
Francisco Lorea-Hernández*
Instituto de Ecología, A.C., Carretera antigua a Coatepec 351, Col. El Haya, 91073 Xalapa, Veracruz, Mexico
*Corresponding author: francisco.lorea@inecol.mx (F. Lorea-Hernández)
Received: 27 February 2024; accepted: 21 January 2025
Abstract
Following a detailed morphological survey of the Licaria collections in several herbaria, various taxonomic entities not recognized before were detected. Here, 11 new species of Licaria from Mesoamerica are described and illustrated. Possible relations to other species in the genus are commented.
Keywords: Lauraceae of Central America; Lauraceae of Mexico; Licaria of Central America; Licaria of Mexico
Nuevas especies de Licaria (Lauraceae) de Mesoamérica
Resumen
Como resultado de un análisis morfológico detallado de las colecciones del género Licaria en diferentes herbarios, se detectaron varias entidades taxonómicas no reconocidas previamente. Aquí se describen e ilustran 11 especies nuevas del género Licaria de la región mesoamericana. Se comentan además las posibles relaciones con otras especies del género.
Palabras clave: Lauraceae de Centroamérica; Lauraceae de México; Licaria de Centroamérica; Licaria de México
Introduction
Licaria is an endemic genus to the Americas represented mostly by medium-sized to large tree species that grow principally in the intertropical region of this part of the world. The hermaphrodite, perigynous flowers with only the third whorl of stamens fertile, whose anthers are bisporangiate, as well as the fruit often seated on a double rimmed cupule, constitute the combination of characters that distinguishes the genus among the Lauraceae. Licaria was last revised by Kurz (2000), who recognized 38 species, including 12 in the Mesoamerican area. That work is an adaptation with minor changes of Kurz doctoral dissertation (Kurz, 1983). Since then, in the lapse of 40 years, subtracting new combinations and synonyms, 31 new species have been described, adding 9 to the flora of Mesoamerica (Burger & van der Werff, 1990; Gómez-Laurito & Cascante, 1999; Gómez-Laurito & Estrada, 2002; Hammel, 1986; van der Werff, 1988, 2009). The impulse given to field work with the onset of the Flora Mesoamericana project resulted in a significant increment of herbarium specimens during the last 2 decades of the past century, but Kurz did not see most of them; those specimens have been frequently the source of new species described afterwards in Licaria. Furthermore, the expanded collections have improved our knowledge about variation of morphology within species and consequently have led to better circumscriptions of them. On the other hand, it is important to say that Kurz missed part of the diversity of Licaria because he relied mainly on what the big museums of Europe and the USA have, but did not search in several of the smaller regional herbaria; some of which preserve important specimens that support the recognition of several species that have elsewhere been synonymized or validate larger geographic distributions of taxa.
Contrary to what is found in most American genera of the Lauracae, Licaria presents a rather wide variation in flower morphology, e.g., in orientation of tepals, extent of stamens fusion, presence and extent of fusion of glands, presence of staminodes, and shape and position of anther openings. These features, together with the presence and type of hairiness on flower parts, as well as position and structure of inflorescence, constitute the basis for the recognition of species in the genus (Kostermans, 1937; Kurz, 2000; Mez, 1889). In the course of the revision of Licaria for the Flora Mesoamericana project, a number of herbarium specimens with distinctive character combinations were found, which had not been recorded previously in the genus. Differences are mainly in flower morphology, but complementary vegetative features or other data emphasize their singularity. In several cases, there are no additional specimens, but the collection on which the description is based. This situation has not been considered a drawback since, as is discussed in every case, the peculiarity of the plants is so patent that there is little chance to think they represent part of the variability of any other known species. The case strengthens the idea that we have as yet an incomplete picture of the extent of diversity and distribution in many taxa. Field work is still important and necessary.
Materials and methods
Specimens identified as Licaria (as well as unidentified, putative lauraceous material) collected in the Mesoamerican area from different herbaria (A, CAS, DS, ENCB, F, GH, HEM, K, MEXU, MICH, MO, NY, P, TEX, US, XAL) were carefully analyzed, both for vegetative and reproductive morphology. Particular attention was paid to floral characters; flowers were dissected using a Zeiss, Stemi DV4 stereomicroscope. Information of vegetative characters was collected directly from dry specimens, while floral morphology was surveyed in rehydrated material. Groups of specimens sharing similar morphology were matched with keys and descriptions of currently accepted species, in order to apply the correct names to them. To prevent misinterpretations, all available type specimens (either from herbaria cited above or accesible on Global Plants database (http://plants.jstor.org/search?plantNam) were also considered during the process of identification. Specimens that did not fit any of the known species were evaluated to determine their singularity; those whose floral and vegetative features combined do not overlap with that of accepted taxa are here proposed as new species. Following the methodology mentioned above, description of vegetative characters is based on dry specimens, while floral characters are described as they look in rehydrated material. In order to apply a standardized nomenclature to characters, applicable information in Radford et al. (1974) was used; particularly, base and apex of leaves, pubescence, tridimensional form of flowers, and shape of tepals were described according to that text.
Descriptions
Licaria breedlovei Lorea-Hern. sp. nov. (Figs. 1, 3)
Diagnosis. Trees similar to L. alata Miranda, but different because of the convex flanks of the base of the leaves, sparsely sericeous lower leaf surface, obloid flowers, densely tomentose tepals abaxially, partially pubescent staminal filaments abaxially and adaxially, densely tomentose hypanthium outside, densely hirsute-tomentose inside, and sparsely puberulent style.
Trees up to 15 m tall; twigs sericeous, soon glabrate, brownish gray or dark brown, lenticellate, conspicuously ridged, the ridges narrow, wing-like (at least when dry), each ridge originates at one extreme of petiole insertion, acropetal. Buds densely sericeous-tomentose. Leaves alternate, petioles 9-10.5(-12) mm long, bi-marginate above, glabrous, round below, sericeous, glabrescent, blades (17.5-)23-28 × (3.5-)4.5-7 cm, narrowly elliptic, pinninerved, secondary veins 11-13 pairs, upper leaf surface glabrous, lower surface sparsely sericeous, glabrescent, leaf apex acuminate, leaf base obtuse, somewhat conduplicate, the basal flanks of the blade convex and projected above the midvein. Inflorescences (7.5-)10-16.5 cm long, axillary to tiny, deciduous bracts, on the proximal section of new twigs, paniculate, sericeous-tomentose, peduncle (2.5-)4-6.5 cm long, sometimes glabrescent, pedicels 0.5-1.2(-2.5) mm long, densely tomentose, hairs yellowish-gray to yellowish-brown. Flowers obloid, tepals conspicuously inflexed, outer tepals 0.8-1 × 1.7-2 mm, very widely ovate, densely tomentose abaxially, slightly papillose toward the apex, glabrous or sometimes with scattered sericeous hairs at the base adaxially, inner tepals 0.6 × 1-1.1 mm, ovate or elliptic, tomentose abaxially, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 1-1.1 mm long, fused throughout, filaments pubescent at the base abaxially, glabrous adaxially or pubescent on the upper section, anthers 0.2-0.3 mm long, glabrous, with apical openings, glands ca. 0.2 mm, free, compressed, glabrous, sometimes reduced or absent, hypanthium ca. 0.8 mm deep, densely tomentose outside, hirsute-tomentose inside, pistil 1.3-1.4 mm long, ovary 0.8-1 mm long, glabrous, style sparsely puberulent. Fruits unknown.
Taxonomic summary
Type. Mexico. Chiapas: municipality of Petalcingo (actually municipality of Tila), steep slope of Ahk’ulbal Nab above Petalcingo, 1,700 m, 28 March 1981, D. E. Breedlove 50394 (holotype CAS; isotype MO 6485834).
Etymology. This species is dedicated to Dr. Dennis E. Breedlove, who spent so much effort in field work documenting the flora of Chiapas, pursuing the aim of elaborating a plant species compendium for that region of Mexico.
Distribution and habitat. So far the species is known only from the type collection, in an area covered by montane rain forest which, according to Breedlove (data from label of type specimen), had a canopy layer reaching 25 – 35 m, and species of the genera Alfaroa, Brunellia, Calatola, Hedyosmum, Matudea, Meliosma, Nectandra, Oreopanax, Quercus, and Turpinia, among others.
Phenology. Regarding the date when the plant was collected, it must have flowers around the end of winter and early days of spring; fruit season is not known, but possibly occurs during winter, for it has been observed that maturation of fruits in most Lauraceae takes around a year after flowering. It seems that the species is not deciduous.
Conservation status. Given the deep environmental degradation that currently prevails in the region where the species is only known, it is suspected that it is critically endangered.
Remarks
The presence of erect, concave tepals, and stamens with extrorse sporangia places L. breedlovei in subgenus Licaria. It is one element in the group (here called Licaria excelsa species group) constituted by L. alata, L. excelsa Kosterm., L. minutiflora (here described), L. pergamentacea W. C. Burger, L. sarukhanii (here described), and L. tomentulosa (here described), as it shares with them morphological features like ridged twigs, large narrow-elliptic leaves (frequently reaching 28-30 cm long), and anthers with sporangia opening apically. Within the group, L. breedlovei is distinguished from the other species by the combination of somewhat conduplicate leaf-base, lower leaf surface persistently sericeous, stamens fused throughout, and sparsely pubescent style. In addition, it differs particularly from L. alata for the obloid flowers (vs. ellipsoid), tepals densely tomentose abaxially (vs. tepals glabrous abaxially), filaments pubescent abaxially and adaxially (vs. filaments glabrous), and hypanthium densely tomentose outside and inside (vs. hypanthium glabrous on both faces).
Licaria dolichopoda Lorea-Hern. sp. nov. (Figs. 2, 3)
Diagnosis. Trees, similar to L. misantlae (Brandegee) Kosterm., glabrous throughout, leaves mostly caudate, inflorescences botryoid, flowers long-pedicellate, without staminodes, and fruit cupule thick, deeply crateriform, its outer rim slightly lobed.
Trees up to 25 m; twigs glabrous, smooth or slightly ribbed, reddish-brown or grayish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (3.5-)6.5-9(-11) mm long, slightly sulcate above, rounded and smooth or slightly ribbed below, glabrous, blades (4.5-)7-11(-13) × (1.5-)2.5-4(-5) cm, elliptic or narrowly elliptic, pinninerved, secondary veins 6-11 pairs, both upper and lower leaf surfaces glabrous, leaf apex caudate, sometimes acuminate, base cuneate to obtuse. Inflorescences 2.5-5 cm long, axillary to tiny, decidous bracts, disposed on very short shoots axillary to leaves, botryoid, or apparently paniculate, due to suppression of terminal bud of the floriferous branchlet, glabrous throughout, peduncle 0.2-1.7 cm long, pedicels (6-)10-13(-16) mm long, glabrous. Flowers widely obovoid to turbinate, greenish-yellow, tepals concave, conspicuously inflexed, outer tepals 0.9-1.3 × 1.1-1.7 mm, widely ovate, blunt cuspidate, glabrous abaxially and adaxially, short-ciliate, inner tepals 0.75-1.1 × 1.1-1.2 mm, ovate to widely ovate, glabrous abaxially and adaxially, short-ciliate, staminodes of whorls I and II absent, stamens of whorl III 0.6-1 mm long, fused throughout, filaments glabrous abaxially and adaxially, sometimes with a few hairs at the base abaxially, anthers ca. 0.1 mm, glabrous, with apical openings, glands ca. 0.4 mm, free, oblong, sometimes almost square, obtuse or sub-acute, glabrous, hypanthium 0.6-0.9 mm deep, glabrous outside and inside, pistil ca. 1.5 mm long, glabrous throughout, ovary 0.7-0.9 mm long. Fruits ca. 27 × 21 mm, ovoid or ellipsoid, cupule ca. 15 × 20 mm, crateriform, clearly bimarginate, inner margin ca. 2 mm tall, erect, outer margin ca. 3.5 mm tall, oblique to slightly reflexed, shallowly lobed, thick, pedicel ca. 5.5 × 4.5 (at the base) and 7 (at the apex) mm, obconic, continuous with the cupule.
Taxonomic summary
Type. Costa Rica. Puntarenas: Reserva Forestal Golfo Dulce, Osa Peninsula, Rancho Quemado, 8°44’ N, 83°36’ W, 200-300 m, 2 May 1988, B. Hammel et al. 16790 (holotype MO; isotypes MEXU, TEX).

Figure 1. Licaria breedlovei Lorea-Hern. sp. nov., general view.
Paratypes. Costa Rica. Guanacaste: Cantón de Tilarán, San Gerardo Abajo, río Caño Negro, Fincas Quesada y Arce, 10°18’40’’ N, 84°50’02’’ W, 1,100-1,200 m, 5 December 1991, E. Bello & E. Cruz 4262 (XAL). Puntarenas: Cantón de Osa, Rincón, filas al margen izquierdo de Quebrada Vaquedano, 8°38’45’’ N, 83°35’25’’ W, 400 m, 21 July 1990, G. Herrera 4000 (XAL); Cantón de Osa, Aguabuena, cuenca media y superior de Quebrada Orito, Rincón, 8°42’40’’ N, 83°31’40’’ W, 400 m, 25 October 1990, G. Herrera 4510 (MEXU); Cantón de Osa, Rancho Quemado, sector oeste, Sierpe, 8°41’00’’ N, 83°35’40’’ W, 350 m, 25 August 1982, J. Marín & G. Marín 499 (MO, XAL).

Figure 2. Licaria dolichopoda Lorea-Hern. sp. nov., general view.
Etymology. The name of this species alludes to the distinctive long pedicels that bear the flowers.
Distribution and habitat. Currently the species is known from 2 rather distant areas in Costa Rica that differ in ecological conditions. One of them, in the province of Guanacaste, is located in the Pacific foothills of the southern end of the Sierra de Guanacaste, while the other, in the province of Puntarenas, is in the lowlands of the Peninsula de Osa toward the southeastern extreme of the country. The first one is covered by pre-montane humid forest, and the second by humid tropical forest.
Phenology. Flowering is apparently distributed in 2 peaks: May-July and October-December. Fruits are only known from October.
Conservation status. There is no information about the abundance of the species in the places where it has been collected, neither about the ecological conditions of the vegetation there. However, the distribution range in the southeast is embedded in the Reserva Forestal Golfo Dulce, by the boundaries with the Parque Nacional Corcovado. Thus, there might not be problems with the persistence of the species in that region. The other point of the species distribution is, according to satellite images, within a region with deeply transformed vegetation, although not too far from the Parque Nacional Volcán Arenal, which could hold the species in its flora.

Figure 3. Inflorescence detail of Licaria breedlovei Lorea-Hern. sp. nov. (above), and Licaria dolichopoda Lorea-Hern. sp. nov. (below). Scale bars 1mm.
Remarks
Because of the erect, concave tepals, and extrorse sporangia of stamens in the flowers, L. dolichopoda must be also considered in the subgenus Licaria. The glabrous condition found in every structure of the plant body (stem, leaves, and flowers), as well as the apical openings of stamens, and the lack of staminodes found in this species places it beside L. eurypaniculata (here described), from which it differs in the botryoid architecture of the inflorescence, and the conspicuous caudate leaves. Collection duplicates of L. dolichopoda have been previously distributed as L. cufodontisii Kosterm. (Herrera 4000), and as L. misantlae (Herrera 4510).
Licaria eurypaniculata Lorea-Hern. sp. nov. (Figs. 4, 6)
Diagnosis. Among the species with glabrous leaves, inflexed tepals, lacking staminodes, as well as apical anther openings, this species is distinguished by paniculate inflorescences where the flowers are not aggregate on the floriferous axes, orbicular flowers, tepal surface glabrous abaxially and adaxially, fused stamens throughout, glabrous hypanthium outside and inside, and glabrous pistil.
Trees 6-7 m tall; twigs glabrous, smooth or slightly ribbed, reddish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (5.5-)7-9(-10.5) mm long, slightly sulcate above, glabrous, blades (7-)11-16(-19.5) × (2-)3-4.5(-6) cm, narrowly elliptic to lanceolate or elliptic, pinninerved, secondary veins 7-9 pairs, both upper and lower leaf surfaces glabrous, leaf apex acuminate to short caudate, base obtuse to cuneate. Inflorescences 10-11 cm long, apparently terminal, if it is axillary to a bract, this one is inconspicuous, paniculate, basal axes longer than the peduncle, glabrous throughout or sparsely puberulent toward the end of the secondary axes, flowers spaced, not aggregate on the axes, peduncle ca. 3 cm long, pedicels 2-3.2 mm long, glabrous or sparsely puberulent. Flowers spheroidal, greenish, tepals conspicuously inflexed, outer tepals ca. 0.5 × 0.9-1.0 mm, widely ovate to depressed ovate, glabrous abaxially and adaxially, inner tepals ca. 0.4 × 0.6-0.7 mm, ovate or almost orbicular, glabrous abaxially and adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.5-0.6 mm long, fused throughout, filaments glabrous outside, sparsely tomentose inside, anthers ca. 0.1 mm, glabrous, with apical openings, glands ca. 0.1 mm, free, widely elliptic or orbicular, glabrous, hypanthium ca. 0.7 mm deep, glabrous outside, glabrous or sparsely pubescent on the distal section inside, pistil ca. 1.1-1.2 mm long, glabrous throughout, ovary ca. 0.6-0.7 mm long. Fruits (not wholly ripe) 15-17 × 12-12.5 mm, ovoid or ellipsoid, cupule 10-11 × 14-15 mm, crateriform, clearly bimarginate, inner margin 1.1-1.3 mm tall, erect, outer margin 0.5-0.7 mm tall, perpendicular to the inner one, pedicel 3-3.6 × 2-2.2 (at the base) and 3.6-4 (at the apex) mm, obconic.
Taxonomic summary
Type. Panama. Bocas del Toro: along road to Chiriquí Grande, c. 10 road miles from continental divide and about 2 miles along road east of highway, 8°45’ N, 82°15’ W, 300 m, 15 April 1987, G. McPherson 10830 (holotype MO; isotype XAL 124596).
Paratype. Panama. Bocas del Toro: along road to Chiriquí Grande, 10 road-miles from continental divide, ca. 2 road-miles along road east of highway, 8°55’04’’ N, 82°10’04’’ W, 300 m, 9 February 1987, G. McPherson 10453 (MO 5048286).
Etymology. The name of this species alludes to the very long basal secondary axes of the inflorescence, which give a broad triangular profile to the panicle.
Distribution and habitat. The only 2 known collections of the species come from the same area in the Atlantic lowlands of western Panama. No information about the vegetation found at the site was recorded, but considering the geographical factors of the place, it is expected to be tropical rain forest.
Phenology. Flowers in spring, and fruits mature in winter or early spring. The plant is not deciduous.
Conservation status. It seems that the species is not frequent in the area where it was found, for it has been collected only twice. On the other hand, the view of the area from satellite images shows that most of the original vegetation has been cleared; therefore, the species might be endangered.
Remarks
There are no species that come close morphologically to L. eurypaniculata. The other species with a general glabrous condition almost throughout the plant body, L. dolichopoda, is very different, in the shape of the leaves and the structure of the inflorescence. Given the erect, concave tepals, and stamens with extrorse sporangia that L. eurypaniculata presents, it is also a member of the subgenus Licaria.
Licaria gibbitepala Lorea-Hern. sp. nov. (Figs. 5, 6)
Diagnosis. Trees similar to L. excelsa, but distinguished by the obovate to oblanceolate leaves, densely yellowish to orange-brown puberulent inflorescences, tepals with basal half conspicuously swollen, puberulent abaxial surface, sericeous adaxial surface at the base, apically pubescent ovary, and pubescent style.
Trees 15-20 m tall, trunk ca. 25 cm DBH, bark smooth, fragrant; twigs hollow, inhabited by ants, glabrous, sparsely lenticellate, slightly ridged, each ridge originating at one extreme of petiole insertion, acropetal. Buds glabrous or partially pubescent. Leaves alternate, petioles 10-12(-14) mm long, bimarginate above, glabrous, blades (20-)24-30 × (9-)11-13 cm, obovate or oblanceolate, sometimes elliptic, pinninerved, secondary veins (12)14-16 pairs, leaf surface glabrous above and below, leaf apex apiculate, sometimes apiculate-acuminate, leaf base obtuse or rounded. Inflorescences 9-15 cm, seemingly terminal, but actually axillary to tiny, deciduous bracts, on the proximal section of new twigs, paniculate, conspicuously puberulent along all axes, hairs yellowish to orange-brown, peduncle (1-)3.5-4.5 cm, sparsely puberulent, glabrescent, pedicels 2.5-5.5(-7) mm, densely puberulent. Flowers widely obovoid, yellowish-green, fragrant, perianth thick, coriaceous, tepals clearly inflexed, their base conspicuously swollen, outer tepals 1.2-1.4 × 1.6-1.9 mm, widely ovate, puberulent abaxially, sericeous-tomentose at the base and toward the margins adaxially, hairs orange-brown, inner tepals 0.8-1 × 1-1.2 mm, ovate, puberulent abaxially, sericeous-tomentose at the base and tomentose at the middle adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.7-0.8 mm long, fused along their filaments, filaments tomentose at the base outside, tomentose throughout inside, hairs orange-brown, anthers ca. 0.2 mm, free or fused just at the base, glabrous, with apical openings, glands ca. 0.3 mm, only 3 given the fusion of adjoining glands, widely oblong, glabrous, hypanthium 1-1.2 mm deep, obconic, densely puberulent outside, hairs yellowish to orange-brown, sericeous-tomentose inside, hairs reddish-brown, pistil 1.6-1.8 mm long, top of the ovary and style pubescent, ovary 0.8-1.2 mm long. Fruits (nearly ripe) 19-23 × 15.5-16.5 mm, ellipsoid, cupule ca. 16.5 × 18.5 mm, cotyliform, sparsely lenticellate, seemingly tri-margined, the 2 regular margins plus the swollen projections of the tepals, inner margin ca. 1.6 mm tall, erect, outer margin 2.2-2.5 mm tall, erect, pedicel 4.5-7 mm long, continuous with the base of the cupule, 3.5 mm diam. at the base.
Taxonomic summary
Type. Costa Rica. Limón: Reserva Biológica Hitoy Cerere, 300 m aguas abajo de la confluencia del río Hitoy con el río Cerere, margen izquierda por la fila que lleva al cerro Bobócora, 9°39’00’’ N, 83°02’45’’ W, 200 m, 20 February 1989, G. Herrera & A. Chacón 2427 (holotype MO, isotype XAL 124445).
Etymology. The name is derived from the conspicuous swollen condition of the base of the tepals; the feature is distinctive.
Distribution and habitat. So far, this species is known only from the southwestern end of Costa Rica. Although no information about the habitat was recorded, there is no doubt that the place lies within the tropical rain forest territory.

Figure 4. Licaria eurypaniculata Lorea-Hern. sp. nov., general view.
Phenology. With flowers and ripe fruits around the end of winter. The species has perennial leaves.
Conservation status. There is no information about the abundance or extent of distribution of the species, but as it is known to grow in the grounds of a nature reserve, it can be expected that it is not under high risk of extinction but certainly endangered.

Figure 5. Licaria gibbitepala Lorea-Hern. sp. nov., general view.
Remarks
Licaria gibbitepala seems to be closely related to L. tomentulosa and, at the same time, to the group of species around L. excelsa. All of them have ridged twigs, large leaves, flowers with erect, concave tepals, no staminodes, and sporangia with apical openings. The hollow twigs, along with the obovate to oblanceolate leaves, and the conspicuous gibbous base of the tepals distinguish L. gibbitepala.
Licaria gracilis Lorea-Hern. sp. nov. (Figs. 7, 9)
Diagnosis. Small trees with glabrous leaves, botryoid, few-flowered inflorescences, glabrous, erect tepals, lacking staminodes, free stamens, anther openings elliptic, lateral, strongly oblique, glands present, glabrous hypanthium outside and inside, glabrous pistil.

Figure 6. Inflorescence detail of Licaria eurypaniculata Lorea-Hern. sp. nov. (above), and Licaria gibbitepala Lorea-Hern. sp. nov. (below). Scale bars 1mm.
Trees to 6 m tall; twigs smooth, dark brown to reddish brown, pruinose, puberulent, glabrescent, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (4-)8-11 mm long, puberulent, soon glabrous, canaliculate above, blades 8.5-13 × 3.5-5.5 cm, elliptic or narrowly elliptic, pinninerved, secondary veins 5-7 pairs, leaf surface glabrous above and below, but the lower surface puberulent in the beginning, leaf apex caudate, sometimes just acuminate, base obtuse to cuneate. Inflorescences 2.5-3.5 cm long, axillary to leaves and to tiny, deciduous bracts, on the proximal section of new twigs, botryoid, racemiform, few-flowered (less than 10 flowers), glabrous throughout or with some hairs toward the end of the peduncle, peduncle 1.8-2 cm long, pedicels 3-4.5 mm long, glabrous. Flowers obovoid, pale yellow, tepals erect to slightly inflexed, concave, outer tepals ca. 0.8 × 1.3 mm, very widely ovate to depressed ovate, glabrous abaxially, with some long, appressed hairs ascending from the base adaxially, inner tepals ca. 0.5 × 0.8 mm, ovate or widely ovate, glabrous abaxially, adaxially like the outer tepals, staminodes of whorls I and II absent, stamens of whorl III ca. 0.7 mm long, free, but very close one to each other, widely ovate in outline, filaments sparsely tomentulose outside and inside, anthers ca. 0.4 mm, tomentulose at the base outside, tomentulose inside along the central line, openings lateral, oblique, glands ca. 0.4 mm, rounded, glabrous, hypanthium ca. 0.7 mm deep, obconic, glabrous inside and outside, pistil ca. 1.7 mm long, glabrous, ovary ca. 0.8 mm long. Fruit unknown.

Figure 7. Licaria gracilis Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Panama. Chiriquí: Punta Burica, El Chorogo, alrededores de la finca de Fernando Chavarría, adyacente al límite fronterizo, cabecera del río San Bartolo, 8°17’07’’ N, 82°58’56’’ W, 395 m, 15 May 2007, J. E. Aranda et al. 3912 (holotype MO 6456198).
Etymology. The name of the species is derived from the attractive view that the slender inflorescences give to the plant.
Distribution and habitat. The species is known only from the location where it was first and last collected. No information about the habitat is mentioned in the data-label of the specimen, except that was collected nearby a ranch. The original vegetation must have been tropical evergreen forest or semi-evergreen forest.
Phenology. Flowers during spring; ripe fruits expected during winter or early spring, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. The species might be (critically) endangered, for most of the land in the area where it was collected has been transformed for diverse agricultural purposes.
Remarks
The few-flowered, botryoid inflorescence, tiny flowers, and very oblique, lateral openings of the sporangia distinguish this species straightaway. There is no species whose general morphology indicates association with L. gracilis. The singular way that the openings of the stamens are displayed resemble that found in Licaria cogolloi van der Werff, and L. caribaea Gómez-Lau. & Cascante, but besides this, there is no other feature that might suggest a relationship to those species. For the features of its flowers, Licaria gracilis belongs to subgenus Licaria.
Licaria minutiflora Lorea-Hern. sp. nov. (Figs. 8, 9)

Figure 8. Licaria minutiflora Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. pergamentacea, but distinct by the presence of hollow twigs, glabrous buds, abaxially densely tomentulose tepals, internally pubescent hypanthium, apically pubescent ovary, and pubescent style.
Trees up to 20 m tall, trunk ca. 30 cm DBH; twigs hollow, inhabited by ants, glabrous, smooth or slightly ridged, each ridge originates at one extreme of petiole insertion, acropetal, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (7-)15-20(-30) mm, glabrous, bimarginate above, blades (11-)20-28(-34.5) × (3-)5-9(-13) cm, narrowly elliptic or narrowly ovate, pinninerved, secondary veins 9-12 pairs, leaf surface glabrous on both sides, leaf apex acute, slightly apiculate, base acute or obtuse. Inflorescences (6-)9-12(-15) cm long, axillary to tiny, deciduous bracts, on the proximal section of new twigs or axillary to leaves, paniculate, tomentulose, peduncle (0.5-)1.5-3(-4) cm long, pedicels (1.2-)2.5-4(-6) mm long, densely tomentulose. Flowers obovoid or ellipsoid, yellowish-green, tepals inflexed, outer tepals 0.5-0.8 × 0.8-1 mm, widely ovate, densely tomentulose abaxially, tepal surface concealed or almost so by hairs, glabrous adaxially, inner tepals 0.5-0.7 × 0.5-0.6 mm, ovate, densely tomentulose abaxially, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.5-0.6 mm long, free or barely united by filament base, filaments tomentose at base outside, glabrous or sparsely pubescent along the medial line inside, anthers ca. 0.2 mm, glabrous, openings apical, glands ca. 0.2 mm, free, rounded, hypanthium 0.6-0.8 mm deep, obconic, densely tomentulose outside, sericeous on upper half inside, hairs yellowish or reddish, pistil ca. 1.4 mm long, top of the ovary and style pubescent, ovary 0.8-1 mm long. Fruits ca. 19.5 × 15-16.5 mm, ellipsoid, cupule 15-16 × 18-20 mm, urceolate, lenticellate, conspicuously bimarginate, inner margin 1.2-1.5 mm tall, outer margin 1-1.2 mm, extended, pedicel 6.5-9 mm long, continuous with the cupule, 2.8-3.6 mm diameter at base.
Taxonomic summary
Type. Costa Rica. Alajuela: camino entre la estación de la Reserva Forestal de San Ramón y el camino a la colonia Palmareña, finca de don Bolívar Ruiz, margen derecha río San Lorencito, 10°12’53’’ N, 84°36’28’’ W, February 1987, G. Herrera 500 (holotype MO 3587621; isotypes MEXU 638804, TEX).
Paratypes. Costa Rica. Alajuela: Reserva Biológica Monteverde, Poco Sol, La Cutacha de San Bosco, 10°22’ N, 84°40’ W, 900 m, 1 April 1989, E. Bello 784 (MEXU 1304162; MO 6142952; XAL 124442); Bosque Eterno de los Niños, Reserva de Arenal, río Peñas Blancas, Quebrada Agua Gata, Finca Villalobos, 10°23’ N, 84°42’ W, 1,000 m, 20 April 1990, E. Bello 2208 (MO 6130824; XAL 124446); Cantón de Upala, Colonia La Libertad, 10°52’ N, 85°17’ W, 300 m, 3 August 1991, Q. Jiménez & G. Rivera 1011 (MO 6117397; XAL 124597). Guanacaste: Parque Nacional Guanacaste, Estación Pitilla, 10°00’15’’ N, 85°25.2’ W, 500 m, 27 May 1989, G. Herrera et al. 2942 (MO; XAL 124428).

Figure 9. Inflorescence detail of Licaria gracilis Lorea-Hern. sp. nov. (above), and Licaria minutiflora Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Etymology. The very small flowers, whose tepals are less than 1 mm long, is the feature on which the species name is based.
Distribution and habitat. This species is known from the northern hills of the Cordillera de Guanacaste (Guanacaste Mountain Range) and the western part of the Cordillera Central (Central Mountain Range), between 200 and1,000 m asl. Prevailing vegetation in the region is tropical evergreen forest and montane rain forest. The species has been collected also in pasture fields. There is a fruiting specimen from the Osa Peninsula (Hammel et al. 16984) that seems to belong to L. minutiflora but until confirmed with a flowering specimen, the presence of the species in this part of the country remains uncertain.
Phenology. Flowers toward the end of winter and early spring; fruits must be ripe around the end of autumn or early winter.
Conservation status. Most specimens of this species have been collected within nature reserves. Therefore, it is considered not threatened, even though its abundance is still unknown.
Remarks
As has been mentioned elsewhere in this paper, Licaria minutiflora is part of the L. excelsa species group. Its general appearance resembles that of L. pergamentacea, for the size and shape of the leaves, structure and hairiness of the inflorescence, and for having small oblong-ellipsoid, not coriaceous flowers. However, besides the presence of hollow twigs, it differs by having flowers with the upper part of the hypanthium homogeneously pubescent inside, and a pubescent pistil.
Licaria ochracea Lorea-Hern. sp. nov. (Figs. 10, 12)

Figure 10. Licaria ochracea Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. multinervis H. W. Kurz, but differing by lower surface indument of leaves composed by 2 types of hairs, the most numerous tomentulose, the fewer sericeous, long inflorescences, terminal or axillary to leaves, with rachis up to 15 cm, fully exserted anthers, clearly stalked glands, conspicuous hypanthium tube projected beyond the insertion point of stamens.
Trees up to 22 m tall, twigs smooth, densely tomentose, hairs initially yellowish-brown, then greysh, sparsely and inconspicuously lenticellate. Buds tomentose. Leaves alternate, petioles (10-)15-20 mm long, tomentose, glabrescent, channeled above, blades 12.5-18.5 × (2.5)3.5-4.5 cm, narrowly elliptic, sometimes narrowly oblanceolate, pinninerved, secondary veins (9)10-13 pairs, leaf surface glabrous above, tomentose below, most hairs sinuous, rather appressed, or patent, fewer hairs straight, appressed, leaf apex acuminate, base narrowly cuneate. Inflorescences (7-)10-18 cm long, axillary to leaves, less frequently terminal, paniculate, densely tomentose, hairs like on twigs, flowers agglomerate at the end of terminal axes, peduncle 0.2-1.5(-3.5) cm long, pedicels (1-)1.5-2 mm long, tomentose. Flowers narrowly oblong, tepals erect, concave, outer tepals, 1-1.3(-1.5) × (1-)1.1-1.3 mm, widely ovate, tomentose abaxially, glabrous adaxially, except for a few long, appressed hairs, coming from the base, inner tepals 1-1.1(-1.4) × 0.8-0.9 mm, ovate, with a pattern of pubescence similar to that of outer tepals, staminodes of whorls I and II absent, stamens of whorl III 3-3.3(-3.8) mm long, coherent along their filaments or even at the base of the anthers, easily separable, filaments tomentose on both faces, anthers 0.9-1.1 mm long, free or coherent at the base, glabrous, completely exserted, openings dorsolateral, glands 0.7-0.8 mm, free, oblanceolate, glabrous, clearly stalked, stalk pubescent, hypanthium 1.2-1.3(-1.6) mm deep, extended 0.3 mm beyond the insertion point of stamens, tomentose outside, densely hirsute-tomentose inside, hairs golden-brown, pistil 3.6-3.9 mm long, sparsely pubescent, at least some hairs on the upper half of the ovary and lower half of the style, ovary 0.9-1.1 mm long. Fruits unknown.

Figure 11. Licaria rufotricha Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Nicaragua. Matagalpa (according to current maps it should be Jinotega): pasture and small woods, tropical premontane forest, Hacienda Santa María del Ostuma, 10 km N of Matagalpa, 1,300 m, 17 July 1978, P. C. Vincelli 756 (holotype MO 2984723; isotypes LL, MEXU 691292).
Etymology. The name refers to the conspicuous orange-brown pubescence of the species on young twigs and main axes of inflorescences.
Distribution and habitat. Only known from the place of type collection, where the Cordillera Isabelia and Cordillera Dariense meet.
Phenology. Flowers in summer; fruit season is not known, but since it has been observed that maturation of fruits in most Lauraceae takes around a year after flowering, possibly ripe fruits are present during spring or early summer.
Conservation status. There is no certainty on the conditions that might be affecting the survival of the species. The type was collected in a private property that used to be preserved for ecotourism activities, but its current situation is unknown. However, since the species has not been collected again, it is suspected to be endangered.
Remarks
The collection on which the description of this species is based was initially identified by Kurz (2000) as L. multinervis; actually it is cited as a paratype of this species. But the impression of being conspecific with this taxon disappears with a detailed survey of the morphology. Important differences are the pubescence on the lower surface of the leaves (simple in L. multinervis vs. made of 2 types of hairs in L. ochracea), position of the inflorescences (axillary to small bracts in the proximal section of new branches in L. multinervis vs. axillary to leaves or terminal in L. ochracea), length of inflorescences (up to 3.5 cm long in L. multinervis vs. 5-7 times longer in L. ochracea), protrusion of anthers (partially exserted in L. multinervis vs. fully exerted in L. ochracea), and projection of hypanthium tube beyond the insertion point of stamens (short in L. multinervis vs. conspicuous in L. ochracea). However, L. multinervis and L. ochracea seem to be more closely related to each other than to the rest of species in the area that present fully exserted stamens, namely L. agglomerata van der Werff, L. capitata (Schltdl. et Cham.) Kosterm., L. nitida van der Werff, and L. vanderwerffii (here described).
Licaria rufotricha Lorea-Hern. sp. nov. (Figs. 11, 12)
Diagnosis. Small trees, distinct for the conspicuously obovoid flowers, aggregate toward the end of the secondary floriferous axes, with inflexed tepals, glabrous abaxially, reddish-brown hirsute-sericeous adaxially, lacking staminodes, stamens fused throughout, reddish-brown hirsute, anther openings apical, hypanthium mostly glabrous outside, reddish-brown hirsute inside, and ovary distally pubescent.
Trees to 12 m tall; twigs smooth or inconspicuously ridged, each ridge originating at one extreme of petiole insertion, acropetal, lenticellate, grayish, densely puberulent, glabrescent. Buds glabrous. Leaves alternate, petioles 10-13 (-17) mm long, slightly channeled above, puberulent, blades (14.5-)16.5-20(-23) × 4-5.5(-9) cm, elliptic or narrowly elliptic, pinninerved, secondary veins 6-7 pairs, leaf surface densely puberulent above when young, glabrescent, rather persistently puberulent below, leaf apex acuminate, base cuneate, sometimes obtuse or narrowly cuneate to indistinctly attenuate. Inflorescences (3-)5-8(-10.5) cm, axillary to tiny, deciduous bracts, on the proximal section of new twigs, or axillary to new leaves, paniculate, flowers aggregate at the end of secondary axes, densely puberulent on peduncle and rachis, glabrate toward the ultimate axes, peduncle (2-)3.5-5.5(-7.5) cm long, pedicels 2-3.5(-4.5) mm long, glabrous or sparsely puberulent. Flowers obovoid or obconic, pale yellow, tepals conspicuously inflexed, outer tepals 1-1.2 × 2-2.4 mm, widely ovate, glabrous abaxially, hirsute-sericeous adaxially, hairs reddish-brown, inner tepals 0.8-1 × 0.8-0.9 mm, ovate or elliptic, glabrous abaxially, generally mostly hirsute adaxially, hairs reddish-brown, surpassing the apical margin of the tepals, making them appear fimbriate, staminodes of whorls I and II absent, stamens of whorl III, 0.6-0.7 mm long, fused throughout, filaments hirsute on both faces, hairs reddish-brown, anthers ca. 0.2 mm, pubescent, except around the margin of the openings, hairs like those of the filaments, glands ca. 0.4 mm, free, rounded or oblate, glabrous, hypanthium 0.8-0.9 mm deep, glabrous or sparsely puberulent outside, hirsute inside, hairs like those of the filaments, pistil 1.7-2 mm long, ovary ca. 0.8 mm long, sparsely pubescent distally, hairs whitish. Fruits unknown.

Figure 12. Inflorescence detail of Licaria ochracea Lorea-Hern. sp. nov. (above), and Licaria rufotricha Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Taxonomic summary
Type. Panama. Panama: 5-10 km NE of Altos de Pacora, on trail at end of road, ca. 750 m, 7 March 1975, S. Mori & J. Kallunki 4977 (holotype MO 2992664).
Etymology. The name given to this species refers to the dense red-brown pubescence in the interior of the flowers, covering the adaxial surface of tepals, stamen filaments, anthers, and hypanthium inside.
Distribution and habitat. Just known from the place where the type was collected (around 25-30 km NE of Panama City), with tropical evergreen forest or semi-evergreen forest as original vegetation.
Phenology. Flowers around the end of winter or early spring; ripe fruits expected during winter, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. Probably the species is endangered. Most of the vegetation in the surroundings of the place where the species was collected has been cut down for cattle raising or agricultural purposes. But the southern boundary of the Parque Nacional Chagres (Chagres National Park) is very close to that place, and the Nature Reserve Cocobolo is a few kilometers ENE of it as well. So, even it has not been collected again, it is hoped the species is still around there.
Remarks
There is no other species in the Mesoamerican area with the distinctive reddish-brown pubescence found in the interior of the Licaria rufotricha flowers. Yellowish to reddish-brown pubescence is present on the filaments of stamens and/or inside the hypanthium of several species, like L. agglomerata, L. excelsa, L. gibbitepala, and L. tomentulosa, but it is never found on the tepals surface. In fact, its general morphology does not suggest a relation to any species in the region.
Licaria sarukhanii Lorea-Hern. sp. nov. (Figs. 13, 15)
Diagnosis. Trees similar to L. alata, but differing by terminal, subsessile inflorescences, longer pedicels (up to 10.5 mm), flowers widely ellipsoid, abaxial tepal surface tomentose at the base and papillose toward the apex, hypanthium densely tomentose outside, sericeous-tomentose inside, and style pubescent.
Trees to 20 m tall, twigs glabrous, grayish to grayish-brown, slightly ridged, each ridge originating at one extreme of petiole insertion, acropetal, sparsely lenticellate. Buds glabrous or pubescent at the apex of the bud scales. Leaves alternate, petioles (11-)13-17(-20) mm long, slightly bimarginate, glabrous, blades (12.5-)20-32(-37) × (5-)7-10(11.5) cm, narrowly elliptic, occasionally oblanceolate, pinninerved, secondary veins (12)14-18 pairs, leaf surface glabrous on both sides, leaf apex acuminate, sometimes acute, base obtuse or acute. Inflorescences 12-22 cm long, terminal, paniculate, tomentose, soon glabrescent, hairs patent or oblique, grey or yellowish-grey, peduncle 0.2-0.4 cm long, pedicels (3-)5-8(-10.5) mm long, glabrous or sparsely tomentose. Flowers broadly ellipsoid, pale green, tepals inflexed, outer tepals 0.8-0.9 × 1.8-2 mm, very widely ovate, concave, tomentose over the basal half, and papillose-pubescent over the distal half abaxially, glabrous or with few straight hairs at base adaxially, inner tepals 0.7-0.9 × 0.4-0.8 mm, ovate, pubescence pattern abaxially like that of outer tepals, but less papillose, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.7-0.9 mm long, fused along their filaments, filaments tomentose along the central line on both faces, anthers 0.2-0.4 mm long, glabrous, openings apical, glands ca. 0.4 mm, oblong, flattened, frequently reduced or absent, glabrous or tomentose on the base abaxially, hypanthium ca. 1 mm deep, densely tomentose outside, hairs sinuous, sericeous-tomentose inside, pistil 1.3-1.6 mm long, ovary 0.8-1.2 mm long, glabrous, style rather densely pubescent. Fruits 27-30 × 17-18 mm, ellipsoid, cupule 15-17.5 × 18-19.5 mm, urceolate, bimarginate, but the margins barely discernible, ca. 0.5 mm, erect, pedicel 4-5 × 3.5-4.5 mm.

Figure 13. Licaria sarukhanii Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Mexico. Chiapas: municipio de Ángel Albino Corzo, Reserva de la Biosfera El Triunfo, aprox. 1.5 km al E de Campamento El Triunfo, sobre el Sendero Bandera, 15°36’ N, 92°50’ W, 1,850 m, 27 April 1993, F. Lorea 5522 (holotype XAL 151063; isotypes [to be distributed]).
Paratypes. Mexico. Chiapas: municipio de Ángel Albino Corzo, Polígono I de la Reserva de la Biosfera El Triunfo, 15°39’ N, 92°48’ W, 1,900 m, 21 February 1993, S. Solórzano 67 (MEXU 754117); Reserva El Triunfo, camino a Cerro de la Bandera, 25 March 1986, M. L. Ávila and V. H. Hernández s/n (MEXU 877857, TEX 146383); Reserva de la Biosfera El Triunfo, sendero Palo Gordo, 15°40’10’’ N, 92°48’42’’ W, 1,990 m, 22 March 2006, F. González-García s/n (XAL 113841).
Etymology. The species is dedicated to José Sarukhán, a Mexican plant ecologist who has played an important role in making a huge amount of information about Mexico’s biodiversity, ecosystem conservation, and sustainable development accessible to the public.
Distribution and habitat. So far, the species is known only from the central region of the Sierra Madre de Chiapas, from 1,800 to 2,000 m asl where the montane rain forest is the dominant type of vegetation.
Phenology. Flowers toward the end of winter and early spring; ripe fruits were found in the same interval of time.
Conservation status. All the collections of the species come from the nature reserve El Triunfo, located in the Sierra Madre de Chiapas, which has persisted largely undisturbed for its almost forty years of existence. Thus, considering the size of the area covered with montane rain forest in the reserve, the species might be regarded as vulnerable.
Remarks
Licaria sarukhanii is part of the L. excelsa species group, with a closer relation to L. breedlovei, because they share tomentose inflorescences and flowers rather widely ellipsoid to obloid. They differ by the conduplicate leaf bases, sparsely sericeous lower leaf surface, conspicuously pedunculate inflorescences axillary to small bracts or leaves, fused anthers, and style sparsely pubescent present in L. breedlovei, in contrast to flat leaf base, glabrous lower leaf surface, terminal, sub-sessile inflorescences, free anthers, and conspicuously pubescent style in L. sarukhanii.
Licaria tomentulosa Lorea-Hern. sp. nov. (Figs. 14, 15)
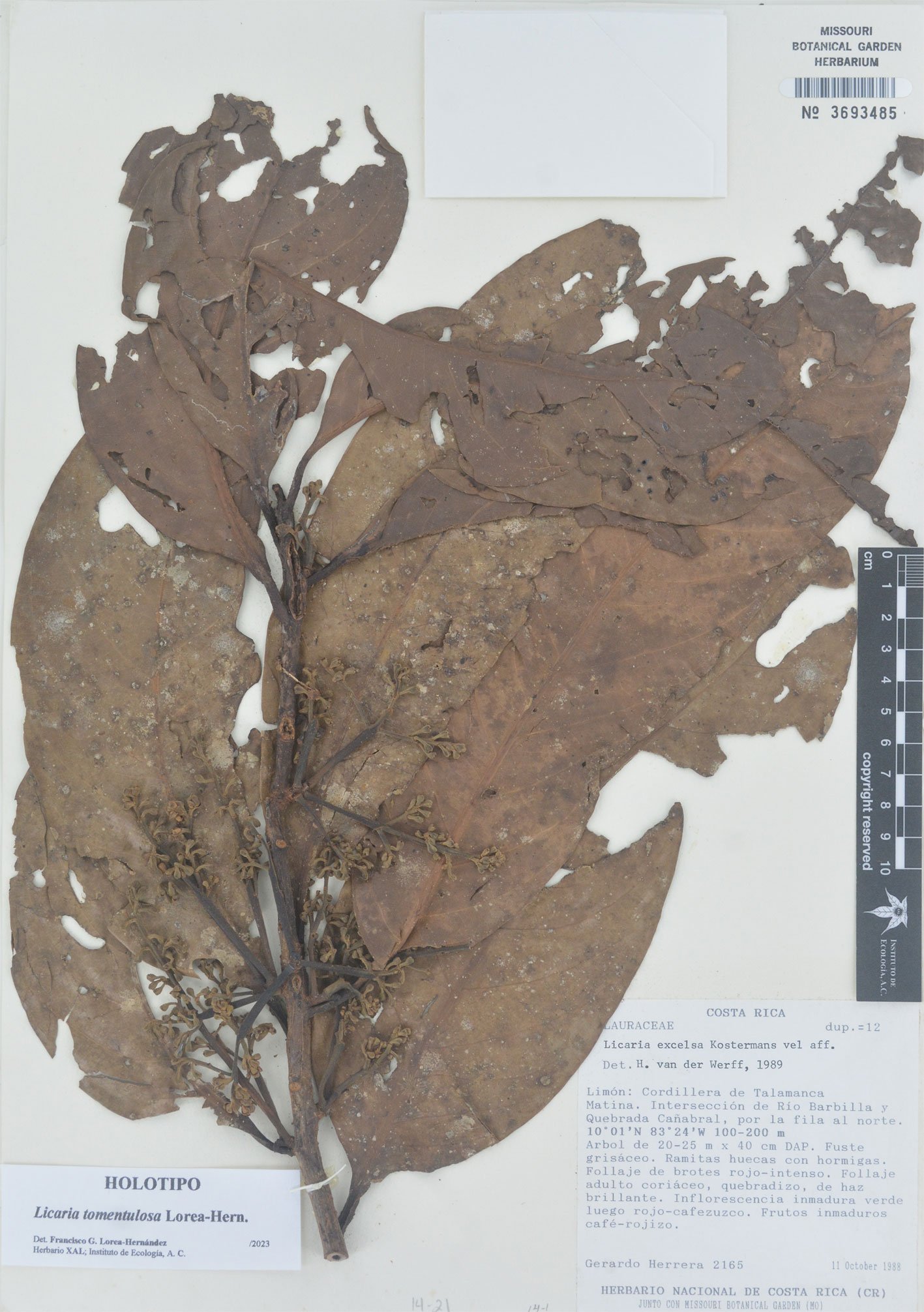
Figure 14. Licaria tomentulosa Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. excelsa, but distinguished by presenting hollow twigs, inflorescences yellowish-brown tomentulose, flowers conspicuously obovoid, tepals densely tomentulose abaxially, and apex of the ovary and style conspicuously pubescent.
Trees up to 25 m tall, and trunk 50 cm DBH, twigs hollow, inhabited by ants, smooth or slightly ridged, glabrous, grayish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (14-)17-22(-24) mm long, bimarginate above, glabrous, blades (22-)25-30(-34) × (6-)8-10(-12) cm, narrowly elliptic or elliptic, pinninerved, secondary veins (9)11-13 pairs, leaf surface glabrous on both sides, leaf apex acute or short acuminate, base obtuse, rounded or shortly attenuate, particularly in leaves close to the twig tips. Inflorescences 6.5-9.5(13.5) cm long, axillary to tiny, deciduous bracts, on very short branches axillary to leaves, paniculate, tomentulose, hairs yellowish-brown, peduncle 2.5-4 cm long, pedicels (1-)2-3.5 mm long, densely tomentulose. Flowers obovoid, thick, coriaceous, greenish-yellow, tepals inflexed, concave, outer tepals 0.7-1 × 1.4-1.7 mm, widely ovate, densely tomentulose abaxially, hairs almost concealing the surface, marginal tomentose bands and some long, appressed hairs at the base adaxially, inner tepals 0.6-0.9 × 0.8-0.9 mm, ovate, sometimes slightly conduplicate, densely tomentulose abaxially, pattern of pubescence adaxially like in the outer tepals, staminodes of whorls I and II absent, stamens of whorl III 0.8-1.1 mm long, fused throughout, filaments tomentose outside on lower half, densely tomentose all over the inner face, hairs reddish-brown, anthers ca. 0.2 mm, glabrous outside, densely tomentose inside, openings apical, glands 0.3-0.4 mm, frequently fused between adyacent stamens, transversely oblong, oblong when free, hypanthium 1-1.2 mm deep, densely tomentulose outside, tomentulose inside, sometimes only in the upper half, hairs mainly reddish-brown, pistil 2-2.3 mm long, ovary 1.1-1.4 mm long, apex of the ovary and style conspicuously pubescent. Fruits (not fully ripe) ca. 13 × 13 mm, spheroidal, cupule 13-14 × 17-18 mm, crateriform, lenticellate, bimarginate, inner margin 0.7-1 mm tall, erect, outer margin 0.5-0.7 mm, erect to spreading, pedicel 4-6 × 3-4 (apex) and 2 mm (base).
Taxonomic summary
Type. Costa Rica. Limón: Cordillera de Talamanca, Matina, intersección de río Barbilla y quebrada Cañabral, por la fila al norte, 10°01’ N, 83°24’ W, 100-200 m, 11 October 1988, G. Herrera 2165 (holotype MO 3693485; isotypes MEXU 529888, 529889, 717774, and 718334; TEX).
Paratypes. Costa Rica. Alajuela: Upala, Bijagua, El Pilón, Cerro La Carmela, entre río Celeste y cabeceras del río Chimurria, 10°43’15’’ N, 84°59’45’’ W, 1,000 m, 11 July 1988, G. Herrera 2056 (MEXU 529875, 717781; MO 3693486; US 3655515). Limón: Reserva Biológica Hitoy Cerere, Valle de la Estrella, sendero a Cerro Bobócara, 9°41’00’’ N, 83°04’20’’ W, 798 m, 17 August 1990, G. Herrera 4115 (MEXU 1304090; MO 6142953; XAL 124444); Reserva Indígena Talamanca, camino a Soki entre la Quebrada Amubri, margen izquierda de río Lari, 9°29’40’’ N, 82°89’40’’ W, 200 m, 29 June 1989, A. Chacón 25 (MO; XAL 124437).
Etymology. The name is for the abundant tomentulose pubescence that covers the axes of the inflorescence, as well as the abaxial surface of tepals and hypanthium.
Distribution and habitat. Known only from Costa Rica, along the Atlantic lowlands adjacent to the Sierra de Talamanca, and the Sierra de Guanacaste. Tropical rain forest used to be the predominant vegetation there, but currently a big amount of land has been changed to agricultural activities or other affairs.
Phenology. Flowers in summer and early autumn; ripe fruits in summer.
Conservation status. Given the deterioration of the habitat, the species might be considered vulnerable or endangered, in spite of its wide range of distribution.
Remarks
Licaria excelsa and L. tomentulosa are much alike. Considering the available material, distinguishing characters seem to work well to separate both species; hollow twigs, as well as type and distribution of pubescence on vegetative and floral parts, are consistently present in all the specimens considered as L. tomentulosa, making it a clear morphological unit, distinct from L. excelsa.
Licaria vanderwerffii Lorea-Hern. sp. nov. (Figs. 16, 17)

Figure 15. Inflorescence detail of Licaria sarukhanii Lorea-Hern. sp. nov. (above), and Licaria tomentulosa Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Diagnosis. Similar to L. agglomerata, but different by the free stamens, white pubescence of stamens, hypanthium densely white tomentose inside, and pistil glabrous.
Shrubs or small trees 1-3 m tall, twigs hollow, smooth, sparsely lenticellate, glabrous, reddish-brown. Buds glabrous. Leaves alternate, petioles ca. 12.5-15 mm long, slightly channeled and bimarginate above, glabrous, blades ca. 15-23 × 6-7.5 cm, narrowly elliptic, pinninerved, secondary veins 9-11 pairs, leaf surface glabrous on both sides, leaf apex acuminate, base obtuse or cuneate. Inflorescences ca. 1.5-2 cm long, axillary to leaves, axillary to tiny, deciduous bracts, on the proximal sections of new twigs too, paniculate, but strongly condensed, with very short rachis and secondary axes, peduncle ca. 0.5 cm long, glabrous, pedicels (1.2-)1.5-2.5(-3) mm long, glabrous. Flowers ellipsoid, slightly swollen in the middle of the hypanthium, greenish, tepals erect, outer tepals 0.5-0.6 × 0.5-0.6 mm, ovate, glabrous abaxially, glabrous adaxially except for some sericeous hairs at the base, inner tepals 0.4-0.5 × 0.2-0.3 mm, ovate, glabrous abaxially, glabrous adaxially or with some sericeous hairs at the base, staminodes of whorls I and II absent, stamens of whorl III 1-1.2 mm long, free, filaments white-tomentose throughout, denser on inside, anthers 0.3-0.4 mm, wholly exserted, glabrous or sparsely tomentose at the base outside, tomentose on lower half inside, openings dorsal, glands absent, hypanthium 1.1-1.3 mm deep, narrowly rhomboid, extending a little beyond the insertion point of stamens, glabrous outside, densely white-tomentose inside, pistil 2-2.3 mm long, glabrous, ovary 1-1.2 mm. Fruit unknown.

Figure 16. Licaria vanderwerffii Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Panama. Darién: Cerro Tacarcuna S slope, 1,250-1,450 m, 26 January 1975, A. Gentry & S. Mori 13920 (holotype MO 2300651; isotype F 1763407).
Etymology. The species is named after Hendrik (Henk) Hessel van der Werff, a prolific scholar of the family Lauraceae in the Neotropics.
Distribution and habitat. So far, only known from the type collection, in the mountains of the southeastern edge of Panama, where premontane rain forest prevails.
Phenology. Flowers in winter; probably ripe fruits expected during autumn or early winter, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. The place of the type collection is within the boundaries of the nature reserve Parque Nacional del Darién, but the altitudinal range within which the specimen was collected stretches barely for 40 km along the reserve. So, the species should be considered endangered.
Remarks
The collection on which the description of L. vanderwerffii is based was cited by Kurz (2000) as L. triandra, but there are several features that separate the former from this species. Whereas L. triandra presents branchlets solid, paniculate inflorescences (up to 6 cm long), stamens fused throughout (or almost so), glands at the base of filaments, anthers partially exserted, and hypanthium glabrous inside, L. vanderwerffii has hollow branchlets, sub-capitate inflorescences (up to 2 cm long), stamens free, no glands at the base of filaments, anthers wholly exserted, and hypanthium densely white tomentose inside.

Figure 17. Inflorescence detail of Licaria vanderwerffii Lorea-Hern. sp. nov. Scale bar 1 mm.
This species is closely related to L. agglomerata; both present small, glomerate inflorescences, ellipsoid to narrowly rhomboid flowers with erect tepals, no staminodes, fully exserted stamens, no glands on filaments, pubescent stamens, and hypanthium tomentose within. However, L. vanderwerffii differs by having stamens free throughout, white tomentum on stamens and (densely on) inner face of hypanthium, and glabrous pistil, against stamens fused by their filaments, brownish-orange tomentum on stamens and upper part of hypanthium inside (sometimes completely glabrous), and puberulent style in L. agglomerata. It is worth to mention that they differ in ecological conditions as well; while L. agglomerata grows in lowland habitats (200-700 m), L. vanderwerffii is a mountain dweller above 1,200 m.
Acknowledgements
I would like to thank the curators of the herbaria cited in the Materials and methods section for allowing the study of specimens of Licaria in their collections. I also thank Jens Rohwer and an anonymous reviewer for their comments and calling my attention to several slips and wrong phrasing in some paragraphs. Eva Piedra assisted me with the preparation of figures. Phil Brewster kindly reviewed the final english version of the manuscript. I thank one of the reviewers who suggested to include the following exsiccata as part of the studied material for several of the species described here, however, it was not possible to see the specimens. So even they correspond to duplicates of the studied material, they are cited here just as a reference for researchers who might need to study them.
L. breedlovei; D. Breedlove 50394, isotype TEX 472525.
L. dolichopoda; B. Hammel et al. 16790, isotypes CR 147811 and CR 2918355; E. Bello & E. Cruz 4262, paratype CR 1596155; G. Herrera 4000, paratype CR 1596509; G. Herrera 4510, paratypes CR 159602, CR 176525 and CR 1561583; J. Marín & G. Marín 499, paratype CR 1596363.
L. gibbitepala; G. Herrera & A. Chacón 2427, isotype CR 1596536.
L. minutiflora; G. Herrera 500, isotype CR 151205; E. Bello 784, paratype CR 1513414; E. Bello 2208, paratype CR 1596135; Q. Jiménez & G Rivera 1011, paratypes CR 1561584 and CR 157780; G. Herrera 2942, paratype CR 1596539.
L. tomentulosa; G. Herrera 2165, isotypes CR 146925 and CR 2921047; G. Herrera 2056, paratypes CR 147034, and CR 2920969; G. Herrera 4115, paratypes CR 1596519, and CR 207025; A. Chacón 25, paratype CR 1596195.
References
Burger, W., & van der Werff, H. (1990). Flora Costaricensis. Family 80 Lauraceae. Fieldiana, Botany, n. s., 23, 1–129.
Gómez-Laurito, J., & Cascante, A. (1999). Licaria caribaea (Lauraceae): a new species from the Caribbean lowlands of Costa Rica. Novon, 9, 199–201. https://doi.org/10.2307/3391798
Gómez-Laurito, J., & Estrada, A. (2002). Licaria leonis (Lauraceae), una nueva especie del Pacífico costarricense, y algunas notas sobre Licaria multinervis H. Kurz. Lankesteriana, 3, 5–9. https://doi.org/10.15517/lank.v2i1.23136
Hammel, B. E. (1986). New species and notes on Lauraceae from the Caribbean lowlands of Costa Rica. Journal of
the Arnold Arboretum, 67, 123–136. https://doi.org/10.5962/bhl.part.27389
Kostermans, A. J. G. H. (1937). Revision of the Lauraceae II. The genera Endlicheria, Cryptocarya (American species) and Licaria. Recueil des Travaux Botaniques Néerlandais, 34, 500–609.
Kurz, H. W. (1983). Fortpflanzungsbiologie einiger Gattungen neotropischer Lauraceen und Revision der Gattung Licaria (Lauraceae) (Ph.D. Thesis). University of Hamburg, Germany.
Kurz, H. W. (2000). Revision der Gattung Licaria (Lauraceae). Mitteilungen aus dem Institut für Allgemeine Botanik in Hamburg, 28/29, 89–221.
Mez, C. (1889). Lauraceae americanae. Jahrbuch des Königlichen Botanischen Gartens und des Botanischen Museums zu Berlin, 5, 1–556.
Radford, A. E., Dickison, W. C., Massey, J. R., & Bell, C. R. (1974). Vascular plant systematics. New York: Harper & Row.
Van der Werff, H. (1988). Eight new species and one new combination of Neotropical Lauraceae. Annals of the Missouri Botanical Garden, 75, 402–419. https://doi.org/10.2307/2399431
Van der Werff, H. (2009). Nine new species of Licaria (Lauraceae) from tropical America. Harvard Papers in Botany, 14, 145–159. https://doi.org/10.3100/025.014.0206


