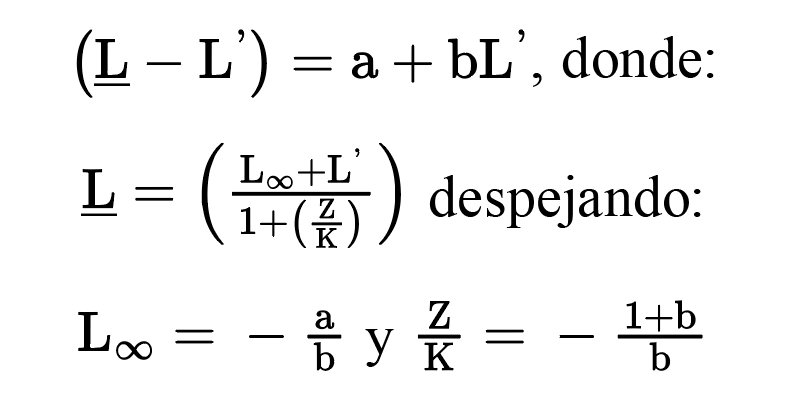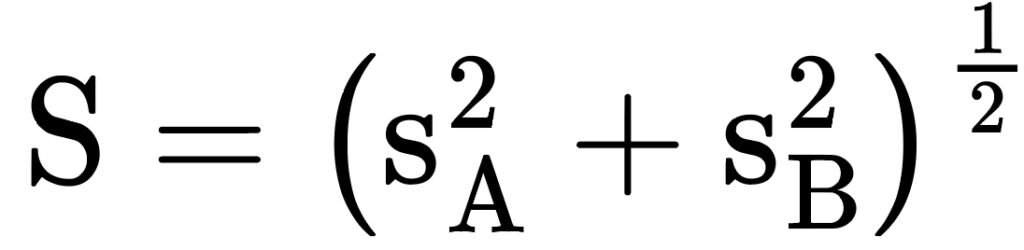Influencia de la variación climática en el crecimiento radial de Pinus montezumae en el Parque Nacional Iztaccíhuatl – Popocatépetl
Influence of climatic variation on the radial growth of Pinus montezumae in Iztaccíhuatl – Popocatépetl National Park
Miguel Ángel Ortega-Cervantes, Liliana Cuapio-Hernández, Alejandro Ismael Monterroso-Rivas y Alejandro Corona-Ambriz *
Universidad Autónoma Chapingo, División de Ciencias Forestales, Km. 38.5 Carretera México-Texcoco, 56230 Chapingo, Texcoco, Estado de México, México
*Autor para correspondencia: acoronaa@chapingo.mx (A. Corona-Ambriz)
Recibido: 27 febrero 2025; aceptado: 1 agosto 2025
Resumen
La variabilidad climática, intensificada por el cambio climático, influye en los patrones de crecimiento de los árboles, afecta su desarrollo y respuesta a condiciones ambientales extremas, el objetivo fue identificar patrones asociados al cambio climático en la región. Se recolectaron y fecharon núcleos de crecimiento. A partir de ésto, se generaron las cronologías de crecimiento radial. Asimismo, se emplearon datos climáticos de reanálisis para identificar los periodos de respuesta clima-crecimiento y calcular las correlaciones entre las cronologías y las variables climáticas. Los resultados mostraron un potencial dendrocronológico de intermedio a alto de Pinus montezumae e indican que la especie responde a la precipitación de primavera y verano, ya que favorecen el incremento anual del crecimiento radial. La temperatura máxima de otoño contribuye al desarrollo de madera temprana al final del ciclo de crecimiento, mientras que la temperatura media y mínima de invierno propicia la generación de madera tardía. Además, se observó que la especie es sensible a eventos de sequía al disminuir el crecimiento radial como se observó en la última década. Estos hallazgos resaltan la importancia de P. montezumae como un indicador del impacto de la variabilidad climática en los bosques de la región.
Palabras clave: Anillos de crecimiento; Anillos de árbol; Bosque de coníferas; Dendrocronología; Respuesta climática
Abstract
Climate variability, intensified by climate change, influences tree growth patterns, affecting their development and response to extreme environmental conditions. This study aimed to identify patterns associated with climate change in the region. From this, radial growth chronologies were generated. Cross-dating was also used to verify the dates and generate chronologies. Reanalysis climate-radial growth response periods and calculate correlations between the chronologies and climate variables. The results showed a moderate to high dendrochronological potential for Pinus montezumae and indicated that the species responds to spring and summer precipitation, which favors annual radial growth. Maximum autumn temperatures contribute to the development of earlywood at the end of the growth cycle, while mean and minimum winter temperatures promote latewood formation. Additionally, the species was sensitive to drought events, as evidenced by reduced radial growth over the past decade. These findings highlight the importance of P. montezumae as an indicator of the impact of climate variability on regional forests.
Keywords: Growth rings; Tree ring; Conifer Forest; Dendrochronology; Climate response
Introducción
El cambio climático es el resultado de las alteraciones de la variabilidad climática natural, atribuida principalmente a actividades humanas, y tiene importantes repercusiones en los ecosistemas terrestres (Cassman y Wood, 2005). De acuerdo con el Panel Intergubernamental sobre Cambio Climático (IPCC), desde aproximadamente 1950 se han registrado cambios significativos en diversos eventos climáticos extremos (IPCC, 2014). Entre estos cambios se incluyen la reducción de temperaturas frías extremas, el aumento de temperaturas cálidas extremas, el incremento de los niveles extremos del mar y el aumento en la frecuencia de precipitaciones intensas en varias regiones, fenómenos que han sido vinculados con la influencia humana.
El cambio climático afecta a los árboles de múltiples maneras, impacta en su crecimiento, reproducción y supervivencia. El aumento de la temperatura y la alteración de los patrones generan estrés hídrico, reduciendo la disponibilidad de agua y afectando la capacidad fotosintética de los árboles. Además, la mayor frecuencia de fenómenos extremos, como sequías e incendios forestales, incrementa la mortalidad de los bosques. Estos cambios también influyen en los ciclos fenológicos, como el inicio del crecimiento anual, y disminuyen la resiliencia de los árboles frente a plagas y enfermedades (Aber et al., 2001; IPCC, 2021).
Ante estos desafíos, surge la necesidad de comprender la influencia de la variabilidad climática en los ecosistemas. La dendrocronología, disciplina que estudia los anillos de crecimiento de los árboles, permite reconstruir eventos climáticos pasados y analizar sus efectos en los bosques (Benito, 2014; Stokes y Smiley, 1996). En particular, una estrategia clave en el estudio del crecimiento estacional de las coníferas es el análisis de los cambios interanuales en el crecimiento radial, centrado en la producción de madera temprana y madera tardía (Meko y Baisan, 2001). La formación de madera temprana está asociada con las condiciones climáticas de invierno y primavera, reflejando el período de mayor tasa de crecimiento (Vaganov et al., 2006), mientras que la producción de madera tardía está influenciada por la disponibilidad de humedad durante el verano y el otoño en regiones con estaciones secas (Howard et al., 2021). La combinación de ambas fases determina el ancho total del anillo de crecimiento.
El crecimiento radial y la formación de anillos han sido ampliamente estudiados en el género Pinus mediante técnicas dendrocronológicas para comprender la respuesta de los bosques a las variaciones climáticas. A nivel mundial, los pinos son reconocidos por formar anillos anuales bien definidos, lo que los convierte en modelos ideales para la reconstrucción climática y el análisis de eventos históricos, como sequías prolongadas y fluctuaciones de temperatura (Fritts, 1976; Osorio-Osorio et al., 2020; Schweingruber, 1988). En este contexto, Pinus montezumae Lamb. es una especie de gran importancia ecológica, ya que forma parte de los bosques templados que actúan como reservorios de biodiversidad y reguladores del ciclo hidrológico (Farjon et al., 1997). Sin embargo, a pesar de su relevancia, su estudio en la región central de México, particularmente en el Estado de México, ha sido limitado. Esta falta de información destaca la necesidad de profundizar en su análisis, lo que permitirá una mejor comprensión de sus respuestas.
El objetivo principal de esta investigación fue analizar la relación entre la variabilidad de precipitación y temperatura, incluyendo eventos climáticos extremos, y el crecimiento de P. montezumae en la Estación Forestal Experimental Zoquiapan mediante métodos dendrocronológicos, con el fin de evidenciar patrones asociados al cambio climático en la región. Este análisis no solo contribuye a un mejor entendimiento de la biología de P. montezumae, sino que también aporta información valiosa sobre la respuesta de esta especie a cambios climáticos en su entorno.
Materiales y métodos
El sitio de estudio se localiza en la Estación Forestal Experimental Zoquiapan (EFEZ), ubicada en la porción norte del Parque Nacional Iztaccíhuatl Popocatépetl, en los límites de Puebla y el Estado de México (19°12’30” – 19°20’00” N, 98°30’00” – 98°42’30” O) (fig. 1). Esta área forma parte del sistema orográfico de la sierra Nevada, en la región centro-oriental de la Faja Volcánica Transmexicana; la altitud varía entre 3,080 a 3,690 m. El clima predominante en la región es templado subhúmedo C (w’’2) (w) (b’) ig con lluvias en verano, la temperatura promedio anual oscila entre 12 y 18 °C, en los meses cálidos puede superar los 20 °C y en invierno desciende a menos de 5 °C (Conanp, 2013). La vegetación está constituida por comunidades de pinos (Pinus hartwegii Lindl., P. montezumae Lamb., P. leiophylla Schltdl. et Cham., P. pseudostrobus Lindl., P. rudis Endl., P. teocote Cham et Schltdl.), Abies (Abies religiosa (Kunth) Schltdl. et Cham.), Alnus (Alnus firmifolia Fernald, A. jorullensis Kunth) y encinos (Quercus laurina Bonpl., Q. rugosa Née., Q. peduncularis Née., Q. crassipes Bonpl.). El sotobosque está compuesto, principalmente, por especies de pasto tipo zacatal de los géneros Festuca L., Muhlenbergia Schreb. y Agrostis L. (Rzedowski, 2006).
La colecta de muestras se realizó en los sitios Chicamula y El Charco, seleccionados por la dominancia de Pinus montezumae. Se eligieron 26 pinos longevos, con fuste recto y sin evidencia visibles de disturbio. A una altura de 1.30 m se extrajeron núcleos de crecimiento utilizando un taladro pressler con un diámetro interno de broca de 5.1 mm; en total se obtuvieron 51 muestras.
Los núcleos de crecimiento fueron colocados en pajillas de plástico perforadas, estas se dejaron secar a temperatura ambiente durante 8 días; después se fijaron con pegamento en molduras de madera acanaladas y se pulieron con lijas de grano grueso (240) a fino (1,200) para facilitar la visibilidad de anillos de crecimiento.
Los análisis se llevaron a cabo en el Laboratorio de Anatomía y Tecnología de la Madera de la División de Ciencias Forestales. El fechado de los anillos de crecimiento se realizó siguiendo técnicas de dendrocronología convencionales (Stokes y Smiley, 1996). El conteo de los anillos se efectuó desde la corteza hacia la médula utilizando un estereoscopio Leica Zoom 2000; durante este proceso se identificaron microanillos y anillos falsos. Posteriormente, se procedió a la medición del ancho total del anillo, así como de la madera temprana y tardía, empleando un medidor de incrementos Carl Zeiss.
Concluida la medición de los anillos de crecimiento, se verificó la calidad del fechado utilizando la librería dplR (dendrochronology program library in R) (Bunn, 2008) del software RStudio versión 2024.04.2 Build 764 (Posit team, 2024). El análisis consistió en correlacionar las series individuales obtenidas de las muestras con la cronología maestra, utilizando segmentos de 50 años con traslapes de 25 años. Este procedimiento permitió identificar y corregir errores en el fechado (Bunn, 2010). Una vez verificado el fechado utilizando las librerías dplR y tidyverse, se generaron las cronologías estándar y residual de anillo total, madera temprana y tardía. A éstas se les aplicó una curva de ajuste (Spline a 10 años) para descartar la varianza a causa de factores asociados a edad y condiciones del sitio (Cook y Kairiukstis, 1990). También se estimaron los parámetros estadísticos de las cronologías —correlación entre series, desviación estándar, autocorrelación de primer orden, sensibilidad media, señal ruido y señal poblacional expresada— (Fritts, 1976).
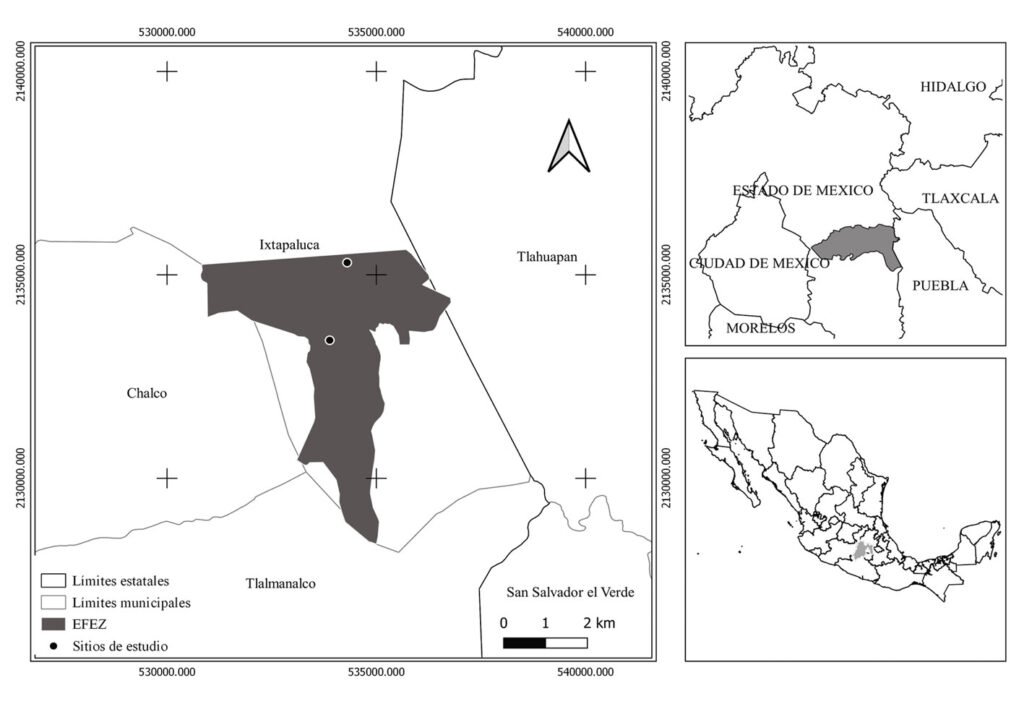
Figura 1. Localización del área de estudio Estación Forestal Experimental Zoquiapan (EFEZ).
En la región cercana a la zona de estudio se ubicaron 6 estaciones meteorológicas; sin embargo, los registros comprenden el periodo de 1961 al 2010, insuficientes para analizar el periodo de interés que se extiende hasta el 2023. Por tanto, se obtuvieron datos de precipitación y temperatura de la base de reanálisis NASA Prediction Of Worldwide Energy Resources (NASA POWER) (NASA POWER, 2023). Estos datos fueron sometidos a un análisis de correlación mediante el coeficiente de Pearson, con el propósito de compararlos con los registros de estaciones meteorológicas y confirmar la similitud y confiabilidad de la información proporcionada por NASA POWER (fig. 2). La vinculación de los eventos de sequía en el Estado de México con el índice de ancho de anillo total de P. montezumae se realizó con indicadores del Tzolkin Monitor de Sequía de Mesoamérica, desarrollado por el Instituto Mexicano de Tecnología del Agua (IMTA, 2020).
Para evaluar la influencia climática en el crecimiento radial de Pinus montezumae, se aplicó un análisis de función de respuesta, el cual consistió en obtener el coeficiente de correlación de Pearson, si los datos tienen una distribución normal en otro caso se obtuvo el coeficiente de Spearman entre las variables climáticas (precipitación total mensual, temperatura máxima, media y mínima) con los índices de las cronologías estandarizadas y residuales correspondientes al ancho de anillo total, madera temprana y madera tardía. Este análisis se hizo para el periodo 1982-2023 utilizando el coeficiente adecuado tras verificar la normalidad de los datos mediante la prueba de Shapiro-Wilk. Esto permitió determinar si el efecto de la variabilidad climática influye en el incremento radial del árbol.
Resultados
Se evaluó la relación entre la variabilidad climática y el crecimiento radial de Pinus montezumae en la Estación Forestal Experimental Zoquiapan. Para ello, se analizaron las correlaciones entre las cronologías estandarizadas y residuales del anillo total, madera temprana y tardía, con las variables climáticas de temperatura (promedio, máxima y mínima) y precipitación mensual. Los 26 árboles de Pinus montezumae presentaron diámetros que oscilaron entre 14 y 120 cm, y edades entre 34 y 123 años. De los 51 núcleos de crecimiento colectados, 45 (88%) fueron fechados con éxito; los restantes se excluyeron debido a la presencia de anillos falsos y ausentes.
Las cronologías del ancho de anillo total (AT), madera temprana (MTe) y madera tardía (MTa) abarcan un periodo común de 123 años, comprendido entre 1901 y 2023. La muestra más longeva data de 1901, mientras que la más reciente corresponde a 1981. El análisis de correlación entre las series de anillos de crecimiento, por segmentos de 50 años con traslapes de 25 años, indican que la calidad de fechado y la datación cruzada es correcta (p ˂ 0.01), superando la correlación mínima r = 0.3281 (tabla 1) establecida por Holmes (1983). Asimismo, los parámetros estadísticos de las series de anillos de crecimiento (tabla 1) respaldan el buen potencial dendroclimático de P. montezumae para estudios del crecimiento radial frente a la variabilidad climática.
Para identificar la función de respuesta del crecimiento radial a las variaciones climáticas, se generaron 2 cronologías: estándar y residual, a partir del índice de ancho de anillo total, madera temprana y madera tardía. Estas cronologías indican que el crecimiento promedio anual del ancho de anillo fue de 1.03 mm (tabla 2) y reflejan tendencias asociadas con periodos húmedos y de sequía. La cronología estándar fue la que determinó mejor la respuesta de P. montezumae a las fluctuaciones del clima, de acuerdo con la media del ancho de anillo (AT= 1.03, MTe = 1.0, MTa = 0.991), sensibilidad media (AT = 0.279, MTe = 0.138, MTa = 0.135), señal ruido (AT = 21.308, MTe = 15.353, MTa = 26.199). Las cronologías del índice de ancho de anillo total (IAA), madera temprana y madera tardía, tanto en su forma estándar (St) como residual (Rs), para P. montezumae comprenden un periodo de 123 años. Ambas series destacaron intervalos en los que el crecimiento radial fue superior a la media (fig. 3A), específicamente en los años 1901-1910, 1913-1917, 1942-1946, 1970-1972. En contraste, se observaron periodos de crecimiento por debajo del promedio durante 1923-1931, 1947-1950, 1961-1965, 1968-1976, 1998-2001 y 2021-2023.
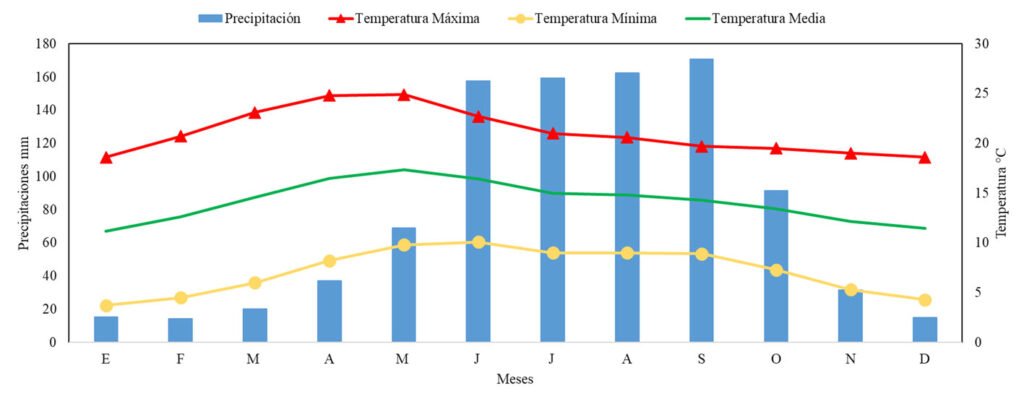
Figura 2. Climograma de la región EFEZ.
Las cronologías estándar y residual de la madera temprana presentaron una marcada variabilidad interanual en el crecimiento radial (fig. 3B). Los periodos con mayor desarrollo de madera temprana corresponden a los años 1901-1910, 1913-1916, 1931-1935, 1943-1947, 1978-1990 y 2002-2012. En contraste, se registró una menor formación de madera temprana durante los periodos 1923-1930, 1936-1942, 1961-1965, 1973-1977 y 1998-2001. Con respecto a las cronologías estándar y residual de la madera tardía también exhibieron una alta fluctuación interanual. Los periodos con mayor formación de madera tardía corresponden a los años 1903- 1904, 1907, 1909-1910, 1912, 1914-1915, 1917, 1923-1926, 1928-1930, 1935, 1937-1939, 1941- 1944, 1949-1951, 1954-1956, 1958-1962, 1969-1971, 1974-1977, 1983-1985, 1992 1995, 1996, 1998, 2003-2006, 2009-2010, 2014-2016 y 2018-2021 (fig. 3C).
Los eventos de sequía descritos por Domínguez (2016) y registrados en el Tzolkin Monitor de Sequía de Mesoamérica se asociaron a la cronología de ancho de anillo (fig. 4) y se observó que aquellos periodos de tiempo donde el crecimiento radial del árbol fue menor a la media anual corresponden a años con eventos de sequía, como son: la sequía de 1908 a 1910, las sequías de 1925, 1935, la de 1948 a 1954 que fue muy severa, 1957, 1960 a 1964, 1969, 1977. También se puede identificar que en el periodo de 1980 al 2023, el crecimiento radial estuvo influenciado por la presencia de sequías anuales de intensidad moderada, destacando el registro en la cronología de sequías severas en 2001, 2006-2012 y de 2018-2023.
Tabla 1
Estadísticos descriptivos de las series de anillos de crecimiento de P. montezumae.
| Parámetro estadístico | Anillo total | Madera temprana | Madera tardía |
| Intercorrelación entre series | 0.657 | 0.601 | 0.673 |
| Sensibilidad media | 0.281 | 0.324 | 0.278 |
| Desviación estándar | 0.875 | 0.758 | 0.223 |
| Autocorrelación de primer orden | 0.529 | 0.518 | 0.520 |
| Relación señal-ruido | 14.621 | 11.159 | 19.533 |
La función de respuesta del crecimiento radial de P. montezumae y la variabilidad del clima se analizó en el periodo común de 1981-2023. El análisis de correlación (coeficiente de Pearson), entre la cronología estandarizada de anillo total y la precipitación mensual, mostró correlaciones tanto positivas como negativas a lo largo del ciclo de crecimiento. La correlación entre el IAA estándar y precipitación mensual fue significativa en julio (r = 0.329, p = 0.033); mientras que, en el índice de madera temprana fue para septiembre del año anterior (r = -0.35, p = 0.049) y julio (r = 0.331, p = 0.032), para madera tardía fue en marzo (r = 0.373, p = 0.015) (fig. 5A). En los análisis de las cronologías residuales y la precipitación, la relación fue similar a la estándar, en el caso del índice de ancho de anillo fue significativa para septiembre del año anterior (r = -0.308, p = 0.046) y julio (r = 0.306, p = 0.048). Con respecto a la madera temprana, en septiembre del año anterior (r = -0.341, p = 0.027) y en madera tardía, en noviembre (r = 0.333, p = 0.031) y marzo (r = 0.373, p = 0.015) (fig. 5B).
La reciprocidad entre los índices de ancho de anillo AT, MTe, MTa y la temperatura (máxima, media y mínima) en el periodo común entre los años 1981 a 2023, reveló correlaciones negativas y positivas. Las asociaciones de los índices de ancho de anillo y la temperatura máxima, media y mínima con valores de coeficientes de correlación estadísticamente significativos (tabla 3), indican que las temperaturas máximas de septiembre y octubre del año anterior favorecieron el crecimiento del anillo total y la formación de madera temprana; sin embargo, en la cronología residual la temperatura máxima de enero afectó de forma negativa el desarrollo de madera tardía (fig. 6A, B).
Tabla 2
Estadísticas descriptivas de las cronologías estandarizada y residual de P. montezumae.
| Parámetros estadísticos | Cronologías | |||||
| Estándar | Residual | |||||
| Anillo total | Madera temprana | Madera tardía | Anillo total | Madera temprana | Madera tardía | |
| Ancho de anillo mm (media) | 1.03 | 1.00 | 0.991 | 0.992 | 0.987 | 0.991 |
| Sensibilidad media | 0.279 | 0.138 | 0.135 | 0.166 | 0.179 | 0.179 |
| Desviación estándar | 0.196 | 0.212 | 0.220 | 0.183 | 0.199 | 0.226 |
| Coeficiente de Gini | 0.110 | 0.120 | 0.124 | 0.104 | 0.113 | 0.124 |
| Autocorrelación de 1er orden | 0.125 | 0.092 | 0.196 | -0.162 | -0.180 | -0.100 |
| Relación señal-ruido | 21.308 | 15.353 | 26.199 | 27.802 | 20.144 | 34.543 |
| Señal expresada de la población (eps) | 0.950 | 0.934 | 0.958 | 0.965 | 0.953 | 0.972 |
Las correlaciones entre la temperatura mínima de octubre, noviembre, diciembre del año anterior con el índice de ancho de anillo de MTa y en diciembre con AT (fig. 6C, D), en la cronología estándar, fueron estadísticamente significativas; mientras que en la cronología residual solo fue significativa para noviembre; lo cual significa que las temperaturas mínimas de invierno favorecen la formación de madera tardía y puede deberse a que los árboles inician su etapa de letargo. Finalmente, la correlación entre la temperatura media de septiembre y octubre del año anterior fue estadísticamente significativa con el índice de madera tardía en la cronología estándar; en la cronología residual, únicamente la temperatura de octubre mostró una relación significativa (fig. 6E, F). Estos resultados reafirman la influencia de las temperaturas de otoño en la formación de madera tardía.
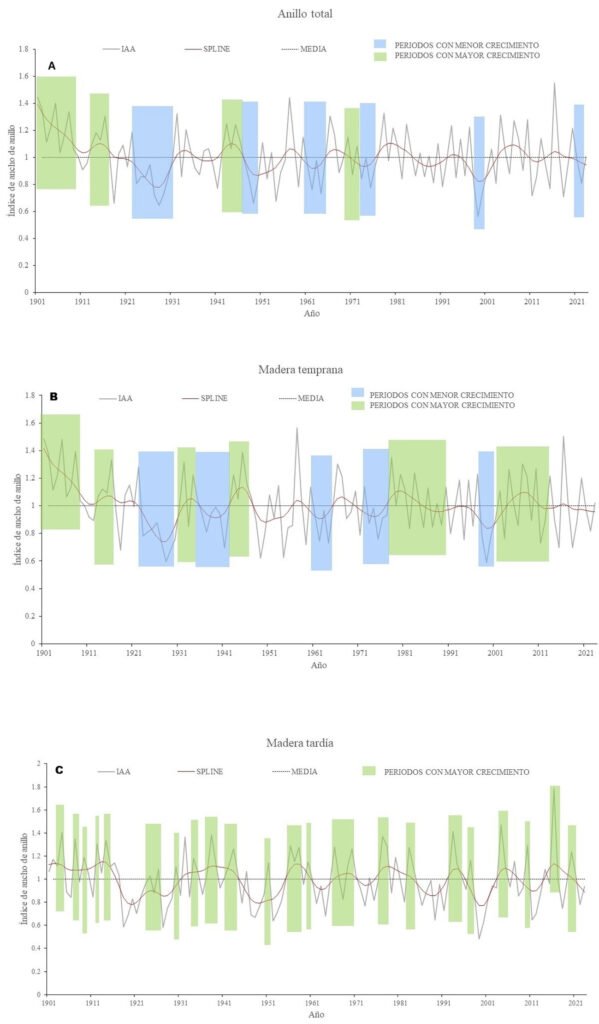
Figura 3. Cronología de índices de ancho de anillo de P. montezumae.
Discusión
Los resultados obtenidos aportan evidencia sobre la respuesta de Pinus montezumae a las condiciones ambientales, ya que se lograron identificar patrones de crecimiento mediante el fechado en 88% de las muestras. El porcentaje es considerado alto, puesto que en estudios dendrocronológicos es común excluir hasta 40% de las muestras debido a irregularidades en los anillos de crecimiento (Cerano-Paredes et al., 2014). La baja presencia de anillos falsos (menos de 5%) encontrada en P. montezumae coincide con lo registrado por Gutiérrez-García y Ricker (2019) y Villanueva-Díaz et al. (2016) para especies que habitan en la alta montaña.
Tabla 3
Coeficientes de correlación de Pearson de la relación entre índices de ancho de anillo y temperatura. * Mes del año anterior al inicio de la temporada de crecimiento.
| Temperatura | Mes | Índices de ancho de anillo de las Cronologías | |||||
| Estándar | Residual | ||||||
| Anillo total | Madera Temprana | Madera Tardía | Anillo total | Madera Temprana | Madera Tardía | ||
| Máxima | Septiembre* | r = 0.365 p = 0.017 | r = 0.353 p = 0.021 | – | r = 0.361 p = 0.018 | r = 0.356 p = 0.020 | – |
| Octubre* | r = 0.326 p = 0.034 | – | – | – | – | – | |
| Enero | – | – | – | – | – | r = -0.318 p = 0.039 | |
| Media | Septiembre* | – | – | r = 0.316 p = 0.041 | – | – | – |
| Octubre* | – | – | r = 0.362 p = 0.018 | – | – | r = 0.310 p = 0.045 | |
| Mínima | Octubre* | – | – | r = 0.323 p = 0.036 | – | – | – |
| Noviembre* | – | – | r = 0.365 p = 0.017 | – | – | r = 0.342 p = 0.026 | |
| Diciembre* | r = 0.318 p = 0.039 | – | r = 0.317 p = 0.040 | – | – | – |
Los valores de intercorrelación entre series fueron de 0.657 (anillo total), 0.601 (madera temprana) y 0.673 (madera tardía), similar a lo obtenido por Cortés-Cortés (2020) para P. montezumae en Coyuca de Catalán, Guerrero (r = 0.57), mayor al valor mínimo de 0 establecido por Holmes (1983), lo cual muestra que las intercorrelaciones de P. montezumae son estadísticamente significativas (p < 0.05). De acuerdo con Grissino-Mayer (2001), los valores de sensibilidad media del ancho total del anillo (0.281), madera temprana (0.324) y madera tardía (0.278) de P. montezumae representan una sensibilidad baja (0.135 – 0.179) e intermedia (0.279) en el ancho total del anillo. Estos valores son menores a los reportados para pináceas del norte de México, donde se obtuvieron valores mayores a 0.5 (Constante-García et al., 2010; Santillán-Hernández et al., 2016) y menores a los obtenidos para P. montezumae en Guerrero (0.31), según lo reportado por Cortés-Cortés (2020).
La autocorrelación de primer orden en los 3 índices de ancho de anillo muestra valores entre 0.092 y 0.196, lo cual es un criterio fundamental en estudios dendrocronológicos, debido a que una baja autocorrelación indica que el crecimiento radial no está significativamente influenciado por las condiciones climáticas del año anterior (Constante-García et al., 2010; Fritts, 1976). Estos resultados muestran que la especie es particularmente adecuada para estudios de reconstrucción climática porque permite registrar con mayor precisión las variaciones climáticas interanuales. De acuerdo con Briffa (1995), una cronología de sitio con un valor de señal expresada de la población (EPS) superior a 0.85, indica que el número de muestras es adecuado; en este estudio, se alcanzaron valores superiores, es decir, valores entre 0.934 y 0.972 para los 3 índices de crecimiento (AT, madera temprana y madera tardía), lo cual confirma la robustez de los datos analizados.
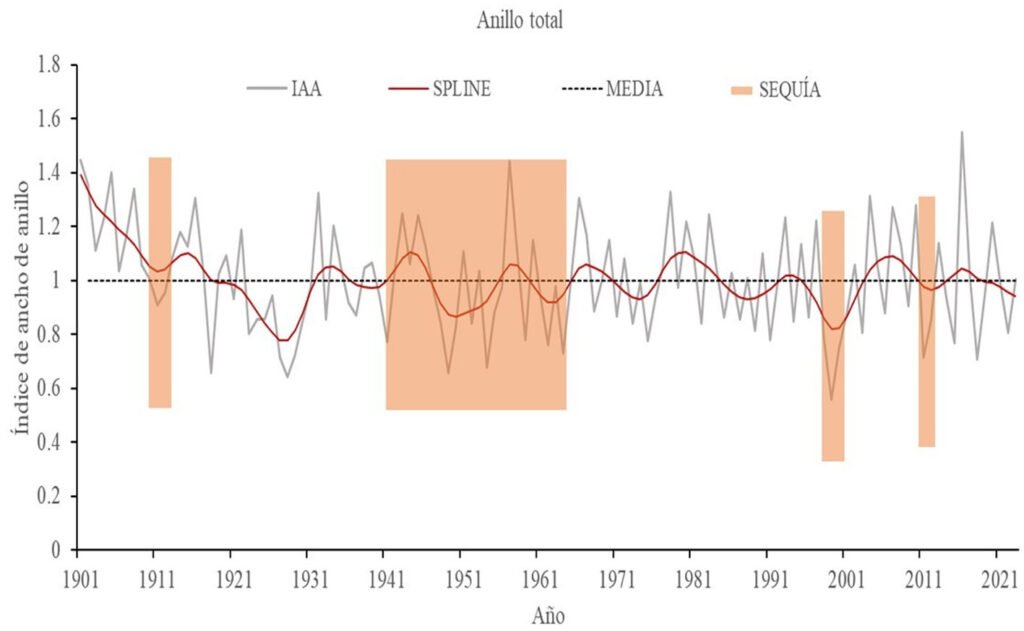
Figura 4. Vinculación de índice de ancho de anillo con eventos de sequía.
Los parámetros dendrocronológicos obtenidos para P. montezumae en este estudio, junto con los resultados reportados para la misma especie (Cortés-Cortés, 2020) y estudios de otras especies del centro del país, reconocidas por su buen potencial en este campo (Cerano-Paredes et al., 2014; Villanueva-Díaz et al., 2015, 2016), respaldan su utilidad en estudios dendrocronológicos. En consecuencia, se concluye que P. montezumae presenta un potencial dendrocronológico de moderado a alto, lo que la hace idónea para investigaciones orientadas a evaluar el impacto del clima y el cambio climático sobre el crecimiento radial. La relación entre el clima y el crecimiento radial indica que la disponibilidad de humedad, particularmente durante el invierno y la primavera, influye positivamente en el crecimiento de la especie. Este comportamiento coincide con un patrón que se presenta desde el sureste de los EUA hasta el centro de México, como lo han señalado diversos estudios (Allende et al., 2016; Cerano-Paredes et al., 2013; St. George et al., 2010; Villanueva-Díaz et al., 2016, 2017). Por lo tanto, las lluvias de baja intensidad, pero de larga duración junto con tasas moderadas de evaporación, permiten la retención de humedad en el suelo, favoreciendo el desarrollo durante los primeros meses de la temporada de crecimiento (Cerano-Paredes et al., 2009, 2011; Cleaveland et al., 2003; Constante-García et al., 2009; Díaz et al., 2002).
Por otra parte, las precipitaciones de verano y otoño del año de crecimiento actual presentan una variabilidad en su relación con las cronologías de P. montezumae; mientras que, los meses de otoño del año previo muestran correlaciones negativas y algunas estadísticamente significativas. Este patrón ha sido documentado también en investigaciones sobre otras especies del género Pinus (Cerano-Paredes et al., 2011, 2013; Chávez-Gándara et al., 2017; López-Hernández et al., 2018). Dicho comportamiento podría explicarse por la abundancia de lluvia durante estos periodos, que supera la capacidad de infiltración del suelo, provoca saturación de sus partículas, genera escurrimientos superficiales y limita así su disponibilidad para el desarrollo del árbol, así como por el hecho de que la mayor parte de los fotosintatos producidos en la temporada de lluvias se asignan al desarrollo de la copa, y se deja en segundo plano el incremento en el diámetro del tronco (Fritts, 1976). No obstante, esta lluvia es de gran importancia, pues parte de ella se almacena en el suelo y puede ser utilizada en la siguiente etapa de crecimiento (Meko et al., 1995).
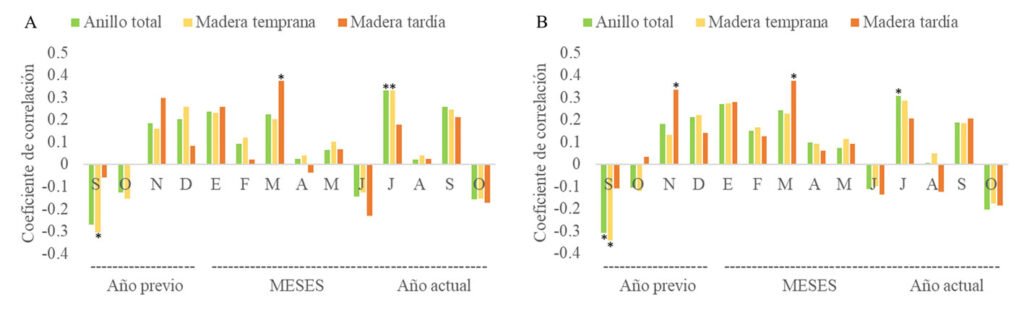
Figura 5. Coeficientes de correlación de precipitación e índices de ancho de anillo: A, estándar; B, residual.
El crecimiento de P. montezumae está principalmente influenciado y regulado por la disponibilidad de humedad durante el otoño del año previo y la primavera del año en curso. Al igual que otras especies estudiadas en investigaciones dendrocronológicas en el norte y centro de México, P. montezumae mostró una respuesta significativamente mayor a la precipitación que a la temperatura (Cerano-Paredes et al., 2013; Constante-García et al., 2010; Villanueva-Díaz et al., 2015, 2017). Los análisis de la temperatura máxima, mínima y media han identificado relaciones positivas entre estos factores y el crecimiento radial de los árboles en diversas partes del mundo, demostrando que la temperatura es un factor determinante en el desarrollo de los bosques, reflejándose en procesos clave, como la reactivación del cambium vascular, la diferenciación celular del xilema y la fotosíntesis, basándose en análisis de índices de ancho de anillo (Antonucci et al., 2017; Li et al., 2017, 2018; Zhu et al., 2016). Este estudio confirma la importancia de la temperatura en el crecimiento del P. montezumae debido a que se logra evidenciar, en algunos meses, el impacto de esta variable. De manera similar a lo observado en el estudio de P. montezumae realizado por Cortés-Cortés (2020), se identificaron principalmente relaciones positivas entre la temperatura máxima y la cronología estandarizada; además, en ambos estudios se observó una disminución en los valores estadísticamente significativos al correlacionarse con la cronología residual.
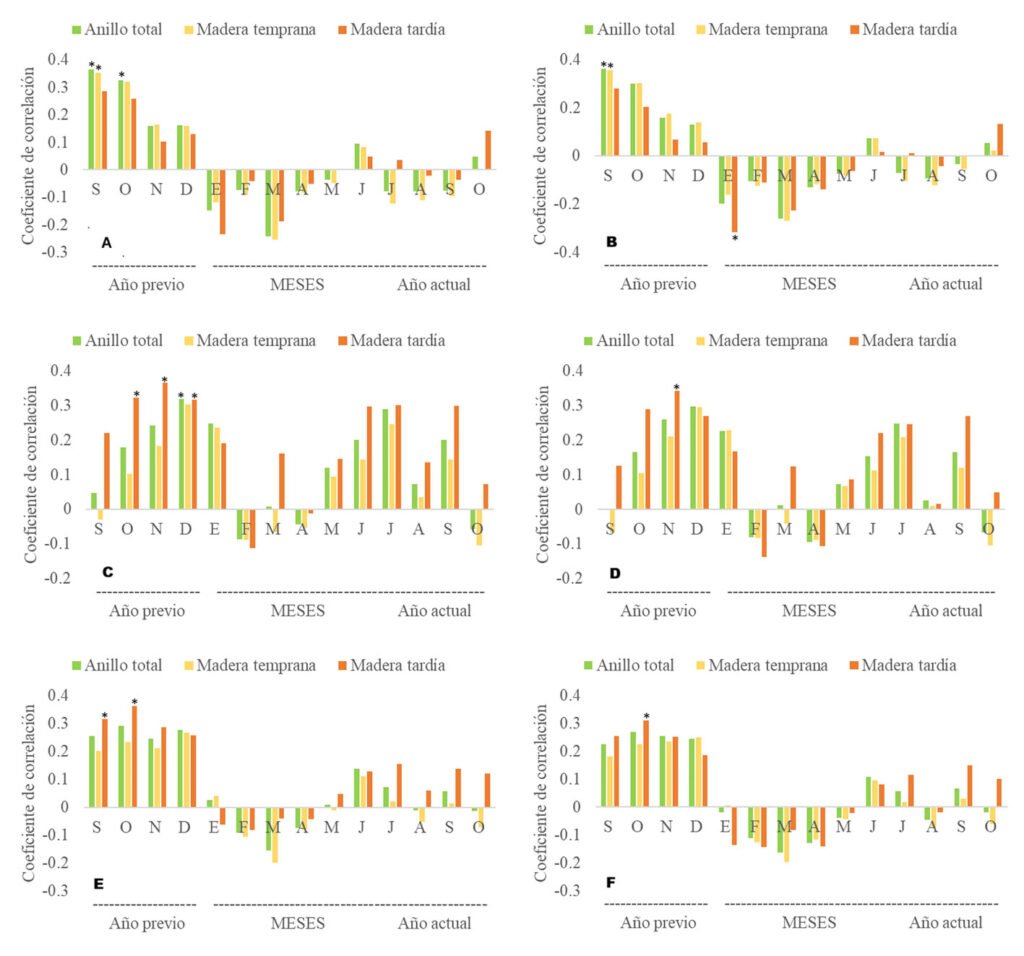
Figura 6. Coeficientes de correlación de temperatura e índices de ancho de anillo: A, relación temperatura máxima-cronología estándar; B, relación temperatura máxima-cronología residual; C, relación temperatura mínima – cronología estándar; D, relación temperatura mínima-cronología residual; E, relación temperatura media-cronología estándar; F, relación temperatura media-cronología residual.
En otros estudios de dendrocronología como el de Pinus hartwegii en el monte Tláloc ubicado también en la Faja Volcánica Transmexicana (Astudillo-Sánchez et al., 2017, 2019) y de Pinus oocarpa Schiede ex Schltdl del ejido Ojo de Agua, La Independencia en Chiapas, se han identificado correlaciones positivas entre las cronologías de ancho de anillo total y la temperatura máxima, éstas sugieren que la temperatura máxima incide en el crecimiento y desarrollo de madera tardía, tanto en la región central del país como en el sur, aunque se trate de especies con diferente rango de distribución (Astudillo-Sánchez et al., 2017, 2019; López-Hernández et al., 2018). La temperatura mínima mostró una influencia positiva en la mayoría de los meses analizados y destacó su efecto sobre la cronología de madera tardía, la cual presentó el mayor número de correlaciones positivas estadísticamente significativas. Estos hallazgos son consistentes con lo reportado en otros estudios dendrocronológicos realizados en México, que han encontrado correlaciones positivas significativas entre las temperaturas invernales y el crecimiento de diversas coníferas (Cerano-Paredes et al., 2012, 2014; Correa-Díaz et al., 2019; Chávez-Gándara et al., 2017; Huante et al., 1991; López-Hernández et al., 2018).
El crecimiento de las especies en respuesta a las temperaturas mínimas se debe, en gran medida, a que estas condiciones favorecen una menor evaporación del suelo y las plantas tienen una mayor disponibilidad de agua para el crecimiento (Cerano-Paredes et al., 2011; Chávez-Gándara et al., 2017). Con relación a la temperatura media, diversos estudios reportan que esta influye de manera negativa en el crecimiento radial (Cortés-Cortés et al., 2020; Manzanilla-Quiñones et al., 2020; Villanueva-Díaz et al., 2016). Sin embargo, en la presente investigación se identificó la presencia de correlaciones positivas, coincidiendo con lo reportado para otras especies de coníferas (Cerano-Paredes et al., 2014; Correa-Díaz et al., 2019). En la cronología residual se observaron correlaciones negativas de enero y mayo, un patrón que también se presentó al correlacionarse con la temperatura máxima. Este comportamiento puede atribuirse tanto al déficit hídrico como a que las temperaturas en la zona de estudio, en ciertos meses, se sitúan fuera del rango óptimo para la fotosíntesis en algunas coníferas, el cual oscila entre 15 y 25 °C, reduciéndose gradualmente en ambos extremos (Huxman et al., 2003; McCullough et al., 2017; Williams et al., 2013).
El crecimiento radial de P. montezumae responde a una interacción entre la temperatura y la precipitación. La disponibilidad de humedad en meses clave, como julio y marzo, junto con las temperaturas máximas en septiembre y octubre del año anterior, regulan conjuntamente la formación del crecimiento anual. Esto sugiere que el aumento en la temperatura puede favorecer el crecimiento solo cuando la precipitación es suficiente para mantener los procesos fisiológicos del árbol. Asimismo, las temperaturas mínimas durante el invierno parecen favorecer la formación de la madera tardía, lo que resalta la importancia de las condiciones térmicas en diferentes fases del ciclo anual. Los resultados coinciden con lo reportado por Harvey et al. (2020), quienes documentaron que el crecimiento radial en bosques templados está influenciado positivamente por inviernos cálidos, pero el efecto favorable del aumento temperatura puede verse limitado si no existe suficiente precipitación durante el verano. Por lo tanto, el impacto del cambio climático en el crecimiento de P. montezumae dependerá de la variación simultánea de la precipitación y la temperatura, por lo que es importante considerar ambas variables para predecir la respuesta de la especie en escenarios futuros.
Agradecimientos
A la Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti) por el otorgamiento de una beca para estudios de maestría al primer autor. A la División de Ciencias Forestales de la Universidad Autónoma Chapingo y al personal responsable de la Estación Forestal Experimental Zoquiapan, por el acceso y apoyo para la realización de esta investigación.
Referencias
Aber, J., Neilson, R. P., McNulty, S., Lenihan, J. M., Bachelet, D. y Drapek, R. J. (2001). Forest processes and global environmental change: predicting the effects of individual and multiple stressors. BioScience, 51, 735. https://doi.org/10.1641/0006-3568(2001)051
Allende, T. C., Mendoza, M. E., Pérez-Salicrup, D. R., Villanueva-Díaz, J. y Lara, A. (2016). Climatic responses of Pinus pseudostrobus and Abies religiosa in the Monarch Butterfly Biosphere Reserve, Central Mexico. Dendrochronologia, 38, 103–116. https://doi.org/10.1016/j.dendro.2016.04.002
Antonucci, S., Rossi, S., Deslauriers, A., Morin, H., Lombardi, F., Marchetti, M. et al. (2017). Large scale estimation of xylem phenology in black spruce through remote sensing. Agricultural and Forest Meteorology, 233, 92–100. https://doi.org/10.1016/j.agrformet.2016.11.011
Astudillo-Sánchez, C. C., Fowler, M. S., Villanueva-Díaz, J., Endara-Agramont, A. R. y Soria-Díaz, L. (2019). Recruitment and facilitation in Pinus hartwegii, a Mexican alpine treeline ecotone, with potential responses to climate warming. Trees, 33, 1087–1100. https://doi.org/10.1007/s00468-019-01844-3
Astudillo-Sánchez, C. C., Villanueva-Díaz, J., Endara-Agramont, A. R., Nava-Bernal, G. E. y Gómez-Albores, M. A. (2017). Climatic variability at the treeline of Monte Tlaloc, Mexico: a dendrochronological approach. Trees, 31, 441–453. https://doi.org/10.1007/s00468-016-1460-z
Benito, F. G. (2014). Archivos climáticos y paleohidrológicos. Introducción a datos proxy y su análisis. Madrid: CSIC- Museo Nacional de Ciencias Naturales.
Briffa, K. R. (1995). Interpreting high-resolution proxy climate data. The example of dendroclimatology. En H. von Storch y A. Navarra (Eds.), Analysis of climate variability, applications of statistical techniques (pp. 77–94). Berlin, Heidelberg: Springer. https://doi.org/10.1007/978-3-662-03167-4_5
Bunn, A. G. (2010). Statistical and visual crossdating in R using the dplR library. Dendrochronologia, 28, 251–258. https://doi.org/10.1016/j.dendro.2009.12.001
Bunn, A.G. (2008). A dendrochronology program library in R (dplR). Dendrochronologia, 26, 115–124. https://doi.org/10.1016/j.dendro.2008.01.002
Cassman, K. G. y Wood, S. (2005) Cultivated systems. En R. Hassan, R. Scholes y N. Ash (Eds.), Ecosystems and human well-being: current state and trends (pp. 745–794). Washington, DC; Millennium Ecosystem Assessment – Island Press.
Cerano-Paredes, J., Villanueva-Díaz J., Cervantes-Martínez, R., Vázquez-Selem, L., Trucios-Caciano, R. y Guerra-de la Cruz, V. (2014). Reconstrucción de precipitación invierno-primavera para el Parque Nacional Pico de Tancítaro, Michoacán. Investigaciones Geográficas, 83, 41–54. https://doi.org/10.14350/rig.35190
Cerano-Paredes, J., Villanueva-Díaz, J., Fulé, P. Z., Arreola-Ávila, J. G., Sánchez-Cohen, I. y Valdez-Cepeda, R. D. (2009). Reconstrucción de 350 años de precipitación para el suroeste de Chihuahua, México. Madera y Bosques, 15, 27–44. https://doi.org/10.21829/myb.2009.1521189
Cerano-Paredes, J., Villanueva-Díaz, J., Valdez-Cepeda, R. D., Méndez-González, J. y Constante-García, V. (2011). Sequías reconstruidas en los últimos 600 años para el noreste de México. Revista Mexicana de Ciencias Agrícolas, 2, 235–249. https://doi.org/10.21829/myb.2009.1521189
Cerano-Paredes, J., Rivera-González, M., Estrada-Ávalos, J., Trucios-Caciano, R. y Ríos-Saucedo, J. C. (2012). Análisis dendrocronológico de Pinus cooperi en Durango, México. Agrofaz, 12, 81–88.
Cerano-Paredes, J., Villanueva-Díaz, J., Cervantes-Martínez, R., Trucios-Caciano, R. y Guerrero-Soto, J. L. (2013). Reconstrucción de sequías fuertes en el Parque Nacional “Pico de Tancítaro”, Michoacán. Revista Chapingo, Serie Zonas Áridas, 2, 57–62. https://doi.org/10.5154/r.rchsza.2012.06.026
Chávez-Gándara, M. P., Cerano-Paredes, J., Nájera-Luna, J. A., Pereda-Breceda, V., Esquivel-Arriaga, G., Cervantes-Martínez, R. et al. (2017). Reconstrucción de la precipitación invierno-primavera con base en anillos de crecimiento de árboles para la región de San Dimas, Durango, México. Bosque, 38, 387–399. https://dx.doi.org/10.4067/S0717-92002017000200016
Cleaveland, M. K., Stahle, D. W., Therrell, M. D., Villanueva-Díaz, J. y Burns, B. T. (2003). Tree-ring reconstructed winter precipitation and tropical teleconnections in Durango, Mexico. Climatic Change, 59, 369–388. https://doi.org/10.1023/A:1024835630188
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2013). Programa de manejo del Parque Nacional Iztaccíhuatl-Popocatépetl. México. Recuperado el 15 agosto, 2024 de: https://conanp.gob.mx/conanp/dominios/iztapopo/documentos/programa_demanejo_izta_popo.pdf
Constante-García, V., Villanueva-Díaz, J., Cerano-Paredes, J., Cornejo-Oviedo, E. H. y Valencia-Manzo, S. (2009). Dendrocronología de Pinus cembroides Zucc. y reconstrucción de precipitación estacional para el Sureste de Coahuila. Ciencia Forestal en México, 34, 17–39.
Constante-García, V., Villanueva-Díaz, J., Cerano-Paredes, J. y Estrada-Ávalos, J. (2010). Parámetros para definir el potencial dendrocronológico. Folleto técnico número 19. Gómez Palacio, Durango: Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias.
Cook, E. R. y Kairiukstis, L. A. (1990) Methods of dendrochronology. Applications in the environmental sciences. Dordrecht, Holanda: International Institute for Applied Systems Analysis. Kluwer Academic Publishers.
Correa-Díaz, A., Silva, L. C. R., Horwath, W. R., Gómez-Guerrero, A., Vargas-Hernández, J., Villanueva-Díaz, J. et al. (2019). Linking remote sensing and dendrochronology to quantify climate-induced shifts in high-elevation forests over space and time. Journal of Geophysical Research: Biogeosciences, 124, 166–183. https://doi.org/10.1029/2018JG004687
Cortés-Cortés, O., Cornejo-Oviedo, E. H., Cerano-Paredes, J., Cervantes-Martínez, R., Flores-López, C. y Valencia-Manzo, S. (2020). Relationship between climate variability and radial growth of Pinus montezumae Lamb. in Coyuca de Catalán, Guerrero. Revista Chapingo, Serie Ciencias Forestales y del Ambiente, 27, 109–126. https://doi.org/10.5154/r.rchscfa.2020.03.012
Díaz, S. C., Therrell, M. D., Stahle, D. W. y Cleaveland, M. K. (2002). Chihuahua (México) winter-spring precipitation reconstructed from tree rings, 1647-1992. Climate Research, 22, 237–244. https://doi.org/10.3354/cr022237
Domínguez, J. (2016). Revisión histórica de las sequías en México: de la explicación divina a la incorporación de la ciencia. Tecnología y Ciencias del Agua, 7, 77–93.
Farjon, A., Pérez-de la Rosa, J. A. y Styles, B. T. (1997). Guía de campo de los pinos de México y América Central. Oxford, UK: The Royal Botanic Gardens, Kew/ Instituto Forestal de Oxford, Universidad de Oxford.
Fritts, H. C. (1976). Tree rings and climate. New York: Academic Press Inc.
Grissino-Mayer, H. D. (2001). Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research, 57, 205–221.
Gutiérrez-García, G. y Ricker, M. (2019). Influencia del clima en el crecimiento radial en cuatro especies de coníferas en la sierra de San Antonio Peña Nevada (Nuevo León, México). Revista Mexicana de Biodiversidad, 90, e902676. https://doi.org/10.22201/ib.20078706e.2019.90.2676
Harvey, J. E., Smiljanić, M., Scharnweber, T., Buras, A., Cedro, A. Cruz-García, R. et al. (2020). Tree growth influenced by warming winter climate and summer moisture availability in northern temperate forests. Global Change Biology, 26, 2505–2518. https://doi.org/10.1111/gcb.14966
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 43, 69–78.
Howard, I. M., Stahle D. W., Torbenson, M. C. A. y Griffin, D. (2021). The summer precipitation response of latewood tree-ring chronologies in the southwestern United States. International Journal of Climatology, 41, 2913–2933. https://doi.org/10.1002/joc.6997
Huante, P., Rincón, E. y Swetnam, T. W. (1991). Dendrochronology of Abies religiosa in Michoacán, Mexico. Tree-Ring Bulletin, 51, 15–28.
Huxman, T. E., Turnipseed, A. A., Sparks, J. P., Harley, P. C. y Monson, R. K. (2003). Temperature as a control over ecosystem CO2 fluxes in a high-elevation, subalpine forest. Oecologia, 134, 537–546. https://doi.org/10.1007/s00442-002-1131-1
IMTA (Instituto Mexicano de la Tecnología del Agua). (2020). Tzolkin Monitor de Sequía de Mesoamérica. Instituto Mexicano de la Tecnología del Agua, Secretaría de Medio Ambiente Recursos Naturales, México. Recuperado el 18 agosto, 2024 de: http://galileo.imta.mx/Sequias/moseq/index.html
IPCC (Intergovernmental Panel on Climate Change). (2014). Climate Change 2014. Synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. Génova, Suiza: IPCC. Disponible: https://archive.ipcc.ch/pdf/assessment-report/ar5/syr/SYR_AR5_FINAL_full_wcover.pdf
IPCC (Intergovernmental Panel on Climate Change). (2021). Climate change 2021. A summary for all. IPCC. Disponible: https://www.ipcc.ch/report/ar6/wg1/downloads/outreach/IPCC_AR6_WGI_SummaryForAll.pdf
Li, M., Duan, J., Wang, L. y Zhu, H. (2018). Late summer temperature reconstruction based on tree-ring density for Sygera Mountain, southeastern Tibetan Plateau. Global and Planetary Change, 163, 10–17. https://doi.org/10.1016/j.gloplacha.2018.02.005
Li, X., Liang, E., Gricar, J., Rossi, S., Čufar, K. y Ellison, A. M. (2017). Critical minimum temperature limits xylogenesis and maintains treelines on the southeastern Tibetan Plateau. Science Bulletin, 62, 804–812. https://doi.org/10.1016/).scib.2017.04.025
López-Hernández, M. I., Cerano-Paredes, J., Valencia-Manzo, S., Cornejo-Oviedo, E. H., Villanueva-Díaz, J., Cervantes-Martínez, R. et al. (2018). Respuesta del crecimiento de Pinus oocarpa a variables climáticas en Chiapas, México. Biología Tropical, 66, 1580–1596. http://dx.doi.org/10.15517/rbt.v66i4.32663
Manzanilla-Quiñones, U., Aguirre-Calderón, O. A., Jiménez-Pérez, J. y Villanueva-Díaz, J. (2020). Sensibilidad climática en anchuras de anillos de crecimiento de Pinus hartwegii: una especie alpina mexicana con potencial dendroclimático. Revista Mexicana de Biodiversidad, 91, e913117. https://doi.org/10.22201/ib.20078706e.2020.91.3117
McCullough, I. M., Davis, F. W. y Williams, A. P. (2017). A range of possibilities: assessing geographic variation in climate sensitivity of ponderosa pine using tree rings. Forest Ecology and Management, 402, 223–233. https://doi.org/10.1016/j.foreco.2017.07.025
Meko, D., Stockton, C. W. y Boggess, W. R. (1995). The tree-ring record of severe sustained drought. JAWRA Journal of the American Water Resources Association, 31, 789–801. https://doi.org/10.1111/j.1752-1688.1995.tb03401.x
Meko, D. M. y Baisan, C. H. (2001). Pilot study of latewood-width of conifers as an indicator of variability of summer rainfall in the North American monsoon region. International Journal of Climatology, 21, 697–708. https://doi.org/10.1002/joc.646
NASA POWER. (2023). Prediction Of Worldwide Energy Resources. National Aeronautics and Space Administration (NASA), Washington D. C. Recuperado el 31 junio, 2024 de: https://power.larc.nasa.gov
Osorio-Osorio, J. A., Astudillo-Sánchez, C. C., Villanueva-Díaz, J., Soria-Díaz, L. y Vargas-Tristán, V. (2020). Reconstrucción histórica de la precipitación en la Reserva de la Biosfera El Cielo, México, mediante anillos de crecimiento en Taxodium mucronatum (Cupressaceae). Revista de Biología Tropical, 8, 818–832. https://doi.org/10.15517/rbt.v68i3.39624
Posit team (2024). RStudio. Integrated development environment for R. Posit Software, PBC, Boston, MA. https://www.posit.co/
Rzedowski, J. (2006). Vegetación de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. http://www.conabio.gob.mx/institucion/centrodoc/doctos/librosdigitales/VegetaciondeMexico/Portadaypaglegales.pdf
Santillán-Hernández, M., Cornejo-Oviedo, E. H., Villanueva-Díaz, J., Cerano-Paredes, J., Valencia-Manzo, S. y Capó-Arteaga, M. Á. (2016). Potencial dendroclimático de Pinus pinceana Gordon en la Sierra Madre Oriental. Madera y Bosques, 16, 17–30. https://doi.org/10.21829/myb.2010.1611177
Schweingruber, F. H. (1988). Tree rings: basics and applications of dendrochronology. Holanda: Dordrecht, Holanda: Kluwer Academic Publishers Group.
St. George, S., Meko, D. M. y Cook, E. R. (2010). The seasonality of precipitation signals embedded within the North American Drought Atlas. The Holocene, 20, 983–988. https://doi.org/10.1177/0959683610365937
Stokes, M. A. y Smiley, T. L. (1996). An introduction to tree-ring dating. Tucson: The University of Arizona Press.
Vaganov, E. A., Hughes, M. K. y Shashkin, A. V. (2006). Growth dynamics of conifer tree rings: images of past and future environments. Berlín, Heidelberg: Springer-Verlag.
Villanueva-Díaz, J., Rubio-Camacho, E. A., Chávez-Durán, A. A., Zavala-Aguirre, J. L., Cerano-Paredes, J. y Martínez-Sifuentes, A. R. (2017). Respuesta climática de Pinus oocarpa Schiede Ex Schetol en el Bosque La Primavera, Jalisco. Madera y Bosques, 24, 1–14. https://doi.org/10.21829/myb.2018.2411464
Villanueva-Díaz, J., Cerano-Paredes, J., Vázquez-Selem, L., Stahle, D. W., Fulé, P. Z., Yocom, L. L. et al. (2015). Red dendrocronológica del pino de altura (Pinus hartwegii Lindl.) para estudios dendroclimáticos en el noreste y centro de México. Investigaciones Geográficas, 86, 5–14. https://doi.org/10.14350/rig.42003
Villanueva-Díaz, J., Vázquez-Selem, L., Gómez-Guerrero, A., Cerano-Paredes, J., Aguirre-González, N. A. y Franco-Ramos, O. (2016). Potencial dendrocronológico de Juniperus monticola Martínez en el Monte Tláloc, México. Revista Fitotecnia Mexicana, 39, 175–185. https://doi.org/10.35196/rfm.2016.2.175-185
Williams, A. P., Allen, C. D., Macalady, A. K., Griffin, D., Woodhouse, C. A., Meko, D. M. et al. (2013). Temperature as a potent driver of regional forest drought stress and tree mortality. Nature Climate Change, 3, 292–297. https://doi.org/10.1038/nclimate1693
Zhu, L., Zhang, Y., Li. Z., Guo, B. y Wang, X. (2016). A 368-year maximum temperature reconstruction based on tree-ring data in the northwestern Sichuan Plateau (NWSP), China. Climate of the Past, 12, 1485–1498. https://doi.org/10.5194/cp-12-1485-2016