
Genetic control of initial growth and physiological response to water stress in Pinus patula provenance-progenies under nursery conditions
Alberto Pérez-Luna a, Javier López-Upton a, *, José Angel Prieto-Ruíz b, Laura Elena Martínez-Nevárez b y José Ángel Sigala-Rodríguez c
a Colegio de Postgraduados, Campus Montecillo, Carretera Federal México-Texcoco Km. 36.5 Montecillo, 56264 Texcoco, Estado de México, México
b Universidad Juárez del Estado de Durango, Facultad de Ciencias Forestales y Ambientales, Río Papaloapan y Bulevar Durango s/n, Col. Valle del Sur, 34120 Durango, Durango, México
c Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Valle del Guadiana, Carretera Durango-El Mezquital Km. 4.5, Camino Real, 34170 Durango, Durango, México
*Autor para correspondencia: jlopezupton@gmail.com (J. López-Upton)
Recibido: 5 septiembre 2025; aceptado: 24 febrero 2026
Resumen
El cambio climático exige identificar procedencias-progenies resistentes al déficit hídrico para plantaciones forestales. El objetivo fue evaluar el control genético del crecimiento y la resistencia a la sequía en plántulas de 9 meses de 20 familias de 5 procedencias de Pinus patula. Antes del déficit hídrico, se midieron altura, diámetro y biomasa seca, además del potencial hídrico y supervivencia después de 10 días sin riego y posterior rehidratación en vivero. Se determinaron correlaciones fenotípicas y genéticas entre variables morfológicas y bioclimáticas de origen. Hubo diferencias significativas entre procedencias y familias, excepto en biomasa de raíz. La altura, diámetro y biomasa aérea se correlacionaron positivamente con la temperatura mínima y negativamente con la altitud. Las familias de sitios húmedos presentaron mayor mortalidad y las de ambientes secos mostraron mayor resiliencia. La heredabilidad fue alta para la altura (0.74) y moderada para las demás variables (0.20-0.58). Se confirmó control genético en la respuesta al estrés. Las correlaciones fenotípicas fueron bajas; se detectaron correlaciones genéticas negativas moderadas entre tamaño y supervivencia, lo que indicó una compensación funcional entre crecimiento y resistencia a la sequía. El origen del germoplasma es importante para producir plantas adaptadas a ambientes restrictivos y anticipar su desempeño en campo.
Palabras clave: Déficit hídrico; Correlaciones genéticas; Heredabilidad; Parámetros genéticos; Resistencia a sequía
Abstract
Climate change demands the identification of provenances-progenies that are drought-resistant for forest plantations. The objective was to evaluate the genetic control of growth and drought resistance in 9-month-old seedlings from 20 families of 5 provenances of Pinus patula. Before the water stress event, height, diameter, dry biomass were measured, as well as water potential and survival after 10 days without irrigation and subsequent rehydration in the nursery. Phenotypic and genetic correlations were determined between morphological and bioclimatic variables of origin. There were significant differences between provenances and families, except for root biomass. Height, diameter, and aerial biomass were positively correlated with minimum temperature and negatively correlated with altitude. Families from humid sites had higher mortality, while those from dry environments showed greater resilience. Heritability was high for height (0.74) and moderate for the other variables (0.20-0.58). Genetic control in the stress response was confirmed. Phenotypic correlations were low; moderate negative genetic correlations were detected between size and survival, indicating a functional trade-off between growth and drought resistance. The origin of the germplasm is important to produce plants adapted to restrictive environments and to anticipate their field performance.
Keywords: Water deficit; Genetic correlations; Heritability; Genetic parameters; Drought resistance
Introducción
Las variaciones en los patrones de temperatura y precipitación por el cambio climático afectan áreas importantes de producción forestal. Este cambio progresivo podría dificultar la elección de las especies y procedencias a plantar. En los programas de reforestación deben incluirse procedencias-progenies de árboles con características sobresalientes productivas en cantidad y calidad, pero con suficiente plasticidad para adaptarse a condiciones ambientales cambiantes (Leibing et al., 2013). En México Pinus patula Schiede ex Schltdl. et Cham. var. patula tiene amplia distribución sobre la sierra Madre Oriental y el este del Eje Neovolcánico Transversal, por lo que debe poseer amplia diversidad genética y plasticidad; además, es el pino de mayor producción forestal con el inicio de programas de mejoramiento genético en México (Flores y Moctezuma-López, 2021; López-Upton et al., 2015).
Los ensayos de procedencias-progenies son útiles para la valoración genética y adaptativa de los progenitores, ya que permiten el mejoramiento de características de interés por medio de la selección de genotipos sobresalientes (White et al., 2007). Una vez depurados los genotipos inferiores, se favorece la obtención de semilla para establecer plantaciones comerciales y de restauración, donde la adaptación a factores adversos es primordial. Se estima que el déficit hídrico fue la principal causa de mortalidad en las reforestaciones establecidas en México entre 2006 y 2014 con 42% del total (Prieto-Ruíz et al., 2016). En ese sentido, los primeros años desde el establecimiento de plantaciones forestales son críticos para su desarrollo, pues las plantas que generan raíces poco robustas y crecimiento aéreo en exceso no son capaces de contrarrestar los efectos provocados por la sequía (Mejía-Bojórquez et al., 2015). Sin embargo, las plantaciones en México se han establecido recientemente en terrenos con vocación agropecuaria en proceso de reconversión forestal, sitios que se caracterizan por ser suelos franco-arenosos que limitan la retención de agua (Salcido-Ruíz et al., 2015).
La evaluación de la resistencia al déficit hídrico en etapas tempranas mediante ensayos en condiciones controladas ofrece ventajas para los estudios de variación genética en especies forestales (Espinoza et al., 2016). El cultivo de plantas jóvenes bajo condiciones controladas reduce la variabilidad no genética y facilita la detección de características heredables entre poblaciones o familias (Acosta-Rico et al., 2024; White et al., 2007). Estos experimentos permiten aplicar de forma precisa tratamientos de sequía y registrar respuestas fisiológicas y morfológicas asociadas con la resistencia, lo que posibilita una selección temprana de genotipos con mayor tolerancia a la sequía. Investigaciones en Pinus greggii var. australis Donahue et Lopez (López-Upton y Muñoz, 1991) y Pinus pinceana Gordon (Martiñón-Martínez et al., 2010) han demostrado la utilidad de este enfoque para predecir el comportamiento de los árboles en condiciones de campo y aportar información clave a los programas de conservación y mejoramiento genético. El preacondicionamiento en vivero, al someter plantas a déficit hídrico mediante la disminución controlada del riego, ayuda a promover la resistencia a condiciones limitantes de humedad en campo (Ávila-Flores et al., 2014) y aunque esta práctica se ha evaluado principalmente en vivero, requiere que sea explorado su efecto en ensayos de procedencias-progenies, tanto en la etapa de producción como en plantaciones.
Los objetivos de este trabajo fueron: 1) evaluar el control genético sobre la altura, diámetro al cuello de la raíz, componentes de biomasa seca, el potencial hídrico como indicadores del estrés hídrico, así como la supervivencia de procedencias y progenies de plántulas de P. patula, 2) determinar las correlaciones fenotípicas y genéticas entre las variables morfológicas y fisiológicas, y 3) evaluar la posible correlación de estas últimas con la ubicación geográfica y las variables climáticas del origen de las familias. Las hipótesis de investigación fueron: 1) que hubiera un control genético moderadamente alto en las variables morfológicas iniciales (altura y diámetro al cuello de la raíz), debido a que la variación en el vigor temprano entre familias está determinada por la herencia de rasgos de crecimiento optimizados para sus sitios de origen, 2) que la correlación genética entre las variables respuesta fuera alta, debido a la acción de genes comunes, 3) que las variables morfológicas medidas previo al déficit hídrico mostrarán una correlación significativa con el clima de origen, lo que sugeriría que las poblaciones de climas más áridos han desarrollado de manera intrínseca un crecimiento más conservador (menor tamaño) como estrategia de adaptación local.
Materiales y métodos
El estudio se realizó en el vivero de la Facultad de Ciencias Forestales y Ambientales de la Universidad Juárez del estado de Durango (24°00’48.38” N, 104°41’03.64” O a 1,860 m snm). El germoplasma empleado fue recolectado de 20 árboles (familias de medios hermanos) de 5 procedencias del centro de México (4 familias por procedencia; tabla 1). Las poblaciones de distribución de los árboles son bosques templados con asociaciones de pino-encino, suelos franco-limosos y precipitación mayor a 800 mm anuales. Las semillas fueron sometidas a estratificación en frío (5 °C) por 5 semanas (Pérez-Luna, Prieto-Ruíz et al., 2024). La siembra se realizó el 16 de diciembre de 2023 en contenedores individuales de 170 mL con un sustrato en proporción 4:3:3 de turba de musgo, aserrín de pino crudo y corteza de pino compostada, mezcla a la que se añadió 10 g L-1 de fertilizante de lenta liberación (8 meses) MulticoteTM (Haifa Group, Israel) 18N-6P-12K + 2MgO más micronutrientes (B, Cu, Fe, Mn, Mo, Zn y S). Posteriormente, se colocaron 2 semillas por contenedor y se cubrieron con la misma mezcla. El inicio de la germinación se observó 3 semanas después de la siembra. El manejo de las plantas consistió en 3 meses en condiciones de invernadero (establecimiento, riego con regadera manual), 3 meses en condición de malla sombra con 60% de retención de luz (crecimiento rápido, riego con manguera) y 3 meses en intemperización (endurecimiento, riego con manguera).
El ensayo se estableció en bloques completos al azar con 15 repeticiones con parcelas de 1 planta de cada familia. A los 9 meses después de la siembra, se midieron en todas las plantas la altura con una regla graduada en cm y el diámetro al cuello de la raíz con un vernier digital Surebilt® (AutoZone Parts Inc., EUA) con aproximación en mm. Posteriormente, el 17 de septiembre de 2024, las plantas se regaron a capacidad de campo y se suspendió el riego por 10 días para inducir el déficit hídrico. Concluido este periodo, se realizaron 5 mediciones del potencial hídrico (Ѱ) en los días: 1 (Ѱ1), 3 (Ѱ3), 6 (Ѱ6), 8 (Ѱ8) y 10 (Ѱ10). Después de la medición del día 10 (el de menor potencial), las plantas se regaron nuevamente a capacidad de campo. Seis días después de este último riego, el 4 de octubre de 2024, se determinó el potencial hídrico de cada planta (día 16, Ѱ16) y se registró la supervivencia como variable dicotómica (planta viva = 1 y muerta = 0). El potencial hídrico se midió prealba a las 6:00 am en acículas del tercio superior de cada planta, con una cámara de presión tipo Scholander M600 de PMS Instruments Co., Corvallis, OR, USA.
Posteriormente, las plantas fueron desenterradas, se separó la parte aérea de la raíz al nivel del cuello y se secaron en una estufa marca Ecoshel® (EUA) modelo 9024a a 60 °C durante 72 horas para determinar la biomasa seca de la parte aérea y de la raíz en una balanza marca Ohaus® (EUA) modelo Pionner PA512 graduada en 0.01 g. En total, se obtuvieron 12 variables de respuesta: altura, diámetro al cuello de la raíz, biomasa seca de la parte aérea, biomasa seca de la raíz, biomasa seca total, 6 registros de potencial hídrico y la supervivencia. La altura y el diámetro al cuello de la raíz se midieron antes del inicio del déficit hídrico, mientras que la medición de los componentes de biomasa fue después de la última medición de Ѱ.
Los supuestos de distribución normal y homocedasticidad de los residuales de las variables continuas (altura, diámetro al cuello de la raíz, componentes de biomasa y mediciones de potencial hídrico) se probaron con los test de Kolmogorov-Smirnov y Levene, respectivamente. En las variables en que fue posible comprobar estos supuestos, se realizaron análisis de varianza (Anova) para detectar diferencias entre ellas a nivel de procedencia y progenie. Cuando se detectaron diferencias estadísticas (p ≤ 0.05), las variables se contrastaron con pruebas de medias de Tukey. Con el procedimiento MIXED de SAS v9.4 (SAS Institute, 2022) se obtuvieron los componentes de varianza, el Anova y la prueba de Tukey.
La estimación de los componentes de varianza para variables continuas con distribución no normal se realizó mediante el método de mínimos cuadrados, el cual proporciona estimadores no sesgados de los parámetros de varianza sin depender del supuesto de normalidad requerido por los métodos de máxima verosimilitud (Searle, 1995). Finalmente, para evaluar la variable de supervivencia, se empleó un modelo lineal generalizado mixto (GLMM) con distribución binomial con el procedimiento GLIMMIX de SAS v9.4 (SAS Institute, 2022). Se usó el siguiente modelo lineal (1):
Yijk = µ + Bi + Pj + BPij + Fk(j) + eijk (1)
| Procedencia | FAM | LAT | LON | ELEV | TMÁX | TMÍN | TMA | PMA | DD5 | ÍS |
|---|---|---|---|---|---|---|---|---|---|---|
| Huayacocotla, Veracruz | 70 | 20°31.8’ | 98°30.0’ | 2,161 | 23.6 | 4.0 | 13.4 | 1,479 | 3,714 | 0.041 |
| Huayacocotla, Veracruz | 77 | 20°33.6’ | 98°31.2’ | 1,954 | 24.7 | 4.2 | 14.2 | 1,308 | 4,001 | 0.048 |
| Huayacocotla, Veracruz | 87 | 20°34.2’ | 98°30.6’ | 1,917 | 24.8 | 4.3 | 14.4 | 1,293 | 4,052 | 0.049 |
| Huayacocotla, Veracruz | 98 | 20°34.8’ | 98°30.0’ | 2,067 | 23.8 | 3.7 | 13.5 | 1,316 | 3,725 | 0.046 |
| Punto central | 20°33.6’ | 98°30.5’ | 2,025 | 24.2 | 4.1 | 13.9 | 1,349 | 3,873 | 0.046 | |
| Acaxochitlán, Hidalgo | 26 | 20°13.2’ | 98°12.0’ | 2,198 | 24.6 | 4.2 | 14.3 | 1,328 | 4,063 | 0.047 |
| Acaxochitlán, Hidalgo | 28 | 20°13.8’ | 98°11.4’ | 2,208 | 24.5 | 4.1 | 14.3 | 1,310 | 4,051 | 0.048 |
| Acaxochitlán, Hidalgo | 29 | 20°14.4’ | 98°13.2’ | 2,225 | 24.5 | 4.1 | 14.3 | 1,192 | 4,040 | 0.053 |
| Acaxochitlán, Hidalgo | 30 | 20°13.2’ | 98°14.2’ | 2,202 | 24.5 | 4.2 | 14.3 | 1,244 | 4,056 | 0.051 |
| Punto central | 20°13.7’ | 98°12.7’ | 2,208 | 24.5 | 4.2 | 14.3 | 1,269 | 4,053 | 0.050 | |
| Zacatlán, Puebla | 1 | 19°58.8’ | 98°00.0’ | 2,288 | 24.1 | 3.3 | 13.9 | 889 | 3,982 | 0.070 |
| Zacatlán, Puebla | 2 | 19°58.4’ | 98°00.0’ | 2,234 | 24.2 | 3.5 | 14.1 | 895 | 4,055 | 0.071 |
| Zacatlán, Puebla | 19 | 20°04.2’ | 98°05.4’ | 2,385 | 24.5 | 3.2 | 13.8 | 1,395 | 3,912 | 0.044 |
| Zacatlán, Puebla | 22 | 20°03.6’ | 98°06.0’ | 2,456 | 24.3 | 2.9 | 13.5 | 1,269 | 3,825 | 0.048 |
| Punto central | 20°01.3’ | 98°02.9’ | 2,341 | 24.3 | 3.2 | 13.8 | 1,112 | 3,944 | 0.058 | |
| Ahuazotepec, Puebla | 5 | 20°03.6’ | 98°06.0’ | 2,429 | 24.3 | 3.0 | 13.6 | 1,248 | 3,862 | 0.049 |
| Ahuazotepec, Puebla | 11 | 20°04.2’ | 98°06.6’ | 2,414 | 24.5 | 3.1 | 13.7 | 1,352 | 3,895 | 0.046 |
| Ahuazotepec, Puebla | 20 | 20°04.8’ | 98°07.2’ | 2,399 | 24.6 | 3.2 | 13.8 | 1,373 | 3,915 | 0.045 |
| Ahuazotepec, Puebla | 21 | 20°04.8’ | 98°06.0’ | 2,300 | 24.7 | 3.6 | 14.1 | 1,471 | 4,033 | 0.043 |
| Punto central | 20°04.0’ | 98°06.5’ | 2,386 | 24.5 | 3.2 | 13.8 | 1,361 | 3,926 | 0.046 | |
| Pinal de Amoles, Querétaro | 3 | 21°07.2’ | 99°40.2’ | 2,622 | 24.9 | 3.1 | 13.8 | 939 | 3,914 | 0.066 |
| Pinal de Amoles, Querétaro | 6 | 21°09.0’ | 99°41.4’ | 2,866 | 23.2 | 2.1 | 12.3 | 1,050 | 3,369 | 0.055 |
| Pinal de Amoles, Querétaro | 9 | 21°07.8’ | 99°40.8’ | 2,641 | 24.8 | 3.0 | 13.7 | 922 | 3,877 | 0.067 |
| Pinal de Amoles, Querétaro | 14 | 21°06.6’ | 99°38.4’ | 2,511 | 25.6 | 3.5 | 14.4 | 974 | 4,125 | 0.065 |
| Punto central | 21°07.7’ | 99°40.2’ | 2,660 | 24.6 | 2.9 | 13.6 | 971 | 3,821 | 0.063 |
Donde Yijk es el valor observado de la planta en la k-ésima familia dentro de la j-ésima procedencia en el i-ésimo bloque, µ la media poblacional, Bi el efecto fijo del i-ésimo bloque (i = 1, 2, …15), Pj el efecto fijo de la j-ésima procedencia, BPij el efecto fijo de la interacción bloque x procedencia, Fk(j) el efecto aleatorio de la k-ésima familia anidada en procedencia (k = 1, 2, …15) ~ ID (0, σ2f), y eijk el error experimental ~ ID (0, σ2e).
Las variables de ubicación geográfica, temperatura, precipitación, índice de sequía y grados del día mayor a 5 °C del punto central de cada procedencia y del punto de cada familia para el periodo 1961-2023 (tabla 1), se obtuvieron con el sistema del programa ClimateNA v5.3 (Wang et al., 2016). Para determinar posibles tendencias de asociación clinal en los patrones de crecimiento entre procedencias, las características morfológicas y la supervivencia se correlacionaron con los valores geográficos y climáticos del punto central. Las correlaciones se generaron con el coeficiente de Spearman mediante el procedimiento CORR de SAS v9.4 (SAS Institute, 2022).
Los componentes de varianza de familias (σ2f), aditiva (σ2A) y del error (σ2e) se estimaron para determinar la heredabilidad en sentido estricto a nivel individual (h2i) (ecuación 2; Falconer y Mackay, 2006) y su error estándar (ecuación 3; Dickerson, 1969). Se utilizó un coeficiente de determinación genética de 3 (σ2A = 3σ2f) (Sorensen y White, 1988) con la ecuación 4 (Morales-González et al., 2013).
Donde: h2i es la heredabilidad individual, σ2f la varianza de familias, EE (h2i) el error estándar, CVGA el coeficiente de variación genética aditiva (%) y la media general para la característica evaluada.
Se calcularon las correlaciones fenotípicas y genéticas entre las características de las plantas evaluadas, así como la correlación de éstas con las variables geográficas y climáticas del origen de las familias. La covarianza de familias se determinó con la ecuación 6 y las correlaciones genéticas y su error fueron estimadas con las ecuaciones 7 y 8, respectivamente (Falconer y Mackay, 2006).
Donde: Covf (x, y) es la covarianza de familias entre las variables “x” e “y”, rg (x, y) la correlación genética entre “x” e “y”, σ2f(x) y σ2f(y) las varianzas de familias de “x” e “y”, σ2f(x+y) la varianza de familias para la suma de las variables (x + y), EE(rg) el error estándar de la correlación genética, r2g(x,y) el cuadrado del coeficiente de correlación genética entre “x” e “y”, EE(h2ix) el error estándar de la heredabilidad individual de “x” (h2ix), y EE(h2iy) el error estándar de la heredabilidad individual de “y” (h2iy).
Las correlaciones fenotípicas entre las variables morfológicas de las plantas y las variables geográficas y climáticas del sitio de origen de las familias (tabla 1) se obtuvieron con el coeficiente de correlación de Spearman con el procedimiento CORR de SAS v9.4 (SAS Institute, 2022).
Resultados
Las variables de altura, diámetro al cuello de la raíz y componentes de biomasa cumplieron con los supuestos de distribución normal y homocedasticidad. Se detectaron diferencias significativas (p < 0.05) en todas las variables de respuesta a nivel de procedencia y familia, excepto en la biomasa seca de la raíz (tabla 2). La mayor altura se presentó en las familias de Huayacocotla, Veracruz (21.9 cm), que superaron en 30.5% el crecimiento total de las familias que presentaron la menor altura (Pinal de Amoles, Querétaro, con 15.2 cm) (fig. 1a). Al igual que la altura total, el diámetro al cuello de la raíz fue mayor en las familias de Huayacocotla (3.26 mm) y menor en las familias de Pinal de Amoles (2.75 mm) (fig. 1b).
El mayor aporte a la biomasa seca total en las familias de Huayacocotla y Acaxochitlán fue el producido por la parte aérea que representó 55 y 54%, respectivamente, mientras que en las familias de Zacatlán y Pinal de Amoles se observó igual aporte de ambos componentes a la biomasa seca total (fig. 1c, d).
La altura de la planta, el diámetro al cuello de la raíz y la biomasa seca aérea mostraron una fuerte correlación con la elevación del punto central de las procedencias (fig. 1a-c). Esta asociación fue más fuerte en la altura y la biomasa seca aérea con respecto a la elevación de las procedencias con el gradiente de variación de 638 m, mientras que, en el diámetro al cuello de la raíz, aunque también existió esa correlación, fue menor. El mejor crecimiento fue en las plantas de procedencias de menor altitud.
Las mismas variables de las plántulas presentaron fuerte correlación con la temperatura mínima: altura (fig. 2a), diámetro al cuello de la raíz (fig. 2b) y biomasa seca aérea (fig. 2c). No se encontró una asociación significativa entre la biomasa seca de la raíz y la temperatura mínima de origen, así como tampoco con las demás variables ambientales evaluadas. Por lo tanto, la variación observada en este componente no puede atribuirse a los gradientes geográficos o climáticos analizados (fig. 2d).
| Variable | Kolmogorov-Smirnov | Levene | Anova |
|---|---|---|---|
| Altura | 0.1508 | 0.9666 | < 0.0001 |
| Diámetro al cuello de la raíz | 0.1185 | 0.5504 | 0.0081 |
| Biomasa seca parte aérea | 0.0753 | 0.2308 | 0.0198 |
| Biomasa seca de la raíz | 0.065 | 0.1624 | 0.8335 |
| Biomasa seca total | 0.2453 | 0.0532 | 0.1055 |
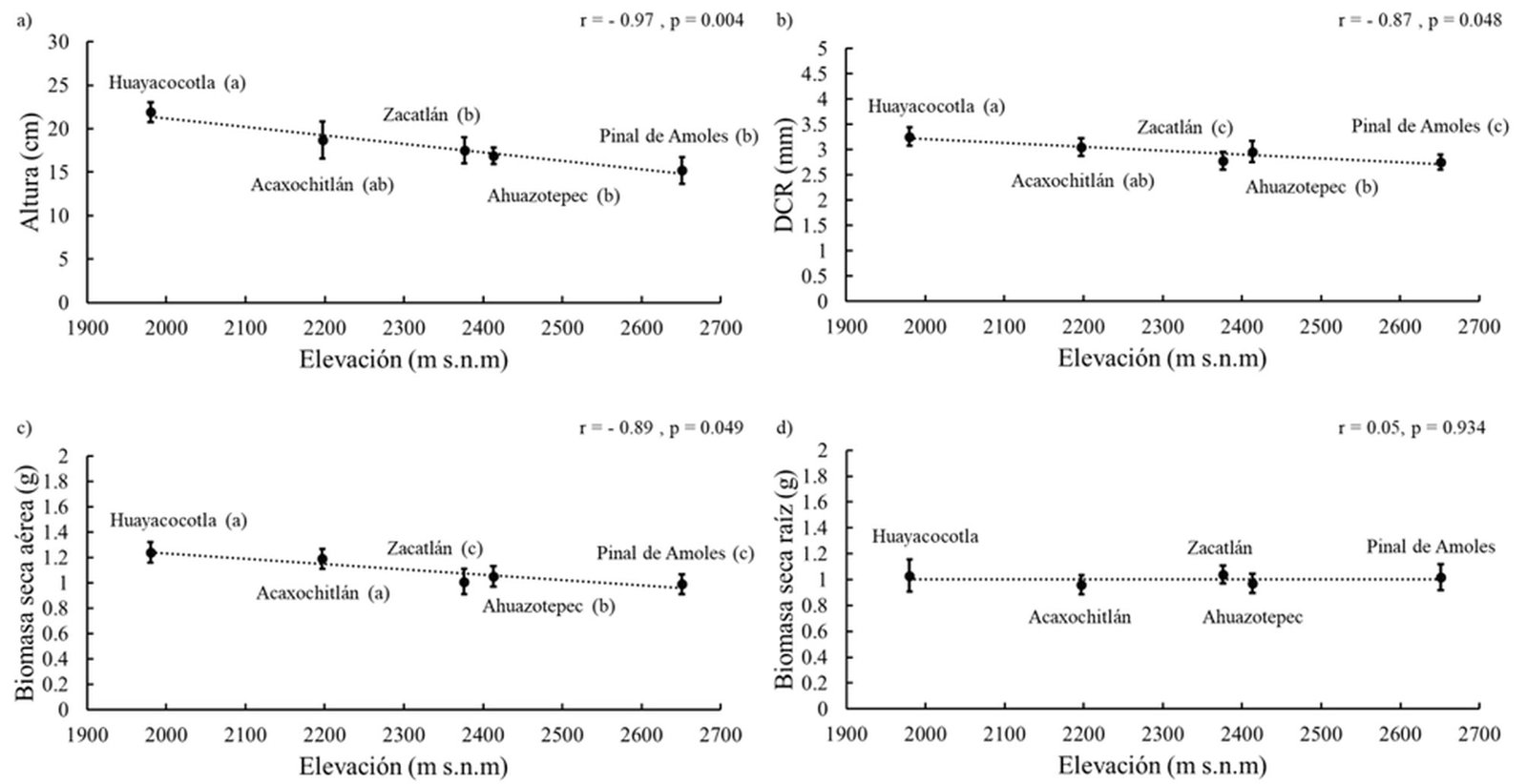
Figura 1. Regresión lineal entre las características morfológicas de Pinus patula y la elevación de las procedencias. a) Altura total; b) diámetro al cuello de la raíz (DCR); c) biomasa seca aérea; d) biomasa seca de raíz. Procedencias ordenadas de menor a mayor elevación. Barras de error representan el error estándar. Letras distintas para la misma variable indican diferencias significativas entre procedencias (Tukey, p ≤ 0.05).
El efecto más pronunciado del estrés hídrico ocurrió en las plantas de familias de Huayacocotla y Acaxochitlán, en las cuales el potencial hídrico disminuyó entre la primera y última evaluación en 1,260 y 1,022%, respectivamente (fig. 3a).
Por otro lado, a pesar de que en todas las procedencias se registraron valores considerados indicadores de estrés hídrico severo (Ѱ < -2.0) según Cleary et al. (2003), 2 familias de Pinal de Amoles (familias 6 y 14) y Huayacocotla (familias 77 y 98) y 1 de Zacatlán (familia 22) mostraron cierta estabilidad y resiliencia (fig. 3b-f). En estos casos las plantas lograron recuperar su potencial hídrico tras el proceso de rehidratación al que se sometieron al finalizar el periodo de déficit hídrico (10-16 días).
El estrés hídrico tuvo un efecto notable sobre la supervivencia, particularmente en familias originarias de sitios de precipitación superior a 1,300 mm anuales (Huayacocotla, tabla 1), donde la mortalidad alcanzó 70%. Esta variable se correlacionó fuertemente con la elevación del punto central de las procedencias (fig. 4a). La correlación entre la supervivencia y la precipitación media anual resultó fuerte, negativa y significativa (fig. 4b).
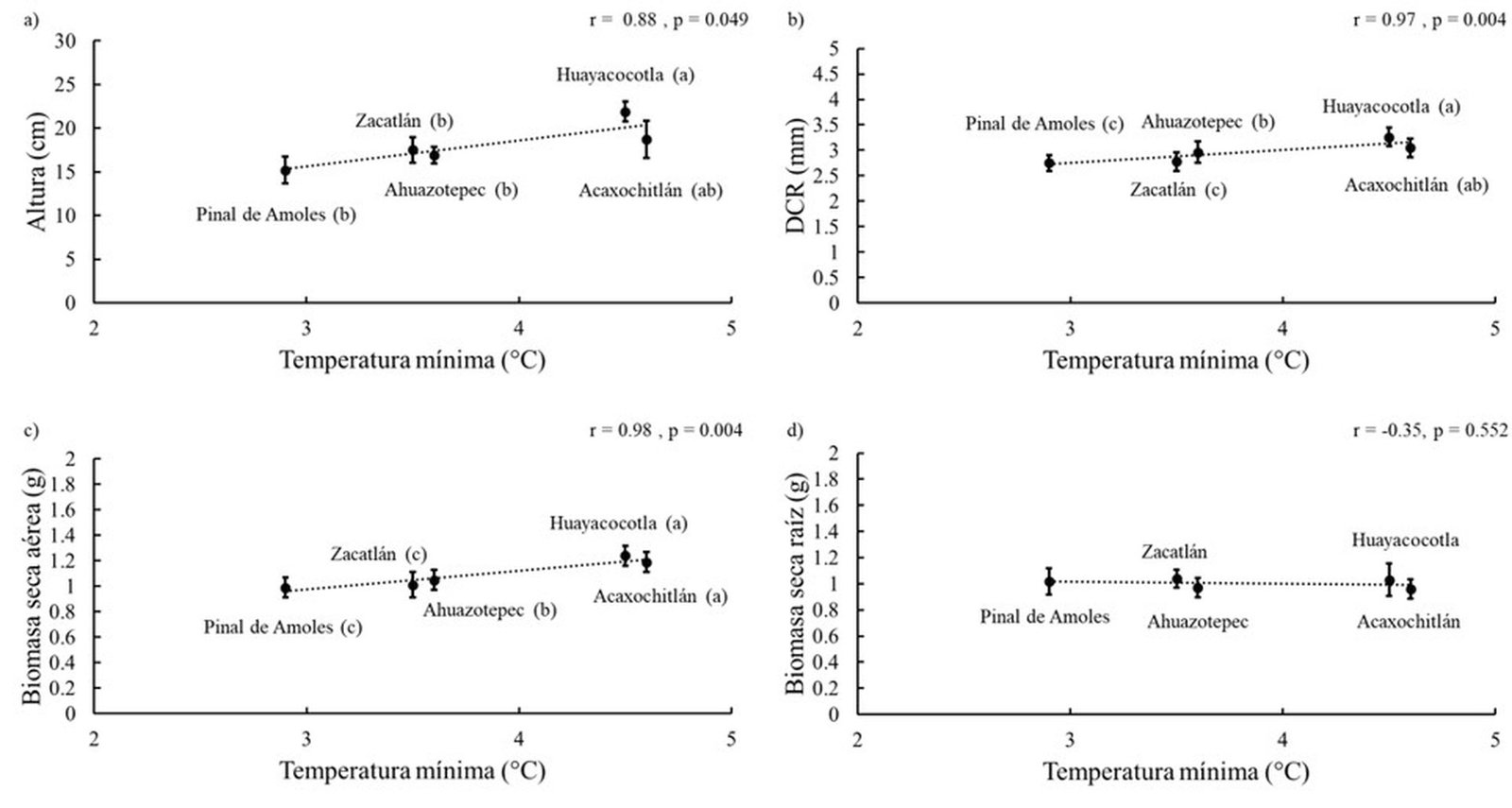
Figura 2. Regresión lineal entre las características morfológicas de Pinus patula y la temperatura mínima de las procedencias. a) Altura total; b) diámetro al cuello de la raíz (DCR); c) biomasa seca aérea; d) biomasa seca de raíz. Procedencias ordenadas de menor a mayor temperatura mínima. Barras de error representan el error estándar. Letras distintas para la misma variable indican diferencias significativas entre procedencias (Tukey, p ≤ 0.05).
La altura de las plantas mostró el mayor control genético. El diámetro al cuello de la raíz, la última medición del potencial hídrico (día 16) y la supervivencia mostraron control genético moderadamente alto (h2i ≥ 0.2). Sin embargo, para el resto de características el control genético fue moderado. La variación genética aditiva fue elevada para la última medición del potencial hídrico y la supervivencia (tabla 3).
Por lo general, las correlaciones genéticas fueron más altas que las correlaciones fenotípicas en todas las variables (tabla 4). A nivel genético, se determinaron correlaciones negativas moderadamente altas entre la altura de las plantas y la supervivencia. Por otra parte, aunque la correlación genética entre biomasa aérea y potencial hídrico fue alta en las 2 primeras mediciones, ésta se anuló bajo déficit hídrico severo (10 días). Asimismo, la correlación fenotípica entre la altura y el diámetro al cuello de la raíz fue fuerte y positiva. La biomasa seca aérea se correlacionó positiva y moderadamente con las variables de altura y diámetro al cuello de la raíz (tabla 4).
Las variables ambientales analizadas (elevación, temperatura mínima, temperatura media anual, precipitación media anual y el índice de sequía) mostraron una fuerte correlación con la mayoría de las variables morfológicas y fisiológicas de las plantas. Sin embargo, la biomasa seca de la raíz y las mediciones del potencial hídrico del día 16 no mostraron esta correlación (tabla 5). Los valores de potencial hídrico se correlacionaron fuerte y positivamente con la elevación del punto central de las procedencias (r ≥ 0.5); no obstante, se apreció una disminución de la correlación conforme disminuyó el potencial (aumento del estrés, tabla 5).
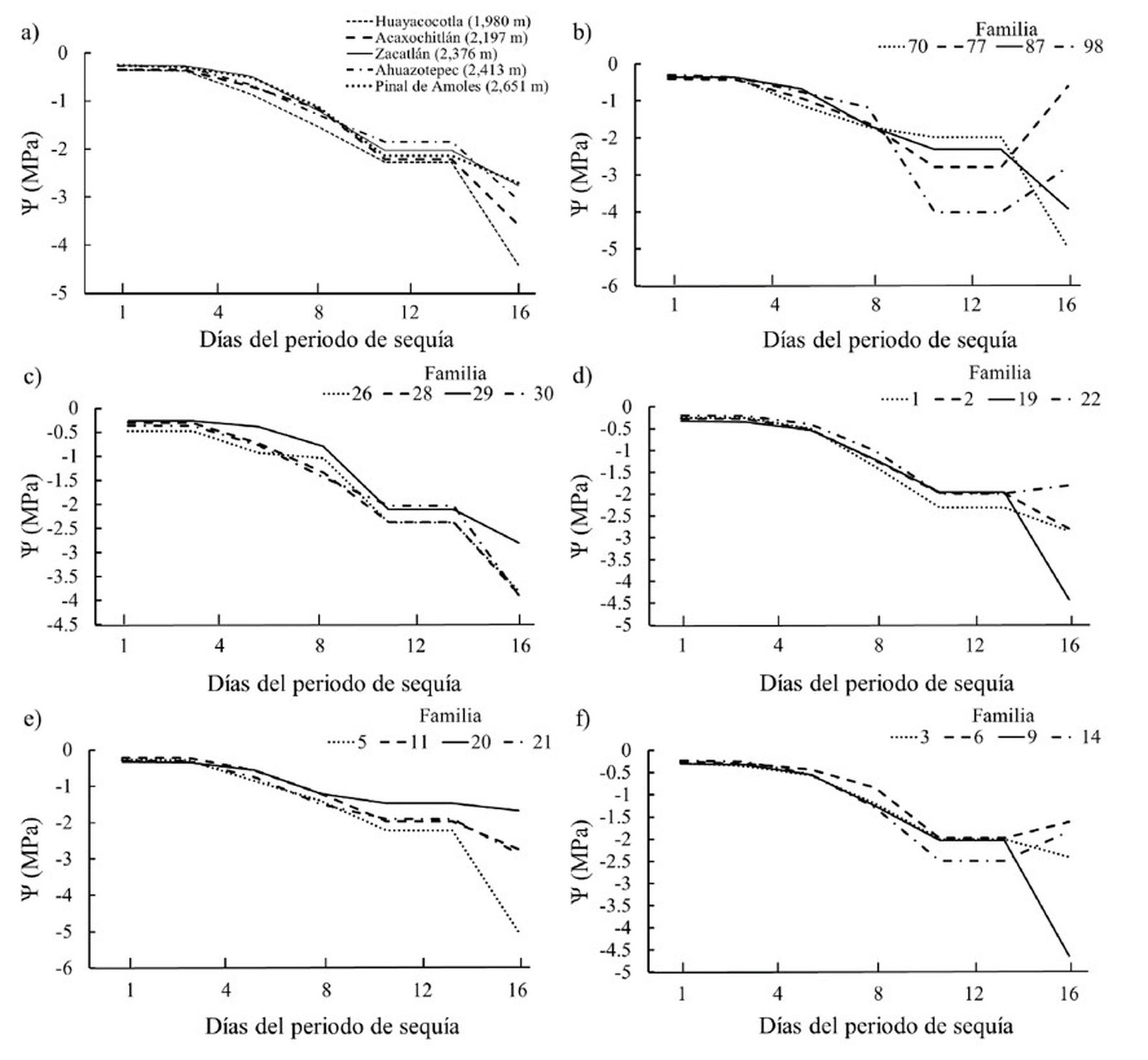
Figura 3. Valores promedio del potencial hídrico de las plantas de Pinus patula sometidas a estrés hídrico a nivel de procedencia y de familia dentro de procedencia. a) Procedencia; b) Huayacocotla; c) Acaxochitlán; d) Zacatlán; e) Ahuazotepec; f) Pinal de Amoles. Procedencias ordenadas de menor a mayor elevación.
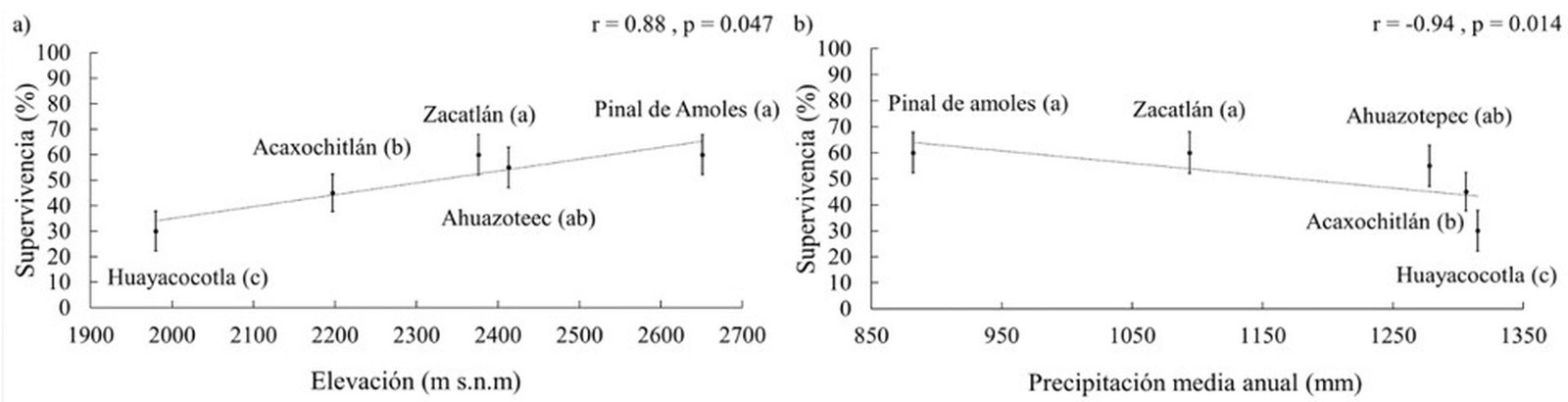
Figura 4. Regresión lineal entre la supervivencia de Pinus patula con la elevación y la precipitación media anual de las procedencias al final del periodo de sequía. a) Correlación supervivencia-elevación, procedencias ordenadas de menor a mayor elevación; b) correlación supervivencia-precipitación media anual, procedencias ordenadas de menor a mayor precipitación. Barras de error representan el error estándar. Letras diferentes indican diferencias significativas entre procedencias (Tukey, p < 0.05).
| Variable | σ2f | σ2e | h2i ± EE | CVGA (%) |
|---|---|---|---|---|
| Altura | 1.51 | 4.64 | 0.74 ± 0.35 | 11.80 |
| Diámetro al cuello de la raíz | 0.01 | 0.16 | 0.30 ± 0.20 | 7.80 |
| Biomasa seca parte aérea | 4.88×10-3 | 0.06 | 0.20 ± 0.12 | 11.10 |
| Biomasa seca de la raíz | 4.45×10-2 | 0.04 | 0.25 ± 0.19 | 11.03 |
| Biomasa seca total | 0.01 | 0.15 | 0.24 ± 0.19 | 9.56 |
| Ѱ3 | 1.72×10-3 | 0.02 | 0.23 ± 0.31 | 26.34 |
| Ѱ6 | 0.01 | 0.12 | 0.25 ± 0.28 | 28.83 |
| Ѱ8 | 0.02 | 0.24 | 0.26 ± 0.28 | 21.07 |
| Ѱ10 | 3.3×10-3 | 0.36 | 0.25 ± 0.19 | 14.92 |
| Ѱ16 | 0.74 | 5.68 | 0.35 ± 0.20 | 49.04 |
| Supervivencia | 0.79 | 3.29 | 0.58 ± 0.42 | 30.07 |
| VAR | ALT | DM | BSA | BSR | BST | Ѱ3 | Ѱ6 | Ѱ8 | Ѱ10 | Ѱ16 | SUP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALT | – | 0.75±0.15* | 0.67±0.11** | -0.02±0.17ns | 0.61±0.13** | -0.50±0.11* | -0.70±0.14* | -0.57±0.18* | -0.42±0.21ns | -0.15±0.30ns | -0.57±0.18* |
| DM | 0.47* | – | 0.80±0.08** | 0.10±0.17ns | 0.72±0.11** | -0.55±0.12** | -0.76±0.12** | -0.42±0.22ns | -0.15±0.24ns | -0.21±0.27ns | -0.59±0.13* |
| BSA | 0.59** | 0.72** | – | -0.12±0.27ns | 0.86±0.05*** | -0.74±0.07*** | -0.64±0.16** | -0.30±0.19ns | -0.04±0.23ns | -0.03±0.23ns | -0.43±0.19* |
| BSR | -0.01ns | 0.21ns | 0.10ns | – | 0.16±0.35ns | -0.20±0.14ns | -0.21±0.19ns | -0.16±0.14ns | -0.03±0.16ns | -0.32±0.23ns | -0.18±0.19ns |
| BST | 0.47* | 0.65** | 0.82*** | 0.51* | – | -0.51±0.14** | -0.49±0.18* | -0.30±0.20ns | -0.01±0.17ns | -0.11±0.21ns | -0.21±0.18ns |
| Ѱ3 | -0.44* | -0.53** | -0.75*** | -0.08ns | -0.54* | – | 0.65±0.10** | 0.30±0.19ns | 0.21±0.22ns | 0.18±0.21ns | -0.51±0.12** |
| Ѱ6 | -0.46* | -0.65** | -0.60** | -0.12ns | -0.46* | 0.71** | – | 0.66±0.15** | 0.30±0.17ns | 0.39±0.23ns | -0.69±0.10** |
| Ѱ8 | -0.38ns | -0.21ns | -0.22ns | -0.14ns | -0.14ns | 0.34ns | 0.57** | – | 0.32±0.15ns | 0.32±0.22ns | -0.61±0.11** |
| Ѱ10 | -0.39ns | -0.18ns | -0.08ns | -0.01ns | -0.08ns | 0.21ns | 0.24ns | 0.27ns | – | 0.05±0.29ns | -0.45±0.21ns |
| Ѱ16 | -0.24ns | -0.16ns | -0.04ns | -0.30ns | -0.02ns | 0.25ns | 0.36ns | 0.47* | 0.08ns | – | -0.55±0.34** |
| SUP | -0.50* | -0.49* | -0.42* | 0.07ns | -0.10ns | -0.55** | -0.64** | -0.61** | -0.47* | -0.68** | – |
| Variable respuesta | LAT | ELEV | TMÁX | TMÍN | TMA | PMA | DD5 | ÍS |
|---|---|---|---|---|---|---|---|---|
| Altura | -0.16ns | -0.91*** | 0.06ns | 0.77*** | 0.37ns | 0.36ns | 0.19ns | -0.37ns |
| DCR | -0.17ns | -0.56** | 0.16ns | 0.57** | 0.35ns | 0.50* | 0.14ns | -0.50* |
| BSA | -0.14ns | -0.70** | 0.12ns | 0.70** | 0.41* | 0.58** | 0.26ns | -0.56** |
| BSR | -0.08ns | 0.03ns | -0.09ns | -0.06ns | -0.02ns | -0.07ns | -0.01ns | 0.06ns |
| BST | -0.16ns | -0.53* | 0.09ns | 0.54** | 0.37ns | 0.42* | 0.26ns | -0.41* |
| Ѱ3 | 0.37ns | 0.57** | -0.24ns | -0.59** | -0.47* | -0.36ns | -0.14ns | 0.34ns |
| Ѱ6 | 0.11ns | 0.55* | -0.11ns | -0.50* | -0.30ns | -0.54* | -0.06ns | 0.52** |
| Ѱ8 | 0.02ns | 0.42* | -0.29ns | -0.42* | -0.37ns | -0.11ns | -0.19ns | 0.11ns |
| Ѱ10 | 0.30ns | 0.33ns | -0.60** | -0.48* | -0.68** | 0.35ns | -0.44* | 0.36ns |
| Ѱ16 | 0.16ns | 0.23ns | 0.03ns | -0.20ns | -0.09ns | -0.15ns | -0.03ns | 0.11ns |
| SUP | 0.01ns | 0.50* | -0.16ns | -0.43* | -0.30ns | -0.46* | -0.03ns | 0.46* |
Discusión
Las plantas mostraron patrones de crecimiento diferenciado de acuerdo con su procedencia; la altura, el diámetro al cuello de la raíz y la biomasa seca aérea fueron mayores conforme su origen fue de menor elevación y mayor temperatura mínima. Esta característica de crecimiento es una respuesta a la mayor disponibilidad de humedad que poseen los sitios de zonas menos elevadas. En contraste, las poblaciones de sitios más elevados, que a la vez presentan menor precipitación, suelen presentar crecimiento más lento para tolerar daños por heladas y sobrevivir bajo condiciones climáticas severas como el déficit hídrico (Bansal et al., 2016; Ishizuka et al., 2015).
Las variables de crecimiento (altura y diámetro) se registraron previo a la fase de estrés para establecer el vigor inicial de las progenies. Los resultados son consistentes con Pérez-Luna, Prieto-Ruíz et al. (2024) en cuanto a la jerarquía de crecimiento de las procedencias, donde las mismas familias de Huayacocotla y Acaxochitlán mostraron los mayores valores promedio en altura y diámetro. No obstante, el presente estudio profundiza en la estructura genética de esta variación. Determinar la heredabilidad en esta etapa permite identificar si las diferencias en el crecimiento inicial (que influyen en la resistencia mecánica y capacidad de almacenamiento de agua) tienen un fundamento genético que justifique la selección de progenies superiores.
Huayacocotla es una procedencia que se caracterizó por un crecimiento en altura superior, lo que la convierte en una opción de alto interés para el establecimiento de plantaciones forestales, que podría acortar el periodo de producción en vivero. Esta capacidad de crecimiento ha sido documentada de manera consistente con familias de P. patula de esa procedencia; por ejemplo, Fuentes-Amaro et al. (2021) reportaron alturas promedio de 27.9 cm a los 6 meses de edad, mientras que Rodríguez-Méndez (2013) registró valores de hasta 34.1 cm a los 9 meses. Sin embargo, este crecimiento acelerado implica mayores requerimientos hídricos, dado que una mayor biomasa aérea se asocia con incrementos en la superficie transpiratoria y, por ende, con una mayor demanda evaporativa, lo que puede ser particularmente complejo bajo condiciones de limitación de humedad.
Los valores observados en el diámetro al cuello de la raíz son similares a los registrados por Pérez-Luna, Prieto-Ruíz et al. (2024) de 3.27 mm en progenies de P. patula de Huayacocotla con 7 meses de edad, aunque en esa investigación las familias de Acaxochitlán presentaron diámetros al cuello de la raíz mayores (3.42 mm). No obstante, los diámetros de las familias de Huayacocotla fueron inferiores a los reportados por Fuentes-Amaro et al. (2021) (3.6 mm en plántulas de P. patula de 6 meses de edad). Cabe destacar que, en ninguna de las investigaciones previamente citadas, se reporta el cálculo de parámetros de control genético.
A diferencia de las variables de crecimiento aéreo, la biomasa seca de la raíz no presentó diferencias significativas entre procedencias. Si bien se ha documentado que familias originarias de ambientes áridos tienden a asignar una mayor biomasa al sistema radical como estrategia adaptativa (Acosta-Rico et al., 2024), en este estudio dicha respuesta no fue evidente. Esta homogeneidad sugiere que, bajo las condiciones de evaluación, la asignación en biomasa radicular fue similar entre progenies, independientemente de la precipitación o temperatura de sus sitios de origen.
En general, los resultados mostraron una biomasa aérea superior a la radical en todas las procedencias, patrón común en el desarrollo inicial de P. patula bajo condiciones de vivero (Martínez-Nevárez et al., 2023; Rodríguez-Méndez, 2013). Este desequilibrio entre la superficie transpiratoria y la capacidad de absorción radicular es un factor determinante en la respuesta al estrés hídrico severo; una mayor proporción de biomasa aérea incrementa la demanda hídrica, lo que podría explicar el rápido descenso de los potenciales hídricos observados en las familias de mayor tasa de crecimiento.
Las diferencias morfológicas entre procedencias confirman que el crecimiento de P. patula depende de su origen; lo anterior, en concordancia con estudios que han determinado mayor crecimiento en plantas de elevaciones inferiores a 2,500 m (Acosta-Rico et al., 2024; Pérez-Luna, Prieto-Ruíz et al., 2024; Rodríguez Méndez, 2013). Este patrón sugiere que las poblaciones de sitios bajos podrían presentar una mayor vulnerabilidad ante eventos de sequía extrema. Dado que estas procedencias son originarias de sitios con mayor precipitación media anual (PMA), es probable que carezcan de mecanismos de eficiencia hídrica como las poblaciones de zonas más secas. En un escenario de migración asistida hacia altitudes mayores para compensar el calentamiento global, estas familias enfrentarían un riesgo dual; el estrés hídrico derivado de su menor capacidad de regulación estomática y la susceptibilidad a daños por heladas, factores críticos que limitarían su éxito de establecimiento fuera de su rango altitudinal óptimo (Sánchez-Mendoza et al., 2023).
Sin embargo, en ambientes muy restrictivos la diferenciación puede atenuarse, como se observó en Pinus canariensis C.Sm. ex DC. en un sitio árido, donde el crecimiento no se asoció con el origen (Climent et al., 2002). En P. patula, la elevación se confirma como un buen predictor del desempeño de las poblaciones, el cual es útil en programas de plantaciones comerciales, como en estrategias de migración asistida y adaptación al cambio climático, al reafirmarse la tendencia de mayor crecimiento en las plantas de menor altitud (Sáenz-Romero et al., 2012).
Los valores de potencial hídrico registrados son comparables a los de otras especies sometidas a estrés hídrico, como Pinus cooperi C.E. Blanco con -0.26 a -3.24 MPa a los 12 días (Prieto-Ruíz et al., 2012) y Pinus engelmannii Carr. -1.22 MPa en promedio con riego cada 8 días (Ávila-Flores et al., 2014). Por otra parte, destaca que una familia de Huayacocotla (familia 77) presentó valores cercanos a 0 tras la rehidratación (fig. 3b), lo que sugiere plasticidad fisiológica y revela la importancia de seleccionar familias específicas en cada procedencia (Escobar-Alonso et al., 2024).
La supervivencia registrada (50%) muestra resultados similares a los documentados por Acosta-Rico et al. (2024), quienes reportaron una supervivencia de 60% en plantas de P. patula de 22 meses de edad con una de sequía de 4 meses en un ensayo de procedencias-progenies en jardín común. Se evidencia que ciertas familias de P. patula presentan baja tolerancia a la sequía. Asimismo, la resistencia parece incrementarse con el aumento del tamaño y edad, siempre que no exista restricción en el crecimiento radical. De hecho, las procedencias de ambientes húmedos mostraron la menor supervivencia. En nuestro caso, la supervivencia estuvo fuertemente influida por el origen del germoplasma; las procedencias de menor elevación registraron mayor mortalidad bajo condiciones de estrés. Este patrón coincide con lo observado en P. canariensis, especie en la cual las procedencias de ambientes húmedos mostraron menor supervivencia frente a la sequía que las de sitios más secos (Climent et al., 2002).
Estos hallazgos permiten caracterizar la capacidad de respuesta de las poblaciones ante condiciones críticas, lo cual es fundamental para el diseño de estrategias que incrementen la supervivencia y reduzcan costos de manejo en programas de plantación. La relevancia de evaluar el estrés hídrico en etapas tempranas radica en que permite identificar genotipos resilientes antes de su establecimiento. Esto es crucial, dado que en condiciones de campo la sequía puede ser el factor determinante de la supervivencia, tal como lo evidenciaron Pérez-Luna, López-Upton et al. (2024) en un ensayo con estas mismas familias en Durango, donde el estrés hídrico extremo explicó 97% de la mortalidad. Contar con estos indicadores fisiológicos en vivero ofrece una base sólida para establecer futuras correlaciones con el desempeño y crecimiento a edades más avanzadas en el sitio de plantación.
El control genético elevado de algunos caracteres a edades tempranas en vivero es común en especies de rápido crecimiento. Barnes y Schweppenhauser (1978) estimaron una heredabilidad individual elevada para altura (h2i = 0.56) en familias de P. patula un año después de la siembra. Resultados similares se han documentado en Pinus wallichiana A.B. Jacks (h2i = 0.93 con 12 meses de edad; Kaur et al., 2022), Pinus radiata D. Don (h2i = 0.33 con 4 meses; Espinoza et al., 2014) y Pinus oocarpa Schiede (h2i = 0.38 con 6 meses; Viveros-Viveros et al., 2005). Sin embargo, en P. patula la heredabilidad de la altura disminuye conforme avanza la edad en ensayos en campo; h2i = 0.23 al año de edad (Pérez-Luna, López-Upton et al., 2024), h2i = 0.21 a los 8 años (Escobar-Sandoval et al., 2018) y 0.16 a los 10 años (Bustillos-Aguirre et al., 2018). Este patrón de reducción también lo observaron los mismos autores en el diámetro.
En el presente estudio, la biomasa de la raíz y la biomasa total mostraron heredabilidades moderadas, similares a las reportadas por Espinoza et al. (2014) en plantas de 4 meses de edad de P. radiata (h2i = 0.28 y 0.36, respectivamente), y estos mismos autores encontraron un menor control genético en la biomasa aérea (h2i = 0.60). En contraste, el potencial hídrico mantuvo heredabilidades altas y constantes a lo largo de las evaluaciones, indicando diferenciación entre familias aún en condiciones restrictivas de sequía y en las condiciones restrictivas del bajo volumen del tubete.
Aunque los parámetros de control genético en vivero no son suficientes para una selección temprana efectiva, resultan útiles si existe correlación con rasgos a mayor edad (Codesido et al., 2012; Espinoza et al., 2014). Así que es importante que las condiciones de desarrollo en vivero sean semejantes a las de campo (Tan et al., 1995). Por otra parte, la supervivencia mostró control genético moderadamente alto y con el mayor coeficiente de variación genética aditiva (CVGA), similar a lo documentado por Acosta-Rico et al. (2024) en un ensayo de P. patula bajo sequía con un CVGA moderado (24.4%).
Se confirmó la asociación fenotípica positiva entre altura, diámetro al cuello de la raíz y biomasa, incluso en la etapa de crecimiento en vivero (Espinoza et al., 2014; Salaya-Domínguez et al., 2012). No obstante, las correlaciones fenotípicas mostraron que las familias con mayor biomasa resultaron más susceptibles al déficit hídrico, probablemente debido a la mayor pérdida de agua por transpiración en plantas de mayor tamaño, efecto acentuado por el volumen limitado de los tubetes pequeños, como los usados en esta investigación (170 mL); este efecto también fue corroborado en P. greggii var. australis (López-Upton y Muñoz-Orozco, 1991).
Asimismo, la correlación genética entre el potencial hídrico y la biomasa seca aérea fue moderadamente alta y negativa en las primeras 2 mediciones (BSA-Ѱ3, rg = -0.74; BSA-Ѱ6, rg = -0.55), lo que indica que ambos rasgos comparten parcialmente una base genética común, y por ser de magnitud moderada, la selección basada en mayor biomasa total produciría parcialmente plántulas más susceptibles al inicio de la sequía. Sin embargo, conforme aumentó la intensidad del déficit hídrico, la correlación genética se volvió nula (Ѱ10: rg = -0.03), esto sugiere que es posible seleccionar familias de mayor biomasa aérea sin comprometer la resistencia en condiciones extremas.
Por otro lado, la correlación genética negativa entre la supervivencia y la biomasa seca aérea (rg = -0.43) indica que los genotipos con mayor desarrollo de hojas y ramas tendieron a una menor supervivencia bajo déficit hídrico, probablemente por una mayor transpiración. De manera similar, la correlación negativa entre la supervivencia y el potencial hídrico sugiere que los genotipos con potenciales hídricos más negativos (capaces de mantener la función fisiológica aun perdiendo agua) tienden a mayor supervivencia. En conjunto, estos resultados sugieren una compensación funcional entre crecimiento aéreo y resistencia a la sequía. Este hallazgo indica que los genotipos con un desarrollo aéreo más restringido poseen una ventaja competitiva bajo estrés hídrico severo, asociada probablemente a una menor demanda transpiratoria. En este sentido, la identificación de familias que presentan estabilidad en su potencial hídrico y una mayor resiliencia es fundamental para el mejoramiento genético, ya que permite seleccionar progenies con mecanismos fisiológicos (como un control estomático eficiente) que aseguran la supervivencia en sitios críticos. Bajo esta perspectiva, el preacondicionamiento mediante ciclos de sequía y rehidratación en vivero adquiere una mayor relevancia, pues más allá de ser una técnica de endurecimiento tradicional (Prieto-Ruíz et al., 2004), funciona como un filtro selectivo que permite expresar y evaluar el potencial genético de resiliencia de las familias antes de su establecimiento en campo.
Al igual que en otros estudios (Acosta-Rico et al., 2024; Bansal et al., 2016; Rodríguez-Méndez et al., 2013), la altura y el diámetro al cuello de la raíz mostraron correlación negativa con la elevación y el índice de sequía, y asociación positiva con la temperatura, lo que indica menor crecimiento en familias de ambientes fríos y secos. Por el contrario, el potencial hídrico se relacionó fuerte y positivamente con la elevación y el índice de sequía, confirmando la mayor resistencia de las plántulas de Pinal de Amoles y Zacatlán, que registraron las tasas de supervivencia más altas. Este comportamiento refleja la plasticidad adaptativa de la especie a condiciones ambientales extremas y sugiere que las poblaciones de mayor altitud podrían requerir periodos más largos de manejo en vivero (Sáenz-Romero et al., 2016).
El origen del germoplasma, en particular la elevación y la temperatura mínima del sitio de procedencia, ejerce efecto sobre las características fenotípicas de P. patula en vivero, bajo un control genético de moderado a alto. Esta estructura genética se manifiesta de forma crítica en la respuesta fisiológica al estrés hídrico, donde las familias con mayor biomasa inicial mostraron una mayor vulnerabilidad al déficit, lo que evidencia un compromiso funcional entre crecimiento y resiliencia. En consecuencia, se concluye que el preacondicionamiento en vivero no debe realizarse de manera uniforme; por el contrario, la selección de familias y el manejo hídrico deben ajustarse según la procedencia. Este enfoque no solo optimiza la producción de planta para ambientes restrictivos, sino que fundamenta el uso de parámetros genéticos para predecir el éxito del establecimiento en programas de migración asistida ante el cambio climático.
Agradecimientos
A la Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti) por la beca otorgada al primer autor para realizar una estancia posdoctoral académica de continuidad (CVU 561686) bajo el marco de la Convocatoria 2022(1) de Estancias Posdoctorales Por México. Al proyecto Conafor-2017-2-291322 “Establecimiento de huertos semilleros asexuales regionales y ensayos de progenie de Pinus patula para la valoración genética de los progenitores” del Fondo Conacyt-Conafor.
Referencias
Acosta-Rico, R., López-Upton, J., Vargas-Hernández, J. J. y Cruz-Huerta, N. (2024). Variación genética en la respuesta de familias de Pinus patula a condiciones de sequía. Bosque (Valdivia), 45, 55–66. https://doi.org/10.4067/s0717-92002024000100055
Ávila-Flores, I. J., Prieto-Ruíz, J. Á., Hernández-Díaz, J. C., Whehenkel, C. A. y Corral-Rivas, J. J. (2014). Preacondicionamiento de Pinus engelmannii Carr. mediante déficit de riego en vivero. Revista Chapingo. Serie Ciencias Forestales y del Ambiente, 20, 237–245. https://doi.org/10.5154/r.rchscfa.2014.02.004
Bansal, S., Harrington, C. A. y St. Clair, J. B. (2016). Tolerance to multiple climate stressors: a case study of Douglas-fir drought and cold hardiness. Ecology and Evolution, 6, 2074–2083. https://doi.org/10.1002/ece3.2007
Barnes, R. D. y Schweppenhauser, M. A. (1978). Pinus patula Schiede and Deppe progeny tests in Rhodesia genetic control of nursery traits. Silvae Genetica, 27, 200–204.
Bustillos-Aguirre, C. V., Vargas-Hernández, J. J., López- Upton, J. y Ramírez-Valverde, G. (2018). Repetibilidad de parámetros genéticos de las características de ramificación en progenies de Pinus patula. Madera y Bosques, 24, e2411131. https://doi.org/10.21829/myb.2018.2411131
Cleary, B., Zaerr, J. y Hamel, J. (2003). Guidelines for measuring plant moisture stress with a pressure chamber. Albany, OR: PMS Instrument Co.
Climent, J., Gil, L., Pérez, E. y Pardos, J. A. (2002). Efecto de la procedencia en la supervivencia de plántulas de Pinus canariensis Sm. en medio árido. Investigación Agraria: Sistemas y Recursos Forestales, 11, 171–180. https://doi.org/10.5424/766
Codesido, V., Zas, R. y Fernández-López, J. (2012). Juvenile– mature genetic correlations in Pinus radiata D. Don. under different nutrient × water regimes in Spain. European Journal of Forest Research, 131, 297–305. https://doi.org/10.1007/s10342-011-0501-9
Dickerson, G. E. (1969). Techniques for research in quantitative animal genetics. En R. R. Johnson (Ed.), Techniques and procedures in animal science research (pp. 36–79). Madison, Wisconsin: American Society of Animal Science.
Escobar-Alonso, S., Vargas-Hernández, J. J., López-Upton, J., García-Campusano, F., Jiménez-Casas, M. y Cruz-Huerta, N. (2024). Genetic variation and phenotypic plasticity in the seasonal shoot growth pattern of Pinus pseudostrobus. New Forests, 55, 1379–1398. https://doi.org/10.1007/s11056-024-10040-2
Escobar-Sandoval, M. C., Vargas-Hernández, J. J., López-Upton, J., Espinosa-Zaragoza, S. y Borja-de la Rosa, A. (2018). Parámetros genéticos de calidad de madera, crecimiento y ramificación en Pinus patula. Madera y Bosques, 24, e2421595. https://doi.org/10.21829/myb.2018.2421595
Espinoza, S. E., Magni, C. R., Santelices, R. E., Ivković, M. y Cabrera, A. M. (2016). Changes in drought tolerance of Pinus radiata in Chile associated with provenance and breeding generation. Annals of Forest Science, 73, 267–275. https://doi.org/10.1007/s13595-015-0498-1
Espinoza, S. E., Martínez, V. A., Magni, C. R., Ivković, M., Santelices, R. E., Guerra, F. P. et al. (2014). Genetic control of growth, biomass allocation, and survival under drought stress in Pinus radiata D. Don seedlings. Tree Genetics & Genomes, 10, 1045–1054. https://doi.org/10.1007/s11295-014-0741-1
Falconer, D. S. y Mackay, T. F. (2006). Introducción a la genética cuantitativa. Zaragoza, España: Editorial Acribia.
Flores, A. y Moctezuma-López, G. (2021). Cosecha de madera de 20 coníferas en zonas de movimiento de germoplasma. Revista Mexicana de Ciencias Forestales, 12, 122–140. https://doi.org/10.29298/rmcf.v12i66.829
Fuentes-Amaro, S. L., Rodríguez-Laguna, R., Razo-Zárate, R., Meza-Rangel, J. y Jiménez-Casas, M. (2021). Variación altitudinal en emergencia y crecimiento inicial de plantas de Pinus patula. Revista Fitotecnia Mexicana, 44, 655–660. https://doi.org/10.35196/rfm.2021.4.655
Ishizuka, W., Ono, K., Hara, T. y Goto, S. (2015). Influence of low- and high-elevation plant genomes on the regulation of autumn cold acclimation in Abies sachalinensis. Frontiers in Plant Science, 6, 890. https://doi.org/10.3389/fpls.2015.00890
Kaur, A., Monga, R., Bhardwaj, D. R. y Sharma, J. (2022). Estimation of genetic parameters of Pinus wallichiana seedlings in the nursery. International Journal of Bio- resource and Stress Management, 13, 578–585. https://doi.org/10.23910/1.2022.2609
Leibing, C., Signer, J., Van Zonneveld, M., Jarvis, A. y Dvorak, W. (2013). Selection of provenances to adapt tropical pine forestry to climate change on the basis of climate analogs. Forests, 4, 155–178. https://doi.org/10.3390/f4010155
López-Upton, J., Donahue, J. K., Plascencia-Escalante, F. O. y Ramírez-Herrera, C. (2005). Provenance variation in growth characters of four subtropical pine species planted in Mexico. New Forests, 29, 1–13. https://doi.org/10.1007/s11056-004-8018-4
López-Upton, J. y Muñoz-Orozco, A. (l99l). Selección familial por tolerancia a sequía en Pinus greggii Engelm. I: evaluación en plántula. Agrociencia, Serie Fitociencia, 2, 111–123.
Martínez-Nevárez, L. E., Prieto-Ruíz, J. Á., Sigala-Rodríguez, J. A., García-Rodríguez, J. L., Martínez-Reyes, M., Carrillo- Parra, A. et al. (2023). Crecimiento y eficiencia en el uso de nutrientes de plantas de Pinus cooperi C. E. Blanco producidas en vivero con un fertilizante de liberación controlada. Terra Latinoamericana, 41, e1707. https://doi.org/10.28940/terra.v41i0.1707
Martiñón-Martínez, R. J., Vargas-Hernández, J. J., López- Upton J., Gómez-Guerrero, A. y Vaquera-Huerta, H. (2010). Respuesta de Pinus pinceana Gordon a estrés por sequía y altas temperaturas. Revista Fitotecnia Mexicana, 33, 239– 248. https://doi.org/10.35196/rfm.2010.3.239
Mejía-Bojórquez, J. M., García Rodríguez, J. L. y Muñoz Flores, H. J. (2015). Evaluación de plantaciones de cuatro especies forestales en el estado de Durango. Reaxion, 2, 8–28.
Morales-González, E., López-Upton, J., Vargas-Hernández, J. J., Ramírez-Herrera, C. y Gil-Muñoz, A. (2013). Parámetros genéticos de Pinus patula en un ensayo de progenies establecido en dos altitudes. Revista Fitotecnia Mexicana, 36, 155–162. https://doi.org/10.35196/rfm.2013.2.155
Pérez-Luna, A., López-Upton, J., Prieto-Ruíz, J. Á. y Madrid- Aispuro, R. E. (2024). Supervivencia y control genético sobre características fenotípicas de un ensayo de progenies de Pinus patula. Revista Fitotecnia Mexicana, 47, 301–309. https://doi.org/10.35196/rfm.2024.3.301
Pérez-Luna, A., Prieto-Ruíz, J. Á., López-Upton, J., Madrid- Aispuro, R. E. y Salcido-Ruíz, S. (2024). Ensayos de germinación y producción de planta de procedencias- progenies de Pinus patula. Bosque (Valdivia), 45, 257–269. https://doi.org/10.4067/S0717-92002024000200257
Prieto-Ruíz, J. Á., Aldrete, A., Hernández-Díaz, J. C. y Goche-Télles, J. R. (2016). Causas de mortalidad de las reforestaciones y propuestas de mejora. En J. Á. Prieto- Ruíz y J. R. Goche-Télles (Eds.), Las reforestaciones en México. Problemática y alternativas de solución (pp. 14– 19). Durango: UJED Editorial.
Prieto-Ruíz, J. Á., Almaraz-Roldán, R. J., Corral-Rivas, J. J. y Díaz-Vázquez, A. (2012). Efecto del estrés hídrico en Pinus cooperi Blanco durante su preacondicionamiento en vivero. Revista Mexicana de Ciencias Forestales, 3, 19–28.
Prieto-Ruíz, J. Á., Cornejo-Oviedo, E. H., Domínguez-Calleros, P. A., Návar-Chaidez, J. D. J., Marmolejo-Monsiváis, J. G. y Jiménez-Pérez, J. (2004). Estrés hídrico en Pinus engelmannii Carr., producido en vivero. Investigación Agraria: Sistemas y Recursos Forestales, 13, 443–451.
Rodríguez-Méndez, C. (2013). Efecto de la fecha de siembra y tamaño de contenedor en el crecimiento de dos especies de pino en vivero (Tesis de maestría en Ciencias). Postgrado Forestal, Colegio de Postgraduados. Montecillo, Texcoco, México.
Sáenz-Romero, C., Lindig-Cisneros, R. A., Joyce, D. G., Beaulieu, J., St. Clair, J. B. y Jaquish, B. C. (2016). Migración asistida de las poblaciones forestales para la adaptación de árboles ante el cambio climático. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 22, 303–323. https://doi.org/10.5154/r.rchscfa.2014.10.052
Sáenz-Romero, C., Martínez-Palacios, A., Gómez-Sierra, J. M., Pérez-Nasser, N. y Sánchez-Vargas, N. M. (2012). Estimación de la disociación de Agave cupreata a su hábitat idóneo debido al cambio climático. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 18, 291–301. https://doi.org/10.5154/r.rchscfa.2011.11.078
Salaya-Domínguez, J. M., López-Upton, J. y Vargas-Hernández, J. J. (2012). Variación genética y ambiental en dos ensayos de progenies de Pinus patula. Agrociencia, 46, 519–534.
Salcido-Ruíz, S., Prieto-Ruíz, J. Á., Santana-Aispuro, E., Chávez-Simental, J. A. y Madrid-Aispuro, R. E. (2021). Supervivencia y crecimiento de Pinus engelmannii Carr. en una reforestación por micorrización y fertilización. Revista Mexicana de Ciencias Forestales, 12, 45–65. https://doi.org/10.29298/rmcf.v12i64.847
Sánchez-Mendoza, J. L., Jiménez-Casas, M., Ramírez-Herrera, C. y Viveros-Viveros, H. (2023). Calidad de semilla y crecimiento de planta en poblaciones y altitudes de Pinus hartwegii Lindl. Revista Mexicana de Ciencias Forestales, 14, 143–165. https://doi.org/10.29298/rmcf.v14i75.1297
Searle, S. R. (1995). An overview of variance component estimation. Metrika, 42, 215–230. https://doi.org/10.1007/BF01894301
Sorensen, F. C. y White, T. L. (1988). Effect of natural inbreeding on variance structure in tests of wind-pollination Douglas- fir progenies. Forest Science, 34, 102–118. https://doi.org/10.1093/forestscience/34.1.102
Tan, W., Blake, T. J. y Boyle, T. J. (1995). Early selection for drought tolerance and relationship to dry weight partitioning in black spruce families. Forest Science, 41, 168–180. https://doi.org/10.1093/forestscience/41.1.168
Viveros-Viveros, H., Sáenz-Romero, C. y Guzmán-Reyna, R. R. (2005). Control genético de características de crecimiento en vivero de plántulas de Pinus oocarpa. Revista Fitotecnia Mexicana, 28, 333–338. https://doi.org/10.35196/rfm.2005.4.333
Wang, T., Hamann, A., Spittlehouse, D. y Carroll, C. (2016). Locally downscaled and spatially customizable climate data for historical and future periods for North America. Plos One, 11, e0156720. https://doi.org/10.1371/journal.pone.0156720
White, T. L., Adams, W. T. y Neale, D. B. (2007). Forest genetics. Oxford: CABI Publishing.


