5–22.
Edmundo González-Santillán a, Laura L. Valdez-Velázquez b, *, Ofelia Delgado-Hernández c, Jimena I. Cid-Uribe d, María Teresa Romero-Gutiérrez e, Lourival D. Possani d
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Arácnidos, Tercer Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Universidad de Colima, Facultad de Ciencias Químicas y Facultad de Medicina, Km 9 Carretera Colima-Coquimatlán, 28400 Coquimatlán, Colima, Mexico
c Instituto Francisco Possenti, Av. Toluca 621, Olivar de los Padres, Álvaro Obregón, 01780 Mexico City, Mexico
d Universidad Nacional Autónoma de México, Instituto de Biotecnología, Avenida Universidad 2001, Colonia Chamilpa, 62210 Cuernavaca, Morelos, Mexico
e Universidad de Guadalajara, Centro Universitario Tlajomulco, Departamento de Innovación Tecnológica, Carretera Tlajomulco – Santa Fé Km. 3.5 No.595 Lomas de Tejeda, 45641 Tlajomulco de Zúñiga, Jalisco, Mexico
Scorpion species diversity in Colima was investigated with a multigene approach. Fieldwork produced 34 lots of scorpions that were analyzed with 12S rDNA, 16S rDNA, COI, and 28S rDNA genetic markers. Our results confirmed prior phylogenetic results recovering the monophyly of the families Buthidae and Vaejovidae, some species groups, and genera. We recorded 11 described species of scorpions and found 3 putatively undescribed species of Centruroides, 1 of Mesomexovis, and 1 of Vaejovis. Furthermore, we obtained evidence that Centruroides elegans, C. infamatus,and C. limpidus do not occur in Colima, contrary to prior reports. Seven genetically different and medically relevant species of Centruroides for Colima are recorded for the first time. We used the InDRE database (Instituto de Diagnóstico y Referencia Epidemiológicos), which contains georeferenced points of scorpions, to estimate the distribution of the scorpion species found in our fieldwork. Finally, we discuss from a biogeographical, ecological, and medical point of view the presence and origin of the 14 scorpion species found in Colima.
Keywords: Barcoding; Holotype; Medical relevance; Microendemic; New species; Species group; Substrate-specialist
Una aproximación multigenes para identificar a las especies de alacranes (Arachnida: Scorpiones) de Colima, México, con cometarios sobre la diversidad de sus venenos
Resumen
La diversidad de especies de alacranes de Colima se investigó utilizando una aproximación multigenes. Del trabajo de campo se obtuvieron 34 lotes de alacranes que fueron analizados con los marcadores 12S rDNA, 16S rDNA, COI, y 28S rDNA. La comparación con trabajos de filogenia previos nos permitió confirmar la monofilia de las familias Buthidae y Vaejovidae, de algunos grupos de especies y géneros. Encontramos 11 especies de alacranes descritas, 3 putativamente nuevas de Centruroides, 1 de Mesomexovis y 1 de Vaejovis. También obtuvimos evidencia de que Centruroides elegans, C. infamatus y C. limpidus no están distribuidos en Colima, como se registró en trabajos anteriores. Reportamos 7 especies genéticamente distintas y de importancia médica para Colima. Usamos la base de datos del InDRE (Instituto de Diagnóstico y Referencia Epidemiológicos) que contiene puntos georreferenciados de alacranes para estimar la distribución de las especies que recolectamos en el campo. Finalmente, discutimos desde una perspectiva biogeográfica, ecológica y de importancia médica las 14 especies de alacranes que reportamos para Colima.
Palabras clave: Código de barras; Holotipo; Importancia médica; Microendémico; Especie nueva; Grupo de especies; Sustrato-especialista
Introduction
The knowledge of scorpion diversity in North America has improved recently (González-Santillán & Prendini, 2013; Goodman, Prendini, Francke et al., 2021; Ponce-Saavedra & Francke, 2019; Santibáñez-López et al., 2014); however, much remains to be discovered. Local-scale inventories may be a solution to unveil species communities that, in turn, can help conform regional faunas. This approach has rarely been applied to study scorpion diversity. Furthermore, few local faunal studies have been conducted in Mexico, and only a handful of them have been published; other revisionary contributions included limited fieldwork effort (Baldazo-Monsivais et al., 2012, 2016, 2017).
The International Barcode of Life (iBOL) has grown as a powerful tool for discovering biodiversity, among other applications (https://ibol.org). Scorpion barcoding studies have permitted the identification and delimitation of species in several regions of the world (Fet et al., 2014, 2016; Goodman, Prendini, & Esposito, 2021; Podnar et al., 2021). Despite the high diversity of scorpions in Mexico —an update by Ponce-Saavedra et al. (2023) comprises 311 scorpion species— only 1 mini-barcoding study has been conducted (Goodman, Prendini, & Esposito, 2021). Herein, we present a second scorpion barcoding study for this country but aim at discovering the components of a local scorpion assembly. Colima exhibits a complex topology comprising littorals with a tropical climate and extreme topological variation from sea level to mountain ranges rising to over 4,000 m in approximately 5,600 km2. Colima’s territory supports a rich local flora and fauna (Ramírez-Ruiz & Bretón-González, 2016). Colima lies between the limits of the Nearctic and Neotropical biogeographical Realms (Fig. 1B). Beyond its complexity, Colima represents an enclosed littoral surrounded by mountain ranges and 3 large rivers that divide the territory into 2 sections (Fig. 1), which makes it a well-defined and manageable geographical unit ideal for studying a unique community of scorpions. Thus far, 2 families, 3 subfamilies, 5 genera, and 12 species of scorpions have been identified in Colima (Table 1). The knowledge of scorpion diversity and evolutionary studies in Mexico has steadily been unveiling one of the vastest biodiversity hotspots in the world. For instance, 2 of the most diverse scorpion families and subfamilies, Vaejovidae (Syntropinae) and Buthidae (Centruroidinae), have been treated in recent phylogenetic and taxonomic analyses (Esposito & Prendini, 2019; González-Santillán & Prendini, 2013, 2015).
This contribution aims to survey the scorpion richness in Colima, using not only the COI-barcoding genetic marker but 2 additional mitochondrial markers and 1 nuclear marker to establish a framework to build a stable and predictable taxonomy. Taking advantage of robust phylogenies produced for the 2 families distributed in Colima, we use these topologies as a baseline for comparison to test the presence of several previously reported species and to taxonomically circumscribe our fresh samples. Unlike other barcoding studies, our approach seeks to unveil the richness within the state instead of focusing on delimiting the species of a taxonomic group of scorpions.
Materials and methods
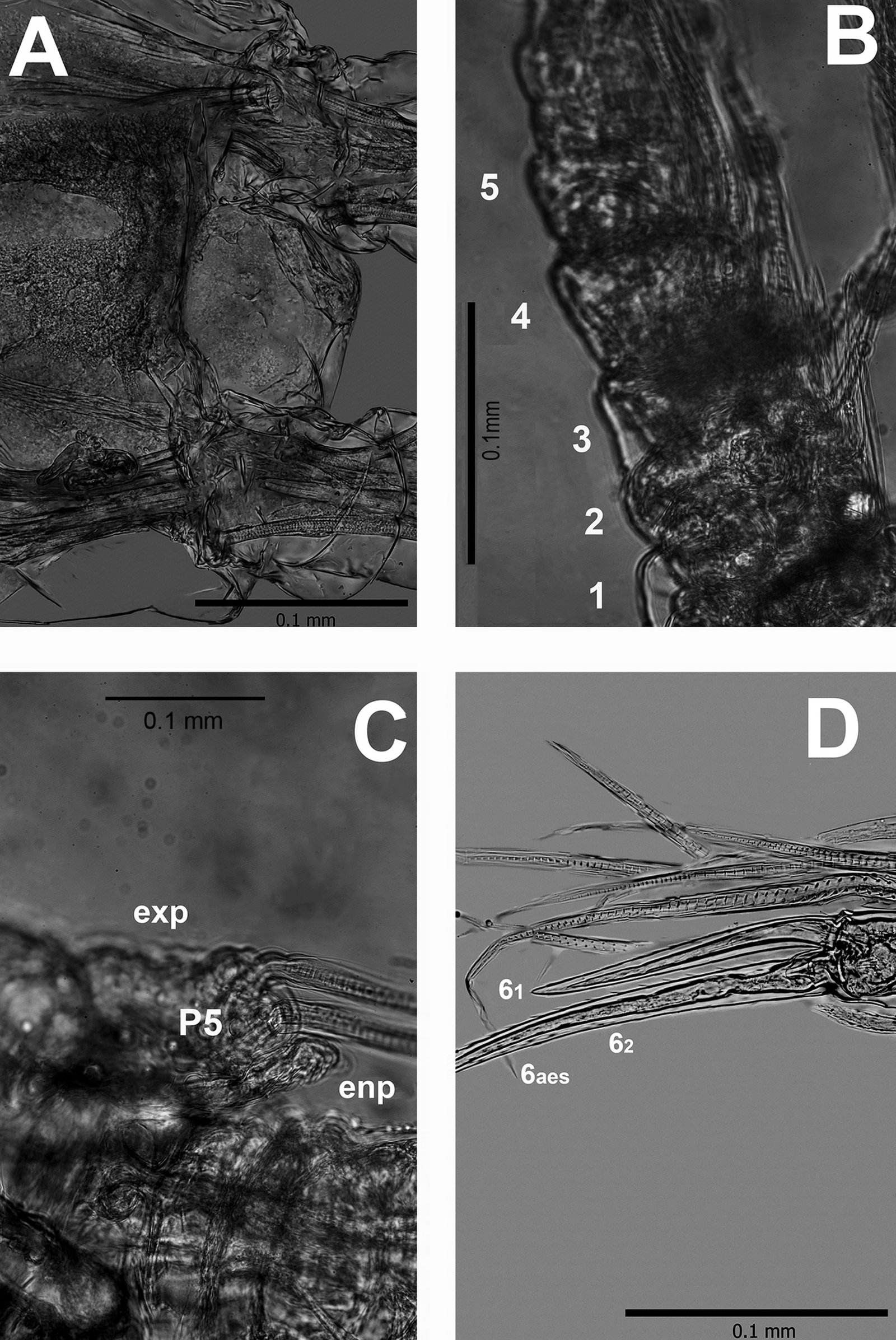
We conducted field collections during May and September 2015-2018 in various ecosystems, including tropical deciduous forest, oak-pine, and tropical forest within the state of Colima, at elevations ranging from 47 to 2,200 m. (Table 2). Logistically, we leveraged our collection sites with the help of private landowners who gave us access to their property. Specimen collection methods included direct collection during the day by moving objects on the ground or by ultraviolet detection during the night. To preserve specimens, ethyl alcohol at 90% was used and stored at -80 °C. Each specimen lot carried a label with coordinates and locality information. We obtained scorpions from 14 localities and sequenced 18 samples of Centruroides from Colima (Table 2).
Table 1
List of families, subfamilies, genera, and species recorded in the state of Colima. *Species of Centruroides cited by Ponce-Saavedra et al. (2016). The species in bold font were not found in Colima in this study. Numbered species were reported by González-Santillán et al. (2019).
11. Vaejovis monticola Sissom, 1989 12. Vaejovis sp. mexicanus group
Figure 1. Map of the west coast of Mexico. A, Orographic and hydrographic elements of Colima (COL) and the surrounding states of Jalisco (JAL) and Michoacán (MIC). 1, Manantlán Sierra; 2, massive Cerro Grande; 3, Colima Volcano; 4, Marabasco or Cihuatlán River; 5, Armería River; 6, Coahuayana River. B, Biogeographical provinces (Morrone et al., 2017). Area within the green line corresponds to the Sierra Madre del Sur and north Colima Volcano Trans Mexican Volcanic Belt province (Morrone et al., 2017) —notice that both provinces are connected in Colima. Area outside the green line corresponds to the Pacific Lowlands province (Morrone et al., 2017). Orographic components are indicated in gray scale from light low elevation to dark high elevation.
Table 2
Collection sites of the scorpion species used in this study. The number within parenthesis after the species name is the number of samples processed from this locality and included in the phylogenetic analyses as terminals. Superscript numbers indicate sources of sequences as follows: 1Bolaños et al. (2019), 2Esposito et al. (2018), 3Esposito and Prendini (2019), 4González-Santillán and Prendini (2015). Cells filled in grey color are samples obtained from GenBank.
Species
Municipality
Locality
Latitude
Longitude
Elevation
Thorellius cristimanus (2)
Comala
La Yerbabuena
19°27′59.55″
-103°41′46.64′′
1,358 m
Centruroides ornatus (2)
Comala
Agosto
19°23′51.74′′
-103°44′03.08′′
1,076 m
Thorellius cristimanus
*Centruroides tecomanus2
Colima
Comunidad La Capacha
19°04′58.40′′
-103°41′24.67′′
656 m
*Centruroides tecomanus2
Colima
Tepames
19°06′22.8′′
-103°59′11.07′′
450 m
Thorellius intrepidus (3)
Coquimatlán
El Palapo
19°11′54.6′′
-103°54′50′′
275 m
*Centruroides tecomanus2
Cuauhtémoc
Camino a Altozano
19°18′30.19′′
-103°40′23.83′′
789 m
Thorellius cristimanus
*Centruroides tecomanus1(2)
Cuauhtémoc
Ocotillo
19°20′00.00′′
-103°39′02.00′′
895 m
*Centruroides tecomanus2
Ixtlahuacán
San Gabriel
18°54′24.48′′
-103°44′05.61′′
462 m
Mesomexovis aff. occidentalis
Centruroides hirsutipalpus
Minatitlán
Minatitlán
19°23′01.73′′
-104°03′35.19′′
703 m
Centruroides possanii (2)
Minatitlán
Terrero
19°26′35.94′′
-103°57′05.67′′
2,200 m
Centruroides possanii (2)
Minatitlán
Mirador el Filete
19°26′40′′
-103°58′10′′
2,137 m
Vaejovis sp. (mexicanus group)
Centruroides sp. 2 (2)
Manzanillo
La central
19°08′38.14′′
-104°26′04.10′′
47 m
*Centruroides tecomanus1
*Centruroides tecomanus2
Centruroides sp. 1
Tecomán
Chanchopa
18°51′58.55′′
-103°44′10.10′′
41 m
Thorellius intrepidus (2)
Villa de Álvarez
Rancho Blanco
19°14′24.71′′
-103°45′49.45′′
455 m
Thorellius intrepidus4
La Huerta (Jal.)
Estación de Biología Chamela
19°30′14.15′′
-105°2′16.50′′
33 m
Centruroides elegans (2)
Centruroides suffusus
Durango (Dgo.)
El Salto, 50 km E Durango
23°45′51.41′′
-105°19′49.16′′
2,847 m
Centruroides limpidus
Iguala (Gro.)
Iguala
18°20′11.87′′
-99°29′29.65′′
823 m
Centruroides sculpturatus
Cumpas (Son.)
18 km NE de Nacozari
30°16.473′
-109°50.070′
930 m
Centruroides noxius
Pantanal (Nay.)
Pantanal
21°25′24.42′′
-104°50′47.89′′
921 m
Centruroides huichol
Nayarit
–
–
–
–
Centruroides infamatus115scrp
Guanajuato (Gto.)
Guanajuato
–
–
–
Centruroides ornatus2LP1822
Tandamangapio (Mich.)
Los Tabanos
19.9749°
– 102.84226°
223 m
Centruroides ornatus3 2003
Michoacán
–
–
–
–
*Centruroides tecomanus11 25scrp
Comala (Col.)
–
–
–
–
*Centruroides tecomanus12 2007
Michoacán
–
–
–
–
Mesomexovis occidentalis4 LP 7056
Acapulco (Gro.)
Cumbres de Llano Largo
16°49.505
-99°49.9990
317 m
Table 2. Continued
Species
Municipality
Locality
Latitude
Longitude
Elevation
Mesomexovis spadix4LP6373
León (Gto.)
San Antonio de Padua
20°34.5170
-100°57.217
–
Mesomexovis subcristatus4LP 2049
Tehuacán (Pue.)
Tehuacán, 2 km east
18°24.0020
-97°22.8670
1435 m
Thorellius cristimanus4LP 5325
Álvaro Obregón (Mich.)
Álvaro Obregón
19°02.3100
-102°58.405
462 m
Thorellius cristimanus4LP 6551
Coquimatlán (Col.)
Road to Coquimatlán, km 71
19°06.7750
-103°51.1850
336 m
Thorellius intrepidus4LP 6377
Comala (Col.)
Comala
19°19.000
-103°45.0000
–
Thorellius intrepidus4LP 6379
Colima (Col.)
Los Ortices
19°06.0468
-103°44.0226
343 m
Vaejovis carolinianus4LP 1576
South Carolina
–
–
–
–
Vaejovis pequeno4LP 6308
Soyopa (Son.)
Sierra El Encinal, 9 km from crossroad on Highway Mex 16 to El Encinal
28°35.4120
-109°27.1480
380 m
Vaejovis rossmani4LP 2027
Hidalgo (Tams.)
Conrado Castillo
23°56.01735
-99°28.04817
–
*Genetically differentiated species.
We also included 8 species from outside the state to test the presence of C. elegans, C. infamatus, and C. limpidus reported previously in the literature (Ponce-Saavedra et al., 2016); C. noxius and C. suffusus for comparative purposes and samples of C. tecomanus and C. ornatus from GenBank with a total of 31 specimens of the genus Centruroides within 4 species groups. Unlike buthids, we obtained 12 samples of vaejovids to include in this analysis. To evaluate the identity of vaejovids, we used the BLAST® suite (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to search for similar sequences deposited in the nucleotide collection database at NCBI. Thorellius has been revised recently (González-Santillán & Prendini, 2018). Therefore, several DNA sequences are available, and fewer sequences of Mesomexovis sp. and Vaejovis sp. were available in GenBank, as they are still unrevised. Using the genetic markers as queries, we obtained 10 additional samples. The total number of taxonomic specimens used for these analyses was 53 (Tables 2, 4).
Genomic DNA was extracted from the legs or pedipalp of specimens using Qiagen Dneasy/trisol method Tissue Kits or a DNAzol Genomic DNA isolation Reagent kit (Molecular Research Center INC, Cincinnati, Oh). We amplified 3 mitochondrial markers, 12S rDNA, 16S rDNA, and the barcode COI, and the nuclear marker 28S rDNA. We performed the Polymerase Chain Reaction with the following thermal profile: an initial denaturation step (3 min at 94°C) followed by 35 cycles including denaturation at 94°C for 30 s, annealing (46-55°C) for 30 s, and extension at 72°C for 30 s, with a final extension step at 72°C for 7 min. The PCR reaction was conducted using PureTaq-Ready-To-Go PCR Beads (GE Healthcare), 2 µl of DNA template, 21 µl of DNA grade H2O, and 1 µl of each direction primer listed in Table 3. We verified PCR products with a 1% agarose-TBE electrophoresis gel stained with CYBR Safe. For purification of the amplified products, we used Ampure Magnetic Beads (Beckman-Coulter) and re-suspended in 40 µl DNA grade water by using a Beckman Coulter Biomek NX 18 robot. Each 8 µl cycle-sequenced reaction mixture included 1 µl of Big Dye, 1 µl of Big Dye Terminating buffer, 1 µl of 3.2 pm primer, and 5 µl of gene amplification product. Cycle-sequenced products were purified with CleanSeq magnetic beads on a Biomex NX robot. Products were re-suspended in EDTA, and 33 µl were processed in an Applied Biosystems, Inc. Prism 3730xl automated DNA sequencer. These products were sequenced with the same primer pairs used for amplification at the Laboratorio de Secuenciación Genómica de la Biodiversidad, at Instituto de Biología and Unidad de Síntesis y Secuenciación de DNA, Instituto de Biotecnología, UNAM. The sequences were edited using Sequencher® version 5.4.6.
Table 3
List of primers used to amplify molecular markers.
Name
Sequence
Reference
12S rDNA
12SAI
AAACTAGGATTAGATACCCTATTAT
Kocher et al. (1989)
12SBI
AAGAGCGACGGGCGATGTGT
Kocher et al. (1989)
16S rDNA
16SA
CGCCTGTTTATCAAAAACAT
Simon et al. (1994)
16SB
CTCCGGTTTGAACTCAGATCA
Simon et al. (1994)
COI
HCO
TAAACTTCAGGGTGACCAAAAAATCA
Folmer et al. (1994)
LCO1
GGTCAACAAATCATAAAGATATTGG
Folmer et al. (1994)
28S rDNA
28SA
GACCCGTCTTGAAGCACG
Nunn et al. (1996)
28SBout
CCCACAGCGCCAGTTCTGCTTACC
Prendini et al. (2005)
Each genetic fragment was aligned separately for all terminals with MAFFT using the online server (https://mafft.cbrc.jp/alignment/server/). Since the number of nucleotides per gene was similar, we used the G-ins-i iterative refinement method, as recommended elsewhere (Katoh et al., 2019; Kuraku et al., 2013), and other parameters were kept default. To select the best fit of the substitution model per partition and conduct the phylogenetic analyses, we used IQ-TREE version 2 (Kalyaanamoorthy et al., 2017; Nguyen et al., 2015) and we estimated branch support with 1,000 replicates of the ultrafast bootstrap (UFBOOT) algorithm (Hoang et al., 2018). Furthermore, each genetic marker was analyzed individually to explore its phylogenetic signal and contribution to the final topology. We conducted concatenated and partitioned analyses, handling all matrices in Mesquite (Maddison & Maddison, 2023). For the COI partition we explored the best codon partition per site, but the results had no effect on the topology. Additionally, to evaluate each marker and nucleotide site within each marker, we calculated the gene (GCF) and site (SCF) concordance factors on the topology that we emphasized in the discussion (Mihn et al., 2020).
Distributional records and maps. We obtained records with geographical coordinates of the scorpion species treated here via the Global Biodiversity Information Facility (GBIF) from the InDRE, responsible in Mexico for epidemiological vigilance (Huerta-Jiménez, 2018), and the records published by Ponce-Saavedra et al. (2015). These records were the basis for the species distribution maps depicted in figures 3 to 6. We used the program QGIS 3.16.6-Hannove (QGIS, 2021) to create the distributional maps. The topological model with the data was from Jarvis et al. (2008), and to draw the political boundaries we used shapefiles obtained from Conabio. The biogeographic regionalization of Mexico into provinces and districts follows Morrone et al. (2017) and Morrone (2019).
Results
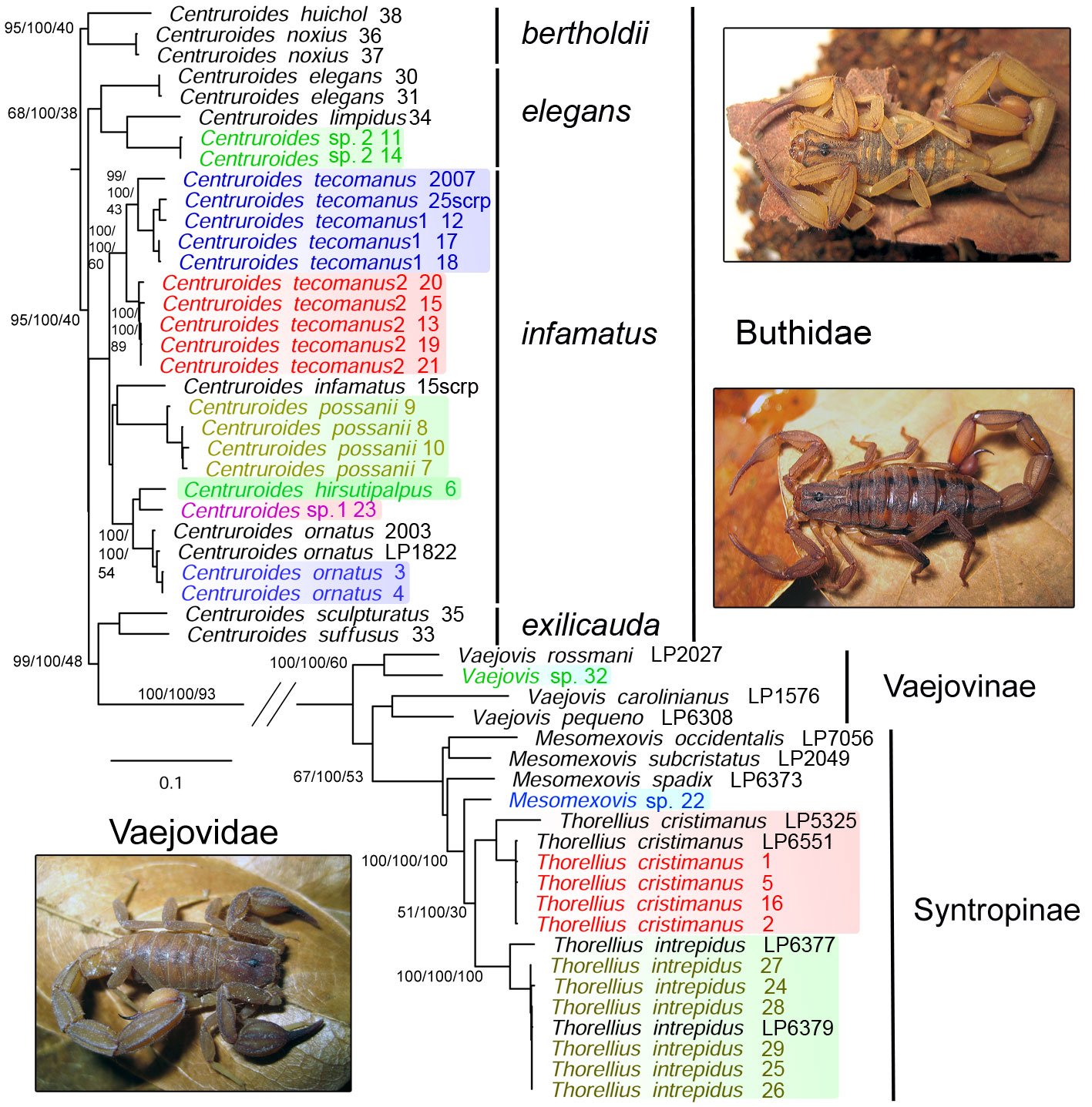
The gene fragments that we obtained are listed in Table 4 and the main statistics of the alignment and concatenated matrix are in Table 5. Our topology produced a clade representing members of the family Buthidae and another clade representing Vaejovidae (Fig. 2). Within buthids, the first clade included C. huichol and C. noxius, component species of the bertholdii species group (Ponce-Saavedra & Francke, 2019), supported by 95% UFBOOT, 100% GCF, and 40% SCF. The next clade included C. elegans, C. limpidus, and a putative undescribed species with lower support values of 68%, 100%, and 38%, respectively, including members of the elegans group. Although with low support, the bulk of species appeared in the third clade comprising species within the infamatus group, with C. tecomanus, C. infamatus, C. possanii, C. hirsuticauda, C. ornatus, and 2 putative new species. The last clade included C. suffusus and C. sculpturatus, whichPonce-Saavedra and Francke (2019) circumscribed within the infamatus and the elegans species group, respectively (Fig. 2). However, the overall topology retrieved herein is concordant with the North American clade of the genus Centruroides (Esposito & Prendini, 2019).
Table 4
Mitochondrial genetic markers 16S, COI, and 12S and nuclear 28S information for samples analyzed in the study. Dash (-) symbols indicate unavailable sequences.
Species
NCBI:txid
16S
12S
COI
28S
Centruroides elegans
217897
Cele_30_16S (PP295377)
Cele_30_12S (PP295301)
Cele_30_COI (PP356615)
Cele_30_28S (PP295328)
Cele_31_16S (PP295378)
Cele_31_12S (PP295302)
Cele_31_COI (PP355194)
Cele_31_28S (PP295329)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Centruroides hirsutipalpus
–
Chir_06_16S (PP295353)
Chir_06_12S (PP295277)
Chir_06_COI (PP356614)
Chir_06_28S (PP295310)
Centruroides huichol
2911785
Chui_38_16S (PP295385)
Chui_38_12S (PP295309)
Chui_38_COI (PP356613)
Chui_38_28S (PP295335)
Centruroides infamatus
42200
MF134694
–
MF134798
MF134763
Centruroides limpidus
29941
Clim_34_16S (PP295381)
Clim_34_12S (PP295305)
Clim_34_COI (PP356612)
Clim_34_28S (PP295332)
Centruroides noxius
6878
Cnox_36_16S (PP295383)
Cnox_36_12S (PP295307)
Cnox_36_COI (PP356611)
Cnox_36_28S (PP295333)
Cnox_37_16S (PP295384)
Cnox_37_12S (PP295308)
Cnox_37_COI (PP356610)
Cnox_37_28S (PP295334)
Centruroides ornatus
2338500
Corn_03_16S (PP295350)
Corn_03_12S (PP295274)
Corn_03_COI (PP355195)
Corn_03_28S (PP295324)
Corn_04_16S (PP295351)
Corn_04_12S (PP295275)
Corn_04_COI (PP356609)
Corn_04_28S (PP295325)
KY981895
KY981799
–
KY982086
MK479042
MK478991
MK479195
MK479144
Centruroides possanii
–
Cpos_10_16S (PP295357)
Cpos_10_12S (PP295281)
–
Cpos_10_28S (PP295314)
Cpos_07_16S (PP295354)
Cpos_07_12S (PP295278)
Cpos_07_COI (PP355196)
Cpos_07_28S (PP295312)
Cpos_08_16S (PP295355)
Cpos_08_12S (PP295279)
Cpos_08_COI (PP356608)
Cpos_08_28S PP295313
Cpos_09_16S (PP295356)
Cpos_09_12S (PP295280)
Cpos_09_COI (PP356607)
Cpos_09_28S (PP295323)
Centruroides sculpturatus
218467
Cscu_35_16S (PP295382)
Cscu_35_12S (PP295306)
Cscu_35_COI (PP356606)
Cscu_35_28S (PP295331)
Centruroides sp. 1
3103037
Csp1_23_16S (PP295370)
Csp1_23_12S (PP295294)
Csp1_23_COI (PP356604)
Csp1_23_28S (PP295311)
Centruroides sp. 2
Csp2_11_16S (PP295358)
Csp2_11_12S (PP295282)
Csp2_11_COI (PP356603)
Csp2_11_28S (PP295327)
Csp_14_16S (PP295361)
Csp_14_12S (PP295285)
Csp_14_COI (PP356605)
Csp_14_28S (PP295326)
Centruroides suffusus
6881
Csu_33_16S (PP295380)
Csu_33_12S (PP295304)
–
Csu_33_28S (PP295330)
Centruroides tecomanus1
1028682
Cte1_12_16S (PP295359)
Cte1_12_12S (PP295283)
Cte1_12_COI (PP356602)
Cte1_12_28S (PP295315)
Cte1_17_16S (PP295364)
Cte1_17_12S (PP295288)
Cte1_17_COI (PP355197)
Cte1_17_28S (PP295320)
Cte1_18_16S (PP295365)
Cte1_18_12S (PP295289)
Cte1_18_COI (PP356601)
Cte1_18_28S (PP295318)
Centruroides tecomanus2
Cte2_13_16S (PP295360)
Cte2_13_12S (PP295284)
Cte2_13_COI (PP356600)
Cte2_13_28S (PP295316)
Cte2_15_16S (PP295362)
Cte2_15_12S (PP295286)
Cte2_15_COI (PP355198)
Cte2_15_28S (PP295317)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Cte2_19_16S (PP295366)
Cte2_19_12S (PP295290)
Cte2_19_COI (PP356599)
Cte2_19_28S (PP295319)
Cte2_20_16S (PP295367)
Cte2_20_12S (PP295291)
Cte2_20_COI (PP356598)
Cte2_20_28S (PP295321)
Cte2_21_16S (PP295368)
Cte2_21_12S (PP295292)
Cte2_21_COI (PP356597)
Cte2_21_28S (PP295322)
Centruroides tecomanus
MF134695
–
MF134799
MF134757
MK479053
MK479002
MK479206
MK479156
Mesomexovis sp.
–
Mesp_22_16S (PP295369)
Mesp_22_12S (PP295293)
Mesp_22_COI
Mesp_22_28S (PP295337)
Mesomexovis occidentalis
1532992
KM274362
KM274216
KM274800
–
Mesomexovis spadix
1532994
KM274221
KM274367
KM274805
KM274659
Mesomexovis subcristatus
1532995
KM274368
KM274222
KM274806
KM274660
Thorellius cristimanus
1533000
Tcri_01_16S (PP295348)
Tcri_01_12S (PP295272)
–
Tcri_01_28S (PP295338)
Tcri_16_16S (PP295363)
Tcri_16_12S (PP295287)
–
Tcri_16_28S (PP295336)
Tcri_02_16S (PP295349)
Tcri_02_12S (PP295273)
–
Tcri_02_28S (PP295339)
Tcri_05_16S (PP295352)
Tcri_05_12S (PP295276)
–
Tcri_05_28S (PP295340)
KM274420
KM274274
KM274858
KM274712
KM274422
KM274276
KM274860
KM274714
Thorellius intrepidus
1533001
Tint_24__16S (PP295371)
Tint_24_12S (PP295295)
Tint_24_COI (PP355193)
Tint_24_28S (PP295341)
Tint_25_16S (PP295372)
Tint_25_12S (PP295296)
Tint_25_COI (PP356616)
Tint_25_28S (PP295342)
Tint_26_16S (PP295373)
Tint_26_12S (PP295297)
Tint_26_COI (PP356617)
Tint_26_28S (PP295343)
Tint_27_16S (PP295374)
Tint_27_12S (PP295298)
Tint_27_COI (PP355192)
Tint_27_28S (PP295344)
Tint_28_16S (PP295375)
Tint_28_12S (PP295299)
Tint_28_COI (PP356618)
Tint_28_28S (PP295345)
Tint_29_16S (PP295376)
Tint_29_12S (PP295300)
Tint_29_COI (PP356619)
Tint_29_28S (PP295346)
KM274424
KM274278
KM274862
–
KM274425
KM274279
KM274863
KM274717
Vaejovis sp.
–
Vasp_32_16S (PP295379)
Vasp_32_12S (PP295303)
Vasp_32_COI (PP356620)
Vasp_32_28S (PP295347)
Vaejovis carolinianus
33322
KM274289
KM274143
KM274727
KM274581
Vaejovis pequeno
1532951
KM274293
KM274147
KM274731
KM274585
Vaejovis rossmani
1532952
KM274294
KM274148
KM274732
KM274586
Table 5
Main statistics of site information per alignment, including parsimony informative sites, (P info) and the concatenated (Conca) alignment of all partitions in a final matrix. The concatenated alignment had 6% missing data.
Partitions
Terminals/ nucleotide
Site information per alignment
Substitution
P info
Invariable
Unique
Constant
Model
16S
53/523
209
285
258
285
K3Pu+F+I+G4
28S
52/538
39
494
96
494
K2P+I
12S
51/393
153
210
212
210
TPM2+F+G4
COI
45/679
220
420
220
420
TIM2+F+G4
Conca
53/2133
621
1,409
786
1,409
simultaneous
Figure 2. Phylogenetic tree from an analysis of 4 concatenated genetic markers, 3 mitochondrial (12S, 16S, and COI) and 1 nuclear (28S). The topology shows families, subfamilies, and species groups. Numbers beside nodes indicate ultrafast bootstrap/genetic concordance factor/site concordance factor. Colored species represent the 11 species found in our collection within the study area and samples grouping with them distributed inside or outside Colima. Scorpion photos: upper right, Centruroides tecomanus from El Palapo, adult male; middle, Centruroides hirsutipalpus from Sierra de Minatitlán, adult female; lower left, Thorellius intrepidus from El Palapo, adult male.
The Vaejovid clade received full support, except SCF (60%), which included 3 genera: Vaejovis, Mesomexovis, and Thorellius (Fig. 2). The genera occurred in a topology that overall resembles that of González-Santillán and Prendini (2013, 2015); while Vaejovis is a genus within Vaejovinae, Mesomexovis and Thorellius are part of the Syntropinae subfamily. Within Syntropinae, only Thorellius was monophyletic, and T. intrepidus received full support. Additionally, we identified 2 putative new species belonging to the genera Vaejovis and Mesomexovis.
Distributional maps and species geographical ranges. We mined 6,965 records of Mexican scorpions from the InDRE database but retained only 1,000 of the species treated herein. We filtered the records using previously published identified species by specialists and the distribution of the species cited for Colima (González-Santillán & Prendini, 2013, 2018; Lourenço & Sissom, 2000; Ponce-Saavedra et al., 2016; Sissom, 2000).
Figure 3. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides elegans (circles), records north of Sierra de Manantlán may be misidentifications; B, distribution of Centruroides tecomanus (circles), records in Guerrero (GRO), Guanajuato (GTO), Jalisco (JAL), and Nayarit (NAY) may be misidentifications; Centruroides tecomanus 1 green cross, and Centruroides tecomanus 2 pink crosses. State abbreviations: AUG, Aguascalientes; COL, Colima; MEX, Estado de México; QRO, Querétaro; SLP, San Luis Potosí; ZAC, Zacatecas.
Figure 4. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database and for B additional records from Ponce-Saavedra et al. (2015). A, Distribution of Centruroides infamatus (circles); B, distribution of Centruroides ornatus (circles), our sample in Colima (cross).
Figure 5. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides hirsutipalpus (red cross), Centruroides sp. 1 (blue cross), Centruroides sp. 2 (pink cross), and Centruroides possanii (green crosses); B, distribution of Mesomexovis aff. occidentalis sequences in this work and a sample from Chamela Jalisco recorded by González-Santillán (2004), conspecific with our samples in green crosses, Vaejovis sp. (mexicanus group) (pink cross).
The distribution of Centruroides species appears to be restricted by physiographical elements. Centruroides elegans and C. tecomanus are restricted to the coastal lands of Jalisco and Colima, respectively (Fig. 3). The Sierra Madre del Sur appears to be the main barrier to the north of their distribution. The records of these species overlap broadly with a visible gap created by the Sierra de Minatitlán (Fig. 3B) either by subsampling or by an effective geographic barrier.
Centruroides ornatus and C. infamatus, on the other hand, are restricted to the Transmexican Volcanic Belt (TVB) (Fig. 4). While the former species has an entire distribution within this province (Fig. 4B), the latter seems to be distributed in patches, one to the north, along the border of the Chihuahuan desert and the TVB, and the second to the south, between the Balsas Depression and the TVB (Fig. 4A).
Centruroides possanii appears to be a microendemic scorpion species component of the fauna restricted to a massive karstic lone mountain, Cerro Grande (González-Santillán et al., 2019). Two putative undescribed species of Centruroides occupy the extreme east and west of the coastal line of Colima, and C. hirsutipalpus appears restricted to the Sierra Minatitlán (Fig. 5A).
Of the vaejovid species, thus far, the only records for Mesomexovis sp. are in Colima, albeit there is one conspecific record in Chamela, Jalisco (González-Santillán, 2004). The putatively undescribed species of Vaejovis appears restricted to Cerro Grande (Fig. 5B), like Centruroides possanii, potentially another microendemic species. Thorellius intrepidus and T. cristimanus are widely distributed within Colima and across the TVB, the Balsas Depression, and the Sierra Madre del Sur (Fig. 6).
Discussion
Of the 2 families found in Colima, Buthidae comprise 7 species of Centruroides,becoming the most diverse (Table 1, Fig. 2). Our multigene analyses suggest that the previously recorded species C. elegans, C. infamatus, and C. limpidus reported by Ponce-Saavedra et al. (2016) might not be part of the scorpion fauna of Colima, as we demonstrate in the following sections.
Figure 6. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Thorellius intrepidus greensquares, our samples in Colima pink crosses; B, distribution of Thorellius cristimanus greensquares, our samples in Colima pink crosses.
Centruroides elegans and C. limpidus were not found in Colima. Centruroides elegans has an obscure taxonomic history. Firstly, its original description is too general and never indicated a precise type locality, but “Mexico” (Fet & Lowe, 2000). Secondly, although some taxonomic works clarified its former subspecies, the nominal taxon identity of C. elegans remains ambiguous. While Lourenço and Sissom (2000) suggested that this species is distributed in Jalisco, later, González-Santillán (2004) concluded that its distributional limits have never been defined with precision. The 2 exemplars of C. elegans collected in Chamela, Jalisco, grouped with members of the elegans group, sisters to C. limpidus from Iguala, Guerrero, and these in turn, were sister to our 2 exemplars identified as Centruroides new sp. 2 11 and 14 from La Central, in the municipality of Manzanillo (Figs. 2). The distributional data from InDRE of C. elegans is limited right at the northern border of Colima, except for 9 records away from that geographic barrier (Fig. 3A), which we hypothesized as a misidentification due to their position outside the Pacific coastline. Our fieldwork produced no sample conspecific to C. elegans but did produce other genetically distant exemplars. Until denser sampling along the northern border of Colima is conducted, we conclude that C. elegans and by corollary C. limpidus might not be part of the scorpion fauna distributed in Colima as suggested by Ponce-Saavedra and Francke (2013) and Ponce-Saavedra et al. (2016).
Centruroides infamatus may not be part of the Colima scorpion fauna. Centruroides infamatus is another obscure taxon with an ambiguous distributional pattern from published data. Despite its inclusion in phylogenetic analyses (Quijano-Ravel et al., 2019; Towler et al., 2001), its taxonomic circumscription and distribution have never been clarified. Among other problems, the original description indicates “type locality unknown” (Fet & Lowe, 2000). Hoffmann (1932) studied the scorpions from Mexico and realized that the specimens from Michoacán agreed with the original description of the species and proposed that its distribution was expanded to Central Mexico from the Pacific Coast in Sinaloa and Colima to Guanajuato. We included 2 exemplars of C. infamatus, 1 from León, Guanajuato (C. infamatus 15 scrp), and 1 terminal from Tandamangapio municipality, Michoacán (C. infamatus LP1822; Esposito et al., 2018). However, the terminal LP1822 grouped with exemplars of C. ornatus (Fig. 2) and Tandamangapio is approximately 16 km from Sahuayo and Jiquilpan, 2 localities recorded in the redescription of C. ornatus (Ponce-Saavedra et al., 2015).
We plotted 612 records from the InDRE database of C. infamatus to compare the distribution of C. ornatus (Fig. 4A). Although northern Michoacán is almost exclusively occupied by C. ornatus, Jalisco and Michoacán present an overlap with C. infamatus (Fig. 4). Additionally, the InDRE database contains records of C. infamatus from Oaxaca, Puebla, Nayarit, Durango, and Sinaloa (not shown), most likely occupied by other species, implying that most records are misidentifications. The species that morphologically could be confused with C. infamatus in the states of Oaxaca and Puebla are C. baergi, C. nigrovariatus, or C. rodolfoi because the overall base color, body size and carapace pigmentation are similar among the species. The same morphological features are present in specimens from Nayarit and Durango, however, the identity of the populations in these two states have never been studied with a molecular approach and may represent a geographically distinct species.
Nevertheless, recently Ponce-Saavedra et al. (2022) described C. baldazoi for Sinaloa, related morphologically with C. infamatus, potentially a species with which it can be mistaken. Although Ponce-Saavedra and collaborators included C. infamatus in the distribution of Sinaloa and Colima, they failed to indicate precise localities. In summary, the distribution of C. infamatus remains unsolved, attested by the wide distribution reported in the InDRE database. Thus, the infamatus species complex requires comprehensive molecular analyses to delimit its taxonomic circumscription and geographical distribution. On the other hand, our samples of C. ornatus 3 and 4 grouped with C. ornatus LP1822 and with C. ornatus 2003 from Morelia, Michoacán (Esposito & Prendini, 2019), forming a monophyletic group (Fig. 2). Since the samples LP1822 and 2003 lie within the area of C. ornatus proposed by Ponce-Saavedra et al. (2015), we concluded that our exemplars are conspecific with C. ornatus and expanded the distribution southwards, drawing a distributional border in northern Colima (Fig. 4B).
In conclusion, we propose that C. infamatus might not be present in Colima, but further sampling and analyses are needed. Despite the morphological similarity between C. infamatus and C. ornatus, our molecular analysis indicates genetic differences and may present a distinctive distributional pattern yet to be drawn with more samples.
Centruroides tecomanus is a species complex. Due to their morphological similarity with C. limpidus, C. tecomanus was described as C. limpidus tecomanus (Hoffmann, 1932), and the author delimited C. tecomanus distribution to the lowlands of Colima and surmised that its distribution extended to the south along the coastline of Michoacán. But, most importantly, Hoffmann mentioned that on the northern coastline, C. elegans substitutes C. tecomanus. In contrast, the distribution recorded in the InDRE database for C. tecomanus implies that these 2 species present a wide area of sympatry on the coast of Jalisco (Fig. 3).
Ponce-Saavedra et al. (2009) proposed C. tecomanus as a “bona fide species” and assumed its distribution includes the coastline of Michoacán, following Hoffmann (1932). In their molecular and morphological analyses, the authors never included exemplars from Tecomán, Colima, the type locality for the species, assuming that the specimens from Michoacán were conspecific with the populations of Tecomán. Furthermore, Quijano-Ravell et al. (2010) extended the distribution of C. tecomanus to 4 localities in Guerrero and a similar number in Jalisco in a montane area.
The phylogenetic tree presented here substantiates 2 clades of C. tecomanus within the monophyletic infamatus species group (Fig. 2). We hypothesize that these clades may represent potentially distinct sympatric species, emphasizing that our topology obtained full support for these clades with multiple samples. Centruroides tecomanus2 appears to be more common and widely distributed in Colima, whereas C. tecomanus1 is less common, with only 2 localities grouped with Centruroides tecomanus 2007 from Michoacán and from the municipality of Comala, Colima, Centruroides tecomanus (Fig. 3B). Our results suggest that the populations distributed in Michoacán may represent a cryptic, undescribed species; consequently, the exemplars reported in Guerrero (Quijano-Ravell et al., 2010) are unlikely to be conspecific with C. tecomanus. The authors’ discovery of new populations in Jalisco and Guerrero were based entirely on their analyses of morphological characters, precisely the most common way of confusing cryptic species.
Considering that the montane border between Colima and Michoacán is occupied by Centruroides romeroi Quijano-Ravell, de Armas, Francke, Ponce-Saavedra, 2019, it is fair to assume that C. tecomanus1 inhabits coastal ranges following the coastline of Michoacán to the Lázaro Cárdenas delta, where it may be substituted by Centruroides bonito Quijano-Ravell, Teruel, Ponce-Saavedra, 2016 or even other undescribed species. The Balsas River has been proposed to be a geographical barrier for several epigean arachnids, such as Amblypygi and Theraphosidae (Mendoza & Francke, 2017; Schramm et al., 2021) and for small mammals (Ruiz-Vega et al., 2018)
Noteworthy is the locality La Central, right on the border between Colima and Jalisco, a few kilometers southeast of the Marabasco River (Fig. 1A), where we collected 3 putatively different species of Centruroides, the 2 morphotypes of C. tecomanus and Centruroides sp. 2, retrieved within the elegans group (Fig. 2). These findings illustrate the complicated patterns of diversification within this buthid genus in Colima. From a taxonomic perspective, if the identity of C. tecomanus is to be clarified, it is now imperative to analyze molecularly exemplars from Tecomán, Colima, which is the type locality of this species.
Thorellius species exhibit a more restricted distribution in Colima. Thorellius intrepidus and T. cristimanus are widely distributed in several states of the Pacific Lowlands (Fig. 6). However, in a recent revision of the genus, 2 species were described, Thorellius wixarika González-Santillán and Prendini, 2018 and Thorellius tekuani González-Santillán and Prendini, 2018 (Fig. 3 of González-Santillán and Prendini, 2018), that are relevant to these analyses. The former occupies the northwestern territory in Nayarit and Jalisco,whereas the latter inhabits the Balsas Depression of Estado de México, Guerrero, and Michoacán. This observation suggests that the InDRE database contains several misidentifications of both T. intrepidus and T. cristimanus. In fact, the T. wixarika and T. tekuani description was in 2018, and the InDRE database is several years older, hence the complete absence of records. Thus, T. intrepidus inhabits Aguascalientes, Colima, Guanajuato, Jalisco, and Michoacán (Fig. 6A); and T. cristimanus, Colima and Jalisco (González-Santillán & Prendini, 2018) (Fig. 6B). From an ecological point of view, we noticed that the InDRE database only has records with elevations below 700 m, implying that these species prefer tropical to subtropical climates in Colima.
González-Santillán (2004) reported Mesomexovis aff. occidentalis as a putative new species for the Biological Station Chamela, Jalisco. Furthermore, González-Santillán and Prendini (2015) conducted a phylogenetic analysis using morphology and mitochondrial and nuclear DNA, resulting in a topology that suggested that this species was not conspecific to Mesomexovis occidentalis, as Williams (1986)identified it. López-Granados (2019) found further morphological evidence to separate this species, but the evidence was never published. The importance of that work is that, for the first time, exemplars of Mesomexovis from Colima that were conspecific with our samples were included in a phylogenetic analysis. Once more, we demonstrate that Mesomexovis sp. 22 is not conspecific with M. occidentalis using molecular evidence, and it requires a formal separation and description (Fig. 2). Unlike Thorellius species in Colima, Mesomexovis is a widespread species inhabiting tropical to montane habitats with a wider range of elevation (Fig. 5B).
The mexicanus group was recently revisited in a monograph with the delimitation of other species groups and the description of 5 new species (Contreras-Félix & Francke, 2019). The paucity of sequences for the mexicanus group in the NCBI database only permitted retrieval of loci for Vaejovis rossmani LP 2027, a species inhabiting the Sierra Madre Oriental in the states of Tamaulipas and Nuevo León and treated in the Contreras-Félix and Francke (2019) monograph. Our analysis retrieved Vaejovis sp. 32 and Vaejovis rossmani LP 2027 together, which suggests membership of the mexicanus group of Vaejovis. Furthermore, Vaejovis sp. 32also matches the morphological diagnosis presented in Contreras-Felix and Francke (2019). Contreras-Félix et al. (2023) recorded Vaejovis santibagnezi Contreras-Felix and Francke, 2019 in Cerro Grande, where we collected our samples (Fig. 5B). However, the authors failed to present morphological evidence to justify this conclusion. We compared our specimens with the geographically closest V. monticola deposited in the CNAN and with V. santibagnezi and found significant morphological differences. Currently, we are preparing a contribution where we propose the description of a new species. Following this idea, we are inclined to think that, like C. possanii, Vaejovis sp. 32 is also microendemic because of its limitation to disperse throughout tropical valleys from the Cerro Grande massif to other mountain ranges. Finally, we submit that such an evident geographical barrier may apply to several epigean, non-volant arthropods such as the scorpions.
Finally, 2 species absent in our fieldwork are Konetontli ilitchi González-Santillán and Prendini 2015 and Vaejovis janssi Williams, 1980. The latter is endemic to the Socorro Island, part of the Revillagigedo Archipelago (Williams, 1980), and K. ilitchi has been found only inside a cave in the vicinity of Coquimatlán, Colima (González-Santillán & Prendini, 2015). These 2 features in the distribution and biology of the species hindered the possibility of collecting and studying them. Future fieldwork may illuminate the taxonomy and distribution of these elusive species. Finally, Vaejovis monticola, another high elevation dweller of the mexicanus species group (Contreras-Felix & Francke 2019) was absent in our collecting trips. Although its type locality cited by Contreras-Felix and Francke (2019) indicates: “Jalisco, northern side of Nevado de Colima” we submit that it may not be on the southern side in Colima, but extensive fieldwork is needed to confirm this hypothesis.
Biogeographical and ecological considerations
There is strong evidence that the ancestors of the North American Centruroides originated in Gondwana and have dispersed to and diversified in that territory via land bridges, vicariance, and rafting over the Atlantic Ocean, over 50 to 20 Mya (Esposito & Prendini, 2019). However, the movement of this genus towards boreal latitudes within North America is not well understood. Although one could suppose that the colonization of such territories appears to be in pulses at different periods due to the overlap in the distribution of the elegans and infamatus species groups, it is beyond the scope of our study to establish a coherent explanation; besides, our data are incomplete to that end. One startling discovery is that the Colima scorpiofauna comprises 7 distinctive, at least genetically, species of Centruroides (Fig. 2). What biotic, abiotic, or ecological factors produce such species diversity? Is it a combination of all these factors? Is it a case of sympatric speciation? Morphologically, it is sometimes difficult to distinguish some of these species. For instance, the 3 species found in La Central were identified initially as C. tecomanus because they exhibit no morphological variation, at least in traditional characters shown in identification keys (Ponce-Saavedra et al., 2016). It is fundamental to conduct morphological studies with this scorpion community to obtain independent information to precisely delimit these species. Unlike other scorpion groups, Centruroides exhibit conservative morphology. For instance, if we compared Centruroides species to species of the Syntropinae subfamily, we find that syntropines are diverse in ecomorphotypic adaptations that promoted morphological diversification (González-Santillán & Prendini, 2013), while in Centruroides, the persistent errant lifestyle maintains morphology without significant variation. It is astonishing to observe, for instance, C. tecomanus living at sea level in tropical deciduous forests and compare it to C. possanii inhabiting pine-oak forests above 2,000 m elevation; as though, the same “Centruroides bauplan” can survive diametrically different environments following a persistent lifestyle. This final assertion leads us to think that the adaptative changes are in the physiology rather than in the morphology of Centruroides.
Thorellius and Mesomexovis (Syntropinae) are substrate-specialists, since the body plan, more robust and armed with setae and spinules on the legs, allow these species to scrape and dig galleries on clayey, fine-grained substrates (González-Santillán & Prendini, 2013, 2018). Such adaptations come with a drawback because the kind of soil restricts the presence of these scorpions; for instance, rocky or shallow soils represent unhospitable habitats for these species. Like other species in the mexicanus group, Vaejovis sp. 32 and Vaejovis monticola Sissom, 1989 are inhabitants of montane habitats within the intricate topography of Colima.
In summary, the scorpion assemblage distributed in Colima includes errant species such as Centruroides without exaggerated adaptations; pelophilous/lapidicolous species such as Thorellius and Mesomexovis, powerful diggers, commonly found underneath rocks or other debris; and scorpions that can be found inside the leaf litter or under nooks of rocks or trees, such as Vaejovis sp. However, it is enigmatic why the Diplocentridae family, well-represented in Mexico, has not been found in Colima. Only in Colima, out of the 3 relatively small states in Mexico, the other 2 being Morelos (Santibáñez-López et al., 2011) and Aguascalientes (Chávez-Samayoa et al., 2022), where similar scorpiofaunistic surveys have been conducted, the genus Diplocentrus is absent. From a biogeographical point of view, if this family is absent in Colima, it is not trivial and requires further investigation.
As a final remark, we hope we have accomplished our aim of urging arachnologists to examine the benefits of local inventories and the potential use of barcodes at the forefront of discovering and documenting local arachnid diversity.
Medical and pharmacological relevance of some scorpions of Colima
From a medical point of view, it is of utmost importance to identify the species of scorpions in an area and to be able to distinguish hazardous from harmless species. Colima is one of the states with the highest incidence rates of mortality and morbidity caused by intoxication by scorpion sting (ISS) (Chowell et al., 2006; González-Santillán & Possani, 2018). By the 1940s, Colima was at the forefront of ISS and deaths in Mexico. However, with the introduction of a safe and effective antivenom in the 1970s, mortality has diminished, although morbidity is still high (González-Santillán & Possani, 2018).
Due to the medical relevance of scorpionism in Mexico, the venom of some buthid scorpions distributed in Colima has been studied extensively (Table 6). We know that mammalian sodium scorpion toxins (NaScTx) are the chief and sometimes most abundant compound responsible for the toxicity of scorpion venom, although potassium, chlorine, and calcium-gated channels, among others, are also affected by toxic peptides (Cid-Uribe et al., 2019; González-Santillán & Possani 2018). Until now, C. hirsutipalpus, C. ornatus, C. possanii, and C. tecomanus have been studied with a proteomic or transcriptomic approach, demonstrating that these venoms are powerful enough to decimate humans (García-Guerrero et al., 2020; García-Villalvazo et al., 2023, Valdez-Velázquez et al., 2016, 2018). Centruroides tecomanus venom contains at least 13 mammalian toxic compounds (Valdez-Velázquez et al., 2016), of which Ct1a is the main mammalian sodium scorpion toxin component, with a molecular weight of 7,591 Da (Martin et al., 1988). The proteomic analysis of C. ornatus toxic components identified 3 major mammalian NaScTx, CO1, CO2, and CO3, with molecular weights of 7,561.2, 7,614.3, and 7,774.9 Da, respectively (García-Guerrero et al., 2020). On the other hand, the published mass fingerprint of C. hirsutipalpus venom has 4 compounds with similar molecular weight to C. tecomanus peptides, suggesting that these species may have similar or identical NaScTx (Valdez-Velázquez et al., 2018). One of the shared compounds found in C. hirsutipalpus corresponds to the molecular weight of Ct1a (Valdez-Velázquez et al., 2013), which means that Ct1a could be responsible for, or at least contribute to, the high toxicity of this species. Although the molecular components with a weight corresponding to the toxic peptides CO1, CO2, and CO3 are not present in C. hirsutipalpus venom, the mass fingerprint of C. tecomanus reported a peptide with similar molecular weight to CO2 toxin present in C. ornatus (Valdez-Velázquez et al., 2013). Our phylogenetic results show that these species belong to the infamatus group, and it is notable that the molecular weight among some toxins is similar. This observation opens the possibility that the toxin diversity of the mammalian NaScTx may have been inherited from a common ancestor. We realize, however, the need for a more in-depth study to unveil the relationship among these toxic peptides to test such a hypothesis. In contrast, a C. possanii crude venom proteomic study resulted in the identification of 18 NaScTx, of which, CpoNatBet09 was identical to Cll2b and Cii1 from C. limpidus and C. infamatus, respectively (Valdez-Velázquez et al., 2013). This example allows us to present the counterpart of the previous one. Centruroides limpidus in our topology grouped with members of the elegans group and C. possanii within the infamatus group (Fig. 2). We can conclude that the exact sequence match between CpoNatBet09 and Cll2b is most likely due to a selective force that produced convergent evolution. Alternatively, this is a deeper inheritance within the ancestors of the infamatus and elegans group.
The comparisons and hypotheses about venom similarities and species assume that all scorpions used to conduct the experiments to obtain the crude venom were correctly identified. However, the discovery of putative species of Centruroides might question some of these results, particularly those of C. tecomanus. The first author has been collaborating to identify most samples of species used in some of the recently published investigation on venom research (Cid-Uribe et al., 2019; García-Guerrero et al., 2020; Romero-Gutiérrez et al., 2017, Valdez-Velázquez et al., 2018, among others). And in some cases, voucher specimens have been deposited in the CNAN, therefore the taxonomic identity for C. hirsutipalpus, C. ornatus, C. possanii can be readily corroborated.
Table 6
Scorpion species with reported LD50 tested on mice at μg/20g mouse weight scale with the named, most abundant, and likely responsible for high toxicity sodium toxins found in the scorpion species of Colima.
Species
LD50
NaScTx
Autor
Centruroides hirsutipalpus Ponce-Saavedra and Francke, 2009
11.7 ± 1.9
26 β-NaTx and 5 α-NaTx (proteome) 71 β-NaTx and 16 α-NaTx (transcriptome)
Valdez-Velázquez et al. (2013, 2016), Ramírez et al. (1988)
Thorellius cristimanus Pocock, 1902
N/A
10 β-NaTx and 3 α-NaTx (transcriptome)
Romero-Gutiérrez et al. (2017)
Because vaejovid scorpions exhibit lower toxin potency, proteomic studies are uncommon. However, the study of the crude venom of Thorellius cristimanus under a transcriptomic and proteomic approachidentified160 potential venom peptides (Romero-Gutiérrez et al., 2017). The authors found a great diversity of channel toxins targeting potassium and calcium ion-channels, numerous enzymes, and NDBP. Nevertheless, none of these toxins appear to affect mammals to endanger their life. Noteworthy are the NDBP compounds with antimicrobial activity (Almaaytah et al., 2014), which potentially may be a source for developing novel commercial antibiotics. More recently, Ibarra-Vega et al. (2023) reported finding the neurotransmitter serotonin and 2 derived or intermediate indoles of serotonin: N-methylserotonin, and bufotenidine. Although serotonin has been reported in other scorpions and the sea snail genus Conus, the other indoles represent the first report in scorpions. The authors proposed that the 3 components are involved in defense and probably prey submission because they produce extreme pain in the victim. Furthermore, N-methylserotonin and bufotenidine showed a similar affinity for serotonin cellular receptors, implying a role in the effect of this neurotransmitter, and thus, it showed promising medical applications.
Integrating taxonomic, distributional, and in our case venom diversity within local scorpion faunas is an interdisciplinary topic uncommon in the literature (Brito & Borges, 2015; Cao et al., 2014), although this exercise has been done in Mexico before (Santibáñez-López et al., 2015). This multidisciplinary work took advantage of a reciprocal illumination exercise. This short synthesis of the diversity of venoms of scorpion of Colima revealed the potential of toxins to be part of the evidence to identify and delimit species (Schaffrath et al., 2018). Moreover, the synthesis provided a glance of the evolution of venoms not only for the medically prominent species of Centruroides but also for vaejovid scorpion species. From the experimental discipline, we learned that it is paramount to maintain vouchers after biochemical or physiological experiments to track down the original material and keep up with the dynamics of taxonomy, a tool requiring empirical information to permit the identification of species to validate past, present, and future discoveries in other biological disciplines.
Acknowledgments
We thank the University of Colima students and Josué López Granados (Facultad de Ciencias, UNAM) for their assistance in collecting scorpions throughout the state of Colima. JICU thanks Conahcyt for granting a postdoctoral scholarship (512560). Collection permits granted by Semarnat included SGPA/DGVS/12063/15, SGPA/DGVS/02139/2022, and FAUT-0305. Finally, we are grateful to the Associate Editor and two reviewers for improving an early version of the manuscript, particularly to Andrés Ojanguren-Afilastro for his critical reading.
Baldazo-Monsivaiz, J. G., Ponce-Saavedra, J., & Flores-Moreno, M. (2012). Los alacranes (Arachnida: Scorpionida) de importancia médica del estado de Guerrero, México. Dugesiana, 19, 143−150.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., & Canché-Aguilar, I. (2017). Los escorpiones (Arachnida: Scorpiones) del municipio de Chilpancingo de los Bravo, estado de Guerrero, México Entomología Mexicana, 4, 21−27.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., Sánchez-Arriaga, J., López-Flores, M., Reyes-Castelán, A. et al. (2016). Los escorpiones (Arachnida: Scorpiones) del municipio de Taxco de Alarcón, del estado de Guerrero, México. Entomología Mexicana, 3, 75−80.
Bolaños, L. M., Rosenblueth, M., de Lara, A. M., Migueles-Lozano, A., Gil-Aguillón, C., Mateo-Estrada, V. et al. (2019). Cophylogenetic analysis suggests cospeciation between the Scorpion Mycoplasma Clade symbionts and their hosts. Plos One, 14, e0209588 https://doi.org/10.1371/journal.pone.0209588
Brito, G., & Borges, A. (2015). A checklist of the scorpions of Ecuador (Arachnida: Scorpiones), with notes on the distribution and medical significance of some species. Journal of Venomous Animals and Toxins Including Tropical Diseases, 21, 1–23. https://doi.org/10.1186%2Fs40409-015-0023-x
Cao, Z., Di, Z., Wu, Y., & Li, W. (2014). Overview of scorpion species from China and their toxins. Toxins, 6, 796–815. https://doi.org/10.3390/toxins6030796
Cid-Uribe, J. I., Meneses E. P., Batista, C. V. F., Ortiz, E., & Possani, L. D. (2019). Dissecting toxicity: The venom gland transcriptome and the venom proteome of the highly venomous scorpion Centruroides limpidus (Karsch, 1879). Toxins, 247, 1−21. https://doi.org/10.3390/toxins11050247
Contreras-Félix, G. A., & Francke, O. F. (2019). Taxonomic revision of the “mexicanus” group of the genus Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae). Zootaxa, 4596, 1−100. https://doi.org/10.11646/zootaxa.4596.1.1
Contreras-Félix, G. A., del Pozo, O. G., & Navarrete-Heredia, J. L. (2023). A new species of Vaejovis from the mountains of west Mexico (Scorpiones: Vaejovidae). Dugesiana, 30, 229−245. https://doi.org/10.32870/dugesiana.v30i2.7306
Chávez-Samayoa, F., Díaz-Plascencia, J. E., & González-Santillán, E. (2022). Two new species of Vaejovis (Scorpiones: Vaejovidae) belonging to the mexicanus group from Aguascalientes, Mexico, with comments on the homology and function of the hemispermatophore. Zoologischer Anzeiger, 298, 148–169. https://doi.org/10. 1016/j.jcz.2022.04.005
Chowell, G., Díaz-Dueñas, P., Bustos-Saldaña, R., Alemán-Mireles, A., & Fet, V. (2006). Epidemiological and clinical characteristics of scorpionism in Colima, Mexico (2000-2001). Toxicon, 47, 753–758. https://doi.org/10.1016/j.toxicon.2006.02.004
Esposito, L. A., & Prendini, L. (2019). Island ancestors and New World biogeography: a case study from the scorpions (Buthidae: Centruroidinae). Scientific Reports, 9, 3500. https://doi.org/10.1038/s41598-018-33754-8
Fet, V., Graham, M. R., Blagoev, G., Karataş, A., & Karataş, A. (2016). DNA barcoding indicates hidden diversity of Euscorpius (Scorpiones: Euscorpiidae) in Turkey. Euscorpius, 216, 1–13.
Fet, V., Graham, M. R., Webber, M. M., & Blagoev, G. (2014). Two new species of Euscorpius (Scorpiones: Euscorpiidae) from Bulgaria, Serbia, and Greece. Zootaxa, 3894, 83–105. https://doi.org/10.11646/zootaxa.3894.1.7
Fet, V., & Lowe G. (2000). Family Buthidae C. L: Koch, 1837. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 54–286). New York: New York Entomological Society.
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
García-Guerrero, I. A., Cárcamo-Noriega, E., Gómez-Lagunas, F., González-Santillán, E., Zamudio, F. Z., Gurrola, G. B. et al. (2020). Biochemical characterization of the venom from the Mexican scorpion Centruroides ornatus, a dangerous species to humans. Toxicon, 15, 27–38. https://doi.org/10.1016/j.toxicon.2019.11.004
García-Villalvazo, P. E., Jiménez-Vargas, J. M., Lino-López, G. J., Meneses, E. P., Bermúdez-Guzmán, M. J., Barajas-Saucedo, C. E. et al. (2023). Unveiling the protein components of the secretory-venom gland and venom of the scorpion Centruroides possanii (Buthidae) through omic technologies. Toxins, 15, 1–20. https://doi.org/10.3390/toxins15080498
Goodman, A. M., Prendini, L., & Esposito, L. A. (2021). Systematics of the arboreal Neotropical ‘thorellii’ clade of Centruroides bark scorpions (Buthidae) and the efficacy of mini-barcodes for museum specimens. Diversity, 13, 441. https://doi.org/10.3390/d13090441
Goodman, A. M., Prendini, L., Francke, O. F., & Esposito, L. A. (2021). Systematic revision of the arboreal neotropical “thorellii” clade of Centruroides Marx, 1890, bark scorpions (Buthidae C. L. Koch, 1837) with descriptions of six new species. Bulletin of the American Museum of Natural History, 452, 1–92.
González-Santillán, E. (2004). Diversidad, taxonomía y hábitat de alacranes. In A. N. García, & R. Ayala (Eds.), Artrópodos de Chamela (pp. 25–35). México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México.
González-Santillán, E., Galán-Sánchez, M. A., & Valdez-Velázquez, L. L. (2019). A new species of Centruroides (Scorpiones, Buthidae) from Colima, Mexico. Comptes Rendus Biologies, 342, 9–10. https://doi.org/10.1016/j.crvi.2019.10.002
González-Santillán, E., & Possani, L. D. (2018). North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Tropica, 187, 264–274. https://doi.org/10.1016/j.actatropica.2018.08.002
González-Santillán, E., & Prendini, L. (2013). Redefinition and generic revision of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, with descriptions of six new genera. Bulletin of the American Museum of Natural History, 382, 1–71.
González-Santillán, E., & Prendini, L. (2015). Phylogeny of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, based on morphology, mitochondrial and nuclear DNA. Cladistics, 31, 341–405. https://doi.org/10.1111/cla.12091
González-Santillán, E., & Prendini, L. (2018). Systematic revision of the North American Syntropine Vaejovid scorpion genera Balsateres, Kuarapu, and Thorellius, with descriptions of three new species. Bulletin of the American Museum of Natural History, 420, 1–81. https://doi.org/10.1206/0003-0090-420.1.1
Hoang, D. T., Chernomor, O., Haeseler, A. von, Minh, B. Q., & Vinh, L. S. (2018) UFBoot2: Improving the ultrafast bootstrap approximation. Molecular Biology and Evolution, 35, 518–522. https://doi.org/10.1093/molbev/msx281
Hoffmann, C. C. (1932). Monografías para la entomología médica de México. Monografía Núm. 2. Los scorpiones de Mexico. Segunda parte, Buthidae. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, 3, 243–361.
Huerta-Jiménez, H. (2018). Actualización de la Colección de Artrópodos con importancia médica (CAIM), Laboratorio de Entomología, InDRE. Version 1.5. Comisión Nacional para el Conocimiento y uso de la Biodiversidad.
Ibarra-Vega, R., Jiménez-Vargas, J. M., Pineda-Contreras, A., Martínez-Martínez, F. J., Barajas-Saucedo, C. E., García-Ortega, H. et al. (2023). Indolealkylamines in the venom of the scorpion Thorellius intrepidus. Toxicon, 233, 107232. https://doi.org/10.1016/j.toxicon.2023.107232
Jarvis, A., Reuter, H. I., Nelson, A. & Guevara, E. (2008). Hole-filled seamless SRTM data V4, International Centre for Tropical Agriculture (CIAT), available from http://srtm.csi.cgiar.org
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., Haeseler, A. von, & Jermiin, L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nature Methods, 14, 587–589. https://doi.org/10.1038/nmeth.4285
Katoh, K., Rozewicki, J., & Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization, Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Kocher, T. D., Thomas, W. K., Meyer, A., Edwards, S. V., Paabo, S., Villablanca, F. X. et al. (1989). Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proceedings of the National Academy of Sciences, 86, 6196–6200. https://doi.org/10.1073%2Fpnas.86.16.6196
Kuraku, S., Zmasek, C. M., Nishimura, O., & Katoh, K. (2013). aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity, Nucleic Acids Research, 41, W22–W28. https://doi.org/10.1093/nar/gkt389
López-Granados, J. (2019). Análisis filogenético del complejo de especies occidentalis del género Mesomexovis González-Santillán y Prendini, 2013 (Scorpiones: Vaejovidae) basado en evidencia morfológica (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. CDMX.
Lourenço, W. R., & Sissom, W. D. (2000). Scorpiones. In B. J. Llorente, E. González, & N. Papavero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia una síntesis de su conocimiento (pp. 115–135). Ciudad de México: Universidad Nacional Autónoma de México.
Maddison, W. P., & Maddison, D. R. (2023). Mesquite: a modular system for evolutionary analysis. Version 3.81. http://www.mesquiteproject.org
Martin, B. M., Carbone, E., Yatani, A., Brown, A. M., Ramírez, A. N., Gurrola, G. B. et al. (1998). Amino acid sequence and physiological characterization of toxins from the venom of the scorpion Centruroides limpidus tecomanus Hoffmann. Toxicon, 26, 785–94. https://doi.org/10.1016/0041-0101(88)90319-4
Mendoza, J., & Francke, O. (2017). Systematic revision of Brachypelma red-kneed tarantulas (Araneae: Theraphosidae), and the use of DNA barcodes to assist in the identification and conservation of CITES-listed species. Invertebrate Systematics, 31, 157–179. https://doi.org/10.1071/IS16023
Minh, B. Q., Hahn, M. W., & Lanfear, R. (2020). New methods to calculate concordance factors for phylogenomic datasets. Molecular Biology and Evolution, 37, 2727–2733. https://doi.org/10.1093/molbev/msaa106
Morrone, J. (2019). Regionalización biogeográfica y evolución biótica de México: encrucijada de la biodiversidad del Nuevo Mundo. Revista Mexicana de Biodiversidad, 90, e902980. https://doi.org/10.22201/ib.20078706e.2019.90.2980
Morrone, J., Escalante, T., & Rodríguez-Tapia, G. (2017). Mexican biogeographic provinces: map and shapefiles. Zootaxa, 4277, 277–279. https://doi.org/10.11646/zootaxa.4277.2.8
Nguyen, L. T., Schmidt, H. A., Haeseler, A. von, & Minh, B. Q. (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Molecular Biology and Evolution, 32, 268–274. https://doi.org/10.1093/molbev/msu300
Nunn, G. B., Theisen, B. F., Chirstensen, B., & Arctander, P. (1996). Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda. Journal of Molecular Evolution, 42, 211–223. https://doi.org/10.1007/bf02198847
Podnar, M., Grbac, I., Tvrtković, N., Hörweg, C., & Haring, E. (2021). Hidden diversity, ancient divergences, and tentative Pleistocene microrefugia of European scorpions (Euscorpiidae: Euscorpiinae) in the eastern Adriatic region. Journal of Zoological Systematics and Evolutionary Research, 59, 1824–1849. https://doi.org/10.1111/jzs.12562
Ponce-Saavedra, J., & Francke, O. F. (2013). Clave para la identificación de especies de alacranes del género Centruroides Marx 1890 (Scorpiones: Buthidae) en el Centro Occidente de México. Biológicas, 15, 52–62.
Ponce-Saavedra, J. y Francke, O. F. (2019). Una especie nueva de alacrán del género Centruroides (Scorpiones: Buthidae) del noroeste de México. Revista Mexicana de Biodiversidad, 90, e902660. https://doi.org/10.22201/ib.20078706e.2019. 90.2660
Ponce-Saavedra, J., Francke, O. F., Cano-Camacho, H., & Hernández-Calderón, E. (2009). Evidencias morfológicas y moleculares que validan como especie a Centruroidestecomanus (Scorpiones, Buthidae). Revista Mexicana de Biodiversidad, 80, 71–84. https://doi.org/10.22201/ib.20078706e.2009.001.585
Ponce-Saavedra, J., Francke, O. F., Quijano-Ravell, A. F., & Santillán, R. C. (2016). Alacranes (Arachnida: Scorpiones) de importancia para la salud pública de México. Folia Entomológica Mexicana, 2, 45–70.
Ponce-Saavedra, J., Jiménez, M. L., Quijano-Ravell, A. F., Vargas-Sandoval, M., Chamé-Vázquez, D., Palacios-Cardiel, C. et al. (2023). The Fauna of Arachnids in the Anthropocene of Mexico, Chapter 2(pp. 18–39) In R. W. Jones, C. P. Ornelas-García, R. Pineda-López, & F. Álvarez (Eds.), Mexican fauna in the Anthropocene. Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-031-17277-9
Ponce-Saavedra, J., Linares-Guillén, J. W., & Quijano-Ravell, A. F. (2022). Una nueva especie de alacrán del género Centruroides Marx (Scorpiones: Buthidae) de la costa Noroeste de México. Acta Zoológica Mexicana (nueva serie), 38, 1–24. https://doi.org/10.21829/azm.2022.3812517
Ponce-Saavedra, J., Quijano-Ravell, A. F., Teruel, R., & Francke, O. F. (2015). Redescription of Centruroides ornatus Pocock, 1902 (Scorpiones: Buthidae), a montane scorpion from Central Mexico. Revista Ibérica de Aracnología, 27, 81–89.
Prendini, L., Weygoldt, P., & Wheeler, W. C. (2005). Systematics of the Damon variegatus group of African whip spiders (Chelicerata: Amblypygi): evidence from behaviour, morphology and DNA. Organisms Diversity & Evolution, 5, 203–236. https://doi.org/10.1016/j.ode.2004.12.004
Quijano-Ravell, A. F., De Armas, L. F., Francke, O. F., & Ponce-Saavedra, J. (2019). A new species of the genus Centruroides Marx (Scorpiones, Buthidae) from western Michoacán state, México using molecular and morphological evidence. Zookeys, 859, 31–48. https://doi.org/10.3897/zookeys.859.33069
Quijano-Ravell, A. F., Ponce-Saavedra, J., Francke, O. F., & Villaseñor-Ramos. M. A. (2010). Nuevos registros y distribución actualizada de Centruroides tecomanus Hoffmann, 1931 (Scorpiones: Buthidae). Ciencia Nicolaita, 52, 179–189.
QGIS.org. (2021). QGIS Geographic Information System. Open-Source Geospatial Foundation Project. http://qgis.org
Ramírez-Ruiz, J. J., & Bretón-González, M. (2016). Fisiografía y geología. In La Biodiversidad en Colima. Estudio de estado (Eds.) (pp. 25–31). México D.F.: Conabio.
Romero-Gutiérrez, T., Peguero-Sánchez, E., Cevallos, M. A., Batista, C. V. F., Ortiz, E., & Possani L. D. (2017). A deeper examination of Thorellius atrox scorpion venom components with omics technologies. Toxins, 9, 399. https://doi.org/10.3390/toxins9120399
Ruiz-Vega, M. L., Hernández-Canchola, G., & León-Paniagua, L. (2018). Molecular systematics and phylogeography of the endemic Osgood’s deermouse Osgoodomys banderanus (Rodentia: Cricetidae) in the lowlands of western Mexico. Molecular Phylogenetics and Evolution, 127, 867–877. https://doi.org/10.1016/j.ympev.2018.06.034
Santibáñez-López, C. E., Francke, O. F., & Córdova-Athanasiadis, M. (2011). The genus Diplocentrus Peters (Scorpiones: Diplocentridae) in Morelos, Mexico. Revista Ibérica de Aracnología, 19, 3–3.
Santibáñez-López, C. E., Francke, O. F., & Prendini, L. (2014). Phylogeny of the North American scorpion genus Diplocentrus Peters, 1861 (Scorpiones: Diplocentridae) based on morphology, nuclear and mitochondrial DNA. Arthropod Systematics & Phylogeny, 72, 257–279. https://doi.org/10.3897/asp.72.e31789
Santibáñez-López, C., Francke, O., Ureta, C., & Possani, L. (2015). Scorpions from Mexico: from species diversity to venom complexity. Toxins, 8, 1–18. https://doi.org/10.3390/toxins8010002
Schramm, F. D., Valdez-Mondragón, A., & Prendini, L. (2021). Volcanism and palaeoclimate change drive diversification of the world’s largest whip spider (Amblypygi). Molecular Ecology, 30, 2872–2890. https://doi.org/10.1111/mec.15924
Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H., & Flook, P. K. (1994). Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Annals of the Entomological Society of America, 87, 651–700. https://doi.org/10.1093/aesa/87.6.651
Sissom, W. D. (2000). Family Vaejovidae Thorell, 1876. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 503–553). New York: New York Entomological Society.
Towler, W. I. I., Ponce-Saavedra, J., Gantenbein, B., & Fet, V. (2001). Mitochondrial DNA reveals a divergent phylogeny in tropical Centruroides (Scorpiones: Buthidae) from Mexico. Biogeographica, 77, 157–172.
Valdez-Velázquez, L. L., Olamendi-Portugal, T., Restano-Cassulini, R., Zamudio, F. Z., & Possani, L. D. (2018). Mass fingerprinting and electrophysiological analysis of the venom from the scorpion Centruroides hirsutipalpus (Scorpiones: Buthidae). Journal of Venom Animal Toxins Including Tropical Diseases, 24, 17. https://doi.org/10.1186/s40409-018-0154-y
Valdez-Velázquez, L. L., Quintero-Hernández, V., Romero-Gutiérrez, M. T., Coronas, F. I., & Possani, L. D. (2013). Mass fingerprinting of the venom and transcriptome of venom gland of scorpion Centruroides tecomanus. Plos One, 8, e66486. https://doi.org/10.1371/journal.pone.0066486
Valdez-Velázquez L. L., Romero-Gutiérrez M. T., Delgado-Enciso, I., Dobrovinskaya, O., Melnikov, V., Quintero-Hernández, V. et al. (2016). Comprehensive analysis of venom from the scorpion Centruroides tecomanus reveals compounds with antimicrobial, cytotoxic, and insecticidal activities. Toxicon, 118, 95-103. https://doi.org/10.1016/j.toxicon.2016.04.046
Williams, S. C. (1980). Scorpions of Baja California, Mexico and adjacent islands. Occasional Papers of the California Academy of Sciences, 135, 1–127.
Williams, S. C. (1986). A new species of Vaejovis from Jalisco, Mexico (Scorpiones: Vaejovidae). Pan-Pacific Entomologist, 62, 355–358.
A new species of whitefly, Aleuropleurocelus tecomastans, is described. The studied specimens were found in the municipality of Acapulco, State of Guerrero, Mexico on Tecoma stans (L.) (Bignoniaceae) leaves. A dichotomous key to identify members of Aleuropleurocelus group ceanothi, defined by a transverse suture of the molt which reaches the submarginal line, is provided. Photographs of morphological pupal structures are given and diagnostic separation to other related species is discussed.
Una especie nueva de Aleuropleurocelus (Hemiptera: Aleyrodidae) y clave para el grupo ceanothi de Mexico
Resumen
Una especie nueva de mosca blanca, Aleuropleurocelus tecomastans, es descrita. Los especímenes estudiados fueron encontradosen el municipio de Acapulco, Estado de Guerrero, México sobre Tecoma stans (L.) (Bignoniaceae). Se provee una clave dicotómica para identificación de especies de Aleuropleurocelus grupo ceanothi, definido por una sutura transversa de la muda que termina en la línea submarginal. Se proporcionan fotografías de las estructuras morfológicas del pupario y se analiza la separación diagnóstica en otras especies relacionadas.
The genus Aleuropleurocelus was described by Drews and Sampson (1956), comprising 8 species and an identification key to all Californian species was included (Drews & Sampson, 1958). Later Dooley et al. (2010) described A. nevadensis Dooley, while Polaszek and Gill (2011) added A. hyptisemoryi Gill to the genus. Nowadays, 19 species of Mexican species of Aleuropleurocelus are known and have been largely studied by Carapia-Ruiz (2023), Carapia-Ruiz (2020a, b), Carapia-Ruiz and Sánchez-Flores (2019a, b), Carapia-Ruiz et al. (2018a, b, 2020, 2023), Sánchez-Flores and Carapia-Ruiz (2018a, b), Sánchez-Flores et al. (2018a, b, 2020, 2021). The genus is segregated into 3 major groups: abnormis (semioval), nigrans, and ceanothi according to Dooley et al. (2010) and Sánchez-Flores et al. (2021).
The ceanothi group, in which the transverse suture of molt ends at the submarginal line is included, A. granulata (Sampson & Drews), and the morphologically related species A. sampsoni Sánchez-Flores & Carapia-Ruiz and A. pseudogranulata Carapia-Ruiz & Sánchez-Flores. While collecting puparia of Aleyrodidae in Acapulco Guerrero, an unknown species of this group with distinctive characters was found. The objective of this contribution is to describe a new species and provide a key to all species of Aleuropleurocelus group ceanothi.
Materials and methods
The specimens were collected on the underside of the leaves of Tecoma stans (L.). in Acapulco, Guerrero. The specimens once taken were retained ethanol and thus transferred to the Laboratorio de Entomología, Escuela de Estudios Superiores de Xalostoc of the Universidad Autónoma del Estado de Morelos, to be fully processed and mounted in permanent preparation with Canada balsam on slides for further study under stereomicroscope and facilitating identification following specialized literature such as Drews and Sampson (1958), Martin (2004, 2005), and Sanchez-Flores et al. (2021). After preparations, specimens were observed under a Motic BA 320 phase contrast optical microscope considering several magnifications: 4X 100X, 400X and 1,000X. The terminology used follows Drews and Sampson (1956) and Martin (2005). The studied specimens are deposited at Colección Nacional de Insectos (CNIN), Instituto de Biología, Universidad Nacional Autónoma de México. México City, Mexico.
Diagnosis. Pupa boat-shaped, transverse molting suture reaches the submarginal line, dorsum and venter black, eye spots with tubercles, abdominal depressions present, middle area of abdominal segments smooth, abdominal VIII setae anterolateral to the vasiform orifice very small.
General form: pupae in situ. Dorsal and ventral surface of pupae black, a thin fringe of wax is distinguished around the body margin.
Specimens on slides: semioval body (Fig. 1). Deflexed submargin. Margin: with 61-68 pairs of marginal teeth with 2-3 acute terminal protrusions (Fig. 2), submargin with a regular band of small tubercles (Fig. 3); subdorsum: with a band of crescent tubercles (Fig. 4). Cephalothorax: longitudinal molting suture with a row of defined tubercles on each side giving the appearance of a zipper, transverse molting suture ends at the apparent margin (submarginal fold), well defined meso-metathoracic suture (Fig. 5), ocular markings (eye spots) indicated by pale coloration and 5-7 pairs of pale tubercles in the subdorsal area near the submarginal line, cephalic setae absent; mid thoracic zone with 2 pairs of setae, the mesothoracic and the metathoracic. Abdomen: dorsum with abdominal segments I-VIII clearly visible in the middle part (Fig. 6). With abdominal depressions in the middle area of the segments (Fig. 7), cuticle in the middle of the abdominal segments smooth. Vasiform orifice: elongated semicordiform (Fig. 8); elevated; operculum with 4 irregular longitudinal furrows and spinules at the apex, completely covering the lingula and most of the vasiform orifice, ring of the orifice defined in its anterior part, abdominal VIII setae anterolateral to the vasiform orifice very small, caudal protuberance developed. Pores, normally as follows: 8 pairs in the cephalic area, 6 on submedian area and 2 posterior to eye spots; 4 pairs in the mesothorax, 2 on submedian area and 2 in the subdorsal area of the mosothorax; medial area of abdominal segments I, III, V, VII with 2 pairs each segment, segment VIII with 2 pairs; subdorsal area of abdominal segments II, IV, V, with 2 pairs of pores in each segment (Fig. 9).
Venter: legs prothoracic, mesothoracic, metathoracic with apical adhesive sac, thoracic adhesive sacs near the base of the first pair of legs, base of the legs with a wide irregular band of 2-4 spinules, thoracic and abdomen cuticle smooth, well defined abdominal setae of segment VIII, posteriorly present a pair of spiracles. Chaetotaxy: a pair of anterior marginal setae present (near marginal teeth), cephalic setae absent, mesothoracic, metathoracic and caudal setae well developed, very small abdominal VIII setae, located anterolateral to the vasiform orifice, and posterior marginal setae small.
Figures 5-8. Aleuropleurocelus tecomastans.5) Thoracic area; 6) submedia and subdorsal area; 7) depression and pore of abdominal segment IV; 8) vasiform orifice. ad = Abdominal depression, cs = caudal seta, mss = mesothoracic seta, mts = metathoracic seta, o = operculum, pp = pore porete, sd = subdorsum, vo = vasiform orifice.
Measurements. Specimens on slides: body 550-650 µm long by 300-400 µm wide. Submargin approximately 45-55 μm. Cephalothorax: 230-280 μm, longitudinal molting suture, 240-360 μm, transverse molting suture 220-250 μm, the metathorax 30-45 μm long in its middle area, cephalic elongated structures of 8-10 µm long by 3-5 µm wide. Abdomen: abdominal segments length for segment I 22-26 μm, segment II 22-26 μm, segment III 25-30 μm, segment IV 26 -30 μm, segment V 27-32 μm, segment VI 26-29 μm, segment VII 23-27 μm, and segment VIII (from suture VII-VIII to vasiform orifice) 40-50 μm, distance from vasiform orifice to apparent margin 25-35 μm; abdominal depressions segments with approximate length in segment I 3-5 μm long by 10-12 μm wide, in segment II 3-4 μm long by 12-15 μm wide, in segment III 3-4 μm long by 10-13 μm wide, in segment IV 3-4 μm long by 10-14 μm wide, in segment V 5-7 μm long by 10-15 μm wide, in segment VI 5-6 μm long by 9-11 μm wide. Vasiform orifice: 46-50 µm long by 32-38 µm of broad at the widest part; operculum 25-30 μm long by 22-26 μm wide. Venter: Prothoracic legs 75-77 μm long, mesothoracic legs 75-78 μm long, metathoracic legs 85-89 μm long, thoracic adhesive sacs of 16-20 μm in diameter, base of the legs spinules 4 μm long and 2 μm wide. Setae: anterior marginal setae approximately 7 μm long, mesothoracic setae approximately 15 μm long, metathoracic setae approximately 20 μm long, abdominal VIII setae 3-4 μm long, caudal seta approximately 20 μm long, and posterior marginal seta approximately 10 μm long.
Type locality: northeast of Acapulco, Guerrero, Mexico
Type material: holotype, puparium: Acapulco, Guerrero, Mexico, in leaves of Tecoma stans (Bignoniaceae), 8-iv-2021, Col. V. E. Carapia-Ruiz, deposited in CNIN (Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City, Mexico), HOM-TIP-170. Paratypes: puparia, same data as holotype, 2 deposited in CNIN, HOM-PAR-171, HOM-PAR-172.
Etymology: the suffix name is based on a combination of the host plant’s scientific name where specimens were associated.
Distribution: Acapulco, Guerrero, Mexico.
Plant associations: Tecoma stans (Bignoniaceae).
Remarks.
Aleuropleurocelus tecomastans is placed within the ceanothi group by the transverse molting suture reaches the apparent margin (submarginal line). The new species is similar in apparence to A. granulate, A. sampsoni,and A. pseudogranulata but can be differentiated by the presence of abdominal depressions which is opposite as shown in A. granulata, A. sampsoni, A. pseudogranulata or A. ceanothi. Also A. asciculatus presents a subdorsal fold which is absent in the other species.
Key to species of Aleuropleurocelus group ceanothi
1. With bands of dense wax on dorsum, pores of double wall on dorsum 2
—Without such bands or pores 3
2. (1) Two bands of dense wax on dorsum (1 thin band on subdorsum and 1 wide band on submedian area ornatus Drews & Sampson
— One band of dense wax on subdorsum sampsoni Sánchez Flores & Carapia-Ruiz
3. (1) Tubercles on submedian surface absent 4
—Tubercles on submedian surface present 8
4. (3) Without evident tubercles on subdorsal surface 5
— With evident tubercles on subdorsal surface 6
5. (4) Longitudinal suture of the molt with tubercles on metathorax laingi Drews & Sampson
— Longitudinal suture of the molt without tubercles on metathorax coachellensis Drews & Sampson
6. (4) Subdorsal fold absent 7
— Subdorsal fold present asciculatus Carapia-Ruiz
7. (6) 2 pair of thoracic adhesive sacs, submarginal band with pores erigonium Carapia-Ruiz
— Without tubercles on the anterior and posterior part of the abdominal segments, abdominal depressions present tecomastans n. sp.
8. (3) With 2 pairs of thoracic adhesive sacs, submedian area with tubercles only on the anterior part of the abdominal segments; marginal teeth without 2 or more evident prostructions 9
— With a pair of thoracic adhesive sacs, submedian area with tubercles on the anterior and posterior part of the abdominal segments; marginal teeth with evident prostructions; transversal suture of the molt curved 10
9. (8) Half posterior of puparium in elliptic form sierra (Sampson)
— Half posterior of puparium in triangular form ceanothi (Sampson)
10. (8) Vasiform orifice semicircular, venter of abdomen with posterior area very narrow, transversal suture of the molt almost straight on median area granulata Sampson & Drews
— Vasiform orifice elongated, clearly more long than wide, venter of abdomen with posterior area triangular in form, transversal suture of the molt curved on median area pseudogranulata Carapia-Ruiz
Acknowledgments
To Paul Brawn and David Ovread of the British Museum of Natural History (BMNH) for facilitating a loan of Aleuropleurocelus to study the specimens.
References
Carapia-Ruiz, V. E. (2020a). Descripción de una especie nueva del género Aleuropleurocelus y nuevos registros para Baja California, México. Southwestern Entomologist, 45, 269–274. https://doi.org/10.3958/059.045.0128
Carapia-Ruiz, V. E. (2020b). Description of a new species of the genus Aleuropleurocelus Drews & Sampson (Hemiptera: Aleyrodidae) and a new country record for a species of the genus from Mexico. Acta Zoológica Mexicana (nueva serie), 36, 1–7. https://doi.org/10.21829/azm.2020.3612184
Carapia-Ruiz, V. E. (2023). Dos especies nuevas de Aleuro- pleurocelus de México, Southwestern Entomologist, 48, 641-648. https://doi.org/10.3958/059.045.0425
Carapia-Ruiz, V. E., & Sánchez-Flores, O. A. (2019a). Descripción de una especie nueva del género Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) de California, Estados Unidos. Acta Zoológica Mexicana (nueva serie), 35, 1–4. https://doi.org/10.21829/azm.2019.3501230
Carapia-Ruiz, V. E., & Sánchez-Flores, O. A. (2019b). Descripción de la primera especie pálida del género Aleuropleurocelus. Southwestern Entomologist, 44, 315–319. https://doi.org/10.3958/059.044.0133
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., & García-Ochaeta, J. F. (2023). A new whitefly species, Aleuropleurocelus petenensis sp. nov. (Hemiptera: Aleyrodidae), from Guatemala. Acta Zoológica Mexicana (nueva serie), 39, 1–8. https://doi.org/10.21829/azm.2023.3912470
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2018a). Descripción de dos especies nuevas del género Aleuropleurocelus Drews y Sampson, 1956 (Hemiptera: Aleyrodidae) de México. Insecta Mundi, 0606, 1–13.
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2018b). Descripción de dos especies nuevas del género Aleuropleurocelus Drews y Sampson, 1956 (Hemiptera: Aleyrodidae) de México. Southwestern Entomologist, 43, 517–526.
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2021). Aleuropleurocelus palidonigrans sp. nov. de Guerrero, México, Southwestern Entomologist, 46, 781–786. https://doi.org/10.3958/059.046. 0320
Dooley, J. W., Lambrecht, S., & Honda, J. (2010). Eight new state records of aleyrodine whiteflies found in Clark County, Nevada and three newly described taxa (Hemiptera: Aleyrodidae, Aleyrodinae). Insecta Mundi, 140, 1−36.
Drews, E. A., & Sampson, W. W. (1956). Tetralicia and a new related genus Aleuropleurocelus (Homoptera: Aleyrodidae). Annals Entomological Society of America, 49, 280–283. https://doi.org/10.1093/aesa/49.3.280
Drews, E. A., & Sampson, W. W. (1958). California aleyrodids of the genus Aleuropleurocelus. Annals of the Entomological Society of America, 51, 120–125.
Martin, J. H. (2004). The whiteflies of Belize (Hemiptera: Aleyrodidae) Part 1 — Introduction and account of the subfamily Aleurodicinae Quaintance and Baker. Zootaxa, 681, 1–119. https://dx.doi.org/10.11646/zootaxa.681.1.1
Martin, J. H. (2005). Whiteflies of Belize (Hemiptera: Aleyrodidae). Part 2 — A review of the subfamily Aleyrodinae Westwood. Zootaxa, 1098, 1–116. https://dx. doi.org/10.11646/zootaxa.1098.1.1
Polaszek, A., & Gill, R. (2011). A new species of whitefly (Hemiptera: Aleyrodidae) and its parasitoid (Hymenoptera: Aphelinidae) from desert lavender in California. Zootaxa, 2750, 51–59. https://doi.org/10.11646/zootaxa.2150.1.5/biota xa.org
Sánchez-Flores, O. A., & Carapia-Ruiz, V. E. (2018a). Nueva especie de Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) y clave para especies del grupo de forma oval. Insecta Mundi, 651, 1–12.
Sánchez-Flores, O. A., & Carapia-Ruiz, V. E. (2018b). Description of two new species of the genus Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) from Mexico. Acta Zoológica Mexicana (nueva serie), 34, 1–9. https://doi.org/10.21829/azm.2018.3412145
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., Coronado-Blanco, J. M., & Ruíz-Cancino, E. (2020). Description of Aleuropleurocelus sampsoni sp. nov. (Hemiptera: Aleyrodidae) from Tamaulipas, Mexico. Florida Entomologist, 103, 274–280. https://doi.org/10.1653/024.103.0219
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., García-Martínez, O., & Castillo-Gutiérrez, A. (2018a). Descripción de una especie nueva del género Aleuropleurocelus de México. Southwestern Entomologist, 43, 257–262. https://doi.org/ 10.3958/059.043.0116
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., García-Martínez, O., & Castillo-Gutiérrez, A. (2018b). Descripción de una especie nueva del género Aleuropleurocelus (Hemiptera: Aleyrodidae) de México. Acta Zoológica Mexicana (nueva serie), 34, 1–6. https://doi.org/10.21829/azm.2018.3412104
Sánchez-Flores, O. A., García-Ochaeta, J. F., Carapia-Ruiz, V. E., Ruiz-Cancino, E., & Coronado-Blanco, J. M. (2021). New species of Aleuropleurocelus Drews and Sampson (Hemiptera: Aleyrodidae) from Guatemala, with a key to species from the oval group. Proceedings of the Entomological Society of Washington, 123, 615–621. https://doi.org/10.4289/0013-8797.123.3.614
Eduardo Suárez-Morales a, *, Karl E. Velázquez-Ornelas b
a El Colegio de la Frontera Sur, Avenida Centenario Km 5.5, 77014 Chetumal, Quintana Roo, Mexico
b Universidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Mazatlán, Joel Montes Camarena s/n, 82040 Mazatlán, Sinaloa, Mexico
Based on deep-water (700-750 m) biological samples obtained from the southern Gulf of California, Pacific coast of Mexico, a new species of the monstrilloid copepod genus Monstrilla Dana, 1849 is described based on a single subadult female collected close to the bottom with an epibenthic sledge. Monstrilla hendrickxi sp. n. is distinguished by a unique combination of characters including: 1) no trace of eyes; 2) strong, thick antennules with segments 2-4 partly fused; 3) strongly developed apical elements on antennular segment 5 and spinous processes on segments 2-5; and 4) bilobed fifth legs with 2 apical setae on the exopodal lobe and a digitiform, unarmed endopodal lobe. The new species exhibits some affinity with surface-dwelling species of Monstrilla that have 2 setae on the exopodal lobes of the fifth legs. This is the fifth record of a species of the order Monstrilloida in the Gulf of California. The present discovery in deep oceanic waters significantly adds to our knowledge of the habitat range of this copepod order and likely anticipate further interesting findings of monstrilloids in deep waters worldwide.
Una especie nueva del género Monstrilla (Copepoda: Monstrilloida: Monstrillidae) del golfo de California, México
Resumen
Con base en muestras biológicas de aguas profundas (700-750 m) obtenidas del sur del golfo de California, costa del Pacífico de México, se describe una especie nueva del género Monstrilla Dana, 1849 con base en una hembra preadulta recolectada con un trineo epibentónico. Monstrilla hendrickxi sp. n. se distingue por una combinación única de caracteres que incluyen: 1) ausencia de estructuras oculares; 2) anténulas robustas, con los segmentos 2-4 parcialmente fusionados; 3) anténula con elementos apicales muy desarrollados en el segmento 5 y procesos espiniformes en los segmentos 2-5; y 4) quinta pata bilobulada, lóbulo exopodal armado con 2 setas apicales, lóbulo endopodal digitiforme, desarmado. La nueva especie tiene afinidad con congéneres de superficie que tienen lóbulos exopodales armados con 2 setas. Este es el quinto registro de monstriloides en el golfo de California. El presente hallazgo aumenta significativamente nuestro conocimiento sobre el intervalo de hábitats de este orden de copépodos y probablemente anticipa más descubrimientos interesantes de monstriloides en aguas profundas de todo el mundo.
Members of the copepod order Monstrilloida Sars, 1901 are, as larvae, endoparasites of benthic marine invertebrates, but their infective early nauplii and their non-feeding reproductive adult and preadult stages are planktonic. The known hosts of the postnaupliar and juvenile stages include benthic polychaetes (species of the families Syllidae, Capitellidae, Serpulidae, and Spionidae), mollusks, and sponges (Huys et al., 2007; Jeon et al., 2018; Suárez-Morales, 2011, 2018; Suárez-Morales et al., 2010, 2014). In plankton, monstrilloids have been reported chiefly from a wide range of shallow coastal habitats including estuaries (Suárez-Morales et al., 2020), coastal embayments (Suárez-Morales, 1994a, b; Suárez-Morales & Gasca, 1990), and coral reefs (Sale et al., 1996), where they can be found in aggregations (Suárez-Morales, 2001). Monstrilloids have been recently reported from rocky shore tidepools as well (Cruz-Lopes de Rosa et al., 2021). Deep oceanic waters, though, would seem to be an unlikely source of monstrilloid specimens for taxonomic study because of the copepods’ limited dispersal capacity in the water column and their need to remain close to their benthic hosts (Suárez-Morales, 2001, 2011, 2018).
The order is currently represented by a single family Monstrillidae Dana, 1849 containing at least 7 valid genera: Monstrilla Dana, 1849; Cymbasoma Thompson, 1888; Monstrillopsis Sars, 1921; Maemonstrilla Grygier & Ohtsuka, 2008; Australomonstrillopsis Suárez-Morales & McKinnon, 2014; Caromiobenella Jeon, Lee & Soh, 2018, and Spinomonstrilla Suárez-Morales, 2019 (Jeon et al., 2016, 2018; Suárez-Morales et al., 2020). Currently, the genus Cymbasoma, with 78 nominal species, is the most diverse within the order (Razouls et al., 2023: Suárez-Morales & McKinnon, 2016; Walter & Boxshall, 2022). The taxonomic examination of a monstrilloid subadult female collected with a deep-water epibenthic sledge operated close to the bottom within a depth range of 700-750 m in oceanic waters of the southern Gulf of California, Mexico, allowed us to recognize this individual as representative of an undescribed species of Monstrilla, which is herein described following upgraded morphological standards and compared with congeneric species (Grygier & Ohtsuka, 1995). This is the third published record of monstrilloid copepods from deep oceanic waters worldwide (see Suárez-Morales & Mercado-Salas, 2023) and the fifth record of monstrilloids in the Gulf of California.
Materials and methods
A subadult female of a monstrilloid copepod was collected with an epibenthic sledge operated in mid-water, close to the bottom off the Pacific coast of northwestern Mexico, in the Gulf of California. The maximum sampling depth was 750 m. Immediately after collection, the organisms from the haul were preserved in 4% formalin with seawater. Sorting of specimens and preliminary observations were made under an Olympus SZ51 stereomicroscope. The single female monstrilloid individual thus found was recognized as a subadult female and tentatively identified as a member of the genus Monstrilla. Further examination was performed under an Olympus BX51 compound microscope with Nomarski DIC optics. Prior to examination, the specimen was partially dissected and swimming legs 1-4 and the cephalothorax, urosome +antennules were mounted in glycerol on glass slides, which were sealed with acrylic nail varnish. The slides have been deposited in the collection of Zooplankton (ECO-CHZ) held at El Colegio de la Frontera Sur (ECOSUR), Unidad Chetumal, in Chetumal, Quintana Roo, Mexico (ECO-CH-Z). Detailed examination allowed us to determine that this specimen represented an undescribed species, which we describe herein following current descriptive standards in monstrilloid taxonomy. The general morphological terminology follows Huys and Boxshall (1991), while the nomenclature of the antennular armature follows Grygier and Ohtsuka (1995).
Diagnosis. Large (2.9 mm) female subadult Monstrilla with robust, cylindrical cephalothorax representing about 60% of total body length and no trace of eyes. Antennules thick, about 1/3 as long as cephalothorax; antennulary segments 2-4 partly fused, segment 3 with modified setae, segments 2, 4, and 5 each furnished with a conical or spinous process. Segments 4-5 partly fused, latter segment with strongly developed apical spiniform elements. Ventral side of genital double-somite carrying short, corrugate ovigerous spines not reaching beyond distal end of caudal rami. Fifth leg bilobed, exopodal lobe bearing 2 subequally long terminal setae and endopodal lobe digitiform, unarmed. Caudal rami each with 6 setae, innermost of which (VI) shortest and outer proximal one (I) longest.
Description of holotype subadult female. Body robust; shape and tagmosis as usual in female Monstrilla (Suárez-Morales 1994a, 2019). Total body length 2.92 mm, measured from anterior end of cephalothorax to posterior margin of anal somite. Cephalothorax length 1.74 mm, representing about 60% of total body length, containing thick egg mass (Fig. 1A). Oral cone located 0.53 of way back along ventral surface of cephalothorax. Cephalic region with weakly produced forehead (Fig. 2A). All 3 cups of naupliar eye with pigment absent (Figs. 1A, B, 2A, 3D). Preoral area with ventral ornamentation of small, nipple-like cuticular processes (nlp) with adjacent fields of integumental wrinkles and anterior cluster of pores (apc) (Figs. 3D, 4B). Antennules slender, relatively short (588 µm) and thick, corresponding to 33% of cephalothorax length and almost 20 % of total body length (Fig. 1A). Antennules indistinctly 5-segmented (1-5 in Fig. 1B), segment 1 separate but segments 2-5 partly fused (Figs. 1B, 3A), with intersegmental division 2-3 marked by weak constriction. In terms of current nomenclature for antennular armature of female monstrilloid copepods (Grygier & Ohtsuka, 1995), element 1 present on first segment (Fig. 3A). Second segment armed as usual with elements 2v1-3, 2d1,2, and long element IId (Figs. 3A, B, 4C), but additionally with conical process on distal inner margin (arrowed in Fig. 3B). Putative third segment with setiform element 3, but pair of short, spiniform elements replacing usual long, setiform elements IIIv and IIId (Fig. 3A). Putative fourth segment armed with elements 4v1-3, long 4aes, and elements IVd and IVv (Fig. 3A), as well as conical spiniform process on inner proximal margin (arrowed in Fig. 3A). Fifth segment carrying setal elements Vv, Vd, Vm, 61, 2, aesthetasc 6aes, plus only 2 unbranched setae of “b” group (b6 and b3) on outer margin, as well as distal spiniform process at insertion of elements 61, 2, and 6aes (Figs. 2B, 3C). First pedigerous thoracic somite incorporated into cephalothorax, succeeding 3 free pedigerous somites each bearing pair of biramous swimming legs, all 3 together accounting for 31.3% of total body length (Fig. 1A). Endopodites and exopodites of swimming legs 1-4 unequal (exopods longer), triarticulate, and with same setal armature in each leg, except exopod of leg 1 with one fewer seta on the distal segment (Fig. 4D).
Armature formula of swimming legs: leg 1: basis 1-0, endopodite 0-1; 0-1; 1-2-2, exopodite I-1; 0-1; I-2-2; legs 2-4: basis 1-0, endopodite 0-1; 0-1; 1-2-2, exopodite I-1; 0-1; I-2-3. Coxae of legs 1-4 unarmed; each pair medially joined by subrectangular intercoxal sclerite about 1.3 times as long as broad with curved distal margin; anterior surface of sclerites 2-4 ornamented with rows of minute hyaline spinules. Basis separated from coxa posteriorly by diagonal articulation, lacking usual basipodal outer seta in legs 2-4. Outer distal corner of first and third exopodal segments of swimming legs 1-4 each with short, slender spiniform element about 1/3 as long as its segment. All natatory setae lightly and biserially plumose except for spiniform seta on outer distal corner of third exopodal segments, this being lightly setulate along inner side and bearing continuous row of small denticles along outer margin (Fig. 4D). Fifth legs medially conjoined, arising ventrally from posterior margin of fifth pedigerous somite (Fig. 4A), each being represented by elongate bilobed structure with its outer (exopodal) lobe armed with 2 subequally long apical setae. Unarmed and smooth inner (endopodal) lobe arising from proximal inner margin of outer lobe, almost reaching distal tip of latter. Urosome short (length = 389 µm long), accounting for 13.1% of total body length and consisting of fifth pedigerous somite, genital double-somite, 2 free abdominal somites, and caudal rami (Figs. 1C, 4A). Genital double somite representing 28.8% of length of urosome. Preanal somite about half as long as anal somite (Fig. 4A). Medial ventral surface of genital double-somite moderately swollen, bearing basally conjoined and posteriorly directed ovigerous spines (Fig. 4A). These spines (os in Figs. 1D, 4A) relatively short, corresponding to 21% of total body length and reaching to mid-length of caudal setae, each narrowing in its distal half to thin, seemingly socketed, seta-like section (Figs. 1D, 4A). Caudal rami subrectangular, 1.25 times as long as wide, moderately divergent, bearing 3 strong and subequally long terminal setae, as usual in genus (Figs. 1D, 4A), among 6 setal elements in all (I-VI), with innermost seta (VI) being shortest and proximal outer seta (I) being longest (Fig. 4A).
Figure 1. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female, digital photos. A, Habitus, dorsal view; B, anterior half of cephalothorax with 5- segmented antennules (1-5), ventral view; C, urosome, ventral view, showing ovigerous spines (os) arising from genital double-somite, fifth legs (P5) with 2 distal setae (1, 2), and caudal setae I-VI; D, distal part of ovigerous spines and caudal rami showing setae I-VI, ventral view.
Figure 2. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female, digital photos. A, Anterior part of cephalothorax showing weakly produced forehead and lack of eyes; B, antennule showing dorsal segmentation; C, fifth leg (P5) showing exopodal (exp) and endopodal (enp) lobes, ventral view; D, apical elements (61, 62, and 6aes) of distal antennular segment.
Figure 3. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female. A, Antennule showing setation labelled in accordance with Grygier and Ohtsuka’s (1995) nomenclature, ventral view; B, similarly labelled setation of second antennular segment, ventral view, also showing segment’s inner distal conical process (arrow); C, distal (fifth) segment of antennule, ventral view, showing similarly labelled setation and distal process (arrow); D, anterior third of cephalothorax, ventral view, showing integumental ornamentation, including nipple-like processes (nlp), anterior pore cluster (apc), and preoral pores (pp). Scales A-D = 100 µm.
Figure 4. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female. A, Urosome, ventral view, showing bilobed fifth leg with exopodal (exp) and endopodal (enp) lobes, ovigerous spines (os), and caudal rami setation (I-VI); B, anterior half of cephalothorax, ventral view, showing unlayed internal egg mass (em), oral cone (oc), and nipple-like processes (nlp); C, antennule segments 1-3 (S1-S3), ventral view, showing setation labelled in accordance with Grygier and Ohtsuka’s (1995) nomenclature, including modified setae IIIv and IIId; D, right leg 1, semi-lateral view, showing setation including basipodal seta (bs) and exopodal (exp) and endopodal (enp) rami. Scales A-D = 100 µm.
Taxonomic summary
Type locality. Southern Gulf of California, Mexico (25°53’15” N, 110°10’08” W). Sampling depth between 710 and 750 m.
Material examined. One subadult female (holotype), partially dissected and mounted on 2 semi-permanent glycerin slides sealed with acrylic nail varnish (ECO-CH-Z 11860). Epibenthic sledge, TALUD XVI B cruise, southern Gulf of California, Mexico, 31 May 2014.
Etymology. The specific name, a masculine genitive eponym, honors Dr. Michel E. Hendrickx (ICMyL-UNAM) for his sustained efforts and achievements in exploring the crustacean fauna of the Gulf of California and the Mexican Pacific.
Male. Unknown.
Host. Unknown.
Remarks
The sampling gear used to collect this specimen was an epibenthic sledge, not a plankton net; the latter is more efficient for capturing planktonic adult monstrilloids in surface waters. Epi-mesopelagic monstrilloids collected by sledge type gears have been reported from depths of 118 and 302 m in the North Atlantic (Suárez-Morales & Mercado-Salas, 2023), but those individuals were badly damaged even though the sledge had an attached plankton collector (Brandt et al., 2014). The specimen recovered from the TALUD XVI-B sledge sample was in reasonably good condition for taxonomic study.
The present subadult female monstrilloid from the Gulf of California can be readily assigned to the genus Monstrilla by its possession of the diagnostic generic features for females, including the presence of 2 somites between the genital double-somite and the anal somite, 6 caudal setae, and the oral cone’s location ventrally at nearly mid-length of the cephalothorax (Isaac, 1975; Suárez-Morales, 1994a). Among the species-level diagnostic features of M. hendrickxi sp. n., the most useful for comparison among congeneric species are the antennular structure and armature, the shape of leg 5, and the absence of eyes. These will be considered in sequence below.
The specifically distinctive characters of M. hendrickxi sp. n. include: 1) eyes and eye-related structures absent; 2) antennules relatively short, robust, representing nearly 33% of cephalothorax length, with segments 2-5 partly fused; 3) segments 2-5 furnished with modified setae or strong spiniform processes, 4) fifth antennulary segment with remarkably long, thick apical elements; 5) fifth leg bilobed, with digitiform endopodal lobe unarmed, exopodal lobe with 2 terminal setae; 6) 6 caudal setae, innermost seta (VI) being shortest, proximal outer seta (I) longest. The finding of subadult monstrillids in the plankton is not unusual and some species have been described from these individuals, including the first described monstrilloid copepod, Thaumaleus typica (Krøyer, 1842) (Grygier, 1994), Monstrilla capitellicola Hartman (1961), M. elongata Suárez-Morales, 1994a, and Monstrilla sp. from Hawaii (Suárez-Morales et al., 2014).
Partial or complete fusion of antennular segments 2-5 is found in several other species of Monstrilla, including M. ilhoii Lee & Chang, 2016, M. mariaeugeniae Suárez-Morales & Islas-Landeros, 1993, M. satchmoi Suárez-Morales & Dias, 2001, M. grandis Giesbrecht, 1891, M. gracilicauda Giesbrecht, 1893, and M. elongata Suárez-Morales, 1994a. None of these species displays the remarkable development of apical elements 61, 62, and 6aes observed in M. hendrickxi sp. n. The only available illustration of an antennule of M. nichollsi Davis, 1949 (= C. helgolandica) (Suárez-Morales pers. obs.) (cf. Nicholls, 1944: fig. 26, as Monstrilla sp.) shows a very long apical element on its fifth segment, which is probably identifiable as the aesthetasc 6aes. Elsewhere on the antennule, no congeneric species has modified setal elements IIId and IIIv on segment 3 or large, spiniform or conical processes on segments 2, 4, and 5 like those described in the new species.
Only a few known species originally described as Monstrilla possess a bilobed fifth leg with 2 setae on the outer (exopodal) lobe, 2 of them have been transferred to the genus Caromiobenella: C. helgolandica (Claus, 1863) and C. hamatapex (Grygier & Ohtsuka, 1995); the other species of Monstrilla sharing this character are M. mariaeugeniae, M. capitellicola, and M. leucopis Sars, 1921. Also, both Monstrilla sp. from Hawaii and M. capitellicola from Southern California (Hartman, 1961), likely represented by subadult individuals, also show only 2 setae on the outer lobe, a character conserved through the copepodiid stages CIII-V (Suárez-Morales et al., 2014). The new species M. hendrickxi differs from C. helgolandica, C. hamatapex, M. capitellicola, and M. leucopis, by its possession of a long, digitiform endopodal lobe, which is absent in these 4 species (Chang, 2014; Grygier & Ohtsuka, 1995; Sars, 1921; Zavarzin & Suárez-Morales, 2024). The corresponding endopodal lobe is clearly shorter in M. capitellicola (Hartman, 1961) than in the new species. Monstrilla wandelii Stephensen, 1913, and M. mariaeugeniae both exhibit a small, unarmed subtriangular endopodal lobe (Nicholls, 1944, fig 26; Park, 1967; Suárez-Morales & Islas-Landeros, 1993), which clearly differs from the elongate, digitiform endopodal lobe observed in M. hendrickxi sp. n.. Monstrilla nichollsi, a synonym of C. helgolandica (Suárez-Morales pers. obs.), was named by Davis (1949) based solely on Nicholl’s (1944, fig. 26) illustration of the fifth leg, thus allowing us to add C. helgolandica to the group of monstrillid species with 2 exopodal setae on the fifth leg exopodal lobe. It should be noted that the armature of the fifth leg exopodal lobe is conservative during the immature stages including the preadult CV (Suárez-Morales et al., 2014); changes in this character at the final molt are unlikely.
Monstrilla hendrickxi sp. n. is the only monstrilloid copepod in which no trace of the naupliar eye is present, although weakly developed visual structures have been observed previously in deep-living species (Suárez-Morales & Mercado-Salas, 2023). This contrasts with the usually well-developed, highly pigmented, three-cup naupliar eyes of most known monstrilloids. Functional eyes are probably extremely important for planktonic adult monstrilloids, allowing them, for example, to migrate to different light conditions in the water column and favor their dispersal (Suárez-Morales, 2018; Suárez-Morales & Gasca, 1990). The weak eye development of deep-living monstrilloids is likely an adaptive consequence of their aphotic habitat.
Monstrilla hendrickxi sp. n. is the fifth species of the copepod order Monstrilloida recorded from the Gulf of California, after Monstrilla gibbosa Suárez-Morales & Palomares-García, 1995, Spinomonstrilla spinosa (Park, 1967) (originally reported as Monstrilla spinosa), Cymbasoma californiense Suárez-Morales & Palomares-García, 1999, and recently M. leucopis Sars, 1921 (Suárez-Morales, 2019; Suárez-Morales & Palomares-García, 1999; Suárez-Morales & Velázquez-Ornelas, 2023).
Acknowledgements
We thank Michel E. Hendrickx (ICMyL-UNAM) for kindly allowing us to examine this specimen. Ship time for the TALUD XVI-B cruise was provided by the Coordinación de la Investigación Científica, UNAM, and partly supported by Conacyt (project # 179467). We also thank to an anonymous reviewer for the corrections made to improve this article.
References
Brandt, A., Havermans, C., Janussen, D., Jörger, K. M., Meyer-Löbbecke, A., Schnurr, S. et al. (2014). Composition and abundance of epibenthic-sledge catches in the South Polar Front of the Atlantic. Deep-Sea ResearchII, 108, 69–75. Doi?
Claus, C.(1863). Die frei lebenden Copepoden mit besonderer Berücksichtigung der Fauna Deutschlands, der Nordsee und des Mittelmeeres. Leipzig: Verlag von Wilhelm Engelmann.
Chang, C. Y. (2014). Two new records of monstrilloid copepods (Crustacea) from Korea. Animal Systematics, Evolution and Diversity, 30, 206–214. https://doi.org/10.5635/ASED. 2014.30.3.206
Cruz-Lopes da Rosa, J., Dias, C. O., Suárez-Morales, E., Weber, L. I., & Gomes-Fischer, L. (2021). Record of Caromiobenella (Copepoda, Monstrilloida) in Brazil and discovery of the male of C. brasiliensis: morphological and molecular evidence. Diversity, 13, 241. https://doi.org/ 10.3390/d13060241
Dana, J. D. (1849). Conspectus crustaceorum, quae in orbis terrarum circumnavigatione, Carolo Wilkes, e classe Reipublicae foederatae duce, lexit et descripsit Jacobus D. Dana. Pars II. Proceedings of the American Academy of Arts and Sciences, 2, 9–61.
Davis, C. C. (1949). A preliminary revision of the Monstrilloida, with descriptions of two new species. Transactions of the American Microscopical Society, 68, 245–255. https://doi.org/10.2307/3223221
Giesbrecht, W. (1891). Elenco dei Copepodi pescati dalla R. Corvetta ‘Vettor Pisani’ secondo la loro distribuzione geografica. Atti della Accademia Nazionale dei Lincei, Classe di Scienze Fisiche Matematiche e Naturali Rendiconti, 4, 276–282.
Giesbrecht, W. (1893). Systematik und Faunistik der pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte, 19, 1–831.
Grygier, M. J. (1994) [dated 1993]. Identity of Thaumatoessa (= Thaumaleus) typica Krøyer, the first described monstrilloid copepod. Sarsia, 78, 235–242. https://doi.org/10.1080/00364827.1993.10413537
Grygier, M. J., & Ohtsuka, S. (1995). SEM observation of the nauplius of Monstrilla hamatapex, new species, from Japan and an example of upgraded descriptive standards for monstrilloid copepods. Journal of Crustacean Biology, 15, 703–719. https://doi.org/10.1163/193724095X00118
Grygier, M. J., & Ohtsuka, S. (2008). A new genus of monstrilloid copepods (Crustacea) with anteriorly pointing ovigerous spines and related adaptations for subthoracic brooding. Zoological Journal of the Linnean Society, 152, 459–506. https://doi.org/10.1111/j.1096-3642.2007.00381.x
Hartman, O. (1961). A new monstrillid copepod parasitic in capitellid polychaetes in southern California. Zoologischer Anzeiger, 167, 325–334.
Huys, R., & Boxshall, G. A. (1991). Copepod evolution. London: Ray Society.
Huys, R., Llewellyn-Hughes, J., Conroy-Dalton, S., Olson, P. D., Spinks, J. N., & Johnston, D. A. (2007). Extraordinary host switching in siphonostomatoid copepods and the demise of the Monstrilloida: Integrating molecular data, ontogeny and antennulary morphology. Molecular Phylogenetics and Evolution, 43, 368–378. https://doi.org/10.1016/j.ympev. 2007.02.004
Isaac, M. J. (1975). Copepoda, sub-order: Monstrilloida. Fiches d’Identification du Zooplancton, 144/145, 1–10.
Jeon, D., Lee, W., & Soh, H. Y. (2018). New genus and two new species of monstrilloid copepods (Copepoda: Monstrillidae): integrating morphological, molecular phylogenetic, and ecological evidence. Journal of Crustacean Biology, 38, 45–65. https://doi.org/10.1093 /jcbiol/rux095
Jeon, D., Lim, D., Lee, W., & Soh, H. Y. (2016). First use of molecular evidence to match sexes in the Monstrilloida (Crustacea: Copepoda), and taxonomic implications of the newly recognized and described partly Maemonstrilla-like females of Monstrillopsis longilobata Lee, Kim & Chang, 2016. Peer J, 13,e4938. https://doi.org/10.7717/peerj.4938
Krøyer, H. (1842). Crustacés. In P. E. Gaimard (Ed.), Atlas de Zoologie. Voyages de la Commission Scientifique du Nord en Scandinavie, en Laponie au Spitzberg et aux Feröe pendant les Anneés 1838, 1839 et 1840 sur la Corvette La Recherche, Commandée par M. Fabvre (pl. 41–43). Arthus Bertrand, Paris.
Lee, J., & Chang, C. Y. (2016). A new species of Monstrilla Dana, 1849 (Copepoda: Monstrilloida: Monstrillidae) from Korea, including a key to species from the North-west Pacific. Zootaxa, 4174, 396–409. https://doi.org/10.11646/zootaxa.4174.1.24
Nicholls, A. G. (1944). Littoral Copepoda from South Australia (II) Calanoida, Cyclopoida, Notodelphyoida, Monstrilloida, and Caligoida. Records of the South Australian Museum, 8, 1–62.
Park, T. S. (1967). Two unreported species and one new species of Monstrilla (Copepoda: Monstrilloida) from the Strait of Georgia. Transactions of the American Microscopical Society, 86, 144–152.
Razouls, C., Desreumaux, N., Kouwenberg, J., & de Bovée, F. (2023). Biodiversity of marine planktonic copepods (morphology, geographic distribution, and biological data [2005-2023]). Sorbonne University, CNRS. http://copepodes.obs-banyuls.fr/en [Accessed April 28, 2022]
Sale, P. F, McWilliams, P. S., & Anderson, D. T. (1996). Composition of the near-reef zooplankton at Heron Reef, Great Barrier Reef. Marine Biology, 34, 59–66. https://doi.org/10.1007/BF00390788
Sars, G. O. (1901). An account of the Crustacea of Norway with short descriptions and figures of all the species Vol. IV. Copepoda Calanoida. Bergen: The Bergen Museum.
Sars, G. O. (1921). An account of the Crustacea of Norway with short descriptions and figures of all the species, Vol.VIII. Copepoda Monstrilloida & Notodelphyoida. Bergen: The Bergen Museum.
Stephensen, K. (1913) Account of the Crustacea and the Pycnogonida collected by Dr. V. Nordmann in the summer of 1911 from northern Stromfjord and Giesecke Lake in West Greenland. Meddeleser om Groenland, 51, 55–77.
Suárez-Morales, E. (1994a). Monstrilla elongata, a new monstrilloid copepod (Crustacea: Copepoda: Monstrilloida) from a reef lagoon of the Caribbean coast of Mexico. Proceedings of the Biological Society of Washington, 107, 262–267.
Suárez-Morales, E. (1994b). Thaumaleus quintanarooensis, a new monstrilloid copepod from the Mexican coasts of the Caribbean Sea. Bulletin of Marine Science, 54, 381–384.
Suárez-Morales, E. (2001). An aggregation of monstrilloid copepods in a western Caribbean reef area: ecological and conceptual implications. Crustaceana, 74, 689–696. https://doi.org/10.1163/156854001750377966
Suárez-Morales, E. (2019). A new genus of the Monstrilloida (Copepoda) with large rostral process and metasomal spines, and redescription of Monstrilla spinosa Park, 1967. Crustaceana, 92, 1099–1112. https://doi.org/10.1163/ 15685403-00003925
Suárez-Morales, E., & Dias, C. O. (2001). Taxonomic report of some monstrilloids (Copepoda: Monstrilloida) from Brazil with description of four new species. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Biologie, 71, 65–81.
Suárez-Morales, E., & Gasca, R. (1990). Variación dial del zooplancton asociado a las praderas de Thalassia testudinum en una laguna arrecifal del Caribe Mexicano. Universidad y Ciencia, 7, 57–64.
Suárez-Morales, E., Harris, L. H., Ferrari, F. D., & Gasca, R. (2014). Late postnaupliar development of Monstrilla sp. (Copepoda: Monstrilloida), a protelean endoparasite of benthic polychaetes. Invertebrate Reproduction & Development, 58, 60–73.
Suárez-Morales, E., & Islas-Landeros, M. E. (1993). A new species of Monstrilla (Copepoda: Monstrilloida) from a reef lagoon off the Mexican coast of the Caribbean Sea. Hydrobiologia, 271, 45–48.
Suárez-Morales, E., & McKinnon, A. D. (2014). The Australian Monstrilloida (Crustacea: Copepoda) I. Monstrillopsis Sars, Maemonstrilla Grygier & Ohtsuka, and Australomonstrillopsis gen. nov. Zootaxa, 3779, 301–340. https://doi.org/10.11646/zootaxa.3779.3.1
Suárez-Morales, E., & McKinnon, A. D., (2016). The Australian Monstrilloida (Crustacea: Copepoda) II. Cymbasoma Thompson, 1888. Zootaxa Monographs, 4102, 1–129. https://doi.org/10.11646/zootaxa.4102.1.1
Suárez-Morales, E., & Mercado-Salas, N. F. (2023). Two new species of Cymbasoma (Copepoda: Monstrilloida: Monstrillidae) from the North Atlantic. Journal of Natural History, 57, 1312–1330. https://doi.org/10.5852/ejt. 2024.917.2395
Suárez-Morales, E., Paiva Scardua, M., & Da Silva, P. M. (2010). Occurrence and histopathological effects of Monstrilla sp. (Copepoda: Monstrilloida) and other parasites in the brown mussel Perna perna from Brazil. Journal of the Marine Biological Association of the United Kingdom, 90, 953–958. https://doi.org/10.1017/S0025315409991391
Suárez-Morales, E., & Palomares-García, R. (1995). A new species of Monstrilla (Copepoda: Monstrilloida) from a coastal system of the Baja California Peninsula, Mexico. Journal of Plankton Research, 17, 745–752. https://doi.org/ 10.1093/plankt/17.4.745
Suárez-Morales, E., & Palomares-García, R. (1999). Cymbasoma californiense, a new monstrilloid (Crustacea: Copepoda: Monstrilloida) from Baja California, Mexico. Proceedings of the Biological Society of Washington, 112, 189–198.
Suárez-Morales, E., Vásquez-Yeomans, L., & Santoya, L. (2020). A new species of the Cymbasoma longispinosum species-group(Copepoda, Monstrilloida, Monstrillidae) from Belize, western Caribbean Sea. Revista Mexicana de Biodiversidad, 91, e913176. https://doi.org/10.22201/ib. 20078706e.2020.91.3176
Suárez-Morales, E., & Velázquez-Ornelas, K. E. (2023). First record of Monstrilla leucopis G.O. Sars, 1921 (Copepoda: Monstrilloida: Monstrillidae) from the Eastern Pacific. Crustaceana, 96, 1183–1190. https://doi.org/10.1163/156854 03-bja10334
Thompson, I. C. (1888). Copepoda of Madeira and the Canary Islands, with descriptions of new genera and species. Zoological Journal of the Linnean Society, 20, 145–156. https://doi.org/10.1111/j.1096-3642.1888.tb01443.x
Zavarzin, D., & Suárez-Morales, E. (2024). First record of Caromiobenella helgolandica (Claus, 1863) (Copepoda: Monstrilloida: Monstrillidae) from the Okhotsk Sea. Crustaceana, 97, 151–157. https://doi.org/10.1163/15685403- bja10345