a Stanford University, Department of Biology, 327 Campus Drive, Stanford, California, 94305 USA
b Universidad Nacional Autónoma de México, Instituto de Biología, Tercer Circuito s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
c University of Helsinki, Faculty of Biological and Environmental Sciences, Ecosystems and Environment Research Programme, Niemenkatu 73, FI-15140, Lahti, Finland
Received: 19 October 2023; accepted: 1 February 2024
Abstract
Native habitat conversion to urban and agricultural areas represents conservation concerns for habitat quality and the breeding success of birds. In tropical areas facing regular deforestation of at-risk habitats, changes may occur to bird and nest predator communities that influence contradictory trends in breeding success. To assess the value of working lands for birds, we placed 100 artificial nests in 5 habitat types of varying human footprint, including a tropical dry forest reserve, a biological research station, croplands, and 2 urban towns. We report a clear decline in survival from the forest to urban towns. Habitat type explained the variation in nest survival probabilities over nest height, elevation, or time of nest exposure. Reducing the structural and compositional contrast of habitat and landscape vegetation between tropical dry forest and working lands represent valuable conservation actions for increasing habitat quality for birds.
No cuentes los huevos antes de que eclosionen: supervivencia diferencial de nidos artificiales de aves en un paisaje antropogénicamente modificado en el oeste de México
Resumen
La conversión de hábitats nativos en áreas urbanas y campos agrícolas representa problemas de conservación para la calidad del hábitat y el éxito reproductivo de las aves. La deforestación constante de hábitats en riesgo puede cambiar las comunidades de aves y los depredadores de los nidos, lo que puede influir en su éxito reproductivo. Para evaluar el valor de los hábitats dentro de un paisaje antropogénicamente modificado en el éxito reproductivo de las aves, colocamos 100 nidos artificiales en 5 hábitats con diferentes niveles de actividades humanas, incluyendo una reserva de bosque tropical caducifolio, una estación de investigación biológica, campos agrícolas y 2 pueblos urbanos. Encontramos una clara disminución en la supervivencia de nidos artificiales desde el bosque tropical caducifolio hasta los pueblos urbanos. El tipo de hábitat fue la variable que mejor explicó la variación en las probabilidades de supervivencia de los nidos artificiales en comparación con la altura del nido, la elevación y el tiempo de exposición del nido. Reducir el contraste dentro del paisaje en la estructura de la vegetación entre la reserva y los hábitats dentro del paisaje modificado representan acciones de conservación importantes para aumentar la calidad del hábitat para las aves.
Palabras clave: Campos agrícolas; Depredación de nidos de aves; Calidad de hábitat; Jalisco; Huevos de plastilina; Selva seca tropical; Urbanización
Introduction
Urbanization and the conversion of native habitat to agricultural land represent key factors in the long-term conservation of bird biodiversity (Aronson et al., 2014; Kehoe et al., 2017). In the tropics, urbanization and agriculture have led to the degradation and destruction of native vegetation and the reconfiguration of landscapes, causing stark contrast in habitat complexity between remnant vegetation and agricultural and urban areas that pose risks to bird biodiversity (Filloy et al., 2019; Fischer et al., 2015; Maas et al., 2016). Biodiverse tropical regions suffer some of the highest rates of urbanization and native habitat transformation (Estrada et al., 2020), which represent pressing challenges for the conservation of bird populations.
A key component of bird biodiversity and population monitoring in tropical landscapes with high rates of natural habitat transformation includes the evaluation of breeding ecology (DeGregorio et al., 2016). Reduced vegetative complexity and the exchange of native plants with non-native plants, both common attributes of agricultural and urban areas (Chace & Walsh, 2006), tend to negatively impact bird species with highly sensitive breeding requirements tied to native vegetation (Maas et al., 2016). Vegetation change in the tropics alters biotic —e.g., nest predation pressure and reduction of nest locations— and abiotic conditions —e.g., increased nest exposure, higher temperatures, and brighter conditions—, leading to potential direct and indirect influences on avian breeding ecology in species that depend on native plants and vegetation structure for nesting (Estrada et al., 2002; Rivera-López & MacGregor-Fors, 2016; Tellería & Díaz, 1995; Zuñiga-Palacios et al., 2021). Meanwhile, certain bird species may be positively impacted by or able to acclimate to novel conditions (DeGregorio et al., 2016; Kurucz et al., 2021; Latif et al., 2012), underlining the semi-permeable ecological filter that is applied to nesting birds in human-modified tropical landscapes and the importance of evaluating bird breeding ecology in different types of transformed land (MacGregor-Fors, 2010; MacGregor-Fors et al., 2022).
Nest predation represents a powerful force on bird breeding success and population dynamics (DeGregorio et al., 2016). A consequence of native habitat conversion to more urban or agricultural areas includes changes to nest predator communities, and the ability of bird species to adapt to these changes will ultimately determine whether disturbed areas offer viable habitat for native levels of biodiversity (DeGregorio et al., 2016; Latif et al., 2012). Urban and agricultural areas tend to have lower vegetation cover as forested habitats, leading to different natural predator abundance and nest visibility to predators, representing powerful determinants of breeding success for bird species that use transformed land (López-Flores et al., 2009; Martin, 1993; Zuñiga-Palacios et al., 2021). More disturbed areas may lead to a reduction of nest predation pressure due to the absence of native nest predators that have a low tolerance for human activity (Kurucz et al., 2021; Pretelli et al., 2023). A possible caveat to lower predation pressure from typical native predators in urban areas include birds that are habitat and foraging generalists (Estrada et al., 2002; Martin, 1995; Rivera-López & MacGregor-Fors, 2016), mammals that are attracted to anthropogenic food sources (Fischer et al., 2012), and increased access to nests by people that manipulate and destroy nests and eggs (López-Flores et al., 2009).
To assess the survivorship of bird nests, we placed artificial bird nests in 5 habitat types with increasing degrees of human disturbance and habitat modification, including 1) a tropical dry forest reserve (TDF hereafter), 2) the Chamela Biological Research Station (CBRS) grounds, embedded in the Chamela-Cuixmala Biosphere Reserve, 3) croplands (CL), 4) Careyes (CAR), a small and heavily built-up town, and 5) Emiliano-Zapata (ZAP), a larger town. While a recent meta-analysis questions the efficacy of artificial nest studies in determining nest survival probabilities in urban areas relative to natural nests (Vincze et al., 2017), the feasibility of finding sufficient numbers of natural nests in heavily built-up urban areas (i.e., outside of urban parks and green spaces) makes the use of artificial nests necessary. We controlled for important variables that may influence predation rates, such as nest size and height, to focus on habitat-level variations in nest survivorship and the impacts of urban and agricultural areas in the working landscape. Such landscapes are common in the tropical areas of Mexico (Levey et al., 2023), where existing reserves are surrounded by a working landscape with non-native vegetation that contrasts highly with native areas (Levey & MacGregor-Fors, 2021; Levey et al., 2021; MacGregor-Fors & Schondube, 2011; Vázquez-Reyes et al., 2017). Efforts to evaluate the impacts on breeding ecology in these working tropical landscapes are needed to supplement a thin body of work (Estrada et al., 2002; López-Flores et al., 2009; Zuñiga-Palacios et al., 2021) and determine the risks that urban and agricultural areas present for breeding birds. We expected nest survivorship to be lower in CL, CAR, and ZAP relative to the conserved TDF reserve and the CBRS due to higher exposure of nests and greater visibility for predators due to reduced vegetation complexity and density (Estrada et al., 2002; López-Flores et al., 2009; Zuñiga-Palacios et al., 2021).
Materials and methods
We conducted our study in a landscape between the Chamela-Cuixmala Biosphere Reserve (19°29’57.5” N, 105°02’41.6” W) and the town Emiliano Zapata (19°23’16.6” N, 104°57’50.1” W) in the Municipality La Huerta (population: 23,258; INEGI, 2020) on the Pacific coast of Jalisco, Mexico (Fig. 1). Historically, native vegetation cover in the region consisted primarily of tropical dry forest, which consists of deciduous forest with a mean canopy height of 12 m, a dense understory (Rzedowski, 2006), and strong phenological changes due to highly seasonal rainfall in the region (Durán et al., 2002). Other forest types exist in areas with more regular water availability, including semi-deciduous (mean canopy height of 20 m) and mangrove forests (Durán et al., 2002). After a period of increased human occupation and agricultural expansion from 1950-1970, large cover of tropical dry forest and other native forest types in lower elevation zones were converted to small towns and agricultural lands, linked by paved and unpaved roads, creating a landscape mosaic of native and non-native vegetation types (Maass et al., 2005).
In this landscape, we selected 5 habitat types with varying degrees of urban and agricultural disturbance for artificial nest placement: 1) TDF, with closed canopy cover and dense understory, 2) CBRS, which consists of moderately built-up 1.4 ha area embedded within the Chamela-Cuixmala Biosphere Reserve, 3) CL, consisting of fields of small, herbaceous plants such as maize (Zeamays), squash (Cucurbita spp.), chili pepper (Capsicum spp.), watermelon (Citrulluslanatus), and beans (Phaseolus spp.) located in the southern edge of the study area (Maass et al., 2005), 4) CAR (19°26’36.15” N, 105°1’49.65” W), a small town with heavy built-up cover, and 5) ZAP, a large town with less built-up cover than CAR (Fig. 1). Both elevations (MSL) of the TDF and CBRS sampling areas were slightly higher than the other habitat types. The TDF and CBRS sampling areas are also in closer proximity to each other than the other sampling locations. We included both habitat categories due to the higher human presence at the Biological Station, the noise generated by people and activities at the station, and a higher density of paved roads that could influence the occupancy of bird and mammals that respond positively to increasing human footprint (Rivera-López & MacGregor-Fors, 2016). Potential bird nest predators in the study region included White-throated Magpie-jays and San Blas jays (Calocitta formosa and Cyanocorax sanblasianus), Great-tailed grackles (Quiscalus mexicanus), mammals (e.g., Nasua narica, domestic dogs and cats, rodents, possums, and Procyon lotor), and diverse reptiles.
We used a mixture of plant fibers, twigs, and mud from the nest location to create bird nests in the shape of open plant fiber nest cups large enough to hold both the clay and quail eggs. We created open cup nests since many species in the study area construct nests in similar ways (e.g., Cyanocorax sanblasianus, Peucaea ruficauda, and Turdus rufopalliatus; Mendoza-Rodríguez et al., 2010) and due to the ease of creating such a nest shape. We placed each nest ~ 2 m above ground to control the tendency of nest height placement to affect predation rates (DeGregorio et al., 2016). We placed 1 Japanese Quail (Coturnixjaponica) commercial egg and 1 clay egg of similar size for a total of 2 eggs in each nest (Bayne et al., 1997; Estrada et al., 2002). We used clay since it is a malleable material that preserves markings from predation events and has negligible influence on predation rates (Bayne & Hobson, 1999; Bayne et al., 1997). We used commercial quail eggs due to their small size and color speckling that best mimicked natural terrestrial bird eggs relative to domestic chicken eggs and their availability in the study region. We used both a real and clay egg to provide stimulus for a wider range of predators than clay eggs alone and to capture predation event evidence if we could not perceive markings on the quail egg from smaller nest predators (Bayne et al., 1997; Estrada et al., 2002). We used rubber gloves to prevent leaving a human scent when handling nest materials and eggs (Estrada et al., 2002).
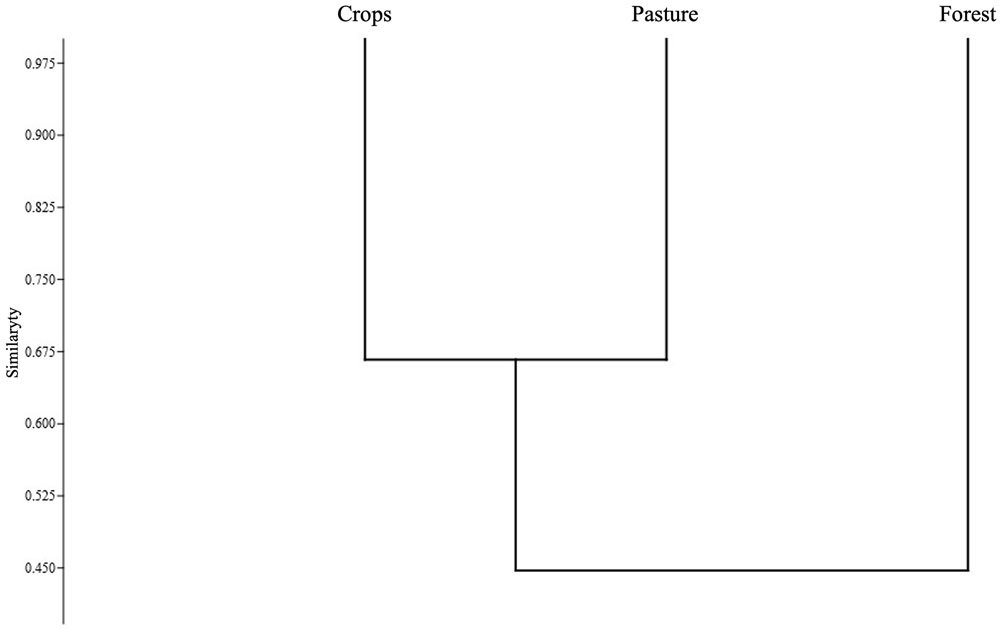
Figure 1. Region of study in the state of Jalisco in western Mexico. We placed artificial bird nests at the localities marked with a black dot and text, including ‘Forests’ (tropical dry forest of the Chamela-Cuixmala Biosphere Reserve), ‘Biology station’ (the Chamela Biological Research Station), ‘Cropfields’ (herbaceous crop plots), ‘Careyes’ (a small, heavily built-up town), and ‘Zapata’ (a large, less built-up town). Nest locations within the marked localities by at least 250 m to increase spatial independence.
We placed 20 artificial nests in each of the 5 habitat types for a total of 100 nests. Nests were exposed for a total of 12 days (April 30 – May 11, 2009), and we checked nests at 3-day intervals for a total of 4 nest visits. We considered nests as failed if the eggs were missing or if there were indications of a predation event on either the clay or quail egg, including scratches, bite marks, or perforations. We removed nests with signs of predation from the sample locations. We considered nests successful if there were no markings on either the clay or quail egg.
Figure 2. Survival probability with 95% confidence intervals from the Known Fate analysis in MARK of the artificial nests in the conserved tropical dry forest (TDF in the figure), Chamela Biological Research Station (CBRS), cropland (CL), the town of Careyes (CAR), and the town of Zapata (ZAP).
We used the program MARK (White & Burnham, 1999) to perform a known fate analysis using our nest check interval to calculate the probability of survivorship of each nest (Dinsmore & Dinsmore, 2007), using the covariables nest height (m), elevation (m asl), habitat type, and time of nest exposure to generate the models. We included the nest height variable in analyses despite controlling the height at 2-m to check for potential interactions with other covariates. We included elevation in our models to account for slight elevation differences between nest site locations and the tendency of lower elevation areas to have higher cover of agricultural and urban areas (Maass et al., 2005). We included habitat type to determine the differences between certain habitat types on artificial nest survival. Finally, we included the time of nest exposure since the likelihood of nest survival is tied to the amount of time eggs are exposed to predators (Dinsmore & Dinsmore, 2007). We ranked the models by parsimony using the adjusted Akaike’s Information Criterion for small sample sizes (AICc; Hurvich & Tsai, 1989). We selected the models that best fit our data by calculating the differences in AICc values (ΔAICc) and choosing those with ΔAICc values less than 2 units from the most parsimonious model (Burnham & Anderson, 2002).
Results
We recorded 82 preyed upon nests of 100 total, including 37 (45.1%) instances of bird predation, 30 (36.6%) instances of unknown predation, 10 (12.2%) instances of egg removal or manipulation by humans, 3 (3.7%) instances of rodent predation, and 2 (2.4%) instances of reptile predation. We recorded 18 nests with no predation signs, with the majority remaining in TDF (44.4%), followed by CBRS (27.8%), CL (22.2%), and ZAP (5.6%). No nests placed in CAR survived the observation period. Nest survival probability was 0.38 (95% CI: 0.26-0.48) in TDF, 0.27 (95% CI: 0.17-0.33) in CBRS, 0.25 (95% CI: 0.15-0.35) in CL, 0.06 (95% CI: 0.01-0.13) in ZAP, and 0.0 in CAR (Fig. 2). The most parsimonious model to explain the variation in nest survival probabilities included the lone covariable habitat, followed closely by the combination of habitat and height (Table 1).
Table 1
Model output from the Known Fate analysis in MARK. Covariates used in the models include habitat, nest height (controlled at 2 m above ground level), elevation, and time of nest exposure.
Model
AICc
ΔAICc
AICc weight
Model likelihood
Parameters
Deviance
Habitat
262.62
0.00
0.44
1.00
5
252.32
Habitat + height
263.96
1.35
0.22
0.51
6
251.55
Habitat + elevation
264.49
1.87
0.17
0.39
6
252.08
Habitat + height + elevation
265.90
3.29
0.09
0.19
7
251.35
Elevation
267.21
4.59
0.04
0.10
5
256.91
Height + elevation
268.74
6.12
0.02
0.05
6
256.32
Time of nest exposure
270.31
7.69
0.01
0.02
4
262.11
Height
272.26
9.64
0
0.01
5
261.96
Discussion
The impacts of bird nest predation along habitat disturbance gradients vary depending on the severity of habitat modification and the biotic and abiotic conditions of transformed land (Vincze et al., 2017). Novel biotic and abiotic conditions in urban and agricultural settings heavily contrast with native habitat, representing important influences on bird breeding success and, ultimately, biodiversity conservation (DeGregorio et al., 2016). We report a clear decline in the survival probabilities of artificial bird nests throughout a gradient of urban intensity between a conserved tropical dry forest and the largest town.
TDF, the most conserved habitat in the disturbance gradient, had the highest artificial nest survival probability among all studied habitats. This finding is consistent with other artificial nest studies from the tropical Americas that show greater vegetation cover offers increased survival odds by concealing nests more effectively from predators, both within forests with seasonal leaf cover (Vega-Rivera et al., 2009) and relative to more open areas (Estrada et al., 2002; López-Flores et al., 2009). TDF contains a dense understory of vegetation and a closed canopy with darker lighting, which may be a key factor in the detection of nests by predators (Estrada et al., 2002; Vázquez et al., 2021). While some studies have found that conserved areas either have similar or lower nest survival probabilities than in urban settings due to changes in predator abundance and composition (DeGregorio et al., 2016; Fischer et al., 2012; Zuñiga-Palacios et al., 2021), local factors in this heterogeneous landscape with various habitat types likely favor ample distribution of potential nest predators (e.g., urban birds, domesticated cats, and dogs) in urban areas (Estrada et al., 2002; López-Flores et al., 2009; Rivera-López & MacGregor-Fors, 2016).
Outside of the conserved TDF habitat, CL showed near-equal nest survival probabilities as the CBRS, which were lower than in TDF. Our results indicate that even small (< 2 ha), moderately built-up areas embedded in conserved habitat may increase the likelihood of nest predation to levels found in agricultural land. Synonymous with development is the opening of forest habitat, leading to new abiotic conditions and biotic stimulus that may influence breeding success in birds (Patten & Smith-Patten, 2012; Shochat et al., 2010). In our study area, CBRS has attracted several bird species that are opportunistic omnivores and often associated with open habitats, such as the Great-tailed Grackle (Quiscalusmexicanus; MacGregor-Fors et al., 2009). Also attracted to this habitat and CL are potential nest predators such as the White-nosed Coati (Nasuanarica) and Common Raccoon (Procyonlotor), which have been documented to predate bird nests (Estrada et al., 2002; Menezes & Marini, 2017; Robinson et al., 2005). Snakes, which occur at similar compositions inside and outside the reserve, may exhibit increased activity at edge habitats (Chalfoun et al., 2002; Suazo-Ortuño et al., 2008; Vetter et al., 2013). These changes to the nest predator communities in CBRS and CL could have important implications on bird breeding success (DeGregorio et al., 2016), and continued urbanization of these areas may continue to decrease nest survival probabilities to the levels of heavily built-up towns.
The built-up areas along the urbanization gradient in our study had significantly lower nest survival probabilities than the other studied habitats. Urbanization and loss of native vegetation have been shown to negatively influence the survival of bird nests in previous studies (Rivera-López & MacGregor-Fors, 2016; Thorington & Bowman, 2003), and a potential mechanism includes the introduction of novel predation pressures, such as domesticated cats (Patterson et al., 2016), dogs (Zuñiga-Palacios et al., 2021) and humans (López-Flores et al., 2009). While it has been shown that urban areas may increase nest survival and breeding success in birds (Fischer et al., 2012; Kurucz et al., 2021), the urban areas in our study area presented an overwhelming amount of novel predation pressures that are not present in the other studied habitats (Chace & Walsh, 2006; López-Flores et al., 2009), highlighting the importance of evaluating changes in the communities of bird nest predators along habitat disturbance gradients (DeGregorio et al., 2016). Conserving and restoring degraded areas within working landscapes and urban centers through measures such as live fencing, remnant forest preservation, and educational programs on bird breeding ecology may provide vital nesting habitat and increase bird breeding success (Bocz et al., 2017; Zuñiga-Palacios et al., 2021).
Acknowledgements
We thank the Estación de Biología Chamela (Instituto de Biología, UNAM) for granting permission to place artificial nests in the biosphere reserve. We thank Carlos Lara for editing and three anonymous reviewers for comments that enhanced the quality and clarity of the manuscript. We thank Michelle García-Arroyo for creating the study area map. DRL received a Master’s scholarship from Conahcyt (grant number 964233) as part of the Posgrado en Ciencias Biológicas of the Universidad Nacional Autónoma de México.
References
Aronson, M. F. J., La Sorte, F. A., Nilon, C. H., Katti, M., Goddard, M. A., Lepczyk, C. A. et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proceedings of the Royal Society B: Biological Sciences, 281, 20133330. https://doi.org/10.1098/rspb.2013.3330
Bayne, E. M., & Hobson, K. A. (1999). Do clay eggs attract predators to artificial nests? Journal of Field Ornithology, 70, 1–7.
Bayne, E. M., Hobson, K. A., & Fargey, P. (1997). Predation on artificial nests in relation to forest type: contrasting the use of quail and plasticine eggs. Ecography, 20, 233–239. https://doi.org/10.1111/j.1600-0587.1997.tb00366.x
Bocz, R., Szép, D., Witz, D., Ronczyk, L., Kurucz, K., & Purger, J. J. (2017). Human disturbances and predation on artificial ground nests across an urban gradient. AnimalBiodiversity Conservation, 40, 153–157. https://doi.org/10.32800/abc.2017.40.0153
Burnham, K. P., & Anderson, D. R. (2002). Model selection and multimodel inference. New York: Springer-Verlag.
Chalfoun, A. D., Thompson, F. R., & Ratnaswamy, M. J. (2002). Nest predators and fragmentation: a review and meta-analysis. Conservation Biology, 16, 306–318.
DeGregorio, B. A., Chiavacci, S. J., Benson, T. J., Sperry, J. H., & Weatherhead, P. J. (2016). Nest predators of North American birds: continental patterns and implications. Bioscience, 66, 655–665. https://doi.org/10.1093/biosci/biw071
Dinsmore, S. J., & Dinsmore, J. J., 2007. Modeling avian nest survival in program MARK. Studies in Avian Biology, 34, 73–83.
Durán, E., Balvanera, P., Lott, E., Segura, G., Pérez-Jiménez, A., Islas, Á. et al. (2002). Estructura, composición y dinámica de la vegetación. In F. A. Noguera, J. H. Vega-Rivera, A. N. García-Aldrete, & M. Quesada-Avendaño (Eds.), Historia natural de Chamela (pp. 443–472). México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México.
Estrada, A., Garber, P. A., & Chaudhary, A. (2020). Current and future trends in socio-economic, demographic and governance factors affecting global primate conservation. PeerJ, 8, e9816. https://doi.org/10.7717/peerj.9816
Estrada, A., Rivera, A., & Coates-Estrada, R. (2002). Predation of artificial nests in a fragmented landscape in the tropical region of Los Tuxtlas, Mexico. BiologicalConservation, 106, 199–209. https://doi.org/10.1016/S0006-3207(01)00246-4
Filloy, J., Zurita, G. A., & Bellocq, M. I. (2019). Bird diversity in urban ecosystems: the role of the biome and land use along urbanization gradients. Ecosystems, 22, 213–227. https://doi.org/10.1007/s10021-018-0264-y
Fischer, J. D., Cleeton, S. H., Lyons, T. P., & Miller, J. R. (2012). Urbanization and the predation paradox: the role of trophic dynamics in structuring vertebrate communities. Bioscience, 62, 809–818. https://doi.org/10.1525/bio.2012.62.9.6
Fischer, J. D., Schneider, S. C., Ahlers, A. A., & Miller, J. R. (2015). Categorizing wildlife responses to urbanization and conservation implications of terminology. ConservationBiology, 29, 1246–1248. https://doi.org/10.1111/cobi.12451
Hurvich, C. M., & Tsai, C. L. (1989). Regression and time series model selection in small samples. Biometrika, 76, 297–307. https://doi.org/10.1093/biomet/76.2.297
Kehoe, L., Romero-Muñoz, A., Polaina, E., Estes, L., Kreft, H., & Kuemmerle, T. (2017). Biodiversity at risk under future cropland expansion and intensification. Nature Ecology and Evolution, 1, 1129–1135. https://doi.org/10.1038/s41559-017-0234-3
Kurucz, K., Purger, J. J., & Batáry, P. (2021). Urbanization shapes bird communities and nest survival, but not their food quantity. Global Ecology and Evolution, 26, e01475. https://doi.org/10.1016/j.gecco.2021.e01475
Latif, Q. S., Heath, S. K., & Rotenberry, J. T. (2012). How avian nest site selection responds to predation risk: testing an ‘adaptive peak hypothesis’. Journal of Animal Ecology, 81, 127–138. https://doi.org/10.1111/j.1365-2656.2011.01895.x
Levey, D. R., Estrada, A., Enríquez, P. L., & Navarro-Sigüenza, A. G. (2021). The importance of forest-nonforest transition zones for avian conservation in a vegetation disturbance gradient in the northern Neotropics. Tropical Conservation Science, 14, 1–14. https://doi.org/10.1177/19400829211008087
Levey, D. R., & MacGregor-Fors, I. (2021). Neotropical bird communities in a human-modified landscape recently affected by two major hurricanes. Avian Conservation and Ecology, 16, art9. https://doi.org/10.5751/ACE-01920-160209
Levey, D. R., Patten, M. A., & Estrada, A. (2023). Bird species occupancy trends in southeast Mexico over 1900–2020: accounting for sighting record absences. Journal of Animal Ecology, 92, 606–618. https://doi.org/10.1111/1365-2656.13871
López-Flores, V., MacGregor-Fors, I., & Schondube, J. E. (2009). Artificial nest predation along a Neotropical urban gradient. Landscape and Urban Planning, 92, 90–95. https://doi.org/10.1016/j.landurbplan.2009.03.001
Maas, B., Karp, D. S., Bumrungsri, S., Darras, K., Gonthier, D., Huang, J. C. C. et al. (2016). Bird and bat predation services in tropical forests and agroforestry landscapes. BiologicalReviews, 91, 1081–1101. http://doi.wiley.com/10.1111/brv.12211
Maass, J. M., Balvanera, P., Castillo, A., Daily, G. C., Mooney, H. A., Ehrlich, P. et al. (2005). Ecosystem services of tropical dry forests: insights from long-term ecological and social research on the Pacific coast of Mexico. Ecology and Society, 10, art17. https://doi.org/10.5751/es-01219-100117
MacGregor-Fors, I. (2010). How to measure the urban-wildland ecotone: redefining ‘peri-urban’ areas. EcologicalResearch, 25, 883–887. https://doi.org/10.1007/s11284-010-0717-z
MacGregor-Fors, I., García-Arroyo, M., & Quesada, J. (2022). Keys to the city: an integrative conceptual framework on avian urban filtering. Journal of Urban Ecology, 8, juac026. https://doi.org/10.1093/jue/juac026
MacGregor-Fors, I., Vázquez, L., Vega-Rivera, J. H., & Schondube, J. E. (2009). Non-exotic invasion of Great-tailed Grackles (Quiscalus mexicanus) in a tropical dry forest reserve. Aredea, 97, 367–369. https://doi.org/10.5253/078.097.0312
Martin, T. E. (1993). Nest predation among vegetation layers and habitat types: revising the dogmas. AmericanNaturalist, 141, 897–913. https://doi.org/10.1086/285515
Martin, T. E. (1995). Avian life history evolution in relation to nest sites, nest predation, and food. EcologicalMonographs, 65, 101–127. https://doi.org/10.2307/2937160
Mendoza-Rodríguez, V., Vega-Rivera, J. H., Medina-Montaño, I., & Campos-Cerda, F. (2010). Response of birds in tropical deciduous forest to Brown-headed Cowbirds (Molothrus ater). The Southwestern Naturalist, 55, 390–393. https://doi.org/10.1894/MH-46.1
Menezes, J. C. T., & Marini, M. Â. (2017). Predators of bird nests in the Neotropics: a review. Journal of Field Ornithology, 88, 99–114. https://doi.org/10.1111/jofo.12203
Patterson, L., Kalle, R., & Downs, C. (2016). Predation of artificial bird nests in suburban gardens of KwaZulu-Natal, South Africa. UrbanEcosystems, 19, 615–630. http://dx.doi.org/10.1007/s11252-016-0526-4
Pretelli, M. G., Cavalli, M., Chiaradia, N. M., Cardoni, A., & Isacch, J. P. (2023). Location matters: survival of artificial nests is higher in small grassland patches and near the patch edge. Ibis, 165, 111–124. https://doi.org/10.1111/ibi.13128
Rivera-López, A., & MacGregor-Fors, I. (2016). Urban predation: a case study assessing artificial nest survival in a Neotro- pical city. UrbanEcosystems, 19, 649–655. http://dx.doi.org/10.1007/s11252-015-0523-z
Robinson, W. D., Styrsky, J. N., & Brawn, J. D. (2005). Are artificial bird nests effective surrogates for estimating predation on real bird nests? A test with tropical birds. Auk, 122, 843–852. https://doi.org/10.1093/auk/122.3.843
Suazo-Ortuño, I., Alvarado-Díaz, M., & Martínez-Ramos, M. (2008). Effects of conversion of dry tropical forest to agricultural mosaic on herpetofaunal assemblages. Conservation Biology, 22, 362-374. https://doi.org/10.1111/ j.1523-1739.2008.00883.x
Shochat, E., Lerman, S. B., Anderies, J. M., Warren, P. S., Faeth, S. H., & Nilon, C. H. (2010). Invasion, competition, and biodiversity loss in urban ecosystems. Bioscience, 60, 199–208. https://doi.org/10.1525/bio.2010.60.3.6
Tellería, J. L., & Díaz, M. (1995). Avian nest predation in a large natural gap of the Amazonian rainforest. Journal of Field Ornithology, 66, 343–351.
Thorington, K. K., & Bowman, R. (2003). Predation rate on artificial nests increases with human housing density in suburban habitats. Ecography, 26, 188–196. https://doi.org/10.1034/j.1600-0587.2003.03351.x
Vázquez-Reyes, L. D., Arizmendi, M. C., Godínez-Álvarez, O. H., & Navarro-Sigüenza, A. G. (2017). Directional effects of biotic homogenization of bird communities in Mexican seasonal forests. Ornithological Applications, 119, 275–288. https://doi.org/10.1650/CONDOR-16-116.1
Vazquez, M. S., Zamora-Nasca, L. B., Rodríguez-Cabal, M. A., & Amico, G. C., 2021. Interactive effects of habitat attributes and predator identity explain avian nest predation patterns. Emu – Austral Ornithology, 121, 250–260. https://doi.org/10.1080/01584197.2021.1928519
Vega-Rivera, J. H., Medina-Montaño, I., Rappole, J. & Campos-Cerda, F. (2009). Testing the importance of nest concealment: does timing matter? Journal of Field Ornithology, 80, 303–307. https://doi.org/10.1111/j.1557-9263.2009.00234.x
Vetter, D., Rücker, G., & Storch, I. (2013). A meta-analysis of tropical forest edge effects on bird nest predation risk: edge effects in avian nest predation. Biological Conservation, 159, 382–395. https://doi.org/10.1016/j.biocon.2012.12.023
Vincze, E., Seress, G., Lagisz, M., Nakagawa, S., Dingemanse, N. J., & Sprau, P. (2017). Does urbanization affect predation of bird nests? a meta-analysis. Frontiers in Ecology and Evolution, 5, 29. https://doi.org/10.3389/fevo.2017.00029
White, G. C., & Burnham, K. P. (1999). Program MARK: survival estimation from populations of marked animals. Bird Study, 46 (Suppl.), S120–S139. https://doi.org/10.1080/00063659909477239
Zuñiga-Palacios, J., Corcuera, P., & Almazán-Núñez, R. C. (2021). Living fences decrease the edge effect on nest predation in a tropical dry forest landscape: evidence from an experiment using artificial nests. Agroforestry Systems, 95, 547–558. https://doi.org/10.1007/s10457-021-00603-z
Mayra R. Cortez-Roldán a, Alejandro Valdez-Mondragón b, *
a Universidad Autónoma de Tlaxcala, Centro Tlaxcala de Biología de la Conducta, Posgrado en Ciencias Biológicas, Carretera Federal Tlaxcala-Puebla, Km. 1.5, 90062 Tlaxcala, Tlaxcala, Mexico
b Centro de Investigaciones Biológicas del Noroeste S.C., Programa Académico de Planeación Ambiental y Conservación, Colección de Aracnología, Km. 1 Carretera a San Juan de La Costa “El Comitán”, 23205 La Paz, Baja California Sur, Mexico
Received: 11 October 2023; accepted: 19 March 2024
Abstract
With 40 of the 149 described species, Mexico harbors the highest diversity of the spider genus Loxosceles. However, knowledge about these spiders’ distribution patterns in a climate change (CC) context is poorly known. In this study, the distributions of 4 species from Central Mexico, Loxosceles malintzi, L. misteca, L. tenochtitlan and L. zapoteca, were estimated and evaluated based on species distribution modeling (SDM) and the possible effects of CC. Two future scenarios were simulated (years 2050 and 2080) to show possible increases or reductions in species distributions. The most important variables that influence the distribution of the species were: isothermality, seasonality of temperature, and precipitation. In the CC scenarios, some species showed a possible increase, specifically, Loxosceles malintzi with an increase in its distribution of 79% by 2050 and 66% by 2080, whereas L. misteca was projected to increase its distribution by 28% for 2050 and 38% for 2080. However, a decrease in the distribution of L. tenochtitlan by 51% for 2050 and 38% for 2080 was projected, as well as a 45% decrease by 2050 and a 40% decrease by 2080 for L. zapoteca.
Efectos del cambio climático y modelado de distribución de especies de Loxosceles (Araneae: Sicariidae) del centro de México
Resumen
Con 40 de 149 especies descritas, México alberga la mayor diversidad de arañas del género Loxosceles. Sin embargo, el conocimiento sobre los patrones de distribución en un contexto de cambio climático (CC) es poco conocido. Se estimaron y evaluaron las distribuciones de 4 especies del centro de México, Loxosceles malintzi, L. misteca, L. tenochtitlan y L. zapoteca, con base en el modelado de distribución de especies (MDE) y posibles efectos del CC. Se simularon 2 escenarios futuros (años 2050 y 2080) para mostrar posibles incrementos o reducciones en las distribuciones de las especies. Las variables más importantes que influyen en la distribución son: la isotérmia, la estacionalidad de la temperatura y las precipitaciones. En los escenarios de CC algunas especies mostraron un posible aumento, Loxosceles malintzi tuvo un aumento proyectado en su distribución de 79% para 2050 y de 66% para 2080, mientras que L. misteca tuvo un aumento de 28% para 2050 y de 38% para 2080. Sin embargo, se proyectó una disminución en la distribución de L. tenochtitlan de 51% para 2050 y de 38% para 2080, así como una disminución de 45% para 2050 y de 40% para 2080 para L. zapoteca.
In North America, the spider genus Loxosceles Heineken and Lowe, 1832 (family Sicariidae Keyserling, 1980) are commonly known as “violin spiders”, “recluse spiders”, or “brown recluse spiders”. The current diversity comprises 149 described species, being one of the spider genera responsible for serious medical problems worldwide due to their venomous bites (Manríquez & Silva, 2009; Ramos-Rodríguez & Méndez, 2008; Sandidge & Hopwood, 2005; Swanson & Vetter, 2009; Vetter, 2008, 2015; Vetter & Bush, 2002; Vetter et al., 2003; WSC, 2024). Recent reviews of the medical aspects regarding Loxosceles bites have revealed that, despite frequent and numerous publications, only 11% of reported bites were verified, and skin necrosis developed in three-quarters of these cases (Taucare-Ríos et al., 2018).
Mexico boasts the highest diversity of violin spiders, with 40 species officially recognized, 38 native species and 2 introduced: Loxosceles reclusa Gertsch & Mulaik, 1940 and Loxosceles rufescens (Dufour, 1820) (Gertsch 1958, 1973; Gertsch & Ennik 1983; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, Solís-Catalán et al., 2018; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018). Some North American synanthropic species, such as L. reclusa from the southern USA, have been widely studied for their biological, medical, and physiological aspects, with their abundances, natural history, and even distributions being analyzed and modeled (Sandidge & Hopwood, 2005; Saupe et al., 2019; Swanson & Vetter, 2009; Vetter, 2005, 2008; Vetter & Barger, 2002; Vetter & Bush, 2002; Vetter et al., 2003). However, these aspects remain poorly studied or not assessed in Mexican species. To date, all 38 states of Mexico have records of at least 1 native or introduced species of Loxosceles. The highest diversity of Loxosceles species in Mexico is mainly concentrated in the northern regions of the country, characterized by warm, dry regions such as deserts and xerophytic shrublands, with the Baja California Peninsula being the most diverse region in the country. The group’s diversity tends to decrease in the southern regions of the country where the habitat is more tropical and sub-tropical, such as in the Yucatán Peninsula which has the lowest species diversity in the country (Navarro-Rodríguez & Valdez-Mondragón, 2020; Valdez-Mondragón, 2020; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, Solís-Catalán et al., 2018; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018; Valdez-Mondragón et al., 2019).
In recent years, climate change (CC) has been accelerated by anthropogenic processes mainly related to the intensification of greenhouse gases (Compagnucci, 2011). The resulting effects on ecosystem processes have led to changes in the distributions of species worldwide, which can be observed in 3 main response patterns: 1) species maintain their areas of distribution (Stranges et al., 2019; Uribe-Botero. 2015); 2) species’ areas of distribution will decrease, and in extreme cases, species might become extinct (Araújo et al., 2006; Romo et al., 2013); and 3) species will expand their geographical distributions (Boorgula et al., 2020; Martínez et al., 2024; Peterson, 2009), which has been even proposed with Loxosceles spiders from North America by Saupe et al. (2011) and also for cosmopolite species such as L. rufescens by Taucare-Ríos et al. (2018). Ecological niche models (ENM) and species distribution models (SDM) have been used to observe and assess the effects of CC (past and future) on species (Costa et al., 2002; Mota-Vargas et al., 2019; Peterson, 2009; Yáñez-Arenas et al., 2016). These methodologies have also been used to assess the potential risks associated with distribution expansions under future climate scenarios of disease-transmitting species (vectors), or those that can cause envenomation/poisoning in humans such as venomous snakes, scorpions or even violin spiders (Brooker et al., 2002; Foley et al., 2008; Lira et al., 2020; Martínez et al., 2024; Saupe et al., 2011; Taucare-Ríos et al., 2018; Yáñez-Arenas et al., 2016).
Climate change is expected to have profound effects on the distribution of venomous species worldwide, including reductions in the biodiversity and changes in patterns of envenomation of humans and even domestic animals (Martínez et al., 2024). Recent studies have projected future changes in species distributions as a consequence of CC, resulting in an increase or decrease of environmentally suitable areas. Vector-associated diseases such as leishmaniasis in Brazil (Lutzomyia, Peterson & Shaw [2003]), malaria in Africa (Anopheles, Peterson [2009]), Chagas in the Americas (Triatoma, Garza et al. [2014]), and Lyme disease and encephalitis in Europe (ticks of the genera Ixodes, Dermacentor, and Rhipicephalus, Gray et al. [2009]) have all been studied under the context of potential CC impacts of their host species’ distributional changes. In the case of venomous animals, ENM research has addressed evolutionary processes such as ecological niche conservatism/divergence between phylogenetically related species. Warren et al. (2008) and Taucare-Ríos (2017) applied such methods to Loxosceles spiders, while Planas et al. (2014) and Taucare-Ríos et al. (2018) used ENM to estimate diversity and observe the evolutionary processes that favored the diversification of L.rufescens in the Mediterranean, as well as the invasion potential of ecological niches globally. The only Loxosceles species from North America to be studied using ENM is L. reclusa, which causes the highest number of necrotic wounds induced by spiders in the USA. Saupe et al. (2011) investigated this species’ present and future distribution patterns and the medical implications within the Unites States.
In Mexico, studies focused on characterizing the ecological niches of Loxosceles species are scarce and often uninformative, especially with regards to species’ distribution areas. In the state of Guerrero, the genus’ distribution covers the entire state; however, no precise information exists on the environmental variables that influence distributions at the species level (Dzul-Manzanilla et al., 2014). The geographical distribution of the genus within Mexico has been shown to be influenced by some bioclimatic variables such as vegetation type, seasonality of temperature, annual temperature range, and isothermality (Cortez-Roldán, 2018; Valdez-Mondragón et al., 2019). At the genus level, SDMs show that Loxosceles is distributed across all biogeographic provinces except for the Sierra Madre Occidental, Soconusco, and Oaxaca (Cortez-Roldán, 2018). However, the broad scale of these analyses may lead to inaccurate or misinterpreted generalizations when assessing patterns and processes at the species level.
In recent years, scientific research on the genus Loxosceles from Mexico has been carried out mainly under an integrative taxonomy approach to describe and delimit species (Navarro-Rodríguez, 2019; Solís-Catalán, 2020; Valdez-Mondragón et al., 2019). However, some aspects such as the effect of CC on the distribution, biogeography, and diversity of these spiders remains unknown. The objective of this work is to estimate the current species distributions and future potential distributions of 4 Loxosceles species from the Central region of Mexico, considering 2 future climate change scenarios to identify possible changes in future distributions.
Materials and methods
Georeferenced localities were obtained from the Colección Nacional de Arácnidos (CNAN) (https://www.ibdata.abaco3.org/web/; https://datosabiertos.unam.mx/), Institute of Biology, Universidad Nacional Autónoma de México (IBUNAM), Mexico City, and Laboratorio de Aracnología (CARCIB), Centro de Investigaciones Biológicas del Noroeste (CIBNOR), La Paz, Baja California Sur, Mexico. To select the species used in the analyses, the following 2 criteria were considered: 1) species inhabiting Central Mexico that have previously been delimited and identified under morphological and molecular evidence (Navarro-Rodríguez, 2019; Navarro-Rodríguez & Valdez-Mondragón, 2020; Solís-Catalán, 2020; Valdez-Mondragón et al., 2019), which represents species with distributions covering the states with the highest population density (INEGI, 2020), as well as the states with the highest number of confirmed cases of loxoscelism from Mexico (Dr. Héctor Cartas, pers. comm.); and 2) adult specimens accurately identified to species level, the species identification using only juvenile specimens is not possible because the sexual primary structures such as male palps and female genitalia in adults are necessary for species identification. For identification of specimens collected and deposited at the CARCIB, dissections of male palps and female seminal receptacles were carried out in 80% ethanol, with tissue around the seminal receptacles cleaned using potassium hydroxide (KOH-10%) for 1 minute. Specimens were identified using the taxonomic keys for North American species by Gertsch and Ennik (1983) and literature of described species from Mexico by Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán (2018), Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán (2018), Valdez-Mondragón et al. (2019). A total of 55 records were used for the 4 analyzed species (Table 1): Loxosceles tenochtitlan Valdez-Mondragón & Navarro-Rodríguez, 2019 (20 records); Loxosceles malintzi Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez & Solís-Catalán, 2018 (19 records); Loxosceles misteca Gertsch, 1958 (8 records), and Loxosceles zapoteca Gertsch, 1958 (8 records). Field trips were carried out from 2016 to 2020 in Mexico City, Guerrero, Estado de México, Morelos, Puebla, and Tlaxcala to collect additional biological material.
The accessibility area (M) sensu Soberón (2010) of the 4 species was delimited based on all records of each species using the biogeographic provinces proposed by Morrone et al. (2017): the Trans-Mexican Volcanic Belt Province (TVB) and the Balsas Basin Province (BBP). The accessibility area M corresponds to the region that has been accessible to the species for relevant periods (Barve et al., 2011; Mota-Vargas et al., 2019; Ortega-Andrade et al., 2015).
Table 1
Species and records of Loxosceles distributed in the Central region of Mexico and used in this study.
We downloaded a set of bioclimatic variables from Cuervo-Robayo et al. (2020) (http://geoportal.conabio.gob.mx/metadatos/doc/html/b19802009gw.html) (Table 2) with a time interval of 1980-2009 and a resolution of 30 seconds (~ 1 km2) to avoid temporality errors with our occurrence data (data between 2010 and 2020) (Mota-Vargas et al., 2019). Four variables that presented spatial errors were excluded from the 19 bioclimatic variables following Escobar et al. (2014): mean temperature of the wettest quarter (BIO8), mean temperature of the driest quarter (BIO9), precipitation of the warmest quarter (BIO18), and precipitation of the coldest quarter (BIO19). Consequently, we used a total of 15 bioclimatic variables (Table 1). The criteria to select the variables was implemented by statistical methods and previous knowledge regarding the species’ ecology and biology. Pearson’s correlation analysis was performed for each species, and bioclimatic variables that showed a correlation r > 0.8 were discarded to avoid an overfit of the models and to reduce the performance of extrapolation (Steen et al., 2017). Variable subsets were created for each species using the R studio v.1.2.5042 “Kuenm” package (Cobos et al., 2019; Marques et al., 2020). For the transfer of the models, 2 CC scenarios were considered: representative concentration pathway (RCP) 4.5 optimal scenario (years 2050 and 2080) and 8.5 catastrophic scenario (2050, 2080) (S1). The RCP provides lower (RCP 4.5) and upper data for future potential greenhouse gas emissions (RCP 8.5). These scenarios were selected because they have been used previously to assess impact, vulnerability, and adaptation in previous studies on Mexican and Central American taxa (Fernández et al., 2016). The global circulation model from which the scenarios were obtained is the MIROC-5, which has been used in several arthropods groups and performs well in projections to future climate change scenarios (Liu & Shi, 2020; Mammola et al., 2018; Peterson et al., 2017; Romero et al., 2018; Roslin et al., 2017). Future climate data was downloaded in a resolution of 30 seconds (~ 1 km2) from CC, Agriculture, and Food Security (CCAFS) (https://ccafs-climate.org/).
Table 2
Environmental variables used in the ecological niche models (Cuervo-Robayo et al., 2020). Bold font indicates those variables used to run the models.
Variable
Description of variables
BIO1
Annual mean temperature
BIO2
Mean diurnal range
BIO3
Isothermally
BIO4
Temperature seasonality
BIO5
Max temperature of warmest month
BIO6
Min temperature of coldest month
BIO7
Temperature annual range
BIO10
Mean temperature of warmest quarter
BIO11
Mean temperature of coldest quarter
BIO12
Annual precipitation
BIO13
Precipitation of wettest month
BIO14
Precipitation of driest month
BIO15
Precipitation seasonality
BIO16
Precipitation of wettest quarter
BIO17
Precipitation of driest quarter
We used the Maxent v.3.3.3 (Maximum Entropy Algorithm) (Phillips et al., 2004) program to the modeling. Maxent predicts probability values (thresholds) of habitat suitability in the study area on a scale from 0 (least suitable) to 1 (most suitable). The parameters for each model were explored in the “Kuenm” package on R studio (Marques et al., 2020). Significance, performance (omission rate), and model complexity were used to choose the optimal parameter settings among the candidate models (Marques et al., 2020). All possible combinations of linear (l), quadratic (q), product (p), threshold (t) and hinge (h) feature types were tested, as well as the regularization multiplier (0.1, 0.3, 0.5, 0.7, 1.0, 5.0, and 10.0) (Marques et al., 2020). For the regularization multipliers a combination of 7 values was used, these influence the level of focus or how closely the obtained output distribution fits. Values < 1.0 produce distributions that are more adjusted to the presence records and, on the other hand, using values > 1.0 produce a more widespread and less localized prediction. Using combinations of the regularization multiplier allows obtaining from simple to more complex models (Phillips et al., 2006). Models were built using all possible combinations between the 16 and 42 subsets of environmental variables.
We evaluated significance (partial ROC) (Peterson et al., 2008), performance (omission error) of E = 5%, and model complexity under Akaike information criterion (AICc) for each candidate model to select the optimal parameter (Warren & Seirfet, 2011). With the optimal models, 10 replications were performed by Bootstrapping with logistic outputs. Logistic models show the probability that the species is present, conditional on environmental variables (Phillips & Dudik, 2008). In cases where there was more than 1 candidate model, the median of all replications was used to obtain the final model and a consensus map was created from the medians.
The models generated by Maxent are expressed in probabilistic values of environmental suitability. Therefore, consensus maps were reclassified in QGIS v.3.8 Zanzibar to create binary maps of potential distributions (0 absence-1 presence) (Costa et al., 2002; Jiménez-García & Peterson, 2019; Waltari & Guralnick, 2009). For the binary maps, we used the “minimum training presence threshold,” a threshold for which at least 1 known presence per species was found. Finally, we calculated the percentage of change in suitable habitat comparing present and future projections in QGIS.
The multivariate environmental similarity surface (MESS) was calculated to quantify the degree of extrapolation for determining the novelty of future climatic conditions relative to current conditions in the calibration area. MESS analyses help predict areas where strict extrapolation may occur (e.g., transfer areas with environmental conditions outside the range of training data). Higher values reflect greater uncertainty, and so caution must be taken when interpreting such areas, as they could be considered an overprediction of the models (Barbosa et al., 2009; Elith et al., 2010) (S2). The analysis was evaluated in the Ntbox package in R Studio (Osorio-Olvera et al., 2020).
Results
The SDM with the best statistical evaluations were obtained for each species (Figs. 1-4). A total of 1,360 models (L. malintzi) and 3,570 models (L. misteca, L. tenochtitlan, and L. zapoteca) were created using different configurations of regularization multiplier and response type, with 16 and 42 environmental data subsets (Table 3). The SDM for the 4 species comprise distributions within the following biogeographic provinces: Trans-Mexican Volcanic Belt (TVB), Balsas Basin (BBP), Chihuahuan Desert (CDP), and Sierra Madre del Sur (SMS) (Figs. 1-4).
Loxosceles malintzi has a potential distribution covering the BBP and SMS biogeographic provinces. These provinces span the states of Morelos, Puebla, Guerrero, Oaxaca, Estado de México, and Michoacán (Fig. 1). The environmental variables that most influenced the models were mean temperature of warmest quarter (BIO10) and precipitation of wettest quarter (BIO16), with 60.2% and 21.1%, respectively (Table 4). The potential distribution of L. misteca is recovered within the biogeographic provinces of BBP, SMS and TVB, including Michoacán, Morelos, Guerrero, and Puebla (Fig. 2). Precipitation of the wettest month (BIO13) had a contribution of 37.2% in the creation of the models (Table 4). Loxoscelestenochtitlan has a potential distribution that includes the provinces of CDP and TVB, including Estado de México, Mexico City, Puebla, Hidalgo, Querétaro, and Guanajuato (Fig. 3). The variable with the greatest contribution to the model was the precipitation of driest month (BIO14) with a 41.8% contribution (Table 4). Finally, L. zapoteca has a potential distribution within the BBP, SMS, and TVB provinces in Guerrero, Morelos, Puebla, Estado de México, and Michoacán (Fig. 4). Similar to L. tenochtitlan, the precipitation of the driest month (BIO14) had the highest contribution to the model at 46.8% (Table 4).
Figures 1-2. Species distribution modeling of Loxosceles species from the Central Mexico region under the Maxent algorithm. 1, Loxosceles malintzi, total distribution: 43,825.508 km2; 2, Loxoscelesmisteca, total distribution: 71,997.320 km2. Biogeographic provinces by Morrone et al. (2017) (black lines).
In transferring the SDM towards the 2 future scenarios, no significant changes in distributions were observed between the RCP 4.5 (2050 and 2080) and RCP 8.5 (2050 and 2080) scenarios in all 4 species (Figs. 5-12, S1). According to the results obtained for the 4 species analyzed in both scenarios, the RCP 4.5 (2050) scenario showed the lowest future environmental conditions, while the RCP 8.5 (2080) scenario had the highest (Figs. 5-12).
Figures 3-4. Species distribution modeling of Loxosceles species from the Central Mexico region under the Maxent algorithm. 3, Loxosceles tenochtitlan, total distribution: 28,836.642 km2; 4, Loxosceles zapoteca, total distribution: 57,911.413 km2. Biogeographic provinces by Morrone et al. (2017) (black lines).
Future distributions of L. malintzi in both scenarios (2050 and 2080) are projected to fall mainly within the BBP province (Figs. 5, 6). By 2050, an increase in the appropriate environmental conditions is projected to the southwest of the BBP including the northern SMS province (Fig. 5), whereas by 2080 a decrease in the species’ distribution is modelled with respect to the current environmental conditions (Fig. 6).
Figures 5-6. Potential distribution of Loxosceles malintzi under future scenarios. 5, Year 2050 (RCP 4.5); 6, year 2080 (RCP 8.5). Biogeographic provinces (Morrone et al., 2017). The plots show the percentage of decrease, increase, and persistence of the current distribution under future climate scenarios.
In both scenarios, a possible decrease between 15-25%, and an increase between 65-80% is observed in the potential distribution, which lies mainly inside the BBP province. Loxosceles malintzi also is projected to persist in environmentally suitable ranges of between 5-10% for both scenarios. The SDM of L. misteca for 2050 shows an increase of 28% towards the northwestern region of the Pacific Lowlands Province (PLP), a zone that is more susceptible to climate change (Fig. 7). In 2080, the projected distribution covers the same areas as the 2050 scenario, while showing a possible tendency to increase 38% towards the southeast of the distribution in the PLP (Fig. 8). The model for L. misteca shows a possible decrease of 13-15% in areas of optimal future environmental conditions, so this species may not have its distribution affected in either climate change scenario, as it shows a potential persistence of 45-47% of its current distribution (Figs. 7, 8).
Figures 7-8. Potential distribution of Loxosceles misteca under future scenarios. 7, Year 2050 (RCP 4.5); 8, year 2080 (RCP 8.5). Biogeographic provinces by Morrone et al. (2017). The plots the percentage of decrease, increase, and persistence of the current distribution under the climate scenarios.
The potential future distributions of L. tenochtitlan under the 2050 and 2080 scenarios cover a greater proportion of areas within the TVB, with changes in distribution appearing to the southwest of the CDP (Figs. 9, 10). These changes under future conditions show similarities to the current conditions, such that the 2 climate change scenarios show considerable areas where adequate environmental conditions could persist for the distribution of the species (20-30%). For L. tenochtitlan, the SDM shows a possible increase of 25-35% in its area of distribution under future projections; however, there is also a possibility of a decrease in the future distribution with respect to the current distribution between 35-50% (Figs. 9, 10). Therefore, this species could restrict its distribution to environmentally stable areas under both climate change scenarios (Figs. 9, 10). For L. zapoteca, both climate change scenarios, 2050 and 2080, suggest that environmental conditions present in the current SDM could be maintained, resulting in a projected increase of 5-10% in its distribution under future environmental conditions (Figs. 11, 12). However, it could also present a decrease between 40-45% in its distribution, so this species could be restricted to environmentally stable areas representing between 45-50% of its current distribution under both climate change scenarios (Figs. 11, 12).
Discussion
Species Distribution Modeling (SDM) has been a powerful approach for understanding how abiotic factors such as temperature, precipitation, and seasonality impact the biogeographic limits of different lineages or species (Graham et al., 2004; Wiens & Graham, 2005). The SDM in this study showed considerable utility in recognizing potential regions where the 4 analyzed species of Loxosceles may be distributed presently and under future scenarios considering different CC scenarios.
The potential distributions of the 4 focal Loxosceles species are mainly affected by the temperature seasonality, isothermality, precipitation seasonality, and precipitation of the wettest quarter. In previous works on Loxosceles spiders, these 4 variables have been shown to influence the spiders’ life cycle, distribution, and survival, increasing populations and food availability throughout the seasons (Canals et al., 2016; Cruz-Cárdenas et al., 2014; Ferreti et al., 2018; Planas et al., 2014; Saupe et al., 2011).
Table 3
Performance metrics of the configuration of the best models for each of the 4 Loxosceles species of the Central Mexican region.
These environmental variables have similarly been important factors in the distribution of other arachnid species, such as harvestmen and mygalomorph spiders (Ferreti et al., 2018; Simó et al., 2014). However, the use of few environmental variables for an ecological niche characterization could generate a slightly restrictive environmental space, causing an over-prediction of potential distribution areas. Therefore, the biology of the species must be considered when choosing environmental variables, including their dispersal capacity, as was proposed by Mota-Vargas et al. (2019). The possible changes of the 4 variables mentioned above under future CC scenarios for the Loxosceles species analyzed, might impact the seasonal life cycle, their dispersal capacity, reproductive seasonality and even the food availability when the distribution would increase (or not), having more ecological competition even with other spiders’ groups.
Table 4
Contribution of the environmental variables used in Maxent for the current distribution of Loxosceles species in the Central Mexican region.
Variable
Contribution (%)
Loxosceles malintzi Mean temperature of warmest quarter (BIO10) Precipitation of wettest quarter (BIO16) Temperature annual range (BIO7) Isothermality (BIO3)
60.2 21.1 12.5 6.2
Loxosceles misteca Precipitation of wettest month (BIO13) Precipitation of driest month (BIO14) Min temperature of coldest month (BIO6) Isothermality (BIO3) Precipitation Seasonality (BIO15)
37.2 21.5 19.6 12.2 9.5
Loxosceles tenochtitlan Precipitation of driest month (BIO14) Precipitation of wettest quarter (BIO16) Precipitation Seasonality (BIO15) Min temperature of coldest month (BIO6) Max temperature of warmest month (BIO5)
41.8 28.1 14.2 10.0 5.9
Loxosceles zapoteca Precipitation of driest month (BIO14) Precipitation Seasonality (BIO15) Precipitation of wettest quarter (BIO16) Temperature Seasonality (BIO4)
46.8 24.2 22.6 6.4
Of the analyzed species, L. malintzi is distributed in the warmest region in the Balsas Basin Province (BBP). This species has previously been recorded from this region by Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, and Solís-Catalán (2018) and Cortez-Roldán (2018). The climate of this area is characterized as semi-warm and sub-humid, correlated with the typical vegetation type of tropical deciduous forests (INEGI, 2017; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018; Valdez-Mondragón et al., 2019). The current distribution of L. malintzi seems to be restricted mainly to sites with high temperatures (22-32 °C), and a stable zone within the BBP is observed under both scenarios for this species (Fig. 1). Loxosceles misteca has a distribution that covers the western region of the BBP, with most records being from karstic caves located in dry and tropical forests, including deciduous forests (Navarro-Rodríguez, 2019; Valdez-Mondragón et al., 2019). Variations in surface temperature and humidity directly influence the availability of food inside such caves, making these variables important factors in the distribution of the species (Cruz-Cárdenas et al., 2014; Mammola et al., 2018). Loxosceles zapoteca has previously been reported in deciduous forests and xeric scrublands within the BBP by Cortez-Roldán (2018), as has L. malintzi and L. misteca. Although these 3 species occupy the same area of distribution, their ecological niches are different, providing specific information to characterize and generate their SDMs that perform better for each species. The SDMs proposed by Cortez-Roldán (2018) for L. malintzi, L. misteca, and L. zapoteca show distributions in the BBP, which is corroborated with the SDMs generated herein.
Figures 9-10. Potential distribution of Loxosceles tenochtitlan under future scenarios. 9, Year 2050 (RCP 4.5); 10, year 2080 (RCP 8.5). Biogeographic provinces by Morrone et al. (2017). The plots the percentage of decrease, increase, and persistence of the current distribution in the climate scenarios.
In the case of L. tenochtitlan, the distribution is unique since this species has synanthropic habits and is associated with human buildings (Valdez-Mondragón et al., 2019). This species is distributed across the Central region of Mexico, encompassing the TVB with a mean elevation of 2,500 m asl. The environmental conditions associated with this species are characterized by temperate and pine-oak forests with temperatures between 20-30 °C and rainfall between 500 and 850 mm per year (Conagua, 2019; Cortez-Roldán, 2018; Juárez-Sánchez, 2019; Navarro-Rodríguez, 2019; Solís-Catalán, 2020; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, Solís-Catalán et al., 2018; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018). Loxosceles malintzi and L. misteca have also been reported in synanthropic environments (Figs. 30-32). In the case of L. malintzi, records have been taken near human settlements, specifically inside houses, between stacked concrete blocks, and in stored objects, near native vegetation (deciduous forest or dry tropical forest) (Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018). Loxosceles misteca has similarly been recorded within human settlements, leading Dzul-Manzanilla et al. (2014) and Cortez-Roldán (2018) to consider it a synanthropic species. Because of this, factors such as urbanized environments and anthropogenic activities must be considered in the preparation of SDMs (Desales-Lara et al., 2013; Maldonado et al., 2016; Quijano-Ravel & Ponce-Saavedra, 2016; Rodríguez-Rodríguez et al., 2015).
Figures 11-12. Potential distribution of Loxosceles zapoteca under future scenarios. 11, Year 2050 (RCP 4.5); 12, year 2080 (RCP 8.5). Biogeographic provinces by Morrone et al. (2017). The plots the percentage of decrease, increase, and persistence of the current distribution under the climate scenarios.
Several species of the genus Loxosceles from North and South America show a high association with human settlements (Cortez-Roldán, 2018; Fischer & Vasconcelos-Neto, 2005; Sandidge & Hopwood, 2005; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, Solís-Catalán et al., 2018; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018; Vetter & Barger, 2002). Undisturbed areas within and around homes, such as basements and storage areas that provide less variable temperature and humidity, can provide optimal conditions for the spiders’ potential prey and therefore can help establish spider populations within human settlements (Cortez-Roldán, 2018; Juárez-Sánchez, 2019; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, Solís-Catalán et al., 2018; Valdez-Mondragón, Cortez-Roldán, Juárez-Sánchez, & Solís-Catalán, 2018). Saupe et al. (2011) hypothesized that urbanization could modify the distribution of Loxosceles species, since a change in natural areas brings changes in temperature and precipitation within urban areas as well. Within other Loxosceles species, anthropogenic activities have been observed to aid the expansion of ranges (e.g., L. rufescens and L.reclusa, both introduced species in Mexico). Planas et al. (2014) and Taucare-Ríos et al. (2018) demonstrated that the potential for L. rufescens to expand to a global distribution is mainly determined by anthropogenic activities.
The possibility that urbanization has modified the distribution of L. malintzi, L. misteca,and L. tenochtitlan cannot be ruled out since similar cases have been observed with other species in which a change in natural areas modifies the temperature and precipitation within urban areas (Ferrelli et al., 2016; Granados, 2011; Saupe et al., 2011). However, the MDE showed the presence of these species in the BBP and TVB zones, provinces whose physiographic, environmental, and biological characteristics allow for the establishment of L. malintzi, L. misteca, L. tenochtitlan, and L. zapoteca (Figs. 30-33).
The SDMs provide a prediction of the potential distribution with respect to current environmental conditions; however, predictions to the future become more ambiguous. Species have been shown to respond to climatic changes throughout their evolutionary histories, and these rates of change are of primary concern for the future distribution of wild species (Peterson & Vieglais, 2001; Root et al., 2003; Walther et al., 2002). Within Loxosceles, Saupe et al. (2011) suggested that the distribution of L. reclusa, a widespread species from southern USA and introduced in northern Mexico, will be affected by climate change.
According to the analyzed species herein, there is a probability that in future scenarios (years 2050 and 2080) the potential distributions of L. misteca and L. malintzi will increase towards southern latitudes into areas where temperatures are projected to increase, allowing the establishment of new populations and further risks of loxoscelism accidents in the future. These potential changes in environmental conditions to the south in the neotropical region have been previously documented to affect other arachnids, such as scorpions of medical-toxicological importance in South America and Mexico (Martínez et al., 2018; Ureta et al., 2020). The effects of CC on L. reclusa suggest a displacement towards northern latitudes in North America and an expansion towards the east and west, where conditions will resemble those of their current niche. This implies that this species will have the capacity to colonize new areas, thereby increasing the risk of human-spider interaction in new regions of the USA (Saupe et al., 2011). The SDMs generated herein differ from those of Saupe et al. (2011) with respect to the projected increase in areas of distribution to the north. This could be due to the fact that the global circulation models (GCM) used with L. reclusa are based on high population growth rates, changes in land use, and energy. In comparison, the MCG MIROC-5 model used in the present work improves the simulation of the average climate, variability, and CC predictions due to anthropogenic radiative forcing that presents corrections in the precipitation parameters (Watanabe et al., 2010).
Future models for L. tenochtitlan and L. zapoteca show a decreasing tendency in geographic range in comparison with the areas predicted by current models, with distributional changes concentrated in the Chihuahuan Desert (CDP) (L. tenochtitlan) and TVB (L. zapoteca) biogeographic provinces restricting the distribution of both species for the years 2050 and 2080. This region is expected to increase in temperature during this time, reducing the suitable sites for these species. At the beginning of the 20th century, temperature was observed to rise non-uniformly and at an accelerated scale, influencing patterns of temperature and precipitation, both important environmental determinants for species distributions (Cuervo-Robayo et al., 2020; Houghton et al., 2001; Solomon et al., 2007). As adequate environmental conditions decrease, species distributions are consequently reduced (Romo & García-Barros, 2013; Romo et al., 2006). The reduction in the distributions of medically important species could have an important impact since it would entail a reduction in the areas with possible envenomation accidents for humans (Hosseinzadeh, 2020; Lira et al., 2020; Ureta et al., 2020).
Several studies have assessed the potential impact of climate change on the geographic ranges of organisms, with studies on arachnids particularly focusing on ticks (Dermacentor and Ixodes) (Boorgula et al., 2020; Gray et al., 2009) and scorpions (Tityus, Odontobuthus and Centruroides) (Hosseinzadeh, 2020; Lira et al., 2020; Martínez et al., 2018; Ureta et al., 2020). Modifications in geographic distributions could be due to variations in future environmental conditions, such as the isothermality, seasonality of temperature, or seasonal precipitation. However, as mentioned by Romo et al. (2013) and worth consideration here, the existence of environmentally suitable areas does not guarantee the survival of a species. Other factors must be considered, such as dependence on a specialized habitat or microhabitat, which would alter the response capacity to climate change (Bravo-Cadena et al., 2011). Similarly, the ability to disperse or colonize new areas is an important factor in a species’ ability to respond to changes (Bravo-Cadena et al., 2011; Pecl et al., 2017; Yáñez-Arenas et al., 2016). Species with greater dispersal capacity and greater niche width (eurytopic) will have a better response to climate change scenarios by increasing populations and geographic distribution (Bravo-Cadena et al., 2011; Martínez-Meyer et al., 2004; Parra-Olea et al., 2005; Vié et al., 2009). However, species with reduced dispersal capacity and limited niche width (stenotopic), such as Loxosceles spiders, will be drastically affected by climate change, reducing populations and geographic distribution areas (Bravo-Cadena et al., 2011; Hill et al., 2002; Schramm et al., 2021; Vié et al., 2009). In this way, low dispersal capacity is a limiting factor in these spiders’ ability to respond to environmental changes. Although larger geographic areas with adequate conditions may arise in future CC scenarios, these may be inaccessible to the spiders, restricting their geographic distribution (McMahon et al., 2011; Pearson & Dawson, 2003).
With regard to the geographical distribution of the Loxosceles species analyzed, 3 response patterns to climate change are proposed: 1) an increase in the occupancy capacity of new areas with optimal conditions for distribution (increase, L. misteca and L. malintzi); 2) a species will present adequacy to the new environmental conditions, so their distribution will not change (persistence, L. misteca, L. malintzi, and L. zapoteca); and 3) barring expansion or persistence, a species will undergo a reduction in the optimal areas for its survival (decrease, L. tenochtitlan and L. zapoteca), and in very extreme cases, the species could become locally extinct.
Loxosceles spiders are ectothermic organisms with a wide range of thermal tolerance, which could allow for adaptability to climate changes through physiological processes (e.g., temperature regulation). A tolerance to temperatures between 4.5-43.5 °C has been reported in Loxosceles species from North and South America (e.g., L. reclusa and L. intermedia Mello-Leitão, 1934 and L. laeta Nicolet, 1849) (Bonnet, 1996; Canals et al., 2016; Magnelli et al., 2016; Sandidge & Hopwood, 2005; Vetter, 2008, 2015). As such, future generations of the analyzed species herein would be expected to respond to temperature regulation (physiological adaptation), making individuals more tolerant of new climatic conditions in future environments as proposed by Saupe et al. (2011), Uribe- Botero (2015), Stranges et al. (2019), and Ryding et al. (2021).
Projections of climate change scenarios should be taken with caution since there may be variability and uncertainty in climatic conditions associated with the GCM (Collevatti et al., 2013; Chefaoui & Serrão, 2017; Elith & Leathwick, 2009; Nogués-Bravo, 2009; Owens et al., 2013). The Multivariate Environmental Similarity Surface Analysis (MESS) does not show different climatic conditions (not analogous) for the calibration area of the models in the different climate change scenarios in this research (S2). However, we observed in the northern region of Mexico where future climatic conditions could be non-analogous that the values of some bioclimatic variables in the GCM might be more extreme than in those variables used to calibrate the model (Guevara et al., 2019).
The magnitude of CC effects on areas of distribution is not only determined by ecological conditions, but also possibly influenced by socioeconomic factors such as migration, human settlements, and change in land use (Gray et al., 2009). Studies on the influence of climate change on the future distributions of species are relevant to the ecology of zoonotic diseases or in regard to envenomations/poisonings by animals of medical-toxicological importance (Boorgula et al., 2020; Saupe et al., 2011).
Acknowledgements
The first author thanks the Master’s Program at the Centro Tlaxcala de Biología de la Conducta (CTBC), Universidad Autónoma de Tlaxcala, Tlaxcala City, and the Consejo Nacional de Humanidades, Ciencias y Tecnologías (Conahcyt) for scholarship support while completing a Master’s Degree in Biological Science. The second author thanks the program “Jóvenes investigadores por México (ex Cátedras Conahcyt)” and Conahcyt for scientific support for the project No. 59. The second author also thanks SEP-Conahcyt for financial support of the project of Basic Science (Ciencia Básica) 2016, No. 282834. We also thank Edmundo González Santillán and Oscar F. Francke (ex-curator) of the Colección Nacional de Arácnidos (CNAN), Instituto de Biología, UNAM, for providing specimen loans. We are grateful to Brett O. Butler for the English language review of the manuscript, and to the reviewers for their comments and suggestions that improved the manuscript. We thank various people who donated specimens and assisted in field collection efforts. Specimens were collected under Scientific Collector Permit FAUT-0309 from the Secretaría de Medio Ambiente y Recursos Naturales (Semarnat), Mexico, provided to Alejandro Valdez-Mondragón.
References
Araújo, M. B., Thuiller, W., & Pearson, R. G. (2006). Climate warming and the decline of amphibians and reptiles in Europe. Journal of Biogeography, 33, 1712‒1728. https://doi.org/10.1111/j.1365-2699.2006.01482.x
Barbosa, A. M., Real, R., & Vargas, J. M. (2009). Transferability of environmental favourability models in geographic space: the case of the Iberian desman (Galemyspyrenaicus) in Portugal and Spain. EcologicalModelling, 220, 747‒754. https://doi.org/10.1016/j.ecolmodel.2008.12.004
Barve, N., Barve, V., Jiménez-Valverde, A., Lira-Noriega, A., Maher, S. P., & Peterson, A. T. (2011). The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecological Modelling, 222, 1810‒1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011
Boorgula, G. D., Peterson, A. T., Foley, D. H., Ganta, R. R., & Raghavan, R. K. (2020). Assessing the current and future potential geographic distribution of the American dog tick, Dermacentor variabilis (Say) (Acari: Ixodidae) in North America. Plos One, 15, e0237191. https://doi.org/10.1371/journal.pone.0237191
Bravo-Cadena, J., Sánchez-Rojas, G., & Gelviz-Gelviz, S. M. (2011). Estudio de la distribución de las especies frente al cambio climático. Cuadernos de Biodiversidad, 35, 12‒18. https://doi.org/10.14198/cdbio.2011.35.03
Brooker, S., Hay, S. I., & Bundy, D. A. (2002). Tools from ecology: ¿useful for evaluating infection risk models? Trends in Parasitology, 18, 70–74. https://doi.org/10.1016/S1471-4922(01)02223-1
Canals, M., Taucare-Ríos, A., Solís, R., & Moreno, L. (2016). Dimorfismo sexual y morfología funcional de las extremidades de Loxosceleslaeta (Nicolet, 1849). Gayana (Concepción), 80, 161‒168. http://dx.doi.org/10.4067/S0717-65382016000200161
Chefaoui, R. M., & Serrão, E. A. (2017). Accounting for uncertainty in predictions of a marine species: integrating population genetics to verify past distributions. Ecological Modelling, 359, 229‒239. https://doi.org/10.1016/j.ecolmodel.2017.06.006
Cobos, M. E., Peterson, A. T., Osorio-Olvera, L., & Jiménez-García, D. (2019). An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecological Informatics, 53, 100983. https://doi.org/10.1016/j.ecoinf.2019.100983
Collevatti, R. G., Terribile, L. C., de Oliveira, G., Lima-Ribeiro, M. S., Nabout, J. C., & Rangel, T. F. (2013). Drawbacks to paleodistribution modelling: the case of South American seasonally dry forests. Journal of Biogeography, 40, 345‒358. https://doi.org/10.1111/jbi.12005
Compagnucci, R. H. (2011). Historia del cambio climático o calentamiento global. Contribuciones Científicas GAEA, 23, 13‒19.
Conagua (Comisión Nacional del Agua). (2019). Reporte del clima en México: reporte anual 2019. Coordinación General del Servicio Meteorológico Nacional de la Comisión Nacional del Agua Tlalpan, Ciudad de México.
Cortez-Roldán, M. R. (2018). Arañas de importancia médica: distribución y modelaje de nicho ecológico de las especies de arañas violinistas del género Loxosceles Heineken y Lowe, 1832 (Araneae, Sicariidae) de México (Bachelor’s Thesis). Facultad de Agrobiología, UAT, Tlaxcala, México.
Costa, J., Peterson, A. T., & Beard, C. B. (2002). Ecologic niche modeling and differentiation of populations of Triatomabrasiliensis Neiva, 1911, the most important Chagas’ disease vector in northeastern Brazil (Hemiptera, Reduviidae, Triatominae). American Journal of Tropical Medicine and Hygiene, 67, 516‒520. https://doi.org/10.4269/ajtmh.2002.67.516
Cruz-Cárdenas, G., López-Mata, L., Villaseñor, J. L., & Ortiz, E. (2014). Potential species distribution modeling and the use of principal component analysis as predictor variables. Revista Mexicana de Biodiversidad, 85, 189‒199. https://doi.org/10.7550/rmb.36723
Cuervo-Robayo, A. P., Ureta, C., Gómez-Albores, M. A., Meneses-Mosquera, A. K., Téllez-Valdés, O., & Martínez-Meyer, E. (2020). One hundred years of climate change in Mexico. Plos One, 15, e0209808. https://doi.org/10.1371/journal.pone.0209808
Desales-Lara, M. A., Francke, O. F., & Sánchez-Nava, P. (2013). Diversidad de arañas (Arachnida: Araneae) en hábitats antropogénicos. Revista Mexicana de Biodiversidad, 84, 291‒305. https://doi.org/10.7550/rmb.31708
Dzul-Manzanilla, F. A., Herrera, L. H., Juárez, O. V., Leyva, E. T., Gutiérrez, C., & Castro J. T. L. (2014). Arañas de importancia médica en el estado de Guerrero. Entomología Mexicana, 1, 909‒913.
Elith, J., & Leathwick, J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics, 40, 677‒697. https://doi.org/10.1146/annurev.ecolsys.110308.120159
Escobar, L. E., Lira-Noriega, A., Medina-Vogel, G., & Peterson, A. T. (2014). Potential for spread of the white-nose fungus (Pseudogymnoascusdestructans) in the Americas: use of Maxent and NicheA to assure strict model transference. Geospatial Health, 9, 221‒229. https://doi.org/10.4081/gh. 2014.19
Fernández, A., Zavala, J., Romero, R., Conde, A. C., & Trejo, I. (2016). Actualización de los escenarios de cambio climático para estudios impactos, vulnerabilidad y adaptación en México y Centroamérica. CDMX: Centro de Ciencias de la Atmosfera, UNAM. https://doi.org/10.7818/ecos.2016.25-1.20
Ferrelli, F., Bustos, M. L., & Piccolo, M. C. (2016). La expansión urbana y sus impactos sobre el clima y la sociedad de la ciudad de Bahía Blanca, Argentina. Estudios Geográficos, 281, 469–489. https://doi.org/10.3989/estgeogr.201615
Ferretti, N. E., Arnedo, M., & González, A. (2018). Impact of climate change on spider species distribution along the La Plata River basin, southern South America: projecting future range shifts for the genus Stenoterommata (Araneae, Mygalomorphae, Nemesiidae). Annales Zoologici Fennici, 55, 123‒133. https://doi.org/10.5735/086.055.0112
Fischer, M. L., & Vasconcellos-Neto, J. (2005). Microhabitats occupied by Loxoscelesintermedia and Loxosceleslaeta (Araneae: Sicariidae) in Curitiba, Paraná, Brazil. Journal of Medical Entomology, 42, 756‒765. https://doi.org/10.1603/0022-2585(2005)042[0756:moblia]2.0.co;2
Foley, D. H., Rueda, L. M., Peterson, A. T., & Wilkerson, R. C. (2008). Potential distribution of two species in the medically important Anophelesminimus complex (Diptera: Culicidae). Journal Medical Entomology, 45, 852‒860. https://doi.org/10.1603/0022-2585(2008)45[852:pdotsi]2.0.co;2
Garza, M., Feria-Arroyo, T. P., Casillas, E. A., Sanchez-Cordero, V., Rivaldi, C. L., & Sarkar, S. (2014). Projected future distributions of vectors of Trypanosomacruzi in North America under climate change scenarios. Plos Neglected Tropical Diseases, 8, e2818. https://doi.org/10.1371/journal.pntd.0002818
Gertsch, W. J. (1958). The spider genus Loxosceles in North America, Central America, and the West Indies. American Museum Novitates, 1907, 1‒46.
Gertsch, W. J. (1967). The spider genus Loxosceles in South America (Araneae, Scytodidae). Bulletin of the American Museum of Natural History, 136, 117‒174.
Gertsch, W. J. & Ennik, F. (1983). The spider genus Loxosceles in North America, Central America, and the West Indies (Araneae, Loxoscelidae). Bulletin of the American Museum of Natural History, 175, 264‒360.
Graham, C. H., Ferrier, S., Huettman, F., Moritz, C., & Peterson, A. T. (2004). New developments in museum-based informatics and applications in biodiversity analysis. Trends in Ecology & Evolution, 19, 497‒503. https://doi.org/10.1016/j.tree.2004.07.006
Granados, J. L. S. (2011). Efectos del crecimiento urbano sobre el sistema climático en la Región Paso del Norte (1975 y 2010). 200 años de urbanización en México. Memorias del XII Seminario-Taller Internacional de la Red Mexicana de Ciudades hacia la Sustentabilidad. Santiago, 16-17 de agosto 2011.
Gray, J. S., Dautel, H., Estrada-Peña, A., Kahl, O., & Lindgren, E. (2009). Effects of climate change on ticks and tick-borne diseases in Europe. Interdisciplinary Perspectives on Infectious Diseases, 2009, 593232. https://doi.org/10.1155/ 2009/593232
Guevara, L., Morrone, J. J., & León-Paniagua, L. (2019). Spatial variability in species’ potential distributions during the last glacial maximum under different global circulation models: Relevance in evolutionary biology. Journal of Zoological Systematics and Evolutionary Research, 57, 113‒126. https://doi.org/10.1111/jzs.12238
Hill, J. K., Thomas, C. D., Fox, R., Telfer, M. G., Willis, S. G., & Asher, J. (2002). Responses of butterflies to twentieth century climate warming: implications for future ranges. Proceedings of the Royal Society of London. Series B: Biological Sciences, 269, 2163‒2171. https://doi.org/10.1098%2Frspb.2002.2134
Hosseinzadeh, M. S. (2020). Potential distribution and effects of climate change on the risk of scorpion sting with endemic and medically important scorpion Odontobuthusdoriae (Thorell, 1876) (Arachnida: Scorpionidae: Buthidae) in Iran. Zoology and Ecology, 30, 109‒115. https://doi.org/10.35513/21658005.2020.2.4
Houghton, J. T., Ding, Y. D. J. G., Griggs, D. J., Noguer, M., van der Linden, P. J., & Dai, X. (2001). Climate change 2001: the scientific basis. The Press Syndicate of the University of Cambridge.
INEGI (Instituto Nacional de Estadística y Geografía). (2017). Guía para la interpretación de cartografía: uso del suelo y vegetación. Escala 1:250 000: serie VI. Instituto Nacional de Estadística y Geografía. México.
INEGI (Instituto Nacional de Estadística y Geografía). (2020). Censo de población y vivienda 2020. Retrieved from Instituto Nacional de Estadística y Geografía on: 12/05/2024. https://www.inegi.org.mx/programas/ccpv/2020/
Jiménez-García, D., & Peterson, A. T. (2019). Climate change impact on endangered cloud forest tree species in Mexico. Revista Mexicana de Biodiversidad, 90, e902781. https://doi.org/10.22201/ib.20078706e.2019.90.2781
Juárez-Sánchez, A. R. (2019). Arañas de importancia médica: estudio ultra morfológico de estructuras somáticas y diagnósticas de arañas violinistas del género Loxosceles Heineken & Lowe, 1832 (Araneae, Sicariidae) de México (Bachelors’ Thesis). Facultad de Agrobiología, UAT, Tlaxcala, México.
Lira, A. F. D. A., Badillo-Montaño, R., Lira-Noriega, A., & de Albuquerque, C. M. R. (2020). Potential distribution patterns of scorpions in north-eastern Brazil under scenarios of future climate change. Austral Ecology, 45, 215‒228. https://doi.org/10.1111/aec.12849
Liu, Y., & Shi J. (2020). Predicting the Potential Global Geographical Distribution of Two Icerya Species under Climate Change. Forests, 11, 684. https://doi.org/10.3390/f11060684
Magnelli, L. M., Peña, H. E., Castillo, G. A. & Ortiz, C. R. (2016). Loxoscelismo local y sistémico. Acta Médica Grupo Ángeles, 14, 36‒38.
Maldonado, C.J., Gutiérrez, C.D.A., & Ponce-Saavedra, J. (2016). Comparación de la riqueza de arañas saltarinas (Araneae: Salticidae) en dos sitios contrastantes en grado de antropización en Morelia Michoacán, México. In REFAMA Libro de Resúmenes del II Congreso de Fauna Nativa en Ambientes Antropizados. Red Temática Biología, Manejo y Conservación de Fauna Nativa en Ambientes Antropizados. Querétaro, Qro. México.
Mammola, S., Goodacre, S. L., & Isaia, M. (2018). Climate change may drive cave spiders to extinction. Ecography, 41, 233‒243. http://dx.doi.org/10.1111/ecog.02902
Manríquez, J. J., & Silva, S. (2009). Loxoscelismo cutáneo y cutáneo-visceral: Revisión sistemática. Revista Chilena de Infectología, 26, 420‒432. http://dx.doi.org/10.4067/S0716-10182009000600004
Marques, R., Krüger, R. F., Peterson, A. T., de Melo, L. F., Vicenzi, N., & Jiménez-García, D. (2020). Climate change implications for the distribution of the babesiosis and anaplasmosis tick vector, Rhipicephalus (Boophilus) microplus. Veterinary Research, 51, 1‒10. https://doi.org/10.1186/s13567-020-00802-z
Martínez, P. A., Andrade, M. A., & Bidau, C. J. (2018). Potential effects of climate change on the risk of accidents with poisonous species of the genus Tityus (Scorpiones, Buthidae) in Argentina. Spatial and Spatio-Temporal Epidemiology, 25, 67‒72. https://doi.org/10.1016/j.sste.2018.03.002
Martínez, P. A., da Fonseca-Teixeira, I. B, Siqueira-Silva, T., da Silva, F. F., Gonzaga-Lima, L. A., Chaves-Silveira, J. et al. (2024). Climate change-related distributional range shifts of venomous snakes: a predictive modelling study of effects on public health and biodiversity. The Lancet Planetary Health, 8,163‒171. https://doi.org/10.1016/S2542-5196(24)00005-6
Martínez-Meyer, E., Peterson, A. T., & Hargrove, W. W. (2004). Ecological niches as stable distributional constraints on mammal species, with implications for Pleistocene extinctions and climate change projections for biodiversity. Global Ecology and Biogeography, 13, 305‒314. https://doi.org/10.1111/j.1466-822X.2004.00107.x
McMahon, S. M., Harrison, S. P., Armbruster, W. S., Bartlein, P. J., Beale, C. M., & Edwards, M. E. (2011). Improving assessment and modelling of climate change impacts on global terrestrial biodiversity. Trends in Ecology & Evolution, 26, 249‒259. https://doi.org/10.1016/j.tree.2011.02.012
Morrone, J. J., Escalante, T., & Rodríguez-Tapia, G. (2017). Mexican biogeographic provinces: Map and shapefiles. Zootaxa, 4277, 277‒279. http://dx.doi.org/10.11646/zootaxa.4277.2.8
Mota-Vargas, C., Encarnación-Luévano, A., Ortega-Andrade, H. M., Prieto-Torres, D. A., Peña-Peniche, A., & Rojas-Soto, O. R. (2019). Una breve introducción a los modelos de nicho ecológico. La biodiversidad en un mundo cambiante: fundamentos teóricos y metodológicos para su estudio. Ciudad de México: Universidad Autónoma del Estado de Hidalgo/ Libermex.
Navarro-Rodríguez, C. I., & Valdez-Mondragón, A. (2020). Description of a new species of Loxosceles Heineken & Lowe (Araneae, Sicariidae) recluse spiders from Hidalgo, Mexico, under integrative taxonomy: morphological and DNA barcoding data (CO1+ ITS2). European Journal of Taxonomy, 704, 1‒30. https://doi.org/10.5852/ejt.2020.704
Navarro-Rodríguez, C. I. (2019). Delimitación de las especies mexicanas de arañas del género Loxosceles Heineken y Lowe (Araneae, Sicariidae) del Centro Occidente de México con evidencia molecular y morfológica (M. Sc. Thesis). Centro Tlaxcala de Biología de la Conducta, Universidad Autónoma de Tlaxcala, Tlaxcala, México.
Ortega-Andrade, H. M., Prieto-Torres, D. A., Gómez-Lora, I., & Lizcano, D. J. (2015). Ecological and geographical analysis of the distribution of the mountain tapir (Tapirus pinchaque) in Ecuador: importance of protected areas in future scenarios of global warming. Plos One, 10, e0121137. http://dx.doi.org/10.1371/journal.pone.0121137
Osorio-Olvera, L., Lira-Noriega, A., Soberón, J., Peterson, A. T., Falconi, M., Contreras-Díaz, R. G. et al. (2020). ntbox: an R package with graphical user interface for modelling and evaluating multidimensional ecological niches. Methods in Ecology and Evolution, 11, 1199‒1206. https://doi.org/10.1111/2041-210X.13452
Owens, H. L., Campbell, L. P., Dornak, L. L., Saupe, E. E., Barve, N., & Soberón, J. (2013). Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecological modelling, 263, 10‒18. https://doi.org/10.1016/j.ecolmodel.2013.04.011
Parra-Olea, G., Martínez-Meyer, E., & de León, G. P. P. (2005). Forecasting climate change effects on salamander distribution in the highlands of Central Mexico 1. Biotropica: The Journal of Biology and Conservation, 37, 202‒208. https://doi.org/10.1111/j.1744-7429.2005.00027.x
Pearson, R. G., & Dawson, T. P. (2003). Predicting the impacts of climate change on the distribution of species: ¿are bioclimate envelope models useful? Global Ecology and Biogeography, 12, 361‒371. https://doi.org/ 10.1046/j.1466-822X.2003.00042.x
Pecl, G. T., Araújo, M. B., & Bell, J. D. (2017). Biodiversity redistribution under climate change: impacts on ecosystems and human well-being. Science, 355, eaai9214. https://doi.org/10.1126/science.aai9214
Peterson, A. T. (2009). Shifting suitability for malaria vectors across Africa with warming climates. BMC Infectious Diseases, 9, 59. https://doi.org/10.1186/1471-2334-9-59
Peterson, A. T., & Shaw, J. (2003). Lutzomyia vectors for cutaneous leishmaniasis in southern Brazil: ecological niche models, predicted geographic distributions, and climate change effects. International Journal of Parasitology, 33, 919‒931. https://doi.org/10.1016/s0020-7519(03)00094-8
Peterson, A. T., Campbell, L. P., Moo-Llanes, D. A., Travi, B., González, C., & Ferro, M. C. (2017). Influences of climate change on the potential distribution of Lutzomyia longipalpissensulato (Psychodidae: Phlebotominae). International Journal of Parasitology, 47, 667‒674. https://doi.org/10.1016/j.ijpara.2017.04.007
Peterson, A. T., Stewart, A., Mohamed, K. I., & Araújo, M. B. (2008). Shifting global invasive potential of European plants with climate change. Plos One, 3,e2441. https://doi.org/10.1371/journal.pone.0002441
Phillips, S. J., & Dudík, M. (2004). A maximum entropy approach to species distribution modeling. Proceedings of the 21st International Conference on Machine Learning. Baniff, Canada.
Phillips, S. J., & Dudík, M. (2008). Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 31, 161‒175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Phillips, S., Anderson, R., & Sphapire, R. (2006). Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190, 231‒259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Planas, E., & Ribera, C. (2014). Uncovering overlooked island diversity: colonization and diversification of the medically important spider genus Loxosceles (Arachnida: Sicariidae) on the Canary Islands. Journal of Biogeography, 41, 1255‒1266. http://dx.doi.org/10.1111/jbi.12321
Quijano-Ravell, A., & Ponce-Saavedra, J. (2016). Géneros de saltícidos (Arachnida: Salticidae: Araneae) urbanos de “Ciudad Caucel”, comisaría de Mérida, Yucatán, México. In REFAMA Libro de Resúmenes del II Congreso de Fauna Nativa en Ambientes Antropizados. Red Temática Biología, Manejo y Conservación de Fauna Nativa en Ambientes Antropizados. Querétaro, Qro. México.
Ramos-Rodríguez, H. G., & Méndez, J. D. (2008). Necrotic araneism. A review of the Loxosceles genus. I. General aspects, distribution and venom composition. Advances in Environmental Biology, 2, 9‒19.
Rodríguez-Rodríguez, S. E., Solís-Catalán, K. P., & Valdez-Mondragón, A. (2015). Diversity and seasonal abundance of anthropogenic spiders (Arachnida: Araneae) in different urban zones of the city of Chilpancingo, Guerrero, Mexico. Revista Mexicana de Biodiversidad, 86, 962‒971. https://doi.org/10.1016/j.rmb.2015.09.002
Romero, G. Q., Gonçalves-Souza, T., Kratina, P., Marino, N. A., Petry, W. K., & Sobral-Souza, T. (2018). Global predation pressure redistribution under future climate change. Nature Climate Change, 8, 1087‒1091. https://www.nature.com/articles/s41558-018-0347-y
Romo, H., García-Barros, E., & Munguira, M. L. (2006). Distribución potencial de trece especies de mariposas diurnas amenazadas o raras en el área ibero-balear (Lepidoptera: Papilionoidea & Hesperioidea). Boletín Asociación Española de Entomología, 30, 25‒49.
Romo, H., Sanabria, P., & García-Barros, E. (2013). Predicción de los impactos del cambio climático en la distribución sobre las especies de Lepidoptera. El caso del género Boloria Moore, 1900 en la península Ibérica (Lepidoptera: Nymphalidae). SHILAP Revista de Lepidopterología, 41, 267‒286.
Root, T. L., Price, J. T., Hall, K. R., Schneider, S. H., Rosenzweig, C., & Pounds, J. A. (2003). Fingerprints of global warming on wild animals and plants. Nature, 421, 57‒60. https://doi.org/10.1038/nature01333
Roslin, T., Hardwick, B., Novotny, V., Petry, W. K., Andrew, N. R., & Asmus, A. (2017). Higher predation risk for insect prey at low latitudes and elevations. Science, 356, 742‒744. https://doi.org/10.1126/science.aaj1631
Ryding, S., Klaassen, M., Tattersall, G. J., Gardner, J. L. & Symonds, M. R. (2021). Shape-shifting: changing animal morphologies as a response to climatic warming. Trends in Ecology & Evolution, 36, 1036‒1048. https://doi.org/10.1016/j.tree.2021.07.006
Saupe, E. E., Papes, M., Selden, P. A., & Vetter, R. S. (2011). Tracking a medically important spider: climate change, ecological niche modeling, and the brown recluse (Loxoscelesreclusa). Plos One, 6, 17‒31. https://doi.org/10.1371/journal.pone.0017731
Schramm, F. D., Valdez-Mondragón, A., & Prendini, L. (2021). Volcanism and palaeoclimate change drive diversification of the world’s largest whip spider (Amblypygi). MolecularEcology, 30, 2872‒2890. https://doi.org/10.1111/mec.15924
Simó, M., Guerrero, J. C., Giuliani, L., Castellano, I., & Acosta, L. E. (2014). A predictive modeling approach to test distributional uniformity of Uruguayan harvestmen (Arachnida: Opiliones). Zoological Studies, 53, 1‒13. http://dx.doi.org/10.1186/s40555-014-0050-2
Solís-Catalán, K. P. (2020). Análisis morfométrico de estructuras sexuales y somáticas de las especies mexicanas de arañas del género Loxosceles Heineken y Lowe (Araneae, Sicariidae) del Centro-Occidente de México (M. Sc. Thesis). Centro Tlaxcala de Biología de la Conducta, Universidad Autónoma de Tlaxcala, Tlaxcala, México.
Solomon, S., Manning, M., Marquis, M., & Qin, D. (2007). Climate change 2007-the physical science basis: working group I contribution to the fourth assessment report of the IPCC. Vol. 4. London: Cambridge University Press.
Steen, V., Sofaer, H. R., Skagen, S. K., Ray, A. J., & Noon, B. R. (2017). Projecting species’ vulnerability to climate change: ¿which uncertainty sources matter most and extrapolate best? Ecology and Evolution, 7, 8841‒8851. https://doi.org/10.1002/ece3.3403
Stranges, S., Cuervo-Robayo, A. P., Martínez-Meyer, E., Morzaria-Luna, H. N., & Reyes-Bonilla, H. (2019). Distribución potencial bajo escenarios de cambio climático de corales del género Pocillopora (Anthozoa: Scleractinia) en el Pacífico oriental tropical. Revista Mexicana de Biodiversidad, 90, 1‒16. https://doi.org/10.22201/ib.20078 706e.2019.90.2696
Taucare-Ríos, A., Nentwig, W., Bizama, G., & Bustamante, R. O. (2018). Matching global and regional distribution models of the recluse spider Loxoscelesrufescens: to what extent do these reflect niche conservatism? Medical and Veterinary Entomology, 32, 490‒496. https://doi.org/10.1111/mve.12311
Ureta, C., González, E. J., Ramírez-Barrón, M., Contreras-Félix, G. A., & Santibáñez-López, C. E. (2020). Climate change will have an important impact on scorpion’s fauna in its most diverse country, Mexico. Perspectives in Ecology and Conservation, 18, 116‒123. https://doi.org/10.1016/j.pecon.2020.04.003
Uribe-Botero, E. (2015). El cambio climático y sus efectos en la biodiversidad en América Latina. Santiago de Chile: UNCEPAL.
Valdez-Mondragón, A. (2020). Expedición a Baja California para colecta de arañas violinistas. Boletín de la Asociación Mexicana de Sistemática de Artrópodos (AMXSA), 4, 11‒16.
Valdez-Mondragón, A., Cortez-Roldán, M. R., Juárez-Sánchez, A. R., Solís-Catalán, K. P., & Navarro-Rodríguez, C. I. (2018). Arañas de importancia médica: arañas violinistas del género Loxosceles en México, ¿qué sabemos acerca de su distribución y biología hasta ahora? Boletín de la Asociación Mexicana de Sistemática de Artrópodos (AMXSA), 2, 14‒24.
Valdez-Mondragón, A., Cortez-Roldán, M. R., Juárez-Sánchez, A. R., & Solís-Catalán, K. P. (2018). A new species of Loxosceles Heineken & Lowe (Araneae, Sicariidae), with updated distribution records and biogeographical comments for the species from Mexico, including a new record of Loxoscelesrufescens (Dufour). Zookeys, 802, 39‒66. https://doi.org/10.3897/zookeys.802.28445
Valdez-Mondragón, A., Navarro-Rodríguez, C. I., Solís-Catalán, K. P., Cortez-Roldán, M. R., & Juárez-Sánchez, A. R. (2019). Under an integrative taxonomic approach: the description of a new species of the genus Loxosceles (Araneae, Sicariidae) from Mexico City. Zookeys, 892, 93–133. https://doi.org/10.3897/zookeys.892.39558
Vetter, R. S. (2005). Arachnids submitted as suspected brown recluse spiders (Araneae: Sicariidae): Loxosceles spiders are virtually restricted to their known distributions but are perceived to exist throughout the United States. Journal of Medical Entomology, 42, 512‒521. https://doi.org/10.1093/jmedent/42.4.512
Vetter, R. S. (2008). Spiders of the genus Loxosceles (Araneae, Sicariidae): a review of biological, medical and psychological aspects regarding envenomations. Journal of Arachnology, 36, 150‒163. http://dx.doi.org/10.1636/RSt08-06.1
Vetter, R. S. (2015). The brown recluse spider. Ithaca and London: Cornell University Press/ Comstock Publishing Associates.
Vetter, R. S., & Barger, D. K. (2002). An infestation of 2,055 brown recluse spiders (Araneae: Sicariidae) and no envenomations in a Kansas home: implications for bite diagnoses in nonendemic areas. Journal of Medical Entomology, 39, 948‒951. https://doi.org/10.1603/0022-2585-39.6.948
Vetter, R. S., & Bush, S. P. (2002). Reports of presumptive brown recluse spider bites reinforce improbable diagnosis in regions of North America where the spider is not endemic. Clinical Infectious Diseases, 35, 442‒445. https://doi.org/10.1086/341244
Vetter, R. S., Cushing, P. E., Crawford, R. L., & Royce, L. A. (2003). Diagnoses of brown recluse spider bites (loxoscelism) greatly outnumber actual verifications of the spider in four western American states. Toxicon, 42, 413‒418. https://doi.org/10.1016/s0041-0101(03)00173-9
Vié, J. C., Hilton-Taylor, C., & Stuart, S. N. (2009). Wildlife in a changing world: an analysis of the 2008 IUCN Red List of threatened species. Gland, Switzerland. IUCN.
Waltari, E., & Guralnick, R. P. (2009). Ecological niche modelling of montane mammals in the Great Basin, North America: examining past and present connectivity of species across basins and ranges. Journal of Biogeography, 36, 148‒161. http://dx.doi.org/10.1111/j.1365-2699.2008.01959.x
Walther, G. R., Post, E., Convey, P., Menzel, A., Parmesan, C., & Beebee, T. J. (2002). Ecological responses to recent climate change. Nature, 416, 389‒395. http://dx.doi.org/ 10.1038/416389a
Warren, D. L., Glor, R. E., & Turelli, M. (2008). Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution: International Journal of Organic Evolution, 62,2868‒2883. https://doi.org/10.1111/j.1558-5646.2008.00482.x
Warren, D. L., & Seifert, S. N. (2011). Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria. EcologicalApplications, 21, 335‒342. https://doi.org/10.1890/10-1171.1
Watanabe, M., Suzuki, T., O’ishi, R., Komuro, Y., Watanabe, S., Emori, S. et al. (2010). Improved climate simulation by MIROC5: Mean states, variability, and climate sensitivity. Journal of Climate, 23, 6312‒6335. https://doi.org/10.1175/2010JCLI3679.1
Wiens, J. J., & Graham, C. H. (2005). Niche conservatism: integrating evolution, ecology, and conservation biology. Annual Review of Ecology, Evolution, and Systematics, 36, 519‒539. https://doi.org/10.1146/annurev.ecolsys.36.102803.095431
World Spider Catalog (WSC). (2024). World Spider Catalog. Version 25. Natural History Museum Bern, available at http://wsc.nmbe.ch
Yáñez-Arenas, C., Peterson, A. T., Rodríguez-Medina, K., & Barve, N. (2016). Mapping current and future potential snakebite risk in the new world. ClimaticChange, 134, 697‒711.
Edmundo González-Santillán a, Laura L. Valdez-Velázquez b, *, Ofelia Delgado-Hernández c, Jimena I. Cid-Uribe d, María Teresa Romero-Gutiérrez e, Lourival D. Possani d
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Arácnidos, Tercer Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Universidad de Colima, Facultad de Ciencias Químicas y Facultad de Medicina, Km 9 Carretera Colima-Coquimatlán, 28400 Coquimatlán, Colima, Mexico
c Instituto Francisco Possenti, Av. Toluca 621, Olivar de los Padres, Álvaro Obregón, 01780 Mexico City, Mexico
d Universidad Nacional Autónoma de México, Instituto de Biotecnología, Avenida Universidad 2001, Colonia Chamilpa, 62210 Cuernavaca, Morelos, Mexico
e Universidad de Guadalajara, Centro Universitario Tlajomulco, Departamento de Innovación Tecnológica, Carretera Tlajomulco – Santa Fé Km. 3.5 No.595 Lomas de Tejeda, 45641 Tlajomulco de Zúñiga, Jalisco, Mexico
Scorpion species diversity in Colima was investigated with a multigene approach. Fieldwork produced 34 lots of scorpions that were analyzed with 12S rDNA, 16S rDNA, COI, and 28S rDNA genetic markers. Our results confirmed prior phylogenetic results recovering the monophyly of the families Buthidae and Vaejovidae, some species groups, and genera. We recorded 11 described species of scorpions and found 3 putatively undescribed species of Centruroides, 1 of Mesomexovis, and 1 of Vaejovis. Furthermore, we obtained evidence that Centruroides elegans, C. infamatus,and C. limpidus do not occur in Colima, contrary to prior reports. Seven genetically different and medically relevant species of Centruroides for Colima are recorded for the first time. We used the InDRE database (Instituto de Diagnóstico y Referencia Epidemiológicos), which contains georeferenced points of scorpions, to estimate the distribution of the scorpion species found in our fieldwork. Finally, we discuss from a biogeographical, ecological, and medical point of view the presence and origin of the 14 scorpion species found in Colima.
Keywords: Barcoding; Holotype; Medical relevance; Microendemic; New species; Species group; Substrate-specialist
Una aproximación multigenes para identificar a las especies de alacranes (Arachnida: Scorpiones) de Colima, México, con cometarios sobre la diversidad de sus venenos
Resumen
La diversidad de especies de alacranes de Colima se investigó utilizando una aproximación multigenes. Del trabajo de campo se obtuvieron 34 lotes de alacranes que fueron analizados con los marcadores 12S rDNA, 16S rDNA, COI, y 28S rDNA. La comparación con trabajos de filogenia previos nos permitió confirmar la monofilia de las familias Buthidae y Vaejovidae, de algunos grupos de especies y géneros. Encontramos 11 especies de alacranes descritas, 3 putativamente nuevas de Centruroides, 1 de Mesomexovis y 1 de Vaejovis. También obtuvimos evidencia de que Centruroides elegans, C. infamatus y C. limpidus no están distribuidos en Colima, como se registró en trabajos anteriores. Reportamos 7 especies genéticamente distintas y de importancia médica para Colima. Usamos la base de datos del InDRE (Instituto de Diagnóstico y Referencia Epidemiológicos) que contiene puntos georreferenciados de alacranes para estimar la distribución de las especies que recolectamos en el campo. Finalmente, discutimos desde una perspectiva biogeográfica, ecológica y de importancia médica las 14 especies de alacranes que reportamos para Colima.
Palabras clave: Código de barras; Holotipo; Importancia médica; Microendémico; Especie nueva; Grupo de especies; Sustrato-especialista
Introduction
The knowledge of scorpion diversity in North America has improved recently (González-Santillán & Prendini, 2013; Goodman, Prendini, Francke et al., 2021; Ponce-Saavedra & Francke, 2019; Santibáñez-López et al., 2014); however, much remains to be discovered. Local-scale inventories may be a solution to unveil species communities that, in turn, can help conform regional faunas. This approach has rarely been applied to study scorpion diversity. Furthermore, few local faunal studies have been conducted in Mexico, and only a handful of them have been published; other revisionary contributions included limited fieldwork effort (Baldazo-Monsivais et al., 2012, 2016, 2017).
The International Barcode of Life (iBOL) has grown as a powerful tool for discovering biodiversity, among other applications (https://ibol.org). Scorpion barcoding studies have permitted the identification and delimitation of species in several regions of the world (Fet et al., 2014, 2016; Goodman, Prendini, & Esposito, 2021; Podnar et al., 2021). Despite the high diversity of scorpions in Mexico —an update by Ponce-Saavedra et al. (2023) comprises 311 scorpion species— only 1 mini-barcoding study has been conducted (Goodman, Prendini, & Esposito, 2021). Herein, we present a second scorpion barcoding study for this country but aim at discovering the components of a local scorpion assembly. Colima exhibits a complex topology comprising littorals with a tropical climate and extreme topological variation from sea level to mountain ranges rising to over 4,000 m in approximately 5,600 km2. Colima’s territory supports a rich local flora and fauna (Ramírez-Ruiz & Bretón-González, 2016). Colima lies between the limits of the Nearctic and Neotropical biogeographical Realms (Fig. 1B). Beyond its complexity, Colima represents an enclosed littoral surrounded by mountain ranges and 3 large rivers that divide the territory into 2 sections (Fig. 1), which makes it a well-defined and manageable geographical unit ideal for studying a unique community of scorpions. Thus far, 2 families, 3 subfamilies, 5 genera, and 12 species of scorpions have been identified in Colima (Table 1). The knowledge of scorpion diversity and evolutionary studies in Mexico has steadily been unveiling one of the vastest biodiversity hotspots in the world. For instance, 2 of the most diverse scorpion families and subfamilies, Vaejovidae (Syntropinae) and Buthidae (Centruroidinae), have been treated in recent phylogenetic and taxonomic analyses (Esposito & Prendini, 2019; González-Santillán & Prendini, 2013, 2015).
This contribution aims to survey the scorpion richness in Colima, using not only the COI-barcoding genetic marker but 2 additional mitochondrial markers and 1 nuclear marker to establish a framework to build a stable and predictable taxonomy. Taking advantage of robust phylogenies produced for the 2 families distributed in Colima, we use these topologies as a baseline for comparison to test the presence of several previously reported species and to taxonomically circumscribe our fresh samples. Unlike other barcoding studies, our approach seeks to unveil the richness within the state instead of focusing on delimiting the species of a taxonomic group of scorpions.
Materials and methods
We conducted field collections during May and September 2015-2018 in various ecosystems, including tropical deciduous forest, oak-pine, and tropical forest within the state of Colima, at elevations ranging from 47 to 2,200 m. (Table 2). Logistically, we leveraged our collection sites with the help of private landowners who gave us access to their property. Specimen collection methods included direct collection during the day by moving objects on the ground or by ultraviolet detection during the night. To preserve specimens, ethyl alcohol at 90% was used and stored at -80 °C. Each specimen lot carried a label with coordinates and locality information. We obtained scorpions from 14 localities and sequenced 18 samples of Centruroides from Colima (Table 2).
Table 1
List of families, subfamilies, genera, and species recorded in the state of Colima. *Species of Centruroides cited by Ponce-Saavedra et al. (2016). The species in bold font were not found in Colima in this study. Numbered species were reported by González-Santillán et al. (2019).
11. Vaejovis monticola Sissom, 1989 12. Vaejovis sp. mexicanus group
Figure 1. Map of the west coast of Mexico. A, Orographic and hydrographic elements of Colima (COL) and the surrounding states of Jalisco (JAL) and Michoacán (MIC). 1, Manantlán Sierra; 2, massive Cerro Grande; 3, Colima Volcano; 4, Marabasco or Cihuatlán River; 5, Armería River; 6, Coahuayana River. B, Biogeographical provinces (Morrone et al., 2017). Area within the green line corresponds to the Sierra Madre del Sur and north Colima Volcano Trans Mexican Volcanic Belt province (Morrone et al., 2017) —notice that both provinces are connected in Colima. Area outside the green line corresponds to the Pacific Lowlands province (Morrone et al., 2017). Orographic components are indicated in gray scale from light low elevation to dark high elevation.
Table 2
Collection sites of the scorpion species used in this study. The number within parenthesis after the species name is the number of samples processed from this locality and included in the phylogenetic analyses as terminals. Superscript numbers indicate sources of sequences as follows: 1Bolaños et al. (2019), 2Esposito et al. (2018), 3Esposito and Prendini (2019), 4González-Santillán and Prendini (2015). Cells filled in grey color are samples obtained from GenBank.
Species
Municipality
Locality
Latitude
Longitude
Elevation
Thorellius cristimanus (2)
Comala
La Yerbabuena
19°27′59.55″
-103°41′46.64′′
1,358 m
Centruroides ornatus (2)
Comala
Agosto
19°23′51.74′′
-103°44′03.08′′
1,076 m
Thorellius cristimanus
*Centruroides tecomanus2
Colima
Comunidad La Capacha
19°04′58.40′′
-103°41′24.67′′
656 m
*Centruroides tecomanus2
Colima
Tepames
19°06′22.8′′
-103°59′11.07′′
450 m
Thorellius intrepidus (3)
Coquimatlán
El Palapo
19°11′54.6′′
-103°54′50′′
275 m
*Centruroides tecomanus2
Cuauhtémoc
Camino a Altozano
19°18′30.19′′
-103°40′23.83′′
789 m
Thorellius cristimanus
*Centruroides tecomanus1(2)
Cuauhtémoc
Ocotillo
19°20′00.00′′
-103°39′02.00′′
895 m
*Centruroides tecomanus2
Ixtlahuacán
San Gabriel
18°54′24.48′′
-103°44′05.61′′
462 m
Mesomexovis aff. occidentalis
Centruroides hirsutipalpus
Minatitlán
Minatitlán
19°23′01.73′′
-104°03′35.19′′
703 m
Centruroides possanii (2)
Minatitlán
Terrero
19°26′35.94′′
-103°57′05.67′′
2,200 m
Centruroides possanii (2)
Minatitlán
Mirador el Filete
19°26′40′′
-103°58′10′′
2,137 m
Vaejovis sp. (mexicanus group)
Centruroides sp. 2 (2)
Manzanillo
La central
19°08′38.14′′
-104°26′04.10′′
47 m
*Centruroides tecomanus1
*Centruroides tecomanus2
Centruroides sp. 1
Tecomán
Chanchopa
18°51′58.55′′
-103°44′10.10′′
41 m
Thorellius intrepidus (2)
Villa de Álvarez
Rancho Blanco
19°14′24.71′′
-103°45′49.45′′
455 m
Thorellius intrepidus4
La Huerta (Jal.)
Estación de Biología Chamela
19°30′14.15′′
-105°2′16.50′′
33 m
Centruroides elegans (2)
Centruroides suffusus
Durango (Dgo.)
El Salto, 50 km E Durango
23°45′51.41′′
-105°19′49.16′′
2,847 m
Centruroides limpidus
Iguala (Gro.)
Iguala
18°20′11.87′′
-99°29′29.65′′
823 m
Centruroides sculpturatus
Cumpas (Son.)
18 km NE de Nacozari
30°16.473′
-109°50.070′
930 m
Centruroides noxius
Pantanal (Nay.)
Pantanal
21°25′24.42′′
-104°50′47.89′′
921 m
Centruroides huichol
Nayarit
–
–
–
–
Centruroides infamatus115scrp
Guanajuato (Gto.)
Guanajuato
–
–
–
Centruroides ornatus2LP1822
Tandamangapio (Mich.)
Los Tabanos
19.9749°
– 102.84226°
223 m
Centruroides ornatus3 2003
Michoacán
–
–
–
–
*Centruroides tecomanus11 25scrp
Comala (Col.)
–
–
–
–
*Centruroides tecomanus12 2007
Michoacán
–
–
–
–
Mesomexovis occidentalis4 LP 7056
Acapulco (Gro.)
Cumbres de Llano Largo
16°49.505
-99°49.9990
317 m
Table 2. Continued
Species
Municipality
Locality
Latitude
Longitude
Elevation
Mesomexovis spadix4LP6373
León (Gto.)
San Antonio de Padua
20°34.5170
-100°57.217
–
Mesomexovis subcristatus4LP 2049
Tehuacán (Pue.)
Tehuacán, 2 km east
18°24.0020
-97°22.8670
1435 m
Thorellius cristimanus4LP 5325
Álvaro Obregón (Mich.)
Álvaro Obregón
19°02.3100
-102°58.405
462 m
Thorellius cristimanus4LP 6551
Coquimatlán (Col.)
Road to Coquimatlán, km 71
19°06.7750
-103°51.1850
336 m
Thorellius intrepidus4LP 6377
Comala (Col.)
Comala
19°19.000
-103°45.0000
–
Thorellius intrepidus4LP 6379
Colima (Col.)
Los Ortices
19°06.0468
-103°44.0226
343 m
Vaejovis carolinianus4LP 1576
South Carolina
–
–
–
–
Vaejovis pequeno4LP 6308
Soyopa (Son.)
Sierra El Encinal, 9 km from crossroad on Highway Mex 16 to El Encinal
28°35.4120
-109°27.1480
380 m
Vaejovis rossmani4LP 2027
Hidalgo (Tams.)
Conrado Castillo
23°56.01735
-99°28.04817
–
*Genetically differentiated species.
We also included 8 species from outside the state to test the presence of C. elegans, C. infamatus, and C. limpidus reported previously in the literature (Ponce-Saavedra et al., 2016); C. noxius and C. suffusus for comparative purposes and samples of C. tecomanus and C. ornatus from GenBank with a total of 31 specimens of the genus Centruroides within 4 species groups. Unlike buthids, we obtained 12 samples of vaejovids to include in this analysis. To evaluate the identity of vaejovids, we used the BLAST® suite (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to search for similar sequences deposited in the nucleotide collection database at NCBI. Thorellius has been revised recently (González-Santillán & Prendini, 2018). Therefore, several DNA sequences are available, and fewer sequences of Mesomexovis sp. and Vaejovis sp. were available in GenBank, as they are still unrevised. Using the genetic markers as queries, we obtained 10 additional samples. The total number of taxonomic specimens used for these analyses was 53 (Tables 2, 4).
Genomic DNA was extracted from the legs or pedipalp of specimens using Qiagen Dneasy/trisol method Tissue Kits or a DNAzol Genomic DNA isolation Reagent kit (Molecular Research Center INC, Cincinnati, Oh). We amplified 3 mitochondrial markers, 12S rDNA, 16S rDNA, and the barcode COI, and the nuclear marker 28S rDNA. We performed the Polymerase Chain Reaction with the following thermal profile: an initial denaturation step (3 min at 94°C) followed by 35 cycles including denaturation at 94°C for 30 s, annealing (46-55°C) for 30 s, and extension at 72°C for 30 s, with a final extension step at 72°C for 7 min. The PCR reaction was conducted using PureTaq-Ready-To-Go PCR Beads (GE Healthcare), 2 µl of DNA template, 21 µl of DNA grade H2O, and 1 µl of each direction primer listed in Table 3. We verified PCR products with a 1% agarose-TBE electrophoresis gel stained with CYBR Safe. For purification of the amplified products, we used Ampure Magnetic Beads (Beckman-Coulter) and re-suspended in 40 µl DNA grade water by using a Beckman Coulter Biomek NX 18 robot. Each 8 µl cycle-sequenced reaction mixture included 1 µl of Big Dye, 1 µl of Big Dye Terminating buffer, 1 µl of 3.2 pm primer, and 5 µl of gene amplification product. Cycle-sequenced products were purified with CleanSeq magnetic beads on a Biomex NX robot. Products were re-suspended in EDTA, and 33 µl were processed in an Applied Biosystems, Inc. Prism 3730xl automated DNA sequencer. These products were sequenced with the same primer pairs used for amplification at the Laboratorio de Secuenciación Genómica de la Biodiversidad, at Instituto de Biología and Unidad de Síntesis y Secuenciación de DNA, Instituto de Biotecnología, UNAM. The sequences were edited using Sequencher® version 5.4.6.
Table 3
List of primers used to amplify molecular markers.
Name
Sequence
Reference
12S rDNA
12SAI
AAACTAGGATTAGATACCCTATTAT
Kocher et al. (1989)
12SBI
AAGAGCGACGGGCGATGTGT
Kocher et al. (1989)
16S rDNA
16SA
CGCCTGTTTATCAAAAACAT
Simon et al. (1994)
16SB
CTCCGGTTTGAACTCAGATCA
Simon et al. (1994)
COI
HCO
TAAACTTCAGGGTGACCAAAAAATCA
Folmer et al. (1994)
LCO1
GGTCAACAAATCATAAAGATATTGG
Folmer et al. (1994)
28S rDNA
28SA
GACCCGTCTTGAAGCACG
Nunn et al. (1996)
28SBout
CCCACAGCGCCAGTTCTGCTTACC
Prendini et al. (2005)
Each genetic fragment was aligned separately for all terminals with MAFFT using the online server (https://mafft.cbrc.jp/alignment/server/). Since the number of nucleotides per gene was similar, we used the G-ins-i iterative refinement method, as recommended elsewhere (Katoh et al., 2019; Kuraku et al., 2013), and other parameters were kept default. To select the best fit of the substitution model per partition and conduct the phylogenetic analyses, we used IQ-TREE version 2 (Kalyaanamoorthy et al., 2017; Nguyen et al., 2015) and we estimated branch support with 1,000 replicates of the ultrafast bootstrap (UFBOOT) algorithm (Hoang et al., 2018). Furthermore, each genetic marker was analyzed individually to explore its phylogenetic signal and contribution to the final topology. We conducted concatenated and partitioned analyses, handling all matrices in Mesquite (Maddison & Maddison, 2023). For the COI partition we explored the best codon partition per site, but the results had no effect on the topology. Additionally, to evaluate each marker and nucleotide site within each marker, we calculated the gene (GCF) and site (SCF) concordance factors on the topology that we emphasized in the discussion (Mihn et al., 2020).
Distributional records and maps. We obtained records with geographical coordinates of the scorpion species treated here via the Global Biodiversity Information Facility (GBIF) from the InDRE, responsible in Mexico for epidemiological vigilance (Huerta-Jiménez, 2018), and the records published by Ponce-Saavedra et al. (2015). These records were the basis for the species distribution maps depicted in figures 3 to 6. We used the program QGIS 3.16.6-Hannove (QGIS, 2021) to create the distributional maps. The topological model with the data was from Jarvis et al. (2008), and to draw the political boundaries we used shapefiles obtained from Conabio. The biogeographic regionalization of Mexico into provinces and districts follows Morrone et al. (2017) and Morrone (2019).
Results
The gene fragments that we obtained are listed in Table 4 and the main statistics of the alignment and concatenated matrix are in Table 5. Our topology produced a clade representing members of the family Buthidae and another clade representing Vaejovidae (Fig. 2). Within buthids, the first clade included C. huichol and C. noxius, component species of the bertholdii species group (Ponce-Saavedra & Francke, 2019), supported by 95% UFBOOT, 100% GCF, and 40% SCF. The next clade included C. elegans, C. limpidus, and a putative undescribed species with lower support values of 68%, 100%, and 38%, respectively, including members of the elegans group. Although with low support, the bulk of species appeared in the third clade comprising species within the infamatus group, with C. tecomanus, C. infamatus, C. possanii, C. hirsuticauda, C. ornatus, and 2 putative new species. The last clade included C. suffusus and C. sculpturatus, whichPonce-Saavedra and Francke (2019) circumscribed within the infamatus and the elegans species group, respectively (Fig. 2). However, the overall topology retrieved herein is concordant with the North American clade of the genus Centruroides (Esposito & Prendini, 2019).
Table 4
Mitochondrial genetic markers 16S, COI, and 12S and nuclear 28S information for samples analyzed in the study. Dash (-) symbols indicate unavailable sequences.
Species
NCBI:txid
16S
12S
COI
28S
Centruroides elegans
217897
Cele_30_16S (PP295377)
Cele_30_12S (PP295301)
Cele_30_COI (PP356615)
Cele_30_28S (PP295328)
Cele_31_16S (PP295378)
Cele_31_12S (PP295302)
Cele_31_COI (PP355194)
Cele_31_28S (PP295329)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Centruroides hirsutipalpus
–
Chir_06_16S (PP295353)
Chir_06_12S (PP295277)
Chir_06_COI (PP356614)
Chir_06_28S (PP295310)
Centruroides huichol
2911785
Chui_38_16S (PP295385)
Chui_38_12S (PP295309)
Chui_38_COI (PP356613)
Chui_38_28S (PP295335)
Centruroides infamatus
42200
MF134694
–
MF134798
MF134763
Centruroides limpidus
29941
Clim_34_16S (PP295381)
Clim_34_12S (PP295305)
Clim_34_COI (PP356612)
Clim_34_28S (PP295332)
Centruroides noxius
6878
Cnox_36_16S (PP295383)
Cnox_36_12S (PP295307)
Cnox_36_COI (PP356611)
Cnox_36_28S (PP295333)
Cnox_37_16S (PP295384)
Cnox_37_12S (PP295308)
Cnox_37_COI (PP356610)
Cnox_37_28S (PP295334)
Centruroides ornatus
2338500
Corn_03_16S (PP295350)
Corn_03_12S (PP295274)
Corn_03_COI (PP355195)
Corn_03_28S (PP295324)
Corn_04_16S (PP295351)
Corn_04_12S (PP295275)
Corn_04_COI (PP356609)
Corn_04_28S (PP295325)
KY981895
KY981799
–
KY982086
MK479042
MK478991
MK479195
MK479144
Centruroides possanii
–
Cpos_10_16S (PP295357)
Cpos_10_12S (PP295281)
–
Cpos_10_28S (PP295314)
Cpos_07_16S (PP295354)
Cpos_07_12S (PP295278)
Cpos_07_COI (PP355196)
Cpos_07_28S (PP295312)
Cpos_08_16S (PP295355)
Cpos_08_12S (PP295279)
Cpos_08_COI (PP356608)
Cpos_08_28S PP295313
Cpos_09_16S (PP295356)
Cpos_09_12S (PP295280)
Cpos_09_COI (PP356607)
Cpos_09_28S (PP295323)
Centruroides sculpturatus
218467
Cscu_35_16S (PP295382)
Cscu_35_12S (PP295306)
Cscu_35_COI (PP356606)
Cscu_35_28S (PP295331)
Centruroides sp. 1
3103037
Csp1_23_16S (PP295370)
Csp1_23_12S (PP295294)
Csp1_23_COI (PP356604)
Csp1_23_28S (PP295311)
Centruroides sp. 2
Csp2_11_16S (PP295358)
Csp2_11_12S (PP295282)
Csp2_11_COI (PP356603)
Csp2_11_28S (PP295327)
Csp_14_16S (PP295361)
Csp_14_12S (PP295285)
Csp_14_COI (PP356605)
Csp_14_28S (PP295326)
Centruroides suffusus
6881
Csu_33_16S (PP295380)
Csu_33_12S (PP295304)
–
Csu_33_28S (PP295330)
Centruroides tecomanus1
1028682
Cte1_12_16S (PP295359)
Cte1_12_12S (PP295283)
Cte1_12_COI (PP356602)
Cte1_12_28S (PP295315)
Cte1_17_16S (PP295364)
Cte1_17_12S (PP295288)
Cte1_17_COI (PP355197)
Cte1_17_28S (PP295320)
Cte1_18_16S (PP295365)
Cte1_18_12S (PP295289)
Cte1_18_COI (PP356601)
Cte1_18_28S (PP295318)
Centruroides tecomanus2
Cte2_13_16S (PP295360)
Cte2_13_12S (PP295284)
Cte2_13_COI (PP356600)
Cte2_13_28S (PP295316)
Cte2_15_16S (PP295362)
Cte2_15_12S (PP295286)
Cte2_15_COI (PP355198)
Cte2_15_28S (PP295317)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Cte2_19_16S (PP295366)
Cte2_19_12S (PP295290)
Cte2_19_COI (PP356599)
Cte2_19_28S (PP295319)
Cte2_20_16S (PP295367)
Cte2_20_12S (PP295291)
Cte2_20_COI (PP356598)
Cte2_20_28S (PP295321)
Cte2_21_16S (PP295368)
Cte2_21_12S (PP295292)
Cte2_21_COI (PP356597)
Cte2_21_28S (PP295322)
Centruroides tecomanus
MF134695
–
MF134799
MF134757
MK479053
MK479002
MK479206
MK479156
Mesomexovis sp.
–
Mesp_22_16S (PP295369)
Mesp_22_12S (PP295293)
Mesp_22_COI
Mesp_22_28S (PP295337)
Mesomexovis occidentalis
1532992
KM274362
KM274216
KM274800
–
Mesomexovis spadix
1532994
KM274221
KM274367
KM274805
KM274659
Mesomexovis subcristatus
1532995
KM274368
KM274222
KM274806
KM274660
Thorellius cristimanus
1533000
Tcri_01_16S (PP295348)
Tcri_01_12S (PP295272)
–
Tcri_01_28S (PP295338)
Tcri_16_16S (PP295363)
Tcri_16_12S (PP295287)
–
Tcri_16_28S (PP295336)
Tcri_02_16S (PP295349)
Tcri_02_12S (PP295273)
–
Tcri_02_28S (PP295339)
Tcri_05_16S (PP295352)
Tcri_05_12S (PP295276)
–
Tcri_05_28S (PP295340)
KM274420
KM274274
KM274858
KM274712
KM274422
KM274276
KM274860
KM274714
Thorellius intrepidus
1533001
Tint_24__16S (PP295371)
Tint_24_12S (PP295295)
Tint_24_COI (PP355193)
Tint_24_28S (PP295341)
Tint_25_16S (PP295372)
Tint_25_12S (PP295296)
Tint_25_COI (PP356616)
Tint_25_28S (PP295342)
Tint_26_16S (PP295373)
Tint_26_12S (PP295297)
Tint_26_COI (PP356617)
Tint_26_28S (PP295343)
Tint_27_16S (PP295374)
Tint_27_12S (PP295298)
Tint_27_COI (PP355192)
Tint_27_28S (PP295344)
Tint_28_16S (PP295375)
Tint_28_12S (PP295299)
Tint_28_COI (PP356618)
Tint_28_28S (PP295345)
Tint_29_16S (PP295376)
Tint_29_12S (PP295300)
Tint_29_COI (PP356619)
Tint_29_28S (PP295346)
KM274424
KM274278
KM274862
–
KM274425
KM274279
KM274863
KM274717
Vaejovis sp.
–
Vasp_32_16S (PP295379)
Vasp_32_12S (PP295303)
Vasp_32_COI (PP356620)
Vasp_32_28S (PP295347)
Vaejovis carolinianus
33322
KM274289
KM274143
KM274727
KM274581
Vaejovis pequeno
1532951
KM274293
KM274147
KM274731
KM274585
Vaejovis rossmani
1532952
KM274294
KM274148
KM274732
KM274586
Table 5
Main statistics of site information per alignment, including parsimony informative sites, (P info) and the concatenated (Conca) alignment of all partitions in a final matrix. The concatenated alignment had 6% missing data.
Partitions
Terminals/ nucleotide
Site information per alignment
Substitution
P info
Invariable
Unique
Constant
Model
16S
53/523
209
285
258
285
K3Pu+F+I+G4
28S
52/538
39
494
96
494
K2P+I
12S
51/393
153
210
212
210
TPM2+F+G4
COI
45/679
220
420
220
420
TIM2+F+G4
Conca
53/2133
621
1,409
786
1,409
simultaneous
Figure 2. Phylogenetic tree from an analysis of 4 concatenated genetic markers, 3 mitochondrial (12S, 16S, and COI) and 1 nuclear (28S). The topology shows families, subfamilies, and species groups. Numbers beside nodes indicate ultrafast bootstrap/genetic concordance factor/site concordance factor. Colored species represent the 11 species found in our collection within the study area and samples grouping with them distributed inside or outside Colima. Scorpion photos: upper right, Centruroides tecomanus from El Palapo, adult male; middle, Centruroides hirsutipalpus from Sierra de Minatitlán, adult female; lower left, Thorellius intrepidus from El Palapo, adult male.
The Vaejovid clade received full support, except SCF (60%), which included 3 genera: Vaejovis, Mesomexovis, and Thorellius (Fig. 2). The genera occurred in a topology that overall resembles that of González-Santillán and Prendini (2013, 2015); while Vaejovis is a genus within Vaejovinae, Mesomexovis and Thorellius are part of the Syntropinae subfamily. Within Syntropinae, only Thorellius was monophyletic, and T. intrepidus received full support. Additionally, we identified 2 putative new species belonging to the genera Vaejovis and Mesomexovis.
Distributional maps and species geographical ranges. We mined 6,965 records of Mexican scorpions from the InDRE database but retained only 1,000 of the species treated herein. We filtered the records using previously published identified species by specialists and the distribution of the species cited for Colima (González-Santillán & Prendini, 2013, 2018; Lourenço & Sissom, 2000; Ponce-Saavedra et al., 2016; Sissom, 2000).
Figure 3. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides elegans (circles), records north of Sierra de Manantlán may be misidentifications; B, distribution of Centruroides tecomanus (circles), records in Guerrero (GRO), Guanajuato (GTO), Jalisco (JAL), and Nayarit (NAY) may be misidentifications; Centruroides tecomanus 1 green cross, and Centruroides tecomanus 2 pink crosses. State abbreviations: AUG, Aguascalientes; COL, Colima; MEX, Estado de México; QRO, Querétaro; SLP, San Luis Potosí; ZAC, Zacatecas.
Figure 4. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database and for B additional records from Ponce-Saavedra et al. (2015). A, Distribution of Centruroides infamatus (circles); B, distribution of Centruroides ornatus (circles), our sample in Colima (cross).
Figure 5. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides hirsutipalpus (red cross), Centruroides sp. 1 (blue cross), Centruroides sp. 2 (pink cross), and Centruroides possanii (green crosses); B, distribution of Mesomexovis aff. occidentalis sequences in this work and a sample from Chamela Jalisco recorded by González-Santillán (2004), conspecific with our samples in green crosses, Vaejovis sp. (mexicanus group) (pink cross).
The distribution of Centruroides species appears to be restricted by physiographical elements. Centruroides elegans and C. tecomanus are restricted to the coastal lands of Jalisco and Colima, respectively (Fig. 3). The Sierra Madre del Sur appears to be the main barrier to the north of their distribution. The records of these species overlap broadly with a visible gap created by the Sierra de Minatitlán (Fig. 3B) either by subsampling or by an effective geographic barrier.
Centruroides ornatus and C. infamatus, on the other hand, are restricted to the Transmexican Volcanic Belt (TVB) (Fig. 4). While the former species has an entire distribution within this province (Fig. 4B), the latter seems to be distributed in patches, one to the north, along the border of the Chihuahuan desert and the TVB, and the second to the south, between the Balsas Depression and the TVB (Fig. 4A).
Centruroides possanii appears to be a microendemic scorpion species component of the fauna restricted to a massive karstic lone mountain, Cerro Grande (González-Santillán et al., 2019). Two putative undescribed species of Centruroides occupy the extreme east and west of the coastal line of Colima, and C. hirsutipalpus appears restricted to the Sierra Minatitlán (Fig. 5A).
Of the vaejovid species, thus far, the only records for Mesomexovis sp. are in Colima, albeit there is one conspecific record in Chamela, Jalisco (González-Santillán, 2004). The putatively undescribed species of Vaejovis appears restricted to Cerro Grande (Fig. 5B), like Centruroides possanii, potentially another microendemic species. Thorellius intrepidus and T. cristimanus are widely distributed within Colima and across the TVB, the Balsas Depression, and the Sierra Madre del Sur (Fig. 6).
Discussion
Of the 2 families found in Colima, Buthidae comprise 7 species of Centruroides,becoming the most diverse (Table 1, Fig. 2). Our multigene analyses suggest that the previously recorded species C. elegans, C. infamatus, and C. limpidus reported by Ponce-Saavedra et al. (2016) might not be part of the scorpion fauna of Colima, as we demonstrate in the following sections.
Figure 6. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Thorellius intrepidus greensquares, our samples in Colima pink crosses; B, distribution of Thorellius cristimanus greensquares, our samples in Colima pink crosses.
Centruroides elegans and C. limpidus were not found in Colima. Centruroides elegans has an obscure taxonomic history. Firstly, its original description is too general and never indicated a precise type locality, but “Mexico” (Fet & Lowe, 2000). Secondly, although some taxonomic works clarified its former subspecies, the nominal taxon identity of C. elegans remains ambiguous. While Lourenço and Sissom (2000) suggested that this species is distributed in Jalisco, later, González-Santillán (2004) concluded that its distributional limits have never been defined with precision. The 2 exemplars of C. elegans collected in Chamela, Jalisco, grouped with members of the elegans group, sisters to C. limpidus from Iguala, Guerrero, and these in turn, were sister to our 2 exemplars identified as Centruroides new sp. 2 11 and 14 from La Central, in the municipality of Manzanillo (Figs. 2). The distributional data from InDRE of C. elegans is limited right at the northern border of Colima, except for 9 records away from that geographic barrier (Fig. 3A), which we hypothesized as a misidentification due to their position outside the Pacific coastline. Our fieldwork produced no sample conspecific to C. elegans but did produce other genetically distant exemplars. Until denser sampling along the northern border of Colima is conducted, we conclude that C. elegans and by corollary C. limpidus might not be part of the scorpion fauna distributed in Colima as suggested by Ponce-Saavedra and Francke (2013) and Ponce-Saavedra et al. (2016).
Centruroides infamatus may not be part of the Colima scorpion fauna. Centruroides infamatus is another obscure taxon with an ambiguous distributional pattern from published data. Despite its inclusion in phylogenetic analyses (Quijano-Ravel et al., 2019; Towler et al., 2001), its taxonomic circumscription and distribution have never been clarified. Among other problems, the original description indicates “type locality unknown” (Fet & Lowe, 2000). Hoffmann (1932) studied the scorpions from Mexico and realized that the specimens from Michoacán agreed with the original description of the species and proposed that its distribution was expanded to Central Mexico from the Pacific Coast in Sinaloa and Colima to Guanajuato. We included 2 exemplars of C. infamatus, 1 from León, Guanajuato (C. infamatus 15 scrp), and 1 terminal from Tandamangapio municipality, Michoacán (C. infamatus LP1822; Esposito et al., 2018). However, the terminal LP1822 grouped with exemplars of C. ornatus (Fig. 2) and Tandamangapio is approximately 16 km from Sahuayo and Jiquilpan, 2 localities recorded in the redescription of C. ornatus (Ponce-Saavedra et al., 2015).
We plotted 612 records from the InDRE database of C. infamatus to compare the distribution of C. ornatus (Fig. 4A). Although northern Michoacán is almost exclusively occupied by C. ornatus, Jalisco and Michoacán present an overlap with C. infamatus (Fig. 4). Additionally, the InDRE database contains records of C. infamatus from Oaxaca, Puebla, Nayarit, Durango, and Sinaloa (not shown), most likely occupied by other species, implying that most records are misidentifications. The species that morphologically could be confused with C. infamatus in the states of Oaxaca and Puebla are C. baergi, C. nigrovariatus, or C. rodolfoi because the overall base color, body size and carapace pigmentation are similar among the species. The same morphological features are present in specimens from Nayarit and Durango, however, the identity of the populations in these two states have never been studied with a molecular approach and may represent a geographically distinct species.
Nevertheless, recently Ponce-Saavedra et al. (2022) described C. baldazoi for Sinaloa, related morphologically with C. infamatus, potentially a species with which it can be mistaken. Although Ponce-Saavedra and collaborators included C. infamatus in the distribution of Sinaloa and Colima, they failed to indicate precise localities. In summary, the distribution of C. infamatus remains unsolved, attested by the wide distribution reported in the InDRE database. Thus, the infamatus species complex requires comprehensive molecular analyses to delimit its taxonomic circumscription and geographical distribution. On the other hand, our samples of C. ornatus 3 and 4 grouped with C. ornatus LP1822 and with C. ornatus 2003 from Morelia, Michoacán (Esposito & Prendini, 2019), forming a monophyletic group (Fig. 2). Since the samples LP1822 and 2003 lie within the area of C. ornatus proposed by Ponce-Saavedra et al. (2015), we concluded that our exemplars are conspecific with C. ornatus and expanded the distribution southwards, drawing a distributional border in northern Colima (Fig. 4B).
In conclusion, we propose that C. infamatus might not be present in Colima, but further sampling and analyses are needed. Despite the morphological similarity between C. infamatus and C. ornatus, our molecular analysis indicates genetic differences and may present a distinctive distributional pattern yet to be drawn with more samples.
Centruroides tecomanus is a species complex. Due to their morphological similarity with C. limpidus, C. tecomanus was described as C. limpidus tecomanus (Hoffmann, 1932), and the author delimited C. tecomanus distribution to the lowlands of Colima and surmised that its distribution extended to the south along the coastline of Michoacán. But, most importantly, Hoffmann mentioned that on the northern coastline, C. elegans substitutes C. tecomanus. In contrast, the distribution recorded in the InDRE database for C. tecomanus implies that these 2 species present a wide area of sympatry on the coast of Jalisco (Fig. 3).
Ponce-Saavedra et al. (2009) proposed C. tecomanus as a “bona fide species” and assumed its distribution includes the coastline of Michoacán, following Hoffmann (1932). In their molecular and morphological analyses, the authors never included exemplars from Tecomán, Colima, the type locality for the species, assuming that the specimens from Michoacán were conspecific with the populations of Tecomán. Furthermore, Quijano-Ravell et al. (2010) extended the distribution of C. tecomanus to 4 localities in Guerrero and a similar number in Jalisco in a montane area.
The phylogenetic tree presented here substantiates 2 clades of C. tecomanus within the monophyletic infamatus species group (Fig. 2). We hypothesize that these clades may represent potentially distinct sympatric species, emphasizing that our topology obtained full support for these clades with multiple samples. Centruroides tecomanus2 appears to be more common and widely distributed in Colima, whereas C. tecomanus1 is less common, with only 2 localities grouped with Centruroides tecomanus 2007 from Michoacán and from the municipality of Comala, Colima, Centruroides tecomanus (Fig. 3B). Our results suggest that the populations distributed in Michoacán may represent a cryptic, undescribed species; consequently, the exemplars reported in Guerrero (Quijano-Ravell et al., 2010) are unlikely to be conspecific with C. tecomanus. The authors’ discovery of new populations in Jalisco and Guerrero were based entirely on their analyses of morphological characters, precisely the most common way of confusing cryptic species.
Considering that the montane border between Colima and Michoacán is occupied by Centruroides romeroi Quijano-Ravell, de Armas, Francke, Ponce-Saavedra, 2019, it is fair to assume that C. tecomanus1 inhabits coastal ranges following the coastline of Michoacán to the Lázaro Cárdenas delta, where it may be substituted by Centruroides bonito Quijano-Ravell, Teruel, Ponce-Saavedra, 2016 or even other undescribed species. The Balsas River has been proposed to be a geographical barrier for several epigean arachnids, such as Amblypygi and Theraphosidae (Mendoza & Francke, 2017; Schramm et al., 2021) and for small mammals (Ruiz-Vega et al., 2018)
Noteworthy is the locality La Central, right on the border between Colima and Jalisco, a few kilometers southeast of the Marabasco River (Fig. 1A), where we collected 3 putatively different species of Centruroides, the 2 morphotypes of C. tecomanus and Centruroides sp. 2, retrieved within the elegans group (Fig. 2). These findings illustrate the complicated patterns of diversification within this buthid genus in Colima. From a taxonomic perspective, if the identity of C. tecomanus is to be clarified, it is now imperative to analyze molecularly exemplars from Tecomán, Colima, which is the type locality of this species.
Thorellius species exhibit a more restricted distribution in Colima. Thorellius intrepidus and T. cristimanus are widely distributed in several states of the Pacific Lowlands (Fig. 6). However, in a recent revision of the genus, 2 species were described, Thorellius wixarika González-Santillán and Prendini, 2018 and Thorellius tekuani González-Santillán and Prendini, 2018 (Fig. 3 of González-Santillán and Prendini, 2018), that are relevant to these analyses. The former occupies the northwestern territory in Nayarit and Jalisco,whereas the latter inhabits the Balsas Depression of Estado de México, Guerrero, and Michoacán. This observation suggests that the InDRE database contains several misidentifications of both T. intrepidus and T. cristimanus. In fact, the T. wixarika and T. tekuani description was in 2018, and the InDRE database is several years older, hence the complete absence of records. Thus, T. intrepidus inhabits Aguascalientes, Colima, Guanajuato, Jalisco, and Michoacán (Fig. 6A); and T. cristimanus, Colima and Jalisco (González-Santillán & Prendini, 2018) (Fig. 6B). From an ecological point of view, we noticed that the InDRE database only has records with elevations below 700 m, implying that these species prefer tropical to subtropical climates in Colima.
González-Santillán (2004) reported Mesomexovis aff. occidentalis as a putative new species for the Biological Station Chamela, Jalisco. Furthermore, González-Santillán and Prendini (2015) conducted a phylogenetic analysis using morphology and mitochondrial and nuclear DNA, resulting in a topology that suggested that this species was not conspecific to Mesomexovis occidentalis, as Williams (1986)identified it. López-Granados (2019) found further morphological evidence to separate this species, but the evidence was never published. The importance of that work is that, for the first time, exemplars of Mesomexovis from Colima that were conspecific with our samples were included in a phylogenetic analysis. Once more, we demonstrate that Mesomexovis sp. 22 is not conspecific with M. occidentalis using molecular evidence, and it requires a formal separation and description (Fig. 2). Unlike Thorellius species in Colima, Mesomexovis is a widespread species inhabiting tropical to montane habitats with a wider range of elevation (Fig. 5B).
The mexicanus group was recently revisited in a monograph with the delimitation of other species groups and the description of 5 new species (Contreras-Félix & Francke, 2019). The paucity of sequences for the mexicanus group in the NCBI database only permitted retrieval of loci for Vaejovis rossmani LP 2027, a species inhabiting the Sierra Madre Oriental in the states of Tamaulipas and Nuevo León and treated in the Contreras-Félix and Francke (2019) monograph. Our analysis retrieved Vaejovis sp. 32 and Vaejovis rossmani LP 2027 together, which suggests membership of the mexicanus group of Vaejovis. Furthermore, Vaejovis sp. 32also matches the morphological diagnosis presented in Contreras-Felix and Francke (2019). Contreras-Félix et al. (2023) recorded Vaejovis santibagnezi Contreras-Felix and Francke, 2019 in Cerro Grande, where we collected our samples (Fig. 5B). However, the authors failed to present morphological evidence to justify this conclusion. We compared our specimens with the geographically closest V. monticola deposited in the CNAN and with V. santibagnezi and found significant morphological differences. Currently, we are preparing a contribution where we propose the description of a new species. Following this idea, we are inclined to think that, like C. possanii, Vaejovis sp. 32 is also microendemic because of its limitation to disperse throughout tropical valleys from the Cerro Grande massif to other mountain ranges. Finally, we submit that such an evident geographical barrier may apply to several epigean, non-volant arthropods such as the scorpions.
Finally, 2 species absent in our fieldwork are Konetontli ilitchi González-Santillán and Prendini 2015 and Vaejovis janssi Williams, 1980. The latter is endemic to the Socorro Island, part of the Revillagigedo Archipelago (Williams, 1980), and K. ilitchi has been found only inside a cave in the vicinity of Coquimatlán, Colima (González-Santillán & Prendini, 2015). These 2 features in the distribution and biology of the species hindered the possibility of collecting and studying them. Future fieldwork may illuminate the taxonomy and distribution of these elusive species. Finally, Vaejovis monticola, another high elevation dweller of the mexicanus species group (Contreras-Felix & Francke 2019) was absent in our collecting trips. Although its type locality cited by Contreras-Felix and Francke (2019) indicates: “Jalisco, northern side of Nevado de Colima” we submit that it may not be on the southern side in Colima, but extensive fieldwork is needed to confirm this hypothesis.
Biogeographical and ecological considerations
There is strong evidence that the ancestors of the North American Centruroides originated in Gondwana and have dispersed to and diversified in that territory via land bridges, vicariance, and rafting over the Atlantic Ocean, over 50 to 20 Mya (Esposito & Prendini, 2019). However, the movement of this genus towards boreal latitudes within North America is not well understood. Although one could suppose that the colonization of such territories appears to be in pulses at different periods due to the overlap in the distribution of the elegans and infamatus species groups, it is beyond the scope of our study to establish a coherent explanation; besides, our data are incomplete to that end. One startling discovery is that the Colima scorpiofauna comprises 7 distinctive, at least genetically, species of Centruroides (Fig. 2). What biotic, abiotic, or ecological factors produce such species diversity? Is it a combination of all these factors? Is it a case of sympatric speciation? Morphologically, it is sometimes difficult to distinguish some of these species. For instance, the 3 species found in La Central were identified initially as C. tecomanus because they exhibit no morphological variation, at least in traditional characters shown in identification keys (Ponce-Saavedra et al., 2016). It is fundamental to conduct morphological studies with this scorpion community to obtain independent information to precisely delimit these species. Unlike other scorpion groups, Centruroides exhibit conservative morphology. For instance, if we compared Centruroides species to species of the Syntropinae subfamily, we find that syntropines are diverse in ecomorphotypic adaptations that promoted morphological diversification (González-Santillán & Prendini, 2013), while in Centruroides, the persistent errant lifestyle maintains morphology without significant variation. It is astonishing to observe, for instance, C. tecomanus living at sea level in tropical deciduous forests and compare it to C. possanii inhabiting pine-oak forests above 2,000 m elevation; as though, the same “Centruroides bauplan” can survive diametrically different environments following a persistent lifestyle. This final assertion leads us to think that the adaptative changes are in the physiology rather than in the morphology of Centruroides.
Thorellius and Mesomexovis (Syntropinae) are substrate-specialists, since the body plan, more robust and armed with setae and spinules on the legs, allow these species to scrape and dig galleries on clayey, fine-grained substrates (González-Santillán & Prendini, 2013, 2018). Such adaptations come with a drawback because the kind of soil restricts the presence of these scorpions; for instance, rocky or shallow soils represent unhospitable habitats for these species. Like other species in the mexicanus group, Vaejovis sp. 32 and Vaejovis monticola Sissom, 1989 are inhabitants of montane habitats within the intricate topography of Colima.
In summary, the scorpion assemblage distributed in Colima includes errant species such as Centruroides without exaggerated adaptations; pelophilous/lapidicolous species such as Thorellius and Mesomexovis, powerful diggers, commonly found underneath rocks or other debris; and scorpions that can be found inside the leaf litter or under nooks of rocks or trees, such as Vaejovis sp. However, it is enigmatic why the Diplocentridae family, well-represented in Mexico, has not been found in Colima. Only in Colima, out of the 3 relatively small states in Mexico, the other 2 being Morelos (Santibáñez-López et al., 2011) and Aguascalientes (Chávez-Samayoa et al., 2022), where similar scorpiofaunistic surveys have been conducted, the genus Diplocentrus is absent. From a biogeographical point of view, if this family is absent in Colima, it is not trivial and requires further investigation.
As a final remark, we hope we have accomplished our aim of urging arachnologists to examine the benefits of local inventories and the potential use of barcodes at the forefront of discovering and documenting local arachnid diversity.
Medical and pharmacological relevance of some scorpions of Colima
From a medical point of view, it is of utmost importance to identify the species of scorpions in an area and to be able to distinguish hazardous from harmless species. Colima is one of the states with the highest incidence rates of mortality and morbidity caused by intoxication by scorpion sting (ISS) (Chowell et al., 2006; González-Santillán & Possani, 2018). By the 1940s, Colima was at the forefront of ISS and deaths in Mexico. However, with the introduction of a safe and effective antivenom in the 1970s, mortality has diminished, although morbidity is still high (González-Santillán & Possani, 2018).
Due to the medical relevance of scorpionism in Mexico, the venom of some buthid scorpions distributed in Colima has been studied extensively (Table 6). We know that mammalian sodium scorpion toxins (NaScTx) are the chief and sometimes most abundant compound responsible for the toxicity of scorpion venom, although potassium, chlorine, and calcium-gated channels, among others, are also affected by toxic peptides (Cid-Uribe et al., 2019; González-Santillán & Possani 2018). Until now, C. hirsutipalpus, C. ornatus, C. possanii, and C. tecomanus have been studied with a proteomic or transcriptomic approach, demonstrating that these venoms are powerful enough to decimate humans (García-Guerrero et al., 2020; García-Villalvazo et al., 2023, Valdez-Velázquez et al., 2016, 2018). Centruroides tecomanus venom contains at least 13 mammalian toxic compounds (Valdez-Velázquez et al., 2016), of which Ct1a is the main mammalian sodium scorpion toxin component, with a molecular weight of 7,591 Da (Martin et al., 1988). The proteomic analysis of C. ornatus toxic components identified 3 major mammalian NaScTx, CO1, CO2, and CO3, with molecular weights of 7,561.2, 7,614.3, and 7,774.9 Da, respectively (García-Guerrero et al., 2020). On the other hand, the published mass fingerprint of C. hirsutipalpus venom has 4 compounds with similar molecular weight to C. tecomanus peptides, suggesting that these species may have similar or identical NaScTx (Valdez-Velázquez et al., 2018). One of the shared compounds found in C. hirsutipalpus corresponds to the molecular weight of Ct1a (Valdez-Velázquez et al., 2013), which means that Ct1a could be responsible for, or at least contribute to, the high toxicity of this species. Although the molecular components with a weight corresponding to the toxic peptides CO1, CO2, and CO3 are not present in C. hirsutipalpus venom, the mass fingerprint of C. tecomanus reported a peptide with similar molecular weight to CO2 toxin present in C. ornatus (Valdez-Velázquez et al., 2013). Our phylogenetic results show that these species belong to the infamatus group, and it is notable that the molecular weight among some toxins is similar. This observation opens the possibility that the toxin diversity of the mammalian NaScTx may have been inherited from a common ancestor. We realize, however, the need for a more in-depth study to unveil the relationship among these toxic peptides to test such a hypothesis. In contrast, a C. possanii crude venom proteomic study resulted in the identification of 18 NaScTx, of which, CpoNatBet09 was identical to Cll2b and Cii1 from C. limpidus and C. infamatus, respectively (Valdez-Velázquez et al., 2013). This example allows us to present the counterpart of the previous one. Centruroides limpidus in our topology grouped with members of the elegans group and C. possanii within the infamatus group (Fig. 2). We can conclude that the exact sequence match between CpoNatBet09 and Cll2b is most likely due to a selective force that produced convergent evolution. Alternatively, this is a deeper inheritance within the ancestors of the infamatus and elegans group.
The comparisons and hypotheses about venom similarities and species assume that all scorpions used to conduct the experiments to obtain the crude venom were correctly identified. However, the discovery of putative species of Centruroides might question some of these results, particularly those of C. tecomanus. The first author has been collaborating to identify most samples of species used in some of the recently published investigation on venom research (Cid-Uribe et al., 2019; García-Guerrero et al., 2020; Romero-Gutiérrez et al., 2017, Valdez-Velázquez et al., 2018, among others). And in some cases, voucher specimens have been deposited in the CNAN, therefore the taxonomic identity for C. hirsutipalpus, C. ornatus, C. possanii can be readily corroborated.
Table 6
Scorpion species with reported LD50 tested on mice at μg/20g mouse weight scale with the named, most abundant, and likely responsible for high toxicity sodium toxins found in the scorpion species of Colima.
Species
LD50
NaScTx
Autor
Centruroides hirsutipalpus Ponce-Saavedra and Francke, 2009
11.7 ± 1.9
26 β-NaTx and 5 α-NaTx (proteome) 71 β-NaTx and 16 α-NaTx (transcriptome)
Valdez-Velázquez et al. (2013, 2016), Ramírez et al. (1988)
Thorellius cristimanus Pocock, 1902
N/A
10 β-NaTx and 3 α-NaTx (transcriptome)
Romero-Gutiérrez et al. (2017)
Because vaejovid scorpions exhibit lower toxin potency, proteomic studies are uncommon. However, the study of the crude venom of Thorellius cristimanus under a transcriptomic and proteomic approachidentified160 potential venom peptides (Romero-Gutiérrez et al., 2017). The authors found a great diversity of channel toxins targeting potassium and calcium ion-channels, numerous enzymes, and NDBP. Nevertheless, none of these toxins appear to affect mammals to endanger their life. Noteworthy are the NDBP compounds with antimicrobial activity (Almaaytah et al., 2014), which potentially may be a source for developing novel commercial antibiotics. More recently, Ibarra-Vega et al. (2023) reported finding the neurotransmitter serotonin and 2 derived or intermediate indoles of serotonin: N-methylserotonin, and bufotenidine. Although serotonin has been reported in other scorpions and the sea snail genus Conus, the other indoles represent the first report in scorpions. The authors proposed that the 3 components are involved in defense and probably prey submission because they produce extreme pain in the victim. Furthermore, N-methylserotonin and bufotenidine showed a similar affinity for serotonin cellular receptors, implying a role in the effect of this neurotransmitter, and thus, it showed promising medical applications.
Integrating taxonomic, distributional, and in our case venom diversity within local scorpion faunas is an interdisciplinary topic uncommon in the literature (Brito & Borges, 2015; Cao et al., 2014), although this exercise has been done in Mexico before (Santibáñez-López et al., 2015). This multidisciplinary work took advantage of a reciprocal illumination exercise. This short synthesis of the diversity of venoms of scorpion of Colima revealed the potential of toxins to be part of the evidence to identify and delimit species (Schaffrath et al., 2018). Moreover, the synthesis provided a glance of the evolution of venoms not only for the medically prominent species of Centruroides but also for vaejovid scorpion species. From the experimental discipline, we learned that it is paramount to maintain vouchers after biochemical or physiological experiments to track down the original material and keep up with the dynamics of taxonomy, a tool requiring empirical information to permit the identification of species to validate past, present, and future discoveries in other biological disciplines.
Acknowledgments
We thank the University of Colima students and Josué López Granados (Facultad de Ciencias, UNAM) for their assistance in collecting scorpions throughout the state of Colima. JICU thanks Conahcyt for granting a postdoctoral scholarship (512560). Collection permits granted by Semarnat included SGPA/DGVS/12063/15, SGPA/DGVS/02139/2022, and FAUT-0305. Finally, we are grateful to the Associate Editor and two reviewers for improving an early version of the manuscript, particularly to Andrés Ojanguren-Afilastro for his critical reading.
Baldazo-Monsivaiz, J. G., Ponce-Saavedra, J., & Flores-Moreno, M. (2012). Los alacranes (Arachnida: Scorpionida) de importancia médica del estado de Guerrero, México. Dugesiana, 19, 143−150.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., & Canché-Aguilar, I. (2017). Los escorpiones (Arachnida: Scorpiones) del municipio de Chilpancingo de los Bravo, estado de Guerrero, México Entomología Mexicana, 4, 21−27.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., Sánchez-Arriaga, J., López-Flores, M., Reyes-Castelán, A. et al. (2016). Los escorpiones (Arachnida: Scorpiones) del municipio de Taxco de Alarcón, del estado de Guerrero, México. Entomología Mexicana, 3, 75−80.
Bolaños, L. M., Rosenblueth, M., de Lara, A. M., Migueles-Lozano, A., Gil-Aguillón, C., Mateo-Estrada, V. et al. (2019). Cophylogenetic analysis suggests cospeciation between the Scorpion Mycoplasma Clade symbionts and their hosts. Plos One, 14, e0209588 https://doi.org/10.1371/journal.pone.0209588
Brito, G., & Borges, A. (2015). A checklist of the scorpions of Ecuador (Arachnida: Scorpiones), with notes on the distribution and medical significance of some species. Journal of Venomous Animals and Toxins Including Tropical Diseases, 21, 1–23. https://doi.org/10.1186%2Fs40409-015-0023-x
Cao, Z., Di, Z., Wu, Y., & Li, W. (2014). Overview of scorpion species from China and their toxins. Toxins, 6, 796–815. https://doi.org/10.3390/toxins6030796
Cid-Uribe, J. I., Meneses E. P., Batista, C. V. F., Ortiz, E., & Possani, L. D. (2019). Dissecting toxicity: The venom gland transcriptome and the venom proteome of the highly venomous scorpion Centruroides limpidus (Karsch, 1879). Toxins, 247, 1−21. https://doi.org/10.3390/toxins11050247
Contreras-Félix, G. A., & Francke, O. F. (2019). Taxonomic revision of the “mexicanus” group of the genus Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae). Zootaxa, 4596, 1−100. https://doi.org/10.11646/zootaxa.4596.1.1
Contreras-Félix, G. A., del Pozo, O. G., & Navarrete-Heredia, J. L. (2023). A new species of Vaejovis from the mountains of west Mexico (Scorpiones: Vaejovidae). Dugesiana, 30, 229−245. https://doi.org/10.32870/dugesiana.v30i2.7306
Chávez-Samayoa, F., Díaz-Plascencia, J. E., & González-Santillán, E. (2022). Two new species of Vaejovis (Scorpiones: Vaejovidae) belonging to the mexicanus group from Aguascalientes, Mexico, with comments on the homology and function of the hemispermatophore. Zoologischer Anzeiger, 298, 148–169. https://doi.org/10. 1016/j.jcz.2022.04.005
Chowell, G., Díaz-Dueñas, P., Bustos-Saldaña, R., Alemán-Mireles, A., & Fet, V. (2006). Epidemiological and clinical characteristics of scorpionism in Colima, Mexico (2000-2001). Toxicon, 47, 753–758. https://doi.org/10.1016/j.toxicon.2006.02.004
Esposito, L. A., & Prendini, L. (2019). Island ancestors and New World biogeography: a case study from the scorpions (Buthidae: Centruroidinae). Scientific Reports, 9, 3500. https://doi.org/10.1038/s41598-018-33754-8
Fet, V., Graham, M. R., Blagoev, G., Karataş, A., & Karataş, A. (2016). DNA barcoding indicates hidden diversity of Euscorpius (Scorpiones: Euscorpiidae) in Turkey. Euscorpius, 216, 1–13.
Fet, V., Graham, M. R., Webber, M. M., & Blagoev, G. (2014). Two new species of Euscorpius (Scorpiones: Euscorpiidae) from Bulgaria, Serbia, and Greece. Zootaxa, 3894, 83–105. https://doi.org/10.11646/zootaxa.3894.1.7
Fet, V., & Lowe G. (2000). Family Buthidae C. L: Koch, 1837. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 54–286). New York: New York Entomological Society.
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
García-Guerrero, I. A., Cárcamo-Noriega, E., Gómez-Lagunas, F., González-Santillán, E., Zamudio, F. Z., Gurrola, G. B. et al. (2020). Biochemical characterization of the venom from the Mexican scorpion Centruroides ornatus, a dangerous species to humans. Toxicon, 15, 27–38. https://doi.org/10.1016/j.toxicon.2019.11.004
García-Villalvazo, P. E., Jiménez-Vargas, J. M., Lino-López, G. J., Meneses, E. P., Bermúdez-Guzmán, M. J., Barajas-Saucedo, C. E. et al. (2023). Unveiling the protein components of the secretory-venom gland and venom of the scorpion Centruroides possanii (Buthidae) through omic technologies. Toxins, 15, 1–20. https://doi.org/10.3390/toxins15080498
Goodman, A. M., Prendini, L., & Esposito, L. A. (2021). Systematics of the arboreal Neotropical ‘thorellii’ clade of Centruroides bark scorpions (Buthidae) and the efficacy of mini-barcodes for museum specimens. Diversity, 13, 441. https://doi.org/10.3390/d13090441
Goodman, A. M., Prendini, L., Francke, O. F., & Esposito, L. A. (2021). Systematic revision of the arboreal neotropical “thorellii” clade of Centruroides Marx, 1890, bark scorpions (Buthidae C. L. Koch, 1837) with descriptions of six new species. Bulletin of the American Museum of Natural History, 452, 1–92.
González-Santillán, E. (2004). Diversidad, taxonomía y hábitat de alacranes. In A. N. García, & R. Ayala (Eds.), Artrópodos de Chamela (pp. 25–35). México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México.
González-Santillán, E., Galán-Sánchez, M. A., & Valdez-Velázquez, L. L. (2019). A new species of Centruroides (Scorpiones, Buthidae) from Colima, Mexico. Comptes Rendus Biologies, 342, 9–10. https://doi.org/10.1016/j.crvi.2019.10.002
González-Santillán, E., & Possani, L. D. (2018). North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Tropica, 187, 264–274. https://doi.org/10.1016/j.actatropica.2018.08.002
González-Santillán, E., & Prendini, L. (2013). Redefinition and generic revision of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, with descriptions of six new genera. Bulletin of the American Museum of Natural History, 382, 1–71.
González-Santillán, E., & Prendini, L. (2015). Phylogeny of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, based on morphology, mitochondrial and nuclear DNA. Cladistics, 31, 341–405. https://doi.org/10.1111/cla.12091
González-Santillán, E., & Prendini, L. (2018). Systematic revision of the North American Syntropine Vaejovid scorpion genera Balsateres, Kuarapu, and Thorellius, with descriptions of three new species. Bulletin of the American Museum of Natural History, 420, 1–81. https://doi.org/10.1206/0003-0090-420.1.1
Hoang, D. T., Chernomor, O., Haeseler, A. von, Minh, B. Q., & Vinh, L. S. (2018) UFBoot2: Improving the ultrafast bootstrap approximation. Molecular Biology and Evolution, 35, 518–522. https://doi.org/10.1093/molbev/msx281
Hoffmann, C. C. (1932). Monografías para la entomología médica de México. Monografía Núm. 2. Los scorpiones de Mexico. Segunda parte, Buthidae. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, 3, 243–361.
Huerta-Jiménez, H. (2018). Actualización de la Colección de Artrópodos con importancia médica (CAIM), Laboratorio de Entomología, InDRE. Version 1.5. Comisión Nacional para el Conocimiento y uso de la Biodiversidad.
Ibarra-Vega, R., Jiménez-Vargas, J. M., Pineda-Contreras, A., Martínez-Martínez, F. J., Barajas-Saucedo, C. E., García-Ortega, H. et al. (2023). Indolealkylamines in the venom of the scorpion Thorellius intrepidus. Toxicon, 233, 107232. https://doi.org/10.1016/j.toxicon.2023.107232
Jarvis, A., Reuter, H. I., Nelson, A. & Guevara, E. (2008). Hole-filled seamless SRTM data V4, International Centre for Tropical Agriculture (CIAT), available from http://srtm.csi.cgiar.org
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., Haeseler, A. von, & Jermiin, L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nature Methods, 14, 587–589. https://doi.org/10.1038/nmeth.4285
Katoh, K., Rozewicki, J., & Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization, Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Kocher, T. D., Thomas, W. K., Meyer, A., Edwards, S. V., Paabo, S., Villablanca, F. X. et al. (1989). Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proceedings of the National Academy of Sciences, 86, 6196–6200. https://doi.org/10.1073%2Fpnas.86.16.6196
Kuraku, S., Zmasek, C. M., Nishimura, O., & Katoh, K. (2013). aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity, Nucleic Acids Research, 41, W22–W28. https://doi.org/10.1093/nar/gkt389
López-Granados, J. (2019). Análisis filogenético del complejo de especies occidentalis del género Mesomexovis González-Santillán y Prendini, 2013 (Scorpiones: Vaejovidae) basado en evidencia morfológica (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. CDMX.
Lourenço, W. R., & Sissom, W. D. (2000). Scorpiones. In B. J. Llorente, E. González, & N. Papavero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia una síntesis de su conocimiento (pp. 115–135). Ciudad de México: Universidad Nacional Autónoma de México.
Maddison, W. P., & Maddison, D. R. (2023). Mesquite: a modular system for evolutionary analysis. Version 3.81. http://www.mesquiteproject.org
Martin, B. M., Carbone, E., Yatani, A., Brown, A. M., Ramírez, A. N., Gurrola, G. B. et al. (1998). Amino acid sequence and physiological characterization of toxins from the venom of the scorpion Centruroides limpidus tecomanus Hoffmann. Toxicon, 26, 785–94. https://doi.org/10.1016/0041-0101(88)90319-4
Mendoza, J., & Francke, O. (2017). Systematic revision of Brachypelma red-kneed tarantulas (Araneae: Theraphosidae), and the use of DNA barcodes to assist in the identification and conservation of CITES-listed species. Invertebrate Systematics, 31, 157–179. https://doi.org/10.1071/IS16023
Minh, B. Q., Hahn, M. W., & Lanfear, R. (2020). New methods to calculate concordance factors for phylogenomic datasets. Molecular Biology and Evolution, 37, 2727–2733. https://doi.org/10.1093/molbev/msaa106
Morrone, J. (2019). Regionalización biogeográfica y evolución biótica de México: encrucijada de la biodiversidad del Nuevo Mundo. Revista Mexicana de Biodiversidad, 90, e902980. https://doi.org/10.22201/ib.20078706e.2019.90.2980
Morrone, J., Escalante, T., & Rodríguez-Tapia, G. (2017). Mexican biogeographic provinces: map and shapefiles. Zootaxa, 4277, 277–279. https://doi.org/10.11646/zootaxa.4277.2.8
Nguyen, L. T., Schmidt, H. A., Haeseler, A. von, & Minh, B. Q. (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Molecular Biology and Evolution, 32, 268–274. https://doi.org/10.1093/molbev/msu300
Nunn, G. B., Theisen, B. F., Chirstensen, B., & Arctander, P. (1996). Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda. Journal of Molecular Evolution, 42, 211–223. https://doi.org/10.1007/bf02198847
Podnar, M., Grbac, I., Tvrtković, N., Hörweg, C., & Haring, E. (2021). Hidden diversity, ancient divergences, and tentative Pleistocene microrefugia of European scorpions (Euscorpiidae: Euscorpiinae) in the eastern Adriatic region. Journal of Zoological Systematics and Evolutionary Research, 59, 1824–1849. https://doi.org/10.1111/jzs.12562
Ponce-Saavedra, J., & Francke, O. F. (2013). Clave para la identificación de especies de alacranes del género Centruroides Marx 1890 (Scorpiones: Buthidae) en el Centro Occidente de México. Biológicas, 15, 52–62.
Ponce-Saavedra, J. y Francke, O. F. (2019). Una especie nueva de alacrán del género Centruroides (Scorpiones: Buthidae) del noroeste de México. Revista Mexicana de Biodiversidad, 90, e902660. https://doi.org/10.22201/ib.20078706e.2019. 90.2660
Ponce-Saavedra, J., Francke, O. F., Cano-Camacho, H., & Hernández-Calderón, E. (2009). Evidencias morfológicas y moleculares que validan como especie a Centruroidestecomanus (Scorpiones, Buthidae). Revista Mexicana de Biodiversidad, 80, 71–84. https://doi.org/10.22201/ib.20078706e.2009.001.585
Ponce-Saavedra, J., Francke, O. F., Quijano-Ravell, A. F., & Santillán, R. C. (2016). Alacranes (Arachnida: Scorpiones) de importancia para la salud pública de México. Folia Entomológica Mexicana, 2, 45–70.
Ponce-Saavedra, J., Jiménez, M. L., Quijano-Ravell, A. F., Vargas-Sandoval, M., Chamé-Vázquez, D., Palacios-Cardiel, C. et al. (2023). The Fauna of Arachnids in the Anthropocene of Mexico, Chapter 2(pp. 18–39) In R. W. Jones, C. P. Ornelas-García, R. Pineda-López, & F. Álvarez (Eds.), Mexican fauna in the Anthropocene. Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-031-17277-9
Ponce-Saavedra, J., Linares-Guillén, J. W., & Quijano-Ravell, A. F. (2022). Una nueva especie de alacrán del género Centruroides Marx (Scorpiones: Buthidae) de la costa Noroeste de México. Acta Zoológica Mexicana (nueva serie), 38, 1–24. https://doi.org/10.21829/azm.2022.3812517
Ponce-Saavedra, J., Quijano-Ravell, A. F., Teruel, R., & Francke, O. F. (2015). Redescription of Centruroides ornatus Pocock, 1902 (Scorpiones: Buthidae), a montane scorpion from Central Mexico. Revista Ibérica de Aracnología, 27, 81–89.
Prendini, L., Weygoldt, P., & Wheeler, W. C. (2005). Systematics of the Damon variegatus group of African whip spiders (Chelicerata: Amblypygi): evidence from behaviour, morphology and DNA. Organisms Diversity & Evolution, 5, 203–236. https://doi.org/10.1016/j.ode.2004.12.004
Quijano-Ravell, A. F., De Armas, L. F., Francke, O. F., & Ponce-Saavedra, J. (2019). A new species of the genus Centruroides Marx (Scorpiones, Buthidae) from western Michoacán state, México using molecular and morphological evidence. Zookeys, 859, 31–48. https://doi.org/10.3897/zookeys.859.33069
Quijano-Ravell, A. F., Ponce-Saavedra, J., Francke, O. F., & Villaseñor-Ramos. M. A. (2010). Nuevos registros y distribución actualizada de Centruroides tecomanus Hoffmann, 1931 (Scorpiones: Buthidae). Ciencia Nicolaita, 52, 179–189.
QGIS.org. (2021). QGIS Geographic Information System. Open-Source Geospatial Foundation Project. http://qgis.org
Ramírez-Ruiz, J. J., & Bretón-González, M. (2016). Fisiografía y geología. In La Biodiversidad en Colima. Estudio de estado (Eds.) (pp. 25–31). México D.F.: Conabio.
Romero-Gutiérrez, T., Peguero-Sánchez, E., Cevallos, M. A., Batista, C. V. F., Ortiz, E., & Possani L. D. (2017). A deeper examination of Thorellius atrox scorpion venom components with omics technologies. Toxins, 9, 399. https://doi.org/10.3390/toxins9120399
Ruiz-Vega, M. L., Hernández-Canchola, G., & León-Paniagua, L. (2018). Molecular systematics and phylogeography of the endemic Osgood’s deermouse Osgoodomys banderanus (Rodentia: Cricetidae) in the lowlands of western Mexico. Molecular Phylogenetics and Evolution, 127, 867–877. https://doi.org/10.1016/j.ympev.2018.06.034
Santibáñez-López, C. E., Francke, O. F., & Córdova-Athanasiadis, M. (2011). The genus Diplocentrus Peters (Scorpiones: Diplocentridae) in Morelos, Mexico. Revista Ibérica de Aracnología, 19, 3–3.
Santibáñez-López, C. E., Francke, O. F., & Prendini, L. (2014). Phylogeny of the North American scorpion genus Diplocentrus Peters, 1861 (Scorpiones: Diplocentridae) based on morphology, nuclear and mitochondrial DNA. Arthropod Systematics & Phylogeny, 72, 257–279. https://doi.org/10.3897/asp.72.e31789
Santibáñez-López, C., Francke, O., Ureta, C., & Possani, L. (2015). Scorpions from Mexico: from species diversity to venom complexity. Toxins, 8, 1–18. https://doi.org/10.3390/toxins8010002
Schramm, F. D., Valdez-Mondragón, A., & Prendini, L. (2021). Volcanism and palaeoclimate change drive diversification of the world’s largest whip spider (Amblypygi). Molecular Ecology, 30, 2872–2890. https://doi.org/10.1111/mec.15924
Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H., & Flook, P. K. (1994). Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Annals of the Entomological Society of America, 87, 651–700. https://doi.org/10.1093/aesa/87.6.651
Sissom, W. D. (2000). Family Vaejovidae Thorell, 1876. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 503–553). New York: New York Entomological Society.
Towler, W. I. I., Ponce-Saavedra, J., Gantenbein, B., & Fet, V. (2001). Mitochondrial DNA reveals a divergent phylogeny in tropical Centruroides (Scorpiones: Buthidae) from Mexico. Biogeographica, 77, 157–172.
Valdez-Velázquez, L. L., Olamendi-Portugal, T., Restano-Cassulini, R., Zamudio, F. Z., & Possani, L. D. (2018). Mass fingerprinting and electrophysiological analysis of the venom from the scorpion Centruroides hirsutipalpus (Scorpiones: Buthidae). Journal of Venom Animal Toxins Including Tropical Diseases, 24, 17. https://doi.org/10.1186/s40409-018-0154-y
Valdez-Velázquez, L. L., Quintero-Hernández, V., Romero-Gutiérrez, M. T., Coronas, F. I., & Possani, L. D. (2013). Mass fingerprinting of the venom and transcriptome of venom gland of scorpion Centruroides tecomanus. Plos One, 8, e66486. https://doi.org/10.1371/journal.pone.0066486
Valdez-Velázquez L. L., Romero-Gutiérrez M. T., Delgado-Enciso, I., Dobrovinskaya, O., Melnikov, V., Quintero-Hernández, V. et al. (2016). Comprehensive analysis of venom from the scorpion Centruroides tecomanus reveals compounds with antimicrobial, cytotoxic, and insecticidal activities. Toxicon, 118, 95-103. https://doi.org/10.1016/j.toxicon.2016.04.046
Williams, S. C. (1980). Scorpions of Baja California, Mexico and adjacent islands. Occasional Papers of the California Academy of Sciences, 135, 1–127.
Williams, S. C. (1986). A new species of Vaejovis from Jalisco, Mexico (Scorpiones: Vaejovidae). Pan-Pacific Entomologist, 62, 355–358.
Joselin Judith Peña-Herrera a, Yury Glebskiy a, b, *, Teresa de Jesús Hernández-Trejo a, Zenón Cano-Santana a
a Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Ecología y Recursos Naturales, Laboratorio de Interacciones y Procesos Ecológicos, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Universidad Nacional Autónoma de México, Facultad de Ciencias, Posgrado en Ciencias Biológicas, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
Received: 14 September 2023; accepted: 6 March 2024
Abstract
Seed dispersal by animals is a key ecosystemic process in many environments; however, it could be compromised or increased in urban environments due to changes in the landscape, the introduction of exotic species, and human activities. This article aims to evaluate the role of ringtails (Bassariscus astutus) as seed dispersers in an urban-natural gradient during low human activity due to the COVID-19 pandemic. Ringtail feces were collected in 3 sampling sites with different levels of urbanization (ranging from 100 to 5% of natural vegetation), and the seeds germinated in germination chambers. Twenty species of plants were dispersed by ringtails, more than reported in previous studies. More seeds were dispersed in natural (7.1 seeds per g) than urbanized (3.2 seeds per g) areas, but diversity and richness were higher in urbanized areas. This suggests that urban environments have a greater diversity, and it could be attributed to the microenvironments created by urban infrastructure and the exotic plants that are established in the area.
Cacomixtles (Bassariscus astutus) como dispersores de semillas en un gradiente urbano bajo condiciones de baja actividad humana por COVID-19
Resumen
La dispersión de semillas por animales es un proceso clave en muchos ecosistemas, pero este se puede ver comprometido o incrementado en ambientes urbanos debido a cambios en el paisaje, introducción de especies exóticas y actividades humanas. El objetivo de este artículo es evaluar el papel del cacomixtle (Bassariscus astutus) como dispersor de semillas en un gradiente urbano-natural durante un periodo de baja actividad humana debido a la pandemia de COVID-19. Se colectaron excretas de cacomixtles en 3 localidades con diferente grado de urbanización (entre 100 y 5% de vegetación natural) y las semillas fueron germinadas en cámaras de germinación. Se registraron 20 especies de plantas dispersadas, más que lo reportado en estudios previos. Más semillas fueron dispersadas en áreas naturales (7.1 semillas por g) que urbanizadas (3.2 semillas por g), pero la riqueza y diversidad fueron mayores en áreas urbanizadas. Ésto sugiere que la diversidad en ambientes urbanos es mayor, lo cual se puede atribuir a los microambientes formados por la infraestructura urbana y las plantas exóticas establecidas en el área.
Palabras clave: Endozoocoria; Ciudad de México; Opuntia; REPSA; Zoocoria
Introduction
Some animals may provide a key ecosystemic service, acting as seed dispersers, which allow many plant species to effectively move their offspring and maintain plant communities across different environments. An example of those animals is the ringtail (Procyonidae: Bassariscus astutus) which is an omnivorous and opportunistic animal that has shown potential to disperse the seeds of a great number of plant species across many ecosystems (Alexander et al., 1994; Rodríguez-Estrella et al., 2000; Rubalcava-Castillo et al., 2020). However, this important service could be diminished in urban ecosystems, where ringtails are very common (Barja & List, 2006; Swanson et al., 2022). Because urban areas offer new sources of food like anthropogenic waste and a variety of exotic plants, ringtails could disperse fewer seeds or seeds of exotic plants. This is an important concern since urban ecosystems are growing fast and thus becoming important areas for the conservation of species and areas on which we rely to obtain ecosystemic services.
Previous studies suggest that seed dispersal is diminished in cities due to a great number of unsuitable habitats (Cheptou et al., 2008) and barriers that obstruct animal movement and thus seed dispersal (Niu et al., 2018). However, animal-plant networks are persistent in cities (Cruz et al., 2013), and plants that rely on animal dispersal tend to have a more successful regeneration of populations than plants that rely on other strategies for dispersal (Niu et al., 2023). At the same time, most studies show that urban ecosystems tend to be more diverse than natural areas due to the great number of exotic species, microenvironments that can host greater plant diversity, and the fact that cities tend to be built in highly diverse locations (Kühn et al., 2004; Wania et al., 2006). Yet all these studies are performed in urban areas with both human activity and urban infrastructure; therefore, the hypothesis that urban areas are diverse due to the exotic plants and microhabitats and not due to “direct-human” seed dispersal (for example, seeds we throw away as garbage) is yet to be proven.
Particularly for the ringtails, Cisneros-Moreno and Martínez-Coronel (2019) found differences in the urban and rural ringtail diets. They report that in urban environments, ringtails consume 11 plant species, and 9 in rural environments. Other studies also show that ringtails commonly consume human-generated waste from trash cans when it is available (Castellanos et al., 2009; Picazo & García-Collazo, 2019). However, all those studies were made under normal human activity, but if it is reduced, the generation of waste could be diminished, affecting the ringtail diet, which in turn could lead to a change in their role as seed dispersers.
Therefore, this article aims to compare the diversity, abundance, and species of seeds dispersed by ringtails in 3 areas with different levels of urbanization during a time of reduced human activity due to the COVID-19 pandemic.
Materials and methods
This study was performed inside the main campus of the Universidad Nacional Autónoma de Mexico, located in Mexico City, Mexico. The campus contains a well-preserved ecological reserve (Reserva Ecológica del Pedregal de San Ángel; henceforth REPSA) and urban areas such as buildings and roads, all of which are surrounded by the Mexico City (Fig. 1). Therefore, a gradient between urban and natural areas can be found in a relatively small area (730 ha; Zambrano et al., 2016) that otherwise shares all environmental characteristics like mean temperature (18.2°C), precipitation (752 mm), original substrate, and vegetation: xerophitic shrubs (Rzedowski, 1954; SMN, 2023).
An important characteristic of this study is that it was performed under lockdown conditions due to the COVID-19 pandemic. Because of the rapid spreading of the virus, most activities on campus were switched to virtual mode; students took lessons from home; maintenance such as cleaning, gardening, and security were kept to a bare minimum; and all research activities had to be done from home, except for some specific cases, such as this study, that required special permission from the university. Those measures were implemented in March 2020 and began to be gradually lifted in the spring of 2022. Under normal conditions, the campus is visited by 166,474 people, and 70,000 vehicles, and 15 tons of waste (excluding gardening products) are generated (Zambrano et al., 2016). However, as a result of the pandemic, human activity on campus such as driving, waste generation and gardening, among others was minimal for 2 years. This gave us the opportunity to study the ringtail seed dispersal in an urban environment without human presence.
The correct identification of ringtail feces is an essential part of this project, since, in our study site, they could be confused with excretes of opossums (Didelphis virginiana). Previous studies suggest that opossums do not use latrines (Aranda, 2000), however, to ensure that latrines are used exclusively by ringtails, we placed camera traps in front of 8 latrines, and animal interactions with those latrines were recorded.
Figure 1. Map of the study location. Blue dots, Faculty (most urbanized area) latrines; red dots, the Institute latrines (semi-urban area); and green dots, the West core (natural area). The lines represent the 178 m around the latrines in which the percentages of the different types of terrain were calculated.
Three areas separated by at least 800 m were chosen to represent the urban-natural gradient: natural, semi-urban, and urban (Fig. 1). The home ranges of this species in the location are small: between 3 and 9.9 ha (Castellanos & List, 2005); therefore, this separation should ensure independence between the treatments. The level of urbanization was based on the percentage of area covered by natural vegetation, altered vegetation, impermeable areas without buildings (mainly roads and parking lots), and buildings found in a 178 m area (Fig. 1, Table 1) around the sampling points (178 m is the radius of the maximum activity area reported for ringtails in this particular location; Castellanos & List, 2005). The natural area (henceforth the West core) is located inside the west core of the REPSA. Vegetation consists mainly of shrubs, and Opuntia cactus is quite common (Cano-Santana, 1994). The semi-urban area (henceforth Institutes) is located around the humanitarian institutes area and consists of a mosaic of spatially located buildings, parking lots, altered vegetation (grass and some cultivated trees, mostly without fruit) and remnants of natural vegetation that surround this area on all sides (Fig. 1, Flores-Morales, 2023). The urban area (henceforth Faculty) is located inside the faculty of sciences and is dominated by tightly packed buildings divided by impermeable areas and gardens. The vegetation is diverse and includes a small amount of native plants such as Opuntia, but mostly consists of grass and introduced trees some of which have fruits that could be consumed by ringtails (Mendoza-Hernández & Cano-Santana, 2009). During the lockdown, gardens were left mostly unattended, and some alimentary plants began to grow (for example, we encountered several tomato plants with ripe fruits). The natural vegetation is located mostly on the edges of this area (Fig. 1).
Table 1
Amount of terrain coverage in the sampling areas. Impermeable areas were considered all areas covered with concrete but without buildings, mainly roads and parking lots.
Natural vegetation (%)
Altered vegetation (%)
Buildings (%)
Impermeable areas (%)
Total area (ha)
West core
100
0
0
0
34.1 ha
Institutes
49.2
24.2
7
19.6
30.7 ha
Faculty
4.9
35
30.5
29.6
15.4 ha
To estimate the number of seeds dispersed by ringtails, we collected excretes from 13 latrines in each location, on January 18th 2022. Since previous studies report that seeds in our study area commonly have dormancy (Glebskiy, 2019), half of each latrine excretes was placed in plastic bags and half in mesh bags (with 1 × 1 mm openings). Both treatments were left in the field (on the soil and without cover) until April 29th 2022 (when the rains and thus the germination period started). The advantage of the plastic bag treatment is that it allows the seeds to experience the temperature changes that are responsible for ending dormancy in most plants and protects the seeds, but limits some other factors like gas exchange and humidity that could potentially contribute to the end of dormancy (Baskin & Baskin, 1998). On the other hand, the mesh bag allows for a better representation of the natural conditions to which seeds are exposed but is susceptible to losing seeds through the mesh and the addition of new seeds from the environment. Both methods were tested.
At the beginning of the rainy season (April 29th 2022) bags were collected and put to germinate in commercial soil (the soil was sterilized by microwaving, and a control with no excretes was used to test the efficiency of sterilization and possible future seed additions) in a germination chamber (25°C, 16 hours of light, and 8 hours of darkness) for 4 months. All plant germinations were recorded and identified to morphospecies; when the plants grew, they were identified to the finest level possible. Species that germinated in the control pot were considered later additions and excluded from the analysis.
Data were analyzed with the R statistical packages: stats (R core team, 2022), dunn.test (Dinno, 2017), fossil (Vavrec, 2011), and vegan (Oksanen et al., 2022). Number of germinated seeds per gram of excrete was calculated. To compare between treatments (3 locations and 2 types of protection bags) Kruskal-Wallis and Dunn tests were performed for the total number of germinated seeds and the number of Opuntia seeds (this was the only species with enough data to analyze independently). A GLIM test (with Poisson distribution) was used to determine if the level of protection (plastic bags = 1, mesh bags = 0) and amount of vegetation (both natural and altered) influenced the amount of total and Opuntia germinated seeds. To compare the richness and diversity of dispersed seeds, we calculated the Shannon diversity and Chao 1 (± 95% confidence interval) richness. The similarity between treatments was measured using the Jaccard and Bray-Curtis indexes.
Results
A total effort of 329 trap nights was performed with 199 independent records (at least 1 hour between sightings) of ringtails, of which 44 times ringtails defecated in the latrine (Fig. 2). Opossums were observed 66 times, and no defecation was observed. At the same time, there were 9 observations of rodents and 7 of birds feeding from the latrines.
A total of 52 bags were recovered from the field (34 plastic, and 18 mesh bags), and were put to germinate. In total, 782 plants of 20 species (Table 2) germinated in this experiment. More seeds per gram of excrete germinated in the mesh bags treatment (U tests; V = 1,225, p < 0.001); however, more Opuntia seeds germinated in the plastic bag treatment (U test; V = 300, p < 0.001; Table 3). The Kruskal-Wallis test showed no differences in the number of germinated seeds between locations in the mesh bags, but there were significant differences for total seed and Opuntia seeds in the plastic bag treatment (p = 0.042 and p = 0.002). According to the Dunn test, there were fewer total seeds in the Faculty area than the West core (p = 0.009) and fewer Opuntia seeds in the Faculty area than the Institutes (p = 0.015) and the West core (p = 0.009).
GLIM analysis showed that both level of protection (-0.818, p < 0.001) and vegetation area (1.279, p < 0.001) are significant predictors for the number of total seeds germinated. For Opuntia seeds, protection (1.674, p < 0.001) and vegetation (3.559, p < 0.001).
Figure 2. a) Ringtail defecating in a latrine; b) a rodent consuming seeds from a latrine; c) a latrine; d) plants germinated from excretes.
We found significant differences in richness (according to Chao1) between the Faculty area and the West core (in plastic bags) and between the Faculty area and Institutes and the West core (in mesh bags; Table 4). The Bray-Curtis dendrogram shows 2 important groups: the mesh bag treatment and the plastic bag (Fig. 3). Jaccard index showed the following results within the same location,Faculty plastic-Faculty mesh: 0.714, Institutes plastic-Institutes mesh: 0.546, West core plastic-West core mesh: 0.571, total plastic-total mesh: 0.65; between locations,plastic bags, Faculty plastic-Institutes plastic: 0.769, Faculty plastic-West core plastic: 0.5, Institutes plastic-West core plastic: 0.643;between locations, mesh bags, Faculty mesh-Institutes mesh: 0.5, Faculty mesh-West core mesh: 0.294, Institutes mesh-West core mesh: 0.333.
Figure 3. Bray-Curtis dendrogram of germination treatments. First letter represents the location, W: west core, I: Institutes, F: Faculty; second letter, the type of bag: M: mesh, P: plastic.
Discussion
The camera trap experiment was designed to prove that the latrines from which we collected excretes belong to ringtails and not opossums since those animals produce very similar feces (Aranda, 2000). Given that we observed 44 events of defecation by ringtails and zero by opossums, it we can conclude that ringtails are the only latrine users. However, at the same time, rodents and birds were seen feeding in those latrines (Fig. 2). The ratio for those visits is 1 visit per 2.8 defecations, and this is important for seed dispersal, since most likely those visitors feed on seeds that are dropped by ringtails and can selectively remove some species. At the same time, it is interesting to consider the trade-off for the rodents that consume seeds from ringtail latrines since they are exposed to predation by the latrine owners. Although it is outside the scope of this research, we consider that the interaction around the latrines is a topic that should be further investigated to better understand the role of these animals as seed dispersers and the interactions that the latrines produce.
The comparison between plastic and mesh bags shows differences between the treatments, especially in the number of seeds that germinated per gram of excrete (Tables 2, 3). This could be attributed to the fact that mesh bags allow the smallest particles to exit the bag, and the total weight of feces diminishes while the number of seeds remains constant. Therefore, the mesh bag treatment is not adequate for estimating the number of seeds per gram of excrete, although data concerning the diversity and richness of seeds is still valid. Given the above and the fact that there were more plastic bag replicas, the data interpretation in this research is based on the plastic bag treatment.
Table 2
Number of seeds per gram of excrete in all treatments for each plant species.
Species
Common name
Faculty
Institutes
West core
Plastic
Mesh
Plastic
Mesh
Plastic
Mesh
Ageratina sp. 1
Snakeroot
0.11
3.19
0.41
2.95
0.12
8.63
Ageratina sp. 2
Snakeroot
0.02
0.09
0.02
0
0
0
Asclepias linaria
Pineneedle milkweed
0
0
0
0
0.13
0
Amaranthaceae
0.33
0.19
0
0
0
0
Bidens sp.
Beggarticks
0
0
0.02
0
0.05
0.61
Cissus sicyoides
Princess vine
0
0
0
0
0.03
0.33
Conyza sp.
Horseweed
0.07
0.22
0.26
0
0
0
Drymaria laxiflora
Chickweed
0.04
0.27
0.12
0.35
0.03
2.68
Eragrostis sp.
Lovegrass
0
0.09
0
0
0
0
Iresine sp.
Bloodleaf
0
0
0
0
0
2.04
Opuntia sp.
Prickly pear
0.23
0.98
2.23
0.59
6.36
0.24
Solanaceae
0.03
0.27
0.08
0
0.1
0.71
Solanum nigrescens
Slender nightshade
0.1
2.79
0.04
2.14
0.08
0
Stevia sp.
Stevia
0
0.09
0
0
0
0
Phytolacca icosandra
Tropical pokeweed
0.46
0.15
0.25
0.22
0.08
0
Poaceae 1
0.02
0.39
0.12
0.07
0.13
0.17
Poaceae 2
0
0
0
0
0
0.25
Unknown sp. 1
0.02
0
0.1
0
0.05
0.33
Unknown sp. 2
0
0
0
0
0.05
0
Unknown sp. 3
1.72
0
0
0
0
0
Table 3.
Number of Opuntia and total seeds dispersed by ringtails in different environments. Values given in seeds per g of excrete ± S.D.
Total seeds
Opuntia seeds
Plastic
Mesh
Plastic
Mesh
West core
7.13 ± 8.06
15.99 ± 13.85
6.36 ± 7.58
0.24 ± 0.3
Institutes
3.64 ± 2.87
6.33 ± 6.61
2.23 ± 2.72
0.59 ± 1.33
Faculty
3.16 ± 7.09
8.72 ± 5.51
0.23 ± 0.75
0.98 ± 2.39
Total
4.7 ± 6.64
10.88 ± 10.26
2.96 ± 5.46
0.58 ± 1.5
GLIM analysis shows a positive relation between the number of seeds dispersed by ringtails and vegetation cover. Therefore, ringtails disperse fewer seeds in urban areas (Table 3); however, diversity and richness tend to be higher in the Faculty, the most urbanized location (Table 4).
Table 4
Richness (Chao1 ± 95% confidence intervals) and diversity (Shannon) of seeds dispersed by ringtails.
Faculty
Institutes
West core
Total
Plastic
Mesh
Total
Plastic
Mesh
Total
Plastic
Mesh
Total
Plastic
Mesh
Total
Observed richness
12
12
14
11
6
11
12
10
14
16
17
20
Chao1
16 ± 2.65
14.67 ± 1.85
18.5 ± 3.9
15 ± 2.65
8 ± 1.87
17 ± 0
12.17 ± 0.34
10.25 ± 0.44
14.5 ± 0.73
16.25 ± 0.44
19.67 ± 1.85
24.5 ± 3.4
Shannon
1.45
1.839
1.889
1.273
1.434
1.466
0.564
1.758
1.122
1.403
2.041
1.779
This is consistent with most previous studies that show a greater diversity of seeds in urban settings (Kühn et al., 2004; Wania et al., 2005), and that ringtails consume more plant species in the cities (Cisneros-Moreno & Martínez-Coronel, 2019). At our location, ringtails disperse fewer seeds in urban areas, but their richness and diversity are higher. The novel contribution of this research is that it was performed in an urban area with very little human activity; therefore, the direct effect of human activities such as waste generation and gardening could not explain the differences between treatments. At the same time, all 3 locations share very similar climatic conditions and originally were the same ecosystem. This suggests that differences between treatments are not due to original conditions (although it has been proven in other locations; Kühn et al., 2004). The high diversity of seeds in urbanized areas can be attributed to the great variety of habitats found in an urban area and the exotic species that have already been established there.
Ringtails are common in urban areas, and Castellanos et al. (2009) suggest they prefer disturbed areas over natural ones. This could be due to food availability in urbanized areas, and previous studies show that ringtails have a greater variety of food in urban areas. For example, Cisneros-Moreno and Martínez-Coronel (2019) show that an urban population consumes 36 food items and a rural population registered 28 items. Another advantage that cities offer is infrastructure. Poglayen-Neuwall and Toweill (1988) mention that rocky habitats favor ringtails, and human-made buildings provide a similar habitat. Interestingly, some data suggests that in the absence of humans, sightings of those animals in urban areas increased (Tzintzun-Sánchez, 2022; pers. obs.). This suggests that the benefits ringtails obtain from cities don’t come directly from human activities since, even without our presence, those animals have the benefits of infrastructure and a more diverse diet. However, further studies are needed to fully prove this hypothesis.
Previous studies have reported that fruits are important in the diet of B. astutus; however, the number of different plant species is generally low (13, ~ 9, 10, Alexander et al. [1994], Harrison [2012], and Rodríguez-Estrella et al. [2000], accordingly). In this study, 20 species of seeds were found and it is noteworthy that the cited articles included all vegetal matter in their analysis while this study only considers seeds that germinated. This suggests that the diversity of plants that are consumed in our site is much higher than usual. This could be the result of higher plant diversity in the location, or that ringtails tend to have a more frugivore diet here, this last hypothesis is supported by personal observations of the authors.
Based on the above, it could be concluded that there are several animals that consume the latrine contents, and this interaction could be important for seed dispersal. Ringtails disperse more seeds in natural areas, but their richness is higher in urban areas even in the absence of human activity.
Acknowledgements
We are thankful to M. E. Muñiz-Díaz for facilitating the germination chambers and her advice during this research, to Y. Martínez-Orea for helping with plant identification, to I. Castellanos-Vargas for technical support, and to the SEREPSA working team for facilitating the permission to perform this research. This project was financially supported by PAPIIT grant IN212121 (“El efecto de la urbanización sobre el tlacuache Didelphis virginiana en un matorral xerófilo de la Ciudad de México”), awarded to ZCS.
References
Alexander, L. F., Verts, B. J., & Farrell, T. P. (1994). Diet of ringtails (Bassariscus astutus) in Oregon. Northwestern Naturalist, 75, 97–101. https://doi.org/10.2307/353683131
Aranda, M. (2000). Huellas y otros rastros de los mamíferos grandes y medianos de México. Cuernavaca, Morelos: Instituto de Ecología, A.C./ la Comisión Nacional para el conocimiento y Uso de la Biodiversidad.
Barja, I., & List, R. (2006). Faecal marking behaviour in ringtails (Bassariscus astutus) during the non-breeding period: spatial characteristics of latrines and single faeces. Chemoecology, 16, 219–222. https://doi.org/10.1007/s00049- 006-0352-x
Baskin, C. C., & Baskin, J. M. (1998). Seeds: ecology, biogeography, and, evolution of dormancy and germination. San Diego: Academic Press.
Cano-Santana, Z. (1994). Flujo de energía a través de Sphenarium purpuracens (Orthoptera: Acrididae) y productividad primaria neta aérea en una comunidad xerófila(Ph.D. Thesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. México D.F.
Castellanos, G., García, N., & List, R. (2009). Ecología del cacomixtle (Bassariscus astutus) y la zorra gris (Urocyon cinereoargenteus). In A. Lot, & Z. Cano (Eds.), Biodiversidad del ecosistema del Pedregal de San Ángel (pp. 371–381). Ciudad de Mexico: Secretaria Ejecutiva de la REPSA-UNAM.
Castellanos, G., & List, R. (2005). Área de actividad y uso de hábitat del cacomixtle (Bassariscus astutus) en “El Pedregal de San Ángel”. Revista Mexicana de Mastozoología, 9, 113–122.
Cheptou, P. O., Carrue, O., Rouifed, S., & Cantarel, A. (2008). Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta. Proceedings of the National Academy of Sciences, 105, 3796–3799. https://doi.org/10.1073/pnas.0708446105
Cisneros-Moreno, C., & Martínez-Coronel, M. (2019). Alimentación del cacomixtle (Bassariscus astutus) en un ambiente urbano y uno agrícola en los valles centrales de Oaxaca. Revista Mexicana de Mastozoología (Nueva Época), 9, 31–43. https://doi.org/10.22201/ie.20074484e.2019.1.1.274
Cruz, J. C., Ramos, J. A., Da Silva, L. P., Tenreiro, P. Q., & Heleno, R. H. (2013). Seed dispersal networks in an urban novel ecosystem. European journal of forest research, 132, 887–897. https://doi.org/10.1007/s10342-013-0722-1
Flores-Morales, I. (2023). Diversidad vegetal y animal de los pedregales remanentes de la Zona de Institutos de Investigaciones en Humanidades de Ciudad Universitaria, Ciudad de México, México (Bachelor’s Thesis). Facultad de Ciencias, UNAM. Mexico City.
Glebskiy, Y. (2019). Efecto del conejo castellano (Sylvilagus floridanus) sobre la comunidad vegetal del Pedregal de San Ángel. (M. Sc. Thesis). Facultad de Ciencias, UNAM. México City.
Harrison, R. L. (2012). Ringtail (Bassariscus astutus) ecology and behavior in central New Mexico, USA. Western North American Naturalist, 72, 495–506. https://doi.org/ 10.3398/064.072.0407
Kühn, I., Brandl, R., & Klotz, S. (2004). The flora of German cities is naturally species rich. Evolutionary Ecology Research, 6, 749–764.
Mendoza-Hernández, P. E., & Cano-Santana, Z. (2009). Elementos para la restauración ecológica de pedregales: la rehabilitación de áreas verdes de la Facultad de Ciencias en Ciudad Universitaria. In A. Lot, & Z. Cano-Santana (Eds.), Biodiversidad del ecosistema del Pedregal de San Ángel (pp. 523–532). Ciudad de México: Secretaría Ejecutiva de la REPSA-UNAM.
Niu, H., Rehling, F., Chen, Z., Yue, X., Zhao, H., Wang, X. et al. (2023). Regeneration of urban forests as influenced by fragmentation, seed dispersal mode and the legacy effect of reforestation interventions. Landscape and Urban Planning, 233, 104712. https://doi.org/10.1016/j.landurbplan.2023.104712
Niu, H. Y., Xing, J. J., Zhang, H. M., Wang, D., & Wang, X. R. (2018). Roads limit of seed dispersal and seedling recruitment of Quercus chenii in an urban hillside forest. Urban Forestry & Urban Greening, 30, 307–314. https://doi.org/10.1016/j.ufug.2018.01.023
Oksanen, J., Simpson, G., Blanchet, F., Kindt, R., Legendre, P., Minchin, P. et al. (2022). vegan: Community Ecology Package. R package version 2.6-2, https://CRAN.R-project.org/package=vegan
Picazo, G. E. R. C., & García-Collazo, R. (2019). Comparación de la dieta del cacomixtle norteño, Bassariscus astutus de un bosque templado y un matorral xerófilo, del centro de México. Biocyt: Biología, Ciencia y Tecnología, 12, 834–845. https://doi.org/10.22201/fesi.20072082.2019.12.68527
Poglayen-Neuwall, I., & Toweill, D. E. (1988). Bassariscus astutus. Mammalian Species, 327, 1–8.
R Core Team (2022). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rodríguez-Estrella, R., Moreno, A. R., & Tam, K. G. (2000). Spring diet of the endemic ring-tailed cat (Bassariscus astutus insulicola) population on an island in the Gulf of California, Mexico. Journal of Arid Environments, 44, 241–246. https://doi.org/10.1006/jare.1999.0579
Rubalcava-Castillo, F. A., Sosa-Ramírez, J., Luna-Ruíz, J. J., Valdivia-Flores, A. G., Díaz-Núñez, V., & Íñiguez-Dávalos, L. I. (2020). Endozoochorous dispersal of forest seeds by carnivorous mammals in Sierra Fría, Aguascalientes, Mexico. Ecology and Evolution, 10, 2991-3003. https://doi.org/10.1002/ece3.6113
Rzedowski, J. (1954). Vegetation of Pedregal de San Ángel. Anales de la Escuela Nacional de Ciencias Biológicas, IPN, Mexico, 8, 59 Endozoochorous 129.
Swanson, A. C., Conn, A., Swanson, J. J., & Brooks, D. M. (2022). Record of an Urban Ringtail (Bassariscus astutus) Outside of its Typical Geographic Range. Urban Naturalist Notes, 9, 1–6.
Tzintzun-Sánchez, C. L. (2022). Efectos de la urbanización en la distribución geográfica y hábitos alimenticios del cacomixtle norteño (Bassariscus astutus) en la Ciudad de México, México (Bachelor’s Thesis). Facultad de Ciencias, UNAM. Ciudad de México.
Wania, A., Kühn, I., & Klotz, S. (2006). Plant richness patterns in agricultural and urban landscapes in Central Germany —spatial gradients of species richness. Landscape and Urban planning, 75, 97–110. https://doi.org/10.1016/j.landurbplan. 2004.12.006
Zambrano, L., Rodríguez-Palcios, S., Pérez-Escobedo, M., Gil-Alarcón, G. Camarena, P., & Lot, A. (2016). Reserva Ecológica del Pedregal de San Ángel: atlas de riesgos. Ciudad de Mexico: Secretaría Ejecutiva de la REPSA-UNAM.
Carolina Granados Mendoza a, *, Miguel Murguía-Romero b, Gerardo A. Salazar a
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Botánica, Circuito Zona Deportiva s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
b Universidad Nacional Autónoma de México, Instituto de Biología, Unidad de Informática para la Biodiversidad, Circuito Zona Deportiva s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
The Instituto de Biología of the Universidad Nacional Autónoma de México (IBUNAM) celebrates 95 years as a leading institution on biodiversity research. Besides developing frontier research, training of human resources, and public science communication, IBUNAM houses 10 National Zoological Collections and the National Herbarium of Mexico (MEXU). The specimens deposited in these collections are the foundation of numerous biological studies, from which ample genetic resources have been generated. Here we present an initial effort to link the specimens deposited at the National Biological Collections housed at IBUNAM to their public genetic resources, using MEXU’s Collection of Types of Vascular Plants as proof of concept. First, a list of the type specimens was retrieved from IBdata, the web system for consulting the records of the biological collections housed at IBUNAM. Then, the interface Entrez Programming Utilities of GenBank was used to search for the available genetic resources associated with the type specimens. New fields were incorporated into IBdata to facilitate access to the identified genetic resources. Future initiatives should promote access to the public metadata (e.g., molecular, morphological) associated to specimens of the biological collections housed at IBUNAM.
Keywords: Metadata; GenBank; IBdata; Type specimens; Digitalized information
¡Es tiempo de celebrar! Vinculando recursos genéticos a las Colecciones Biológicas Nacionales mexicanas custodiadas por el Instituto de Biología de la Universidad Nacional Autónoma de México
Resumen
El Instituto de Biología de la Universidad Nacional Autónoma de México (IBUNAM) cumple 95 años como institución líder en investigación de la biodiversidad. Además de desarrollar ciencia de frontera, formación de recursos humanos y comunicación pública de la ciencia, el IBUNAM alberga 10 Colecciones Zoológicas Nacionales y el Herbario Nacional de México (MEXU). Los ejemplares depositados en estas colecciones son fundamento de numerosos estudios biológicos, de los cuales se han generado amplios recursos genéticos. Aquí se presenta un primer esfuerzo para vincular los ejemplares depositados en las Colecciones Biológicas Nacionales albergadas en el IBUNAM con sus recursos genéticos públicos, utilizando como prueba de concepto la Colección de Tipos de Plantas Vasculares del MEXU. Primero, se recuperó una lista de especímenes tipo de IBdata, el sistema web que permite consultar los registros de las colecciones biológicas alojadas en el IBUNAM. Luego, se utilizó la interfaz Entrez Programming Utilities de GenBank para buscar los recursos genéticos disponibles asociados a los especímenes tipo. Se incorporaron nuevos campos a IBdata para facilitar el acceso a los recursos genéticos identificados. Iniciativas futuras deberían promover el acceso a los metadatos públicos (e.g., moleculares, morfológicos) asociados a los especímenes de las colecciones biológicas albergadas en el IBUNAM.
The Instituto de Biología of the Universidad Nacional Autónoma de México (IBUNAM) celebrates its 95th anniversary this year. Faculty and students at IBUNAM are devoted to the study, conservation, and sustainable use of the biota of Mexico, but also from other regions of the world. The research performed at IBUNAM touches on virtually all branches of the tree of life and uses a wide variety of methodological and analytical tools to discover, describe, document, and understand biological diversity. Among other research institutions in Mexico, IBUNAM stands out for housing several National Biological Collections, including 10 National Zoological Collections and the National Herbarium of Mexico (MEXU). The specimens deposited at the IBUNAM’s National Biological collections are the foundation of a myriad of taxonomic, evolutionary, ecological, biogeographic, social, and conservation studies, from which an enormous amount of associated data (hereafter referred to as “metadata”) is generated.
Every day, these collections are actively consulted, both in person and virtually through IBdata (http://ibdata4.ib.unam.mx), a web system for consulting the records of the National Biological Collections housed at IBUNAM (Murguía-Romero et al., 2024). Under UNAM’s open data policy (http://www.datosabiertos.unam.mx/informacion/terminosdeuso.html), IBdata currently provides free, easy, and continuous access to digitalized information of over 1.7 million biological specimens, allowing the dissemination of knowledge and transdisciplinary research, thus benefiting the scientific, governmental, and educational society sectors, as well as private users. For each physical specimen, the digitalized information available in IBdata usually includes high-resolution digital images along with data on the locality where the specimen was collected (including geographic coordinates, when available), date of collecting, collector(s), as well as notes on habitat, morphological, and socio-cultural aspects recorded by the collectors.
One of the commonly generated metadata are genetic resources, which are often made publicly available through the International Nucleotide Sequence Database Collaboration (INSDC; https://www.insdc.org/), which includes 3 international databases that exchange data every day, namely the DNA Databank of Japan (DDBJ; https://www.ddbj.nig.ac.jp/index-e.html), the European Nucleotide Archive (ENA; https://www.ebi.ac.uk/ena/browser/), and GenBank of the USA National Center for Biotechnology Information (NCBI; https://www.ncbi.nlm.nih.gov/genbank/). When properly submitted, these genetic resources include information about the voucher specimens of the genetic data, as well as where the specimens are deposited. Access to such genetic information is essential for the sustainable use and conservation of global biodiversity (Cowell et al., 2022).
Here we used the information of MEXU’s Collection of Types of Vascular Plants available in IBdata (10,972 records) to search and link the specimens to their public genetic resources available at GenBank. For this, we downloaded all the type records and built URL calls for the interface Entrez Programming Utilities (E-utilities; https://www.ncbi.nlm.nih.gov/books/NBK25501/) of GenBank. Query searches used the species’ scientific name and the collection number assigned by the collector or the unique identifier of the specimens (MEXU’s catalogue number) as in the following example: “https://eutils.ncbi.nlm.nih.gov/entrez/eutils/esearch.fcgi?db=nucleotide&term=%22Agave%20isthmensis%22[organism]+AND+(4177+OR+628489)”.
In cases where the collection number contained non-numerical characters or spaces, query searches used instead the main collector’s last name (only the first last name was used when 2 last names were present). URL calls were submitted to the nucleotide NCBI database, with a 1 s delay between each search request to avoid overload of the NCBI E-utility servers, with the Bulk URL Opener extension of Google Chrome. Calls with hits were saved, and the corresponding query translations were used to download the associated GenBank accession numbers. Retrieved accession numbers were merged into a text file to perform a NCBI Batch Entrez search (https://www.ncbi.nlm.nih.gov/sites/batchentrez). The resulting records were further filtered with a custom filter using the flag “MEXU.” The filtered results were revised individually to confirm their association with one vascular plant type specimen deposited at MEXU. To facilitate linking back the type specimens to their associated genetic metadata, IBdata records of type specimens having genetic information available at GenBank were complemented with a new data field called “GenBank”, which displays a list of available molecular markers and their corresponding GenBank accession numbers. Additionally, a web link leading to the corresponding NCBI records was added as an additional data field named “GenBank Search” in IBdata (Fig. 1).
Our search identified 71 GenBank accession numbers corresponding to 23 angiosperm species representing 20 genera, 7 families, and 5 orders (Table 1). Type specimens found associated with genetic resources at GenBank can be easily accessed in IBdata through the “Simple Search” option and the keyword “genbanksearch”. The sequenced molecular markers include 18 plastid regions: genes accD, atpA, matK, ndhF, psbA, rbcL, and ycf1; the introns of the genes rpl16, trnL; the intergenic spacers rpl20-rps12, rpl32-trnL, rps16-trnQ, trnS-trnG, and ycf6-psbM; the regions trnD-trnT, trnH-psbA, trnK-matK, and trnL-trnF; and different portions of the nuclear-ribosomal Internal Transcribed Spacer (ITS) region. Each type specimen had 1 to 7 associated sequences, being the most frequently sequenced markers the plastid regions trnL-trnF and trnK-matK and the nuclear-ribosomal ITS region. Most sequenced type specimens were collected in the 1980’s, 1990’s, and 2000’s. However, the sequenced isotype specimen of Epiphyllum chrysocardium Alexander (Cactaceae) was collected in 1951 (Fig. 1). Authors of the sequences of the latter type specimen indicated us that the plant tissue used for DNA extraction came indeed from the type collection (MacDougall, 198), but from a division maintained under cultivation at the Botanical Garden of IBUNAM, explaining why sequencing was achieved from such an “old” specimen. Although explicitly stated for only 19 accessions, all the markers recovered seem to have been generated through capillary (Sanger) sequencing.
Previous studies have stressed the importance of open access to the digitalized information of type specimens (Nicolson et al., 2023), which are key reference elements of scientific names. The value of both the digitalized information of type specimens and the genetic information derived from them increases when both elements can be linked and easily accessed. Including genetic sequences from type specimens into molecular taxonomic studies often plays an important role in the circumscription of taxa or their placement at a particular place of the tree of life. Explicit recognition of the inclusion of genetic sequences from type specimens in molecular studies can promote the progress of molecular systematics and taxonomy (Chakrabarty, 2010).
Table 1
MEXU’s types of vascular plants with available genetic information at GenBank.
Caesalpinia oyamae synonym of Erythrostemon oyamae (Sotuyo & G.P. Lewis) Gagnon & G.P. Lewis
KX373079.1, KX379300.1
Hawkins, 23
Holotype
Phaseolus albescens McVaugh ex R. Delgad. & A. Delgado
AF115150.1, DQ445955.1
Delgado, 1705
Holotype
Platymiscium calyptratum M. Sousa & Klitg.
EU735872.1, EU735933.1, EU735990.1, EU736047.1
Tenorio, 126
Holotype
Harpalyce torresii São-Mateus & M. Sousa
PP250089.1, PP238799.1
Téllez, 950
Paratype
Figure 1. IBdata “Summary data sheet of the specimen” of the isotype of Epiphyllum chrysocardium (MEXU’s catalogue number: 72938) showing within a red rectangle two new implemented fields, “GenBank” and “GenBank Search”, which link the specimen to its available genetic information in GenBank.
Given the improvements in sequencing technology, sequencing of type and non-type herbarium specimens should seek to incorporate more efficient sequencing strategies that maximize the amount of generated sequence data. The combination of supervised sampling of herbarium specimens with high-throughput DNA sequencing and bioinformatics has given rise to “herbariomics” i.e., the access to genome-scale genetic information from specimens maintained in herbaria. Such an approach opens the possibility of incorporating in genomic, phylogenomic, and population genetic studies taxa that otherwise may not be accessible, such as extinct or extremely rare species, or species that live in places difficult to access or subjected to regulations for collecting (Davis, 2023; Strijk et al., 2020). The wealth of already available, potential sources of new genomic information is informed by the recent report by Thiers (2023) on the world’s herbaria, based on data from the Index Herbariorum (https://sweetgum.nybg.org/science/ih/): the 3,567 active herbaria in the world hold over 396.7 million specimens. It should be a priority to incorporate those valuable sources of already collected, curated specimens in world-wide initiatives such as the “global biodiversity cyberbank” (Wen et al., 2017), aimed at integrating all the existing resources to promote free access and generation of information on biological diversity.
Cowell, C., Paton, A., Borrell, J. S., Williams, C., Wilkin, P., Antonelli, A. et al. (2022). Uses and benefits of digital sequence information from plant genetic resources: Lessons learnt from botanical collections. Plants, People, Planet, 4, 33–43. https://doi.org/10.1002/ppp3.10216
Murguía-Romero, M., Serrano-Estrada, B., Salazar, G. A., Sánchez-González, G. E., Melo-Samper, U., Gernandt, D. S. et al. (2024). The IBdata Web System for Biological Collections: design focused on usability. Biodiversity Informatics, 18, 1–12. https://doi.org/10.17161/bi.v18i.20516
Nicolson, N., Trekels, M., Groom, Q. J., Knapp, S., & Paton, A. J. (2023). Global access to nomenclatural botanical resources: Evaluating open access availability. Plants, People, Planet, 5, 899–907. https://doi.org/10.1002/ppp3.10438
Strijk, J. S., Binh, H. T., Ngoc, N. V., Pereira, J. T., Slik, J. W. F., Sukri, R. S. et al. (2020). Museomics for reconstructing historical floristic exchanges: divergence of stone oaks across Wallacea. Plos One, 15, e0232936. https://doi.org/10.1371/journal.pone.0232936
Wen, J., Harris, A., Ickert-Bond, S. M., Dikow, R., Wurdack, K., & Zimmer, E. A. (2017). Developing integrative systematics in the informatics and genomic era and calling for a global Biodiversity Cyberbank. Journal of Systematics and Evolution, 55, 308–321. https://doi.org/10.1111/jse.12270
A new species of whitefly, Aleuropleurocelus tecomastans, is described. The studied specimens were found in the municipality of Acapulco, State of Guerrero, Mexico on Tecoma stans (L.) (Bignoniaceae) leaves. A dichotomous key to identify members of Aleuropleurocelus group ceanothi, defined by a transverse suture of the molt which reaches the submarginal line, is provided. Photographs of morphological pupal structures are given and diagnostic separation to other related species is discussed.
Una especie nueva de Aleuropleurocelus (Hemiptera: Aleyrodidae) y clave para el grupo ceanothi de Mexico
Resumen
Una especie nueva de mosca blanca, Aleuropleurocelus tecomastans, es descrita. Los especímenes estudiados fueron encontradosen el municipio de Acapulco, Estado de Guerrero, México sobre Tecoma stans (L.) (Bignoniaceae). Se provee una clave dicotómica para identificación de especies de Aleuropleurocelus grupo ceanothi, definido por una sutura transversa de la muda que termina en la línea submarginal. Se proporcionan fotografías de las estructuras morfológicas del pupario y se analiza la separación diagnóstica en otras especies relacionadas.
The genus Aleuropleurocelus was described by Drews and Sampson (1956), comprising 8 species and an identification key to all Californian species was included (Drews & Sampson, 1958). Later Dooley et al. (2010) described A. nevadensis Dooley, while Polaszek and Gill (2011) added A. hyptisemoryi Gill to the genus. Nowadays, 19 species of Mexican species of Aleuropleurocelus are known and have been largely studied by Carapia-Ruiz (2023), Carapia-Ruiz (2020a, b), Carapia-Ruiz and Sánchez-Flores (2019a, b), Carapia-Ruiz et al. (2018a, b, 2020, 2023), Sánchez-Flores and Carapia-Ruiz (2018a, b), Sánchez-Flores et al. (2018a, b, 2020, 2021). The genus is segregated into 3 major groups: abnormis (semioval), nigrans, and ceanothi according to Dooley et al. (2010) and Sánchez-Flores et al. (2021).
The ceanothi group, in which the transverse suture of molt ends at the submarginal line is included, A. granulata (Sampson & Drews), and the morphologically related species A. sampsoni Sánchez-Flores & Carapia-Ruiz and A. pseudogranulata Carapia-Ruiz & Sánchez-Flores. While collecting puparia of Aleyrodidae in Acapulco Guerrero, an unknown species of this group with distinctive characters was found. The objective of this contribution is to describe a new species and provide a key to all species of Aleuropleurocelus group ceanothi.
Materials and methods
The specimens were collected on the underside of the leaves of Tecoma stans (L.). in Acapulco, Guerrero. The specimens once taken were retained ethanol and thus transferred to the Laboratorio de Entomología, Escuela de Estudios Superiores de Xalostoc of the Universidad Autónoma del Estado de Morelos, to be fully processed and mounted in permanent preparation with Canada balsam on slides for further study under stereomicroscope and facilitating identification following specialized literature such as Drews and Sampson (1958), Martin (2004, 2005), and Sanchez-Flores et al. (2021). After preparations, specimens were observed under a Motic BA 320 phase contrast optical microscope considering several magnifications: 4X 100X, 400X and 1,000X. The terminology used follows Drews and Sampson (1956) and Martin (2005). The studied specimens are deposited at Colección Nacional de Insectos (CNIN), Instituto de Biología, Universidad Nacional Autónoma de México. México City, Mexico.
Diagnosis. Pupa boat-shaped, transverse molting suture reaches the submarginal line, dorsum and venter black, eye spots with tubercles, abdominal depressions present, middle area of abdominal segments smooth, abdominal VIII setae anterolateral to the vasiform orifice very small.
General form: pupae in situ. Dorsal and ventral surface of pupae black, a thin fringe of wax is distinguished around the body margin.
Specimens on slides: semioval body (Fig. 1). Deflexed submargin. Margin: with 61-68 pairs of marginal teeth with 2-3 acute terminal protrusions (Fig. 2), submargin with a regular band of small tubercles (Fig. 3); subdorsum: with a band of crescent tubercles (Fig. 4). Cephalothorax: longitudinal molting suture with a row of defined tubercles on each side giving the appearance of a zipper, transverse molting suture ends at the apparent margin (submarginal fold), well defined meso-metathoracic suture (Fig. 5), ocular markings (eye spots) indicated by pale coloration and 5-7 pairs of pale tubercles in the subdorsal area near the submarginal line, cephalic setae absent; mid thoracic zone with 2 pairs of setae, the mesothoracic and the metathoracic. Abdomen: dorsum with abdominal segments I-VIII clearly visible in the middle part (Fig. 6). With abdominal depressions in the middle area of the segments (Fig. 7), cuticle in the middle of the abdominal segments smooth. Vasiform orifice: elongated semicordiform (Fig. 8); elevated; operculum with 4 irregular longitudinal furrows and spinules at the apex, completely covering the lingula and most of the vasiform orifice, ring of the orifice defined in its anterior part, abdominal VIII setae anterolateral to the vasiform orifice very small, caudal protuberance developed. Pores, normally as follows: 8 pairs in the cephalic area, 6 on submedian area and 2 posterior to eye spots; 4 pairs in the mesothorax, 2 on submedian area and 2 in the subdorsal area of the mosothorax; medial area of abdominal segments I, III, V, VII with 2 pairs each segment, segment VIII with 2 pairs; subdorsal area of abdominal segments II, IV, V, with 2 pairs of pores in each segment (Fig. 9).
Venter: legs prothoracic, mesothoracic, metathoracic with apical adhesive sac, thoracic adhesive sacs near the base of the first pair of legs, base of the legs with a wide irregular band of 2-4 spinules, thoracic and abdomen cuticle smooth, well defined abdominal setae of segment VIII, posteriorly present a pair of spiracles. Chaetotaxy: a pair of anterior marginal setae present (near marginal teeth), cephalic setae absent, mesothoracic, metathoracic and caudal setae well developed, very small abdominal VIII setae, located anterolateral to the vasiform orifice, and posterior marginal setae small.
Figures 5-8. Aleuropleurocelus tecomastans.5) Thoracic area; 6) submedia and subdorsal area; 7) depression and pore of abdominal segment IV; 8) vasiform orifice. ad = Abdominal depression, cs = caudal seta, mss = mesothoracic seta, mts = metathoracic seta, o = operculum, pp = pore porete, sd = subdorsum, vo = vasiform orifice.
Measurements. Specimens on slides: body 550-650 µm long by 300-400 µm wide. Submargin approximately 45-55 μm. Cephalothorax: 230-280 μm, longitudinal molting suture, 240-360 μm, transverse molting suture 220-250 μm, the metathorax 30-45 μm long in its middle area, cephalic elongated structures of 8-10 µm long by 3-5 µm wide. Abdomen: abdominal segments length for segment I 22-26 μm, segment II 22-26 μm, segment III 25-30 μm, segment IV 26 -30 μm, segment V 27-32 μm, segment VI 26-29 μm, segment VII 23-27 μm, and segment VIII (from suture VII-VIII to vasiform orifice) 40-50 μm, distance from vasiform orifice to apparent margin 25-35 μm; abdominal depressions segments with approximate length in segment I 3-5 μm long by 10-12 μm wide, in segment II 3-4 μm long by 12-15 μm wide, in segment III 3-4 μm long by 10-13 μm wide, in segment IV 3-4 μm long by 10-14 μm wide, in segment V 5-7 μm long by 10-15 μm wide, in segment VI 5-6 μm long by 9-11 μm wide. Vasiform orifice: 46-50 µm long by 32-38 µm of broad at the widest part; operculum 25-30 μm long by 22-26 μm wide. Venter: Prothoracic legs 75-77 μm long, mesothoracic legs 75-78 μm long, metathoracic legs 85-89 μm long, thoracic adhesive sacs of 16-20 μm in diameter, base of the legs spinules 4 μm long and 2 μm wide. Setae: anterior marginal setae approximately 7 μm long, mesothoracic setae approximately 15 μm long, metathoracic setae approximately 20 μm long, abdominal VIII setae 3-4 μm long, caudal seta approximately 20 μm long, and posterior marginal seta approximately 10 μm long.
Type locality: northeast of Acapulco, Guerrero, Mexico
Type material: holotype, puparium: Acapulco, Guerrero, Mexico, in leaves of Tecoma stans (Bignoniaceae), 8-iv-2021, Col. V. E. Carapia-Ruiz, deposited in CNIN (Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City, Mexico), HOM-TIP-170. Paratypes: puparia, same data as holotype, 2 deposited in CNIN, HOM-PAR-171, HOM-PAR-172.
Etymology: the suffix name is based on a combination of the host plant’s scientific name where specimens were associated.
Distribution: Acapulco, Guerrero, Mexico.
Plant associations: Tecoma stans (Bignoniaceae).
Remarks.
Aleuropleurocelus tecomastans is placed within the ceanothi group by the transverse molting suture reaches the apparent margin (submarginal line). The new species is similar in apparence to A. granulate, A. sampsoni,and A. pseudogranulata but can be differentiated by the presence of abdominal depressions which is opposite as shown in A. granulata, A. sampsoni, A. pseudogranulata or A. ceanothi. Also A. asciculatus presents a subdorsal fold which is absent in the other species.
Key to species of Aleuropleurocelus group ceanothi
1. With bands of dense wax on dorsum, pores of double wall on dorsum 2
—Without such bands or pores 3
2. (1) Two bands of dense wax on dorsum (1 thin band on subdorsum and 1 wide band on submedian area ornatus Drews & Sampson
— One band of dense wax on subdorsum sampsoni Sánchez Flores & Carapia-Ruiz
3. (1) Tubercles on submedian surface absent 4
—Tubercles on submedian surface present 8
4. (3) Without evident tubercles on subdorsal surface 5
— With evident tubercles on subdorsal surface 6
5. (4) Longitudinal suture of the molt with tubercles on metathorax laingi Drews & Sampson
— Longitudinal suture of the molt without tubercles on metathorax coachellensis Drews & Sampson
6. (4) Subdorsal fold absent 7
— Subdorsal fold present asciculatus Carapia-Ruiz
7. (6) 2 pair of thoracic adhesive sacs, submarginal band with pores erigonium Carapia-Ruiz
— Without tubercles on the anterior and posterior part of the abdominal segments, abdominal depressions present tecomastans n. sp.
8. (3) With 2 pairs of thoracic adhesive sacs, submedian area with tubercles only on the anterior part of the abdominal segments; marginal teeth without 2 or more evident prostructions 9
— With a pair of thoracic adhesive sacs, submedian area with tubercles on the anterior and posterior part of the abdominal segments; marginal teeth with evident prostructions; transversal suture of the molt curved 10
9. (8) Half posterior of puparium in elliptic form sierra (Sampson)
— Half posterior of puparium in triangular form ceanothi (Sampson)
10. (8) Vasiform orifice semicircular, venter of abdomen with posterior area very narrow, transversal suture of the molt almost straight on median area granulata Sampson & Drews
— Vasiform orifice elongated, clearly more long than wide, venter of abdomen with posterior area triangular in form, transversal suture of the molt curved on median area pseudogranulata Carapia-Ruiz
Acknowledgments
To Paul Brawn and David Ovread of the British Museum of Natural History (BMNH) for facilitating a loan of Aleuropleurocelus to study the specimens.
References
Carapia-Ruiz, V. E. (2020a). Descripción de una especie nueva del género Aleuropleurocelus y nuevos registros para Baja California, México. Southwestern Entomologist, 45, 269–274. https://doi.org/10.3958/059.045.0128
Carapia-Ruiz, V. E. (2020b). Description of a new species of the genus Aleuropleurocelus Drews & Sampson (Hemiptera: Aleyrodidae) and a new country record for a species of the genus from Mexico. Acta Zoológica Mexicana (nueva serie), 36, 1–7. https://doi.org/10.21829/azm.2020.3612184
Carapia-Ruiz, V. E. (2023). Dos especies nuevas de Aleuro- pleurocelus de México, Southwestern Entomologist, 48, 641-648. https://doi.org/10.3958/059.045.0425
Carapia-Ruiz, V. E., & Sánchez-Flores, O. A. (2019a). Descripción de una especie nueva del género Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) de California, Estados Unidos. Acta Zoológica Mexicana (nueva serie), 35, 1–4. https://doi.org/10.21829/azm.2019.3501230
Carapia-Ruiz, V. E., & Sánchez-Flores, O. A. (2019b). Descripción de la primera especie pálida del género Aleuropleurocelus. Southwestern Entomologist, 44, 315–319. https://doi.org/10.3958/059.044.0133
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., & García-Ochaeta, J. F. (2023). A new whitefly species, Aleuropleurocelus petenensis sp. nov. (Hemiptera: Aleyrodidae), from Guatemala. Acta Zoológica Mexicana (nueva serie), 39, 1–8. https://doi.org/10.21829/azm.2023.3912470
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2018a). Descripción de dos especies nuevas del género Aleuropleurocelus Drews y Sampson, 1956 (Hemiptera: Aleyrodidae) de México. Insecta Mundi, 0606, 1–13.
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2018b). Descripción de dos especies nuevas del género Aleuropleurocelus Drews y Sampson, 1956 (Hemiptera: Aleyrodidae) de México. Southwestern Entomologist, 43, 517–526.
Carapia-Ruiz, V. E., Sánchez-Flores, O. A., García-Martínez, O., & Castillo-Gutiérrez, A. (2021). Aleuropleurocelus palidonigrans sp. nov. de Guerrero, México, Southwestern Entomologist, 46, 781–786. https://doi.org/10.3958/059.046. 0320
Dooley, J. W., Lambrecht, S., & Honda, J. (2010). Eight new state records of aleyrodine whiteflies found in Clark County, Nevada and three newly described taxa (Hemiptera: Aleyrodidae, Aleyrodinae). Insecta Mundi, 140, 1−36.
Drews, E. A., & Sampson, W. W. (1956). Tetralicia and a new related genus Aleuropleurocelus (Homoptera: Aleyrodidae). Annals Entomological Society of America, 49, 280–283. https://doi.org/10.1093/aesa/49.3.280
Drews, E. A., & Sampson, W. W. (1958). California aleyrodids of the genus Aleuropleurocelus. Annals of the Entomological Society of America, 51, 120–125.
Martin, J. H. (2004). The whiteflies of Belize (Hemiptera: Aleyrodidae) Part 1 — Introduction and account of the subfamily Aleurodicinae Quaintance and Baker. Zootaxa, 681, 1–119. https://dx.doi.org/10.11646/zootaxa.681.1.1
Martin, J. H. (2005). Whiteflies of Belize (Hemiptera: Aleyrodidae). Part 2 — A review of the subfamily Aleyrodinae Westwood. Zootaxa, 1098, 1–116. https://dx. doi.org/10.11646/zootaxa.1098.1.1
Polaszek, A., & Gill, R. (2011). A new species of whitefly (Hemiptera: Aleyrodidae) and its parasitoid (Hymenoptera: Aphelinidae) from desert lavender in California. Zootaxa, 2750, 51–59. https://doi.org/10.11646/zootaxa.2150.1.5/biota xa.org
Sánchez-Flores, O. A., & Carapia-Ruiz, V. E. (2018a). Nueva especie de Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) y clave para especies del grupo de forma oval. Insecta Mundi, 651, 1–12.
Sánchez-Flores, O. A., & Carapia-Ruiz, V. E. (2018b). Description of two new species of the genus Aleuropleurocelus Drews y Sampson (Hemiptera: Aleyrodidae) from Mexico. Acta Zoológica Mexicana (nueva serie), 34, 1–9. https://doi.org/10.21829/azm.2018.3412145
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., Coronado-Blanco, J. M., & Ruíz-Cancino, E. (2020). Description of Aleuropleurocelus sampsoni sp. nov. (Hemiptera: Aleyrodidae) from Tamaulipas, Mexico. Florida Entomologist, 103, 274–280. https://doi.org/10.1653/024.103.0219
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., García-Martínez, O., & Castillo-Gutiérrez, A. (2018a). Descripción de una especie nueva del género Aleuropleurocelus de México. Southwestern Entomologist, 43, 257–262. https://doi.org/ 10.3958/059.043.0116
Sánchez-Flores, O. A., Carapia-Ruiz, V. E., García-Martínez, O., & Castillo-Gutiérrez, A. (2018b). Descripción de una especie nueva del género Aleuropleurocelus (Hemiptera: Aleyrodidae) de México. Acta Zoológica Mexicana (nueva serie), 34, 1–6. https://doi.org/10.21829/azm.2018.3412104
Sánchez-Flores, O. A., García-Ochaeta, J. F., Carapia-Ruiz, V. E., Ruiz-Cancino, E., & Coronado-Blanco, J. M. (2021). New species of Aleuropleurocelus Drews and Sampson (Hemiptera: Aleyrodidae) from Guatemala, with a key to species from the oval group. Proceedings of the Entomological Society of Washington, 123, 615–621. https://doi.org/10.4289/0013-8797.123.3.614
Universidad Nacional Autónoma de México, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Antigua carretera a Pátzcuaro No. 8701, Colonia Ex Hacienda de San José de La Huerta, 58190, Morelia, Michoacán, Mexico
The mutualism between Ficus species (Moraceae) and their pollinating wasps (Agaonidae) is a widely recognized coevolutionary model. In Ficus species from the Paleotropic, it has been determined that the ostiole acts as a morphological filter that affects the head dimensions of pollinating female wasps. Here, for the first time, the allometric relationship between ostiole size (diameter and length) and the shape of the head (length/width) of pollinating wasps is quantitatively explored in 6 Neotropical Ficus species (3 sect. Americanae and 3 sect. Pharmacosycea). In the case of sect. Americanae, wasp head shape was significantly correlated only with ostiole length, while in sect. Pharmacosycea both ostiole variables were correlated with head shape. The ordination analysis (NMDS) clearly reflected associations of these traits in species for both sections. The results support what has been interpreted in previous studies as reciprocal evolution between the analyzed traits, which contribute, along with other morphological and ecological traits, to the specificity between Ficus species and their pollinating wasps.
¿Qué tan relevante es la relación entre el tamaño del ostíolo y la forma de la cabeza de las avispas en la interacción mutualista Ficus-Agaonidae?
Resumen
El mutualismo entre Ficus (Moraceae) y sus avispas polinizadoras (Agaonidae) es un modelo coevolutivo ampliamente reconocido. En especies de Ficus del Paleotrópico, se ha determinado que el ostíolo actúa como un filtro morfológico que afecta las dimensiones de la cabeza de las hembras polinizadoras. Por primera vez se explora, cuantitativamente, la relación alométrica entre el tamaño del ostíolo (diámetro y largo) y la forma de la cabeza (largo/ancho) de las avispas polinizadoras en 6 especies de Ficus neotropicales (3 sect. Americanae y 3 sect. Pharmacosycea). En el caso de sect. Americanae, la forma de la cabeza de las avispas se correlacionó significativa y únicamente con el largo del ostíolo, mientras que en la sect. Pharmacosycea, ambas variables del ostíolo se correlacionaron con la forma de la cabeza. El análisis de ordenación (NMDS) reflejó claramente asociaciones de estos atributos por especie para ambas secciones. Los resultados apoyan lo expuesto en estudios previos de una evolución recíproca entre los atributos analizados, lo que contribuye, en conjunto con otros atributos morfológicos y ecológicos, a la especificidad entre Ficus y sus avispas polinizadoras.
The interaction between species of the genus Ficus L. (Moraceae) and wasps of the family Agaonidae Walker (Hymenoptera) is a classic example of a mutualistic interaction, in which the phenomena of co-diversification, co-speciation, and co-evolution are well documented (Borges, 2021; Bronstein, 1988; Jousselin et al., 2003; Rønsted et al., 2005; Wang et al., 2019; Wiebes, 1979) and is estimated to date to ca. 74.9 Ma (60.0-101.9 Ma; Cruaud et al., 2012; Zhang et al., 2019). For many years it was assumed that 1 Ficus species was pollinated exclusively by 1 wasp species (e.g., Ramírez, 1970; Wiebes, 1966, 1979), but a growing amount of evidence contradicts this assumption (Cook & Rasplus, 2003; Cook & Segar, 2010; Herre et al., 2008; Machado et al., 2005; Molbo et al., 2003; Oldenbeuving et al., 2023; Peng et al., 2008; Su et al., 2008; Weiblen, 2002).
Regardless of the degree of specificity, this ancestral and close mutualistic relationship involves complex morphological modifications (Clement et al., 2020; Galil & Eisikowich, 1968). The flowers of Ficus species are found inside an urn-shaped inflorescence (syconium). The development of the syconium traditionally has been divided into 5 phases (Galil & Eisikowich, 1968; Ibarra-Manríquez et al., 2012; Verkerke, 1989): A) floral or female, B) receptive or female, C) interfloral, D) male), and E) post floral. Pollinators can only access the flowers during phase B, through the ostiole, which is an apical opening in the syconium made up of numerous overlapping bracts (Castro-Cárdenas et al., 2022; Machado et al., 2013; van Noort & Compton, 1996; Verkerke, 1989). The ostiole plays a fundamental role in the specificity of pollinating wasps among the different sections of the genus Ficus, since it acts as a physical barrier, blocking the entry of non-pollinating wasps and/or regulating the size of the pollinating wasps that are able to reach the syconial cavity (Chen et al., 2001; Janzen, 1979; Jousselin et al., 2003; Liu et al., 2011; Ramírez, 1974; Verkerke, 1989).
One of the morphological modifications recorded for Agaonidae wasps was suggested by Ramírez (1974), who proposed that the shape of the head of female wasps presents various modifications that facilitate their passage through the ostiole, indicating that heads that are subquadrangular or subhemispherically flattened (usually as long as broad across the eyes) have been recorded in Ficus species in different groups (e.g., subgen. Ficus or the sections Americanae or Conosycea), whose syconia present spirally arranged ostiolar bracts. On the other hand, wasps with thin elongated heads have been recorded in Ficus species possessing syconia with linearly arranged ostiole bracts, which mostly point downwards (e.g., sections Galoglychia or Pharmacosycea).
van Noort & Compton (1996) evaluated the convergence in head shape between wasps from 2 subfamilies (Agaonidae and Sycoecinae), pollinators and non-pollinators, respectively, of the same Ficus species (subg. Urostigma sect. Galoglychia). These authors suggested that selection would have favored convergence in the shape of the head of both lineages of wasps, due to the pressure derived from achieving a successful entry of pollinating females into the syconium cavity. Furthermore, they emphasized that the length of the ostiole is the main factor in determining this trait in wasps, since the arrangement of the ostiole bracts in sect. Galoglychia is constant between species (Verkerke, 1989). However, the authors suggested that confirmation of their results required obtaining measurements of the ostiole during phase B of syconium development since the measurements used in the analysis were obtained from taxonomic literature. This is an important proposal, considering that there are records of Ficus species in which the size of the syconium differ among the different stages of development (Delgado-Pérez et al., 2020; Piedra-Malagón et al., 2019). Finally, Jousselin et al. (2003) reconstructed the evolution of the shape of the ostiole (spiral or linear) and the shape of the head of pollinating wasps (short or elongate) using molecular markers only (ITS and ETS) and concluded that both traits are correlated throughout the genus Ficus.
The genus Ficus is represented in the Neotropics by the subgenera Pharmacosycea (Miq.) Miq. and Spherosuke Raf. (Berg, 1989; Pederneiras et al., 2015), whose sections are endemic to the American continent (Pharmacosycea (Miq.) Griseb. and Americanae (Miq.) Corner, respecti- vely). The diversity of section Pharmacosycea is estimated to be 20 species, which are exclusively pollinated by wasps of the genus Tetrapus Mayr 1885, whose heads are longer than wide. The sect. Americanae includes a larger number of species (ca. 120) whose pollinators belong to the genus Pegoscapus Cameron 1906, which possess subquadrangular or subhemispherical heads (Berg, 1989; Jousselin et al., 2003; Ramírez, 1974).
The main objective of the present study was to determine the relationship between the size of the ostiole in species belonging to the sections of Ficus and the shape of the heads of pollinating wasps, a relationship that has not been previously analyzed quantitatively. The inclusion of Ficus species belonging to the 2 Neotropical sections is relevant, since it will allow a further evaluation between fig and fig wasp traits from a phylogenetic context (Jousselin et al., 2003).
Materials and methods
Fieldwork was carried out in the NW portion of the Los Tuxtlas region in the state of Veracruz, Mexico (Fig. 1). This region forms part of the Upper Tertiary and Middle Quaternary volcanic complex and is composed of basaltic andesite and basalt rock (Verma et al., 1993) and is bordered by Holocene lava flows (Nelson & Gonzalez-Caver, 1992). The study area reaches elevations between 100 and 600 m. The predominant climate is warm and humid, with rain all year round (A), an average annual temperature between 24.1 and 25 °C, and an average annual precipitation of 1,939 to 3,928 mm (Fig. 2; SMN, 2010). Rainfall is more frequent and abundant during the summer, whereas a drier season occurs between March to May, the latter being the driest month (SMN, 2010; Soto, 2004). The species richness of vascular plants documented for the Los Tuxtlas region is 2,548 species (2,391 native; 307 are endemic to Mexico), distributed in 208 families and 1,018 genera (Villaseñor et al., 2018). The families with the highest number of species are Fabaceae (212), Orchidaceae (171), Asteraceae (150), Rubiaceae (143), and Poaceae (82). Villaseñor et al. (2018) point out that the predominant biome in the region is the humid tropical forest. The location of the individuals of Ficus sampled in the present study are indicated in Figure 1.
The collection and measurement of syconia and pollinating wasps. Three species were chosen from each section of Ficus, which represented the size range of the syconia described for both sections in Mexico (Ibarra-Manríquez et al., 2012): F. apollinaris Dugand, F. insipida Willd., and F. yoponensis Desv. from sect. Phamacosycea, and F. colubrinae Standl., F. isophlebia Standl., and F. obtusifolia Kunth from sect. Americanae. Detailed descriptions and illustrations of the species can be consulted in Ibarra-Manríquez et al. (2012), Cornejo-Tenorio et al. (2019), and Hernández-Esquivel et al. (2020).
The material collected included 5 individuals per Ficus species with syconia in the development phases B and D (Galil & Eisikowitch, 1968; Ibarra-Manríquez et al., 2012). The collected syconia for phase B allowed us to measure the size of the ostiole at the time when the pollinating wasps enter the syconium, whereas for the syconium in phase D (male phase), we were able to capture the emerging pollinating wasps. In phase B syconia, the ostioles were dissected and fixed and stored in 70% ethanol. Phase D syconia were placed in airtight bags until the pollinating wasps emerged and then each wasp was collected with a small brush, fixed, and stored in 70% ethanol. Wasps were identified to genus level using identification keys (Bouček, 1993; Rasplus & Soldati, 2006) and classified into morphospecies (Fig. 3). We measured the ostioles for a total of 150 syconia and 170 individuals of pollinating wasps.
Figure 1. Study site localization and sampling area. A) Veracruz state (black area) in Mexico; B) location of the Los Tuxtlas Region (black dot) within Veracruz, Mexico; C) syconium and wasps’ collection points are shown in the Los Tuxtlas Region and a zoom of the vegetation cover is shown in areas for each species included in the study (D – I). Points were placed on base maps (OpenStreetMap and World Imagery) of ArcGIS® software.
The size of the ostiole was recorded using 3 measurements: 2 diameters of the area that is covered by the superficial ostiolar bracts (Fig. 4A, B), and the length, which indicates the distance between the internal and superficial ostiolar bracts (Fig. 4C, D). The head of pollinating wasp individuals were dissected and measured under a Zeiss stereoscopic microscope model Axio Zoom V16. We recorded the length (the distance between the posterior margin of the head and the apex of the jaw) and the width at maximum eye level (Fig. 4E, F). Both measurements allowed us to determine the shape of the wasp’s head (average head length to head width ratio), as described by van Noort & Compton (1996).
Data analysis. We calculated the Pearson correlation coefficients between diameter 1, diameter 2, and length of the ostiole. We also applied generalized linear models and carried out multiple comparisons using the ‘multcomp’ package to assess differences in ostiole diameter and Agaonidae wasps’ length/width ratio among species of each section. We performed a Nonmetric Multidimensional Scaling (NMDS) analysis to examine the multivariate relationship of the ostiole proportions (diameters and length) and head size (length and width) among species of each section of Ficus. We applied the decostand function of ‘Vegan’ to obtain a standardized matrix constructed with values to reduce the extremes values effect (Oksanen et al., 2019). We implemented NMDS with Bray-Curtis’s dissimilarity. Finally, we added convex hulls with the ordihull function of ‘vegan’ to highlight point clusters based on species. Since NMDS uses rank order information, this technique provides a highly flexible quantitative method that allows distribution-free inferences and, because it has a high flexibility in the number of observations, it is particularly useful with unbalanced experimental designs to explore the proximities and resemblances in the structure of the data among groups (Tong, 1992). Therefore, these analyses allowed us to identify the structure and association degree of ostiole-wasp heads among species of each section. All the analyses were conducted using R software.
Results
We found a positive correlation between the 2 diameters of the ostiole, both for sect. Americanae (0.99, p < 0.000) and for sect. Pharmacosycea (0.96, p < 0.001). In the latter section, a high positive correlation was also obtained between the length of the ostiole and both diameters (0.93, p < 0.000), a relationship that was lower for sect. Americanae (diameter 1, 0.37 and diameter 2, 0.39, p < 0.01). Figure 5 shows that the diameter of the ostiole differs significantly between the 3 species included in each section (Americanae, F2, 73 = 2156922, p < 0.000; Pharmacosycea, F2, 74 = 3.81, p < 0.000). The same result was found for the species from both sections when considering the length of the ostiole (Americanae, F2, 71 = 0.74, p < 0.000; Pharmacosycea, F2, 74 = 1.88, p < 0.000).
Figure 2. Ombrothermal diagrams of meteorological stations in the Los Tuxtlas Region, Veracruz. The data correspond to 30 years of records from the SMN (2010).
Figure 3. Heads of pollinating wasps. A) Pegoscapus sp. 1 (Ficus colubrinae); B) Pegoscapus sp. 2 (F. isophlebia); C) Pegoscapus sp. 3 (F. obtusifolia); D) Tetrapus sp. 1 (F. apollinaris); E) Tetrapus sp. 2 (F. insipida); F) Tetrapus sp. 3 (F. yoponensis). Scale bar: 200 μm.
Figure 4. Variables considered in the study. A-B) Frontal view of the ostiole; D1, diameter 1 and D2, diameter 2; C-D) longitudinal section of the ostiole; E-F) frontal view of the head of pollinating wasps; L, length of the head (from the protuberances of the clypeal margin to the mandible) and W, width of the head (Interocular distance). A, C) Ficus colubrinae; B) F. yoponensis; D) F. insipida; E) Pegoscapus sp. 1; F) Tetrapus sp. 1. Scale bars: a, c, d = 500 μm, b = 250 μm, e = 50 μm, f = 100 μm.
Figure 5. Diameter of the ostiole in the Americanae and Pharmacosycea sections. Syconia by species (25). Different letters indicate significant differences at a p < 0.001 level.
A positive correlation was recorded between the length and width of the head of the pollinating wasps in both genera; however, the relationship was greater in Pegoscapus than in Tetrapus (rho = 0.85 and 0.75, respectively). The wasps in this latter genus have elongated heads and differ from those recorded in the Pegoscapus species, which are subquadrangular (Fig. 6). For the case of sect. Americanae, the head proportion of F. isophlebia wasps differed significantly from F. colubrinae and F. obtusifolia (F2, 37 = 0.079, p < 0.000), while for sect. Pharmacosycea, differences were also found between the 3 species analyzed (F2, 74 = 0.291, p < 0.000) (Fig. 6).
In the correlation analysis between the size of the ostiole (diameter 1 and length) and the shape of the wasp’s head, sect. Americanae only showed significance for ostiole length (0.6, p < 0.000), while for Pharmacosycea both ostiole variables are correlated, although with low values (diameter, 0.35, p < 0.01; length – 0.25, p < 0.05). The ordination analysis (NMDS) reflects associations by species (Fig. 7). For sect. Americanae (stress = 0.01), the species with smaller ostioles (F. colubrinae and F. isophlebia) are found at 1 end of the ordination, with a greater width in the dispersion of the data for F. isophlebia. Likewise, in sect. Pharmacosycea (stress = 0.02), the species with the smallest ostiole size (F. apollinaris and F. yoponensis) are located at 1 end of the ordination, distant from F. insipida, which shows greater variation in the traits analyzed.
Figure 6. Head length/width of wasps by section. Distict letters indicate significant differences at p < 0.001 level.
Discussion
The present study is the first to quantitatively analyze the relationship between the dimensions of the ostiole and the shape of the head of pollinating wasps for the Neotropical sections of the genus Ficus and it contributes to support the reciprocal evolution in their mutualism. Here, it is important to point out that our results were obtained during phase B of syconia development, which was not the case in the study by van Noort & Compton (1996), who used measurements of syconia size from the literature. In addition to the above, Castro-Cárdenas et al. (2022) hypothesized that the ostiole could play a selective filter role that only allows the entry of a particular pollinating wasp morphospecies into the syconium, as has been suggested in previous studies (Dunn, 2020; Herre, 1989; Jandér et al., 2016; Liu et al., 2011). The differences found for the diameter and length of the ostioles among the species studied in both sections (Fig. 5) support this hypothesis.
As regards to the shape of the heads in the Agaonidae wasps proposed by Ramírez (1974), for the genera recorded in America, that information coincides with our results, since the species belonging to the genus Pegoscapus presented subquadrangular heads, while those of Tetrapus were clearly more elongated than wide (Figs. 3, 6). With respect to the 2 categories in wasp head shape (short heads, with a ratio ≥ 1; elongate, ratio ≤ 1), proposed by Jousselin et al. (2003), the ranges of variation found in the species evaluated (Americanae, 0.9-1.2; Pharmacosycea, 0.9-1.6), do not coincide exactly, although short and elongate are equivalent terms to those proposed by Ramírez (1974). The close relationship between the metrics of the ostiole and the head of the pollinating wasps in the American sections (Fig. 7) coincides with previous studies of a reciprocal evolution between both mutualistic partners (Cook & Segar, 2010; Galil, 1977; Ramírez, 1974; Weiblen, 2002). Furthermore, interspecific variation in head shape suggests a closer modification, at the species level, in response to ostiole morphology, as has been argued in various previous studies (Cook & Rasplus, 2003; Cook & Segar, 2010; Jousselin et al., 2003; Liu et al., 2011; Ramírez, 1974; Rasplus et al., 2020; van Noort & Compton, 1996). Similar patterns have been reported for other genera of the family Agaonidae, such as Alfonsiella Waterston, Allotriozoon Grandi, Courtella Kieffer, Elisabethiella Grandi, and Pleistodontes Saunders (Dunn et al., 2008; van Noort & Compton, 1996).
The close correlation recorded between the shape of the head of the pollinating wasps and the size of the ostiole coincides with that proposed by Cruaud et al. (2012) and van Noort & Compton (1996), who indicate that these traits are part of a complex mechanism (the lock-and-key shapes of fig ostioles and wasp heads), that determines the specificity between Ficus species and their pollinating wasps. As previously mentioned, this mechanism may explain the role of the ostiole, not only as a physical filter for pollinators but also as a chemical filter, since it is associated with the emission of volatiles for the attraction of legitimate pollinating wasps (Castro-Cárdenas et al., 2022; Okamoto & Su, 2021; Wang et al., 2013). This mechanism is considerably important since it has been shown that closely related species can emit different volatile compounds to attract their pollinating wasps (Okamoto & Su, 2021; Oldenbeuving et al., 2023; Souto-Vilarós et al., 2018).
Figure 7. Grouping based on the characteristics of the diameters and length of the ostiole and the shape of the head of the wasps in the species of the Ficus sections included in the study.
A critical aspect for the advancement in the understanding of the connection between the head shape of pollinating wasps and the dimensions of the ostiole is to carry out studies with a greater geographic scope, since the taxonomy of Ficus species in the Neotropical sections is not completely resolved (Berg, 1989; Ibarra-Manríquez et al., 2012). Global studies are required to resolve the taxonomic complexes that have been detected (Berg, 2007; Hernández-Esquivel et al., 2020; Pederneiras et al., 2023). These studies need to be performed together with a precise identification of the pollinating wasps, since there is evidence of Ficus species with more than 1 wasp pollinator (Jackson et al., 2008; Machado et al., 2005; Molbo et al., 2003; Su et al., 2008), some of which can be cryptic (Haine et al., 2006; Molbo et al., 2003). Future studies where these elements can be integrated are thus required to validate and estimate the possible contribution of each trait in this intriguing mutualism.
Acknowledgements
The first author acknowledges scholarship provided by Consejo Nacional de Humanidades, Ciencias y Tecnologías (Conahcyt) and the DGAPA-UNAM (Dirección General de Asuntos del Personal Académico – Universidad Nacional Autónoma de México) for the award of a postdoctoral fellowship grant. A. Navarrete-Segueda thanks the support received from the Consejo Nacional de Humanidades, Ciencias y Tecnologías through the 2022 (1) postdoctoral fellowship grant of the “Estancias Posdoctorales por México – Académica”. We thank Ma. Guadalupe Cornejo Tenorio for her help in measuring the wasp heads and Iván Leonardo Ek Rodríguez for their collaboration in the fieldwork. We are grateful to Silvia Espinosa Matías and Orlando Hernández Cristóbal for technical support with SEM pictures. Thanks to the Laboratorio Nacional de Análisis y Síntesis Ecológica (LANASE) of the ENES Morelia, UNAM, for their microscopy facilities and the Estación de Biología Tropical de Los Tuxtlas, Instituto de Biología, UNAM, for logistical support. We appreciate the support of Rosamond Coates in translating the English version of the manuscript. Finally, we are also grateful to two reviewers for their valuable comments to improve our manuscript.
Berg, C. C. (2007). Proposals for treating four species complexes in Ficus subgenus Urostigma section Americanae (Moraceae). Blumea, 52, 295–312. https://doi.org/10.3767/000651907X609034
Bronstein, J. L. (1988). Mutualism, antagonism, and the fig-pollinator interaction. Ecology, 69, 1298-1302. https://doi.org/10.2307/1941287
Borges, R. M. (2021). Interactions between figs and gall-inducing fig wasps: Adaptations, constraints, and unanswered questions. Frontiers in Ecology and Evolution, 9, 685542. https://doi.org/10.3389/fevo.2021.685542
Castro-Cárdenas, N., Vázquez-Santana, S., Teixeira, S. P., & Ibarra-Manríquez, G. (2022). The roles of the ostiole in the fig-fig wasp mutualism from a morpho-anatomical perspective. Journal of Plant Research, 135, 739–755. https://doi.org/10.1007/s10265-022-01413-9
Chen, Y.-R., Chou, L.-S., & Wu, W.-J. (2001). Regulation of fig wasps entry and egress: The role of ostiole of Ficusmicrocarpa L. Formosan Entomology, 21, 171–182, https://doi.org/10.6661/TESFE.2001014
Clement, W. L., Bruun Lund, S., Cohen, A., Kjellberg, F., Weiblen, G. D., & Rønsted, N. (2020). Evolution and classification of figs (Ficus) and their close relatives (Castilleae) united by involucral bracts. Botanical Journal of the Linnean Society, 193, 316–339. https://doi.org/10.1093/botlinnean/boaa022
Cook, J. M., & Rasplus J.-Y. (2003). Mutualists with attitude: Coevolving fig wasps and figs. Trends in Ecology & Evolution, 18, 241–248. https://doi.org/10.1016/S0169-5347(03)00062-4
Cruaud, A., Ronsted N., Chantarasuwan, B., Chou, L. S., Clement, W. L., Couloux, A. et al. (2012). An extreme case of plant-insect codiversification: figs and fig-pollinating wasps. Systematic Biology, 61, 1029–1047. https://doi.org/10.1093/sysbio/sys068
Cornejo-Tenorio, G., Ibarra-Manríquez, G., & Sinaca-Colín, S. (2019). Flora de Los Tuxtlas. Guía ilustrada. Ciudad de México: Universidad Nacional Autónoma de México.
Delgado-Pérez, G., Vázquez-Santana, S., Cornejo-Tenorio, G., & Ibarra-Manríquez, G. (2020). Morfoanatomía de las fases de desarrollo del sicono de Ficus tuerckheimii (subg. Spherosuke, sect. Americanae, Moraceae). Botanical Sciences, 98, 570–583. https://doi.org/10.17129/botsci.2631
Dunn, D. W. (2020). Stability in fig tree-fig wasp mutualisms: How to be a cooperative fig wasp. Biological Journal of the Linnean Society, 130, 1–17. https://doi.org/10.1093/biolinnean/blaa027
Dunn, D. W., Yu, D. W., Ridley, J., & Cook, J. M. (2008). Longevity, early emergence and body size in a pollinating fig wasp – Implications for stability in a fig-pollinator mutualism. Journal of Animal Ecology, 77, 927–935. https://doi.org/10.1111/j.1365-2656.2008.01416.x
Galil, J., & Eisikowitch, D. (1968). On the pollination ecology of Ficussycomorus in East Africa. Ecology, 49, 259–269. https://doi.org/10.2307/1934454
Haine, E. R., Martin, J., & Cook, J. M. (2006). Deep mtDNA divergences indicate cryptic species in a fig-pollinating wasp. BMC Evolutionary Biology, 6, 83. https://doi.org/10.1186/1471-2148-6-83
Hernández-Esquivel, K, B., Piedra-Malagón, E. M., Cornejo-Tenorio, G., Mendoza-Cuenca, L., González-Rodríguez, A., Ruiz-Sánchez, E. et al. (2020). Unraveling the extreme morphological variation in the neotropical Ficusaurea complex (subg. Spherosuke, sect. Americanae, Moraceae). Journal of Systematics and Evolution, 58, 263–281. https://doi.org/10.1111/jse.12564
Herre, E. A. (1989). Coevolution of reproductive characteristics in 12 species of New World figs and their pollinator wasps. Experientia, 45, 637–647. https://doi.org/10.1007/BF01975680
Herre, E. A., Jandér, K. C., & Machado, C. A. (2008). Evolutionary ecology of figs and their associates: recent progress and outstanding puzzles. Annual Review of Ecology, Evolution, and Systematics, 39, 439–458. https://doi.org/10.1146/annurev.ecolsys.37.091305.110232
Ibarra-Manríquez, G., Cornejo-Tenorio, G., González-Castañeda, N., Piedra-Malagón, E. M., & Luna, A. (2012). El género Ficus L. (Moraceae) en México. Botanical Sciences, 90, 389–452. https://doi.org/10.17129/botsci.472
Jackson, A. P., Machado, C. A., Robbins, N., & Herre, E. A. (2008). Multi-locus phylogenetic analysis of Neotropical figs does not support co-speciation with the pollinators: the importance of systematic scale in fig/wasp cophylogenetic studies. Symbiosis, 45, 57–72.
Jandér, K. C., Dafoe, A., & Herre, E. A. (2016). Fitness reduction for uncooperative fig wasps through reduced offspring size: A third component of host sanctions. Ecology, 97, 2491–2500. https://doi.org/10.1002/ecy.1471
Jousselin, E., Rasplus, J. Y., & Kjellberg, F. (2003). Convergence and coevolution in a mutualism: Evidence from a molecular phylogeny of Ficus. Evolution, 57, 1255–1269. https://doi.org/10.1111/j.0014-3820.2003.tb00334.x
Liu, C., Yang, D.-R., & Peng, Y.-Q. (2011). Body size in a pollinating fig wasp and implications for stability in a fig-pollinator mutualism. Entomologia Experimentalis et Applicata, 138, 249–255. https://doi.org/10.1111/j.1570-7458.2011.01096.x
Machado, A. F. P., de Souza, A. M., & Leitão, C. A. E. (2013). Secretory structures at syconia and flowers of Ficusenormis (Moraceae): A specialization at ostiolar bracts and the first report of inflorescence colleters. Flora, 208, 45–51. https://doi.org/10.1016/j.flora.2012.12.005
Machado, C. A., Robbins, N., Gilbert, M. T. P., & Herre, E. A. (2005). Critical review of host specificity and its coevolutio- nary implications in the fig/fig-wasp mutualism. Proceedings of the National Academy of Sciences, 102 (suppl._1), 6558–6565. https://doi.org/10.1073/pnas.0501840102
Molbo, D., Machado, C. A., Sevenster, J. G., Keller, L., & Herre, E. A. (2003). Cryptic species of fig-pollinating wasps: Implications for the evolution of the fig-wasp mutualism, sex allocation, and precision of adaptation. Proceedings of the National Academy of Sciences, 100, 5867–5872. https://doi.org/10.1073/pnas.0930903100
Nelson, S. A., & Gonzalez-Caver, E. (1992). Geology and K-Ar dating of the Tuxtla Volcanic Field, Veracruz, Mexico. Bulletin of Volcanology, 55, 85–96. https://doi.org/10.1007/BF00301122
Okamoto, T., & Su, Z.-H. (2021). Chemical analysis of floral scents in sympatric Ficus species: highlighting different compositions of floral scents in morphologically and phylogenetically close species. Plant Systematics and Evolution, 307, 1–12. https://doi.org/10.1007/s00606-021-01767-y
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D. et al. (2019). Vegan: Community ecology package. In R package version 2.5-6 (p. 296). https://doi.org/10.4135/9781412971874.n145
Oldenbeuving, A., Gómez-Zúniga, A., Florez-Buitrago, X., Gutiérrez-Zuluaga, A. M., Machado, C. A., Van Dooren, T. J. M. et al. (2023). Field sampling of fig pollinator wasps across host species and host developmental phase: Implications for host recognition and specificity. Ecology and Evolution, 13, e10501. https://doi.org/10.1002/ece3.10501
Pederneiras, L. C., Carauta, J. P. P., Neto, S. R., & de Freitas, V. (2015). An overview of the infrageneric nomenclature of Ficus (Moraceae). Taxon, 64, 589–594. http://dx.doi.org/10.12705/643.12
Pederneiras, L. C., Zamengo, H. B., Plata-Castro, A. D., Romaniuc-Neto, S., & de Freitas, V. (2023). Ficus sect. Americanae ser. Kinuppii: a ramiflorous group of Neotropical fig trees. Brittonia, 75, 249-268. https://doi.org/10.1007/s12228-023-09750-2
Peng, Y. Q., Duan, Z. B., Yang, D. R., & Rasplus, J. Y. (2008). Cooccurrence of two Eupristina species on Ficus altissima in Xishuangbanna, SW China. Symbiosis, 45, 9–14.
Piedra-Malagón, E. M., Hernández-Ramos, B., Mirón-Monterrosas, A., Cornejo-Tenorio, G., Navarrete-Segueda, A., & Ibarra-Manríquez, G. (2019). Syconium development in Ficus petiolaris (Ficus, sect. Americanae, Moraceae) and the relationship with pollinator and parasitic wasps. Botany, 97, 190–203. https://doi.org/10.1139/cjb-2018-0095
Ramírez, B. W. (1974). Coevolution of Ficus and Agaonidae. Annals of the Missouri Botanical Garden, 61, 770–780. https://doi.org/10.2307/2395028
Rasplus, J.-Y., Rodriguez, L. J., Sauné, L., Peng, Y.-Q., Bain, A., Kjellberg, F. et al. (2020). Exploring systematic biases, rooting methods and morphological evidence to unravel the evolutionary history of the genus Ficus (Moraceae). Cladistics, 37, 402–422. https://doi.org/10.1111/cla.12443
Rasplus, J. Y., & Soldati, L. (2006). Familia Agaonidae. In: F. Fernández y M. J. Sharkey, (Eds.) Introducción a los Hymenoptera de la Región Neotropical (pp. 683-698). Bogotá: Sociedad Colombiana de Entomología & Universidad Nacional de Colombia.
Rønsted, N., Weiblen, G. D., Cook, J. M., Salamin, N., Machado, C. A., & Savolainen, V. (2005). 60 million years of co-divergence in the fig-wasp symbiosis. Proceedings of the Royal Society London B, 272, 2593–2599. https://doi.org/10.1098/rspb.2005.3249
SMN (Servicio Metereológico Nacional). (2010). Información climatológica por estado (document on line). Servicio Meteorológico Nacional. Available at: https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado?estado=ver
Soto, M. (2004). El Clima. In S. Guevara, J. Laborde, & G. Sánchez-Río (Eds.), Los Tuxtlas. El paisaje de la sierra. Xalapa: Instituto de Ecología A.C./ Unión Europea.
Souto-Vilarós, D., Proffit, M., Buatois, B., Rindos, M., Sisol, M., Kuyaiva, T. et al. (2018). Pollination along an elevational gradient mediated both by floral scent and pollinator compatibility in the fig and fig-wasp mutualism. Journal of Ecology, 106, 2256–2273. https://doi.org/10.1111/1365-2745.12995
Su, Z. H., Iino, H., Nakamura, K., Serrato, A., & Oyama, K. (2008). Breakdown of the one-to-one rule in Mexican fig-wasp associations inferred by molecular phylogenetic analysis. Symbiosis, 45, 73–81.
Tong, S. T. Y. (1992). The use of non-metric multidimensional scaling as an ordination technique in resource survey and evaluation: A case study from southeast Spain. Applied Geography, 12, 243–260. https://doi.org/10.1016/0143-6228(92)90042-L
van Noort, S., & Compton, S. G. (1996). Convergent evolution of agaonine and sycoecine (Agaonidae, Chalcidoidea) head shape in response to the constraints of host fig morphology. Journal of Biogeography, 23, 415–424. https://doi.org/10.1111/j.1365-2699.1996.tb00003.x
Verma, S. P., Salazar-V., A., Negendack, J. F. W., Milán, M., Navarro, L. I., & Besch, T. (1993). Características petrográfica y geoquímicas de elementos mayores del Campo Volcánico de Los Tuxtlas, Veracruz, México. Geofísica Internacional, 32, 237–248. https://doi.org/10.22201/igeof.00167169p.1993.32.2.558
Villaseñor, J. L., Ortiz, E., & Campos-Villanueva, A. (2018). High richness of vascular plants in the tropical Los Tuxtlas Region, Mexico. Tropical Conservation Science, 11, 1–12. https://doi.org/10.1177/1940082918764259
Wang, A. Y., Peng, Y.-Q., Harder, L. D., Huang, J.-F., Yang, D.-R., Zhang, D.-Y. et al. (2019). The nature of interspecific interactions and co-diversification patterns, as illustrated by the fig microcosm. New Phytologist, 224, 1304–1315. https://doi.org/10.1111/nph.16176
Wang, G., Compton, S. G., & Chen, J. (2013). The mechanism of pollinator specificity between two sympatric fig varieties: a combination of olfactory signals and contact cues. Annals of Botany, 111, 173–181. https://doi.org/10.1093/aob/mcs250
Zhang, Q., Onstein, R. E., Little, S. A., & Sauquet, H. (2019). Estimating divergence times and ancestral breeding systems in Ficus and Moraceae. Annals of Botany, 123, 191–204. https://doi.org/10.1093/aob/mcy159
The genus Myianoetus Oudemans(Acari: Histiostomatidae) is commonly associated with carrion, utilizing flies (Diptera) from various families as a means of dispersal through phoresy. The objective of this paper is to present a new association between Myianoetus sp. mites and Calliphoridae flies and discuss its relevance in forensic sciences. Samples were collected in 3 locations in the Prepuna ecoregion of Jujuy, Argentina. Specimens were captured using necrotraps baited with cow lung. Flies carrying phoretic mites were separated and identified to a specific level, while mites were counted and identified at the lowest possible taxonomic level. Compsomyiops fulvicrura (Robineau-Desvoidy) (Diptera: Calliphoridae) was the only species that presented attached mites, with an average intensity of 12.26 mites per fly. The mites carried by C. fulvicrura were identified as deutonymphs of Myianoetus sp., with a prevalence of 2.56% of infested flies. Significant differences in the abundance of flies with mites were observed between locations and seasons. This article represents the first contribution to knowledge on the specific association between Myianoetus sp. and C. fulvicrura. These findings in forensic ecology are relevant for their potential contribution and application in the development of more precise methods in specific forensic cases.
Keywords: Astigmata; Diptera; Forensic Acarology; Phoresy; New report
Asociación de Myianoetus sp. (Acari: Histiostomatidae) con la mosca necrófaga Compsomyiops fulvicrura (Diptera: Calliphoridae), en la ecoregión Prepuna (Jujuy: Argentina)
Resumen
El género Myianoetus Oudemans (Acari: Histiostomatidae) suele asociarse a la carroña utilizando moscas (Diptera) de distintas familias como medio de dispersión, a través de la foresia. El objetivo de este trabajo fue presentar una nueva asociación entre Myianoetus sp. con moscas Calliphoridae y discutir su alcance dentro de las ciencias forenses. Las muestras se recolectaron en 3 localidades de la Prepuna jujeña, Jujuy, Argentina. Los especímenes se capturaron mediante necrotrampas cebadas con pulmón vacuno. Las moscas con ácaros se separaron y determinaron a nivel específico; los ácaros fueron numerados e identificados al nivel taxonómico más bajo posible. Compsomyiops fulvicrura (Diptera: Calliphoridae) fue la única especie que presentó ácaros adheridos, con una intensidad media de 12.26 ácaros por mosca. Los ácaros fueron identificados como deutoninfas de Myianoetus sp. y se determinó una prevalencia de 2.56% de moscas infestadas. Se observaron diferencias significativas en la abundancia de moscas con ácaros entre las localidades y estaciones analizadas. Este artículo representa el primer aporte al conocimiento sobre la asociación específica entre Myianoetus sp. y C. fulvicrura. Estos hallazgos sobre ecología forense son relevantes por su potencial contribución y aplicación al desarrollo de métodos más precisos en casos forenses determinados.
Palabras clave:Astigmata; Diptera; Acarología forense; Foresia; Nuevo reporte
Introduction
Carcasses present limited and ephemeral biocenosis made up of diverse organisms that often comprise complex food webs (Braig & Perotti, 2009; Perotti et al., 2010). Many Diptera species actively participate in the cadaveric decomposition process in which the Calliphoridae and Sarcophagidae families, along with Coleoptera are often investigated because of their large number, persistence and capacity to act as hosts to diverse mites that use them for dispersion by phoresis (Camerick, 2010; Perotti & Braig, 2009; Perotti et al., 2010).
Mites present morphological and physiological adaptations to serve phoresy during adult and nymphal stages. These adaptations are documented in the order Mesostigmata, in the suborder Prostigmata and in the infraorder Astigmatina (Oribatida) (Perotti et al., 2010). Astigmata mites are specialists in irregular or ephemeral habitats which they colonize through a deutonymphal heteromorphic stage known as hypopus which is specialized for phoresy (OConnor, 2009). Astigmatid deutonymphs are morphologically simplified, have lost the mouth and chelicerae, have greatly reduced the remainder of the gnathosoma, and have suckers on the paraproctal region for efficient phoretic attachment. The body is strongly dorsoventrally flattened, heavily sclerotized and much more resistant to desiccation than other stages of the life cycle (Farish & Axtell, 1971; OConnor, 1982). The conditions needed to reach this stage may involve genetic factors and physicochemical factors from the environment (Greenberg & Carpenter, 1960).
Astigmatid mites are particularly important for the 3 areas of forensic entomology: urban, stored product pests and medico-legal (Catts & Goff, 1992; Perotti & Braig, 2019). Nevertheless, they often go unnoticed because of their small size. Moreover, their analysis is limited because of difficulties in species identification, lack of specific knowledge and misuse of forensic methodology (OConnor, 2009; Perotti et al., 2010). Numerous species of mites are compulsory or facultative inhabitants of carrion. They are found not only in legal cases that involve human carcasses (Pimsler et al., 2016; Rai et al., 2020; Russell et al., 2004; Saloña-Bordas & Perotti, 2015); but also, in experimental studies concerning faunal succession in animal remains (Arnaldos et al., 2005; Barton et al., 2014; Centeno & Perotti, 1999; Heo et al., 2021).
In Argentina, the only record of the presence of phoretic mites associated with decomposing remains were the preliminary observations of Centeno and Perotti (1999), in which they found mites of the genus Myianoetus Oudemans (Astigmata: Histiostomatidae) associated with a specimen of Morellia sp. (Muscidae). In order to contribute to the further study on phoretic relations between mites and arthropods, this paper presents a new association between mites and Diptera from the Calliphoridae family in Prepuna of Jujuy, Argentina, and discusses its relevance and importance within the forensic sciences.
Materials and methods
The collection of Diptera specimens was carried out in the following locations: Tres Cruces (22°55’06.01” S, 65°35’13.58” W), Humahuaca (23°12’14.27” S, 65°20’54.90” W), and Tumbaya (23°51’27.79” S, 68°28’03.31” W) (Fig. 1a-c). These locations are part of the Monte Province, Prepuna District in the province of Jujuy, Argentina. Two sampling campaigns were carried out, one during the dry season (June, July, and August) and the other during the wet season (December, January, and February) between 2016 and 2018. Each location was equipped with 18 traps, totaling 54 traps across all sampling locations, totaling 108 traps per year.
Figure 1. Location of the study area. The map depicts the region corresponding to the Monte Province, Prepuna District, in the province of Jujuy, Argentina. Study sites are located in the following localities: a) Tres Cruces, b) Humahuaca, and c) Tumbaya (Photos by Fernández Salinas, M. L.).
To obtain specimens in good condition for identification, traps were made following Hwang and Turner (2005) (Fig. 2). A modified cone trap, based on a soft drink bottle with a baited target, was constructed. The bottle traps were assembled using two 3 L clear plastic soft drink bottles with a diameter of 11.5 cm, along with a black acrylic container 11 cm measured in diameter and 13 cm in depth. Consisted of 2 parts, the upper collection chamber and the lower bait chamber. The collection chamber was formed from the bottles cut 24 cm and 12 cm from the top respectively, one pushed inside the other (so that the bottle’s spout acts as a funnel and prevents flies from escaping). The bait chamber was made with a black container so the flies were drawn upwards, into the transparent collection chamber. The 2 halves of the trap were push-fitted together and secured by strips of waterproof adhesive tape. To facilitate the entry of flies 4 holes of approximately 0.8 cm in diameter were made around the bait chamber. A 125 cm³ plastic container with the bait was placed at the base of this container. A feeding substrate made of 100 g of cow’s lung was used. A distance of approximately 100 m was maintained between the traps because of the competitive nature of the colonizing species and were separated from the floor as they were hung at a minimum height of 1.5 m to avoid the attack of scavenger mammals. It was placed in a closed recipient which was subjected to a warm temperature between 15 °C and 30 °C, during 60 hours, for sufficient time to decompose. The traps were left in place for 7 consecutive days.
The captured specimens were put in Kahn tubes with 70% alcohol and they were transported to the Institute of Altitude Biology (INBIAL), San Salvador de Jujuy, Jujuy, Argentina.
Figure 2. Design of the bottle necrotrap baited with cow lung.
Flies that presented mites were counted, separated by sex and identified to its most specific level using keys and revisions from Olea and Mariluis (2013), Whitworth (2014), and Mulieri et al. (2014). The flies were photographed “in situ” using a Canon 5D Mark IV camera, 3 extension tubes for macro photography and a Canon 85 mm 1.8 lens illuminated with a Godox AD200 flash and a Godox V860 flash. Afterward, the specimens were sent to the Parasitic Zoonoses Laboratory, National University of Mar del Plata, Mar del Plata, Buenos Aires, Argentina. Each fly was individually examined, and the number of mites per fly and their attachment sites on the host flies were determined. The mites were then removed with the assistance of fine-tipped needles. From selected mite specimens, permanent preparations in Hoyer’s medium were made. The remaining specimens were identified from temporary preparations after being cleared in lactic acid using an open slide technique in order to be observed under the optic microscope (Olympus CX31). Taxonomic identification was done at a genus level using diagnostic keys (Dindal, 1990). Mites were photographed with a Sony Powershot DSC-P200 camera. The photographs were edited with Adobe Photoshop CS.
Table 1
Percentage of prevalence and mean intensity of Myianoetus sp. associated with Compsomyiops fulvicrura, in 3 locations of Jujuy, Argentina.
Location
Season
Nº of flies with attached mites
Total Nº of flies
Nº of mites
Prevalence (%)
Mean Intensity
Tumbaya
Dry
26
661
243
3.93
9.34
Wet
0
85
0
0
0
Humahuaca
Dry
40
1,517
478
2.63
11.95
Wet
0
205
0
0
0
Tres Cruces
Dry
18
624
156
2.88
8.83
Wet
10
582
276
1.71
27.3
The abundance of flies with attached mites was analyzed using Generalized Linear Models (GLM) through the software InfoStat (Di Rienzo et al., 2020). In the model, the 3 study locations were considered as fixed effects while the seasons were treated as random variables. Variance heterogeneity was adjusted using the VarExp variance function, and models were selected according to Akaike (AIC) and Bayesian (BIC) criteria. Subsequently, a Fisher’s LSD test (α = 0.05) of adjusted means and standard errors was conducted to evaluate differences between locations, following the methods described. Prevalence and mean intensity were calculated as indicated by Bush et al. (1997) and Margolis et al. (1982). Prevalence was calculated as the number of flies infected with phoretic mites, divided by the number of flies examined in a sample, and was expressed as a percentage. The mean intensity of phoresy was defined as the total number of phoretic mites of a particular species found in a sample, divided by the number of host flies.
For further taxonomic studies, voucher species were deposited as slide-mounted specimens in the Entomological Collection “Dr. Lilia Estela Neder”, Institute of Altitude Biology (INBIAL), National University of Jujuy, Jujuy, Argentina (INBIAL C 15000; INBIAL C 15001).
Results
A total of 9,454 Calliphoridae individuals were collected. They spanned 5 genera and 12 species: Calli- phora vicina (Robineau-Desvoidy), Chlorobrachycoma versicolor (Bigot), Chrysomya albiceps (Wiedemann), Chrysomya megacephala (Fabricius), Cochliomyia mace-llaria (Fabricius), Cochliomya hominiborax (Coquerel), Compsomyiops fulvicrura (Robineau-Desvoidy), Com-psomyiops sp., Lucilia cuprina (Wiedemann), Lucilia sericata (Meigen), Sarconesia chlorogaster (Wiedemann), Sarconesiopsis magellanica (Le Guillou). The most abundant species were C. albiceps and C. fulvicrura with 4,651and 3,674 individuals respectively. C. fulvicrura was the only species that had mites attached to its body (Fig. 3). These mites primarily attached themselves to the thorax and head regions and were identified as deutonymphs of Myianoetus sp. (Figs. 4, 5, 6). The individuals found exhibit morphological similarities to the deutonymphs of Myianoetus muscarum (Linnaeus) (OConnor et al., 2015). However, they differ from this species by possessing dorsal hysterosomal setae of approximately equal length to the exobothridial setae, unlike M. muscarum, where the hysterosomal setae are less than half the length of the exobothridial setae. Given that this characteristic is diagnostic of M. muscarum, we hypothesize that the specimens uncovered in this study may represent a yet undescribed species. Out of the total number of C. fulvicrura individuals, 94 carried phoretic mites (83 females and 11 males), representing a prevalence of 2.56% (Table 1). A total of 1,153 mites were counted, which corresponds to a mean intensity of 12.26 mites/fly (1-89 rank) (Fig. 7). The majority of mites (76%) were found during the dry season in all 3 studied locations. However, in Tres Cruces, mites were also found during the wet season (Table 1).
Table 2
Summary of generalized linear model (GLM) analysis results and model fitting parameters. Significance levels (p values) and variance function parameters, model fitting parameters including number of observations (N), Akaike information criterion (AIC), Bayesian information criterion (BIC), the log probability, the standard deviation (Sigma) and the coefficient of determination (R2) are shown.
Effects
p-value
Variance function parameters
Location
< 0.0001 **
Season
0.0105 *
-0.27 (dry)
-0.09 (wet)
Model tuning: N = 6, AIC = 31.83, BIC = 22.68, LogLik = -8.91, Sigma = 4.82, R2 = 0.74
Table 3
Results of the Fisher’s LSD test (α = 0.05): adjusted means and standard errors for the 3 locations under study. Common letters indicate that the means do not differ significantly (p > 0.05).
Location
Means
SE
Humahuaca
20.84
1.98
A
Tumbaya
6.84
1.98
B
Tres Cruces
1.16
1.98
C
The GLM analysis revealed significant differences in the abundance of flies with attached mites among the study locations (p < 0.0001) and a significant effect of seasonality (p = 0.0105) (Table 2).
Figure 3. Deutonymphs of Myianoetus sp. (yellow arrow) between the thorax and abdomen of Compsomyiops fulvicrura.
Additionally, subsequent Fisher’s LSD analysis revealed statistically different groups among the study locations. A higher mean abundance of flies with attached mites was observed in Humahuaca, followed by Tumbaya and Tres Cruces (Table 3). It is noteworthy that the highest variance parameter for the dry season (-0.27) compared to the wet season (-0.09) suggests that these differences are primarily attributed to this time of the year, between June and August.
The genus Myianoetus comprises more than 40 species widespread throughout the world (OConnor et al., 2015), most known only from deutonymphs phoretic on Diptera. In this work, the association between deutonymphs of Myianoetus sp. with C. fulvicrura is described for the first time. Up to present, there are reports of deutonymphs from the Myianoetus that have been found associated with various Diptera families: Sphaeroceridae (Fain et al., 1980), Muscidae (Centeno & Perotti, 1999; Greenberg & Carpenter 1960; Negm & Alatawi, 2011; Pimsler et al., 2016), Calliphoridae (Greenberg & Carpenter 1960; Miranda & Bermúdez, 2008) and Heleomyzidae (Zamec & Košel, 2014). Evidence obtained from lab experiments further described the phoretic interaction of the hypopi of Myianoetus muscarum with Muscina stabulans Fallen (Diptera: Muscidae), Stomoxys calcitrans Linnaeus (Diptera: Muscidae), Lucilia sericata (Diptera: Calliphoridae) and Musca domestica Linnaeus (Diptera: Muscidae) (Greenberg & Carpenter, 1960). Additionally, in a study carried out in Texas, USA, by Pimsler et al. (2016), a great number of M. muscarum individuals associated with Synthesiomyia nudiseta (Wulp) (Diptera: Muscidae) were collected in 3 human corpses.
Among the Calliphoridae species collected, C. albiceps stood out as the most abundant. However, deutonymphs of Myianoetus sp. were exclusively phoretically associated with C. fulvicrura. The statistical differences observed in the abundances of flies with attached mites among the different studied locations suggest that these were influenced by the dry season. Therefore, the preference for C. fulvicrura could be associated with seasonal variation, as it was more abundant during the dry season, contrasting with C. albiceps, which showed a preference for the wet season. These trends were notable in Tumbaya and Humahuaca, where C. albiceps was the dominant species, while in Tres Cruces, the abundance of this species was very low, with C. fulvicrura being the dominant species in both seasons in that area. Additionally, it is plausible that this choice is related to the chemical attraction of mites to volatile substances released by the puparia of C. fulvicrura, as demonstrated in the studies by Greenberg and Carpenter (1960). These observations were reflected in the prevalence values, which indicated higher values during the dry season in all 3 locations, compared to the wet season.
Figure 5. Dorsal view of Myianoetus sp. (scale = 0.05 mm). The yellow arrow indicates the hysterosomal setae (ex) and exobothridial setae (in).
Figure 6. Dorsal view of legs I and II of Myianoetus sp. (scale = 0.1 mm). The yellow arrow shows the bifurcate empodial claw, characteristic of the genus, present on legs I-III.
Figure 7. Abundance frequency (AF) histogram of Myianoetus sp. deutonymphs associated with Compsomyiops fulvicrura individuals.
The lack of interaction between Myianoetus sp. with other species and its demonstrated affinity with C. fulvicrura suggest that these mites can be phoretically selective in the Prepuna environment. At genus or species level, mites have micro-habitat specific requirements, being excellent specific environmental indicators, offering themselves as potentially one of the most informative pieces of biological trace evidence collected from a crime scene (Perotti & Braig, 2019). This may explain events of corpse location, of relocation, a link to a suspect and a possible connection between a suspect and a victim or a crime scene (Hani et al., 2018; Kamaruzaman et al., 2018; Szeleczl et al., 2018). The specificity and abundance of mites, coupled with the intensity of phoresy, could contribute to estimating more precise post-mortem intervals (PMI) (Miranda & Bermúdez, 2008; Rodrigueiro & Prado, 2004; Russell et al., 2004). In addition, Perotti and Braig (2009) suggested that the presence of a specific phoretic mite (for example Myianoetus sp.) may confirm the presence of its host (for example C. fulvicrura), even when the host is already gone.
Given that mites are a valuable forensic tool, it is crucial to deepen the understanding of the biology and ecology of the species involved. To expand this knowledge, it is necessary to continue registering and investigating new species and their phoretic associations under various climatic and biogeographical conditions.
Acknowledgements
We would like to thank Pablo A. Martínez for his critical analysis and recommendations for our manuscript, Mario A. Linares for the Myianoetus sp.photograph and Ismael Acosta for the C. fulvicrura photograph.
References
Arnaldos, M. I., García, M. D., Romera, E., Presa, J. J., & Luna, A. (2005). Estimation of post-mortem interval in real cases based on experimentally obtained entomological evidence. Forensic Science International, 149, 57–65. https://doi.org/10.1016/j.forsciint.2004.04.087
Barton, P. S., Weaver, H. J., & Manning, A. D. (2014). Contrasting diversity dynamics of phoretic mites and beetles associated with vertebrate carrion. Experimental and Applied Acarology, 63, 1–13. https://doi.org/10.1007/s10493-013-9758-7
Bush, A. O., Lafferty, K. D., Lotz, J. M., & Shostak, A. W. (1997). Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. The Journal of Parasitology, 83, 575–583. https://doi.org/10.2307/3284227
Camerik, A. M. (2010). Phoresy revisited. In M. Sabelis, & J. Bruin (Eds.), Trends in Acarology (pp. 333–336). Dordrecht: Springer. https://doi.org/10.1007/978-90-481-9837-5_53
Centeno, N. D., & Perotti, M. A. (1999). Ácaros vinculados a procesos de descomposición de cadáveres y sus posibles asociaciones foréticas. In Actas y Trabajos de la XIX Reunión Argentina de Ecología, 1999. Tucumán, Argentina.
Di Rienzo J. A., Casanoves, F., Balzarini, M. G., González, L., Tablada, M., & Robledo, C. W. (2020). InfoStat versión 2020. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
Dindal, D. L. (1990). Soil biology guide. New York: John Wiley and Sons.
Fain, A., Britt, D. P., & Molyneux, D. H. (1980). Myianoetus copromyzae sp. nov. (Acari, Astigmata, Anoetidae) phoretic on Copromyza atra (Meigen 1830) in Scotland. Journal of Natural History, 14, 401–403. https://doi.org/ 10.1080/00222938000770341
Farish, D. J., & Axtell, R. C. (1971). Phoresy redefined and examined in Macrocheles muscaedomesticae (Acarina: Macrochelidae). Acarologia, 13, 16–29.
Hani, M., Thieven, U., & Perotti, M. A. (2018). Soil bulb mites as trace evidence for the location of buried money. Forensic Science International, 292, e25–e30. https://doi.org/10.1016/j.forsciint.2018.09.016
Heo, C. C., Teel, P. D., & OConnor, B. M. (2021). Acari community in association with delayed pig carrion decomposition. Experimental and Applied Acarology, 85, 223–246. https://doi.org/10.1007/s10493-021-00676-6
Hwang, C., & Turner, B. D. (2005). Spatial and temporal variability of necrophagous Diptera from urban to rural areas. Medical and Veterinary Entomology, 19, 379–391. https://doi.org/10.1111/j.1365-2915.2005.00583.x
Kamaruzaman, N. A. C., Mašán, P., Velásquez, Y., González-Medina, A., Lindström, A., Braig, H. R. et al. (2018). Macrocheles species (Acari: Macrochelidae) associated with human corpses in Europe. Experimental and Applied Acarology, 76, 453–471. https://doi.org/10.1007/s10493-018-0321-4
Margolis, L., Esch, G. W., Holmes, J. C., Kuris, A. M., & Schad, G. A. (1982). The use of ecological terms in parasitology (report of an ad hoc committee of the American Society of Parasitologists). Journal of Parasitology, 68, 131–133. https://doi.org/10.2307/3281335
Miranda, R., & Bermúdez, S. (2008). Ácaros (Arachnida: Acari) asociados con moscas Calliphoridae (Diptera: Oestroidea) en tres localidades de Panamá. Revista Colombiana de Entomología, 34, 192–196. https://doi.org/10.25100/socolen.v34i2.9287
Mulieri, P. R., Mariluis, J. C., & Patitucci, L. D. (2014). Calliphoridae. In S. Roig-Juñent, L. E. Claps, & J. J. Morrone (Eds.), Biodiversidad de artrópodos argentinos, Vol. 4 (pp. 463–474). INSUE, Universidad Nacional de Tucumán, San Miguel de Tucumán, Argentina.
Negm, M. W., & Alatawi, F. J. (2011). Four new records of mites (Acari: Astigmata) phoretic on insects in Riyadh, Saudi Arabia. Journal of the Saudi Society of Agricultural Sciences, 10, 95–99. https://doi.org/10.1016/j.jssas.2011.04.001
OConnor, B. M. (2009). Astigmatid mites (Acari: Sarcoptiformes) of forensic interest. Experimental and Applied Acarology, 49, 125–133. https://doi.org/10.1007/s10493-009-9270-2
OConnor, B. M., Pimsler, M. L., Owings, C. G., & Tomberlin, J. K. (2015). Redescription of Myianoetus muscarum (Acari: Histiostomatidae) associated with human remains in Texas, USA, with designation of a neotype from Western Europe. Journal of Medical. Entomology, 52, 539–550. https://doi.org/10.1093/jme/tjv045
Olea, S. M., & Mariluis, J. C. (2013). The genus Calliphora (Diptera: Calliphoridae) in Argentina, with the first records of C. lopesi Mello 1962. Revista de la Sociedad Entomológica Argentina, 72, 99–104.
Perotti, M. A., & Braig, H. R. (2009). Phoretic mites associated with animal and human decomposition. Experimental and Applied Acarology, 49, 85–124. https://doi.org/10.1007/s10493-009-9280-0
Perotti, M. A., Braig, H. R., & Goff, M. L. (2010). Phoretic mites and carcasses: Acari transported by organisms associated with animal and human decomposition. In J. Amendt, M. Goff, C. Campobasso, & M. Grassberger (Eds.), Current concepts in forensic Entomology (pp. 69–91). Dordrecht: Springer. https://doi.org/10.1007/978-1-4020-9684-6_5
Perotti, M. A., & Braig, H. R. (2019). Acarology in Crimino-Legal Investigations. In J. Byrd, & J. Tomberlin (Eds.), Forensic Entomology, 3rd. Edition (pp. 461–473). Boca Raton: CRC Press. https://doi.org/10.4324/9781351163767-22
Pimsler, M. L., Owings, C. G., Sanford, M. R., OConnor, B. M., Teel, P. D., Mohr, R. M., & Tomberlin, J. K. (2016). Associa- tion of Myianoetus muscarum (Acari: Histiostomatidae) with Synthesiomyia nudiseta (Wulp) (Diptera: Muscidae) on human remains. Journal of Medical Entomology, 53, 290–295. https://doi.org/10.1093/jme/tjv203
Rai, J., Amendt, J., Bernhardt, V., Pasquerault, T., Lindström, A., & Perotti, M. A. (2020). Mites (Acari) as a relevant tool in trace evidence and postmortem analyses of buried corpses. Journal of Forensic Sciences, 65, 2174–2183. https://doi.org/10.1111/1556-4029.14506
Rodrigueiro, T. S. C., & Prado, A. P. (2004). Macrocheles muscaedomesticae (Acari, Macrochelidae) and a species of Uroseius (Acari, Polyaspididae) phoretic on Musca domestica (Diptera, Muscidae): effects on dispersal and colonization of poultry manure. Iheringia. Série Zoologia, 94, 181–185. https://doi.org/10.1590/S0073-47212004000200011
Russell, D. J., Schulz, M. M., & OConnor, B. M. (2004). Mass occurrence of astigmatid mites on human remains. Abhandlungen und Berichte des Naturkundmuseums Görlitz, 76, 51–56.
Saloña-Bordas, M. I., & Perotti, M. A. (2015). Acarología forense. Ciencia Forense, 12, 91–112.
Szelecz, I., Lösch, S., Seppey, C. V. W., Lara, E., Singer, D., Sorge, F. et al. (2018). Comparative analysis of bones, mites, soil chemistry, nematodes and soil micro-eukaryotes from a suspected homicide to estimate the post-mortem interval. Scientific Reports, 8, 25. https://doi.org/10.1038/s41598-017-18179-z
Whitworth, T. (2014). A revision of the Neotropical species of Lucilia Robineau- Desvoidy (Diptera: Calliphoridae). Zootaxa, 3810, 1–76. https://doi.org/10.11646/zootaxa.3810. 1.1
Zamec, R., & Košel, V. (2014). A new species of mite (Acari: Histiostomatidae) phoretic on Gymnomus caesius (Diptera: Heleomyzidae) from Vlčie Diery cave. Biologia, 69, 916–919. https://doi.org/10.2478/s11756-014-0387-3
Omer José Jiménez-Ortega a, d, Keiner L. Tílvez b, Joselin Castro-Palacios a, Andrés García c, *, Gabriel R. Navas a, Julio Abad Ferrer-Sotelo e, Dilia Naranjo-Calderón e, Juan Gabriel Díaz-Castellar e, Víctor Buelvas-Meléndez e
a Universidad de Cartagena, Campus San Pablo, Grupo de Investigación en Hidrobiología, Programa de Biología, Carrera 50#24-120, Zaragocilla, Cartagena de Indias, Colombia.
b Universidad de Cartagena, Campus San Pablo, Grupo de Investigación en Biología Descriptiva y Aplicada, Carrera 50#24-120, Zaragocilla, Cartagena de Indias, Colombia
c Universidad Nacional Autónoma de México, Instituto de Biología, Estación de Biología Chamela, Apartado postal 21, 48980 San Patricio, Jalisco, México
d Parque Temático Vivarium del Caribe-Fundación Archosauria zona norte km 15, Provincia de Cartagena, Bolívar, Colombia
e Santuario de Flora y Fauna Los Colorados, Parques Nacionales Naturales de Colombia, Carrera 8# 9-20 Plaza Olaya Herrera, San Juan Nepomuceno, Bolívar, Colombia
Received: 28 October 2023; accepted: 18 March 2024
Abstract
This study aimed to determine anuran diversity and the use of microhabitats in 3 vegetation covers in the Santuario de Flora y Fauna Los Colorados. Five field trips of 6 days each were made, 2 days and 2 nights in each cover: forest, pasture, and crop. Sampling was carried out with the visual encounter inspection technique under a randomized design by random walks with manual capture. A total of 19 species were recorded, 14 in the forest, 13 in pasture, and 12 in crop. Pasture and crop were the vegetation covers with the greatest similarity of species. This work updates the list of anuran species recorded in the management plan of the Santuario de Flora y Fauna Los Colorados 2018-2023. The greatest number of anuran species was associated with leaf litter, “jagüeyes”, and soils. The transformation of the landscape as a result of agriculture and cattle ranching generated changes in the richness, abundance, composition, and use of microhabitats of the anurans present in the Santuario de Flora y Fauna Los Colorados.
Diversidad de anuros y uso de microhábitats en tres coberturas vegetales del Santuario de Flora y Fauna Los Colorados, Caribe colombiano
Resumen
Este estudio tuvo como objetivo determinar la diversidad de anuros y el uso de microhábitats en 3 coberturas vegetales en el Santuario de Flora y Fauna Los Colorados. Se hicieron 5 salidas de campo de 6 días cada una, 2 días y 2 noches en cada una: bosque, potrero y cultivo. Se realizaron muestreos con la técnica de inspección por encuentro visual, bajo el diseño aleatorizado por caminatas al azar con captura manual. Se registraron 19 especies, 14 de ellas en bosque, 13 en potrero y 12 en cultivo, siendo el potrero y el cultivo las coberturas con mayor similitud de especies. Este trabajo actualiza el listado de las especies de anuros registrados en el Plan de manejo del Santuario de Flora y Fauna Los Colorados 2018-2023. El mayor número de especies de anuros se encontró asociado a la hojarasca, el jagüey y los suelos. La transformación del paisaje producto de la agricultura y la ganadería genera cambios en la riqueza, abundancia, composición y uso de microhábitats de los anuros presentes en el Santuario de Flora y Fauna Los Colorados.
Seasonally tropical dry forests (STDF here after) in Colombia are distributed mainly in the inter-Andean valleys and the Caribbean region (García et al., 2014), the latter being one of the regions with the best conserved areas of this ecosystem (Pizano et al., 2014; Rodríguez et al., 2012). However, Etter et al. (2008) point out that indiscriminate deforestation for various anthropogenic activities such as agriculture and livestock have generated large reductions in forest cover over time. This, combined with other activities such as mining and urban development (Cristal et al., 2020; Galván-Guevara et al., 2015; Jiménez et al., 2018), cause biological and ecological interactions to deteriorate, and the functionality of the ecosystem is compromised (Thomson et al., 2017), which is why Colombian STDFs have been classified as critically endangered (CR) (Etter et al., 2017). Consequently, it is a strategic ecosystem for conservation study due to its high risk of disappearing, strongly threatening the local fauna and the people who depend directly and indirectly on the ecosystem services it provides (Andrade, 2011).
One of most sensitive groups to forest transformation is amphibians, including anurans, which are highly dependent on humid places or sites with high water availability since most of their species have indirect development, permeable skin, and anamniote-type eggs (O’Malley, 2007). The spatial distribution and microhabitats use by anurans depend on the physiological requirements of each organism, and the available resources (Urbina-Cardona et al., 2006; Zug et al., 2009), as suggested by several studies showing many anuran species prefer forested areas (Cáceres-Andrade & Urbina-Cardona, 2009; García-R et al., 2005; Román-Palacios et al., 2016). Consequently, these species may be affected by anthropogenic disturbance, forest fragmentation, and loss (Cáceres-Andrade & Urbina-Cardona, 2009).
Forest transformation is among the main factors affecting anuran communities (Cáceres-Andrade & Urbina-Cardona, 2009; Marín et al., 2017; Romero, 2013; Vargas & Bolaños-L, 1999), causing around 38% of Colombian amphibians to be included under a category of endangered species and positioning Colombia as the country with the highest number of threatened species according to the second global review of amphibians (Re:wild, 2023). A study carried out by Duarte-Marín et al. (2018) in 3 habitats of the Selva de Florencia National Natural Park estimated that the covers with greater vegetation (forest and pine forest) presented greater richness and diversity of anurans than those covers with less vegetal complexity (pastures). This means land use and changes in vegetation cover are factors that influence amphibian species richness and diversity. Therefore, species that are not adapted to the new environmental conditions created by landscape transformation are eliminated from the assembly, negatively affecting the ecosystem processes in which they had intervention (Díaz et al., 2006).
Additionally, forest fragmentation has created barriers that prevent anuran dispersal, resulting in a decrease in their genetic diversity (De Sá, 2005). Furthermore, it has generated changes in the composition and abundance of anurans to an extent that depends on the levels of disturbance (Acuña-Vargas, 2016), with an increase in the penetration of light and winds along the perimeter of a forest remnants, coming from non-forest environments such as pastures, with the subsequent changes in microclimates (Echeverry et al., 2006; Galván-Guevara et al., 2015; Laurence & Gascon, 1997), phenomena known as the edge effect ( Rojas & Pérez-Peña, 2018). However, studies such as Blanco and Bonilla (2010) show that some transformed areas provide a greater number of microenvironments due to the modifications made by humans (e.g., creation of jagüeyes) and record greater richness and abundance of anurans species when compared to less transformed areas, which is known as intermediate disturbance theory (Conell, 1978). However, it must be considered that the species found in these areas have extensive plasticity to tolerate the environmental and structural gradients generated by anthropogenic disturbance, that is, they are resilient (Cáceres-Andrade & Urbina-Cardona, 2009).
Based on the above, the general objective of this research was to determine the diversity of anurans and their use of microhabitats in 3 vegetation covers within the Los Colorados Flora and Fauna Sanctuary (SFF Los Colorados), an important protected area in the Caribbean region of Colombia, which contributes to the understanding of how amphibians respond to changes in land use for agriculture and livestock, in order to provide information that can be useful for environmental entities to determine management and conservation policies for these organisms in landscape fragments.
The specific objectives are: 1) to determine the richness, abundance, diversity, and composition of anurans in 3 vegetation covers that are representative of the Los Colorados SFF; 2) to describe the use of the microhabitat by the species in each vegetation cover; 3) to analyze and compare the relationship between precipitation and environmental temperature with the richness, abundance, and diversity of species in each vegetation cover and; 4) to analyze and compare the alpha and beta diversity of anuran species in each vegetation cover.
We expect to record differences between the 3 types of vegetation cover, hypothesizing that due to the greater heterogeneity of an ecosystem in better conservation condition such as the tropical forest, it will register a greater richness and diversity of species and its species composition will differ with respect to the other covers. While the use of the microhabitat by the species will differ in each cover and will depend on the variety of natural or anthropogenic substrates existing in each site.
Materials and methods
Montes de María is a subregion of the Colombian Caribbean. It is located between the departments of Sucre and Bolívar with an area of 6,297 km2, of which 3,719 km2 belong to the department of Bolívar (Aguilera-Díaz, 2013; Herazo et al., 2017). It integrates several municipalities, among which is San Juan Nepomuceno, where the SFF Los Colorados is located at 9°56’06.7” N, 75°06’48.7” W (Fig. 1) with an area of 1,041.96 ha, an average high temperature of 28 °C and an elevation of 23 m asl (Jiménez et al., 2018). Due to the seasonality of rainfall in the region, 3 seasons can be identified, each lasting 4 months and including the dry season (December to April), the transition season (little rain, May to August), and the rainy season (abundant rain, September to November). The average precipitation is around 1,643mm with a monthly average of 137mm (Rangel & Carvajal-Cogollo, 2012).
SFF Los Colorados is composed of a small mountain system formed by sedimentary rocks, in which the largest and most important STDF relic of Montes de María is located (Jiménez et al., 2018). This ecosystem has humid forest components, which is why it is considered a place of high species diversity (IAVH, 1998). Its hydrographic system is made up of 2 streams: Cacaos and Salvador, located on the south and north sides, respectively; it also has a large number of ravines that flow into these streams (Jiménez et al., 2018). There are 6 land uses within the SFF Los Colorados (Fig. 1), which are in descending order by their percentage of coverage, forest (66.36%), agricultural areas (17.29%), pastures (12.01%), herbaceous and shrubby vegetation areas (3.51%), urbanized areas (0.80%), and mining extraction areas (0.02%). The exact age of the crop areas is unknown; this area has historically been agricultural, even before 1977 when the SFF Los Colorados was declared as a protected natural area. However, for about 10 years these areas have been in the succession stage towards shrublands because they were purchased and practically little cultivated. There are only crops at the sampling point where yam (Dioscorea) or tuber is grown. The only management that is done with these crops is slash-and-burn. With respect to livestock, none of the pasture areas in the sampling sites have more than 40 heads of livestock. No fertilizers or other types of agrochemicals or pesticides are used.
SFF Los Colorados faces 2 main problems in the conservation of their natural environments. The first is an occupancy rate close to 30% of its surface (3 neighborhoods and 11 properties). The second is the inadequate environmental planning outside the protected area that has generated a transformation of the landscape because of cattle ranching, agriculture, forest plantations, mining activities, the presence of a national highway as a limit, and the proximity to a municipal seat of 25,000 inhabitants (Jiménez et al., 2018).
A two-day prospecting visit was carried out at the SFF Los Colorados in November 2021 to inspect the site and locate the sampling points. Subsequently, 5 field trips of 6 days each were carried out (2 days and 2 nights in each cover: forest, pasture, and crop) during the months of January, February, March, April, and June 2022. In this way, sampling was carried out during the dry and transition season, that is, under conditions of no rain (February to April) or very little rain (June). In these months 2 researchers and 2 officials from the SFF Los Colorados carried out daytime (8:00 -10:00 am) and nighttime (6:00-8:00 pm) outings with a constant speed route, for a sampling effort of 160 man-hours in each coverage for a total of 480 man-hours.
The visual encounter inspection technique was used to locate and record anuran species and their abundance, under the randomized design of random walks (Crump & Scott, 2001) and manual capture of individuals (Aguirre-León, 2011; Manzanilla & Péfaur, 2000). The identification of anuran species in each cover (forest, pasture, and crop) was based on regional taxonomic keys (Ballesteros-Correa et al., 2019; Cuentas et al., 2002; Dunn, 1994), supported by field guides with photographs (Meza-Tílvez et al., 2018; Salvador & Gómez-Sánchez, 2018), and databases (Acosta-Galvis, 2021).
The 3 selected coverages were described following the CORINE Land Cover methodology adapted for Colombia (IDEAM et al., 2008) as follows: forest is an area made up mainly of tree elements of native or exotic species, trees being woody plants with a single main trunk or in some cases with several stems, which also have a defined and semi continuous canopy. In the study area, trees reach a height greater than 5m, and watercourses with a width of less than 50m were found. Pasture includes lowlands covered with grasses and some scattered trees with a height greater than 5 m, which are located on hills and flat pastures in warm climates. Crops are areas dedicated primarily to the production of food, fiber, and other raw materials with permanent, transitional, or annual crops of avocado, chili and cassava. Temporary yam crops are mainly found in the study area.
To describe microhabitat used by anurans, the number of individuals of each species observed in one of the substrate types (leaf litter, branch, trunk, sites with the presence of water, rock, soil, herbaceous or shrubby vegetation) were recorded (Cáceres-Andrade & Urbina-Cardona, 2009).
Figure 1. Location of the Los Colorados Flora and Fauna Sanctuary; source National Natural Parks of Colombia, with permission granted by SFF Los Colorados.
All observed species were photographed and at least 1 individual per species was collected, anesthetized with 2% xylocaine gel on the head and belly, and sacrificed (McDiarmid, 1994). To avoid tissue necrosis, they were prepared and fixed with 10% formalin (McDiarmid, 1994; Simmons & Muñoz-Saba, 2005), then placed in a suitable position in a container that had a white absorbent paper impregnated with 10% formalin. Distinctive characteristics were then observed. Finally, they were preserved in 70% ethanol (Cortez-F et al., 2006). The collected material was deposited in the Armando Dugand Gnecco collection of the Universidad del Atlántico, with the following catalog numbers: UARC-Am-00508, UARC-Am-00509, UARC-Am-00510, UARC-Am-00511, UARC- Am-00512, UARC-Am-00513, and UARC-Am-00514. The collecting permit was granted by the regional environmental authority called the Regional Autonomous Corporation of the Canal del Dique (Cardique), and the permit number is the resolution number 0751 of June 27, 2014. In addition, through the research endorsement approved by National Parks of Colombia, No. 20212000004933, October 25, 2021.
Information on the number of species and their abundance in each cover and climatic season was stored in an Excel. To confirm sampling was carried out on dates with the typical characteristics of the climatic seasons (rainy and dry), we graphed and compared statistically (ANOVA) the average precipitation and temperature for the months in which the sampling was carried out based on data obtained from the Institute of Hydrology, Meteorology and Environmental Studies (IDEAM) of the Guamo-Bolívar Station (Retrieved on July 19, 2022, from: http://www.ideam.gov.co/web/atencion-y-participacion-ciudadana/pqrs).
To detect significant differences in alpha diversity (richness, abundance, Simpson, Shannon), the Kruskall-Wallis or ANOVA tests were applied, depending on the normality of the data using the Shapiro-Wilk test and homogeneity of variances using Levene’s test (p < 0.05).
Alpha diversity was determined as the species richness for each coverage (Moreno, 2001), and was evaluated using Chao 1, 2, and Jack 1 estimators in EstimateS v. 9.1 (Villareal et al., 2004). In addition, bootstrap was used, which is useful to determine richness with a high number of rare species (Colwell & Coddington, 1994; Magurran, 2004). On the other hand, the diversity of anurans was estimated for each cover using the Shannon-Wiener index in the program PAST v. 4.03 (Hammer et al., 2001), and dominance using the Simpson index, where values close to 0 were considered as low levels of dominance and those close to 1 as high levels of dominance (Clarke et al., 2014).
To evaluate the turnover of anuran species between different covers (forest, pasture, and crops), the Jaccard index was used because it relates the number of shared species to the total number of exclusive species (Villareal et al., 2004). The range of values goes from 0 in the case of no shared species, to 1 when the covers have the same species composition (Moreno, 2001). From the estimator, a dendrogram was constructed in PAST v. 4.03.
To analyze the use of microhabitats, a graph was constructed where the percentage of use of each microhabitat by species and cover was established, to observe in each cover which microhabitats were most used by each species of anuran. Data were plotted in Excel.
Results
In total 1,269 individuals belonging to 19 species and 1 casual record (not included in this analysis) were recorded and grouped into 13 genera and 7 families (Table 1). Hylidae and Leptodactylidae were the families with the greatest species recorded, 8 and 6, respectively whereas only 1 species was recorded for Microhylidae and Phyllomedusidae.
Table 1
Taxonomic list and number of anuran individuals recorded in forest, crop, and pasture cover in the Los Colorados Flora and Fauna Sanctuary.
Family
Species
Forest
Crops
Pasture
Bufonidae
Rhinella horribilis (Wiegmann, 1833)
29
48
89
Rhinella humboldti (Spix, 1824)
2
97
101
Ceratophryidae
Ceratophrys calcarata (Boulenger, 1890)*
Dendrobatidae
Dendrobates truncatus (Cope, 1861, “1860”)
173
Hylidae
Boana platanera (Escalona et al., 2021)
23
3
4
Boana pugnax (Schmidt, 1857)
3
90
Dendropsophus ebraccatus (Cope, 1874)
1
Dendropsophus microcephalus (Cope, 1886)
2
4
59
Scarthyla vigilans (Solano, 1971)
3
Scinax cf. rostratus (Peters, 1863)
5
14
Scinax cf. ruber (Laurenti, 1768)
1
2
Trachycephalus typhonius (Linnaeus, 1978)
6
1
7
Leptodactylidae
Engystomops pustulosus (Cope, 1864)
190
15
12
Leptodactylus fuscus (Schneider, 1799)
2
55
Leptodactylus insularum (Barbour, 1906)
12
1
81
Leptodactylus poecilochilus (Cope, 1862)
44
Leptodactylus savagei (Heyer, 2005)
17
Pleurodema brachyops (Cope, 1869, “1868”)
38
10
Microhylidae
Elachistocleis panamensis (Dunn et al., 1948)
19
Phyllomedusidae
Phyllomedusa venusta (Duellman & Trueb, 1967)
1
5
* Species recorded casually outside the sampled coverage, which is not included in the analyses of our study.
Table 2
Richness estimators and percentages of representativeness with respect to the number of anuran species recorded in the 3 coverages of the SFF Los Colorados.
Richness estimator
Cover
Forest
Pasture
Crops
Species recorded
14
13
12
Chao 1
15.00 (86.7%)
13.00 (100.0%)
12.33 (97.3%)
Chao 2
14.68 (88.6%)
13.00 (100.0%)
12.90 (93.0%)
Jack 1
16.70 (77.8%)
13.90 (93.5%)
14.70 (81.6%)
Bootstrap
15.40 (84.3%)
13.57 (95.8%)
13.32 (90.1%)
Alpha diversity was highest in forest (14 species), followed by pasture (13 species) and crop (12 species). The species accumulation curves in the 3 coverages based on the Chao 1, Chao 2, and bootstrap estimators allowed estimating a number of species similar to that recorded in the field and an efficiency in the sampling carried out with a representativeness greater than 80%. The Jack 1 estimator for forest indicates a representativeness of 77.8%, and for pasture and crops greater than 80% (Table 2, Fig. 2). In the singleton and doubleton curves (Fig. 2), a decreasing behavior is observed for the pasture and crop covers, indicating little probability of finding new anuran species in these covers. For forest, the doubleton curve shows an ascending behavior, indicating a probability of finding more species in this cover.
Figure 2. Accumulation curves of anuran species for the 3 coverages of the Los Colorados Flora and Fauna Sanctuary.
Figure 3. Box plots of richness (A), abundance (B), Shannon Index (C), and Simpson (D) for each of the 3 vegetation covers.
Figure 4. Histogram and box plot of daily ambient temperature across the sampling months; temperature (A, B), rainfall (C).
According to the determined Shannon-Wiener index, the diversity for forest cover was 1,631, crops 1,677, and pasture 2,107. On the other hand, Simpson’s index estimated a dominance of 0.726 for the forest, crops 0.746, and pasture 0.858. When comparing the metrics recorded in each vegetation cover (Fig. 3), the pastures registered on average the greatest richness, abundance, and diversity. The average richness was similar between the crops and the forest; however, the variation was greater in the crops. In contrast, the average and variation of abundance was greater in the forest than in the crops. The average species diversity was lowest in forests, intermediate in crops, and highest in pastures. There were statically differences of all metrics among vegetation cover; richness (one-way ANOVA, F = 5.456, df = 2, p > 0.05), abundance (H(χ2) = 4.63, p > 0.05), Shannon (one-way ANOVA, F = 16.71, df = 2, p > 0.05), and Simpson (H(χ2) = 15.97, p > 0.05).
When we graph the monthly fluctuations of ambient temperature and precipitation (Fig. 4), it is evident that during the days and months of sampling, precipitation was little or none (monthly average from 1.3 mm in January to 7.4 mm in April) whereas monthly average temperature tended to increase from 21.5 °C to 23.3 °C from January to March and from 24.0 to 24.5 °C from April to June. These daily temperature records showed significant monthly differences (ANOVA, F = 67.1, df5 = 5, df2 = 79.8, p < 0.001). There were no significant monthly fluctuations with respect to precipitation (ANOVA, F = 1.78, df5 = 5, df2 = 78.8, p > 0.05). The species richness tended to be higher in April and June in all 3 vegetation covers (Fig. 5) whereas abundance was higher in the forest in March and higher for pastures and crops during April and June. Species diversity (Shannon and Simpson) in the forest was higher in March whereas in both crops and pastures it was higher in June (Fig. 5).
Based on the Jaccard similarity index, crop and pastures presented a greater degree of similarity (Fig. 6), that is, a greater number of shared species. The forest presented the greatest dissimilarity in species composition with respect to the crop and the pasture, having a greater number of unique species (D. truncatus, D. ebraccatus, L. poecilochilus, and L. savagei), which are shown in Table 1.
It was observed that the microhabitat most used by anurans in the forest was leaf litter. The species most associated with this type of microhabitat were D. truncatus and L. poecilochilus (Fig. 7); these species were only recorded in this coverage (Table 1). In the pasture, the highest record of species was found in bare soils and jagüeyes, with L. fuscus, L. insularum, and B. pugnax being the most associated with the latter, while R. horribilis and R. humboldti were observed mainly in bare soil (Fig. 8). In addition, these species presented the highest number of records of individuals in this coverage (Table 1). Finally, in the crop coverage, the microhabitat with the highest number of anuran records was bare soil (Fig. 9), this microhabitat was used most frequently by R. humboldti, R. horribilis, and P. brachyops which were the species with the highest number of individuals recorded; this microhabitat was also used by E. panamensis, which was the only species present in this cover (Table 1).
Discussion
In this study, 19 species of anurans and 1 casual record were identified, for a total of 20 species, this being a slightly smaller number than the 21 species recorded in the SFF Los Colorados 2018-2023 Management Plan (Jiménez et al., 2018). Craugastor raniformis (Boulenger, 1896), Pseudopaludicola pusilla (Ruthven, 1916), and Lectodactylus fragilis (Brocchi, 1877) were not observed in our study, possibly due to lack of sampling in some areas of the SFF Los Colorados. Their occurrence cannot be ruled out, since they were recorded by Acosta-Galvis (2012) in the Montes de María. This study reports C. raniformis in the forest, in ravines (on rocks), on leaf litter, and in shrubby vegetation; P. pusilla in crop areas and on the edge of plain forests, on sandy substrates and in cracks after rains; L. fragilis in flat areas, around seasonal ponds, and near swamps.
Scarthyla, D. ebraccatus, and L. savagei are added to the anuran fauna of the SFF Los Colorados, which shows that it is necessary to continue carrying out studies in the subregions of STDF, including the protected areas of the plains of the Caribbean region, valleys of the Magdalena and Cauca Rivers, Catatumbo, and enclaves of the Patía Valley. Amphibian diversity is poorly known due to the lack of biological studies (Urbina-Cardona et al., 2014).
Sampling carried out in the first 3 months of the year (January, February, and March) regularly corresponds to the dry season (Rangel & Carvajal-Cogollo, 2012). However, rains occurring in these months is a consequence of the effects of the La Niña phenomenon in Colombia for 2022 (Guzmán-Ferraro & García, 2022).
Figure 4. Histogram and box plot of daily ambient temperature across the sampling months; temperature (A, B), rainfall (C).
Increases were observed in the specific richness and in the recorded number of individuals as rainfall increased (especially in April and June), so it was considered that the rainfall regime prior to sampling played an important role in the observation of anurans. These increases in richness and mainly in the number of individuals are attributed to higher activity and the reproductive strategies of some species, which take advantage of the rains to reproduce and lay eggs in temporary ponds. The rains caused greater activity and detectability of some species that were observed vocalizing in small ponds that had formed and cow dams.
Figure 5. Monthly trend of average richness (A), abundance (B), and diversity (C, D) for each vegetation cover type.
Only some amplexuses were recorded but we did not record nesting or reproduction events. Some of the species have explosive activity, which generates an increase in the number of individuals, as is the case of R. horribilis and other species (Vargas-Salinas et al., 2019); some other species vocalizing included Engytomops pustulosus in some ponds and Dendropsophus microcephalus in the emerging vegetation around the cow dams. However, it is worth mentioning that the frequency and intensity of the La Niña phenomenon due to climate change could alter the reproductive times of anurans, causing many species to have early reproduction, which would bring about temporal overlaps of the species that would generate changes in the structure of the assembly (Lawyer & Morin, 1993).
Figure 6. Jaccard similarity dendrogram for the anuran samples from the SFF Los Colorados.
As we expect, there were differences of species richness among vegetation covers. The forest recorded greater richness of anurans than the productive systems (pasture and crops). This is mainly attributed to the greater availability of humid microhabitats and the vegetaion complexity, since there are species that require dense vegetation cover and abundant leaf litter. For example, the oviposition of D. truncatus occurs in humid leaf litter (Cárdenas-Ortega et al., 2019), so different studies record it abundantly in forested areas (Burbano-Yandi et al., 2016; De la Ossa et al., 2016, 2011; Posso-Peláez et al., 2017). On the other hand, pasture and crop are covers with less complexity in the vegetal structure, generating changes in the composition of the anuran assemblages (Cortés-Gómez et al., 2013), such as the reduction in richness, which is closely linked to the reproductive modes of each species (Almeida-Gomes & Rocha, 2015). These same changes in richness in covers with different degrees of disturbance have been recorded in different studies carried out in the Middle Magdalena Valley, in Meta, and in Florencia (Burbano-Yandi et al., 2016; Cáceres-Andrade & Urbina-Cardona, 2009; Duarte-Marín et al., 2018). However, total and monthly average species richness and diversity tended to be higher in the pastures and the lowest in the forests, which registered the greatest monthly variation, recorded the higher species richness and diversity in April and June. The greater diversity of species recorded in pastures may be associated with the lower complexity of the vegetation structure of this habitat, which allows anurans to be easier to detect, while the greater structural complexity of forests and crops decreases detectability of the anurans. Additionally, the presence of jagüeyes in pastures, as sites with availability of water and constant humidity necessary for the survival of the anurans, contributed to the greatest number of records of individuals in this vegetation coverage. Leptodactylus fuscus, L. insularum, and B. pugnax had greater abundance in cow dams of the pasture, since they make postures close to bodies of water (Carvajal-Cogollo et al., 2019; Ortega-Chinquilla et al., 2019).
Figure 7. Use of microhabitats by species in the forest cover of the SFF Los Colorados.
Jagüeyes are considered important for many species in disturbed areas because they permanently provide water resources, which can be used to increase water uptake and reduce evaporation rates (Urbina-Cardona et al., 2014), in addition to be used by species with reproductive modes associated with this resource (Cardozo & Caraballo, 2017). On the other hand, bare soils were mostly used by R. horribilis and R. humboldti, which are species that are commonly found in disturbed areas (Acosta-Galvis, 2012), these have physical characteristics (tuberculated skin) and physiological characteristics that allow them to adapt to exploit this microhabitat (Cáceres-Andrade & Urbina-Cardona, 2009), for this reason, they were found with greater abundance in pastures and crops.
Dendrobates truncatus, D. ebraccatus, L. poecilochilus, and L. savagei are species that were recorded only in the forest, similar to what has been reported by other anuran assemblage studies, where they are not only recorded in forested areas, but also in wetlands (Acosta-Galvis et al., 2006; Angarita et al., 2015; Burbano-Yandi et al., 2016). On the other hand, S. vigilans was only recorded in pastures; in this study the species was observed mainly around bodies of water, specifically on emergent vegetation, in sympatry with D. microcephalus (Muñoz-Guerrero et al., 2007; Fonseca- Pérez et al., 2017). Finally, E. panamensis was only present in crops, although in the study carried out by Blanco-Torres et al. (2015), it was also recorded in pastures. This is a species identified as a leaf miner (Cuentas et al., 2002), which has possibly been the reason why it was observed near the cracks produced by cassava plantations. It is due to all the above that the species similarity analysis showed that the forest differs with respect to the other 2 covers, which are noticeably more similar to each other. Just as we expected, the forest differs in species composition.
Figure 8. Use of microhabitats by species in the pasture cover of the SFF Los Colorados.
The species diversity index values reported in our study are similar to those obtained in a nearby area located in Meta, Colombia (Cáceres-Andrade and Urbina-Cardona, 2009) where they reported values of 1.4 for humid forest, 1.43 in pastures and 1.9 in sugarcane crops. On the other hand, Román-Palacios et al. (2016) estimated a low Shannon-Wiener index in the Magdalena Medio for forest and quarry (0.92 and 1.74, respectively), while for the lake, they estimated an intermediate diversity (2.03). The forest value was very far from that estimated in this work, which may indicate that the SFF Los Colorados forest has an important conservation status that benefits the anurafauna.
On the other hand, Simpson’s index estimated high dominance for the 3 plant covers. This dominance may be associated with the microhabitats they offer; for example, the forest offered important microhabitats (numerous ponds and abundant leaf litter) for the development of E. pustulosus and D. truncatus, which made these species dominant in this cover. On the other hand, the crop was dominated mainly by P. brachyops, R. humbolbti, and R. horribilis; these species have terrestrial habits, tolerant to landscape transformations and abundant open environments (Acosta-Galvis, 2012; Rodríguez-Molina, 2004). Finally, the pasture was dominated by species of the genus Rhinella and Leptodactylus, where the latter has reproductive modes associated with foam nests, allowing them to conquer and be abundant in anthropized environments (Alcaide et al., 2012).
Jaccard’s similarity analysis for the anurans of the SFF Los Colorados indicates a grouping between pasture and crop cover due to the percentage of shared species (66.6%). This result may be associated with the fact that both covers are intervened areas, with a vegetation structure different from that of the forest and host generalist species (e.g., P. brachyops, B. pugnax, L. fuscus) that can share in greater quantities, while the forest, due to the resources it offers, may have species that do not tolerate landscape transformations (e.g., D. truncatus), being restricted only to forested areas (Cáceres-Andrade & Urbina-Cardona, 2009).
Figure 9. Use of microhabitats by species in the crop cover of the SFF Los Colorados.
The results of this research indicate that the transformation of the landscape because of the agricultural economy of the Montes de María, based mainly on cultivation and the raising of animals (Aguilera-Díaz, 2013), generated changes in the wealth, abundance, composition and use of microhabitats in anuran assemblages. Therefore, this knowledge is important to create concrete tools for the management and conservation of these organisms in the protected area and its surroundings, such as maintaining native vegetation and layers of leaf litter in productive systems, conserving lentic and lotic water sources, and reducing the use of agrochemicals, among others (Urbina-Cardona et al., 2015).
This research constitutes the baseline to evaluate the long-term response of anurans to ecological restoration processes and initiatives led by the SFF Los Colorados team in transformed areas of the protected area. This research constitutes the baseline to evaluate the long-term response of anurans to ecological restoration processes and initiatives led by the SFF Los Colorados team in transformed areas of the protected area. Results that could be useful in future studies where reference ecosystems (conserved areas) and disturbed areas in the process of restoration are used, to determine if these protected areas are achieving the expected objectives and if they are contributing to the conservation of anurans (Urbina-Cardona et al., 2015). Additionally, this study updates the list of anuran species in the protected area, pointing out those to a specific coverage and those shared among covers (forest, pasture, and crops), which can be useful to define those that may be vulnerable to fragmentation of the habitat or to be included as conservation target values (VOC) of the SFF Los Colorados in the construction of future Management Plans.
Acknowledgments
We thank the Hydrobiology research group of the University of Cartagena for providing their equipment for the development of this research. Likewise, to the University of Cartagena, for the financial support through resolution number 01878 of 2022. To IDEAM for providing information on the environmental variables for 2022. To Gabriel R. Navas-S, Dr. Andrés García, and Vivarium del Caribe for financial support and suggestions for carrying out this study. Likewise, to the technical and administrative team of the Los Colorados Flora and Fauna Sanctuary for their support in sampling, logistics, loan of facilities, and management of the research guarantee. To Joselin Castro-Palacios for his support in the implementation of the methodology of this work and to Adolfo A. Mulet-Paso for his suggestions in the identification of the species. To David Gernandt, for his revision to the text which improved substantially.
References
Acosta-Galvis, A. R. (2012). Anfibios de los enclaves secos del área de influencia de los montes de María y la ciénaga de La Caimanera, departamento de Sucre, Colombia. Biota Colombiana, 13, 211–255.
Acosta-Galvis, A. R. (2021). Lista de anfibios de Colombia. BATRACHIA. Retrieved on June 17, 2021 from: http://www.batrachia.com
Acosta-Galvis, A. R., Huertas-Salgado, C., & Rada, M. (2006). Aproximación al conocimiento de los anfibios en una localidad del Magdalena medio (Departamento de Caldas, Colombia). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 30, 291–303. https://doi.org/10.18257/raccefyn.30(115).2006.2250
Acuña-Vargas, J. C. (2016). Anfibios y reptiles asociados a cinco coberturas de la tierra, municipio de Dibulla, La Guajira, Colombia. Acta Zoológica Mexicana, 32,133–146. https://doi.org/10.21829/azm.2016.322940
Aguilera-Díaz, M. M. (2013). Montes de María: una subregión de economía campesina y empresarial. Cartagena: CEER.
Aguirre-León, G. (2011). Métodos de estimación, captura y contención de anfibios y reptiles. Manual de Técnicas para el estudio de la Fauna. In S. López-González y C. López-González (Eds.), Manual de técnicas para el estudio de fauna (pp. 61–85). Querétaro: Universidad Autónoma de Querétaro, A.C.
Alcaide, A. P., Ponssa, M. L., Alcaide, F. P., & Alcaide, M. (2012). Histología de ovario en hembras vitelogénicas de Leptodactylus latinasus (Anura, Leptodactylidae). Acta Zoológica Lilloana, 56, 44–53.
Almeida-Gomes, M., & Rocha, C. F. (2015). Habitat loss reduces the diversity of frog reproductive modes in an Atlantic Forest fragmented landscape. Biotropica, 47, 113–118. https://doi.org/10.1111/btp.12168
Andrade, M. G. (2011). Estado del conocimiento de la biodiversidad en Colombia y sus amenazas. Consideraciones para fortalecer la interacción ciencia-política. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 35, 491–508.
Angarita, M. O., Montes-Correa, A. C., & Renjifo, J. M. (2015). Amphibians and reptiles of an agroforestry system in the Colombian Caribbean. Amphibian and Reptile Conservation, 8, 33–52.
Blanco, A., & Bonilla, M. (2010). Partición de microhábitats entre especies de Bufonidae y Leiuperidae (Amphibia: Anura) en áreas con bosque seco tropical de la región Caribe-Colombia. Acta Biológica Colombiana, 15,47–60.
Blanco-Torres, A., Duré, M., & Bonilla, M. A. (2015). Observaciones sobre la dieta de Elachistocleis pearsei y Elachistocleis panamensis en dos áreas intervenidas de tierras bajas del norte de Colombia. Revista Mexicana de Biodiversidad, 86,538–540. https://doi.org/10.1016/j.rmb.2015.04.031
Ballesteros-Correa, J., Vidal-Pastrana, C., & Ortega-León, A. M. (2019). Anfibios de Córdoba, Colombia. Córdoba: Fondo Editorial de Córdoba.
Burbano-Yandi, C. E., Gómez-Díaz, M. A., Gómez-Figueroa, A., Velásquez-Trujillo, D. A., & Bolívar-García, W. (2016). Ensamblaje de anfibios presentes en un bosque seco y en sistemas productivos, Valle Medio del Magdalena, Victoria y La Dorada, Caldas, Colombia. Revista de Ciencias, 20, 81–93.
Cáceres-Andrade, S. P., & Urbina-Cardona, J. N. (2009). Ensamblajes de Anuros de sistemas productivos y bosques en el Piedemonte Llanero, departamento del Meta, Colombia. Caldasia, 31, 175–195.
Cárdenas-Ortega, M. S., Gutiérrez-Cárdenas, P. D., Cifuentes-Ortegón, M., & Patiño-Gallego, A. M. (2019). Dendrobates truncatus (Cope, 1861) Rana venenosa de rayas amarillas. Catálogo de Anfibios y Reptiles de Colombia, 5, 32–41.
Cardozo, J., & Caraballo, P. (2017). Fauna anura (Amphibia: Anura) asociada a jagüeyes en dos localidades de la región Caribe colombiana. Revista Colombiana de Ciencia Animal-RECIA, 9, 39–47. https://doi.org/10.24188/recia.v9.nS.2017.519
Carvajal-Cogollo, J. E., Bernal-González, V., Paternina-Hernández, A., Muñoz-Ávila, J. N., & Vargas-Salinas, F. (2019). Uso de hábitat y reglas de ensamble: patrones y mecanismos. In M. H. Restrepo-Domínguez, E. Vera-López, Y. Bolívar-Suárez, S. G. Numpaque-Piracoca, O. Y. Acuña-Rodríguez, Z. Z. Ojeda-Pérez et al. (Eds.), Biología de los anfibios y reptiles en bosque tropical del norte de Colombia (pp. 297–338). Tunja: Editorial UPTC.
Clarke, K. R., Gorley, N. R., Somerfield, P. J., & Warwick, R. M. (2014). Change in marine communities: an approach to statistical analysis and interpretation. Plymouth: PRIMER-E.
Colwell, R. K., & Coddington, J. A. (1994). Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 345,101–118. https://doi.org/10.1098/rstb.1994.0091
Cortés-Gómez, A. M., Castro-Herrera, F., & Urbina-Cardona, J. N. (2013). Small changes in vegetation structure create great changes in amphibian ensembles in the Colombian Pacific rainforest. Tropical Conservation Science, 6, 749–769. https://doi.org/10.1177/194008291300600604
Cortez, F. C., Suárez-Mayorga, A. M., & López-López, F. J. (2006). Preparación y preservación de material científico. In A. Angulo, J. V. Rueda-Almonacid, J. V. Rodríguez-Mahecha y E. La Marca (Eds.), Técnicas de inventario y monitoreo para los anfibios de la región tropical andina (pp. 173–218). Bogotá: Panamericana Formas e Impreso S.A.
Cristal, A., Sánchez, E., Romero, J., Leyva, J., Andrade, W., & Buelvas, C. (2020). ¿Qué hemos logrado con el proyecto de Conectividades Socio-Ecosistémicas? La evolución de la metodología de las 4Ps: avances y retos. In A. Cristal, M. Peña y J. Ferrer-Sotelo (Eds.), El proyecto de conectividades Socio-Ecosistémicas en los Montes de María, 2013–2020 (pp. 63–87). Bogotá-Colombia: Fundación Herencia Ambiental Caribe.
Crump, M., & Scott, N. (2001). Relevamiento por encuentros visuales. In W. R. Heyer, M. A. Donnelly, R. W. Diarmid, L. C. Hayek, & M. S. Foster (Eds.), Medición y monitoreo de la diversidad biológica: métodos estandarizados para anfibios (pp. 80–86). Chubut, Argentina: Editorial Universitaria de la Patagonia.
Cuentas, D., Borja, R., Lynch, J. D., & Renjifo, J. M. (2002). Anuros del departamento del Atlántico y norte de Bolívar. Barranquilla: Editorial Cencys.
De Sá, R. O. (2005). Crisis global de biodiversidad: importancia de la diversidad genética y la extinción de anfibios. Agrociencia, 9, 513.
De la Ossa, J., Contreras-Gutiérrez, J. C., & Campillo-Castro, J. (2011). Censo de Dendrobates truncatus (Anura, Dendrobatidae) en la reserva forestal protectora Serranía de Coraza, Montes de María, Sucre, Colombia. Revista Colombiana de Ciencia Animal-RECIA, 3, 339–343. https://doi.org/10.24188/recia.v3.n2.2011.407
Díaz, S., Fargione, J., Chapin, F. S., & Tilman, D. (2006). Biodiversity loss threatens human well-being. Plos Biology, 4, 1300–1305. https://doi.org/10.1371/journal.pbio.0040277
Duarte-Marín, S., González-Acosta, C., & Vargas-Salinas, F. (2018). Estructura y composición de ensamblajes de anfibios en tres tipos de hábitat en el Parque Nacional Natural Selva de Florencia, Cordillera Central de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 42, 227–236. https://doi.org/10.18257/raccefyn.631
Dunn, E. R. (1944). Los géneros de anfibios y reptiles de Colombia. Caldasia, 2,497–529.
Echeverry, M. A., & Rodríguez, J. M. (2006). Análisis de un paisaje fragmentado como herramienta para la conservación de la biodiversidad en áreas de bosque seco y subhúmedo tropical en el municipio de Pereira, Risaralda Colombia. Scientia et Technica, 12, 405–410.
Etter, A., Andrade, A., Saavedra, K., Amaya, P., Cortés, J., Pacheco, C. et al. (2017). Lista Roja de Ecosistemas de Colombia, 2,1–6. https://doi.org/10.13140/RG.2.2.10861.08165
Etter, A., McAlpine, C., & Possingham, H. (2008). Historical patterns and drivers of landscape change in Colombia since 1500: a regionalized spatial approach. Annals of the Association of American Geographers, 98, 2–23. https://doi.org/10.1080/00045600701733911
Fonseca-Pérez, K. A., Molina, C., & Tárano, Z. (2017). Diet of Dendropsophus microcephalus and Scarthyla vigilans (Anura: Hylidae) at a locality in north-western Venezuela with notes on microhabitat occupation. Papéis Avulsos de Zoologia, 57,93–104. https://doi.org/10.11606/0031-1049.2017.57.07
Fox, B. J., Taylor, J. E., Fox, M. D., & Williams, C. (1997). Vegetation changes across edges of rainforest remnants. Biological Conservation, 82, 1–13. https://doi.org/10.1016/S0006-3207(97)00011-6
Galván-Guevara, S., Ballut-Dajud, G., & De La Ossa, J. (2015). Determinación de la fragmentación del bosque seco del arroyo Pechelín, Montes de María, Caribe, Colombia. Biota Colombiana, 16, 149–157.
García, R. J. C., Castro, H. F., & Cárdenas, H. H. (2005). Relación entre la distribución de anuros y variables del hábitat en el sector La Romelia del Parque Nacional Natural Munchique (Cauca, Colombia). Caldasia, 27, 299–310.
García, H., Corzo, G., Isaacs, P., & Etter, A. (2014). Distribución y estado actual de los bosques remanentes del bioma de bosque seco tropical en Colombia: insumos para su gestión. In C. Pizano, & H. García (Eds.), El bosque seco tropical en Colombia (pp. 228–251). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Guzmán-Ferraro, M., & García, G. (2022). El fenómeno de La Niña persistirá hasta casi la mitad del año. CVC. Retrieved on January 14th, 2022 from: http://www.cvc.gov.co/boletin-prensa-007-2022
Hammer, Ø., Harper, D. A., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4, 1–9.
Herazo, F., Mercado, J., & Mendoza, H. (2017). Estructura y composición florística del bosque seco tropical en los Montes de María (Sucre-Colombia). Ciencia en Desarrollo, 8,71–8.
Instituto Alexander von Humboldt. (1998). El bosque seco tropical en Colombia B-sT. Grupo de Exploraciones y Monitoreo Ambiental. Bogotá, Colombia.
IDEAM (Instituto de Hidrología, Meteorología y Estudios Ambientales), IGAC (Instituto Geográfico Agustín Codazzi), & CORMAGDALENA (Corporación Autónoma Regional del río Grande de La Magdalena). (2008). Mapa de cobertura de la Tierra Cuenca Magdalena-Cauca: Metodología CORINE Land Cover adaptada para Colombia a escala 1:100.000. Bogotá: IDEAM/ IGAC/ CORMAGDALENA.
Jiménez, B., De la Rosa, N., & Naranjo, D. (2018). Plan de manejo del Santuario de Flora y Fauna Los Colorados. Parques Nacionales Naturales de Colombia. Santa Marta: Dirección Territorial Caribe.
Laurance, W. F., & Gascon, C. (1997). How to creatively fragment a landscape. Conservation Biology, 11, 577–579.
Lawler, S., & Morin, P. (1993). Temporal overlap, competition, and priority effects in larval anurans. Ecology, 71, 174–182. https://doi.org/10.2307/1939512
Magurran, A. (2004). Measuring biological diversity. Malden: Blackwell Publishing.
Manzanilla, J., & Péfaur, J. E. (2000). Consideraciones sobre métodos y técnicas de campo para el estudio de anfibios y reptiles. Revista de Ecología Latinoamericana, 7,17–30.
Marín, A. M., Ríos, L., Ríos, L., & Almario, J. (2017). Impacto de la actividad ganadera sobre el suelo en Colombia. Ingeniería y Región, 17, 1–12. https://doi.org/10.25054/issn.2216-1325
Meza-Tílvez, K., Mulet-Paso, A., & Zambrano-Cantillo, R. (2018). Fauna del Jardín Botánico de “Guillermo Piñeres” de Cartagena, Turbaco, Colombia: Anfibios y reptiles. Versión 1. Chicago, Illinois: Field Museum.
Moreno, C. E. (2001). Métodos para medir la biodiversidad. Volumen 1. Zaragoza: CYTED, ORCYT/ UNESCO & SEA.
Muñoz-Guerrero, J., Serrano, V. H., & Ramírez-Pinilla, M. P. (2007). Uso de microhábitat, dieta y tiempo de actividad en cuatro especies simpátricas de ranas hílidas neotropicales (Anura: Hylidae). Caldasia, 29, 413–425.
McDiarmid, R. W. (1994). Preparing amphibians as scientific specimens. In W. R. Heyer, M. A. Donnelly, R. W. McDiarmid, L. C. Hayek, & M. S. Foster (Eds.), Measuring and monitoring biological diversity. standard methods for amphibians (pp: 289–296). Washington D.C.: Smithsonian Institution Press.
O’Malley, B. (2007). Anatomía y fisiología clínica de animales exóticos. Zaragoza: Servet.
Ortega-Chinquilla, J., Méndez-Narváez, J., Carvajalino-Fernández, J., & Galindo-Uribe, D. (2019). Ecofisiología. In F. Vargas-Salinas, J.A. Muñoz-Avila & M.E. Morales-Puente (Eds.), Biología de los anfibios y reptiles en bosque tropical del norte de Colombia (pp. 297–338). Tunja: Editorial UPTC.
Pizano, C., Gonzáles, R., Gonzáles, M., Castro-Lima, R., Rodríguez, N., Idárraga, A. et al. (2014). Plantas de los bosques secos de Colombia. In C. Pizano, & H. García (Eds.), El bosque seco tropical en Colombia (pp. 228–251). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Posso-Peláez, C., Blanco-Torres, A., & Gutiérrez-Moreno, L. C. (2017). Uso de microhábitats, actividad diaria y dieta de Dendrobates truncatus (Cope, 1861) (Anura: Dendrobatidae) en bosque seco tropical del norte de Colombia. Acta Zoológica Mexicana, 33, 490–502.
Rangel, C. J. O., & Carvajal-Cogollo, J. E. (2012). Clima de la región Caribe Colombiana. In J. O. Rangel (Ed.), Colombia diversidad biótica XII: la región Caribe de Colombia (pp. 67–129). Bogotá: Instituto de Ciencias Naturales.
Re:wild, Synchronicity Earth, IUCN SSC Amphibian Specialist Group. (2023). State of the World’s Amphibians: The Second Global Amphibian Assessment. Texas: Re:wild. Electronic source at the IUCN website
Rodríguez-Molina, C. R. (2004). Reproducción de Pleurodema brachyops (Anura: Lectodactylidae) en los llanos del Estado Apure, Venezuela. Memoria de la Fundación La Salle Ciencias Naturales, 2002, 117–125.
Rodríguez, G. M., Banda, K., Reyes, S. P., & Estupiñán-González, A. C. (2012). Lista comentada de las plantas vasculares de bosques secos prioritarios para la conservación en los departamentos de Atlántico y Bolívar (Caribe colombiano). Biota Colombiana, 13, 7–39.
Rojas, R. R., & Pérez-Peña, P. E. (2018). Evidencia preliminar del efecto borde en anfibios de la Reserva Nacional Pucacuro, al norte de la Amazonía peruana. Revista del Instituto de Investigaciones de la Amazonía Peruana, 27, 55–67.
Romero, H. G. (2013). Deforestación en Colombia: retos y perspectivas. In F. Dane (Ed.), El desafío del desarrollo sustentable en América Latina (pp. 123–142). Río de Janeiro: SOPLA.
Román-Palacios, C., Fernández-Garzón, S., Hernández, M., Ishida-Castañeda, J., Gallo-Franco, W., & Bolívar-García, A. (2016). Uso de microhábitat por anuros en un fragmento de bosque seco intervenido del Magdalena Medio, Guarinocito, Caldas. Boletín Científico Centro de Museos Museo de Historia Natural, 20, 181–196. https://doi.org/10.17151/bccm.2016.20.2.14
Salvador, J., & Gómez, D. (2018). Reptiles y anfibios del departamento del Atlántico, Colombia. Versión 3. Bogotá, Colombia: Field Museum-Museo de Historia Natural ANDES.
Stuart, S. N., Chanson, J., Cox, N.A., & Young, B. E. (2006). Estado global de los anfibios. In A. Angulo, J. V. Rueda-Almonacid, J. V. Rodríguez-Mahecha, & E. La Marca (Eds.), Técnicas de inventario y monitoreo para los anfibios de la región tropical andina (pp. 19–41). Bogotá: Panamericana Formas e Impreso S.A.
Simmons, J. E., & Muñoz-Saba, Y. (Eds.). (2005). Cuidado, manejo y conservación de las colecciones biológicas. Bogotá: Universidad Nacional de Colombia.
Thompson, P. L., Rayfield, B., & González, A. (2017). Loss of habitat and connectivity erodes species diversity, ecosystem functioning, and stability in metacommunity networks. Ecography, 40, 98–108. https://doi.org/10.1111/ecog.02558
Urbina-Cardona, J. N., Olivares-Pérez, M., & Reynoso, V. H. (2006). Herpetofauna diversity and microenvironment correlates across a pasture-edge-interior ecotone in tropical rainforest fragments in the Los Tuxtlas Biosphere Reserve of Veracruz, Mexico. Biological Conservation, 132,61–75. https://doi.org/10.1016/j.biocon.2006.03.014
Urbina-Cardona, J. N., Arturo-Navas, C., Gonzales, I., Gómez-Martínez., M. J., Llano-Mejía, J., Medina-Rangel, G. F. et al. (2014). Determinantes de la distribución de los anfibios en el bosque seco tropical de Colombia: herramientas para su conservación. In H. Pizano, & H. García (Eds.), El bosque seco tropical en Colombia (pp. 169–195). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Urbina-Cardona, J. N., Bernal, E. A., Giraldo-Echeverry, N., & Echeverry-Alcendra, A. (2015). El monitoreo de herpetofauna en los procesos de restauración ecológica: indicadores y métodos. Monitoreo a procesos de restauración ecológica, aplicado a ecosistemas terrestres. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humbold.
Vargas-Salinas, F., Angarita-Sierra, T., Ospinal, L. A., Rocha-Úsuga, A., & Rueda-Solano, L. (2019). Comunicación y ecología reproductiva. In F. Vargas-Salinas, J.A. Muñoz-Avila & M.E. Morales-Puente (Eds.), Biología de los anfibios y reptiles en bosque tropical del norte de Colombia (pp. 297–338). Tunja: Editorial UPTC.
Vargas, S. F., & Bolaños, L. M. E. (1999). Anfibios y reptiles presentes en hábitats perturbados de selva lluviosa tropical en el bajo Anchicayá, Pacífico colombiano. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 23,499–511.
Villareal, H. M., Álvarez, M., Córdoba-Córdoba, S., Escobar, F., Fagua, G., Gast, F. et al. (2004). Manual de métodos para el desarrollo de inventarios de biodiversidad. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Zug, G. R., Vitt, L., & Caldwell, J. P. (2001). Herpetology: an introductory biology of amphibians and reptiles. San Diego: Academic Press.
Eduardo Suárez-Morales a, *, Karl E. Velázquez-Ornelas b
a El Colegio de la Frontera Sur, Avenida Centenario Km 5.5, 77014 Chetumal, Quintana Roo, Mexico
b Universidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Mazatlán, Joel Montes Camarena s/n, 82040 Mazatlán, Sinaloa, Mexico
Based on deep-water (700-750 m) biological samples obtained from the southern Gulf of California, Pacific coast of Mexico, a new species of the monstrilloid copepod genus Monstrilla Dana, 1849 is described based on a single subadult female collected close to the bottom with an epibenthic sledge. Monstrilla hendrickxi sp. n. is distinguished by a unique combination of characters including: 1) no trace of eyes; 2) strong, thick antennules with segments 2-4 partly fused; 3) strongly developed apical elements on antennular segment 5 and spinous processes on segments 2-5; and 4) bilobed fifth legs with 2 apical setae on the exopodal lobe and a digitiform, unarmed endopodal lobe. The new species exhibits some affinity with surface-dwelling species of Monstrilla that have 2 setae on the exopodal lobes of the fifth legs. This is the fifth record of a species of the order Monstrilloida in the Gulf of California. The present discovery in deep oceanic waters significantly adds to our knowledge of the habitat range of this copepod order and likely anticipate further interesting findings of monstrilloids in deep waters worldwide.
Una especie nueva del género Monstrilla (Copepoda: Monstrilloida: Monstrillidae) del golfo de California, México
Resumen
Con base en muestras biológicas de aguas profundas (700-750 m) obtenidas del sur del golfo de California, costa del Pacífico de México, se describe una especie nueva del género Monstrilla Dana, 1849 con base en una hembra preadulta recolectada con un trineo epibentónico. Monstrilla hendrickxi sp. n. se distingue por una combinación única de caracteres que incluyen: 1) ausencia de estructuras oculares; 2) anténulas robustas, con los segmentos 2-4 parcialmente fusionados; 3) anténula con elementos apicales muy desarrollados en el segmento 5 y procesos espiniformes en los segmentos 2-5; y 4) quinta pata bilobulada, lóbulo exopodal armado con 2 setas apicales, lóbulo endopodal digitiforme, desarmado. La nueva especie tiene afinidad con congéneres de superficie que tienen lóbulos exopodales armados con 2 setas. Este es el quinto registro de monstriloides en el golfo de California. El presente hallazgo aumenta significativamente nuestro conocimiento sobre el intervalo de hábitats de este orden de copépodos y probablemente anticipa más descubrimientos interesantes de monstriloides en aguas profundas de todo el mundo.
Members of the copepod order Monstrilloida Sars, 1901 are, as larvae, endoparasites of benthic marine invertebrates, but their infective early nauplii and their non-feeding reproductive adult and preadult stages are planktonic. The known hosts of the postnaupliar and juvenile stages include benthic polychaetes (species of the families Syllidae, Capitellidae, Serpulidae, and Spionidae), mollusks, and sponges (Huys et al., 2007; Jeon et al., 2018; Suárez-Morales, 2011, 2018; Suárez-Morales et al., 2010, 2014). In plankton, monstrilloids have been reported chiefly from a wide range of shallow coastal habitats including estuaries (Suárez-Morales et al., 2020), coastal embayments (Suárez-Morales, 1994a, b; Suárez-Morales & Gasca, 1990), and coral reefs (Sale et al., 1996), where they can be found in aggregations (Suárez-Morales, 2001). Monstrilloids have been recently reported from rocky shore tidepools as well (Cruz-Lopes de Rosa et al., 2021). Deep oceanic waters, though, would seem to be an unlikely source of monstrilloid specimens for taxonomic study because of the copepods’ limited dispersal capacity in the water column and their need to remain close to their benthic hosts (Suárez-Morales, 2001, 2011, 2018).
The order is currently represented by a single family Monstrillidae Dana, 1849 containing at least 7 valid genera: Monstrilla Dana, 1849; Cymbasoma Thompson, 1888; Monstrillopsis Sars, 1921; Maemonstrilla Grygier & Ohtsuka, 2008; Australomonstrillopsis Suárez-Morales & McKinnon, 2014; Caromiobenella Jeon, Lee & Soh, 2018, and Spinomonstrilla Suárez-Morales, 2019 (Jeon et al., 2016, 2018; Suárez-Morales et al., 2020). Currently, the genus Cymbasoma, with 78 nominal species, is the most diverse within the order (Razouls et al., 2023: Suárez-Morales & McKinnon, 2016; Walter & Boxshall, 2022). The taxonomic examination of a monstrilloid subadult female collected with a deep-water epibenthic sledge operated close to the bottom within a depth range of 700-750 m in oceanic waters of the southern Gulf of California, Mexico, allowed us to recognize this individual as representative of an undescribed species of Monstrilla, which is herein described following upgraded morphological standards and compared with congeneric species (Grygier & Ohtsuka, 1995). This is the third published record of monstrilloid copepods from deep oceanic waters worldwide (see Suárez-Morales & Mercado-Salas, 2023) and the fifth record of monstrilloids in the Gulf of California.
Materials and methods
A subadult female of a monstrilloid copepod was collected with an epibenthic sledge operated in mid-water, close to the bottom off the Pacific coast of northwestern Mexico, in the Gulf of California. The maximum sampling depth was 750 m. Immediately after collection, the organisms from the haul were preserved in 4% formalin with seawater. Sorting of specimens and preliminary observations were made under an Olympus SZ51 stereomicroscope. The single female monstrilloid individual thus found was recognized as a subadult female and tentatively identified as a member of the genus Monstrilla. Further examination was performed under an Olympus BX51 compound microscope with Nomarski DIC optics. Prior to examination, the specimen was partially dissected and swimming legs 1-4 and the cephalothorax, urosome +antennules were mounted in glycerol on glass slides, which were sealed with acrylic nail varnish. The slides have been deposited in the collection of Zooplankton (ECO-CHZ) held at El Colegio de la Frontera Sur (ECOSUR), Unidad Chetumal, in Chetumal, Quintana Roo, Mexico (ECO-CH-Z). Detailed examination allowed us to determine that this specimen represented an undescribed species, which we describe herein following current descriptive standards in monstrilloid taxonomy. The general morphological terminology follows Huys and Boxshall (1991), while the nomenclature of the antennular armature follows Grygier and Ohtsuka (1995).
Diagnosis. Large (2.9 mm) female subadult Monstrilla with robust, cylindrical cephalothorax representing about 60% of total body length and no trace of eyes. Antennules thick, about 1/3 as long as cephalothorax; antennulary segments 2-4 partly fused, segment 3 with modified setae, segments 2, 4, and 5 each furnished with a conical or spinous process. Segments 4-5 partly fused, latter segment with strongly developed apical spiniform elements. Ventral side of genital double-somite carrying short, corrugate ovigerous spines not reaching beyond distal end of caudal rami. Fifth leg bilobed, exopodal lobe bearing 2 subequally long terminal setae and endopodal lobe digitiform, unarmed. Caudal rami each with 6 setae, innermost of which (VI) shortest and outer proximal one (I) longest.
Description of holotype subadult female. Body robust; shape and tagmosis as usual in female Monstrilla (Suárez-Morales 1994a, 2019). Total body length 2.92 mm, measured from anterior end of cephalothorax to posterior margin of anal somite. Cephalothorax length 1.74 mm, representing about 60% of total body length, containing thick egg mass (Fig. 1A). Oral cone located 0.53 of way back along ventral surface of cephalothorax. Cephalic region with weakly produced forehead (Fig. 2A). All 3 cups of naupliar eye with pigment absent (Figs. 1A, B, 2A, 3D). Preoral area with ventral ornamentation of small, nipple-like cuticular processes (nlp) with adjacent fields of integumental wrinkles and anterior cluster of pores (apc) (Figs. 3D, 4B). Antennules slender, relatively short (588 µm) and thick, corresponding to 33% of cephalothorax length and almost 20 % of total body length (Fig. 1A). Antennules indistinctly 5-segmented (1-5 in Fig. 1B), segment 1 separate but segments 2-5 partly fused (Figs. 1B, 3A), with intersegmental division 2-3 marked by weak constriction. In terms of current nomenclature for antennular armature of female monstrilloid copepods (Grygier & Ohtsuka, 1995), element 1 present on first segment (Fig. 3A). Second segment armed as usual with elements 2v1-3, 2d1,2, and long element IId (Figs. 3A, B, 4C), but additionally with conical process on distal inner margin (arrowed in Fig. 3B). Putative third segment with setiform element 3, but pair of short, spiniform elements replacing usual long, setiform elements IIIv and IIId (Fig. 3A). Putative fourth segment armed with elements 4v1-3, long 4aes, and elements IVd and IVv (Fig. 3A), as well as conical spiniform process on inner proximal margin (arrowed in Fig. 3A). Fifth segment carrying setal elements Vv, Vd, Vm, 61, 2, aesthetasc 6aes, plus only 2 unbranched setae of “b” group (b6 and b3) on outer margin, as well as distal spiniform process at insertion of elements 61, 2, and 6aes (Figs. 2B, 3C). First pedigerous thoracic somite incorporated into cephalothorax, succeeding 3 free pedigerous somites each bearing pair of biramous swimming legs, all 3 together accounting for 31.3% of total body length (Fig. 1A). Endopodites and exopodites of swimming legs 1-4 unequal (exopods longer), triarticulate, and with same setal armature in each leg, except exopod of leg 1 with one fewer seta on the distal segment (Fig. 4D).
Armature formula of swimming legs: leg 1: basis 1-0, endopodite 0-1; 0-1; 1-2-2, exopodite I-1; 0-1; I-2-2; legs 2-4: basis 1-0, endopodite 0-1; 0-1; 1-2-2, exopodite I-1; 0-1; I-2-3. Coxae of legs 1-4 unarmed; each pair medially joined by subrectangular intercoxal sclerite about 1.3 times as long as broad with curved distal margin; anterior surface of sclerites 2-4 ornamented with rows of minute hyaline spinules. Basis separated from coxa posteriorly by diagonal articulation, lacking usual basipodal outer seta in legs 2-4. Outer distal corner of first and third exopodal segments of swimming legs 1-4 each with short, slender spiniform element about 1/3 as long as its segment. All natatory setae lightly and biserially plumose except for spiniform seta on outer distal corner of third exopodal segments, this being lightly setulate along inner side and bearing continuous row of small denticles along outer margin (Fig. 4D). Fifth legs medially conjoined, arising ventrally from posterior margin of fifth pedigerous somite (Fig. 4A), each being represented by elongate bilobed structure with its outer (exopodal) lobe armed with 2 subequally long apical setae. Unarmed and smooth inner (endopodal) lobe arising from proximal inner margin of outer lobe, almost reaching distal tip of latter. Urosome short (length = 389 µm long), accounting for 13.1% of total body length and consisting of fifth pedigerous somite, genital double-somite, 2 free abdominal somites, and caudal rami (Figs. 1C, 4A). Genital double somite representing 28.8% of length of urosome. Preanal somite about half as long as anal somite (Fig. 4A). Medial ventral surface of genital double-somite moderately swollen, bearing basally conjoined and posteriorly directed ovigerous spines (Fig. 4A). These spines (os in Figs. 1D, 4A) relatively short, corresponding to 21% of total body length and reaching to mid-length of caudal setae, each narrowing in its distal half to thin, seemingly socketed, seta-like section (Figs. 1D, 4A). Caudal rami subrectangular, 1.25 times as long as wide, moderately divergent, bearing 3 strong and subequally long terminal setae, as usual in genus (Figs. 1D, 4A), among 6 setal elements in all (I-VI), with innermost seta (VI) being shortest and proximal outer seta (I) being longest (Fig. 4A).
Figure 1. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female, digital photos. A, Habitus, dorsal view; B, anterior half of cephalothorax with 5- segmented antennules (1-5), ventral view; C, urosome, ventral view, showing ovigerous spines (os) arising from genital double-somite, fifth legs (P5) with 2 distal setae (1, 2), and caudal setae I-VI; D, distal part of ovigerous spines and caudal rami showing setae I-VI, ventral view.
Figure 2. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female, digital photos. A, Anterior part of cephalothorax showing weakly produced forehead and lack of eyes; B, antennule showing dorsal segmentation; C, fifth leg (P5) showing exopodal (exp) and endopodal (enp) lobes, ventral view; D, apical elements (61, 62, and 6aes) of distal antennular segment.
Figure 3. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female. A, Antennule showing setation labelled in accordance with Grygier and Ohtsuka’s (1995) nomenclature, ventral view; B, similarly labelled setation of second antennular segment, ventral view, also showing segment’s inner distal conical process (arrow); C, distal (fifth) segment of antennule, ventral view, showing similarly labelled setation and distal process (arrow); D, anterior third of cephalothorax, ventral view, showing integumental ornamentation, including nipple-like processes (nlp), anterior pore cluster (apc), and preoral pores (pp). Scales A-D = 100 µm.
Figure 4. Monstrilla hendrickxi sp. n., from the Gulf of California, holotype female. A, Urosome, ventral view, showing bilobed fifth leg with exopodal (exp) and endopodal (enp) lobes, ovigerous spines (os), and caudal rami setation (I-VI); B, anterior half of cephalothorax, ventral view, showing unlayed internal egg mass (em), oral cone (oc), and nipple-like processes (nlp); C, antennule segments 1-3 (S1-S3), ventral view, showing setation labelled in accordance with Grygier and Ohtsuka’s (1995) nomenclature, including modified setae IIIv and IIId; D, right leg 1, semi-lateral view, showing setation including basipodal seta (bs) and exopodal (exp) and endopodal (enp) rami. Scales A-D = 100 µm.
Taxonomic summary
Type locality. Southern Gulf of California, Mexico (25°53’15” N, 110°10’08” W). Sampling depth between 710 and 750 m.
Material examined. One subadult female (holotype), partially dissected and mounted on 2 semi-permanent glycerin slides sealed with acrylic nail varnish (ECO-CH-Z 11860). Epibenthic sledge, TALUD XVI B cruise, southern Gulf of California, Mexico, 31 May 2014.
Etymology. The specific name, a masculine genitive eponym, honors Dr. Michel E. Hendrickx (ICMyL-UNAM) for his sustained efforts and achievements in exploring the crustacean fauna of the Gulf of California and the Mexican Pacific.
Male. Unknown.
Host. Unknown.
Remarks
The sampling gear used to collect this specimen was an epibenthic sledge, not a plankton net; the latter is more efficient for capturing planktonic adult monstrilloids in surface waters. Epi-mesopelagic monstrilloids collected by sledge type gears have been reported from depths of 118 and 302 m in the North Atlantic (Suárez-Morales & Mercado-Salas, 2023), but those individuals were badly damaged even though the sledge had an attached plankton collector (Brandt et al., 2014). The specimen recovered from the TALUD XVI-B sledge sample was in reasonably good condition for taxonomic study.
The present subadult female monstrilloid from the Gulf of California can be readily assigned to the genus Monstrilla by its possession of the diagnostic generic features for females, including the presence of 2 somites between the genital double-somite and the anal somite, 6 caudal setae, and the oral cone’s location ventrally at nearly mid-length of the cephalothorax (Isaac, 1975; Suárez-Morales, 1994a). Among the species-level diagnostic features of M. hendrickxi sp. n., the most useful for comparison among congeneric species are the antennular structure and armature, the shape of leg 5, and the absence of eyes. These will be considered in sequence below.
The specifically distinctive characters of M. hendrickxi sp. n. include: 1) eyes and eye-related structures absent; 2) antennules relatively short, robust, representing nearly 33% of cephalothorax length, with segments 2-5 partly fused; 3) segments 2-5 furnished with modified setae or strong spiniform processes, 4) fifth antennulary segment with remarkably long, thick apical elements; 5) fifth leg bilobed, with digitiform endopodal lobe unarmed, exopodal lobe with 2 terminal setae; 6) 6 caudal setae, innermost seta (VI) being shortest, proximal outer seta (I) longest. The finding of subadult monstrillids in the plankton is not unusual and some species have been described from these individuals, including the first described monstrilloid copepod, Thaumaleus typica (Krøyer, 1842) (Grygier, 1994), Monstrilla capitellicola Hartman (1961), M. elongata Suárez-Morales, 1994a, and Monstrilla sp. from Hawaii (Suárez-Morales et al., 2014).
Partial or complete fusion of antennular segments 2-5 is found in several other species of Monstrilla, including M. ilhoii Lee & Chang, 2016, M. mariaeugeniae Suárez-Morales & Islas-Landeros, 1993, M. satchmoi Suárez-Morales & Dias, 2001, M. grandis Giesbrecht, 1891, M. gracilicauda Giesbrecht, 1893, and M. elongata Suárez-Morales, 1994a. None of these species displays the remarkable development of apical elements 61, 62, and 6aes observed in M. hendrickxi sp. n. The only available illustration of an antennule of M. nichollsi Davis, 1949 (= C. helgolandica) (Suárez-Morales pers. obs.) (cf. Nicholls, 1944: fig. 26, as Monstrilla sp.) shows a very long apical element on its fifth segment, which is probably identifiable as the aesthetasc 6aes. Elsewhere on the antennule, no congeneric species has modified setal elements IIId and IIIv on segment 3 or large, spiniform or conical processes on segments 2, 4, and 5 like those described in the new species.
Only a few known species originally described as Monstrilla possess a bilobed fifth leg with 2 setae on the outer (exopodal) lobe, 2 of them have been transferred to the genus Caromiobenella: C. helgolandica (Claus, 1863) and C. hamatapex (Grygier & Ohtsuka, 1995); the other species of Monstrilla sharing this character are M. mariaeugeniae, M. capitellicola, and M. leucopis Sars, 1921. Also, both Monstrilla sp. from Hawaii and M. capitellicola from Southern California (Hartman, 1961), likely represented by subadult individuals, also show only 2 setae on the outer lobe, a character conserved through the copepodiid stages CIII-V (Suárez-Morales et al., 2014). The new species M. hendrickxi differs from C. helgolandica, C. hamatapex, M. capitellicola, and M. leucopis, by its possession of a long, digitiform endopodal lobe, which is absent in these 4 species (Chang, 2014; Grygier & Ohtsuka, 1995; Sars, 1921; Zavarzin & Suárez-Morales, 2024). The corresponding endopodal lobe is clearly shorter in M. capitellicola (Hartman, 1961) than in the new species. Monstrilla wandelii Stephensen, 1913, and M. mariaeugeniae both exhibit a small, unarmed subtriangular endopodal lobe (Nicholls, 1944, fig 26; Park, 1967; Suárez-Morales & Islas-Landeros, 1993), which clearly differs from the elongate, digitiform endopodal lobe observed in M. hendrickxi sp. n.. Monstrilla nichollsi, a synonym of C. helgolandica (Suárez-Morales pers. obs.), was named by Davis (1949) based solely on Nicholl’s (1944, fig. 26) illustration of the fifth leg, thus allowing us to add C. helgolandica to the group of monstrillid species with 2 exopodal setae on the fifth leg exopodal lobe. It should be noted that the armature of the fifth leg exopodal lobe is conservative during the immature stages including the preadult CV (Suárez-Morales et al., 2014); changes in this character at the final molt are unlikely.
Monstrilla hendrickxi sp. n. is the only monstrilloid copepod in which no trace of the naupliar eye is present, although weakly developed visual structures have been observed previously in deep-living species (Suárez-Morales & Mercado-Salas, 2023). This contrasts with the usually well-developed, highly pigmented, three-cup naupliar eyes of most known monstrilloids. Functional eyes are probably extremely important for planktonic adult monstrilloids, allowing them, for example, to migrate to different light conditions in the water column and favor their dispersal (Suárez-Morales, 2018; Suárez-Morales & Gasca, 1990). The weak eye development of deep-living monstrilloids is likely an adaptive consequence of their aphotic habitat.
Monstrilla hendrickxi sp. n. is the fifth species of the copepod order Monstrilloida recorded from the Gulf of California, after Monstrilla gibbosa Suárez-Morales & Palomares-García, 1995, Spinomonstrilla spinosa (Park, 1967) (originally reported as Monstrilla spinosa), Cymbasoma californiense Suárez-Morales & Palomares-García, 1999, and recently M. leucopis Sars, 1921 (Suárez-Morales, 2019; Suárez-Morales & Palomares-García, 1999; Suárez-Morales & Velázquez-Ornelas, 2023).
Acknowledgements
We thank Michel E. Hendrickx (ICMyL-UNAM) for kindly allowing us to examine this specimen. Ship time for the TALUD XVI-B cruise was provided by the Coordinación de la Investigación Científica, UNAM, and partly supported by Conacyt (project # 179467). We also thank to an anonymous reviewer for the corrections made to improve this article.
References
Brandt, A., Havermans, C., Janussen, D., Jörger, K. M., Meyer-Löbbecke, A., Schnurr, S. et al. (2014). Composition and abundance of epibenthic-sledge catches in the South Polar Front of the Atlantic. Deep-Sea ResearchII, 108, 69–75. Doi?
Claus, C.(1863). Die frei lebenden Copepoden mit besonderer Berücksichtigung der Fauna Deutschlands, der Nordsee und des Mittelmeeres. Leipzig: Verlag von Wilhelm Engelmann.
Chang, C. Y. (2014). Two new records of monstrilloid copepods (Crustacea) from Korea. Animal Systematics, Evolution and Diversity, 30, 206–214. https://doi.org/10.5635/ASED. 2014.30.3.206
Cruz-Lopes da Rosa, J., Dias, C. O., Suárez-Morales, E., Weber, L. I., & Gomes-Fischer, L. (2021). Record of Caromiobenella (Copepoda, Monstrilloida) in Brazil and discovery of the male of C. brasiliensis: morphological and molecular evidence. Diversity, 13, 241. https://doi.org/ 10.3390/d13060241
Dana, J. D. (1849). Conspectus crustaceorum, quae in orbis terrarum circumnavigatione, Carolo Wilkes, e classe Reipublicae foederatae duce, lexit et descripsit Jacobus D. Dana. Pars II. Proceedings of the American Academy of Arts and Sciences, 2, 9–61.
Davis, C. C. (1949). A preliminary revision of the Monstrilloida, with descriptions of two new species. Transactions of the American Microscopical Society, 68, 245–255. https://doi.org/10.2307/3223221
Giesbrecht, W. (1891). Elenco dei Copepodi pescati dalla R. Corvetta ‘Vettor Pisani’ secondo la loro distribuzione geografica. Atti della Accademia Nazionale dei Lincei, Classe di Scienze Fisiche Matematiche e Naturali Rendiconti, 4, 276–282.
Giesbrecht, W. (1893). Systematik und Faunistik der pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte, 19, 1–831.
Grygier, M. J. (1994) [dated 1993]. Identity of Thaumatoessa (= Thaumaleus) typica Krøyer, the first described monstrilloid copepod. Sarsia, 78, 235–242. https://doi.org/10.1080/00364827.1993.10413537
Grygier, M. J., & Ohtsuka, S. (1995). SEM observation of the nauplius of Monstrilla hamatapex, new species, from Japan and an example of upgraded descriptive standards for monstrilloid copepods. Journal of Crustacean Biology, 15, 703–719. https://doi.org/10.1163/193724095X00118
Grygier, M. J., & Ohtsuka, S. (2008). A new genus of monstrilloid copepods (Crustacea) with anteriorly pointing ovigerous spines and related adaptations for subthoracic brooding. Zoological Journal of the Linnean Society, 152, 459–506. https://doi.org/10.1111/j.1096-3642.2007.00381.x
Hartman, O. (1961). A new monstrillid copepod parasitic in capitellid polychaetes in southern California. Zoologischer Anzeiger, 167, 325–334.
Huys, R., & Boxshall, G. A. (1991). Copepod evolution. London: Ray Society.
Huys, R., Llewellyn-Hughes, J., Conroy-Dalton, S., Olson, P. D., Spinks, J. N., & Johnston, D. A. (2007). Extraordinary host switching in siphonostomatoid copepods and the demise of the Monstrilloida: Integrating molecular data, ontogeny and antennulary morphology. Molecular Phylogenetics and Evolution, 43, 368–378. https://doi.org/10.1016/j.ympev. 2007.02.004
Isaac, M. J. (1975). Copepoda, sub-order: Monstrilloida. Fiches d’Identification du Zooplancton, 144/145, 1–10.
Jeon, D., Lee, W., & Soh, H. Y. (2018). New genus and two new species of monstrilloid copepods (Copepoda: Monstrillidae): integrating morphological, molecular phylogenetic, and ecological evidence. Journal of Crustacean Biology, 38, 45–65. https://doi.org/10.1093 /jcbiol/rux095
Jeon, D., Lim, D., Lee, W., & Soh, H. Y. (2016). First use of molecular evidence to match sexes in the Monstrilloida (Crustacea: Copepoda), and taxonomic implications of the newly recognized and described partly Maemonstrilla-like females of Monstrillopsis longilobata Lee, Kim & Chang, 2016. Peer J, 13,e4938. https://doi.org/10.7717/peerj.4938
Krøyer, H. (1842). Crustacés. In P. E. Gaimard (Ed.), Atlas de Zoologie. Voyages de la Commission Scientifique du Nord en Scandinavie, en Laponie au Spitzberg et aux Feröe pendant les Anneés 1838, 1839 et 1840 sur la Corvette La Recherche, Commandée par M. Fabvre (pl. 41–43). Arthus Bertrand, Paris.
Lee, J., & Chang, C. Y. (2016). A new species of Monstrilla Dana, 1849 (Copepoda: Monstrilloida: Monstrillidae) from Korea, including a key to species from the North-west Pacific. Zootaxa, 4174, 396–409. https://doi.org/10.11646/zootaxa.4174.1.24
Nicholls, A. G. (1944). Littoral Copepoda from South Australia (II) Calanoida, Cyclopoida, Notodelphyoida, Monstrilloida, and Caligoida. Records of the South Australian Museum, 8, 1–62.
Park, T. S. (1967). Two unreported species and one new species of Monstrilla (Copepoda: Monstrilloida) from the Strait of Georgia. Transactions of the American Microscopical Society, 86, 144–152.
Razouls, C., Desreumaux, N., Kouwenberg, J., & de Bovée, F. (2023). Biodiversity of marine planktonic copepods (morphology, geographic distribution, and biological data [2005-2023]). Sorbonne University, CNRS. http://copepodes.obs-banyuls.fr/en [Accessed April 28, 2022]
Sale, P. F, McWilliams, P. S., & Anderson, D. T. (1996). Composition of the near-reef zooplankton at Heron Reef, Great Barrier Reef. Marine Biology, 34, 59–66. https://doi.org/10.1007/BF00390788
Sars, G. O. (1901). An account of the Crustacea of Norway with short descriptions and figures of all the species Vol. IV. Copepoda Calanoida. Bergen: The Bergen Museum.
Sars, G. O. (1921). An account of the Crustacea of Norway with short descriptions and figures of all the species, Vol.VIII. Copepoda Monstrilloida & Notodelphyoida. Bergen: The Bergen Museum.
Stephensen, K. (1913) Account of the Crustacea and the Pycnogonida collected by Dr. V. Nordmann in the summer of 1911 from northern Stromfjord and Giesecke Lake in West Greenland. Meddeleser om Groenland, 51, 55–77.
Suárez-Morales, E. (1994a). Monstrilla elongata, a new monstrilloid copepod (Crustacea: Copepoda: Monstrilloida) from a reef lagoon of the Caribbean coast of Mexico. Proceedings of the Biological Society of Washington, 107, 262–267.
Suárez-Morales, E. (1994b). Thaumaleus quintanarooensis, a new monstrilloid copepod from the Mexican coasts of the Caribbean Sea. Bulletin of Marine Science, 54, 381–384.
Suárez-Morales, E. (2001). An aggregation of monstrilloid copepods in a western Caribbean reef area: ecological and conceptual implications. Crustaceana, 74, 689–696. https://doi.org/10.1163/156854001750377966
Suárez-Morales, E. (2019). A new genus of the Monstrilloida (Copepoda) with large rostral process and metasomal spines, and redescription of Monstrilla spinosa Park, 1967. Crustaceana, 92, 1099–1112. https://doi.org/10.1163/ 15685403-00003925
Suárez-Morales, E., & Dias, C. O. (2001). Taxonomic report of some monstrilloids (Copepoda: Monstrilloida) from Brazil with description of four new species. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Biologie, 71, 65–81.
Suárez-Morales, E., & Gasca, R. (1990). Variación dial del zooplancton asociado a las praderas de Thalassia testudinum en una laguna arrecifal del Caribe Mexicano. Universidad y Ciencia, 7, 57–64.
Suárez-Morales, E., Harris, L. H., Ferrari, F. D., & Gasca, R. (2014). Late postnaupliar development of Monstrilla sp. (Copepoda: Monstrilloida), a protelean endoparasite of benthic polychaetes. Invertebrate Reproduction & Development, 58, 60–73.
Suárez-Morales, E., & Islas-Landeros, M. E. (1993). A new species of Monstrilla (Copepoda: Monstrilloida) from a reef lagoon off the Mexican coast of the Caribbean Sea. Hydrobiologia, 271, 45–48.
Suárez-Morales, E., & McKinnon, A. D. (2014). The Australian Monstrilloida (Crustacea: Copepoda) I. Monstrillopsis Sars, Maemonstrilla Grygier & Ohtsuka, and Australomonstrillopsis gen. nov. Zootaxa, 3779, 301–340. https://doi.org/10.11646/zootaxa.3779.3.1
Suárez-Morales, E., & McKinnon, A. D., (2016). The Australian Monstrilloida (Crustacea: Copepoda) II. Cymbasoma Thompson, 1888. Zootaxa Monographs, 4102, 1–129. https://doi.org/10.11646/zootaxa.4102.1.1
Suárez-Morales, E., & Mercado-Salas, N. F. (2023). Two new species of Cymbasoma (Copepoda: Monstrilloida: Monstrillidae) from the North Atlantic. Journal of Natural History, 57, 1312–1330. https://doi.org/10.5852/ejt. 2024.917.2395
Suárez-Morales, E., Paiva Scardua, M., & Da Silva, P. M. (2010). Occurrence and histopathological effects of Monstrilla sp. (Copepoda: Monstrilloida) and other parasites in the brown mussel Perna perna from Brazil. Journal of the Marine Biological Association of the United Kingdom, 90, 953–958. https://doi.org/10.1017/S0025315409991391
Suárez-Morales, E., & Palomares-García, R. (1995). A new species of Monstrilla (Copepoda: Monstrilloida) from a coastal system of the Baja California Peninsula, Mexico. Journal of Plankton Research, 17, 745–752. https://doi.org/ 10.1093/plankt/17.4.745
Suárez-Morales, E., & Palomares-García, R. (1999). Cymbasoma californiense, a new monstrilloid (Crustacea: Copepoda: Monstrilloida) from Baja California, Mexico. Proceedings of the Biological Society of Washington, 112, 189–198.
Suárez-Morales, E., Vásquez-Yeomans, L., & Santoya, L. (2020). A new species of the Cymbasoma longispinosum species-group(Copepoda, Monstrilloida, Monstrillidae) from Belize, western Caribbean Sea. Revista Mexicana de Biodiversidad, 91, e913176. https://doi.org/10.22201/ib. 20078706e.2020.91.3176
Suárez-Morales, E., & Velázquez-Ornelas, K. E. (2023). First record of Monstrilla leucopis G.O. Sars, 1921 (Copepoda: Monstrilloida: Monstrillidae) from the Eastern Pacific. Crustaceana, 96, 1183–1190. https://doi.org/10.1163/156854 03-bja10334
Thompson, I. C. (1888). Copepoda of Madeira and the Canary Islands, with descriptions of new genera and species. Zoological Journal of the Linnean Society, 20, 145–156. https://doi.org/10.1111/j.1096-3642.1888.tb01443.x
Zavarzin, D., & Suárez-Morales, E. (2024). First record of Caromiobenella helgolandica (Claus, 1863) (Copepoda: Monstrilloida: Monstrillidae) from the Okhotsk Sea. Crustaceana, 97, 151–157. https://doi.org/10.1163/15685403- bja10345