Fausto Roberto Méndez-de la Cruz a, *, Rodrigo Macip-Ríos b y Gabriel Barrios-Quiroz c
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Apartado Postal 70-153, 04510 Ciudad de México, México
b Universidad Nacional Autónoma de México, Escuela Nacional de Estudios Superiores, Unidad Morelia, Antigua Carretera a Pátzcuaro Núm. 8701, Col. Ex Hacienda San José La Huerta, 58190 Morelia, Michoacán, México
c Ecología y Conservación Dharma A.C., Carretera Federal Tepoztlán-Yautepec Km 7.5, 62520 Tepoztlán, Morelos, México
*Autor para correspondencia: faustomendez6@gmail.com (F.R. Méndez-de la Cruz)
Recibido: 11 octubre 2024; aceptado: 8 noviembre 2024
“De niño acompañaba a mi mamá, que daba clases en una escuela de Xochimilco y me gustaba ir al patio de la escuela que estaba junto a un bosque. Ese bosque estaba lleno de muchos animales, entre ellos chapulines, mariposas, plantas diversas y árboles enormes, me encantaba perderme en ese bosque lleno de vida”. Así comenzaba la admiración por la vida de un pionero de la herpetología en México, el Dr. Gustavo Casas Andreu.
El Dr. Casas-Andreu (Gus, como le llamábamos sus amigos más cercanos) fue una figura destacada en el campo de la herpetología, con una carrera que abarcó más de 6 décadas. Nacido en la Ciudad de México el 15 de mayo (día del maestro) de 1943, realizó sus estudios de educación básica y media superior en la Escuela Militarizada México. Desde temprana edad mostró una pasión por los reptiles y anfibios, lo que lo llevó a estudiar Biología en la Facultad de Ciencias de la Universidad Nacional Autónoma de México, donde sería alumno de ilustres personajes de la biología en México como el Maestro Rafael Martín del Campo y Sánchez y el Dr. Eucario López Ochoterena. Su interés por los anfibios y reptiles, particularmente por los cocodrilos, un grupo de reptiles que lo cautivó por su complejidad y su importancia ecológica, lo motivaron para realizar investigaciones pioneras que ampliaron nuestro entendimiento sobre su comportamiento, ecología y conservación.
A lo largo de su vida, Gustavo Casas comenzó su acercamiento con los reptiles trabajando como Investigador y jefe de la sección de herpetología en estudios con anfibios y reptiles acuáticos, del entonces Instituto Nacional de Investigaciones Biológico Pesqueras, perteneciente, en su momento, a la Secretaría de Industria y Comercio (1964-1971). Su tesis de licenciatura la desarrolló con las tortugas dulceacuícolas de México bajo la asesoría del Maestro Rafael Martín del Campo y Sánchez, misma que defendió en 1967. Siguiendo con su interés en la investigación en los reptiles, junto con Manuel Guzmán hicieron una recopilación del estatus de los cocodrilos de México, la cual se publicó como un número del Boletín del Instituto Nacional del Instituto Biológico Pesquero en 1970. Obtuvo la Maestría en Ciencias por la Facultad de Ciencias en el año 1973, por examen general de conocimientos.
Gustavo Casas ingresó como investigador al Instituto de Biología en 1974. Aquí empezó el trabajo curatorial de la colección de anfibios y reptiles. Posteriormente, realizó su tesis doctoral con el tema “Anfibios y reptiles de la costa suroeste del estado de Jalisco con aspectos de ecología y biogeografía”, la cual defendió en 1982, bajo la asesoría del Dr. Ismael Ferrusquía Villafranca. Como parte de su labor como curador de la Colección de Anfibios y Reptiles, en 1982 comienza un proyecto para representar la herpetofauna del país en lo que sería la Colección Nacional de Anfibios y Reptiles, y encabeza un grupo que colecta por todo el país. En esos momentos, la colección tenía representado menos de 5.6% de la herpetofauna del país. El proyecto que inició Gustavo Casas terminó en 1988, dando como resultado una representación aproximada de 60% de la herpetofauna de México en la colección del Instituto de Biología, convirtiéndose en una de las colecciones más importantes de México y del mundo.
Además de su trabajo como curador y biólogo de campo, Gustavo Casas fue un profesor dedicado y un mentor generoso en varias universidades de México. Su entusiasmo por la ciencia y su habilidad para transmitir conocimientos inspiraron a numerosas generaciones de herpetólogos y biólogos de la Universidad Nacional Autónoma de México, Universidad Autónoma Metro- politana y Universidad Autónoma del Estado de México. Sus clases y conferencias eran conocidas por su claridad, profundidad y el fervor con que abordaba cada tema, dejando una marca duradera en aquellos que tuvimos el privilegio de aprender de él. Dirigió 34 de tesis de licenciatura, 7 de maestría y 4 de doctorado. Entre los alumnos que desarrollaron tesis en diferentes niveles se encuentran: Oscar Flores Villela, Miriam Benabib Nisembaum, Aurelio Ramírez Bautista, Héctor Gadsden Esparza, Fausto R. Méndez de la Cruz, Elizabeth Arévalo Marín, Dolores Huacuz Elías, Rodrigo Macip Ríos, Anahí Guizado Rodríguez y Gabriel Barrios Quiroz, entre muchos otros, dejando un legado académico que ha trascendido y sigue dando frutos en generaciones presentes de herpetólogos.
Publicó 115 artículos de investigación, 40 de divulgación científica y 9 libros, que abarcan diversos temas, desde claves de identificación, herpetofaunas regionales, historia de la herpetología, etnoherpetología, biología de la reproducción y conservación de anfibios y reptiles. Entre los libros publicados se encuentran “Estado de las investigaciones sobre cocodrilos mexicanos”, “Anfibios y Reptiles de México”, “Nombres estándar en español, en inglés y nombres científicos de los anfibios y reptiles de México”, “Cómo hacer una colección de anfibios y reptiles”, “Sinopsis de datos biológicos y ecológicos del cocodrilo de pantano Crocodylus moreletii”, y otros. Su obra puede consultarse en el siguiente enlace: https://scholar.google.com/citations?user=jGqOlAgAAAAJ&hl=es
Gustavo Casas también fue un defensor incansable de la protección del hábitat natural de los cocodrilos y trabajó activamente con organizaciones nacionales e internacionales para promover políticas de conservación eficaces. Fue parte del Grupo de Especialistas de Cocodrilos de la IUCN, de grupos de asesores de la Conabio, además fue miembro fundador de varias sociedades científicas en México, como la Sociedad Mexicana de Zoología (SMZ), donde fue el primer presidente, de la Sociedad de Historia Natural, la Sociedad Herpetológica Mexicana (SHM), la Asociación para la Investigación y Conservación de Anfibios y Reptiles (AICAR), y más recientemente, del Colegio de Biólogos. Recibió homenajes (en vida y posmortem) de la SMZ, de la Sociedad Latinoamericana de Herpetología (en vida), de la SHM (en vida y se designó como miembro honorario) y de AICAR (en vida y posmortem, además de ser designado como miembro honorario).
Gustavo Casas fue un agradable conversador, cuando los temas que se tocaban eran diversos, desde cuestiones académicas hasta humanas y cotidianas, siempre con la oportunidad de aprender y reflexionar. Imposible olvidar las gratas charlas de sobremesa que se prolongaban hasta muy noche. Es importante mencionar que el Congreso de Herpetología de 1987, en donde participó como organizador local y reunió a las 2 sociedades de herpetólogos estado- unidenses más importantes (Herpetologists’ League y la Society for the Study of Amphibians and Reptiles) y al Comité Herpetológico Mexicano (precursor de la Sociedad Herpetológica Mexicana), sigue siendo recordada por la gran camaradería que se vivió en el Hotel Mocambo, sede del congreso. Después de una larga vida académica productiva, se jubiló el 31 de octubre de 2017 de la UNAM, que fue su alma mater e institución académica principal.
Gustavo Casas, el profesor, académico, mentor y amigo, falleció el 1 de septiembre de 2024 a los 81 años de edad. Su memoria y legado viven en el impacto duradero que ha tenido en el estudio de los anfibios y reptiles de México, así como en el corazón de todos aquellos que tuvimos el privilegio de conocerlo y que hoy continuamos con la eterna tradición de generar nuevo conocimiento, difundirlo y enseñar lo aprendido.
Le sobreviven su esposa Xóchitl y sus hijos, Gustavo, Adrián, Gustavo Antonio y Sara Sofia, además de una legión de colegas, estudiantes y amigos que lo apreciamos profundamente.
Meghan I. Zolá-Rodríguez a, Mariana Cuautle b, *, Marco Daniel Rodríguez-Flores c, Citlalli Castillo-Guevara b
a Akumal Monkey Sanctuary & Rescued Animals, Camino a Uxuxubi s/n Predio Santa Pilar Lote 16, 77776 Akumal, Quintana Roo, Mexico
b Universidad Autónoma de Tlaxcala, Centro de Investigación en Ciencias Biológicas, Km 10.5 Carretera Tlaxcala-San Martín Texmelucan, 90120 San Felipe Ixtacuixtla, Tlaxcala, Mexico
c Blue Marlin Conservation (Conservation Diver), Sunset Beach Gili Air, Gili Indah, Pemenang, North Lombok Regency, West Nusa Tenggara 83355, Indonesia
This study examines the impact of disturbance on the food preferences and dominance of an ant community in a temperate ecosystem in Mexico. The study focused on 2 types of vegetation: native oak forest and induced grassland (disturbed vegetation). Observations were conducted to record the food elements carried by ants to their nests. These data were analyzed using x2 tests. Tuna and honey baits were placed near the nests to record the presence of ants in 5-minute periods. We used a binomial model to determine whether the probability of finding an ant foraging at the baits was affected by vegetation type, bait type, and/or ant species. Additional baits were used to determine the ant dominance indices. T-tests and ANOVAs were used to compare dominance indices between vegetation types, baits, and ant species. No significant differences were observed in food preferences between vegetations. However, some species showed a preference for honey (i.e., carbohydrates), which could be limited in ground-level environments. Ants showed a submissive behavior in both vegetation types. This research shows that ants could optimize their nutrient intake, enabling them to survive efficiently even when facing disturbances, instead of increasing dominance.
Impacto del disturbio en las preferencias alimentarias y dominancia de las hormigas (Hymenoptera: Formicidae) en un bosque templado de México
Resumen
Este estudio examina el impacto del disturbio en las preferencias alimentarias y dominancia de una comunidad de hormigas en un ecosistema templado en México, en bosque de encino nativo y pastizal inducido (vegetación perturbada). Se registraron los alimentos transportados por las hormigas a sus nidos. Estos datos fueron analizados utilizando pruebas de x2. Se colocaron cebos de atún y miel cerca de los nidos para registrar la presencia de hormigas. Utilizamos un modelo binomial para determinar si la probabilidad de encontrar una hormiga en los cebos se veía afectada por la vegetación, cebo o especie de hormiga. Los índices de dominancia se determinaron usando cebos. Se emplearon pruebas t y Anova para comparar los índices de dominancia entre tipos de vegetación, cebos y especies de hormigas. No hubo diferencias significativas en las preferencias alimentarias entre tipos de vegetación, pero algunas especies mostraron una preferencia por la miel (carbohidratos), que podría ser un recurso limitado a nivel del suelo. Las hormigas mostraron un comportamiento sumiso en ambos tipos de vegetación. Esta investigación muestra que las hormigas podrían optimizar su ingesta de nutrientes, permitiéndoles sobrevivir bajo condiciones de disturbio, en lugar de aumentar su dominancia.
Palabras clave:Nidos de hormigas; Índice de dominancia; Hábitos alimenticios; Hipótesis de compensación; Carbohidratos; Proteínas
Introduction
Land use change stands as one of the primary factors contributing to global change and its adverse effects on biodiversity (Ellis et al., 2010; Foley et al., 2005; Sala et al., 2000). Land use change has reduced biodiversity through the loss, modification, and fragmentation of habitats; degradation of soil and water; and overexploitation of native species (Foley et al., 2005), and its effects depend strongly on the type, severity, frequency and timing of disturbance (Foley et al., 2005; White & Jentsch, 2001). Ants are one of the most dominant insects both ecologically and numerically (Rico-Gray & Oliveira, 2007; Schultheiss et al., 2022; Toro et al., 2012) that participate in different ecological processes —e.g., nutrient recycling, soil formation, decomposition, seed dispersion (Toro et al., 2012). Ants are model organisms to study the effect of disturbance because they respond to environmental change (Agosti et al., 2000; Andersen, 2000). It is often observed that disturbances favor behaviorally dominant ants, including invasive species. Vonshak and Gordon (2015), observed that native ant richness was highest in natural habitats, and alien species richness was highest in urban habitats, along an urban-rural gradient in the San Francisco Bay area. Nevertheless, the specific effect will depend on factors like the type of disturbance, including fires, floodings, or even treefall gaps (Cerdá et al., 2013). Hoffman and Andersen (2003) studied the response of ant functional groups to disturbance finding that the dominant Dolichoderinae and “hot climate specialists” tend to be favoured by low levels of disturbance. The opportunist and generalized Myrmicinae have wide habitat tolerances but are sensitive to competitive interactions, while “cryptic species” and “specialist predators” are highly sensitive to disturbance.
Feeding habits and foraging strategies represent vital life history traits among ants, playing a significant role in their ecological dynamics. These traits are contingent upon the availability of resources within their respective habitats (Andersen, 2000; Davidson, 2005), and the ensuing competition for those resources (Arnan et al., 2012). The feeding habits of ants play a pivotal role in determining their ecological function within the ecosystem (Spotti et al., 2015). However, despite their significance, the food habits and preferences of ants remain relatively understudied and represent one of the least understood aspects of their biology (Houdria et al., 2015).
The effects of disturbance (e.g., land use change) on ant feeding habits (especially foraging strategies) have not been studied in depth (Castillo-Guevara et al., 2019; Hernández-Flores et al., 2016; Radnan et al., 2018), and the way ants respond to habitat complexity can provide clues as to how ants could respond to disturbance. When the external disturbing factors are of high intensity (natural-fires or anthropogenic-deforestation), the disturbances can initiate a directional regression manifested as gradual or rapid simplification of the horizontal and vertical structure of a community, leading to the replacement of complex communities by a simpler one (Łaska, 2001). The level of habitat complexity can influence ant foraging patterns. For instance, higher complexity may decrease interspecific interactions and ant recruitment or minimize the trade-off between resource discovery and dominance (Parr & Gibb, 2012).
Examining the effect of soil surface complexity on food exploitation in the context of change from grassland to shrubland in Australia, Radnan et al. (2018) discovered that substrate complexity (wood debris, leaf litter, or no substrate) influenced the discovery time, ant size, and monopolization index of tuna and honey baits within testing arenas. However, at a larger scale of vegetation type, the effect was not observed. Similarly, Castillo-Guevara et al. (2019) found that the dominance level was similar in natural oak vegetation in comparison to an agricultural area in a temperate community; however, ant foraging strategies differed between the 2 communities.
Competition has been regarded as the primary factor influencing the structure of ant communities (Cerdá et al., 2013). Through the use of aggressive strategies, ants exert influence on the spatial distribution, abundance, and behavior of other ants. These strategies encompass a range of tactics, including the deployment of repellent chemicals and the establishment of territories (Cerdá et al., 2013). Moreover, ants can establish interspecific dominance hierarchies primarily based on variations in food collection behavior and aggressiveness. While there are various proposals for categorizing such hierarchies, the classification proposed by Vepsäläinen and Pisarski (1982) and Savolainen et al. (1989), as described by Cerdá et al. (2013), is considered the most well-defined hierarchical system from an ecological perspective. This classification system is founded on the aggressive behavior of ants and its impact on other ant species, and includes 3 categories: the dominant, subdominant and subordinate category. Dominant species exhibit highly aggressive behavior, exhibit numerical predominance over other species, fiercely defend their territories, and establish mutual exclusivity within their ecological communities. Subdominant species, while not actively defending territories, display a remarkable propensity for aggressively safeguarding their food resources (Cerdá et al., 2013). Lastly, subordinate species, characterized by small colonies devoid of recruitment systems, employ strategies to avoid physical confrontations with other colonies and species. Nonetheless, they exhibit a strong commitment to defending their nests against potential threats (Cerdá et al., 2013). Different foraging strategies, such as the subordinate species’ ability to discover resources before dominant species (dominance-discovery trade-off, Fellers, 1987) or their capacity to forage across a broader temperature range compared to dominant species —dominance-thermal tolerance trade-off (Fellers, 1989), can contribute to the coexistence of ants within a community.
The present study focuses on the analysis of ant feeding habits and dominance hierarchy in response to land use change within a temperate ecosystem located in central Mexico. A habitat with the native oak forest vegetation (complex habitat), was compared to a nearby area where the land use had been altered to induced grassland (simplified habitat). The study hypothesized: i) it is expected that in the oak forest, food items transported to the nest will be more varied than in the induced grassland. This variation at the species level is also anticipated due to the change in land use. The conversion from a complex habitat (oak forest) to a simplified habitat (induced grassland) will likely provide fewer resources variety for the ants; ii) it is expected that as a result of the disturbance in the induced grassland, ants will be more generalized in low heterogeneous habitats due to the dominance of generalist ant species. Additionally, the arrival times at the baits will be shorter due to the absence of leaf litter; iii) a lower dominance index is expected in the induced grassland than in the oak forest, due to the presence of subdominant species and the modification of the dominance hierarchy system by the land use change.
Materials and methods
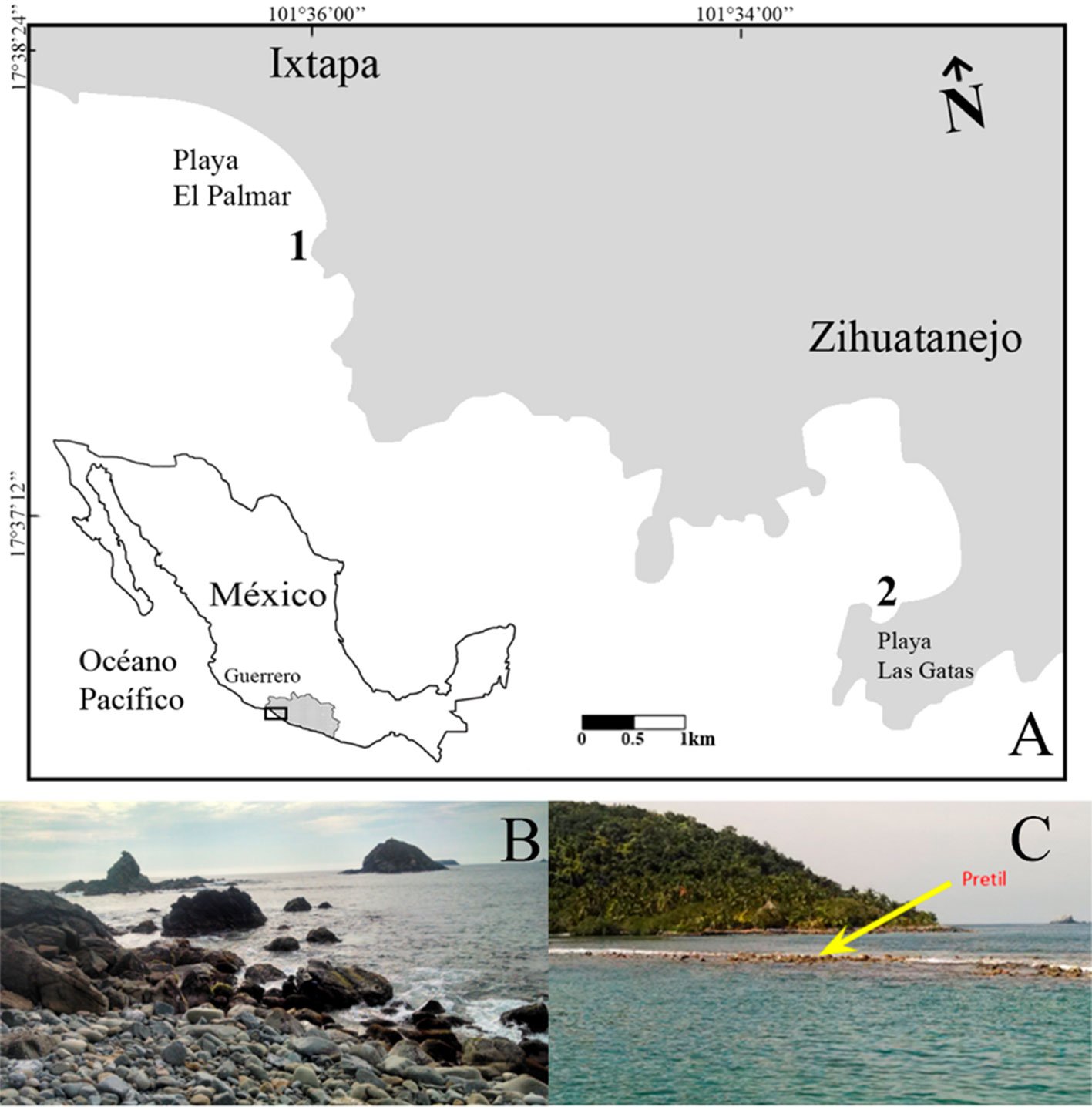
The study was conducted within “Flor del Bosque” State Park, located in the municipality of Amozoc de Mota, in the state of Puebla. The coordinates of this protected reserve are 19°00’00”-19°01’50” N, 98°20’35”-98°20’53” W. The State Park encompasses an area of 664.03 hectares, characterized by altitudes ranging from 2,225 to 2,400 m asl. The annual average temperature in this region fluctuates between 14 °C and 16 °C, with the majority of rainfall occurring during the summer months, contributing to an average annual precipitation of 750 to 950 mm. It is important to note that the park experiences a distinct dry season lasting approximately 6 months, from November to April, as reported by Costes et al. (2006). The native vegetation of the reserve primarily consists of oak forest [Quercus castanea (Née), Q. laurina (Bonpl.), Q. laeta (Liebm.) (Fagaceae)]. However, human disturbances have led to the presence of induced grassland areas and, to a lesser extent, eucalyptus plantations [Eucalyptus spp. L´Hér) (Myrtaceae)] within the park, as documented by Costes et al. (2006). Given that the disturbed vegetation within the reserve primarily consisted of induced grassland, our research aimed to establish a meaningful comparison between native and altered vegetation.
Ant communities were surveyed once a month during specific periods (April, August and October 2015, and January to March 2016). The survey methodology involved the establishment of 6 transect plots (400 m × 20 m), with 3 plots located in the oak forest and 3 in the induced grassland. The spatial distribution of these transects can be observed in Figure 1. To locate ant nests, the transects were traversed, and various substrates such as leaf litter, stones, trunks, and branches were lifted and examined. Additionally, a total of 5 tuna baits and 5 honey baits were placed along the transects at 10 m intervals to attract ants and to follow them to locate their nest. The nests found during the survey were georeferenced for accurate spatial documentation. A 5-minute observation period was designated for each nest, wherein all food items transported by the ants to their nests were recorded. These observations were conducted between 9:00 a.m. and 3:00 p.m. on the sampling days. Each recorded item was subsequently categorized into one of the following groups: 1) plant elements, encompassing any component of plants except for seeds, 2) seeds, and 3) arthropods. After each observation period, 2 bait samples, enclosed within Petri dishes, were positioned in close proximity to the nests (approximately 10 cm), maintaining a distance of approximately 5 centimeters from each other. Various studies exploring the feeding habits, preferences, and foraging strategies of ants have employed diverse bait types. Typically, these baits consist of carbohydrates such as honey and other sources representing proteins such as tuna (Houdria et al., 2015; Lynch et al., 1980; Spotti et al., 2015; Trigos-Peral et al., 2016). The bait materials employed in the study consisted of honey and tuna, which were consistently provided in a standardized amount of one tablespoon, equivalent to approximately 5 grams. To quantify the number of individual ants on each type of food resource, we placed baits and observed them for 5 min. Ants responded very quickly to the baits; therefore, the observation time was limited to 5 min. Within this duration, the number of individuals was recorded for each bait type. After the observation periods, 1 to 3 ants were collected from each nest using a vacuum cleaner or tweezers and then preserved in Eppendorf tubes containing 70% ethanol. In the laboratory, the ants were separated, mounted, and identified to the genus level using the key by Mackay and Mackay (1989). In certain instances, the species identification was feasible by comparing the collected specimens with those present in the Entomological Collection of the Universidad de las Américas Puebla (UDLAP).
To analyze the food preferences of the ant community, we considered 3 factors. First, we determined whether the number of foragers carrying plants, seeds, or arthropods to the nest differed between the whole ant communities of each vegetation type or among ant species. We used the appropriate contingency tables and chi-square tests for such comparisons. Second, we determined whether the presence of foragers at each bait during each minute of the 5-minute period was influenced by vegetation type, bait type (honey or tuna), ant species or interactions among these factors. We used a binomial model to evaluate this response variable, with “ant presence” coded as 1 and “absence” as 0. To select the most relevant explanatory variables, we employed a stepwise forward approach. We began with the simplest model and successively added each response variable. The significance of adding each variable was assessed using an x2 test, comparing the previous model with the new model. If a variable was found to be statistically significant, it was retained in the model. Otherwise, it was removed, and we proceeded with the next explanatory variable. This stepwise procedure allowed us to build the final model with the most significant factors explaining the presence of foragers at each bait. No overdispersion was detected in the selected model. Post-hoc analyses were conducted when significant differences were detected. The statistical analysis of data was performed using R software (R Core Team, 2022).
We evaluated the dominance hierarchy of the ants in each vegetation type (oak forest and induced grassland), ant species, and bait types using the same transects as for the detection of the nests (Fig. 1), during October 2015 and January, February and March (2016). At each transect, 9 sampling points were established on the ground, spaced 10 m apart. Each sampling point consisted of a pair of Petri dishes with baits: one with honey and one with tuna, placed less than 5 cm apart. This resulted in a total of 540 baits (5 replicates × 6 transects × 9 sampling points × 2 baits; 2 samplings in February). These types of baits have been widely used for ant dominance hierarchy studies (Dáttilo et al., 2014; Parr & Gibb, 2012; Trigos-Peral et al., 2016).
For the first 3 sets of bait, we recorded the arrival times of the ants during a one-hour observation period. Most of the ant species were identified in the field. For the ants that were not identified, a few individuals (2 to 3 specimens) were collected. In the case of the remaining 6 sets of bait, they were filled with water. Ants that fell onto the Petri dish after 2 h were collected. The presence of water did not deter the ants from visiting, but it allowed us to determine which ants had been attracted to the baits. The baits were placed and retrieved from different transects in the field between 9:00 a.m. and 5:00 p.m. The order of the transects was changed on different sampling days to mitigate the potential impact of the time of day. Ant specimens collected from the baits were preserved in 70% ethanol and transported to the laboratory for further identification.
Figure 1. Localization of the transects used in the study zone (mapped by Luna F.).
We chose numerical dominance to assess the dominance hierarchy of the ants —i.e., ordering of ant species based on their numerical or behavioral dominance by vegetation types (Andersen, 1992; Cerdá et al., 1997; Stuble et al., 2017), ant species and bait type. As numerical and behavioral dominance are highly correlated, this method has been well-established and documented in the ant literature (Dáttilo et al., 2014; Dejean & Corbara, 2003; Parr, 2008; Parr & Gibb, 2012; Santini et al., 2007). This method indicates which species are consistently present at the baits, and which ones dominate the baits numerically and thus monopolize them (Parr, 2008). We represented numerical dominance using the numerical dominance index (DI) for each morphospecies calculated by the formula: DI = (Di)/(Di + Si), where, Di is the number of baits monopolized by the species of ant i, and Si is the number of baits that the species of ant i used but did not monopolize. Baits were considered to be monopolized when more than 5 individuals (workers and/or soldiers) of the same morphospecies were using the resource without the presence of other morphospecies. This measure (more than 5 individuals) takes into consideration that in temperate climates, ants are less abundant, and recruitment is considered weaker than in tropical environments where the index has been widely more (Santini et al., 2007). Therefore, dominant morphospecies are those that find and monopolize a larger proportion of the food resources in a given environment. The value of the index ranges from 0 (completely submissive species) to 1 (totally dominant species) and is similar to the “monopolization index” used in other studies (Dáttilo et al., 2014; Fellers, 1987; Parr & Gibb, 2012; Santini, et al., 2007). In this study, ant species with a DI lower than 0.5 were classified as submissive.
The arrival times of the ants were compared using survival curves using the survival package of the R Software program. The one-hour and 2-hour baits were used to calculate the ID of the ant species. To compare the ID between vegetation and bait types, t-tests were performed. To compare the DI between ant species, a one-way ANOVA test was performed after applying a square root transformation to meet the normality requirements. These analyses were performed using the program STATVIEW 5.0 (Abacus Concepts Inc., 1996).
Results
Fourteen morphospecies of ants were recorded in this study belonging to 11 ant genera (Table 1). For the analysis of the results from the nests and the baits placed near them, 2 morphospecies of Pheidole were identified. However, due to the low number of records for each morphospecies across different vegetation types, Pheidole sp. 1 and Pheidole sp. 2 were combined for the analysis. In the oak forest, 2 out of 19 records corresponded to Pheidole sp. 1, and in the grassland, 4 out of 15 records corresponded to Pheidole sp. 2. Since Pheidole species often share similar feeding habits and are functionally similar (Agosti, 2000; Andersen, 2000), these 2 morphospecies were grouped to increase statistical power, and the analysis was conducted at the genus level. In contrast, another Pheidole species, Pheidole sp. 3, was recorded in the baits used to determine the DI and was analyzed independently due to its different context of occurrence. The rest of the analyses were conducted at the species level.
In the case of the nests, we found 9 morphospecies distributed among 34 nests, 19 of these were located in the oak forest and 18 in the induced grassland (Table 2). No significant differences were observed in the types of food elements those individual ants transported to their nest when comparing different vegetation types (x2 = 2.13, df = 2, p = 0.34). In the oak forest considering all ant species, there were no significant differences in the number of individuals carrying elements from different categories (x2 = 0.03, df = 1, p = 0.86) (Fig. 2). Of the genera of ants observed carrying food elements to their nests, only 2 genera were observed with a single type of food resource: Prenolepis imparis (Say) only carried arthropods and Dorymyrmex insanus (Buckley) only seeds.
Table 1
List of ant species recorded in this study in “Flor del Bosque” State Park in Puebla, Mexico.
Conversely, ants of the genus Pheidole spp. carried a greater quantity of plant elements compared to the other categories (x2 = 10.4, df = 2, p = 0.006); only 2 of these individuals were observed carrying arthropods, and none were recorded carrying seeds. No significant differences were found for D. insanus (x2 = 2, df = 2, p = 0.37), P. barbatus (x2 = 1.6, df = 2, p = 0.450), nor P. imparis (x2 = 4, df = 2, p = 0.135) among the 3 food categories. Only P. barbatus carried all 3 types of food (Fig. 2).
Considering all ant species, there were no significant differences in the number of individuals carrying different food elements in the induced grassland (x2 = 0.66, df = 1, p = 0.41, Fig. 2). Among the observed ant species that transported food types to their nests, P.barbatus was found to carry both plant elements and seeds. Additionally, Dorymyrmex insanus and Pheidole spp. were observed carrying food from all 3 categories (Fig. 2). Pheidole spp. (x2 = 10.33, df = 2, p = 0.006) and P. barbatus (x2 < 30.33, df = 2, p < 0.001), carried a higher number of plant elements. For D. insanus no significant differences were found in the number of elements of each type (x2 < 3.77, df = 2, p = 0.15) (Fig. 2).
Figure 2. Number of individuals of the different ant species recorded carrying any food resource to their nest. DORY: Dorymyrmex insanus, PHEI: Pheidole spp., POGO: Pogonomyrmex barbatus, PREN: Prenolepis impairs.
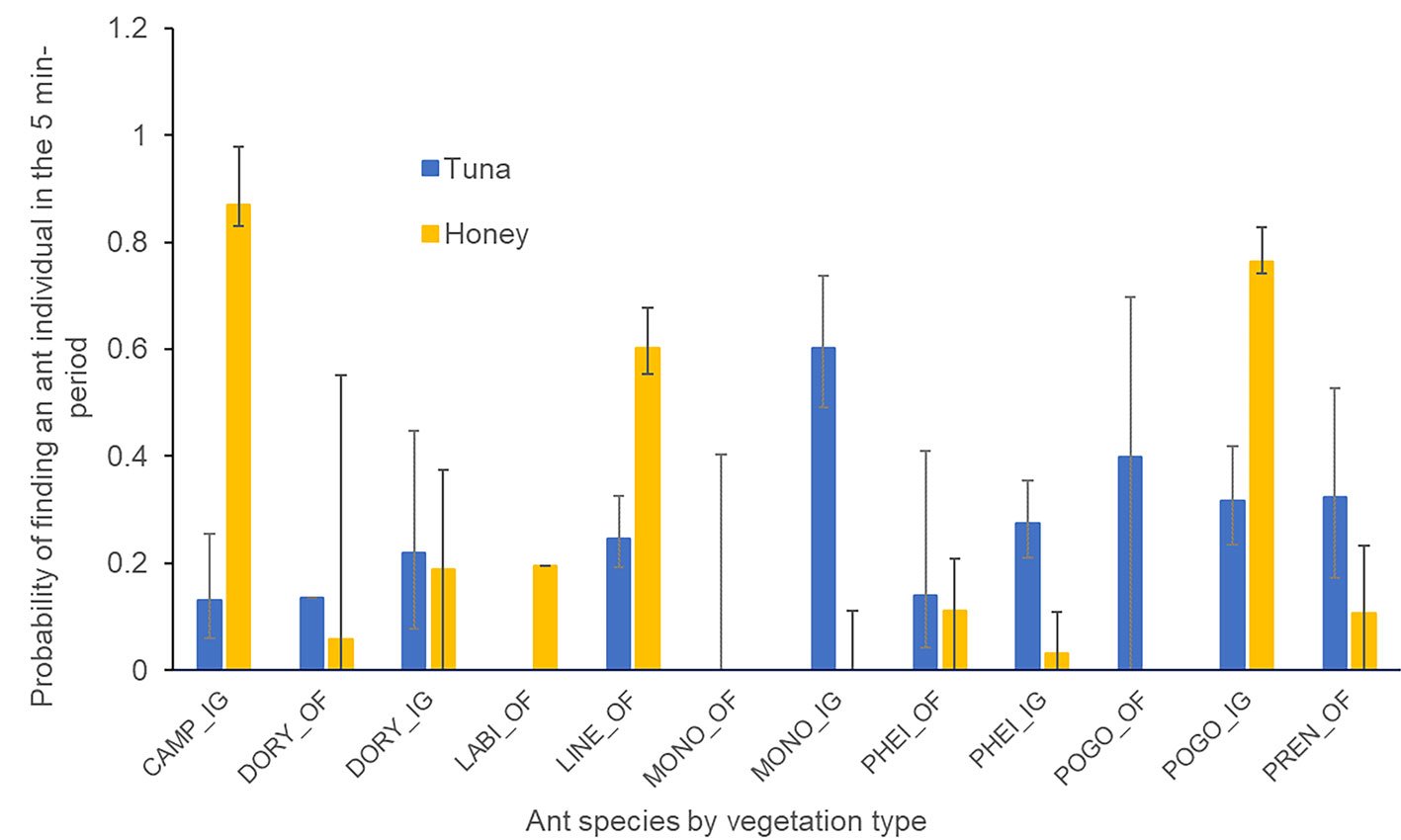
In the induced grassland, 5 ant species were attracted to both types of baits, and 1 species was found in the tuna baits. In the oak area, 5 ant species were observed in both types of baits, and 1 species was attracted to the honey bait (Fig. 3). The probability of presence of a forager during a given minute of the observation period depended on the vegetation type (x2= 8.73, df = 1, p = 0.03), the ant species (x2= 59.64, df = 7, p < 0.001), the interaction between the ant species and the bait (x2= 57.96, df = 8, p < 0.001), and the interaction between the vegetation type, the ant species, and the bait (x2= 26.12, df = 7, p < 0.001). The Tukey contrast test for the vegetation*species*bait interaction showed (Z > 3.83, p < 0.042) that the presence of Linepithema dispertitum (Forel) on the honey bait, in the oak forest, was more likely than the presence of Pheidole spp. on either the tuna or honey bait in the oak forest, Pheidole spp. on the honey bait in the induced grassland, and P. impairs on the honey bait in the oak forest (Fig. 3). The probability of Camponothus rubrithorax (Forel) being present on the honey bait in the induced grassland, was greater than that of D. insanus in either the tuna or honey bait in the induced grassland, Pheidole spp. on either the tuna or honey bait in the oak forest, as well as its presence in the honey bait in the induced grassland and P. imparis in the honey bait in the oak forest (Fig. 3). The probability that P. barbatus was present on the honey bait in the induced grassland, was greater than the presence of Pheidole spp. in either the tuna or honey bait and P. imparis in the honey bait in the oak forest (Fig. 3). Finally, the presence of Pheidole spp. on the tuna bait was more likely than its presence on the honey bait in the induced grassland (Fig. 3).
Figure 3. Probability of finding an ant forager on the honey or tuna baits at the nest of the different ant species registered in the Oak Forest (OF) and the Induced Grassland (IG) in the 5-minute period. CAMPO: Camponotus rubrithorax, DORY: Dorymyrmex insanus, LABI: Labidus coecus, LINE: Linepithema dispertitum, MONO: Monomorium ebenium, PHEI: Pheidole spp., POGO: Pogonomyrmex barbatus, PREN: Prenolepis imparis.
Figure 4. Survival curves depicting the arrival times of ant foragers on the baits (tuna or honey) during a one-hour observation. No significant differences were found in the arrival times among ant species.
No significant differences were found in the arrival times between the vegetation types (x2 = 0.4, df = 1, p = 0.5). Moreover, no significant differences were found in the arrival times among the different ant species (x2 = 7.6, df = 7, p = 0.4) or the type of bait (x2 = 0.01, df = 1, p = 0.9; Fig. 4).
In the dominance index baits 12 ant morphospecies were recorded and different ant morphospecies were recorded in each of the vegetation types (Table 3). When comparing the average dominance index (DI) by vegetation type (mean ± SE, n; oak forest DI = 0.372 ± 0.071, 33; induced grassland DI = 0.251 ± 0.052, 44), was not statistically different (t= 1.091; df = 75; p = 0.2787), which would suggest a submissive behavior in both types of vegetation. Average dominance index by ant species indicates that M. ebenium, P. imparis and Pheidole sp. 3 behave as submissive in both vegetation types (t = 0.590, df = 20, p = 0.5617; t = -0.237, df = 10, p = 0.8171; t = -0.315, df = 11, p = 0.7590, respectively) (Table 4). It was not possible to compare the DI between vegetation types for Temnothorax sp., T. augusti, C. rubrithorax, N. austroccidua, L. dispertitum, D. insanus or P. pallidus, because they were only present in one of the vegetation types (Table 4).
The average dominance index for each ant species was low suggesting that all of them displayed submissive behaviors (F8,68 = 1.949, p = 0.06). Nylanderia austroccidua and L. dispertitum had the highest dominance indices, although it should be noted that N. austroccidua only had one record and L. dispertitum had 3 records. Additionally, Monomorium ebenium showed a tendency to behave as dominant (Table 4).
When comparing the mean dominance index by bait type, no significant differences were found (t = 1.023, df = 110, p = 0.3088; mean ± SE, n; tuna = 0.382 ± 0.060, 55; honey = 0.295 ± 0.053, 57). This indicates that ants exhibited submissive behavior in both types of bait.
According to the average dominance index per ant species in relation to the bait type, Pheidole sp. 3, P. imparis, M. ebeninum, C. rubrithorax, and D. insanus exhibited a submissive behavior in both types of baits (t = 0.888, df = 13, p = 0.3907; t= 0.988, df = 15, p = 0.3388; t = 0.189, df = 32, p = 0.8513; t = -0.918, df = 24, p = 0.3675, t = 2.390, df = 5, p = 0.0624, respectively) (Table 5) and L. dispertitum exhibited a dominant behavior in both types of baits (t = 0.421, df = 3, p = 0.7021).
Temnothorax sp. and P. pallidus were not compared statistically due to the low incidence registered; nonetheless, they presented a submissive behavior index in both types of bait. Neither T. augusti nor N. austroccidua were included in this analysis, as they were only present on the honey bait. In none of the species was there a difference in behavior between the baits.
Table 2
Number of nests found in the oak forest and the induced grassland.
Species
Oak forest
Induced grassland
Camponotus rubrithorax
0
3
Dorymyrmexinsanus
4
6
Labidus coecus
1
0
Linepithema dispertitum
2
0
Pheidole spp.
7
6
Pogonomyrmex barbatus
1
2
Prenolepis imparis
3
0
Monomorium ebenium
1
1
Discussion
This study found that certain ant species (e.g., C. rubrithorax, L. dispertitum, P. barbatus) exhibited a preference for honey within specific vegetation types when compared to the rest of the ant community. The compensation hypothesis (Davidson, 2005; Kaspari & Yanoviak, 2001) predicts that the attractiveness of a nutrient to an organism is higher the more limiting it is. It is possible that this result is related to the fact that sugar is generally less available on the ground strata than protein (Kaspari & Yanoviak, 2001; Kaspari et al., 2012).
However, when considering the overall community level, no noticeable food preferences were found between the ant communities in the oak forest and the induced grassland vegetation. This lack of preference was evident both in the items carried to their nests and the resources provided on the baits. There were no significant differences in arrival times at the baits between vegetation types. This similarity can be attributed to ants experiencing soil-level heterogeneity similarly in both oak forests and induced grasslands (see below).
The ant communities in both habitats were composed of ant species displaying submissive behavior. This characteristic could be attributed to stress factors such as low temperatures in the oak forest and disturbances in the induced grassland, which potentially reduce ant competition. In the oak forest, low temperatures may favor the presence of cold climate specialists that can forage at low temperatures without the need to display a dominant behavior. In the induced grassland, disturbances may favor the presence of generalist species that take advantage of the absence of dominant species such as the dominant Dolichoderinae (see below). This study emphasizes the significance of ant species’ response to their environment and their adaptability in dealing with disturbances.
Our findings do not provide support for vegetation-scale disparities in food preferences, first and second hypotheses, which proposed greater differences in the food items being carried to the nest in the oak forest and reduced food preferences in the induced grassland. There were no differences between vegetation types in the number of seeds, plants, or arthropods taken by the ant foragers to their nests. Similarly, there was not a preference for a specific food bait (honey or tuna) between the oak forest and induced grassland ant communities. These findings are consistent with previous studies conducted by Radnan et al. (2018) and Castillo-Guevara et al. (2019), which did not identify differences in food preferences or foraging strategies between natural and disturbed vegetation at the community level. However, our observations did reveal that certain ant species exhibited preferences for specific types of food, within particular vegetation types, in accordance with the first hypothesis and second hypothesis but at the species level, which proposed an influence of ant species on food preferences. The specific preferences of these ant species are discussed in detail below.
Table 3
Visit frequency (i.e., number of foragers) per species in each vegetation type and functional groups they belong to (Andersen, 2000). GM = Generalized Myrmicinae, CCS = cold climate specialist, SC = subordinate Camponotini, TCS = tropical climate specialist, O = opportunist.
Subfamily
Specie
Funtional group
Oak forest
Induced grassland
Myrmicinae
Monomorium ebenium
GM
753
167
Myrmicinae
Pheidole sp. 3
GM
14
236
Myrmicinae
Temnothorax sp.
CCS
5
0
Myrmicinae
Temnothorax augusti
CCS
0
1
Formicinae
Prenolepis imparis
CCS
289
11
Formicinae
Camponotus rubrithorax
SC
0
351
Formicinae
Nylanderia austroccidua
TCS
0
30
Dolichoderinae
Linepithema dispertitum
CCS
190
0
Dolichoderinae
Dorymyrmex insanus
O
0
26
Pseudomyrmecinae
Pseudomyrmex pallidus
TCS
0
10
Total
1,251
832
Table 4
Average numerical dominance index (DI) (mean ± SE, n) per ant morphospecies. (-) Unregistered species.
Specie
Oak forest
Induced grassland
ID
Monomorium ebenium
0.4 ± 0.1, 13
0.3 ± 0.1, 9
0.4 ± 0.0, 22
Pheidole sp.3
0.2 ± 0.2, 4
0.2 ± 0.1, 9
0.2 ± 0.1, 13
Temnothorax sp.
0 ± 0, 3
–
0 ± 0, 3
Temnothorax augusti
–
0, 1
0 ± 0, 1
Prenolepis imparis
0.3 ± 0.1, 10
0.5 ± 0.5, 2
0.3 ± 0.1, 12
Camponotus rubrithorax
–
0.1 ± 0.0, 15
0.1 ± 0.0, 15
Nylanderia austroccidua
–
1, 1
1, 1
Linepithema dispertitum
0.7 ± 0.1, 3
–
0.7 ± 0.1, 3
Dorymyrmex insanus
–
0.2 ± 0.1, 4
0.3 ± 0.1, 4
Pseudomyrmex pallidus
–
0 ± 0, 3
0 ± 0, 3
Pheidole spp., in the induced grassland, were observed carrying a greater quantity of plant elements to their nests in both vegetation types. However, it exhibited a preference for the tuna bait in induced grassland, possibly indicating a supplementary dietary preference. Given the extensive diversity within the Pheidole genus, which encompasses 900 species described worldwide (Wilson, 2003), it is not feasible to categorize them based on a specific food habit and our results suggest that it is omnivorous (Table 6). Nevertheless, these results should be interpreted with caution due to the grouping of species.
Camponotus rubrithorax and P. barbatus, in the induced grassland, as well as L. dispertitum, in the oak forest, were more frequently observed foraging the honey bait compared to other ant species. Camponotus (Mayr) is a genus known for its nectarivorous habits and consumption of other sweet secretions, such as honeydew which coincides with its preference for honey found in the present study (Nettimi & Iyer, 2015). In this study, L. dispertitum predominantly consumed honey, although it is a generalist forager species capable of consuming other types of food as well (Table 5), in the study site it is found exclusively in the oak forest (Cuautle et al., 2016).
Despite Pogonomyrmex (Mayr) has been recognized as a granivorous genus (Pirk & López-de Casenave, 2014), this study provided new insights into the foraging preferences of P. barbatus, revealing a notable inclination towards carbohydrates and plant elements within the induced grassland. Moreover, we even registered individuals transporting arthropods to their nests. These findings (Table 5) strongly indicate that certain species within the Pogonomyrmex genus exhibit a generalist foraging behavior. Other species such as D. insanus, which used resources more intensively in the induced grassland, showed no preference for any of the baits or items taken to the nest, which coincides with the generalist forager behavior, observed in open and disturbed habitats by Cuezzo and Guerrero (2012).
Table 5
Average numerical dominance index (DI) (mean ± SE, n) per ant species in both bait types (tuna, honey). (-) Unregistered species.
Species
Tuna
Honey
Monomorium ebenium
0.4 ± 0.1, 18
0.4 ± 0.1, 16
Pheidole sp.3
0.4 ± 0.2, 7
0.1 ± 0.1, 8
Temnothorax sp.
0, 1
0 ± 0, 2
Temnothorax augusti
–
0, 1
Prenolepis imparis
0.5 ± 0.1, 7
0.3 ± 0.1, 10
Camponotus rubrithorax
0.1 ± 0.0, 14
0.2 ± 0.0, 12
Nylanderia austroccidua
–
1, 1
Linepithema dispertitum
0.7 ± 0.2, 2
0.6 ± 0.3, 3
Dorymirmex insanus
0.6 ± 0.3, 3
0 ± 0, 4
Pseudomyrmex pallidus
0 ± 0, 3
0, 1
Table 6
The biology and ecology of the ant genera found in this study, in the Flor del Bosque State Park (Agosti et al., 2000; AntWiki, n.d.).
Genera
Microhabitat
Food habits
Camponotus
Ground nesting, decaying wood and in trees
Generalist foragers
Dorymyrmex
————–
Generalist foragers
Linepithema
————–
Generalist foragers
Labidus
Epigeous,bivouacs
Predators (Army ants)
Monomorium
————–
Generalized foragers, harvesters
Nylanderia
Nest in leaf litter, soil, or in rotten wood
Generalist foragers
Pheidole
Soil or decaying wood
Granivores or omnivores
Pogonomyrmex
Ground nesting
Generalist foragers and granivores
Prenolepis
————–
Generalist predators
Pseudomyrmex
Mostly arboreal (nesters and foragers), few epigaeic
Nesting in ground, and under stones, in wood, and in trees
Generalized foragers and parasites
The previous results (e.g., C. rubrithorax, Pheidole spp., D. insanus) support the reported food habits of specific ant genera. Nonetheless, it is also possible to interpret these results as ants taking nutrients from the baits that are not currently available or that are limited within their community to supplement their diet (Compensation hypothesis) (Davidson, 2005; Kaspari & Yanoviak, 2001). In this study, significant differences in ant species presence on baits were primarily associated with the presence of ant species on honey baits. This finding aligns with the compensation hypothesis (Davidson, 2005; Kaspari & Yanoviak, 2001), which posits that the utility of a resource remains constant across species and varies solely with availability. The hypothesis predicts a singular limiting resource that is locally in shortest supply. Consequently, habitats with relatively high protein availability should attract ants more inclined towards carbohydrates, and vice versa. Previous research has demonstrated that litter ant communities are limited by carbohydrates, whereas ant arboreal communities face protein limitations in tropical regions (Kaspari & Yanoviak, 2001; Kaspari et al., 2012). Although this study was conducted in a temperate environment, the results might be related to the usual scarcity of sugar in the ground strata compared to protein.
Our results do not align with the second hypothesis, which speculated faster arrival times in the induced grassland. We did not find significant differences in arrival times among vegetation types, ant species, or bait types. Ants could be experiencing similar heterogeneities at the soil level between the oak forest and induced grassland. While it is expected that the presence of more litter in the oak forest could hinder ant movement, within the induced grassland, the vegetation morphology itself (long grass species) could be interfering with ant movement. For instance, Hernández-Flores et al. (2016) observed that the foraging performance of P. barbatus was reduced due to the presence of herbaceous vegetation in plots where regeneration after grazing was permitted.
Dominance hierarchy. Disturbance typically favors the presence of generalist and opportunistic species, hence, we expected to find a lower dominance index within the induced grassland (third hypothesis). Nonetheless, we did not find enough evidence to support this hypothesis as we registered ant communities consisted of submissive species in both habitats. Additionally, the ants did not show a tendency to dominate a specific resource, neither at the bait level nor at the species level, suggesting a lack of food preferences. These results are consistent with the findings of Castillo-Guevara et al. (2019), who registered no significant differences in dominance indices between a native oak forest and an altered agricultural land. Moreover, the authors reported intermediate to low values of dominance within the ant communities of each vegetation type. Our findings could be attributed to the presence of other factors that could have overshadowed the role of competition in shaping the organization of ant communities. For example, a review by Parr and Gibb (2012), which encompassed data from 3 continents, indicated that the trade-off between discovery and dominance occurs primarily when parasitoids are present. In environments without parasitoids, species with high discovery abilities tend to also be dominant (Parr & Gibb, 2012). While our study did not specifically assess the discovery-dominance trade-off, the observed lack of differences in arrival times and similar dominance values among the ants suggest that dominance was not a prominent factor in our study sites. It is possible that factors such as low temperatures in the oak forest, or disturbance in the induced grassland, may have played a role in relaxing dominance. According to Andersen (2000), in disturbed vegetation like the induced grassland, it is expected to observe the presence of subdominant ant species that exploit the absence of dominant species from the native vegetation, such as dominant Dolichoderinae. The findings obtained during this study align with this prediction and provide support for it. Notwithstanding, the absence of ants with high dominance indices in the studied communities does not necessarily indicate a complete lack of dominance hierarchy. In each community, the ants can still be ordered based on their DI. For example, in the oak forest, L. dispertitum had the highest DI value (0.7 DI), M. ebenium had an intermediate value (0.4 DI) and Temnothorax spp. had the lowest value (0.0 DI).
Land use change did not seem to influence food preferences or foraging strategies at the community level. However, we observed an effect at species level, indicating that individual ant species exhibited specific food preferences. Carbohydrates could be the limiting resource in the oak forest and the induced grassland litter ant communities, as some ant species showed preference for honey baits. Although competition is typically considered a key factor in understanding food preferences among ants, it is noteworthy that both, the natural oak forest and induced grassland, were predominantly populated by submissive ant species. Therefore, it appears that other factors instead of competition may be playing a role in shaping food preferences within these communities. In conclusion, our study reveals that ant species may exhibit preferences for specific foods, which could be limited in their environment. The ability of ants to respond to available resources enables them to optimize their nutrient intake, as well as adapt and persist under variable conditions, including disturbances. Understanding the distinct dietary preferences and foraging strategies of ant species within functional groups will provide valuable insights into their ecological roles and potential impacts on ecosystem dynamics. Such investigations would enhance our ability to predict the responses of ants to diverse forms of disturbances in an anthropized world.
Acknowledgements
We appreciate the assistance provided by the authorities of “Flor del Bosque” State Park, coordinator Enrique Martínez Romero (M.S.) and director Mario Alberto Romero Guzmán (MVZ). We would also like to thank Florencio Luna Castellanos for his support with the fieldwork. This study was financed by Consejo Nacional de Humanidades Ciencias y Tecnologías (Conahcyt) as part of a grant awarded to Mariana Cuautle (223033).
References
Abacus Concepts Inc. (1996). Abacus Concepts, Stat View Reference. Berkeley, California.
Agosti, D., Majer, J. D., Alonso, L. E., & Schultz, T. R. (2000). Ants: standard methods for measuring and monitoring bio- diversity. Washington D.C.: Smithsonian Institution Press.
Andersen, A. (2000). A global ecology of rainforest ants: functional groups in relation to environmental stress and disturbance. In D. Agosti, J. D. Majer, L. E. Alonso, & T. R. Schultz (Eds.), Ants: standard methods for measuring and monitoring biodiversity (pp. 25–34). Washington D.C.: Smithsonian Institution Press.
Andersen, A.N. (1992). Regulation of “momentary” diversity by dominant species in exceptionally rich ant communities of the Australian seasonal tropics. The American Naturalist, 40, 401–420. https://www.journals.uchicago.edu/doi/abs/10. 1086/285419
Arnan, X., Cerdá, X., & Retana, J. (2012). Distinctive life traits and distribution along environmental gradients of dominant and subordinate Mediterranean ant species. Oecologia, 170, 489–500. https://doi.org/10.1007/s00442-012-2315-y
Castillo-Guevara, C., Cuautle, M., Lara, C., & Juárez-Juárez, B. (2019). Effect of agricultural land-use change on ant dominance hierarchy and food preferences in a temperate oak forest. PeerJ, 7, e6255. https://peerj.com/articles/6255/
Cerdá, X., Arnan, X., & Retana, J. (2013). Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecological News, 18, 131–147. https://doi.org/10.25849/myrmecol.news_018:131
Cerdá, X., Retana, J., & Cros S. (1997). Thermal disruption of transitive hierarchies in Mediterranean ant communities. Journal of Animal Ecology, 66, 363–374.
Costes-Quijano, R., Meza, A. R., Macías-Juárez, A., Berriel-Mastreta, C. A., Cortés-Atilano, B., Martínez-Romero, L. E. et al. (2006). Plan de manejo Parque Ecológico Recreativo General Lázaro Cárdenas “Flor del Bosque”. [Management Plan of the recreative ecological Park “General Lázaro Cárdenas “Flor del Bosque”]. Ciudad de México: Gobierno del Estado de Puebla/ Secretaría de Medio Ambiente y Recursos Naturales.
Cuautle, M., Vergara, C., & Badano, E. (2016). Comparison of ant community diversity and functional group composition associated to land use change in a seasonally dry oak forest. Neotropical Entomology, 45, 170–9. https://doi.org/ 10.1155/2012/516058
Cuezzo, F., & Guerrero, R. J. (2012). The Ant Genus Dorymyrmex Mayr (Hymenoptera: Formicidae: Dolicho- derinae) in Colombia. Psyche, 51605, 1–24. https://doi.org/ 10.1155/2012/516058
Dáttilo, W., Díaz-Castelazo, C., & Rico-Gray, V. (2014). Ant dominance hierarchy determines the nested pattern in ant-plant networks. Biological Journal of the Linnean Society, 113, 405–414. https://doi.org/10.1111/bij.12350.
Davidson, D. W. (2005). Ecological stoichiometry of ants in a New World rain forest. Oecologia, 142, 221–231. https://doi.org/10.1007/s00442-004-1722-0
Dejean, A., & Corbara, B. (2003). A review of mosaics of dominant ants in rainforests and plantations. In Y. Basset, V. Novotny, S. E. Miller, & R. L. Kitching (Eds.), Arthropods of tropical forests: spatio-temporal dynamics and resource use in the canopy (pp 341–347). Cambridge: Cambridge University Press.
Ellis, E. C., Goldewijk, K. K., Siebert S., Lightman, D., & Ramankutty, N. (2010). Anthropogenic transformation of the biomes, 1700 to 2000. Global Ecology and Biogeo- graphy, 19, 589–606. https://doi.org/10.1111/j.1466-8238.20 10.00540.x
Fellers, J. H. (1987). Interference and exploitation in a guild of Woodland ants. Ecology, 68, 1466–1478. https://doi.org/ 10.2307/1939230
Fellers, J. H. (1989). Daily and seasonal activity in woodland ants. Oecologia, 78, 69–76. https://doi.org/10.1007/BF0037 7199
Foley, J. A., DeFries, R., Asner, G. P., Barford C., Bonan, G., Carpenter, S. R. et al. (2005). Global consequences of land use. Science, 309, 570–574. https://doi/10.1126/science.1111772
Hoffmann, B. D., & Andersen, A. N. (2003). Responses of ants to disturbance in Australia, with particular reference to functional groups. Austral Ecologyl, 28, 444–464. –https://doi.org/10.1046/j.1442-9993.2003.01301.x
Houdria, M., Salas-López, A., Orivel. J., Bluthgen, N., & Menzel, F. (2015). Dietary and temporal niche differentiation in tropical ants – can they explain local ant coexistence? Biotropica, 47,208–217. https://doi.org/10.1111/btp.12184
Hernández-Flores, J., Osorio-Beristain. M., & Martínez-Garza. C. (2016). Ant foraging as an indicator of tropical dry forest restoration. Environmental Entomology, 45, 991–994. https://doi.org/10.1093/ee/nvw054
Kaspari, M., Donoso, D., Lucas, J. A., Zumbusch, T., & Kay, A. D. (2012). Using nutritional ecology to predict community structure: a field test in Neotropical ants. Ecosphere, 3,93. https://doi.org/10.1890/ES12-00136.1
Kaspari, M., & Yanoviak, S. P. (2001). Bait use in tropical litter and canopy ants-evidence of differences in nutrient limitation. Biotropica, 33,207–211. https://doi.org/10.1646/ 0006-3606(2001)033[0207:BUITLA]2.0.CO;2
Łaska, G. (2001). The disturbance and vegetation dynamics: a review and an alternative framework. Plant Ecology, 157, 77–99. https://doi.org/10.1023/A:1013760320805
Lynch, J. F., Balinsky, E. C., & Vail, S. G. (1980). Foraging patterns in three sympatric forest ant species, Prenolepis imparis, Paratrechina melanderi and Aphaenogaster rudis (Hymenoptera: Formicidae). Ecological Entomology, 5,353–371. https://doi.org/10.1111/j.1365-2311.1980.tb01160.x
Mackay, W., & Mackay, E. (1989). Clave de los géneros de hormigas en México (Hymenoptera: Formicidae). El Paso, Texas: The University of Texas.
Nettimi, R. P., & Iyer, P. (2015). Patch fidelity in Camponotus compressus ants foraging on honeydew secreted by treehoppers. Current Science, 109, 362–366.
Parr, C. L. (2008). Dominant ants can control assemblage species richness in a South Africa savanna. Journal of Animal Ecology, 77, 1191–1198. https://doi.org/10.1111/j.13 65-2656.2008.01450.x
Parr, L., & Gibb, H. (2012). The discovery-dominance trade-off is the exception, rather than the rule Journal of Animal Ecology, 81, 233–241. https://doi.org/10.1111/j.1365-2656. 2011.01899.x
Pirk, G. I., & López-de Casenave, J. (2014). Effect of harvester ants of the genus Pogonomyrmex on the soil seed bank around their nests in the central Monte desert, Argentina. Ecological Entomology, 39, 610–619. https://doi.org/10.1111/een.12140
R Core Team (2022). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/
Radnan, G. N., Gibb, H., & Eldridge, D. J. (2018). Soil surface complexity has a larger effect on food exploitation by ants than a change from grassland to shrubland. Ecological Entomology, 43, 379–388. https://doi.org/10.1111/een.12510
Rico-Gray, V., & Oliveira, P. S. (2007). The ecology and evolution of ant-plant interactions. Chicago: University of Chicago Press.
Sala, O. E., Chapin, F. S., Armesto, J. J., Berlow, E., Bloomfield, J., Dirzo. R. et al. (2000). Global biodiversity scenarios for the year 2100. Science, 287,1770–1774. https://doi.org/10.1126/science.287.5459.17
Santini, G., Tucci, L., Ottonetti, L., & Frizzi, L. (2007). Competition trade-offs in the organisation of a Medite- rranean ant assemblage. Ecological Entomology, 32, 319–326. https://doi.org/10.1111/j.1365-2311.2007.00882.x
Savolainen, R., Vepsäläinen, K., & Wuorenrinne, H. (1989). Ant assemblages in the taiga biome: testing the role of territorial wood ants. Oecologia, 81, 481–486. https://doi.org/10.1007/BF00378955
Manuel Palomo-Morales a, b, *, Luis Enrique Sánchez-Ramos c, d, Luis Gerardo Herrera-Montalvo e, Adolfo G. Navarro-Sigüenza d, f y Pedro Luis Valverde b
a Universidad Autónoma Metropolitana, Doctorado en Ciencias Biológicas y de la Salud, Av. Ferrocarril San Rafael Atlixco, Núm. 186, Col. Leyes de Reforma 1 A Sección, 09310 Ciudad de México, México
b Universidad Autónoma Metropolitana-Iztapalapa, Departamento de Biología, Av. Ferrocarril San Rafael Atlixco, Núm. 186, Col. Leyes de Reforma 1 A Sección, 09310 Ciudad de México, México
c Universidad Nacional Autónoma de México, Instituto de Geociencias, Blvd. Juriquilla 3001, La Mesa, 76230 Juriquilla, Querétaro, México
d Universidad Nacional Autónoma de México, Facultad de Ciencias, Museo de Zoología, Apartado postal 70-399, 04510 Ciudad de México, México
e Universidad Nacional Autónoma de México, Instituto de Biología, Estación de Biología Chamela, Carretera Federal Barra de Navidad-Puerto Vallarta Km 59, 48854 Chamela, Jalisco, México
f Universidad Nacional Autónoma de México, Unidad Multidisciplinaria de Docencia e Investigación, Facultad de Ciencias-Campus Juriquilla, Blvd. Juriquilla 3001, La Mesa, 76230 Juriquilla, Querétaro, México
*Autor para correspondencia: manuelpm.biol@gmail.com (M. Palomo-Morales)
Recibido: 25 junio 2024; aceptado: 15 agosto 2024
Resumen
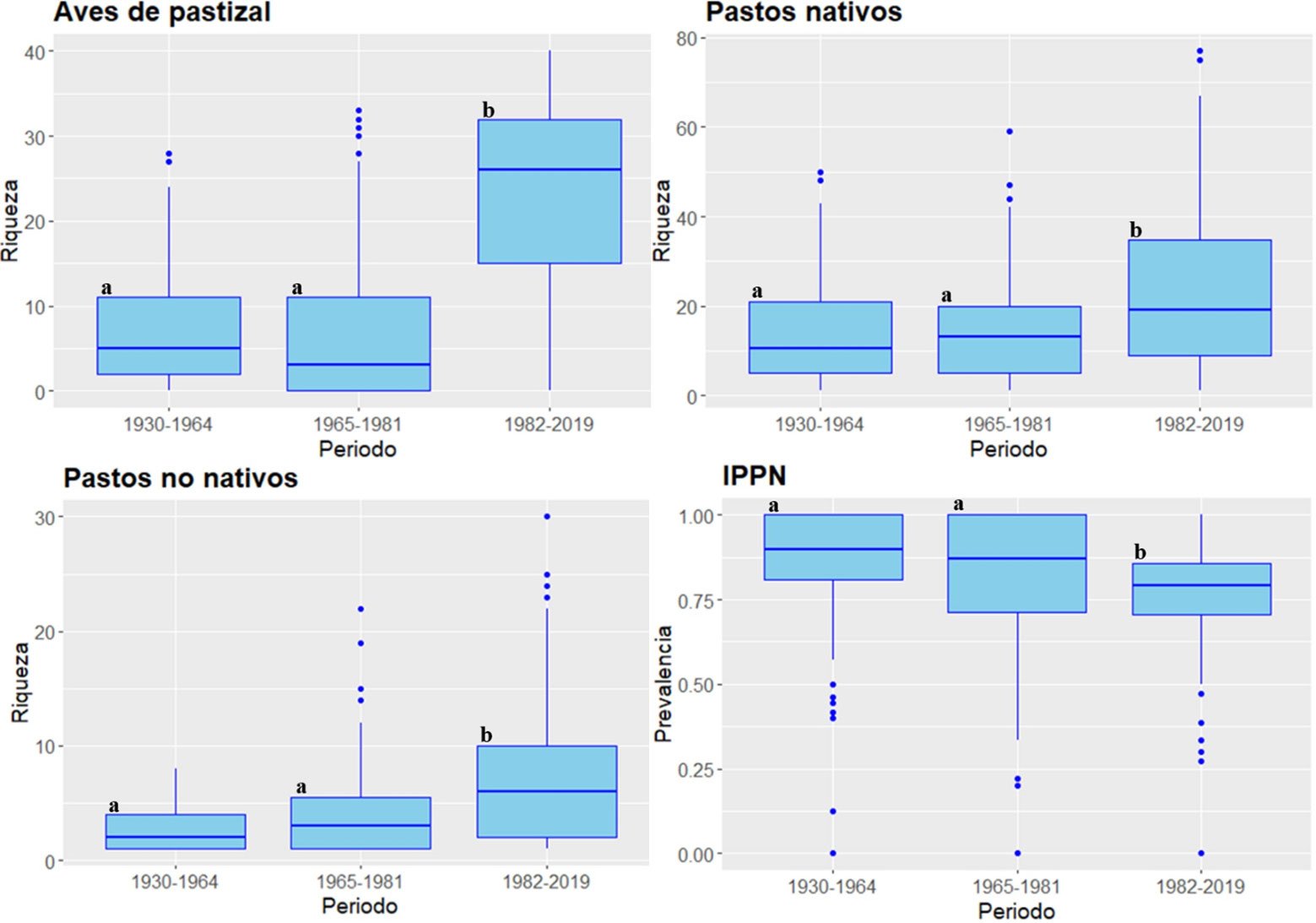
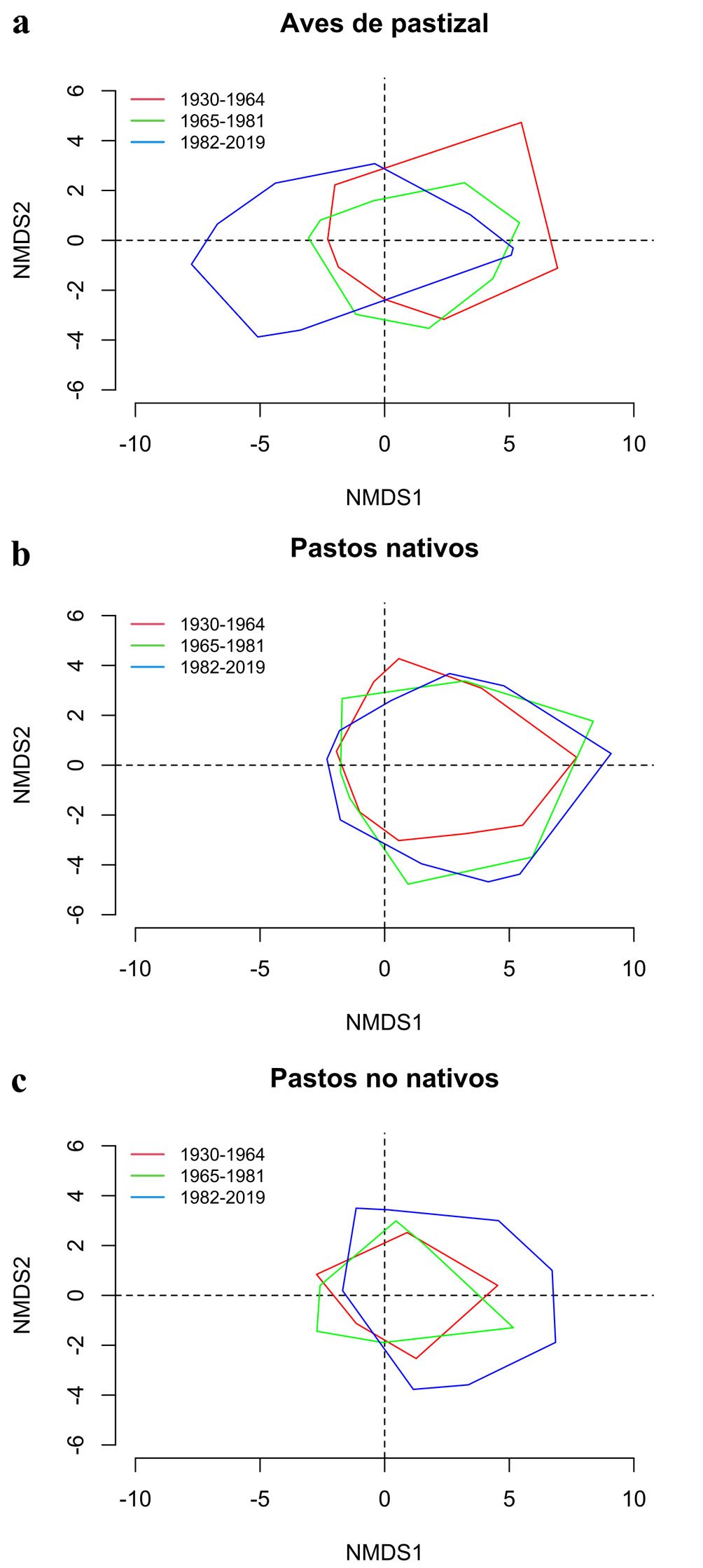
Evaluamos la diversidad de pastos nativos, pastos no nativos y aves de pastizal en el Desierto Chihuahuense en un contexto histórico-ambiental de presión antropogénica en los periodos de 1930-1964, 1965-1981 y 1982-2019. Analizamos la riqueza regional, la diversidad beta y la composición de 41 especies de aves de pastizal, 149 especies de pastos nativos y 60 especies de pastos no nativos, así como la prevalencia de pastos nativos y su correlación con las aves de pastizal en cada periodo. Los resultados mostraron una reducción de la diversidad beta en los 3 ensambles a lo largo del tiempo, sin relaciones significativas entre el ensamble de aves y la prevalencia de pastos nativos en el último periodo. Esto sugiere que la distribución actual de las aves de pastizal es independiente del tipo de pasto a nivel regional. La expansión de pastos no nativos en el Desierto Chihuahuense, favorecida por políticas públicas a lo largo del tiempo, ha conformado un ensamble de pastizales mixtos, lo que actualmente no afecta la composición de aves de pastizal a escala regional. Sugerimos realizar evaluaciones locales para comprender mejor la relación entre las aves de pastizal y los distintos pastos presentes en la región.
Palabras clave: Diversidad beta; Presencia-ausencia; Ensamble de especies; Índice de prevalencia; NMDS
Influence of the introduction of non-native grasses on the species richness of bird assemblages in the Chihuahuan Desert region: a temporal analysis
Abstract
We assessed the diversity of native grasses, non-native grasses, and grassland birds in the Chihuahuan Desert in a historical-environmental context of anthropogenic pressure during the periods 1930-1964, 1965-1981, and 1982-2019. We analyzed the regional richness, beta diversity, and composition of 41 grassland bird species, 149 native grass species, and 60 non-native grass species, as well as the prevalence of native grasses and their correlation with grassland birds in each period. The results showed a reduction in beta diversity in the 3 assemblages over time, with no significant relationships between bird assemblage and native grass prevalence in the last period. This suggests that the current distribution of grassland birds is independent of grass type at the regional level. The expansion of non-native grasses in the Chihuahuan Desert, favored by public policies over time, has formed an assemblage of mixed grasslands, which currently does not affect the composition of grassland birds at the regional level. We suggest conducting local assessments to better understand the relationship between grassland birds and the different grasses present in the region.
Keywords: Beta diversity; Presence-absence; Species assemblage; Prevalence rate; NMDS
Introducción
Las especies introducidas de animales y plantas alteran la composición y ecología de las comunidades biológicas, tanto local como regionalmente (Davis, 2003). En particular, las investigaciones sobre plantas introducidas e invasoras se han centrado en cuantificar y describir sus impactos a diversas escalas, desde poblaciones y especies, hasta comunidades y ecosistemas (Hejda et al., 2009; Pyšek et al., 2012). Con el tiempo, muchas especies de plantas introducidas se han expandido y pueden, potencialmente, acelerar la extirpación de poblaciones de especies nativas de forma localizada y, eventualmente, aumentar la homogeneidad biótica del hábitat, lo que reduce la diferenciación de las floras a nivel regional (Qian y Ricklefs, 2006).
Las especies de pastos (familia Poaceae) que han sido introducidas en diferentes regiones son el principal agente que ha perturbado los pastizales nativos de extensas regiones del mundo, como ha sucedido por ejemplo en las praderas de California, las Grandes Planicies del centro de los EUA, así como en varias regiones del centro de Australia, entre otras (Gibson, 2009). Los efectos de la invasión de pastos exóticos dependen en parte de la gravedad de cualquier cambio ambiental o del régimen de perturbación humana, como el pastoreo excesivo, la sequía, la frecuencia de incendios, el pisoteo del ganado, la fragmentación del hábitat y la cercanía con tierras agrícolas que favorecen su dispersión (Chaneton et al., 2002; Gibson, 2009; Hamilton, 1999; Weaver et al., 1996). Algunas de las consecuencias de las invasiones por estos pastos incluyen la simplificación de la estructura de la vegetación, la reducción de la biodiversidad al disminuir el hábitat preferente de muchas especies de vida silvestre, así como la alteración de los regímenes naturales de incendios y ciclo de nutrientes, entre otras (Wied et al., 2020).
En Norteamérica, las regiones de pastizales naturales ocupan una importante porción de la superficie continental (Gibson, 2009). Se pueden encontrar áreas importantes de este tipo de vegetación desde Canadá hasta el centro de México (APCP, 2010), albergando una gran biodiversidad a lo largo de su extensión. En México este tipo de vegetación ocupa, aproximadamente, 118,320 km2 (Challenger, 1998), las áreas más importantes se encuentran en extensas zonas del norte, principalmente en el Desierto Chihuahuense (Desmond y Montoya, 2006). Existe también una gran diversidad de condiciones en las que los pastizales naturales se presentan en el país, como por ejemplo los pastizales submontanos del Eje Neovolcánico Transversal (dispersos en zonas como la sierra nevada del sistema Popocatépetl-Iztaccíhuatl, el nevado de Colima, y el volcán Ajusco), entre otros (Challenger y Soberón, 2009). Otras zonas de pastizal húmedo se pueden encontrar en áreas de selva tropical con distribución restringida en Tabasco y Campeche (Challenger, 1998). Sin embargo, también existe una considerable extensión de pastizales inducidos dispersa por el país derivado de actividades ganaderas desde hace varias décadas (Challenger y Soberón, 2009).
Ante la presencia cada vez mayor de pastizales inducidos, el sistema de pastizales nativos del Desierto Chihuahuense, pese a su fisonomía y estructura aparentemente simples, representa una región de gran importancia para la conservación de los pastizales semiáridos, por la considerable riqueza de especies vegetales y presencia de pastos endémicos (Challenger y Soberón, 2009; Desmond y Montoya, 2006). Además, la extensión de estos pastizales se ha reconocido como relevante para una gran diversidad de taxones animales, que incluyen tanto aves de pastizal residentes (y algunas endémicas; e.g., Spizella wortheni) como migratorias, que son compartidas con Canadá y EUA (Macías-Duarte et al., 2009). Factores como la deforestación, la fragmentación del hábitat y el reemplazo de las comunidades de pastos nativos por paisajes agrícolas, han contribuido en el deterioro a gran escala de los pastizales, siendo a la vez los responsables de la disminución de las poblaciones de aves de pastizal (Brennan y Kuvlesky, 2005). Adicionalmente, se puede considerar que las zonas semiáridas del norte de México son de las menos susceptibles a la invasión de plantas no nativas debido a un menor desarrollo urbano, en contraste con los enormes centros urbanos en el centro y sur de México (del Val et al., 2015). Dentro de esta región, la gran extensión de pastizales naturales ha sido ocupada por varias especies de pastos no nativos que se han establecido desde principios del siglo XX (Melgoza-Castillo et al., 2014; Valenzuela-Núñez et al., 2009). Las zonas de pastizales invadidas por gramíneas no nativas proveen una menor cantidad de hábitats disponibles y, por lo tanto, menos recursos en comparación con aquellas zonas de pasto nativo que soportan un mayor conjunto de aves de pastizal (George et al., 2013). Por lo tanto, se vuelve necesario mantener y restaurar las condiciones originales de los pastizales para evitar la reducción de las poblaciones de aves residentes y migratorias de pastizal (Macías-Duarte et al., 2017).
En México, la introducción y expansión de especies de pastos exóticos obedece, probablemente, a la aplicación de políticas públicas impulsadas para promover la producción cárnica (Challenger y Soberón, 2009). En la historia del desarrollo agropecuario de México se pueden distinguir 3 periodos principales que implican cambios radicales en las prácticas ganaderas y que, por consiguiente, guiaron la transformación de los pastizales naturales (Chauvet-Sánchez, 1999). El primero, de 1930 a 1964 se caracterizó por la sustitución de las importaciones ganaderas para incentivar el mercado nacional, los intentos de erradicación de la tuberculosis y la fiebre aftosa en el ganado, así como la consolidación de la medicina veterinaria en México (Cervantes-Sánchez y Román de Carlos, 2001; Chauvet-Sánchez, 1999). El segundo periodo refleja la consolidación del mercado ganadero de exportación de cárnicos de manera constante, lo que dio lugar al auge ganadero nacional entre 1965-1981 con la ganaderización de la agricultura caracterizado por la sobreexplotación y agotamiento de los pastizales (Chauvet-Sánchez, 1999). Posteriormente, la eliminación de las políticas proteccionistas y el incremento de importación de cárnicos en el inicio de las políticas neoliberales dieron lugar al tercer periodo considerando la decadencia ganadera de 1982 hasta la actualidad (Chauvet-Sánchez, 1999). Cabe mencionar que durante este periodo surgen políticas de manejo y conservación de pastizales en varios sitos del Desierto Chihuahuense, como consecuencia del auge de la ciencia de la ecología y una mayor preocupación por la conservación de los recursos naturales (Hoth, 2012). Además, el cambio de modelo económico posterior a la firma del Tratado de Libre Comercio de América del Norte (TLCAN) produjo una menor presión sobre el pastizal al reducir el tiempo de cría del ganado para su engorde en EUA y posterior retorno a nuestro país como productos cárnicos procesados (Carrera-Chávez et al., 2010; Chauvet-Sánchez, 1999).
Estas etapas en el país enmarcan el tiempo en que la presión antropogénica pudo ser relevante para favorecer la introducción y expansión de pastos no nativos dentro del Desierto Chihuahuense, sobre todo porque estas especies de pastos, una vez establecidas, se expandieron con el tiempo ampliando su área de distribución geográfica (Guo et al., 2006). Comprender la dinámica temporal de cambio en la biodiversidad vegetal sobre las especies nativas es crucial para las estrategias de conservación a una escala regional. Estos cambios se han abordado a través de análisis de la diversidad beta (Qian, 2008; Qian y Ricklefs, 2006), entendida como los cambios en la composición de especies en ámbitos espacial, temporal y ambiental (Calderón-Patrón y Moreno, 2019).
Para evaluar el efecto de las especies de gramíneas introducidas sobre la composición de especies nativas en sistemas de pastizal (Eskelinen y Harrison, 2015; Joner et al., 2021; Szentes et al., 2012), la diversidad beta puede valorarse a través de índices de similitud con análisis basado en 2 componentes: el recambio y el anidamiento (Baselga, 2010; Calderón-Patrón y Moreno, 2019). Ecológicamente, estos componentes pueden reflejar una selección de especies por el ambiente o por procesos de dispersión, así como una relación entre las dinámicas de pérdida o colonización de especies (Baselga, 2010). Por ejemplo, en los pastizales centrales de Norteamérica, la diversidad beta mostró cambios en la composición de especies de aves a lo largo de un gradiente de disponibilidad de pastizales, lo que refleja la pérdida del hábitat original en el espacio tras varios años de modificación antropogénica (Pavlacky et al., 2022). Por otro lado, también se ha descrito que una partición de la diversidad total entre tipos de pastizales de una región puede mostrar el comportamiento del recambio total de la composición de aves de pastizal (Żmihorski et al., 2016).
La diversidad beta ofrece una herramienta adecuada para medir la disimilitud en la composición de especies debido a factores como el arreglo espacial, las restricciones espaciales o históricas, así como el gradiente de riqueza de especies (Baselga, 2010). Esto permite explorar los patrones de diversidad beta a lo largo del tiempo (Almeida-Neto et al., 2011), por lo que resulta valioso para evaluar el efecto de la introducción de pastos no nativos sobre la diversidad de aves de pastizal en el Desierto Chihuahuense.
Partiendo del contexto histórico-ambiental de la región antes descrito, se espera en el presente estudio que el recambio composicional de pastos nativos por pastos no nativos influya en la pérdida de especies de aves de pastizal. El objetivo de este estudio fue analizar el efecto de la introducción de pastos no nativos en la riqueza de aves de pastizal en el Desierto Chihuahuense, durante los 3 periodos descritos (1930-1964, 1965-1981 y 1982-2019) con distintas prácticas agropecuarias con la finalidad de evaluar si las políticas económicas han tenido una influencia sobre la diversidad de la biota asociada a esos ecosistemas. Especialmente, se pretende analizar si el incremento de los cambios en la gestión agrícola a largo plazo ha propiciado cambios en la diversidad de aves de pastizal (Chamberlain, 2000; Rosenberg et al., 2019). De manera específica, se plantearon los siguientes objetivos particulares: 1) obtener la diversidad beta regional de los pastos nativos y no nativos, así como de aves de pastizal en la región; 2) evaluar si existen diferencias en la composición de especies de los ensambles de aves, pastos nativos y no nativos, entre los 3 periodos propuestos; 3) evaluar la relación de los componentes de la diversidad beta (pérdida/ganancia de especies y si las especies son únicas o compartidas) de pastos nativos y no nativos sobre la composición de aves de pastizal; 4) probar si existe una correlación entre la prevalencia de pastos nativos y las aves de pastizal. Los resultados de esta investigación aportarán un enfoque novedoso para entender la influencia del cambio composicional en los pastizales (nativos/no nativos) sobre la diversidad de aves, resultado de las actividades de manejo planteadas en los planes de desarrollo agropecuario mexicano en el último siglo.
Materiales y métodos
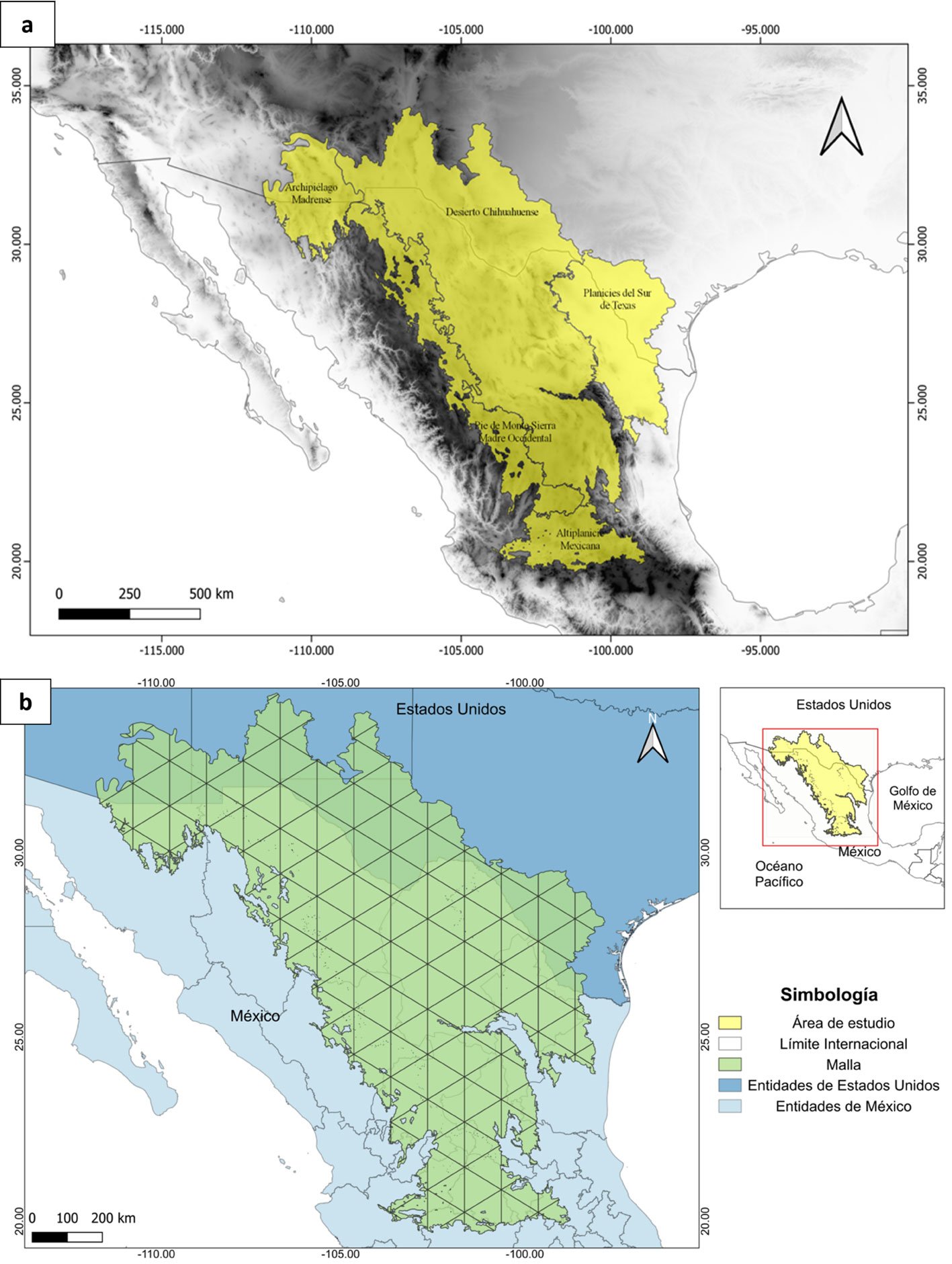
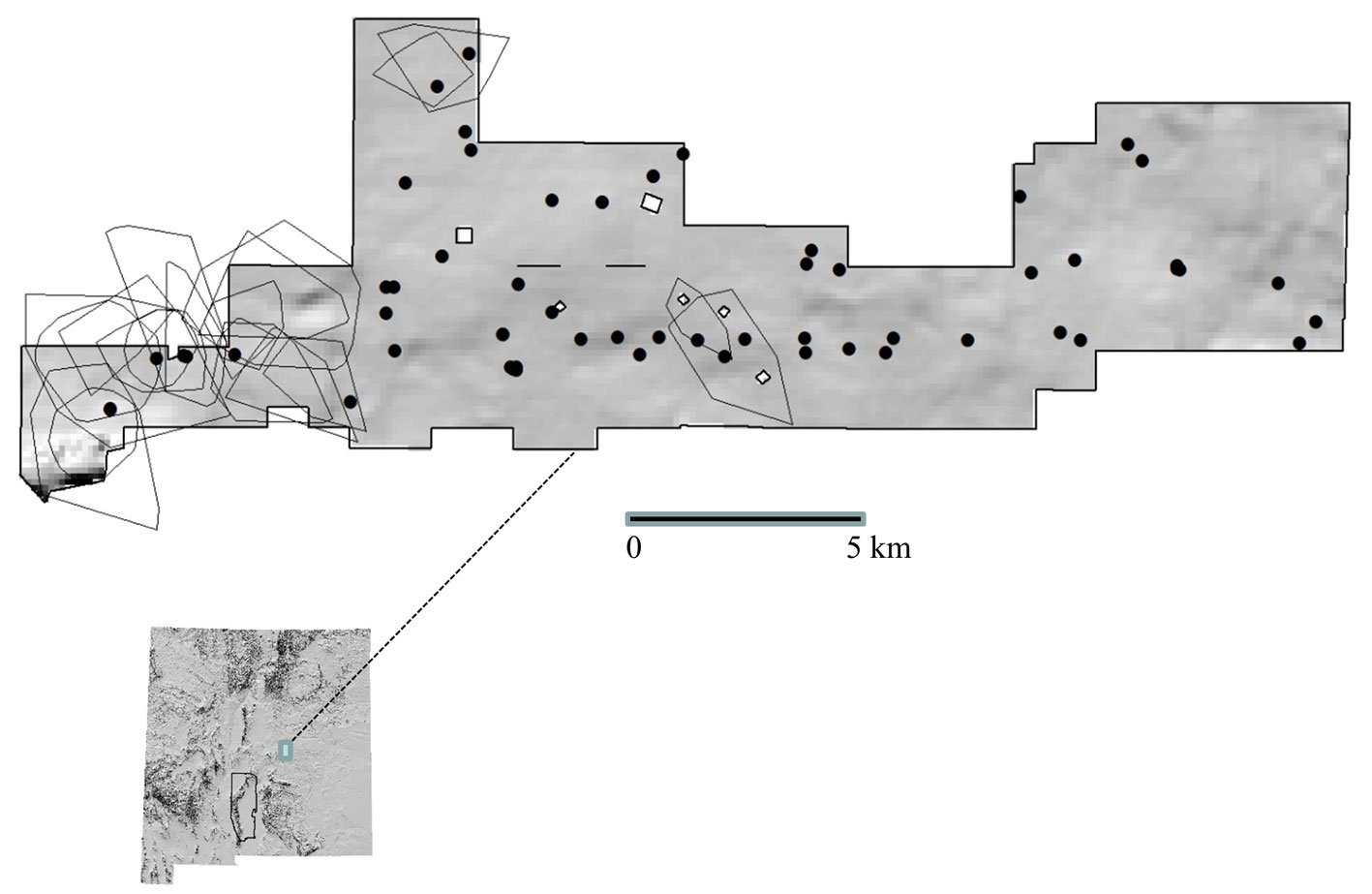
El área de estudio son los pastizales semiáridos que se extienden a lo largo del Desierto Chihuahuense en EUA y México, destacando que estos pastizales varían considerablemente en la composición de pastos nativos (González-Medrano, 2012). Además, en esta región se ha registrado la presencia de diversas especies de aves de pastizal residentes y migratorias (Levandoski y Panjabi, 2014; Ruvalcaba-Ortega y Panjabi, 2018; Vickery et al., 1999). La delimitación del área de estudio (fig. 1a) se estableció utilizando el sistema de información geográfica QGis 3.16, a partir de la unión de los polígonos correspondientes a las ecorregiones de la CCA (Comisión de Cooperación Ambiental) extraídas del mapa de la North American Terrestrial Ecoregions-Level III (Wiken et al., 2011): 1) Archipiélago Madrense, 2) Desierto Chihuahuense, 3) Planicies del Sur de Texas, 4) Pie de Monte Sierra Madre Occidental y 5) Altiplanicie Mexicana (fig. 1a). Estas ecorregiones fueron elegidas por la presencia de pastizales naturales y son consideradas como los pastizales cálidos de Norteamérica (Brown y Makings, 2014).
A partir del polígono del área de estudio, se construyó en QGis 3.16 una malla de celdas triangulares con una resolución geográfica de 1° de longitud por celda (aprox. 111.1 km2). La forma de las celdas fue elegida de entre varias alternativas, como cuadrícula (e.g., Rojas-Soto et al., 2003) o hexágonos (e.g., Sánchez-Ramos et al., 2018), optando por las celdas triangulares debido a que fueron las que mejor se ajustaron al área del polígono, y el nivel de resolución fue el óptimo para organizar los registros de especies para algunos de los análisis posteriores (fig. 1b).
Para obtener los registros de pastos nativos y no nativos (material suplementario: tabla 1S) se tomó como base el Catálogo de las gramíneas malezas nativas e introducidas de México (Melgoza-Castillo et al., 2014; Sánchez-Ken et al., 2012). Considerando a las especies presentes dentro del área del polígono, se seleccionaron 149 especies de pastos nativos y 60 especies de pastos no nativos. Para la nomenclatura taxonómica se siguió a The World Checklist of Vascular Plants(WCVP; Govaerts, 2022).
Para obtener los registros de las aves de pastizal de acuerdo con la literatura científica, se seleccionaron las especies que previamente han sido catalogadas como dependientes de este tipo de hábitat, ya sea por utilizar estos sitios como áreas reproductivas o importantes en sus rutas migratorias dentro del Desierto Chihuahuense (Levandoski y Panjabi, 2014; Ruvalcaba-Ortega y Panjabi, 2018; material suplementario: tabla 2S). En total, se obtuvieron los registros de 41 especies de aves (residentes y migratorias) asociadas a pastizales, que se determinaron siguiendo la nomenclatura de American Ornithological Society (AOS, Chesser et al., 2022).
Figura 1. a, Ecorregiones extraídas del mapa de North American Terrestrial Ecoregions —Level III (Wiken et al., 2011) con las que se definió el polígono del Desierto Chihuahuense (amarillo); b, rejilla de 1 minuto de arco (verde) sobre el área de estudio usada para realizar las matrices de presencia-ausencia.
Los datos primarios de presencia de las especies seleccionadas de aves, pastos nativos y no nativos, se extrajeron del explorador de registros de la Global Biodiversity Information Facility (GBIF, https://www.gbif.org), utilizando el polígono del área de estudio para delimitar espacialmente la consulta (los enlaces de descarga [DOI] de las consultas se encuentran en el material suplementario: tabla 1S). Se utilizó el intervalo de tiempo de 1930 a 2019, correspondiente a los 3 periodos del desarrollo agropecuario mexicano en el último siglo. Los datos de presencia disponibles permitieron contar con un registro histórico confiable del recambio espacial de las especies de la región. No fue posible establecer una base previa a este intervalo debido a la escasez de registros de pastos no nativos antes de 1930. Los registros obtenidos fueron reorganizados en 3 bases de datos (pastos nativos, pastos no nativos y aves de pastizal) construidas en un sistema de gestión de bases de datos (Microsoft Access®). En cada una de las bases de datos se realizó una depuración de las georreferencias repetidas y los registros mal georreferenciados con respecto al polígono, con el fin de eliminar las inconsistencias generadas por la captura de datos. Por último, se consideraron los intervalos de tiempo de 1930 a 1964, 1965 a 1981 y 1982 a 2019 para detectar los posibles cambios temporales en la composición de especies de aves, consecuencia de la introducción de especies de pastos no nativos.
Con la finalidad de evaluar la calidad del muestreo de los diferentes taxones, se generaron curvas de acumulación de las especies de aves, pastos nativos y pastos no nativos para cada periodo. Estas curvas representan el número acumulado de especies encontradas al aumentar el tamaño de muestra (e.g., el aumento del esfuerzo de muestreo en el tiempo para cada periodo). Los estimadores de las curvas de acumulación de especies por grupo taxonómico y periodo se obtuvieron a partir de las matrices de presencia-ausencia con el paquete EstimateS 9.1 (Colwell, 2013). Adicionalmente, se estimaron los estadísticos de Chao2 para evaluar la completitud del muestreo (Colwell, 2013).
Diversidad beta
Índices de disimilitud. Con los datos de presencia-ausencia, se exploraron los cambios en la composición de especies en las celdas durante los 3 periodos mediante el cálculo de la diversidad β de Sorensen (βsor aditivo) (Baselga, 2010). Esta métrica está compuesta por el recambio, medido por la diversidad β de Simpson (βsim), y por el anidamiento medido por la diversidad β anidada (βnes) (Baselga, 2010). El primer componente corresponde a cuántas especies de un sitio son reemplazadas por especies diferentes de otros sitios (valores de 0 = todas las especies son iguales y 1 = todas las especies son diferentes), mientras que el segundo componente se refiere a la pérdida gradual de especies de un sitio a otro (valores de 0 = pérdida total de especies y 1 = ganancia total de especies); es decir, un sitio es un subconjunto del otro (Baselga, 2010). Estos componentes se consideran un reflejo de los procesos de selección de especies por factores ambientales, así como una dinámica de pérdida o colonización de especies (Calderón-Patrón y Moreno, 2019; Vellend, 2010). Estos índices se calcularon en la paquetería “betapart” de R por medio de betapair a partir de las matrices de presencia-ausencia (Baselga et al., 2018). Con estos índices se midió la variación en la composición de especies de pastos nativos, pastos no nativos y aves de pastizal en cada uno de los 3 periodos (Baselga y Orme, 2012; Llanes-Quevedo et al., 2024).
Riqueza de aves de pastizal, pastos nativos y no nativos, y prevalencia de pastos nativos. Para evaluar las diferencias de la composición de especies de aves de pastizal y la composición de pastos nativos y no nativos entre los 3 periodos, se obtuvo la riqueza total de pastos nativos y no nativos por celda por periodo. Estas riquezas totales fueron utilizadas para calcular un índice de prevalencia de especies de pastos nativos (IPPN). Para cada celda por periodo, este índice sintetiza la riqueza de especies de pastos nativos y no nativos en una variable (adaptado de Legendre, 2019). El IPPN se calcula como: PN / (PN + PNN), donde PN es el número de especies de pastos nativos y PNN es el número de especies de pastos no nativos. Este índice varía de 0 (cuando todas las especies son pastos no nativos) a 1 (cuando todas las especies son pastos nativos). Las celdas sin registros de especies de pastos nativos o no nativos se eliminaron. Adicionalmente, para cada celda se obtuvo la riqueza total de aves de pastizal por periodo. Para evaluar las diferencias en la riqueza total de aves, pastos nativos, pastos no nativos, así como en el IPPN entre periodos, se utilizaron análisis de varianza de Kruskal-Wallis (KW). En cada análisis, el periodo con 3 niveles (1930-1964, 1965-1981 y 1982-2019) fue considerado como factor (Zar, 2010). Los análisis de KW se realizaron utilizando la función de kruskal.test() del paquete R (Kloke y McKean, 2015). En el caso de detectarse diferencias significativas, se realizaron pruebas “post-hoc” de Wilcoxon con la probabilidad ajustada de Bonferroni (α = 0.05/3 = 0.0166; Wright, 1992). Las posibles relaciones entre el IPPN y la riqueza total de aves de pastizal en cada uno de los 3 periodos, se analizaron con correlaciones de Spearman (rs) utilizando la función cor.test() del paquete R (Hollander y Wolfe, 1973). Estas variables fueron transformadas al arcoseno√ y log10+1, respectivamente (Zar, 2010). Adicionalmente, se visualizó y exploró la composición de cada conjunto de especies (i.e., aves de pastizal, pastos nativos y pastos no nativos) en los 3 periodos con análisis de escalamiento multidimensional no métrico (NMDS) con la función metaMDS() de la paquetería Vegan de R (Oksanen et al., 2013). Se utilizó la matriz transpuesta de presencia/ausencia (especies × sitios) de las especies de aves de pastizal, pastos nativos y pastos no nativos de cada uno de los 3 periodos con base en el coeficiente de distancia euclidiana (Palacio et al., 2020). La visualización por medio del análisis NMDS nos permite explorar y presentar, de manera más accesible, el efecto de los 3 periodos.
Resultados
El total de datos descargados que se obtuvieron para los 3 conjuntos de especies fue de 2,345,226 registros para aves de pastizal, 37,750 registros para pastos nativos y 8,815 para pastos no nativos. Al reorganizar, depurar, ajustar geográficamente y dividir los datos para cada uno de los 3 periodos correspondientes, se obtuvieron en total 328,809 registros de las 41 especies de aves de pastizal, 25,429 registros de las 149 especies de pastos nativos y 5,555 de las 60 especies de pastos no nativos. El número total de especies y conjunto de registros para cada uno de los periodos, así como la disposición espacial de los mismos, se puede consultar en el material suplementario (fig. 1S; tabla 3S). La mayor parte (93.33%) de los registros recolectados en GBIF correspondientes a las especies tanto de pastos nativos como no nativos, proceden de colecciones científicas y, en menor medida, de observaciones en campo o artículos de investigación (6.77%). Por el contrario, los registros de aves de pastizal proceden, en su mayor parte (97.16%), de observaciones de campo, seguido de investigaciones publicadas o de colecciones científicas (2.84%).
Se observaron patrones consistentes en la riqueza de especies en pastizales a lo largo de los 3 periodos analizados. En cada periodo, las curvas de acumulación indicaron que la mayor diversidad de especies se observó en pastos nativos, seguida de los pastos no nativos y las aves de pastizal (fig. 2). Los estimadores Chao2 respaldaron estos patrones con tamaños de muestra de diversidad distintos para cada conjunto. Para los pastos nativos, se estimó un registro de aproximadamente 94.7%, 90.4% y 99.8% de la riqueza total esperada en cada uno de los 3 periodos, respectivamente. Para pastos no nativos, las estimaciones fueron de 84.9%, 96.1%, y 96.9%, respectivamente. Para aves de pastizal, se estimó un registro de alrededor de 97.6% de la riqueza de especies en el primer periodo, mientras que, en los periodos siguientes, se estimó en 100% de la riqueza de especies, aunque esta estimación perfecta podría deberse a características particulares del estimador y al número fijo de especies a analizar.
El índice de disimilitud (βsor) mostró una tendencia en la disminución en los 3 ensambles a lo largo de los 3 periodos, lo que indica que aumentó la similitud entre las celdas que conforman el área de estudio para aves de pastizal, pastos nativos y no nativos (tabla 1). El índice de recambio (βsim) también disminuyó entre el primer periodo y el último, principalmente para las aves de pastizal (tabla 1). Por su parte, el índice de anidamiento (βsne) incrementó en todos los casos (tabla 1). Los resultados indican que, en la actualidad, las celdas comparten más especies entre sí y el recambio de especies ha disminuido para el último periodo.
Tabla 1
Índices de diversidad beta (βsor = βsim + βnes) obtenidos con beta.pair() de aves de pastizal, pastos nativos y pastos no nativos para cada periodo. x = Media y DE = desviación estándar.
Periodo
Aves de pastizal
Pastos nativos
Pastos no nativos
1930-1964
x
DE
x
DE
x
DE
βsor
0.74
0.21
0.83
0.16
0.81
0.23
βsim
0.50
0.36
0.66
0.29
0.70
0.38
βsne
0.24
0.27
0.16
0.22
0.12
0.20
1965-1981
x
DE
x
DE
x
DE
βsor
0.73
0.23
0.81
0.16
0.85
0.20
βsim
0.40
0.39
0.64
0.28
0.69
0.38
βsne
0.33
0.32
0.17
0.22
0.15
0.25
1982-2019
x
DE
x
DE
x
DE
βsor
0.35
0.21
0.74
0.18
0.77
0.19
βsim
0.09
0.11
0.52
0.28
0.55
0.36
βsne
0.26
0.23
0.23
0.23
0.22
0.27
Figura 2. Curvas de acumulación de especies de aves de pastizal (azul), pastos nativos (verde) y pastos no nativos (rojo) para los periodos de a) 1930-1964, b) 1965-1981 y c) 1982-2019.
Los análisis revelaron diferencias significativas de la riqueza de aves de pastizal (H = 196.24, g.l. = 2, p = 0.0001), pastos nativos (H = 35.78, g.l. = 2, p = 0.0001), pastos no nativos (H = 50.74, g.l. = 2, p = 0.0009) y el IPPN (H = 47.40, g.l. = 2, p = 0.0005) entre periodos (fig. 3). Estos hallazgos indican un efecto significativo a lo largo del tiempo en cada uno de los grupos taxonómicos evaluados, lo que refleja posibles cambios en la composición y estructura de las comunidades biológicas en respuesta a factores de manejo específicos o ambientales de cada período. La riqueza de aves de pastizal del periodo 1982-2019 fue significativamente mayor (p < 0.0001) con respecto a los periodos 1930-1964 y 1965-1981. Similarmente, la riqueza de pastos nativos y no nativos del periodo 1982-2019 fue significativamente mayor en comparación con aquella del periodo 1930-1964 (p = 0.0001) y 1965-1981 (p = 0.0013). Por otro el contrario, el IPPN del periodo 1982-2019 fue significativamente menor en comparación con el del periodo de 1930-1964 (p < 0.0001) y 1965-1981 (p = 0.0003).
Figura 3. Diagramas de caja de la riqueza de aves de pastizal, pastos nativos, pastos no nativos y el índice de prevalencia de pastos nativos (IPPN) para cada periodo. Periodos identificados con letras diferentes indican diferencias significativas (p ≤ 0.0166) con el resto a partir de pruebas “post-hoc” de Wilcoxon (ver detalles en Materiales y métodos).
Hubo correlaciones negativas débiles entre el IPPN y la riqueza de aves de pastizal en los periodos de 1930-1964 (rs = -0.1553, p = 0.0400) y 1965-1981 (rs = -0.1784, p = 0.0100). Sin embargo, no se encontró una correlación significativa entre ambas variables (rs = 0.0321, p = 0.6600) en el periodo de 1982-2019. La proyección de IPPN sobre la región muestra que durante el periodo de 1930-1964, el ensamble regional de pastos estaba conformado principalmente por pastos nativos, en contraste con una reducida prevalencia de pastos nativos para el periodo de 1982-2019 (fig. 4).
Finalmente, los NMDS revelaron una variación en la composición y riqueza regional del ensamble de especies tanto de aves como de pastos nativos y no nativos entre los periodos estudiados (fig. 5). Se observó una discontinuidad en la composición de especies de aves de pastizal a lo largo del tiempo, con una marcada contracción durante el periodo de 1965-1981 y una distribución más dispersa en el periodo de 1982-2019, indicando cambios abruptos en la estructura del ensamble (fig. 5a). En contraste, se observó un cambio gradual en la composición de especies de pastos nativos, con un incremento en la distribución del ensamble en varios sitios de la región con una posible contracción entre periodos (fig. 5b). Por último, se observó un marcado aumento en la composición del ensamble de pastos no nativos, especialmente notable durante el periodo de 1982-2019 (fig. 5c). Este patrón de cambio confirma la expansión significativa del ensamble de pastos no nativos en comparación con los periodos anteriores, en el mismo sentido que la proyección del IPPN para el último periodo (fig. 4). Estos resultados sugieren que el periodo de 1982-2019 muestra cambios en el ensamble de especies respecto a los otros periodos en términos del IPPN y la riqueza de especies. Sin embargo, esta tendencia no se mantuvo de manera consistente en el último periodo.
Discusión
Este estudio representa un acercamiento a escala regional sobre la influencia de los pastos no nativos en los pastizales del Desierto Chihuahuense, sugiriendo cambios en la composición general de las aves de pastizal, atribuibles al considerable cambio en la riqueza de especies y composición taxonómica de los pastos nativos influenciados por los procesos agropecuarios a lo largo del tiempo. Esto no involucra únicamente esta región, ya que debido a su conectividad, es fundamental para las aves migratorias provenientes de las praderas templadas y de pastos mixtos de las Grandes Llanuras Centrales de Norteamérica (Comer et al., 2018). Aunado a lo anterior, en esta región se han identificado procesos de diversificación y dispersión de diversos grupos de vertebrados terrestres y de plantas vasculares (Savage, 1982). En cuanto a las dinámicas ecológicas, estos hábitats también son importantes para diversas especies de animales, como el perrito de las praderas de cola negra (Cynomys ludovicianus), considerada especie clave en la dinámica de pastizales del Desierto Chihuahuense (Detling, 2013). Nuestro estudio aporta evidencia sobre cómo la implementación histórica de políticas públicas pudo modelar de forma positiva la diversidad beta de pastos no nativos y negativamente para los pastos nativos.
Los índices de disimilitud de diversidad βsor (tabla 1) revelaron que los pastos nativos y no nativos están caracterizados por un marcado reemplazo, que se explica por una ausencia de anidamiento a lo largo de los 3 periodos analizados. En cambio, las aves de pastizal cambian este patrón solo en el último periodo al aumentar el anidamiento. Este patrón sugiere que el impacto de las diferentes políticas de aprovechamiento de pastizales a lo largo del tiempo puede influir en la diversidad beta a nivel regional. El reemplazo de especies de pastos nativos y no nativos puede estar más vinculado a eventos de manejo que varían de manera diferencial en cada periodo (Chauvet-Sánchez, 1999; Hoth, 2012), además de otros procesos como los gradientes ambientales (Baselga, 2010).
Figura 4. Proyección del índice de prevalencia de pastos nativos (IPPN) sobre los sitios para cada periodo. La escala de colores corresponde a los valores de IPPN. Valores de 0 indican ensambles dominados completamente por pastos no nativos y 1 cuando dominan completamente los pastos nativos.
Por otro lado, los valores constantes de reemplazo en los 2 tipos de pastos a lo largo del tiempo, no necesariamente se asocian con procesos de homogeneización biótica, debido a la pérdida de especies de plantas nativas frente a las no nativas (Alignier y Baudry, 2016). Aunque esto puede ocurrir en un nivel alfa local, el nivel beta regional es imperceptible, como puede ser nuestro caso. Por ejemplo, se ha observado que los cambios en la diversidad taxonómica de pastizales a nivel alfa local se asocian con la presión por pastoreo, mientras que un nivel beta regional, la diversidad filogenética se asocia con gradientes ambientales, los cuales corresponden a procesos históricos y evolutivos (Wang et al., 2019). Aunque la diversidad filogenética no fue objeto de nuestro estudio, esta observación nos ayuda a entender por qué en nuestros conjuntos de especies no se logran apreciar cambios considerables a lo largo del tiempo, puesto que los periodos de estudio son relativamente cortos históricamente. En el caso de las aves de pastizal, el patrón de diversidad beta sugiere una homogenización de las comunidades, atribuible a la disminución de la disimilitud en el último periodo. Esto indica que las diferencias en la riqueza de especies entre los sitios también han disminuido, lo cual puede atribuirse al incremento del patrón de anidamiento a lo largo del tiempo detectado desde el periodo de 1965-1981. Estas evidencias pueden estar influenciadas por el incremento de la presión debido a la expansión de la ganadería a partir de 1965 (Chauvet-Sánchez, 1999; Hoth, 2012), ya que desde entonces existía un considerable sobrepastoreo de los pastizales del norte del país (CFAN, 1965).
Figura 5. Análisis de escalamiento multidimensional no métrico (NMDS) de la composición de especies de a) aves de pastizal, b) pastos nativos y c) pastos no nativos. Los polígonos dentro de cada gráfico corresponden al espacio que ocupa la composición de especies en 2 dimensiones.
Periodo de 1930-1964
Nuestros resultados evidenciaron una mayor estabilidad en la riqueza estimada de aves de pastizal con respecto de los pastos nativos y no nativos en la región durante este periodo. Esto puede atribuirse en gran parte al amplio conocimiento taxonómico de las aves en México (Navarro-Sigüenza et al., 2014), pero también a que la selección de especies de aves de nuestro universo de análisis fue restringido y limitado a aquellas que dependen del pastizal para eventos clave de su ciclo de vida. Por otro lado, los pastos nativos tuvieron una mejor representación con respecto a los pastos no nativos. Al examinar la proyección del IPPN y los NMDS (figs. 4, 5b), es claro que la región estaba predominantemente conformada por ensambles de pastos nativos. La poca representación del ensamble de pastos no nativos pudo deberse a que algunas de las introducciones se dieron desde inicios del siglo XX, al ser originalmente introducidas en el suroeste de EUA (Herrera, 2001), por lo que su presencia pudo haberse pasado por alto en este periodo.
Se debe tener en cuenta que durante las décadas de 1930 y 1940, los esfuerzos gubernamentales estaban más centrados en establecer la tenencia de la tierra entre propiedad comunal y propiedad privada (González-Navarro, 1969), así como en recuperar los niveles de producción ganadera previos a la revolución de 1910 (de S. Lopes, 2008). La correlación significativa del IPPN y la riqueza de aves de pastizal, aunque baja y negativa, sugiere que el periodo en que se tiene la mayor prevalencia por parte de pastos nativos influyó poco sobre la riqueza de aves de pastizal del Desierto Chihuahuense. Se debe tener en cuenta que se introdujeron especies de pastos de manera directa en la región a partir de 1956 con la intención de mejorar la producción de forraje para el ganado por parte de la Oficina de Estudios Especiales del Gobierno de México (Ortega et al., 2013). Como incentivo adicional, durante este periodo se logró la consolidación de la práctica veterinaria pecuaria a nivel nacional con la erradicación de la fiebre aftosa en 1956 (Chauvet-Sánchez, 1999). Estos factores serían fundamentales para estimular la producción cárnica dentro de zonas de pastizales y serían la antesala de la expansión de pastos no nativos dentro del Desierto Chihuahuense.
Periodo de 1965-1981
La riqueza de especies estimada para el periodo 1965-1981 muestra que es cuando se detecta una mayor presencia de pastos no nativos en el Desierto Chihuahuense. Sin embargo, esto se refleja mínimamente en el IPPN (fig. 4) y en los análisis NMDS correspondientes a pastos no nativos. No obstante, se pudo observar que el ensamble de pastos en la región ya no está conformado únicamente por pastos nativos en todos los sitios (fig. 5b, c). Por su parte, el ensamble de aves de pastizal exhibe a nivel regional una contracción de su NMDS (fig. 5a), lo que implica una menor riqueza de aves de pastizal en varios sitios con respecto al periodo anterior.
Lo antes mencionado y la correlación negativa entre la riqueza de las aves de pastizal con el IPPN sugiere que durante el auge ganadero entre 1965 y 1981, se desencadenó un efecto que favoreció en algunos sitios la presencia de pastos no nativos, mientras que para las aves sucedió lo contrario. Esto coincide con lo observado en los pastizales centrales de EUA, donde la disminución de la diversidad de aves de tierras de cultivo se correlacionó con cambios significativos en las prácticas agrícolas entre 1966 y 1980 (Stanton et al., 2018). Además, por monitoreos estandarizados en gradientes de pastizal nativo en EUA (Wiens, 1973), se sabe que el grupo de aves de pastizal presentó la mayor pérdida poblacional atribuible al incremento de la agricultura y la urbanización en zonas de pastizal desde la década de 1970 (Rosenberg et al., 2019). Durante este periodo, varias zonas de pastizal sobrepastoreadas en el Desierto Chihuahuense presentaban erosión leve o avanzada, lo que provocó pérdida de hábitat (CFAN, 1965; Royo et al., 2005). En los últimos años de este periodo, comienzan a aparecer investigaciones que analizan el impacto de estas especies como malezas nocivas sobre el hábitat original (Espinosa-García y Villaseñor, 2017). Esto pudo ser una respuesta a la permisiva explotación de los pastizales durante la ganaderización de la agricultura en varias zonas de pastizal para la producción cárnica (Chauvet-Sánchez, 1999), incluido el Desierto Chihuahuense. Los pastos nativos parecen responder de manera diferente, ya que los NMDS (fig. 5b) muestran que con el tiempo se tiene una mejor representación del ensamble de especies al ir incrementado entre las 2 dimensiones, pese a que la producción cárnica durante el auge ganadero se mantuvo constante hasta la apertura comercial con el TLCAN en 1992 (Chauvet-Sánchez, 1999). Estos pueden ser efectos a largo plazo, atribuibles al auge ganadero (Chauvet-Sánchez, 1999), cuyo uso como recurso pudo haber sido generalizado y no exclusivo para el consumo del ganado.
Periodo de 1982-2019
Finalmente, la diversidad de especies observada en el periodo de 1982-2019 siguió la tendencia del periodo anterior siendo más alta en pastos nativos, seguida de pastos no nativos y aves de pastizal. Sin embargo, los pastos no nativos empiezan a alcanzar una estabilidad para el final del periodo, lo que puede ser resultado de la colaboración entre EUA y México para la conservación de los pastizales, así como del mejoramiento de las prácticas ganaderas dentro del Desierto Chihuahuense (Hoth, 2012). Pese a esto, la riqueza de pastos no nativos ha llegado a representar poco más de la tercera parte de la totalidad de especies de pastos que se pueden encontrar dentro del Desierto Chihuahuense.
Las curvas para los 3 periodos dentro de toda la región en general revelan el importante esfuerzo de registro taxonómico que se viene llevando desde principios del siglo XX, no solo en la región sino en todo territorio mexicano (Llorente-Bousquets y Ocegueda, 2008). Todas las pruebas “post-hoc” evidencian las discrepancias de este periodo con respecto a los anteriores en términos de riqueza de los 3 conjuntos de especies, así como del IPPN. No obstante, es importante considerar, sobre todo para las aves de pastizal, la mayor densidad de registros en la porción territorial norteamericana lo que pudo influir en que este periodo difiera en comparación con los anteriores. Sin embargo, la ausencia de correlación entre la riqueza de aves y el IPPN exhibe que la representación del ensamble de aves de pastizal no es una variable relacionada con el comportamiento del ensamble de pastos nativos. Esto sugiere la ausencia de un efecto sobre la riqueza de aves por la expansión de los pastos no nativos, aunque se ha detectado que los ensambles de aves presentan frecuentemente cambios en la abundancia relativa de las especies, pero no en la riqueza, en zonas homogenizadas por pastos no nativos (Harris et al., 2019), especialmente relacionados al reordenamiento de las abundancias y no al recambio en la riqueza de especies (Jones et al., 2017; Rosenberg et al., 2019).
La falta de correlación entre la riqueza de aves y el IPPN podría deberse a: 1) que las especies de aves tienen una mayor capacidad de persistir en cualquiera de las configuraciones de pastizal presentes dentro de la región (Ellis-Felege et al., 2013); 2) que su distribución es más amplia de la que abarca nuestra resolución, debido a que la región tiene conectividad con pastizales norteamericanos (Vickery et al., 1999) y 3) finalmente porque son un grupo biológico con alta capacidad de movilidad dentro de la región (Comer et al., 2018). La proyección del IPPN (fig. 4) y los NMDS correspondientes a pastos nativos y no nativos (fig. 5b, c) sugieren que el ensamble de pastos no nativos se encuentra en prácticamente los mismos sitios de pastizal nativo. Es posible que los pastos no nativos se hayan visto influenciados positivamente debido a que la constante degradación de los pastizales en la región por sobrepastoreo del periodo anterior, contribuyó a la expansión regional y que en parte, nuestros resultados son un reflejo de la enorme fragmentación del hábitat (Hoth, 2012).
Por otro lado, los esfuerzos de conservación en varias zonas de pastizal del Desierto Chihuahuense durante el periodo más reciente (APCP, 2010; Aguirre et al., 2007), en un inicio no fueron especialmente selectivos sobre las especies de pastos que conforman el hábitat, pudiendo contribuir al incremento en la riqueza de especies, tanto de pastos nativos como no nativos. En este sentido, una parte considerable del ensamble de pastos no nativos presentes en la región pudieron originarse de pastizales nativos degradados o bien de la deforestación de zonas con arbustos y árboles (Buisson et al., 2022). Por lo tanto, las especies de pastos no nativos se establecieron aleatoriamente en toda la región a partir de tierras agrícolas (Rzedowski y Calderón, 1990; Rzedowski et al., 1995). Esta expansión produjo ensambles de pastos más reducidos en cada sitio en comparación con zonas con pastos nativos (Flanders et al., 2006), pero que finalmente, en la actualidad se deben considerar como parte de la flora de la región (Brown y Makings, 2014).
Nuestros resultados sugieren que la introducción de especies de pastos no nativos no fue un evento único y que el aumento en la presencia de estas especies, con el paso del tiempo, fue en los mismos sitios en los que se detectaron los pastos nativos, lo que denota la necesidad de priorizar los esfuerzos de conservación selectivos para los pastos nativos. Además, se requiere analizar si la cobertura de otros tipos de hábitat, como el incremento de matorrales espinosos sobre la configuración de pastizales en el Desierto Chihuahuense, ha cambiado dentro de la región.
Nuestros resultados indican cambios en la diversidad beta de aves de pastizal, de pastos nativos y de pastos no nativos a nivel regional. Considerando la resolución espacial y temporal utilizada en este estudio, nuestros análisis sugieren que estos cambios pueden ser atribuibles a las diferentes políticas económicas de cada periodo. No obstante, se destaca la importancia de analizar a las especies de pastos no nativos en un nivel de diversidad alfa, con énfasis en una posible respuesta de los ensambles de aves de pastizal, posiblemente asociada a las particularidades de cada comunidad. También subrayamos la necesidad de identificar sitios prioritarios para la gestión y conservación selectiva de pastos nativos en zonas de pastizal con el objetivo de mitigar posibles alteraciones en el ensamble de aves de pastizal en la región.
Agradecimientos
El presente artículo forma parte de los requisitos para la obtención del grado de Doctor de MP-M en el programa de posgrado de Ciencias Biológicas y de la Salud de la Universidad Autónoma Metropolitana, quien cuenta con una beca del Conahcyt (CVU 718731). Agradecemos el apoyo financiero del programa PAPIIT-UNAM IN214621 (a AGN-S) y Proyecto UAM 143.05.27 (a PLV). A Alejandro Gordillo, Alexander Llanes, Ronald Fernández, Reinhard Matadamas, Rodolfo Flores, Itzel Estrada, José Zazueta, Carlos Lara y dos revisores anónimos, por su apoyo logístico y valiosos comentarios durante el desarrollo de esta investigación. LES-R agradece a DGAPA-UNAM por la beca posdoctoral.
Referencias
Aguirre, C., Hoth, J. y Lafón, A. (Eds.). (2007). Estrategia para la Conservación de Pastizales del Desierto Chihuahuense (ECOPAD). Repórte técnico. https://doi.org/10.13140/RG.2. 2.36081.74089
Alignier, A. y Baudry, J. (2016). Is plant temporal beta diversity of field margins related to changes in management practices? Acta Oecologica, 75, 1–7. http://dx.doi.org/10.1016/j.actao. 2016.06.008
Almeida-Neto, M., Frensel D. M. B. y Ulrich, W. (2011). Rethinking the relationship between nestedness and beta diversity: a comment on Baselga (2010). Global Ecology and Biogeography, 21, 772–777. https://doi.org/10.1111/j. 1466-8238.2011.00709.x
APCP (Áreas Prioritarias de Conservación Pastizal). (2010). Comisión para Cooperación Ambiental (CCA). http://www.cec.org/es/atlas-ambiental-de-america-del-norte/areas-pri oritarias-de-conservacion-pastizales-2010/
Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, 19, 134–143. https://doi.org/10.1111/j.1466-82 38.2009.00490.x
Baselga, A. y Orme, C. D. L. (2012). betapart: an R package for the study of beta diversity. Methods in Ecology and Evolution, 3, 808–812. https://doi.org/10.1111/j.2041-210X.2012.00224.x
Baselga, A., Orme, D., Villeger, S., De Bortoli, J., Leprieur, F., Logez, M. et al. (2018). Package ‘betapart’: Partitioning Beta Diversity into Turnover and Nestedness Components. Ver 1.5. 1. CRAN.
Brennan, L. A. y Kuvlesky Jr., W. P. (2005). North American grassland birds: an unfolding conservation crisis? The Journal of Wildlife Management, 69, 1–13. https://doi.org/10.2193/0022-541X(2005)069<0001:NAGBAU>2.0.CO;2
Brown, D. E. y Makings, E. (2014). Desert plants. Tucson:The University of Arizona for the Boyce Thompson Arboretum.
Buisson, E., Archibald, S., Fidelis, A. y Suding, K. N. (2022). Ancient grasslands guide ambitious goals in grassland restoration. Science, 377, 594–598. https://doi.org/10.1126/science.abo4605
Calderón-Patrón, J. M. y Moreno, C. E. (2019). Diversidad beta basada en índices de disimilitud: Su partición en componentes de recambio y diferencias en riqueza. En C. E. Moreno (Ed.), La biodiversidad en un mundo cambiante: fundamentos teóricos y metodológicos para su estudio (pp. 203–222). Ciudad de México: Universidad Autónoma del Estado de Hidalgo/ Libermex.
Guo, B., Cortázar-Martínez, A. y Ayala-Garay, A. V. (2010). Competitividad de la ganadería bovina de carne en México. En B. A. Cavallotti-Vázquez, C. F. Marcof-Álvarez y B. Ramírez-Valverde (Eds.), Los grandes retos para la ganadería: hambre, pobreza y crisis ambiental. (pp. 3–14). Estado de México: Universidad Autónoma Chapingo. México.
Cervantes-Sánchez, J. M. y Román de Carlos, A. M. (2001). Consecuencias de la epizootia de la fiebre aftosa en la medicina veterinaria y la zootecnia mexicanas (1946–1955). Imagen Veterinaria, 1, 17–20.
CFAN (Consejo de Fomento Agropecuario del Norte de México). (1965). Estudio integral preliminar sobre la ganadería de la zona norte de la República Mexicana. Tomo IV. Ciudad de México: Consejo de Fomento Agropecuario del Norte de México/ COPARMEX.
Challenger, A. (1998). Utilización y conservación de los ecosistemas terrestres de México: pasado, presente y futuro. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Instituto de Biología, UNAM/ Agrupación Sierra Madre. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Challenger, A. y Soberón, J. (2009). Los ecosistemas terrestres. En J. Soberón, G. Halffter y J. Llorente-Bousquets (Eds.), Capital natural de México, Vol. 1. Conocimiento actual de la biodiversidad (pp. 88–105). Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Chamberlain, D. E., Fuller, R. J., Bunce, R. G., Duckworth, J. C., y Shrubb, M. (2000). Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. Journal of Applied Ecology, 37, 771–788. https://doi.org/10.1046/j.1365-2664.2000.00548.x
Chaneton, E. J., Perelman, S. B., Omacini, M. y León, R. J. (2002). Grazing, environmental heterogeneity, and alien plant invasions in temperate Pampa grasslands. Biological Invasions, 4, 7–24.
Chauvet-Sánchez, M. (1999). La ganadería bovina de carne en México: del auge a la crisis. México D.F.: Universidad Autónoma Metropolitana.
Chesser, R. T., Billerman, S. M., Burns, K. J., Cicero, J. C., Dunn, L., Hernández-Baños, B. E. et al. (2022). Check-list of North American birds. American Ornithological Society. Recuperado 19 diciembre, 2022 de: https://checklist.americanornithology.org/taxa/
Colwell, R. K. (2013). Estimates: statistical estimation of species richness and shared species from samples. Version 9. User’s Guide and Application.
Comer, P. J., Hak, J. C., Kindscher, K., Muldavin, E. y Singhurst, J. (2018). Continent-scale landscape conservation design for temperate grasslands of the Great Plains and Chihuahuan Desert. Natural Areas Journal, 38, 196–211. https://doi.org/10.3375/043.038.0209
Davis, M. A. (2003). Biotic globalization: does competition from introduced species threaten biodiversity? Bioscience, 53, 481–489. https://doi.org/10.1641/0006-3568(2003)053[0481: BGDCFI]2.0.CO;2
del Val, E., Balvanera, P., Castellarini, F., Espinosa-García, F. J., Murguía, M. y Pacheco, C. (2015). Identifying areas of high invasion risk: a general model and an application to Mexico. Revista Mexicana de Biodiversidad, 86, 208–216. https://doi.org/10.7550/rmb.44743
Desmond, M. y Montoya, J. A. (2006). Status and distribution of Chihuahuan Desert grasslands in the United States and Mexico. En X. Basurto y D. Hadley (Eds.), Grasslands ecosystems, endangered species, and sustainable ranching in the Mexico-US borderlands: Conference proceedings. RMRS-P-40 (pp. 17–25). Fort Collins, CO: US Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Detling, J. K. (2013). Do prairie dog compete with livestock? En J. Hoogland (Ed.), Conservation of the black-tailed prairie dog: saving North America’s western grasslands (pp. 65–88). Washington D.C.: Island Press.
Ellis-Felege, S. N., Dixon, C. S. y Wilson, S. D. (2013). Impacts and management of invasive cool-season grasses in the Northern Great Plains: challenges and opportunities for wildlife. Wildlife Society Bulletin, 37, 510–516. https://doi.org/10.1002/wsb.321
Eskelinen, A. y Harrison, S. (2015). Erosion of beta diversity under interacting global change impacts in a semi-arid grassland. Journal of Ecology, 103, 397–407. https://doi.org/10.1111/1365-2745.12360
Espinosa-García, F. J. y Villaseñor, J. L. (2017). Biodiversity, distribution, ecology and management of non-native weeds in Mexico: a review. Revista Mexicana de Biodiversidad, 88, 76–96. https://doi.org/10.1016/j.rmb.2017.10.010
Flanders, A. A., Kuvlesky Jr, W. P., Ruthven III, D. C., Zaiglin, R. E., Bingham, R. L., Fulbright, T. E. et al. (2006). Effects of invasive exotic grasses on south Texas rangeland breeding birds. The Auk, 123, 171–182. https://doi.org/10.1093/auk/ 123.1.171
George, A. D., O’Connell, T. J., Hickman, K. R. y Leslie Jr, D. M. (2013). Food availability in exotic grasslands: a potential mechanism for depauperate breeding assemblages. The Wilson Journal of Ornithology, 125, 526–533. https://doi.org/10.1676/13-003.1
Gibson, D. J. (2009). Grasses and grassland ecology. Nueva York: Oxford University Press.
González-Medrano, F. (2012). Las zonas áridas y semiáridas de México y su vegetación. México D.F.: Instituto Nacional de Ecología.
González-Navarro, M. (1969). La tenencia de la tierra en México. Cahiers du monde hispanique et luso-brésilien, 12, 115–134.
Govaerts, R. (2022). The World Checklist of Vascular Plants (WCVP). En O. Bánki, Y. Roskov, M. Döring, G. Ower, L. Vandepitte, D. Hobern et al. (Eds.), Catalogue of Life Checklist (10.0). Kew: The Royal Botanic Gardens. https://doi.org/10.48580/dfqt-4nz
Guo, Q., Qian, H., Ricklefs, R. E. y Xi, W. (2006). Distributions of exotic plants in eastern Asia and North America. Ecology Letters, 9, 827–834. https://doi.org/10.1111/j.1461-0248.2006. 00938.x
Hamilton, J. G., Holzapfel, C. y Mahall, B. E. (1999). Coexistence and interference between a native perennial grass and non-native annual grasses in California. Oecologia, 121, 518–526. https://doi.org/10.1007/s004420050958
Harris, M. A., Greve, M., Voysey, M. D. y Jamison, S. L. (2019). Changes in bird assemblages because of vegetation homogenisation in communal livestock systems. Koedoe: African Protected Area Conservation and Science, 61, 1–18. https://doi.org/10.4102/koedoe.v61i1.1543
Hejda, M., Pyšek, P. y Jarošík, V. (2009). Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 97, 393–403. https://doi.org/10.1111/j.1365-2745.2009.01480.x
Herrera, Y. (2001). Las gramíneas de Durango. Durango, México: Instituto Politécnico Nacional/ Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Editorial Filo de Agua.
Hollander, M. y Wolfe, D. A. (1973). Nonparametric statistical methods. New York: John Wiley and Sons.
Hoth, J. (2012). Buenas prácticas ganaderas en México. Montreal: Comisión para la Cooperación Ambiental de América del Norte (CCA).
Joner, F., Giehl, E. L. y Pillar, V. D. (2021). Functional and taxonomic alpha and beta diversity responses to burning grasslands in southern Brazil. Journal of Vegetation Science, 32, e13060. https://doi.org/10.1111/jvs.13060
Jones, S. K., Ripplinger, J. y Collins, S. L. (2017). Species reordering, not changes in richness, drives long-term dynamics in grassland communities. Ecology Letters, 20, 1556–1565. https://doi.org/10.1111/ele.12864
Kloke, J., McKean, J. W. y McKean, J. W. (2015). Nonparametric statistical methods using R. Boca Raton: CRC Press.
Legendre, P. (2019). A temporal beta-diversity index to identify sites that have changed in exceptional ways in space–time surveys. Ecology and Evolution, 9, 3500–3514. https://doi.org/10.1002/ece3.4984
Levandoski, G. y Panjabi, A. (2014). Las aves de pastizal. En Conabio (Ed.), La biodiversidad en Chihuahua: estudio de estado (pp. 128–134). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Llanes-Quevedo, A., Sánchez-Ramos, L. E. y Navarro-Sigüenza, A. G. (2024). Patrones históricos y actuales de diversidad y relaciones biogeográficas de la avifauna residente de los bosques tropicales de México. Revista Mexicana de Biodiversidad, 95, e955341. https://doi.org/10.22201/ib.2007 8706e.2024.95.5341
Llorente-Bousquets, J. y Ocegueda, S. (2008). Estado del conocimiento de la biota. En Conabio (Ed.), Capital natural de México, Vol. I. Conocimiento actual de la biodiversidad (pp. 283–322). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Macías-Duarte, A., Montoya, A. B., Méndez-González, C. E., Rodríguez-Salazar, J. R., Hunt, W. G. y Krannitz, P. G. (2009). Factors influencing habitat use by migratory grassland birds in the state of Chihuahua, Mexico. The Auk, 126, 896–905. https://doi.org/10.1525/auk.2009.08251
Macías-Duarte, A., Panjabi, A. O., Strasser, E. H., Levandoski, G. J., Ruvalcaba-Ortega, I., Doherty, P. F. et al. (2017). Winter survival of North American grassland birds is driven by weather and grassland condition in the Chihuahuan Desert. Journal of Field Ornithology, 88, 374–386. https://doi.org/10.1111/jofo.12226
Melgoza-Castillo, A., Balandrán Valladares, M. I., Mata-González, R. y Pinedo Álvarez, C. (2014). Biología del pasto rosado Melinis repens (Willd.) e implicaciones para su aprovechamiento o control: Revisión. Revista Mexicana de Ciencias Pecuarias, 5, 429–442.
Navarro-Sigüenza, A. G., Rebón-Gallardo, M. F., Gordillo-Martínez, A., Townsend-Peterson, A., Berlanga-García, H. y Sánchez-González, L. A. (2014). Biodiversidad de aves en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), S476–S495. https://doi.org/10.7550/rmb.41882
Ortega, S. J. A., Ibarra-Flores, F. A., Melgoza, A., González-Valenzuela, E. A., Martin-Rivera, M. H., Ávila-Curiel, J. M. et al. (2013). Exotic grasses and wildlife in northern Mexico. Wildlife Society Bulletin, 37, 537–545. https://doi.org/10.1002/wsb.325
Palacio, F. X., Apodaca, M. J. y Crisci, J. V. (2020). Análisis multivariado para datos biológicos: teoría y su aplicación utilizando el lenguaje R. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara.
Pavlacky Jr, D. C., Green, A. W., George, T. L., Iovanna, R., Bartuszevige, A. M., Correll, M. D. et al. (2022). Landscape-scale conservation mitigates the biodiversity loss of grassland birds. Ecological Applications, 32, e2548. https://doi.org/10.1002/eap.2548
Pyšek, P., Jarošík, V., Hulme, P. E., Pergl, J., Hejda, M., Schaffner, U. et al. (2012). A global assessment of invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Global Change Biology, 18, 1725–1737. https://doi.org/10.1111/j.1365-2486.2011.02636.x
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D. et al. (2022). Vegan. Community Ecology Package: Ordination, Diversity and Dissimilarities. 3.4.
Qian, H. y Ricklefs, R. E. (2006). The role of exotic species in homogenizing the North American flora. Ecology Letters, 9, 1293–1298. https://doi.org/10.1111/j.1461-0248. 2006.00982.x
Qian, H. (2008). A latitudinal gradient of beta diversity for exotic vascular plant species in North America. Diversity and Distributions, 14, 556–560. https://doi.org/10.1111/j.1472- 4642.2008.00466.x
Rojas-Soto, O. R., Alcántara-Ayala, O. y Navarro-Sigüenza, A. G. (2003). Regionalization of the avifauna of the Baja California Peninsula, Mexico: a parsimony analysis of endemicity and distributional modelling approach. Journal of Biogeography, 30, 449–461. https://doi.org/10.1046/j.1365- 2699.2003.00836.x
Rosenberg, K. V., Dokter, A. M., Blancher, P. J., Sauer, J. R., Smith, A. C., Smith, P. A. et al. (2019). Decline of the North American avifauna. Science, 366, 120–124. https://doi.org/ 10.1126/science.aaw1313
Royo, M., Melgoza, A., Santos, J. S., Carrillo, R., Jurado, P., Gutiérrez, R. et al. (2005). La salud de los pastizales medianos en los estados de Chihuahua y Zacatecas. II Simposio Internacional de Manejo de Pastizales. Memorias. Zacatecas, México.
Ruvalcaba-Ortega I. y Panjabi, A. O. (2018). Aves de pastizal del Desierto Chihuahuense. En Conabio (Ed.), La biodiversidad en Coahuila. Estudio de Estado, vol. II. (pp. 375–384). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Gobierno del Estado de Coahuila de Zaragoza.
Rzedowski, J. y Calderón, de R. G. (1990). Nota sobre el elemento africano en la flora adventicia de México. Acta Botanica Mexicana, 12, 21–24. https://doi.org/10.21829/abm12.1990.602
Rzedowski, J., Calderón, de R. G. y Galván, R. (1995). Los pastizales calcífilos del estado de Guanajuato. Flora del Bajío y de Regiones Adyacentes. Fascículo Complementario IX. Pátzcuaro, Mich.: Instituto de Ecología, A.C. – Centro Regional del Bajío.
de S. Lopes, M. A. (2008). Revolución y ganadería en el norte de México. Historia Mexicana, 57, 863–910. https://www.jstor.org/stable/25139806
Sánchez-Ken, J. G., Zita-Padilla, G. A. y Mendoza-Cruz, M. (2012). Catálogo de las gramíneas malezas nativas e introducidas de México. México D.F.: Conacofi.
Sánchez-Ramos, L. E., Gordillo-Martínez, A., Gutiérrez-Arellano, C. R., Kobelkowsky-Vidrio, T., Ríos-Muñoz, C. A. y Navarro-Sigüenza, A. G. (2018). Bird diversity patterns in the Nuclear Central American Highlands: a conservation priority in the Northern Neotropics. Tropical Conservation Science, 11, 1940082918819073. https://doi.org/10.1177/1940082918819073
Savage, J. M. (1982). The enigma of the Central American herpetofauna: dispersals or vicariance? Annals of the Missouri Botanical Garden Press, 69, 464–547. https://doi.org/10.2307/2399082
Stanton, R. L., Morrissey, C. A. y Clark, R. G. (2018). Analysis of trends and agricultural drivers of farmland bird declines in North America: A review. Agriculture, Ecosystems & Environment, 254, 244–254. https://doi.org/10.1016/j.agee. 2017.11.028
Szentes, S., Sutyinszki, Z., Szabó, G., Zimmermann, Z., Házi, J., Wichmann, B. et al. (2012). Grazed Pannonian grassland beta-diversity changes due to C4 yellow bluestem. Central European Journal of Biology, 7, 1055–1065.
Valenzuela-Núñez, L. M., Mojica-Guerrero, A. S., González-Barrios, J. L. y Trucíos-Caciano, R. (2009). Evolución del matorral en el sur del Área Natural Protegida Sierra de Lobos, Guanajuato. Tecnociencia Chihuahua, 3, 131–137.
Vellend, M. (2010). Conceptual synthesis in community ecology. The Quarterly Review of Biology, 85, 183–206. https://doi.org/10.1086/652373
Vickery, P. D., Tubaro, P. L., Cardoso-da Silva, J. M., Peterjohn, B. G., Herkert, J. R. y Cavalcanti, R. B. (1999). Conservation of grassland birds in the Western Hemisphere. Studies in Avian Biology, 19, 2–26.
Wang, Y., Wang, J., Chen, C., Li, J. y Chu, J. (2019). Grazing plays an important role in structuring alpha and beta components of taxonomic, functional, and phylogenetic diversity in semiarid sandy land of northern China. Global Ecology and Conservation, 20, e00790. https://doi.org/10.1016/j.gecco. 2019.e00790
Weaver, T., Payson, E. M. y Gustafson, D. L. (1996). Prairie ecology —the shortgrass prairie. En F. B. Samson y F. L. Knopf (Ed.), Prairie Conservation. Washington D.C.: Island Press.
Wied, J. P., Perotto-Baldivieso, H. L., Conkey, A. A., Brennan, L. A. y Mata, J. M. (2020). Invasive grasses in South Texas rangelands: historical perspectives and future directions. Invasive Plant Science and Management, 13, 41–58.
Wiens, J. A. (1973). Pattern and process in grassland bird communities. Ecological Monographs, 43, 237–270. https://doi.org/10.2307/1942196
Wiken, E., Jiménez-Nava, F. y Griffith, G. (2011). North American terrestrial ecoregions —Level III. Montreal, Canada: Commission for Environmental Cooperation.
Wright, S. P. (1992). Adjusted p-values for simultaneous inference. Biometrics, 48, 1005–1013. https://doi.org/10.2307/2532694
Zar, J. H. (2010). Biostatistical analysis. Upper Saddle River, New Jersey: Pearson Education Inc.
Żmihorski, M., Pärt, T., Gustafson, T. y Berg, Å. (2016). Effects of water level and grassland management on alpha and beta diversity of birds in restored wetlands. Journal of Applied Ecology, 53, 587–595. https://doi.org/10.1111/1365-2664.12588
Louis C. Bender a, Jon C. Boren b, Shad Cox c, Erik Joaquín Torres-Romero d, e, *
a New Mexico State University, Department of Extension Animal Sciences and Natural Resources, PO Box 30003 MSC 3AE, Las Cruces, New Mexico 88003, USA
b New Mexico State University, Cooperative Extension Service, PO Box 30003 MSC 3AE, Las Cruces, New Mexico 88003, USA
c New Mexico State University, Corona Range and Livestock Research Center, PO Box 392, Corona, New Mexico 88318, USA
d Universidad Politécnica de Puebla, Ingeniería en Biotecnología, Tercer Carril del Ejido, Serrano s/n, San Mateo Cuanalá, Juan C. Bonilla, 72640 Puebla, Mexico
e Tecnológico Nacional de México campus Zacapoaxtla, Subdirección de Investigación y Posgrado, División de Biología, Carretera Acuaco-Zacapoaxtla Km. 8, Col. Totoltepec, 73680 Zacapoaxtla, Puebla, Mexico
Deer in northern temperate environments show behavioral and physiological adaptations to conserve energy during winter, including decreased movements. Whether these behaviors persist in warmer temperate environments such as the arid Southwest has received little consideration. We compared daily movements as estimated by continuous-time movement models and minimum subdaily (4 h) straight-line movements of adult female mule deer between winter and spring-autumn seasons in south-central New Mexico. Deer moved less during winter daily (2.90 vs. 4.34 km/d) and subdaily (302 vs. 409 m). Similarly, for deer for which movement data for successive seasons were available, movements between successive seasons were less during the winter (daily = -1.05 km/d; subdaily = -91 m) than the following or preceding spring-autumn. Our results support conservation of decreased movements during winter in the less extreme winters of the arid Southwest. Because some proximate stimuli (i.e., deep snow, very cold temperatures) associated with energy conservation behaviors are lacking in the arid Southwest, our results further support low forage quality and availability being the primary drivers of this behavior.
Keywords: Energy conservation; Movements; Mule deer; New Mexico
Disminución de los movimientos de las hembras adultas de venado bura durante el invierno en el árido suroeste de América del Norte
Resumen
Los ciervos en entornos templados del norte muestran adaptaciones de comportamiento y fisiológicas para conservar energía durante el invierno, incluyendo una disminución en sus movimientos. Se ha explorado poco si estos comportamientos persisten en ambientes templados más cálidos, como el suroeste árido. Comparamos los movimientos diarios, mediante modelos de movimiento continuo y movimientos mínimos en línea recta subdiarios (4 horas) de hembras adultas de venado bura entre las estaciones de invierno y primavera-otoño en el centro-sur de Nuevo México. Los ciervos se movieron menos durante el invierno, tanto diario (2.90 vs. 4.34 km/día) como subdiario (302 vs. 409 m). Además, para ciervos con datos de movimiento en estaciones sucesivas, los movimientos en invierno fueron menores (diarios = -1.05 km/día, subdiarios = -91 m) en comparación con la primavera-otoño previa o siguiente. Nuestros resultados respaldan la disminución de los movimientos durante el invierno en los inviernos menos extremos del suroeste árido. Dado que algunos estímulos inmediatos (por ejemplo, nieve profunda, temperaturas muy frías) asociados con comportamientos de conservación de energía están ausentes en el suroeste árido, es evidente que nuestros resultados apoyan que la baja calidad y disponibilidad de forraje son los principales factores que impulsan este comportamiento.
Palabras clave: Conservación de energía; Movimientos; Venado bura; Nuevo México
Introduction
Deer in northern temperate environments of North America employ a complex energy conservation strategy during winter, incorporating multiple behavioral and physiological adaptations, including reducing activity (i.e., movements) and limiting feeding while relying on endogenous reserves (Alldredge et al., 1974; Short, 1981; Verme & Ullrey, 1984); growing a highly insulative pelage (Jacobsen, 1980); limiting vascular circulation to the extremities (Parker & Robbins, 1984); and lowering metabolic rate to slow the rate of loss of body reserves (Short, 1981; Silver et al., 1971; Verme & Ullrey, 1984). This strategy is considered primarily an adaptation to conserve energy in response to low forage availability and quality, as well as increased costs of movement associated with snow (Short, 1981; Verme & Ullrey, 1984). Conserving energy by minimizing radiant and convective heat loss was also believed to drive other behaviors such as yarding under dense conifer forest canopies (Marchinton & Hirth, 1984). While initially thought to be a response to extreme cold (Marchinton & Hirth, 1984), yarding likely relates to decreased costs of movement because of reduced snow depths as the presumed temperature-moderating influence of forest canopy (i.e., thermal cover) has been shown to have no real effect on deer condition (Cook et al., 1998; Freddy, 1984). Many of the behavioral aspects (at least) of the winter energy conservation strategy are not invariant, however, and can be affected by proximate stimuli. For example, both movements and feeding periods are reduced less if winter conditions are less severe (Bartmann & Bowden, 1984; Verme & Ullrey, 1984).
Whether these energy conservation behaviors persist in warmer temperate ranges such as the arid southwestern USA and Mexico has received little consideration. In the arid Southwest, deer similarly experience winter seasonality in terms of temperature and precipitation differences (Krausman et al., 1990; McKinney, 2003; Marshal et al., 2008), which affects forage availability and quality (Kemp, 1983; Krausman et al., 1990; McKinney, 2003; Short, 1981). Consequently, mule deer lose most of their endogenous reserves over winter (Bender et al., 2012; Bender & Hoenes, 2017). However, snow is relatively rare and short-lived in much of the arid Southwest, and winter temperatures are higher than in northern environments (Table 1). Therefore, aside from decreased forage availability and quality, many of the potential proximate stimuli (i.e., deep snow, very cold temperatures) associated with energy conservation behaviors are lacking in the arid Southwest. Moreover, because winter is less extreme in terms of minimum temperatures and particularly snowfall, availability of forage may also be less limiting, although forage quality constraints are similarly severe (Bender, 2020; Kemp, 1983; Krausman et al., 1990; McKinney et al., 2003). Consequently, behavioral responses associated with the winter energy conservation strategy may be less pronounced or absent in the arid Southwest.
Table 1
Long-term range of monthly mean high and low temperatures (oC) and monthly snowfall (cm) during Dec.-Feb. and Mar.-Nov. at the Corona Range and Livestock Research Center (CRLRC), Corona, New Mexico USA, and the Cusino Wildlife Research Station, Shingleton, Michigan USA. Cusino was selected as a comparison because of the volume and depth of deer nutritional, physiological, and behavioral research conducted there (Verme & Ullrey, 1984). Also presented are the range of conditions on the CRLRC study area for 2005-2008 study period.
Months
Climatic variable
CRLRC
Cusino
Study
Dec.-Feb.
Mean high temperatures
6.7-8.9
-2.8- -1.7
6.4-13.0
Mean low temperatures
-5.6- -4.4
-11.7- -8.3
-6.3- -1.9
Mean snow accumulations
13-23
69-109
7.3-17.1
Mar.-Nov.
Mean high temperatures
11.7-28.3
1.7-22.8
14.2-29.1
Mean low temperatures
-1.1-13.3
-6.7-13.3
-0.5-13.8
Mean snow accumulations
0-13
0-48
0-5.1
If energy conservation behaviors are maintained in the arid Southwest, mule deer (Odocoileus hemionus) should move less during the winter, conserving body reserves in the face of lower quality and less abundant forage even if the impacts of winter weather are less severe on forage availability and costs of movement. Thus, our goal was to contrast short-term movements between winter and spring-autumn seasons for adult female mule deer in a Chihuahuan desert-short grass prairie habitat in New Mexico, USA to determine whether deer reduce movements during winter as predicted by the winter energy conservation strategy. Specifically, we compared minimum daily and subdaily movement distances of adult female mule deer between winter and spring-autumn.
Materials and methods
Our study was conducted on the Corona Range and Livestock Research Center (CRLRC; 34°15’36” N, 105°24’36” W), an 11,290-ha ranch owned and operated by New Mexico State University and located approximately 22.5 km east of Corona, New Mexico (Fig. 1). CRLRC has an average elevation of 1,900 m asl; mean annual precipitation is 40 cm, 87% of which occurs in the Mar.-Nov. period. Snowfall totals < 74 cm annually. Climate of the CRLRC shows distinct seasonality, although the magnitude of seasonal differences in winter is less than seen in northern temperate deer habitats (Table 1).
Topography of the CRLRC is mostly rolling. Vegetation includes perennial grassland, with scattered sparse to dense pinyon (Pinus edulis) and one-seed juniper (Juniperus monosperma) woodlands and a few shrublands. Free water was abundant and comparably available in both winter and spring-autumn seasons because of numerous permanent water developments, ≥ 1 of which were present within or adjacent to annual home ranges of study deer (Fig. 1). Deer on the CRLRC do not migrate between distinct summer and winter ranges.
We captured and collared ≥ 2.5-year-old female mule deer with GPS/VHF radio-collars (Advanced Telemetry Solution, Asanti, Minnesota, USA) programmed to record a position fix every 4 h, early-December 2005-2007, and April, 2006-2007, as part of a larger study of mule deer ecology including other VHF-only radio-collared individuals (Bender et al., 2011, 2013). Deer were captured using a helicopter by aerial net-gunning or darting with 1.5-1.8 mg of carfentanil citrate and 50-75 mg of xylazine hydrochloride per deer. We aged deer as yearling or adult by tooth wear and replacement (Robinette et al., 1957), determined lactation status (Bender et al., 2011), and treated deer with antibiotics, vitamin E/selenium, vitamin B, and an 8-way Clostridium bacterin to help alleviate capture stress. Following processing, immobilants were antagonized with naltrexone and tolazoline.
We defined seasons as winter = Dec-Feb and spring-autumn = Mar-Nov. These seasons corresponded with both typical seasonal and phenological patterns on the CRLRC and the Chihuahuan desert-short grass prairie habitats of the arid Southwest in general, as well as important periods in the annual cycle of female mule deer in the arid Southwest (Bender et al., 2011, 2012). We estimated daily movements using continuous-time movement modeling (see Fleming et al., 2014, 2016), using the Speed/Distance analysis in ctmmweb (https://ctmm.shinyapps.io/ctmmweb/) (Calabrese et al., 2016, 2021). We used only locations with 3D fixes and DOP < 2, as these had an accuracy of < 3 m in our study area. We also determined minimum subdaily movements, defined as the straight-line distance moved between successive 4 h locations, and calculated seasonal means for each deer.
Figure 1. Topographic hillshade showing locations of annual home ranges of adult female mule deer (Bender et al., 2013), and locations of permanent water sources on the Corona Range and Livestock Research Center (CRLRC), east-central New Mexico, USA.
We compared movement distances (km from continuous-time movement models [ctmms]; m between successive subdaily locations) between seasons using PROC GLIMMIX in SAS 9.4 (SAS, 1988), using individual deer as a random effect. We also compared mean daily and subdaily movements between successive seasons for individual deer for which we had both winter and subsequent spring-autumn data, or spring-autumn and subsequent winter data, available. We determined mean seasonal differences in movement distances for each successive time period for each individual, and used bootstrapping with N = 1,000 iterations to determine the probability that mean movement distances differed seasonally for this subset of data (see Efron & Tibshirani, 1993).
Additionally, because lactating females enter winter in poorer condition than do dry females and condition subsequently converges between the 2 classes over winter (including on the CRLRC; Bender & Hoenes, 2017), lactation status might affect desire or need to forage and thus movements of deer. However, because deer condition was very low on CRLRC during our study (i.e., lactating females were able to accrue only ≤ 5.7% percent body fat annually at the annual peak in late autumn; Bender et al., 2011, 2013), our GPS/VHF collared sample contained ≤ 2 lactating females each year, and thus we were unable to meaningfully include lactation status in our analyses. Consequently, we explored any potential effect of lactation status on movements by determining the percentile movement distances of lactating females relative to the frequency distribution of dry female movement distances to determine whether lactating females were closer to the mean or extremes of the range of dry females.
Lastly, because our GPS collared sample comprised a limited proportion of the overall radio-collared sample, we compared annual, spring-autumn, and winter home range sizes (Bender et al. 2013; N = 18-27 collared females annually) to see whether movements of our GPS sample were representative of radio-collared deer in general, comprised of N = 18-27 VHF-only collared adult females for each season (Bender et al., 2011, 2013). For this we compared annual and seasonal home range sizes of the 2 classes (GPS/VHF and VHF-only) using PROC GLM (SAS, 1988), specifically testing the year × class interaction. We visually located all deer (i.e., both GPS/VHF and VHF-only) via ground tracking of VHF signals a minimum of once per week with additional location emphasis on spring-autumn locations, and mapped locations using the Geographic Information System software package ArcGIS 10.0 (Environmental Systems Research Institute, Redlands, California, USA) (Bender et al., 2011, 2013). We constructed 95% minimum convex polygon (MCP) annual and seasonal home ranges after determining the minimum number of locations to adequately estimate seasonal home range size by plotting size as a function of number of locations (Bender et al. 2013; Kie et al. 1996). For this comparison we used only VHF visual locations of GPS/VHF collared deer so that both GPS/VHF and VHF-only samples were comprised of comparable data.
Results
We collected GPS movement data for 37 seasonal ranges (Table 2), winter 2005-6 through winter 2007-8, from 6-10 GPS/VHF collared adult females annually (mean = 336 and 472 locations per deer for winter and spring-autumn ranges, respectively). For all deer, the OUF-anisotropic movement model provided the best fit (i.e., AICc < 2 vs. all other models) of deer movements. The OUF-anisotropic movement model is the most general of ctmms, and includes a home range, correlated positions, correlated velocities, and movements varying by direction (Calabrese et al., 2021; Fleming et al., 2014).
Table 2
Mean distance moved by adult female mule deer during winter (Dec.-Feb.) and spring-autumn (Mar.-Nov.) seasons as estimated by continuous-time movement modeling (daily) and subdaily straight line movements between successive 4 h locations (subdaily) on the Corona Range and Livestock Research Center, east-central New Mexico, 2005-2007.
Daily (km)
Subdaily (m)
Season
Distance
SE
N
Distance
SE
N
Winter
2.90
0.24
24
302
19
24
Spring-Autumn
4.34
0.43
13
409
44
13
Deer moved less during the winter (Table 2) for both daily ctmms (F1,20 = 6.0; p = 0.024) and minimum subdaily straight-line distances (F1,20 = 4.8; p = 0.041); in both cases the magnitude of differences varied among individual deer (t15 < -2.72; p < 0.016). Similarly, for deer for which movement data for successive seasons were available, movements between successive seasons were always less during the winter season (p (winter < spring-autumn) = 1.000) than the following or preceding spring-autumn season (Table 3).
Additionally, movement distances of lactating females (mean = 42nd percentile; range = 34-55th percentile) were always closer to the average than the extremes of the frequency distribution of movement distances of dry females for each period. Last, neither annual, spring-autumn, or winter home MCP range sizes (N = 7-16 per period) differed between GPS/VHF and VHF-only collared deer (F5,39 ≤ 0.94; p ≥ 0.599).
Discussion
Despite much less snow and warmer winter temperatures, mule deer females moved less during winter regardless of movement period (i.e., daily, subdaily), supporting the maintenance of this energy conservation behavior in mule deer in the arid Southwest (Verme & Ullrey, 1984). Because deep snow cover and very cold temperatures are typically lacking in the arid Southwest, our results further support low forage quality and availability being the primary drivers of this behavior in deer (Verme & Ullrey, 1984; see below). This latter was reflected by the poor nutritional condition of lactating adult females in the study area (i.e., <5.7% body fat annually at the annual peak in late autumn; Bender et al., 2011, 2013), and in the arid Southwest in general (Bender et al., 2007, 2011, 2012; Bender, 2020), as well as the significant losses in condition seen over winter (Bender & Hoenes, 2017; Hoenes, 2008), despite mule deer likely requiring lower quality forage than white-tailed deer (O. virginianus) (Staudenmaier et al., 2022).
While lack of deep snow cover may result in relative forage availability being less impacted during winter in the arid Southwest, senesced forages are still of very low quality, similar to northern environments (Kemp, 1983; Krausman et al., 1990; McKinney, 2003). Low forage quality in winter was reflected in the condition dynamics of deer on the CRLRC; for example, dry females lost on average 32% of body fat reserves and 38% of rump body condition score over winter (L. Bender, unpublished data), even though their condition in late autumn-early winter was already low relative to other southwestern populations (Bender et al., 2007, 2011, 2012). The nutritional condition of deer is driven primarily by forage quality (Bender, 2020; National Research Council, 2007; Tollefson et al., 2010; Verme & Ullrey, 1984; Wakeling & Bender, 2003), illustrating that deer in the arid Southwest face similar constraints in terms of limited nutrient gains from forage intake as do deer in northern environments during winter.
Table 3
Mean movement distances (x) and mean differences (D) in mean distance moved daily as predicted by continuous-time movement modeling (daily) and subdaily straight line movements between successive 4 h locations (subdaily) of adult female mule deer for which movement data for successive seasons were available between winter (Dec.-Feb.) and spring-autumn (Mar.-Nov.) on the Corona Range and Livestock Research Center, east-central New Mexico, 2005-2007. p = Probability that seasonal differences differ from 0; N = number of seasonal comparisons.
Period
x Summer
x Winter
D
90% CI
p
N
Subdaily
400.2
309.6
-90.6 m
-123- -63
1.000
20
Daily
4.21
3.16
-1.05 km
-1.53- -0.63
1.000
20
Deer in northern environments do face additional energetic challenges associated with persistent snow cover, which can limit forage availability (by making location and acquisition of food more difficult; Hovey & Harstad, 1992), diet quality (due to reduced forage quality and availability; McKinney, 2003; Osborn & Jenks, 1998), and increase loss of endogenous reserves (because of increased costs of moving through snow; Bunnell et al., 1990; Mattfeld, 1973). However, while deer in the arid Southwest are less influenced by snow-depth related challenges, northern deer do not face the lack of free water experienced by most deer populations in arid environments because of persistent snow cover in northern environments. Lack of water can present an energetic cost to deer in the arid Southwest, as mule deer may increase movement distances to access water (Heffelfinger, 2006), and winter is much drier than spring-autumn in Chihuahuan desert and short-grass prairie habitats of the arid Southwest (e.g., 87% of precipitation occurs during spring-autumn on the CRLRC).
Because of permanent water developments, water was comparably available seasonally on the CRLRC; water developments were accessible from all deer home ranges, so mule deer did not need to alter their movements in response to seasonal changes in availability of water. Hence, need or preference for free water likely had a negligible effect on deer movements on the CRLRC, unless presence of temporary sources (ephemeral pools, etc.) during the summer monsoon reduce deer movements during spring-autumn because of increased availability. Thus, despite water being effectively controlled in our study, deer still showed less movements during winter. This again supports decreased movements during winter being most influenced by the lack of energetic benefit from seeking and foraging on low quality senesced forage.
Cold temperatures are often thought to influence the winter conservation strategy, despite demonstrated lack of benefit of thermal cover in winter to deer (Cook et al., 1998; Freddy, 1984), the pronounced effect of solar radiation on warming deer (Cook et al., 1998; Parker & Gillingham, 1990; Parker & Robbins, 1984), and deer movements and tolerance of exposure to cold (including bedding in the open) except during the most extreme conditions when high quality forage is available (Moen, 1968; Verme & Ullrey, 1984). Mule deer possess a low thermal critical zone (ca. -20 oC) and show greater tolerance of cold than do white-tailed deer, including a lower metabolic rate response to decreasing temperatures (Mautz et al., 1985; Parker & Robbins, 1984). Hence, they are less affected by even extreme cold, which is seldom the case in the arid Southwest where mean low temperatures seldom approach their lower thermal tolerance (Table 1). Conversely, the upper thermal critical level of mule deer in winter pelage (5 ºC; Mautz et al., 1985) is lower than average high temperatures during winter on our study area (6.7-8.9 ºC; Table 1) and much of the arid Southwest. This would require active metabolic activity or behaviors (e.g., panting, etc.) by mule deer to cool themselves, or possibly limiting movements and bedding under shade (although the energetic benefits of the latter are questionable; Cook et al., 1998). Consequently, if temperature affected movements during the winter in our study area, deer would be more likely to decrease movements because of heat stress, not cold stress.
Lastly, while at least one behavioral aspect of the winter energy conservation strategy is seen in mule deer in the arid Southwest, to what degree other adaptations are conserved is unknown. Mule deer in the arid Southwest do develop a highly insulative pelage in winter (Heffelfinger, 2006), but the extent that they may decrease metabolic rate or regulate vascular circulation to the extremities (or need to, in light of the more moderate temperatures) is unknown. Additionally, while lactating females showed a tendency to move less relative to dry females on the CRLRC, whether this is typical is unknown because of our small sample of lactating females. However, given that most females on CRLRC were in poorer condition than females elsewhere in the arid Southwest (Bender et al., 2007, 2011, 2012; Bender & Hoenes, 2017), if very low condition (such as results from lactation) increases movements during winter, this increase would likely have been seen in all CRLRC females regardless of lactation status. Moreover, GPS/VHF and VHF-only collared females showed similar movements (as indexed by home range sizes) seasonally and annually on CRLRC, indicating that movements of GPS/VHF collared females reflected females in general. Although small sample sizes (3-4) precluded including lactation status as an interactive term in the contrast of GPS/VHF and VHF-only collared females, for all females lactation status had no effect on annual or seasonal home range sizes (p ≥ 0.599).
Acknowledgments
Support for this project was provided by the U.S. Forest Service-Rocky Mountain Research Station and the New Mexico State University Cooperative Extension Service and Agricultural Experimental Station. All activities were in accordance with NMSU IACUC Permit No. 2005-023. E.J.T.-R. was supported by a postdoctoral fellowship from Consejo Nacional de Humanidades, Ciencias y Tecnologías (Conahcyt-Mexico).
References
Alldredge, A. W., Lipscomb, J.F., & Whicker, F.W. (1974). Forage intake rates of mule deer estimated with fallout cesium-137. The Journal of Wildlife Management, 38, 508–516. https://doi.org/10.2307/3800882
Bartmann, R. M., & Bowden, D. C. (1984). Predicting mule deer mortality from weather data in Colorado. Wildlife Society Bulletin, 12, 246–248. https://doi.org/10.1002/jwmg.956
Bender, L. C. (2020). Elk, deer, and pinyon-juniper: needs, what works, and what doesn’t. In K. Malcolm, B. Dykstra, K. Johnson, D. Lightfoot, E. Muldavin, & M. Ramsey (Compilers), Symposium proceedings on piñon-juniper habitats: status and management for wildlife – 2016. Proceedings RMRS-P-77. Fort Collins, Colorado: U.S. Forest Service, Rocky Mountain Research Station.
Bender, L. C., & Hoenes, B. D. (2017). Costs of lactation to body condition and future reproduction of free-ranging mule deer Odocoileus hemionus (Cervidae). Mammalia, 81, 329–338. https://doi.org/10.1515/mammalia-2015-0143
Bender, L. C., Lomas, L. A., & Browning, J. (2007). Condition, survival, and cause-specific mortality of mule deer in northcentral New Mexico. Journal of Wildlife Management, 71, 1118–1124. https://doi.org/10.2193/2006-226
Bender, L. C., Boren, J. C., Halbritter, H., & Cox, S. (2011). Condition, survival, and productivity of mule deer in semiarid grassland-woodland in east-central New Mexico. Human-Wildlife Interactions, 5, 276–286. https://doi.org/10.26077/60n0-ks48
Bender, L. C., Hoenes, B. D., & Rodden, C. L. (2012). Factors influencing survival of desert mule deer in the greater San Andres Mountains, New Mexico. Human-Wildlife Interactions, 6, 245–260. https://doi.org/10.26077/h5bg-1829
Bender, L. C., Boren, J. C., Halbritter, H., & Cox, S. (2013). Effects of site characteristics, pinyon-juniper control, and precipitation on habitat quality for mule deer on the Corona Range and Livestock Research Center. Human-Wildlife Interactions, 7, 47–59. https://digitalcommons.usu.edu/hwi/vol7/iss1/5
Bunnell, F. L., Hovey, F. W., McNay, R. S., & Parker, K. L. (1990). Forest cover, snow conditions, and black-tailed deer sinking depths. Canadian Journal of Zoology, 68, 2403–2408. https://doi.org/10.1139/z90-333
Calabrese, J. M., Fleming, C. H., & Gurarie, E. (2016). ctmm: an R package for analyzing animal relocation data as a continuous-time stochastic process. Methods in Ecology and Evolution, 7, 1124–1132. https://doi.org/10.1111/2041-210X.12559
Calabrese, J. M., Fleming, C. H., Noonan, M. J., & Dong, X. (2021). ctmmweb: a graphical user interface for autocorrelation-informed home range estimation. Wildlife Society Bulletin, 45, 162–169. https://doi.org/10.1002/wsb.1154
Cook, J. G., Irwin, L. L., Bryant, L. D., Riggs, R. A., & Thomas, J. W. (1998). Relations of forest cover and condition of elk: a test of the thermal cover hypothesis in summer and winter. Wildlife Monographs, 141, 1–61.
Efron, B., & Tibshirani, R. J. (1993). An introduction to the bootstrap. New York: Chapman & Hall.
Fleming, C. H., Fagan, W. F., Mueller, T., Olson, K. A., Leimgruber, P., & Calabrese, J. M. (2014). From fine-scale foraging to home ranges: a semivariance approach to identifying movement modes across spatiotemporal scales. American Naturalist, 183, E154–E167. http://doi.org/10.1086/675504
Fleming, C. H., Fagan, W. F., Mueller, T., Olson, K. A., Leimgruber, P., & Calabrese, J. M. (2016). Estimating where and how animals travel: an optimal framework for path reconstruction from autocorrelated tracking data. Ecology, 97, 576–582. https://doi.org/10.1890/15-1607.1
Freddy, D. J. (1984). Quantifying capacity of winter ranges to support deer – evaluation of thermal cover used by deer. Denver, CO: Wildlife Research Report, Colorado Division of Wildlife, USA.
Heffelfinger, J. (2006). Deer of the Southwest: a complete guide to the natural history, biology, and management of Southwestern mule deer and white-tailed deer. Texas A&M University, College Station, Texas.
Hoenes, B. D. (2008). Identification of factors limiting desert mule deer populations in the greater San Andres Mountains of southcentral New Mexico (Thesis). New Mexico State University, Las Cruces.
Hovey, F. W., & Harestad, A. S. (1992). Estimating effects of snow on shrub availability for black-tailed deer in southwestern British Columbia. Wildlife Society Bulletin, 20, 308–313.
Jacobsen, N. K. (1980). Differences of thermal properties of white-tailed deer pelage between seasons and body regions. Journal of Thermal Biology, 5, 151–158. https://doi.org/10.1016/0306-4565(80)90014-5
Kemp, P. R. (1983). Phenological patterns of Chihuahuan desert plants in relation to the timing of water availability. Journal of Ecology, 71, 427–436.
Kie, J. G., Baldwin, J. A., & Evans, C. J. (1996). CALHOME: a program for estimating animal home ranges. Wildlife Society Bulletin, 24, 342–344.
Krausman, P. R., Ordway, L. L., Whiting, F. M., & Brown, W. H. (1990). Nutritional composition of desert mule deer forage in the Picacho Mountains, Arizona. Desert Plants, 10, 32–34.
Marchinton, R. L., & Hirth, D. H. (1984). Behavior. In L. K. Halls (Ed.), White-tailed deer ecology and management. Pennsylvania: Stackpole Books.
Marshal, J. P., Krausman, P. R., & Bleich, V. C. (2008). Body condition of mule deer in the Sonoran Desert is related to rainfall. Southwestern Naturalist,53, 311–318. https://doi.org/10.1894/CJ-143.1
Mattfeld, G. F. (1973). The effect of snow on the energy expenditure of walking white-tailed deer. In Transactions of the 30th Northeast Fish and Wildlife Conference, Dover, Vermont, Spring, April 1973.
Mautz, W. W., Pekins, P. J., & Warren, J. A. (1985). Cold temperature effects on metabolic rates of white-tailed, mule, and black-tailed deer in winter coat. In P. F. Fennessy, & K. R. Orew (Eds.), The biology of deer. Royal Society of New Zealand Bulletin, 22, 453–457.
McKinney, T. (2003). Precipitation, weather, and mule deer. In J. C. Jr. deVos, M. R. Conover, & N. E. Headrick (Eds.), Mule deer conservation. Issues and management strategies. Logan, Utah: Jack H. Berryman Press.
Moen, A. N. (1968). Energy exchange of white-tailed deer, western Minnesota. Ecology, 49, 676–682. https://doi.org/10.2307/1935531
National Research Council (US). (2007). Committee on Nutrient Requirements of Small Ruminants. Nutrient requirements of small ruminants: sheep, goats, cervids, and New World camelids. Washington D.C.: National Academy Press.
Osborn, R. G., & Jenks, J. A. (1998). Assessing dietary quality of white-tailed deer using fecal indices: effects of supplemental feeding and area. Journal of Mammalogy, 79, 437–447. https://doi.org/10.2307/1382974
Parker, K. L., & Robbins, C. T. (1984). Thermoregulation in mule deer and elk. Canadian Journal of Zoology,62, 1409–1422. https://doi.org/10.1139/z84-202
Parker, K. L., & Gillingham, M. P. (1990). Estimates of critical thermal environments for mule deer. Journal of Range Management,43, 73–81. http://doi.org/10.2307/3899126
Robinette, W. L., Jones, D. A., Rogers, G., & Gashwiler, J. S. (1957). Notes on tooth development and wear for Rocky Mountain mule deer. Journal of Wildlife Management, 21, 134–153. https://doi.org/10.2307/3797579
SAS. (1988). SAS/STAT user’s guide. SAS Institute Incorporated, Cary, North Carolina.
Short, H. L. (1981). Nutrition and metabolism. In O. C. Wallmo, R. E. McCabe, & L. R. Jahn (Eds.), Mule and black-tailed deer of North America. Lincoln: University of Nebraska Press.
Silver, H., Holter, J. B., Colovox, N. F., & Hayes, H. H. (1971). Effect of falling temperature on heat production in fasting white-tailed deer. Journal of Wildlife Management, 35, 37–46.
Staudenmaier, A. R., Shipley, L. A., Camp, M. J., Forbey, J. S., Hagerman, A. E., Brandt, A. E. et al. (2022). Mule deer do more with less: comparing their nutritional requirements and tolerances with white-tailed deer. Journal of Mammalogy, 103, 178–195. https://doi.org/10.1093/jmammal/gyab116
Tollefson, T. N., Shipley, L. A., Myers, W. L., Keisler, D. H., & Dasgupta, N. (2010). Influence of summer-autumn nutrition on body condition and reproduction in lactating mule deer. Journal of Wildlife Management, 74, 974–986. https://doi.org/10.2193/2008-529
Verme, L. J., & Ullrey, D. E. (1984). Physiology and nutrition. In L. K. Halls (Ed.), White-tailed deer: Ecology and management. Harrisburg, Pennsylvania: Stackpole Books.
Wakeling, B. F., & Bender, L. C. (2003). Influence of nutrition on mule deer biology and ecology. In J. C. Jr. deVos, M. R. Conover, & N. E. Headrick (Eds.), Mule deer conservation. Issues and management strategies. Logan, Utah: Jack H. Berryman Press.
Oscar Pérez-Flores a, b, *, Santiago Zaragoza-Caballero b
a Universidad Nacional Autónoma de México, Escuela Nacional de Estudios Superiores, Laboratorio Nacional de Análisis y Síntesis Ecológica, Antigua Carretera a Pátzcuaro No. 8701, 58190 Morelia, Michoacán, Mexico
b Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Apartado postal 70-153, 04510 Ciudad de México, Mexico
The genus Telegeusis Horn is the most diverse of the family Telegeusidae, with 16 described species, most of them distributed in Mexico (13 species). Herein, we describe 4 new species based on material collected from Tamaulipas, Mexico: T. cieloensis sp. nov., T. estebani sp. nov., T. flavafusca sp. nov., and T. luismartinezi sp. nov. Distinguishing characters from closely related species are given and discussed, and the key for the species of the genus is updated.
Especies nuevas de Telegeusis (Coleoptera: Telegeusidae) del norte de México
Resumen
El género Telegeusis Horn es el más diverso dentro de la familia Telegeusidae, con 16 especies descritas, muchas de ellas distribuidas en México (13 especies). En el presente trabajo, describimos 4 especies nuevas con base en material colectado de Tamaulipas, México: T. cieloensis sp. nov., T. estebani sp. nov., T. flavafusca sp. nov. y T. luismartinezi sp. nov. Se proporcionan y analizan los caracteres diagnósticos de especies estrechamente relacionadas, y se actualiza la clave para las especies del género.
Palabras clave: Telegeúsidos; Región Neotropical; Taxonomía; Coloración
Introduction
The small family Telegeusidae (Coleoptera: Elateroidea) contains 23 species distributed in 5 genera: Telegeusis Horn (16 spp.), Pseudotelegeusis Wittmer (4 spp.), Pseudokarumia Pic (1 sp.), Stenodrilus López-Pérez & Zaragoza-Caballero (1 sp.), and Platydrilus López-Pérez & Zaragoza-Caballero (1 sp.) (López-Pérez & Zaragoza-Caballero, 2021). In Mexico 17 species and 4 genera have been recorded (López-Pérez & Zaragoza-Caballero, 2021; Zaragoza-Caballero, 2008, 2015), becoming the most diverse country for the family.
Telegeusidae are easily recognized by their remarkably long apical labial and maxillary palpomeres (Lawrence, 2010; Miller, 2002). Only males have been described, and little is known about the ecology and biology of immature stages (Lawrence, 2010). Commonly, adult specimens are collected with light traps or interception traps (e.g., Malaise) (López-Pérez & Zaragoza-Caballero, 2021; Zaragoza-Caballero, 2008, 2015; Zaragoza-Caballero & Rodríguez-Vélez, 2011). The taxonomic position of Telegeusidae within Elateroidea has been controversial, placing them either as a subfamily of Oemethidae or as an independent family (Kundrata et al., 2014; López-Pérez & Zaragoza-Caballero, 2021; Ramsdale, 2010). Herein we treat them as a family.
Species of Telegeusis Horn, 1895 are recognized by their labial palpi 2-3 segmented, last palpomere greatly enlarged and elongated, similar to maxillary palpi; and antennae filiform (Ivie, 2002). The 16 currently recognized species are distributed from southern USA to Panama, with 13 species recorded for Mexico (Zaragoza-Caballero, 2008, 2015; Zaragoza-Caballero & Rodríguez-Vélez, 2011). The length of last maxillary and labial palpi, length of elytra, and integument coloration have been used to separate species within the genus (Zaragoza-Caballero, 2015; Zaragoza-Caballero & Rodríguez-Vélez, 2011). In this work, we describe 4 new species of Telegeusis from Mexico and update the key for the species.
Materials and methods
Specimens studied were collected with a blacklight and Malaise traps in El Cielo Biosphere Reserve, Tamaulipas, Mexico. The morphological terminology and classification are based on Ivie (2002), Zaragoza-Caballero (2015), and Zaragoza-Caballero and Rodríguez-Vélez (2011). Morphological structures were studied using a Zeiss Stemi DV-4 stereomicroscope. Measurements are in millimeters. Photographs were taken with a Zeiss® AxioZoom V16 stereomicroscope, with a Zeiss® AxioCam MRc5, 5 megapixels camera, controlled by ZEN (Zeiss Efficient Navigation) application. Specimens are deposited in the Colección de Insectos, UAEM, Morelos, Mexico (CIUM), and the Colección Nacional de Insectos, Instituto de Biología, UNAM, CDMX, Mexico (CNIN). All label data were transcribed verbatim as appearing on the respective labels.
Diagnosis. Similar to T. debilis Horn, 1895 but differing in the length of head which is slightly longer than wide (twice longer than wide in T. debilis); maxillary and labial palpi slightly longer than antennae (shorter than antennae in T. debilis); elytra reaching posterior third of abdomen (reaching the middle in T. debilis).
Holotype male (CNIN). Size: length 4.1 mm; width 0.75 mm. Color: head, base of elytra and legs yellowish brown; meso and metaventrite orange; antennae, pronotum, posterior part of elytra and abdomen reddish brown; apical palpomeres brown. Head: flat, 1.2 times longer than wide, at eye level, distance between outer margins of eyes slightly wider than pronotum (0.62 mm). Interocular area convex, forming pronounced gibba centrally (Fig. 1C). Distance between eyes and pronotum 0.85 times the length of eye. Integument shiny, shagreened, finely punctate; each puncture with an erect, yellow seta; setae longer behind eyes; with strongly deep tentorial pits. Antennal tubercles prominent, convex (Fig. 1C). Inter-antennal area concave, 2.25 times as wide as length of scape. Antennae longer (1.2 mm) than last maxillary palpomere (1.05 mm) (Fig. 1A, B); scape slightly widened. Eyes prominent, slightly longer (0.18 mm) than wide (0.15 mm); interocular distance 3.3 times width of eyes. Frons concave, anterior margin concave. Clypeus with margin slightly sinuous. Maxillary and labial palpi with 3 and 2 palpomeres, respectively (Fig. 1D). Thorax: pronotum wider (0.62 mm) than long (0.45 mm), width less than length of head (Fig. 1A, C); anterior margin straight, posterior margin convex, sides slightly constricted at middle; disk glossy, convex centrally, with 1 elongated gibbosity on each side; integument finely, sparsely punctate, with erect, sparse orange setae. Scutellum shiny, finely punctate, with sparse setae, posterior margin truncate. Elytra 6.45 times as long as wide (base width) (Fig. 1A), subparallel on basal half, then becoming narrower toward rounded apex; integument coarsely punctate, granulate on posterior third; each puncture with an erect, yellow seta. Metathoracic wings 2.35 times longer than wide (Fig. 1E), anterior radial vein (RA) well sclerosed, posterior radial 2 (PR2) present, cubital 2 (CU2) slightly curved, elongated. Legs finely punctate, with dense yellow setae, yellowish-brown on apical half of tibiae. Coxae conical; procoxae long. Trochanters elongate. Femora fusiform, flattened; metafemora longer and wider (Fig. 1A, B). Tibiae elongate, with 2 apical spines; metatibia longer and slightly widened toward apex. Tarsomere I shorter in length than II-IV together; claws simple. Abdomen: shiny, integument shagreened with fine punctures, each puncture with an erect, yellow seta (Fig. 1B).
Figure 1. Telegeusis cieloensis sp. nov., holotype male: A) dorsal habitus; B) ventral habitus; C) head, dorsal view; D) palpi, ventral view; E) paratype, metathoracic wing. Scale bars = 1 mm (A-B, E); 0.5 mm (C-D).
Variation. The type material varies in total length from 4.08 to 5.86 mm, and in width from 0.7 mm to 0.93 mm.
Taxonomic summary
Etymology. The specific epithet is derived from El Cielo Biosphere Reserve where the holotype was collected, combined with the Latin suffix –ensis, meaning belonging to.
Type material. Holotype male: Mexico: Tamaulipas, Gómez Farías, Reserva de la Biosfera El Cielo, Malaise trap, 23°08’01.1” N, 99°13’11.6” W, 19/Abril/2021, 1,600 m snm, M. Janda et al. leg. (CNIN). Paratypes. Same data as holotype except: 23°05’56.2” N, 99°12’17.1” W, 31/Enero/2020, 1,200 m snm (CNIN, 1); 23°08’09.4” N, 99°12’53.1” W, 01/Septiembre/2020, 1,800 m snm (CNIN, 2); 23°05’56.2” N, 99°12’17.1” W, 16/Septiembre/2020, 1,200 m snm (CIUM, 2); light trap, 23°05’16.5” N, 99°11’59.6” W, 10/Octubre/2020, 1,200 m snm (CNIN, 1); light trap, 23°07’39.3” N, 99°12’30.6” W, 10/Octubre/2020, 1,400 m snm (CIUM, 2); 23°05’56.2” N, 99°12’17.1” W, 10/Diciembre/2020, 1,200 m snm (CIUM, 1); 23°05’56.2” N, 99°12’17.1” W, 19/Abril/2021, 1,200 m snm (CNIN, 1); 23°08’01.1” N, 99°13’11.6” W, 19/Abril/2021, 1,600 m snm (CNIN, 1); 23°08’01.1” N, 99°13’11.6” W, 05/Mayo/2021, 1,600 m snm (CNIN, 1); light trap, 23°09’05.3” N, 99°12’53.8” W, 17/Julio/2021, 1,400 m snm (CNIN, 1).
Diagnosis. Similar to T. moroni Zaragoza-Caballero, 2015 determined by the color and length of maxillary palpi, but it differs with prominent eyes (small in T. moroni); and with 2 labial palpomeres (3 in T. moroni). The speciesalso resembles T. cieloensis sp. nov. but differs mainly by a darker coloration (lighter in T. cieloensis), and the length of elytra, just surpassing the middle of elytra (reaching last third in T. cieloensis).
Holotype male (CNIN). Size: length 5.1 mm; width 0.92 mm. Color: mostly brown; somewhat orangish-brown on frons and vertex; antennae orangish brown; pedicel, meso-, metaventrite, scutellum and legs orange; apical palpomeres dark brown. Head: flat, 1.1 times longer than wide, at eye level, distance between outer margins of eyes slightly wider than pronotum (0.7 mm). Interocular area slightly convex, forming small gibba (Fig. 2C). Distance between eyes and pronotum 0.92 times length of eye. Integument shiny, shagreened, finely punctate; each puncture with an erect, orange seta; setae longer around eyes; with conspicuous tentorial pits. Antennal tubercles prominent, slightly convex (Fig. 2C). Inter-antennal area concave; 2.75 times as wide as length of scape. Antennae just longer (1.65 mm) than last maxillary palpomere (1.57 mm) (Fig. 2A, B); scape somewhat prominent. Eyes prominent, longer (0.27 mm) than wide (0.2 mm); interocular distance 2.9 times width of eyes. Frons slightly concave, anterior margin concave. Clypeus with margin slightly sinuous. Maxillary and labial palpi with 3 and 2 palpomeres, respectively (Fig. 2D). Thorax: pronotum wider (0.76 mm) than long (0.58 mm), width less than length of head; anterior margin straight, posterior margin convex, sides strongly constricted at middle; disk glossy, convex centrally (strongly convex on anterior half), with 1 elongated gibbosity on each side; integument finely, sparsely punctate, with erect, sparse orange setae. Scutellum shiny, finely punctate, with sparse setae, posterior margin truncate. Elytra 6.3 times as long as wide (base width) (Fig. 1A), subparallel on anterior 2/3, slightly constricted about this area, becoming narrower towards rounded apex, reaching apex of abdominal ventrite IV; integument coarsely punctate, strongly granulate on posterior third; each puncture with an erect, orange seta. Metathoracic wings 2.2 times longer than wide (Fig. 2E), anterior radial vein (RA) narrowed, cubital 2 (CU2) curved centrally. Legs finely punctate, with dense orange, orange-brown setae. Coxae conical. Trochanters elongate. Femora fusiform, flattened; metafemora slightly longer (Fig. 2A, B). Tibiae elongate, with 2 apical spines: metatibia slightly longer. Tarsomere I shorter in length than II-IV together; claws simple. Abdomen: shiny, integument shagreened with fine punctures, each puncture with an erect, orange seta, slightly sparser at middle (Fig. 2B).
Variation. Type material varies in total length from 4.38 mm to 5.68 mm, and in width from 0.78 mm to 1.0 mm.
Taxonomic summary
Etymology. This species is named after our friend Esteban Berrones from Alta Cima, El Cielo Biosphere Reserve, for his dedication to protect the biodiversity of El Cielo.
Type material. Holotype male: Mexico: Tamaulipas, Gómez Farías, Reserva de la Biosfera El Cielo, Malaise trap, 23°05’56.2” N, 99°12’17.1” W, 19/Abril/2021, 1,200 m snm, M. Janda et al. leg. (CNIN). Paratypes. Same data as holotype except: 23°07’48.6” N, 99°12’44.3” W, 01/Septiembre/2020, 1,400 m snm (CIUM, 1); 23°05’56.2” N, 99°12’17.1” W, 19/Abril/2021, 1,200 m snm (CNIN, 2); 23°07’48.6” N, 99°12’44.3” W, 04/Junio/2021, 1,400 m snm (CIUM, 1); light trap, 23°05’02.0” N, 99°11’50.4” W, 17/Julio/2021, 1,200 m snm (CNIN, 1).
Figure 2. Telegeusis estebani sp. nov., holotype male: A) dorsal habitus; B) ventral habitus; C) head, dorsal view; D) palpi, ventral view; E) paratype, metathoracic wing. Scale bars = 1 mm (A-B, E); 0.5 mm (C-D).
Diagnosis. It may be distinguished by the wide, gibbous interocular area, which in other species is narrower and only gibbous centrally. Telegeusis flavafusca sp. nov. is similar to T. hidalguensis Zaragoza-Caballero, 2015 but differs as follows: integument mostly yellowish-brown (mostly pale yellow in T. hidalguensis); elytra shortened, reaching base of abdominal ventrite 3 (longer in T. hidalguensis, surpassing middle of abdomen).
Holotype male (CNIN). Size: length 4.4 mm; width 0.75 mm. Color: mostly yellowish brown, darker on some areas; mouthparts reddish brown; posterior half of elytra, and apical palpomeres brown. Head: flat, 1.2 times longer than wide, at eye level, distance between outer margins of eyes slightly wider than pronotum (0.48 mm). Interocular area convex, forming conspicuous gibba (Fig. 3C). Distance between eyes and pronotum 1.4 times length of eye. Integument shiny and shagreened, finely, sparsely punctate, coarser behind eyes; each puncture with an erect, orange seta; setae longer around the eyes. Antennal tubercles slightly pronounced (Fig. 3C). Inter-antennal area concave, 1.7 times as wide as length of scape. Antennae longer (1.2 mm) than the last maxillary palpomere (0.82 mm) (Fig. 3A, B); scape unwidened. Eyes slightly prominent, longer (0.23 mm) than wide (0.11 mm); interocular distance 4.25 times width of eyes. Frons almost straight; anterior margin concave. Clypeus with very pronounced median tooth at anterior margin. Maxillary and labial palpi with 3 and 2 palpomeres, respectively (Fig. 3D). Thorax: pronotum just wider (0.48 mm) than long (0.46 mm), width less than length of head; anterior margin straight, posterior margin convex, sides strongly constricted at middle; disk glossy, slightly convex centrally, with 1 irregular, elongated gibbosity on each side; integument finely, sparsely punctate, with erect, sparse orange setae. Scutellum shiny, finely punctate, with few setae, posterior margin truncate. Elytra 4.15 times as long as wide (base width) (Fig. 3A), subparallel on anterior 2/3, slightly narrower in posterior third, apex rounded, reaching base of abdominal ventrite III; integument coarsely punctate, strongly granulate on posterior third; each puncture with erect, orange seta. Metathoracic wings 2.35 times longer than wide (Fig. 3E), anterior radial vein (RA) distinctly sclerosed, cubital 2 (CU2) curved, jugal lobe widened. Legs finely punctate, with sparse orange setae from coxae to femora, and dense orangish-brown setae on tibiae and tarsi. Coxae conical; metacoxae shorter. Trochanters not elongated. Femora fusiform, flattened; metafemora slightly longer (Fig. 3A, B). Tibiae elongate, with 2 apical spines: metatibia slightly longer. Tarsomere I shorter in length than II-IV together; claws simple. Abdomen: shiny, integument shagreened with fine punctures, each puncture with an erect, orange seta, slightly sparser at middle (Fig. 3B).
Variation. Type material varies in total length from 3.58 to 4.54 mm, and in width from 0.66 to 0.8 mm. One specimen has integument slightly lighter.
Taxonomic summary
Etymology. From Latin “flavus” (yellow) and “fuscus” (brown), referring to the main coloration of integument.
Type material. Holotype male: Mexico: Tamaulipas, Gómez Farías, Reserva de la Biosfera El Cielo, Malaise trap, 23°08’03.5” N, 99°11’16.5” W, 1/Septiembre/2020, 1,000 m snm, M. Janda et al. leg. (CNIN). Paratypes. Same data as holotype except 23°08’03.5” N, 99°11’16.5” W, 1/Septiembre/2020, 1,000 m snm (CIUM, 1); 23°08’08.2” N, 99°10’54.5” W, 05/Abril/2021, 800 m snm (CNIN, 2).
Diagnosis. The coloration resembles T. moroni Zaragoza-Caballero, 2015, T. brachypalpus Zaragoza-Caballero, 2015 and T. granulatus Zaragoza-Caballero & Rodríguez-Vélez, 2011. It differs from T. moroni by the labial palpi 2-segmented (3-segmented in T. moroni); from T. brachypalpus by the length of maxillary and labial palpi, about half of antennae (reaching basal third of antennae in T. brachypalpus; and from T. granulatus by the head shagreened (granulated in T. granulatus), and elytral shortened, about 3.5 times as long as wide (about 6 times as long as wide in T. granulatus).
Holotype male (CNIN). Size: length 4.2 mm; width 0.8 mm. Color: mostly brown; mouthparts reddish brown; frons, meso and metaventrite, base of elytra, trochanters, femora orange brown; apical palpomeres dark brown. Head: flat, 1.25 times longer than wide, at eye level, distance between outer margins of eyes slightly wider than pronotum (0.6 mm). Interocular area slightly convex, forming small gibba (Fig. 4C). Eyes-pronotum distance 1.45 times length of eye. Integument shiny, shagreened, finely punctate, coarser behind eyes; each puncture with an erect, orange seta; setae longer around eyes. Antennal tubercles slightly pronounced (Fig. 4C). Inter-antennal area concave; 1.95 times as wide as length of scape. Antennae longer (1.22 mm) than last maxillary palpomere (0.63 mm) (Fig. 4A, B); scape prominent. Eyes reduced, longer (0.21 mm) than wide (0.12 mm); interocular distance 5.25 times width of eyes. Frons slightly concave; anterior margin concave. Clypeus with very pronounced median tooth at anterior margin. Maxillary and labial palpi with 3 and 2 palpomeres respectively (Fig. 4D). Thorax: pronotum wider (0.56 mm) than long (0.46 mm), width less than length of head; anterior margin straight, posterior margin convex, sides slightly constricted at middle; disk glossy, slightly convex centrally, with 1 elongated gibbosity on each side; integument finely, sparsely punctate, with erect, sparse orange setae. Scutellum shiny, coarsely punctate, with few setae, posterior margin truncate. Elytra 3.55 times as long as wide (base width) (Fig. 4A), subparallel on anterior half, gradually narrowing towards rounded apex, reaching apex of abdominal ventrite III; integument coarsely punctate, strongly granulate on posterior third; each puncture with an erect, orange seta.
Figure 3. Telegeusis flavafusca sp. nov., holotype male: A) dorsal habitus; B) ventral habitus; C) head, dorsal view; D) palpi, ventral view; E) paratype, metathoracic wing. Scale bars = 1 mm (A-B, E); 0.5 mm (C-D).
Metathoracic wings 2.3 times longer than wide (Fig. 4E), anterior radial vein (RA) distinctly sclerosed, cubital 2 (CU2) slightly curved. Legs finely punctate, with sparse orange setae from coxae to femora, and dense orangish-brown setae on tibiae and tarsi. Coxae conical; metacoxae shorter. Trochanters not elongated. Femora fusiform, flattened; metafemora slightly longer (Fig. 3A, B). Tibiae elongate, somewhat arched, with 2 apical spines; metatibia longer. Tarsomere I shorter in length than II-IV together; claws simple. Abdomen: shiny, integument shagreened with fine punctures, each puncture with an erect, orange seta (Fig. 4B).
Figure 4. Telegeusis luismartinezi sp. nov., holotype male: A) dorsal habitus; B) ventral habitus; C) head, dorsal view; D) palpi, ventral view; E) paratype, metathoracic wing. Scale bars = 1 mm (A-B, E); 0.5 mm (C-D).
Variation. Type material varies in total length from 4.12 to 5.06 mm, and in width from 0.76 to 0.95 mm.
Taxonomic summary
Etymology. This species is named after our friend Luis Martínez from Gómez Farías, El Cielo Biosphere Reserve, for his dedication to protect the biodiversity of El Cielo.
Type material. Holotype male: Mexico: Tamaulipas, Gómez Farías, Reserva de la Biosfera El Cielo, Malaise trap, 23°05’56.2” N, 99°12’17.1” W, 19/Marzo/2020, 1,200 m snm, M. Janda et al. leg. (CNIN). Paratypes. Same data as holotype except 23°05’56.2” N, 99°12’17.1” W, 19/Marzo/2020, 1,200 m snm (CNIN, 2; CIUM, 2); light trap, 23°05’48.3” N, 99°11’59.4” W, 19/Abril/2021, 1,200 m snm (CNIN, 1).
Key to species of Telegeusis (modified from Zaragoza-Caballero, 2015)
1. Elytra less than 4 times as long as wide 2
1’ Elytra more than 4 times as long as wide 10
2. Elytra short, not reaching hind coxae, almost as long as wide T. schwarzi Barber, 1952
2’ Elytra at least reaching hind coxae, longer than wide 3
3. Integument mostly from dark brown to black 4
3’ Integument mostly from yellow to amber 6
4 Maxillary and labial palpi at most reaching half of the antennae T. luismartinezi sp. nov.
4’ Maxillary and labial palpi surpassing half of the antennae 5
5. Occipital region, genae, gula, clypeus, and pronotum orange; total length 4.5 mm T. texensis Fleenor & Taber, 2001
5’; Integument dark brown, pronotum and basal part of the legs yellow; total length 3.5 mm T. panamensis Alan & Huntton, 1969
6. Maxillary and labial palpomeres longer than antennae 7
6’ Maxillary and labial palpomeres commonly no longer than antennae 8
7. Integument mostly yellow; last maxillary and labial palpomeres yellow; total length 7.0 – 8.0 mm T. nunifer Martin, 1931
7’ Integument mostly amber; last maxillary and labial palpomeres black; total length 3.3 – 4.6 mm T. austellus Zaragoza-Caballero & Rodríguez-Vélez, 2011
8. Maxillary and labial palpi black, shorter than antennae T. boreios Zaragoza-Caballero & Rodríguez-Vélez, 2011
8’ Maxillary and labial palpi brown, as long or longer than antennae 9
9. Eyes small; maxillary and labial palpi as long as antennae T. sonorensis Zaragoza-Caballero & Rodríguez-Vélez, 2011
9’ Eyes prominent; maxillary and labial palpi longer than antennae T. chamelensis Zaragoza-Caballero, 1975
10. Integument yellow to amber 11
10’ Integument brown to black 16
11. Elytra more than 5 times as long as wide 12
11’ Elytra less than 5 times as long as wide 13
12. Head twice as long as pronotum T. debilis Horn, 1895
12’ Head as long as pronotum T. cieloensis sp. nov.
13. Maxillary and labial palpi black, with 4 and 3 palpomeres respectively, longer than antennae T. glessum Zaragoza-Caballero & Rodríguez-Vélez, 2011
13’ Maxillary and labial palpi yellow or amber, with 3 and 2 or 2 and 1 palpomeres respectively, shorter than antennae 14
14. Eyes small; maxillary and labial palpi with 2 and 1 palpomeres respectively T. dissimilis Zaragoza-Caballero, 2015
14’ Eyes prominent; maxillary and labial palpi with 3 and 2 palpomeres respectively 15
15. Integument mostly yellow; head about as long as wide; maxillary and labial palpi reaching half of antennae T. hidalguensis Zaragoza-Caballero, 2015
15’ Integument mostly amber; head longer than wide; maxillary and labial palpi reaching apical third of antennae T. flavafusca sp. nov.
16. Maxillary and labial palpi with 4 and 3 palpomeres respectively 17
16’ Maxillary palpi with 3 palpomeres; and labial palpi with 3 or 2 palpomeres 18
17. Integument mostly dark brown; head shagreened; elytra 4.5 times as long as wide T. orientalis Zaragoza-Caballero, 1990
17’ Integument black; head granulated; elytra almost 6.5 times as long as wide T. granulatus Zaragoza-Caballero & Rodríguez-Vélez, 2011
18. Maxillary and labial palpi with 3 palpomeres T. moroni Zaragoza-Caballero, 2015
18’ Maxillary and labial palpi with 3 and 2 palpomeres respectively 19
19. Last maxillary palpomere about as long as antennae; elytral length more than 6 times as long as wide T. estebani sp. nov.
19’ Last maxillary palpomere reaching half of the antennae; last labial palpomere shortened; elytral length less than 6 times as long as wide T. brachypalpus Zaragoza-Caballero, 2015
Discussion
Most of the species of Telegeusis Horn, 1895 which have been recorded in northern Mexico and USA, show a Nearctic distributional pattern with a disjunct Neotropical distribution (Zaragoza-Caballero, 2015; Zaragoza-Caballero & Rodríguez-Vélez, 2011); however, this may be due to lack of sampling (e.g., they are not recorded in most of Central America) and does not mean that they are not equally or mostly distributed in the Neotropical region. Interestingly, this distributional species pattern is similar to those of North American Omethidae, one of the proposed sister groups of telegeusids (Bocak et al., 2018; Ramsdale, 2010).
Telegeusis are commonly recognized by their 2-3 segmented labial palpi, with the last palpomere greatly elongated, similar to maxillary palpi; and antennae with short-filiform antennomeres (Ivie, 2002). These characters are used to distinguish most species of the genus, mainly the length of last labial and maxillary palpomeres, compared with antennal length (Zaragoza-Caballero, 2015; Zaragoza-Caballero & Rodríguez-Vélez, 2011). One other important character used in the specific diagnoses of Telegeusis is the integument coloration, which is greatly consistent across the species. In different groups of insects, the color of the integument is associated to the environment, altitudinal (commonly darker coloration as altitude increases) gradients or feeding habits (Badejo et al., 2020; Stanbrook et al., 2021). However, in Telegeusis does not appear to be related to elevation, since there are species with dark and light colors in both low and high areas. Regarding the environment, we found the new species coexisting in the same habitat (cloud forest), between 1,000 and 1,600 m, but nothing else is known about their biology.
Although we confirm the usefulness of the color pattern in Telegeusis, we recommend combining it with other characters (e.g., elytral length, head sculpture). However, further studies may help to corroborate the informativeness of this type of character under a phylogenetic framework.
Acknowledgements
The first author is indebted with the local community of the Reserva de la Biosfera El Cielo for their hospitality and facilities provided during fieldwork. We thank Esteban Berrones and Luis Martínez for their support in the fieldwork. To Susana Guzmán (IBUNAM) for helping with the use of the Zeiss equipment in LANABIO. This research was founded by the Consejo Nacional de Humanidades, Ciencias y Tecnologías DICB No.435282471, and Universidad Nacional Autónoma de México PAPIIT IN206818 and IN218520.
References
Badejo, O., Skaldina, O., Gilev, A., & Sorvari, J. (2020). Benefits of insect colours: a review from social insect studies. Oecologia, 194, 27–40. https://doi.org/10.1007/s00442-020-04738-1
Ivie, M. A. (2002). The transfer of Pseudokarumia Pic from Dascillidae (Karumiinae) to Telegeusidae, with a key to world genera of Telegeusidae. The Coleopterist Bulletin, 56, 582–584. https://doi.org/10.1649/0010-065X(2002)056[0582:TTOPPF]2.0.CO;2
Kundrata, R., Kundrata, R., Bocakova, M., & Bocak, L. (2014). The comprehensive phylogeny of the superfamily Elateroidea (Coleoptera: Elateriformia). Molecular Phylo- genetics and Evolution, 76, 162–171. https://doi.org/10.1016/j.ympev.2014.03.012
Lawrence, J. F. (2010). Telegeusidae Leng, 1920. In R. A. B. Leschen, R. G. Beutel, & J. F. Lawrence (Eds.), Handbook of Zoology, Arthropoda: Insecta, Coleoptera, beetles morphology and systematics, Vol. 2 (pp. 123–126). Berlin, New York: Walter de Gruyter. https://doi.org/10.1515/9783110911213.123
López-Pérez, S., & Zaragoza-Caballero, S. (2021). Two new genera of Telegeusidae (Coleoptera) from Mexico. Revista Mexicana de Biodiversidad, 92, e923613. https://doi.org/10.22201/ib.20078706e.2021.92.3613
Miller, S.R. (2002). Telegeusidae Leng, 1920. In R. H. Arnett, M. C. Thomas, P. E. Skelley, & H. F. D. Phil (Eds.), American beetles, Volume II: Polyphaga: Scarabaeoidea through Curculionoidea (pp. 179–180). Gainsville, Florida: CRC Press.
Ramsdale, A. S. (2010). Omethidae LeConte, 1861. In R. A. B. Leschen, R. G. Beutel, & J. F. Lawrence (Eds.), Handbook of zoology, Arthropoda: Insecta, Coleoptera, beetles morphology and systematics, Vol. 2 (pp. 149–153). Berlin, New York: Walter de Gruyter. https://doi.org/10.1515/9783110911213.149
Stanbrook, R. A., Harris, W. E., Wheater, C. P., & Jones, M. (2021). Evidence of phenotypic plasticity along an altitudinal gradient in the dung beetle Onthophagus proteus. PeerJ, 9, e10798. https://doi.org/10.7717/peerj.10798
Zaragoza-Caballero, S. (2008). Nueva especie de Pseudotelegeusis (Coleoptera: Telegeusidae) del estado de Colima, México. Revista Mexicana de Biodiversidad, 79, 369–372. https://doi.org/10.22201/ib.20078706e.2008.002.573
Zaragoza-Caballero, S. (2015). Nuevos telegeúsidos (Coleoptera: Telegeusidae) de Hidalgo, México. Dugesiana, 22, 179–186. https://doi.org/10.32870/dugesiana.v22i2.4730
Zaragoza-Caballero, S., & Rodríguez-Vélez, B. (2011). Five new species of Telegeusis Horn, 1895 (Coleoptera: Telegeusidae) from Mexico, with a key to the species. Zootaxa, 2917, 59–68. https://doi.org/10.11646/zootaxa.2917.1.4
Gabriela Diaz a, b, *, Vanesa Pellegrini-Piccini a, Liliana Moreno d, Martín Palma c, e, Vanesa Bentancourt c y Valeria Corbalán f
a Universidad Nacional de Cuyo-Sede Malargüe, Campus Educativo Municipal, Facultad de Ciencias Exactas y Naturales, Rosario Vera Peñaloza y Beltrán, 5613 Malargüe, Mendoza, Argentina
b Universidad Nacional de Cuyo, Instituto de Ingeniería y Ciencias Aplicadas a la Industria-CONICET, Facultad de Ciencias Aplicadas a la Industria, Bernardo de Yrigoyen Núm. 375, 5600 San Rafael, Mendoza, Argentina
c Instituto de Educación Física Núm. 9-016 “Jorge E. Coll” Dirección General de Escuelas-Sede Malargüe, Tecnicatura en Conservación de la Naturaleza, Campus Educativo Municipal, Rosario Vera Peñaloza y Beltrán, 5613 Malargüe, Mendoza, Argentina
d Universidad Nacional de San Luis, Facultad de Química Bioquímica y Farmacia, Ejército de los Andes Núm. 950, 5700 San Luis, Argentina
e Ministerio de Ambiente y Desarrollo Sustentable de la Provincia de Mendoza, Dirección de Recursos Naturales Renovables, Delegación Malargüe, San Martín Norte Núm. 352, 5613 Malargüe, Mendoza, Argentina
f Instituto Argentino de Investigaciones de Zonas Áridas (CCT Mendoza-CONICET), Av. Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, Argentina
*Autor para correspondencia: gdiaz@infoar.net (G. Diaz)
Recibido: 02 octubre 2023; aceptado: 15 agosto 2024
Resumen
La ranita del Pehuenche, Alsodes pehuenche, es endémica de los Andes centrales de Argentina y Chile, ha sido categorizada en peligro crítico por la UICN y entre sus amenazas se encuentran la ruta internacional que atraviesa los arroyos que habita, la presencia del hongo quitridio, los salmónidos exóticos invasores, el ganado y el cambio climático. El objetivo de este trabajo fue evaluar el estado actual de conservación de A. pehuenche en el valle Pehuenche para conocer tendencias poblacionales, el impacto de las amenazas y futuras acciones de manejo. Se realizaron 14 salidas de campo durante 3 temporadas (2021-2023) y se muestrearon 12 arroyos usando la técnica de encuentro visual nocturno. Se delimitaron y nombraron 7 subpoblaciones: Nacientes, del Límite, Pichintur, Rial Rojas, Nueva, Campanaria y Cajón Largo. Los resultados muestran conteos de adultos (5.82 en 200 m2 y 13.64 por hora) y de larvas (6.24 en 200 m2 y 17.76 por hora). Éstos no variaron significativamente entre temporadas, pero fueron mayores en enero y febrero. Con base en la conectividad y las amenazas, los índices del estado de conservación permiten priorizar las subpoblaciones como unidades de conservación, de las cuales la del Límite requiere esfuerzos más urgentes.
Palabras clave: Especie amenazada; Encuentro visual; Conectividad; Subpoblaciones; Priorización de conservación
Population monitoring and conservation status of the Pehuenche frog (Alsodes pehuenche) in the valle Pehuenche, Mendoza, Argentina
Abstract
The Pehuenche spiny-chest frog, Alsodes pehuenche, is endemic to the Central Andes of Argentina and Chile. It has been categorized as critically endangered by the IUCN and its threats include the international road that crosses the streams inhabited by the species, the presence of the chytrid fungus, invasive exotic salmonids, livestock, and climate change. The objective of this work was to evaluate the current conservation status of A. pehuenche in the Pehuenche Valley as a basis for understanding population trends, the impact of threats, and future management actions. Fourteen field trips were conducted during 3 seasons (2021-2023) and 12 streams were sampled using the nocturnal visual encounter technique. Seven subpopulations were delimited and named: Nacientes, del Límite, Pichintur, Rial Rojas, Nueva, Campanaria, and Cajón Largo. The results show counts of adults (5.82 in 200 m2 and 13.64 per hour) and larvae (6.24 in 200 m2 and 17.76 per hour). These did not vary significantly between seasons but were higher in January and February. According to connectivity and threats, the conservation status indices allow us to prioritize the subpopulations as conservation units, with del Límite being the one that requires the most urgent efforts.
Los anfibios son el grupo de vertebrados más amenazados de nuestro planeta, varias son las causas responsables de la disminución de sus poblaciones (Grant et al., 2020; Green et al., 2020; Luedtke et al., 2023). Más de 45% de la diversidad de anfibios del mundo se distribuyen en el Neotrópico (Kacoliris et al., 2022). Alrededor de 25% de las especies de Argentina son endémicas (Vaira et al., 2017), 37% de ellas en Argentina y Chile se encuentran en disminución, mientras que del 22% de las especies no se conoce su tendencia poblacional (Kacoliris et al., 2022). Las amenazas más importantes con las que se asocian la disminución poblacional o extinciones locales son los depredadores invasores, enfermedades emergentes y la ganadería (Kacoliris et al., 2022; Velasco et al., 2016).
La ranita del Pehuenche, Alsodes pehuenche, fue descrita por Cei (1976). Luego de estudios citogenéticos realizados entre 1983 y 2003 (Cuevas y Formas, 2003), se renueva el interés sobre la especie en Argentina debido a las obras viales sobre la ruta internacional ARG145 – CH115, cuya pavimentación desvió el curso de 5 afluentes del arroyo Pehuenche con presencia de la especie (Corbalán et al., 2010). La ranita del Pehuenche es una especie endémica y su distribución está restringida a los Andes centrales de Argentina y Chile (Corbalán et al., 2010, 2023; Correa et al., 2013, 2018, 2020). Habita arroyos de montaña en ecosistemas de vegas o mallines entre 2,150 y 2,825 m snm (Corbalán et al., 2023). Los arroyos poseen lechos pedregosos y una fina capa de sedimentos. La especie presenta dimorfismo sexual y como otras especies de Alsodes, tiene larvas de desarrollo prolongado con juveniles y adultos de hábitos acuáticos (Cei, 1976, 1980; Herrera y Velázquez, 2016a; Úbeda, 2021). Las larvas son de gran tamaño y pasan al menos 4 años en los cuerpos de agua permanentes hasta completar el ciclo larval (Corbalán et al., 2014). Se ha reportado la puesta de huevos ocultos bajo rocas o en oquedades en las márgenes de los arroyos (Corbalán et al., 2014; Piñeiro et al., 2020). Estas cavidades son utilizadas también como refugio por los adultos (Cei, 1980; Correa et al., 2013; Herrera y Velázquez, 2016b).
Los datos de conteos disponibles corresponden a muestreos de tramos cortos de arroyos en Argentina y Chile (Corbalán et al., 2010, 2023; Correa et al., 2013). La coexistencia de individuos en una cavidad durante el día sugiere densidades elevadas en una categoría de microhábitat (Correa et al., 2013). Han ocurrido eventos de mortalidad y se han reportado nuevas poblaciones (Corbalán et al., 2023; Correa et al., 2018), lo cual hace necesaria una evaluación actualizada de su estatus de conservación y tendencia poblacional.
Las evaluaciones del estado de conservación de la ranita del Pehuenche la han colocado en la categoría más alta de amenaza: “en peligro” por la Asociación Herpetológica Argentina (Vaira et al., 2012), cuarta en orden de prioridad entre los 58 anfibios evaluados de Chile (Vidal et al., 2024) y “en peligro crítico” por la Unión Internacional para la Conservación de la Naturaleza (IUCN, 2019), según los criterios B1ab basados en su extensión de presencia estimada y su disminución continua estimada, provocada principalmente por la pavimentación de la ruta. El área de ocupación de la especie (AOO sensu UICN) estimada actualmente es de 4.84 km2 y su extensión de presencia (EOO sensu UICN) 497.9 km2 (Corbalán et al., 2023). En cuanto a las amenazas consideradas en la categorización de IUCN (2019), se enumeran el desvío de los cursos de agua por la construcción de la ruta, el impacto del ganado, el cambio climático, la presencia del hongo quitridio (Batrachochytrium dendrobatidis) y la depredación por salmónidos exóticos invasores tales como la trucha arcoíris (Oncorhynchus mykiss)y trucha marrón (Salmo trutta) (Corbalán et al., 2023; Ghirardi et al., 2014; Zarco et al., 2020).
Si bien los avances en el conocimiento de la distribución de A. pehuenche han sido importantes en los últimos años, aún se desconocen aspectos básicos de la ecología, sistemática, reproducción, comportamiento y estrategias ecofisiológicas de esta especie en los humedales de altura. Por tratarse de una especie endémica y amenazada, la historia de vida y demografía son fundamentales para la evaluación del estatus de conservación (Luja et al., 2015). Los programas de seguimiento de poblaciones son necesarios para identificar y detectar disminuciones que amenacen la persistencia de poblaciones y deben llevarse a cabo en un marco de gestión adaptativa que permita realizar monitoreos que maximicen la detección y minimicen el esfuerzo (Pollock, 2006, Yoccoz et al., 2001). A su vez, la definición de subpoblaciones es una herramienta útil que permite definir su estado actual y priorizar acciones de conservación (Velasco, 2018).
El objetivo de este trabajo fue evaluar el estado de conservación actual de A. pehuenche en el valle Pehuenche. Esta información es fundamental para estimar tendencias poblacionales a largo plazo, evaluar el impacto de las amenazas y el éxito de futuras acciones de manejo. En este trabajo se muestran los primeros datos del programa de monitoreo iniciado en 2021 en el valle Pehuenche. A partir del mismo, se definen y delimitan subpoblaciones como unidades de conservación sobre las que se deben priorizar las acciones.
Materiales y métodos
El área de estudio corresponde al valle Pehuenche, en el lado argentino de la zona limítrofe entre Argentina y Chile; forma parte de los Andes centrales, en el departamento de Malargüe, suroeste de la provincia de Mendoza. Las precipitaciones anuales son de 400 a 600 mm (Rivera et al., 2018) y están influidas por los vientos provenientes del Pacífico sur, creando un gradiente de precipitación y humedad de oeste a este con fuertes nevadas en invierno (Garreaud et al., 2009). Durante la primavera-verano, el deshielo alimenta los humedales, denominados localmente vegas o mallines, donde se asientan familias con su ganado en los puestos, denominados reales o riales. El área es considerada un corredor ecológico y cultural trashumante de gran importancia (Llano et al., 2021).
La cuenca del arroyo Chico en Argentina incluye las subcuencas donde se distribuye A. pehuenche: arroyos Pehuenche y Callao (Corbalán et al., 2023). El arroyo Chico es afluente del río Grande en el departamento de Malargüe, provincia de Mendoza. En Chile, el área de distribución de la especie se ubica en la cuenca del río Maule, con 2 subpoblaciones posiblemente aisaldas: laguna del Maule y Lo Aguirre (Correa et al., 2013, 2018).
Durante 3 temporadas se muestrearon 14 arroyos tributarios del arroyo Pehuenche (fig. 1). Doce de los 14 arroyos fueron muestreados exhaustivamente y en los 2 restantes (B0 y A8), solo se constató la presencia de la especie. La especie fue observada en la desembocadura de los arroyos de primer orden en el arroyo Pehuenche.
Siguiendo a Corbalán et al. (2010) y Prado et al. (2019), los arroyos de primer orden que desembocan en el arroyo Pehuenche se distinguen, según su ubicación respecto a este último, en arroyos A que se ubican hacia el sur y son atravesados por la ruta Núm. 145, mientras que los arroyos B se ubican al norte del arroyo Pehuenche (fig. 1). A los arroyos con bifurcaciones y cursos de agua paralelos a menos de 100 m de distancia se los denominó con el mismo nombre. Durante el estudio, se tomaron datos de temperatura del agua con termómetro de mercurio, así como pH, oxígeno disuelto y conductividad con sonda multiparamétrica Lutron WA-2017SD, y de temperatura ambiente mediante la aplicación yr (Jensen et al., 2007). La conductividad fue baja, entre 0 y 181.56 μS/cm (n = 65), el valor medio del pH estuvo cercano a la neutralidad 6.41 ± 1.2, variando entre 3.29 y 8.25 (n = 77) y la concentración de oxígeno disuelto varió entre 1.3 y 15.8 mg/l (n = 16). La temperatura ambiente durante los muestreos osciló entre 2 y 21 ºC (n = 89), y la temperatura del agua entre 2 y 17.6 ºC (n = 75).
Se realizaron un total de 14 salidas de campo con una duración de 2 a 4 días/noches (tabla 1). Las 14 salidas se distribuyeron en 3 temporadas; cada una se inicia con el deshielo (octubre-noviembre-diciembre) y finaliza con la caída de las primeras nevadas (marzo-abril). En enero de 2021 se iniciaron muestreos preliminares diurnos y nocturnos consistentes en encuentros visuales sin manipulación de individuos. Los muestreos nocturnos iniciaron luego de la puesta del sol que coincide con el pico de actividad de los individuos adultos. Se optó por continuar con los muestreos nocturnos debido a que no se obtuvieron diferencias significativas en la cantidad de larvas entre el día y la noche (W = 191, p = 0.859).
Figura 1. Arroyos muestreados en la cuenca del valle Pehuenche, distribuidos en 7 subpoblaciones locales: I. Nacientes, II. del Límite, III. Pichintur, IV. Rial Rojas, V. Nueva, VI. Campanaria, VII. Cajón Largo. Los arroyos de cada subpoblación se indican con el mismo color.
Se contabilizaron adultos, juveniles y larvas usando la técnica de encuentro visual (Crump y Scott, 1994). Los transectos fueron de rumbo variable siguiendo el curso del arroyo y el ancho fue de 1 m a cada lado del arroyo. Cada transecto se realizó con 2 a 5 observadores. Siguiendo las recomendaciones de Pereyra et al. (2021), al menos un observador tenía experiencia previa y las salidas se programaron en días con condiciones meteorológicas similares: días sin lluvias, mayormente despejados y con vientos leves a moderados.
Para los estadios maduros se registró sexo (macho, hembra e indeterminado). Se registró punto GPS y hora de inicio y de finalización del transecto. Se calcularon las abundancias relativas como individuos/área e individuos/tiempo. El área de los arroyos se consideró como el tramo muestreado en metros multiplicado por 2 m de ancho. Los individuos contabilizados estuvieron a no más de 1 m de distancia del curso del arroyo, cuando éste está claramente definido, y solo se observaron más dispersos en las zonas húmedas de vegas. Para el cálculo de individuos/tiempo se usó el tiempo en minutos de la duración del muestreo (Pereyra et al., 2021).
Adicionalmente, se realizaron muestreos en transectos de 800 m de longitud en los márgenes de la calzada: 10 en la temporada estival de 2021 y 5 en las temporadas siguientes. Este registro se realizó ya que en enero de 2018 se observaron individuos deshidratados o muertos, adyacentes a un cordón o bordillo de 20 cm de altura y 15 cm de ancho.
Se definieron subpoblaciones, siguiendo a Velasco (2018), formadas por un grupo de arroyos que contienen individuos de A. pehuenche, con hábitat adecuado (sensu Corbalán et al., 2010; Correa et al., 2013) y cuya distancia entre ellos no fue mayor a 500 m lineales. De este modo, los 14 arroyos iniciales fueron agrupados en 7 subpoblaciones (fig. 1). Los nombres de las subpoblaciones se relacionan con referencias a las características del paisaje donde se encuentran, excepto la subpoblación denominada Nueva. Se calculó el índice de conectividad (IC) entre subpoblaciones de acuerdo con la fórmula de Lin (2009), modificada por Velasco (2018): IC = A1*(1/D1*T1) + A2*(1/D2*T2), donde Ai es el área (= tamaño poblacional) de la población local vecina hacia el lado i, Di es la distancia a la población local vecina hacia el lado i y Ti la presencia de truchas en hábitat intermedios (presencia multiplica por 2, ausencia multiplica por 1). Para estimar el área correspondiente a cada subpoblación, se usaron las coordenadas GPS de los muestreos de campo y en el caso de las 2 subpoblaciones donde solo se constató presencia, se estimó tamaño del área en Google Earth Pro.
Tabla 1
Temporadas y subpoblaciones en las que se realizaron los muestreos de Alsodes pehuenche. Se indican la cantidad de muestreos realizados en la ruta y los nombres de los arroyos (tipos A, B; P: arroyo Pehuenche) muestreados por subpoblación.
Temporada
Mes
Quincena primera (1) segunda (2)
Ruta
Subpoblaciones
I
II
III
IV
V
VI
VII
1 (2021)
enero
1
2
B0
A1, A2
enero
2
6
A1, A2, A3, A4
A5
A7
febrero
2
1
A1, A2
B2
B3
abril
2
1
A1, A2
A5
A8
2 (2021-2022)
noviembre
2
1
A1, B1
B2
A5
A7
febrero
1
A2
B3
marzo
1
A1
B2
marzo
2
B1
B3
3 (2022-2023)
noviembre
2
1
A5
diciembre*
1
1
A1, A3, B1
B2, B2
A5-B3
A6
A7
enero
1
1
A2, P
A5, B3, P
A6, P
A7, P
febrero
2
1
A1, P
A7, P
marzo
2
A3, P
B4, P
B6, P
Las amenazas que se registraron durante los conteos se consideraron de manera cualitativa, en una escala de 0 a 3, de acuerdo con la intensidad de la amenaza (Velasco, 2018). En la tabla 2 se muestran las amenazas consideradas: 1) la presencia de salmónidos que fueron registrados por encuentro visual; 2) la sequía observada de arroyos que podría ser reversible según la época del monitoreo y cárcavas permanentes cuya profundidad va en aumento; 3) la presencia de ganado, principalmente vacuno y caprino, el primero es el que podría causar mayor impacto debido al mayor uso de las vegas; 4) mortalidad de individuos por causas indeterminadas; 5) la ruta, considerada como una barrera para la dispersión, ya sea porque las alcantarillas no fueron diseñadas como pasos de fauna o porque la altura del cordón o bordillo construido sobre la misma es muy alto para el libre tránsito de ranas juveniles y adultas. Y a pesar de que la ruta representa una barrera, la construcción posterior de rampas, con el objetivo de mitigar este impacto, se considera una acción concreta de conservación. Se registró, además, la presencia de depredadores no acuáticos potenciales y de otras especies de anfibios, pero ninguno de estos casos se considera como amenaza, aunque podrían representar interacciones interespecíficas negativas.
El estado de conservación (EC) relativo de cada subpoblación se obtuvo a partir de la sumatoria de los valores asignados al índice de conectividad y las amenazas (Velasco, 2018). Los valores fueron estandarizados (ECE = EC subpoblación – promedio de todos los EC/desviación estándar de todos los EC) y el valor resultante se multiplicó por -1. Así, los valores negativos de ECE quedaron relacionados con situaciones menos favorables. Se establecieron 2 categorías de prioridades de conservación: baja a media (valores positivos de ECE) y alta (valores negativos de ECE). Los valores altos son los que requieren medidas más urgentes de manejo (Velasco, 2018).
Tabla 2
Variables consideradas para evaluar estado de conservación de las subpoblaciones de Alsodes pehuenche en el valle Pehuenche. IC: Índice de conectividad. Valores 0 a 3 son los que se le asignan a cada variable.
Variables
0
1
2
3
Área (m2)
7,500-10,000
5,000-7,500
2,500-5,000
1,000-2,500
IC1
7.51-10
5.1-7.5
2.51-5
0-2.5
Amenazas
Efecto especies exóticas: salmónidos
sin
presencia registrada
presencia registrada en más de un conteo
presencia registrada y presencia de anfibio/s con signo de depredación
Fragmentación por la ruta
Sin ruta
Sin cordón o bordillo
Con cordón o bordillo y con rampas
Con cordón o bordillo y sin rampas
Pérdida de hábitat por sequía/ cárcavas
Sin evidencias de sequía ni cárcavas
Algún arroyo parcialmente seco
Algún arroyo parcialmente seco y/o con presencia cárcavas
Algún arroyo completamente seco y/o presencia de individuos muertos por sequía
Efecto del ganado
Sin ganado
Con caprinos
Con vacunos
Con vacunos y cabras
Indeterminada
Sin registro de individuos muertos sin causa aparente
Registro de individuos con manchas en la piel
Registro de individuos con manchas y uno muerto sin causa aparente
Registro de más de un individuo muerto sin causa aparente
1 = Índice de conectividad calculado de acuerdo con Lin (2009), modificado por Velasco (2018).
Se realizaron análisis de los conteos con R versión 4.1.3 y los análisis estadísticos consideraron 0.05 de nivel de significación de alfa. La normalidad de los datos se verificó con la prueba de Shapiro-Wilk. La cantidad de individuos (adultos, juveniles y larvas) por área y por hora no se distribuyó normalmente (W < 0.82934, p < 0.001), por lo que se usaron las pruebas estadísticas no paramétricas de Mann-Whitney-Wilcoxon y de Kruskal-Wallis para 2 grupos y más de 2 grupos, respectivamente. Se evaluó estadísticamente si hubo diferencias entre adultos y juveniles considerados en conjunto versus larvas. También se evaluaron diferencias en relación con variables temporales (temporadas, meses) y espaciales (entre arroyos A y B y entre subpoblaciones). Cuando se obtuvieron diferencias significativas con la prueba de Kruskal-Wallis, se usó el paquete conover.test versión 1.1.5 (2017), basado en Conover-Iman Test para comparaciones múltiples. En todos los casos, los resultados se expresan en mediana, el rango intercuartílico (IQR) como medida de dispersión, los valores mínimos y máximos (min-max) y el tamaño de la muestra (n).
Resultados
Los transectos muestreados alcanzaron un total de 51,292 m lineales recorridos en 181 horas con un esfuerzo de muestreo 12.35 horas/persona. De los 14 arroyos estudiados, todos tuvieron presencia de la especie y el área total fue de 0.04 km². Más de la mitad corresponde a los arroyos denominados A (56.2%).
Cuando se analizaron los conteos respecto de la cantidad de observadores, la correlación de Spearman resultó en valores de rho cercanos a 0, lo que sugiere que no hay correlación lineal (p > 0.05), tanto para individuos postmetamórficos (por área: rho = -0.0427, n = 99; por hora: rho = 0.0975, n = 99), como para larvas (por área: rho = -0.0297, n = 92; por hora rho = -0.0063, n = 92).
A lo largo del periodo de muestreo se observaron 5.82 adultos y juveniles/área (IQR = 9.27, 0-45 min-max, n = 99) y 13.64 adultos y juveniles/hora (IQR = 30.29, 0-108 min-max, n = 99); 6.24 larvas/área (IQR = 31, 0-175 min-max, n = 92) y 17.76 larvas/hora (IQR = 65, 0-367 min-max, n = 92). La comparación entre estadios muestra menos postmetamórficos que larvas, tanto para los conteos de individuos por área (W = 12.524, p < 0.001) como por hora (W = 26.918, p < 0.001).
Tabla 3
Prioridades de conservación para las subpoblaciones de Alsodes pehuenche en el valle Pehuenche, con base en el tamaño de la población, la conectividad y las amenazas.
Sub-población
Área (m2)
IC
Amenazas
ECE
PC1
Ruta
Sequía/cárcavas
Ganado
Muertes indet.
Salmónidos
I. Nacientes
4,044
8.41
0
0
2
0
1
0.88
Baja
II. del Límite
5,744
8.39
2
3
2
2
3
-1.26
Muy alta
III. Pichintur
5,002
9.91
0
0
2
3
2
0.08
Media
IV. Rial Rojas
6,208
6.05
1
3
2
1
2
-0.73
Alta
V. Nueva
2,160
9.61
1
0
2
3
0
-0.19
Media
VI. Campanaria
5,860
5.26
1
0
1
3
3
-0.46
Media
VII. Cajón Largo
9,550
6.64
0
0
1
0
0
1.69
Muy baja
1 PC: Prioridad de conservación calculada de acuerdo con Velasco (2018).
Las comparaciones entre las 3 temporadas no mostraron diferencias significativas para ninguno de los grupos estudiados adultos y juveniles/área (H = 1.9935, df = 2, p = 0.369), adultos y juveniles/hora (H = 1.3599, df = 2, p = 0.5066), larvas/área (H = 0.13936, df = 2, p = 0.933) y larvas por hora (H = 0.6051, df = 2, p = 0.7389). Entre meses, la diferencia no fue significativa para larvas (H = 4.5278, df = 5, p = 0.4762 por área, H = 4.0986, df = 5, p = 0.5353 por hora), en cambio para los postmetamórficos solo fue significativa para los conteos por área (H = 13.97, df = 5, p = 0.0158). Los análisis post hoc indican menores cantidades de adultos y juveniles en febrero respecto a diciembre (p = 0.0075) y enero (p = 0.0021), como también en marzo respecto a enero (p = 0.0185, fig. 2).
Figura 2. Variación mensual de la abundancia de adultos y juveniles (individuos/200 m2) de Alsodes pehuenche. Los meses con diferentes códigos de letras indican diferencias significativas (p < 0.05).
Figura 3. Abundancia de Alsodes pehuenche (individuos/200 m2) entre 5 subpoblaciones. A, Adultos y juveniles; B, larvas. Las subpoblaciones con diferentes códigos de letras indican diferencias significativas (p < 0.05).
De los 14 arroyos del valle Pehuenche con presencia de A. pehuenche, los arroyos A tienen un área de ocupación de 49% respecto de los arroyos B con 51%. Y en los 12 arroyos donde se realizaron conteos por área, éstos fueron mayores en los arroyos A que en los B, tanto para adultos y juveniles (H = 31.977, df = 2, p < 0.001), como para larvas (H = 10.42, df = 2, p = 0.0055).
Se reconocieron 7 subpoblaciones en el valle Pehuenche: Nacientes, del Límite, Pichintur, Rial Rojas, Nueva, Campanaria y Cajón Largo (fig. 1). De éstas, se tienen datos de conteo de 5 (todas, excepto Nacientes y Cajón Largo). Cuando se comparan estas 5 subpoblaciones, se encuentran diferencias significativas tanto para adultos y juveniles/área (H = 17.436, df = 4, p = 0.0016), como para larvas/área (por área H = 23.639, df = 4, p < 0.001). Se registraron más adultos y juveniles en las subpoblaciones Rial Rojas y Nueva, y más larvas en Rial Rojas (fig. 3). Sin embargo, estas diferencias no son significativas cuando se analizan los datos de postmetamórficos/hora (H = 8.8053, df = 5, p = 0.1171) y larvas/hora (H = 4.0986, df = 5, p = 0.5353).
El único lugar donde se encontraron solo larvas fue en una charca de la subpoblación del Límite. Éstas coexistían con larvas y adultos de rana de 4 ojos (Pleurodema bufoninum). Además, esta última especie ha sido observada eventualmente coexistiendo con A. pehuenche en las subpoblaciones Rial Rojas, Campanaria y del Límite. Otras especies registradas durante los muestreos, que se identifican como posibles depredadores de A. pehuenche son el zorro gris (Lycalopex gymnocercus), el chiñe o zorrino (Conepatus chinga), la gaviota capucho café (Chroicocephalus maculipennis) y el águila mora (Geranoaetus melanoleucus).
Aunque la detección de huevos no fue objeto de este estudio, dado que no se realizó búsqueda activa en oquedades ni se manipularon hembras, en noviembre 2022 se observó una masa de 20 huevos aproximadamente, flotando en una pequeña poza de la subpoblación Rial Rojas en época de deshielo. Los huevos no eran pigmentados y cada uno midió entre 6 y 7 mm de diámetro.
Los índices de conectividad calculados para cada una de las subpoblaciones se muestran en la tabla 3. El arroyo Pehuenche representa la principal conexión entre subpoblaciones. En este arroyo se encontraron individuos adultos en la desembocadura de arroyos de primer orden pertenecientes a las subpoblaciones del Límite, Rial Rojas, Nueva y Campanaria (tabla 1). Si bien Cajón Largo tiene relativamente bajo índice de conectividad, el área de esta subpoblación es la mayor de todas.
Se registraron 24 individuos muertos durante el periodo de estudio: 1) 6 individuos adultos —2 hembras, 2 machos y 2 adultos indeterminados— en el mes de enero de la temporada estival 2021 y 2 en la temporada 2022-2023 en los muestreos realizados en la ruta internacional ARG145; 2) 3 individuos —1 adulto indeterminado y 2 larvas— encontrados muertos por desecación sobre el curso de un arroyo de Rial Rojas que mostró una disminución abrupta del caudal luego del deshielo entre noviembre y diciembre 2022; 3) 4 adultos —3 machos y 1 indeterminado— encontrados con signos de depredación (patas traseras lastimadas o mutiladas) al costado o dentro del curso del arroyo donde se registra presencia de salmónidos, esto es, en las subpoblaciones del Límite y Campanaria; 4) 6 adultos —2 machos y 4 indeterminados—, 1 juvenil indeterminado y 2 larvas encontrados muertos sin causa evidente, dentro del curso del arroyo o próximo a éste.
Finalmente, los valores de ECE negativos usados como indicadores, permitieron identificar a las subpoblaciones del Límite y Rial Rojas como unidades de conservación prioritarias, de las cuales la del Límite requiere esfuerzos más urgentes.
Discusión
Corbalán et al. (2010) reportaron 355 individuos de A. pehuenche en 2,500 m lineales (equivalente a 14 individuos/200 m2) en un muestreo realizado en 2008 en arroyos de la subpoblación del Límite. Si se compara ese valor con el obtenido en este estudio para los mismos arroyos en el mismo mes en 2021 (8 individuos/200 m2), el primero es mayor, pero se encuentra dentro del intervalo de variación de este estudio. Cabe mencionar que se usaron 2 técnicas diferentes para el conteo (búsqueda activa de individuos en oquedades durante el día y encuentro visual nocturno, con 3 observadores en los 2 casos), por lo que, con la información disponible actualmente, solo la mortalidad de individuos reportada desde 2008 permite estimar una tendencia poblacional decreciente para esta subpoblación.
En 2 arroyos muy cercanos a la subpoblación del Límite en Chile, Correa et al. (2013) reportaron 20 y 24 adultos/h o 15 y 66 adultos/200 m². Estos últimos datos corresponden a muestreos nocturnos realizados en marzo de 2012, con 4 observadores. La cantidad máxima de adultos por área observados por Correa et al. (2013) es mayor al máximo obtenido en este estudio, lo cual puede deberse a un error de muestreo en solo 30 m lineales, o bien a fluctuaciones poblacionales naturales (Kissel et al., 2020). En arroyos de Rial Moreno, a 21 km al NE de la población del valle Pehuenche, Corbalán et al. (2023) reportaron 26 adultos/hora y 4 juveniles/hora en un muestreo nocturno realizado en enero de 2019, los cuales se encuentran dentro de valores obtenidos en el valle Pehuenche en este estudio.
Durante las 3 temporadas consecutivas muestreadas en este trabajo, no se detectaron cambios significativos para ningún estadio de desarrollo de A. pehuenche. Esto puede deberse a que se trata de un periodo corto en términos de monitoreo. Por lo tanto, es necesario mantener los muestreos para conocer tendencias poblacionales a largo plazo. Por otro lado, la detección de individuos por encuentro visual a lo largo de los arroyos, podría compararse con otros estudios enfocados al monitoreo poblacional de la especie para lograr establecer un óptimo en términos de esfuerzo de muestreo. Dado que estas investigaciones dependen del presupuesto disponible y de limitaciones logísticas (Joseph et al., 2006) principalmente ligadas a la elevada demanda de horas/persona en un ambiente de alta montaña, se debe buscar una estrategia que permita satisfacer tanto la demanda de datos pertinentes para la gestión, como la obtención de información ecológica de calidad (Stephens et al., 2015; Yoccoz et al., 2001).
En relación con la primera acción de conservación llevada a cabo en la población del valle Pehuenche, que consistió en la construcción de 800 m de cordón o bordillo en la ruta y 2 cámaras de infiltración para evitar que la sal vertida en la ruta llegue a los arroyos, provocó la deshidratación y muerte de 18 individuos que fueron encontrados en diciembre de 2017, y 50 en enero de 2018. Posiblemente, la construcción a posteriori de rampas entre el cordón o bordillo y la calzada en febrero de 2018, ha evitado que se vuelvan a registrar mortalidades tan abruptas. Si bien nunca pudo constatarse el uso de las rampas por parte de la especie, las mismas habrían mejorado la conexión entre las secciones superior e inferior de los arroyos. Esto demuestra la necesidad de seguimiento de las acciones de conservación que se llevan a cabo. Pero, por otro lado, si bien la ruta pudo haber provocado una disminución poblacional (Corbalán et al., 2010; IUCN, 2019), el número elevado de individuos muertos registrado en este estudio alerta sobre la necesidad de avanzar en el conocimiento de otras amenazas, como la depredación por salmónidos invasores, el cambio climático y la infección por el hongo quitridio (Batrachochytrium dendrobatidis) de los anfibios.
En este trabajo, de todos los estadios evaluados, las larvas fueron las más abundantes, las más estables en el tiempo y presentaron el mismo patrón de variación que los adultos entre arroyos A y B, y subpoblaciones (fig. 3). Al igual que lo reportado por Corbalán et al. (2010, 2023), se encontraron larvas de diferentes tamaños conviviendo en pozas, pero también se observaron en rápidos, remansos y debajo de la vegetación acuática. Estos resultados son coherentes con la estrategia de especies con desarrollo larval plurianual (Úbeda, 2021). Por otro lado, la menor cantidad de juveniles registrada respecto de adultos podría ser un problema de detectabilidad por su tamaño (Petrovana y Schmidt, 2019), podría corresponder a una segregación por microhábitats según el estadio (Gonwouo et al., 2022), o bien, ser evidencia de una dinámica poblacional particular solo en esta fase de su ciclo de vida (Kissel et al., 2020).
Los muestreos preliminares realizados de día y de noche demuestran que hay arroyos permanentes que solo tienen adultos. Los estudios a futuro podrán determinar las causas que posiblemente puedan estar asociadas con aspectos fisicoquímicos del agua, o a la dispersión de individuos. La información actual permite suponer que las rutas de dispersión de A. pehuenche con base en sus hábitos acuáticos, son los mismos arroyos. Los registros de individuos secos en la ruta, principalmente entre diciembre y enero, al igual que la mayor cantidad de adultos y juveniles registrados en estos meses (fig. 2), podrían estar indicando una mayor dispersión en esa temporada del año.
Los arroyos del valle Pehuenche son los mejores conocidos hasta el momento y el área que ocupan representa solo 6% del área de distribución de A. pehuenche (487.9 km²) reportada por Corbalán et al. (2023). De la misma manera, habiendo considerado en nuestro estudio casi todos los arroyos presentes en el valle Pehuenche, representa 0.8% del área de ocupación total para la especie (4.84 km²; Corbalán et al., 2023). El espacio sin ocupación efectiva de la especie, actualmente, es muy grande (Corbalán et al., 2023), aunque en el pasado pudo haber estado presente en un área mayor si se comparan los registros de presencia en Cajón Grande (Cei y Roig, 1965; Corbalán et al., 2023). Si se considera desde la descripción de la especie en 1965, A. pehuenche ha reducido su área de distribución histórica, lo cual puede deberse a diferentes causas. La depredación por salmónidos en la actualidad podría ser la causa de que la especie haya quedado restringida solo a las zonas altas de las cuencas. Por ello, es necesario conocer la distribución de salmónidos e identificar dónde coexisten estos peces y la rana. Donde esto ocurre en el valle Pehuenche, se registraron adultos muertos de A. pehuenche con signos de depredación. Así, los salmónidos representan una barrera para la dispersión de las ranas, reduciendo la conectividad entre subpoblaciones y dentro de ellas entre arroyos A y B (tabla 3). Como se ha demostrado para otras especies de anfibios, la consecuencia es la disminución del intercambio genético o demográfico (e.g., Kacoliris et al., 2022; Velasco, 2018). Por lo tanto, para conservar las subpoblaciones se requiere asegurar su conectividad, tanto entre las secciones inferior y superior de los arroyos A impactados por la ruta, como dentro de las subpoblaciones entre los arroyos A y B, y entre las subpoblaciones conectadas a través del arroyo Pehuenche.
Las observaciones de campo sugieren que los individuos se distribuyen de manera agrupada a lo largo de los cursos de agua y vegas, no solo las larvas en las pozas, sino también los adultos a lo largo de los arroyos. Esto debe ser evaluado en la etapa reproductiva a inicios de la temporada y al final de ésta, cuando el caudal disminuye y algunos cauces se secan total o parcialmente, donde solo quedan pozas pequeñas con agua. Si este efecto es acentuado por la disminución de cobertura permanente de nieve en la cuenca del Río Grande (Aumassanne et al., 2019), es posible que más arroyos, que todavía mantienen agua hasta el final de la temporada, se sequen como ha ocurrido en los arroyos A3 y A4 de la población del Límite, aumentando así la fragmentación del hábitat. Si A4 no se hubiera secado, las 3 subpoblaciones (del Límite, Pichintur y Rial Rojas) estarían conectadas, formando una sola subpoblación (fig. 1).
Si bien gran parte de las amenazas han sido identificadas (Corbalán et al., 2010, 2023), la mayoría no han sido cuantificadas. Con base en las observaciones realizadas durante los muestreos en este estudio, se sistematizaron las amenazas para cada arroyo y cada subpoblación en el valle Pehuenche (tablas 2, 3). Como resultado de la priorización, las subpoblaciones del Límite y Rial Rojas se identifican como prioritarias para iniciar acciones concretas de conservación. Es posible que ambas sean fragmentos de una población mayor y que hayan estado unidas cuando el arroyo intermedio no estaba seco. En la del Límite se podría iniciar la mitigación de la depredación por salmónidos y reducir los efectos de la ruta. Es necesario continuar con el seguimiento y cuantificación de las amenazas, considerar las aún no evaluadas, tales como las que pueden estar generando las actividades turísticas, el vertido de sal en la ruta, crecidas extraordinarias y otras potenciales como el tendido de líneas de alta tensión, planificada desde el Maule (Chile) al río Diamante (San Rafael, Argentina).
El avance logrado en 20 años de estudios sobre la biología y conservación de este anfibio endémico de Argentina y Chile nos brinda herramientas para sostener acciones concretas de manejo que alivien alguna de las amenazas y que aseguren la viabilidad de las poblaciones a largo plazo. Se espera que un programa de monitoreo anual permita avanzar en el conocimiento de las tendencias poblacionales a largo plazo. Las subpoblaciones del valle Pehuenche son las mejor conocidas hasta el momento y han sufrido disminuciones, fragmentaciones y también extinciones locales. La priorización de su estado de conservación brinda herramientas para implementar acciones necesarias a corto plazo.
Agradecimientos
Por la colaboración en el trabajo de campo, se agradece a estudiantes de la Tecnicatura en Conservación de la Naturaleza Sede Malargüe del IEF Núm. 9-016, especialmente a Julián Rodríguez, Armando Barros, Francisco Jofré y Pablo Lucero; también a estudiantes voluntarios y Graciela Ríos. A Karen Olate por la elaboración del mapa. Y a un revisor anónimo por sus aportes a la primera versión del manuscrito y a los dos revisores que realizaron aportes importantes para mejorar el manuscrito. La investigación se logró gracias al financiamiento del proyecto SIIP 06/M003 Resol. 3978/2022 UNCUYO y aportes personales de investigadoras y estudiantes.
Referencias
Aumassanne, C. M., Beget, M. E., Di Bella, C. M., Oricchio, P. y Gaspari, F. J. (2019). Cobertura de nieve en las cuencas de los ríos Grande y Barrancas (Argentina) y su relación con la morfometría. Revista de Investigación Agropecuaria, 45, 394–403.
Cei, J. M. (1976). Remarks on some neotropical amphibians of the genus Alsodes from southern Argentina. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 117, 159–164.
Cei, J. M. (1980). Amphibians of Argentina. Monitore Zoologico Italiano, N.S. Monografia 2. Florencia: Universitá degli studi di Firenze.
Cei, J. M. y Roig, V. G. (1965). The systematic status and biology of Telmatobius montanus Lataste (Amphibia: Leptodactylidae). Copeia, 4, 421–425.
Corbalán, V., Debandi, G. y Martínez, F. (2010). Alsodes pehuenche (Anura: Cycloramphidae): past, present and future. Cuadernos de Herpetología, 24, 17–23.
Corbalán, V., Debandi, G., Martínez, F. y Úbeda, C. (2014). Prolonged larval development in the critically endangered Pehuenche’s Frog Alsodes pehuenche: implications for conservation. Amphibia–Reptilia, 35, 283–292. http://dx.doi. org/10.1163/15685381-00002951
Corbalán V., Debandi G., Literas S., Álvarez L., Rivera J. A., Dopazo J. et al. (2023). Newly discovered sites and potential threats for the critically endangered frog, Alsodes pehuenche, in Southern South America. Herpetological Conservation and Biology, 18, 48–56.
Correa, C., Pastenes, L., Iturra, P., Calderón, P., Vásquez, D., Lam, N. et al. (2013). Confirmation of the presence of Alsodes pehuenche Cei, 1976 (Anura, Alsodidae) in Chile: morphological, chromosomal and molecular evidence. Gayana, 77, 117–123. http://dx.doi.org/10.4067/S0717-65382013000200006
Correa, C., Zepeda, P., Lagos, N., Salinas, H., Palma, R. E. y Vásquez, D. (2018). New populations of two threatened species of Alsodes (Anura, Alsodidae) reveal the scarce biogeographic knowledge of the genus in the Andes of central Chile. Zoosystematics and Evolution, 94, 349–358.
Correa, C. J. Morales, C. Schussler y J. C. Ortiz. (2020). An enigmatic population of Alsodes (Anura, Alsodidae) from the Andes of central Chile with three species-level mitochondrial lineages. Mitochondrial DNA Part A DNA Mapping, Sequencing, and Analysis, 31, 25–34. https://doi.org/10.1080/24701394.2019.1704744
Crump, M. L. y Scott, N. J. (1994). Visual encounter surveys. En W. R.Heyer, M. A. Donnelly, R. W. Mc Diarmid, L. A. C. Hayek y M. S. Foster (Eds.). Measuring and Monitoring Biological Diversity Standard Methods for Amphibians (pp. 84–92). Washington D.C.: Smithsonian Institution Press.
Cuevas, C. C. y Formas, J. R. (2003). Cytogenetic analysis of four species of the genus Alsodes (Anura: Leptodactylidae) with comments about the karyological evolution of the genus. Hereditas, 138, 138–147. https://doi.org/10.1034/j.1601-5223.2003.01677.x
Garreaud, R., Vuille, M., Compagnucci, R. y Marengo, J. (2009). Present-day south american climate. Palaeogeography, Palaeoclimatology, Palaeoecology, 281, 180–195. http://dx. doi.org/10.1016/j.palaeo.2007.10.032
Ghirardi, R., Levy, M. G., López, J. A., Corbalán, V., Steciow, M. M. y Perotti, M. G. (2014). Endangered amphibians infected with the chytrid fungus Batrachochytrium dendrobatidis in austral temperate wetlands from Argentina. Herpetological Journal, 24, 129–133.
Gonwouo, N. L. Schäfer, M. y Tsekané, S. J. (2022). Goliath frog (Conraua goliath) abundance in relation to frog age, habitat, and human activity. Amphibian and Reptile Conservation, 16, e319.
Grant, E. H. C., Miller, D. A. y Muths, E. (2020). A synthesis of evidence of drivers of amphibian declines. Herpetologica, 76, 101–107. https://doi.org/10.1655/0018-0831-76.2.101
Green D. M., Lannoo M. J., Lesbarrères D. y Muths E. (2020). Amphibian population declines: 30 years of progress in confronting a complex problem. Herpetologica, 76, 97–100. https://doi.org/10.1655/0018-0831-76.2.97
Herrera, F. y Velásquez, N. A. (2016a). Dimorfismo sexual en Alsodes pehuenche Cei 1976 (Amphibia, Anura, Alsodidae). Boletín Chileno de Herpetología, 3, 4–6.
Herrera, F. y Velásquez, N. A. (2016b). Uso de cuevas en Alsodes pehuenche Cei 1976 (Amphibia, Anura, Alsodidae). Boletín Chileno de Herpetología, 3, 17–20.
Jensen, I. S., Haugen, V. F. y Skålin, R. (2007). Yr. Norwegian Meteorological Institute and the Norwegian Broadcasting Corporation. Recuperado el 15 de agosto, 2024 de https://www.yr.no/nb
Joseph, L. N., Field, S. A., Wilcox, C. y Possingham, H. P. (2006). Presence-absence versus abundance data for monitoring threatened species. Conservation Biology, 20, 1679–1687. https://doi.org/10.1111/j.1523-1739.2006.00529.x
Kacoliris, F. P., Berkunsky, I., Acosta, J. C., Acosta, R., Agostini, M. G., Akmentins, M. et al. (2022). Current threats faced by amphibian populations in the southern cone of South America. Journal for Nature Conservation, 69, 126254. https://doi.org/10.1016/j.jnc.2022.126254
Kissel, A. M., Tenan, S. y Muths, E. (2020). Density dependence and adult survival drive dynamics in two high elevation amphibian populations. Diversity, 12, 478.
Lin, J. P. (2009). The functional linkage index: a metric for measuring connectivity among habitat patches using least-cost distances. Journal of Conservation Planning, 5, 28–37.
Llano, C., Durán, V., Gasco, A., Reynals, E. y Zárate, M. S. (2021). Traditional puesteros´ perceptions of biodiversity in semi-arid Southern Mendoza, Argentina. Journal of Arid Environments, 192, 104553. https://doi.org/10.1016/j.ja ridenv.2021.104553
Luedtke, J. A., Chanson, J., Neam, K., Hobin L., Maciel, A. O., Catenazzi, A. et al. (2023). Ongoing declines for the world’s amphibians in the face of emerging threats. Nature, 622, 308–314. https://doi.org/10.1038/s41586-023-06235-w
Luja, V., Rodríguez-Estrella, H. R., Schaub, M. y Schmidt, B. R. (2015). Among-population variation in monthly and annual survival of the Baja California treefrog, Pseudacris hypochondriaca curta, in desert oases of Baja California Sur, Mexico. Herpetological Conservation and Biology, 10, 112–122.
Pereyra, L. C., Etchepare, E. y Vaira, M. (2021). Manual de técnicas y protocolos para el relevamiento y estudio de anfibios de Argentina. San Salvador de Jujuy: Editorial de la Universidad Nacional de Jujuy.
Petrovana, S. O. y Schmidt, B. R. (2019). Neglected juveniles; a call for integrating all amphibian life stages in assessments of mitigation success (and how to do it). Biological Conservation, 236, 252–260. https://doi.org/10.1016/j.biocon. 2019.04.016
Piñeiro, A., Fibla, P., López, C., Velásquez, N. y Pastenes, L. (2020). Characterization of an Alsodes pehuenche breeding site in the Andes of central Chile. Herpetozoa, 33, 21–26. https://doi.org/10.3897/herpetozoa.33.e49268
Pollock, J. E. (2006). Detecting population declines over large areas with presence-absence, time-to-encounter, and count survey methods. Conservation Biology, 20, 882–892. https://doi.org/10.1111/j.1523-1739.2006.00559.x
Prado, W. S., Meriggi, J., Martínez, F. y Corbalán, V. (2019). Ampliación del área de distribución de Alsodes pehuenche en Argentina. Revista del Museo de La Plata, 4 (Suplem.), 1R–117R.
Rivera, J. A., Marianetti, G. y Hinrichs, S. (2018). Validation of CHIRPS precipitation dataset along the Central Andes of Argentina. Atmospheric Research, 213, 437–449. https: //doi.org/10.1016/j.pce.2022.103184
Stephens, P. A., Pettorelli, N., Barlow, J., Whittingham, M. J. y Cadotte, M. W. (2015). Management by proxy? The use of indices in applied ecology. Journal of Applied Ecology, 52, 1–6. https://doi.org/10.1111/1365-2664.12405
Úbeda, C. (2021). Estrategias reproductivas, hábitats y otros aspectos ecológicos de los anfibios altoandinos en la vertiente oriental de la Cordillera de los Andes. Boletín Chileno de Herpetología, 8, 10–21.
UICN (Unión Internacional para la Conservación de la Naturaleza), Species Survival Commission Amphibian Specialist Group. (2019). Alsodes pehuenche. IUCN Red List of threatened species, 2019. https://www.iucnredlist.org
Vaira, M., Akmentins, M., Attademo, M., Baldo, D., Barrasso, D., Barrionuevo, S. et al. (2012). Categorización del estado de conservación de los anfibios de la República Argentina. Cuadernos de Herpetología, 26 (Suplem. 1), 131–159.
Vaira, M., Pereyra, L. C., Akmentins M. S. y Bielby, J. (2017). Conservation status of Amphibians of Argentina: An update and evaluation of national assessments. Amphibian y Reptile Conservation, 11, e135.
Velasco, M. A. (2018). Dinámica poblacional y conservación de la ranita del Valcheta (Pleurodema somuncurense) (Cei, 1969), Patagonia, Argentina (Tesis doctoral). Universidad Nacional de La Plata, La Plata, Buenos Aires.
Velasco, M. A., Kacoliris, F. P., Berkunsky, I., Quiroga, S. y Williams, J. D. (2016). Current distributional status of the critically endangered Valcheta frog: implications for conservation. Neotropical Biology and Conservation, 11, 110–113. http://dx.doi.org/10.4013/nbc.2016.112.08
Vidal, M. A., Henríquez, N., Torres-Díaz, C., Collado, G. y Acuña-Rodríguez, I. S. (2024). Identifying strategies for effective biodiversity preservation and species status of Chilean amphibians. Biology, 13, 169. https://doi.org/10.33 90/biology13030169
Yoccoz, N. G., Nichols, J. D. y Boulinier, T. (2001). Monitoring of biological diversity in space and time. Trends in Ecology and Evolution, 16, 446–453. https://doi.org/10.1016/S0169- 5347(01)02205-4
Zarco, A., Corbalán, V. y Debandi, G. (2020). Depredación por truchas arcoíris invasoras en la rana de pecho espinoso Pehuenche, en peligro crítico. Journal of Fish Biology, 98, 878-880. https://doi.org/10.1111/jfb.14401
Fernando Arriola-Álvarez a, Luis Gabriel Aguilar-Estrada a, *, Lucía Álvarez-Castillo b, Ivette Ruiz-Boijseauneau a y Dení Rodríguez a
a Universidad Nacional Autónoma de México, Facultad de Ciencias, Laboratorio de Ficología (Biodiversidad Marina), Circuito Exterior s./n., Coyoacán, 04510 Ciudad de México, México
b Universidad Nacional Autónoma de México, Facultad de Ciencias, Posgrado en Ciencias del Mar y Limnología, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
*Autor para correspondencia: lgae@ciencias.unam.mx (L.G. Aguilar-Estrada)
Recibido: 7 julio 2022; aceptado: 11 septiembre 2024
Resumen
Las macroalgas intermareales proporcionan alimento y refugio para diferentes organismos. El objetivo de este trabajo fue analizar los bivalvos asociados a macroalgas. Se realizaron muestreos en enero, mayo, julio y noviembre de 2014, se recolectaron manualmente 72 muestras de macroalgas y sus bivalvos asociados dentro de cuadros de 400 cm2 en 2 localidades de Guerrero: playas El Palmar y Las Gatas. La estructura comunitaria de los bivalvos se determinó a partir de la riqueza específica, composición, abundancia, distribución e índices comunitarios: diversidad de Shannon, equidad de Pielou y dominancia de Simpson. Cada especie de macroalga (59 spp.) se asoció con la propuesta de grupos morfofuncionales. Se analizó la cobertura de macroalgas, abundancia de bivalvos y sedimento retenido. Del total de individuos (873), se reconocieron 17 especies de bivalvos. El índice de Shannon fue de 2.15 bits/individuo. Los bivalvos se asociaron a 3 grupos morfofuncionales de macroalgas. La abundancia de bivalvos y los sedimentos retenidos disminuyeron por mes, mientras que la abundancia, cobertura y sedimentos disminuyeron al aumentar el nivel de marea. Estudios como este proporcionan información importante para el conocimiento de la diversidad costera, en este caso de una zona turística en Guerrero.
Community structure of bivalves (Mollusca: Bivalvia) associated with intertidal macroalgae of Guerrero, Mexico
Abstract
Intertidal macroalgae provide food and shelter for different organisms. The objective of this work was to analyze the bivalves associated with macroalgae. Sampling was carried out in January, May, July, and November 2014, 72 samples of macroalgae and their associated bivalves were manually collected within 400 cm2 in 2 locations in Guerrero: El Palmar and Las Gatas beaches. The community structure of bivalves was determined from specific richness, composition, abundance, distribution, and community indices: Shannon diversity, Pielou evenness and Simpson dominance. Each macroalgal species (59 spp.) was associated with proposed morphofunctional groups. Macroalgal cover, bivalve abundance and retained sediment were analyzed. Of the total number of individuals (873), 17 bivalve species were recognized. The Shannon index was 2.15 bits/individual. Bivalves were associated with 3 morphofunctional groups of macroalgae. Bivalve abundance and retained sediment decreased by month, while abundance, cover, and sediment decreased with increasing tide level. Studies like this provide important information for understanding coastal diversity, in this case of a tourist area in Guerrero.
Keywords: Malacofauna; Tropical Mexican Pacific; Species richness; Sediments
Introducción
En el Pacífico tropical mexicano se han descrito 2 patrones principales de circulación de corrientes oceánicas: primavera (marzo-abril) y otoño (septiembre-octubre), que generan variaciones espacio-temporales como las temporadas de lluvias y secas o fenómenos climatológicos (Baumgartner y Christensen, 1985; Pérez, 2013; Vega et al., 2008; Wyrtki, 1966). La zona intermareal rocosa es un sitio de transición entre los ambientes terrestre y marino, la cual se encuentra sujeta a cambios constantes de las variables ambientales como la oscilación de la marea, la intensidad lumínica, el viento, las variaciones en la salinidad y la temperatura (Salazar-Vallejo y González, 1990; Vassallo et al.,2014); ésto genera un hábitat heterogéneo con diversos microambientes en donde varios grupos de organismos pueden desarrollarse (Flores-Garza etal., 2011, 2014). Los organismos más frecuentes en esta zona son las macroalgas (Lee, 2008), mismas que proporcionan refugio y alimento para numerosos grupos de invertebrados (García-Robledo et al.,2008; Jover-Capote y Diez, 2017; Moreno, 1995; Steneck y Watling, 1982; Yang et al., 2007). Las macroalgas son un ambiente espacialmente heterogéneo, lo que hace posible que puedan albergar distintos grupos de invertebrados a lo largo del tiempo (Benedetti-Cecchi et al., 2001; Olabarria y Chapman, 2001). Los anfípodos, poliquetos y moluscos son los grupos más importantes al interior de la comunidad de macroalgas, ya que representan 70% de la abundancia en éstas (Aguilera, 2011; Colman, 1940).
Dentro del phylum Mollusca, los bivalvos son la segunda clase más representativa (Gosling, 2015). En la zona intermareal pueden vivir adheridos a diversos sustratos como rocas, arena o macroalgas (García-Cubas y Reguero, 2007). Algunos organismos que conforman la clase Bivalvia son sésiles y tienen diferentes estrategias en cuanto a sus tipos de alimentación: suspensívoros o detritívoros (Coan y Valentich-Scott, 2006). Además, generan redes de mucus para atrapar las partículas que flotan en la columna de agua (Jorgensen, 1996; Ward et al., 1998), por lo que desempeñan papeles ecológicos importantes en los cuerpos de agua; por sus hábitos de vida, son un grupo de especial interés en los estudios ecológicos (Lozada, 2015; Vega et al., 2008).
En general, los trabajos sobre moluscos en México son numerosos. En el de Sánchez (2014) se mencionó que existen alrededor de 47 contribuciones tomando en cuenta las costas del Atlántico y del Pacífico. En Guerrero se cuenta con alrededor de 30 estudios malacológicos (Gama, 2019), los cuales en su mayoría se han orientado a conocer la riqueza y composición de especies (Flores, 2004; Flores-Rodríguez et al., 2012; Lesser, 1984; López-Rojas et al., 2017). Gran parte (70%) de los trabajos en este estado se han realizado en el área de Acapulco y se han analizado distintos aspectos de la comunidad de moluscos, incluyendo a los bivalvos (Barba-Marino et al., 2010; Castro-Mondragón et al.,2016; Flores-Garza et al., 2010, 2011, 2012, 2014; Flores-Rodríguez et al.,2003; Galeana-Rebolledo et al., 2012, 2018; Garcés, 2011; Kuk-Dzul et al.,2019; Torreblanca, 2010; Torreblanca-Ramírez et al., 2012; Valdés-González etal.,2004; Villegas-Maldonado et al., 2007; Villalpando, 1986).
En la parte norte de Guerrero, que incluye Ixtapa-Zihuatanejo, se han realizado estudios ecológicos y sobre ciclos reproductivos de moluscos (Baqueiro, 1979; Flores-Rodríguez et al.,2007; Salcedo-Martínez et al.,1988), o sobre especies de importancia comercial como el de Cerros-Cornelio et al. (2021), quienes mencionaron 24 especies de moluscos, de las cuales 13 son de la clase Bivalvia. Por su parte, en la costa sur de Guerrero existen 9 trabajos malacológicos; Flores-Garza et al. (2007) analizaron la densidad de Plicopurpura columellaris (Lamarck, 1816) y su malacofauna asociada, reportando 34 especies de moluscos, de las cuales 7 fueron bivalvos.
Entre los estudios realizados en las costas de Guerrero, resalta el de Salcedo-Martínez et al. (1988) por ser el primero sobre la relación entre las macroalgas e invertebrados de Zihuatanejo, donde los moluscos, en especial la clase Gastropoda, fueron el componente mayoritario (38.72%). Existen algunos trabajos sobre la asociación alga-molusco en Ixtapa-Zihuatanejo (Aguilar-Estrada et al., 2017, 2022; Cisneros, 2016; Gama-Kwick et al., 2021; Quiroz-González et al., 2020); sin embargo, dichos estudios están enfocados a otras clases de moluscos (gasterópodos y quitones); por lo que, el conocimiento sobre la relación de las macroalgas y bivalvos es escaso. La presente contribución tiene como objetivo aportar conocimiento de la estructura comunitaria de los moluscos bivalvos asociados a macroalgas, en un ciclo anual en la zona intermareal rocosa de Ixtapa-Zihuatanejo en Guerrero.
Materiales y métodos
Se realizaron 4 salidas de campo durante enero, mayo, julio y noviembre de 2014 a Ixtapa-Zihuatanejo, Guerrero, con el propósito de observar los posibles cambios de la estructura comunitaria de los bivalvos asociados a las macroalgas en la zona. Las comunidades de macroalgas se recolectaron en la zona intermareal rocosa en 2 localidades: playa El Palmar en Ixtapa (17°39’0.4” N, 101°36’2.79” O) y el pretil de playa Las Gatas, al interior de la bahía de Zihuatanejo (17°37’22.07” N, 101°33’4.85” O) (fig. 1A). La recolección de ejemplares se realizó con un permiso otorgado por la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA) (Registro Nacional de Pesca y Acuacultura -DF00000208).
Playa El Palmar es un sitio expuesto, se encuentra frente al complejo turístico Ixtapa y la playa está formada por litoral rocoso y arenoso, tiene una longitud de 2.7 km. Este lugar consta de un relieve heterogéneo, compuesto por riscos y morros de diferente tamaño y forma irregular, la zona intermareal rocosa tiene una amplitud aproximada de 2 m (Aguilar, 2017). En esta playa se han descrito patrones con una circulación del agua dominante hacia el norte a lo largo de la zona, por lo que el oleaje es intenso en la parte norte (Trasviña y Andrade, 2002) (fig. 1B).
Playa Las Gatas es un sitio protegido porque se localiza al interior de la bahía de Zihuatanejo; tiene una extensión de 350 m y se compone principalmente de arenas, fragmentos de coral y rocas (Cisneros, 2016; López, 1993). Paralelo a la línea de costa se encuentra el “pretil”, que es un conglomerado de rocas apiladas, irregulares y de tamaños variables (Aguilar, 2017; Cisneros, 2016; Urbano, 2004). La zona intermareal tiene una amplitud aproximada de 1 m (López, 1993). La presencia de este conglomerado hace que el oleaje sea menor al interior de la playa (Aguilar, 2017) (fig. 1C).
Figura 1. A, Ubicación de la zona de estudio en Ixtapa-Zihuatanejo; B, playa El Palmar; C, playa Las Gatas.
En cada localidad, 3 muestras de macroalgas se recolectaron manualmente de forma aleatoria con una espátula (Bakus, 2007), dentro de cuadros de 20 × 20 cm (0.04 m2) por cada nivel de marea: intermareal bajo (3), medio (3) y alto (3); en total, se obtuvieron 72 muestras provenientes de 36 cuadros en playa Las Gatas y de 36 cuadros en playa El Palmar. Las muestras de las comunidades de macroalgas y moluscos asociados se preservaron en una mezcla de formaldehído al 4% con agua de mar, neutralizada con borato de sodio y glicerina; después se trasladaron al Laboratorio de Ficología (Biodiversidad Marina) de la Facultad de Ciencias de la Universidad Nacional Autónoma de México (UNAM).
En el laboratorio, se calculó la cobertura de cada especie de macroalga en cm2 al colocar cada muestra sobre un área delimitada de 20 × 20 cm. Para su identificación taxonómica se tomaron en cuenta las características morfológicas externas, como tipo de talo y tipo de ramificación, e internas a partir de cortes anatómicos transversales de ejes, frondas y, de estar presentes, de estructuras reproductivas; los especímenes y cortes fueron observados bajo microscopios estereoscópico y óptico (Zeiss). La identificación taxonómica de los ejemplares de macroalgas se realizó utilizando literatura especializada para macroalgas del océano Pacífico (Abbott, 1999; Abbott y Hollenberg, 1976; Dawson, 1949, 1953, 1954, 1960, 1961, 1963; Dawson y Beaudette, 1959; Rodríguez et al.,2008; Taylor, 1945). La actualización de la nomenclatura se hizo a partir de la base de datos de Algaebase (Guiry y Guiry, 2024) y, con base en ella, se elaboró una lista sistemática de las especies de macroalgas. De cada comunidad de macroalgas se separó el sedimento retenido por las algas después de la medición de la cobertura para cada especie y se midió su peso húmedo con una balanza digital, modelo OBI.
De cada muestra, se extrajeron de forma manual todos los moluscos de la clase Bivalvia. Los ejemplares con concha y parte blanda (vivos) fueron identificados al nivel taxonómico más bajo posible, género o especie, dependiendo del estado de conservación de cada ejemplar, a partir de la observación de las características morfológicas de la concha, con apoyo de un microscopio estereoscópico. La identificación taxonómica de las especies se hizo con literatura malacológica especializada para la zona del océano Pacífico oriental (Coan et al., 2000; Keen, 1971). Se elaboró una lista sistemática con base en la propuesta de Bouchet et al. (2010) para los niveles suprafamiliares y la actualización de nomenclatura se realizó a partir de la base de datos de World Register of Marine Species (WORMS) para género y especie (Horton et al., 2024).
Los ejemplares de moluscos y macroalgas recolectados fueron depositados en la colección “Invertebrados asociados a macroalgas”, en proceso de registro, con número de inventario para bivalvos (INV-1531 a INV-1638) del Laboratorio de Ficología (Biodiversidad Marina) de la Facultad de Ciencias, UNAM y las macroalgas se depositaron en la colección del Herbario de la Facultad de Ciencias (FCME) con número de catálogo (PTM-10534 a PTM-10558; PTM-10577 a PTM-10604; PTM-10613 a PTM-10630; PTM-10640 a PTM-10648).
Se elaboró una curva de acumulación de las especies de bivalvos asociados a macroalgas recolectadas en las localidades de estudio, con la finalidad de conocer la cantidad de especies que faltaría encontrar y recolectar en dichas localidades. Con los datos de riqueza de especies (S) y abundancia de bivalvos (N) de ejemplares vivos (concha y parte blanda), se estimaron los índices de Shannon (H´) y de diversidad máxima (H’ max) para cada fecha de muestreo, ya que éstos corresponden a los miembros de la comunidad en el momento de muestreo (Aguilar-Estrada et al., 2014); H´ fue expresado en bits/individuo (Magurran, 2004). Se calculó el índice de equidad de Pielou (J´) y el índice de dominancia de Simpson (D) (Moreno, 2001). Estos índices permiten hacer comparaciones cuantitativas y cualitativas entre estudios o zonas ya que se han utilizado como referente mínimo para describir la estructura comunitaria de un lugar (Aguilar-Estrada et al., 2014).
Para evaluar la normalidad de los datos de abundancia de bivalvos y de los índices de diversidad se realizaron pruebas de Shapiro-Wilk (W) (Siegel, 1990). Posteriormente, se realizaron pruebas de Levene (F) para comprobar la homogeneidad de varianzas para los datos no normales y prueba de Bartlett para los datos con distribución normal (Bartlett, 1937; Levene, 1960). Los índices de diversidad de Shannon fueron analizados usando una prueba de “t de student” para evaluar si existían diferencias estadísticamente significativas entre los meses de muestreo. Estos análisis se realizaron utilizando la paquetería Vegan Versión 2.5-6 (Oksanen et al., 2019) en el programa R Studio 2023.12.0+369 (R Core Team, 2023).
Los datos de abundancia de bivalvos no fueron normales de acuerdo con las pruebas de normalidad, por ello, se realizaron análisis de estadística no paramétrica en el software PRIMER v6 + add on Permanova (PRIMER-E Ltd., Plymouth, UK) (Anderson et al., 2008; Clarke y Gorley, 2006). Los datos fueron transformados con 4√ y con ellos se calculó una matriz de similitud a partir del índice de Bray-Curtis. A partir de dicha matriz, se realizó un análisis de escalamiento multidimensional no métrico (nMDS) con la finalidad de observar la distribución de las abundancias de bivalvos en el área de estudio. Se incluyó la abundancia de las especies dominantes en forma de vectores azules superpuestos en el gráfico nMDS para facilitar la interpretación de las abundancias de especies dominantes en las distintas localidades y niveles de marea, donde el círculo azul representa la variación de la abundancia. Las especies dominantes que se seleccionaron en este análisis son las que estuvieron presentes en todas las muestras con abundancias mayores a la media.
Además, se realizaron análisis de varianza permutacionales (Permanova) para determinar si existían diferencias significativas en la abundancia de bivalvos (variable dependiente) con respecto a los factores utilizados: localidad con 2 niveles (El Palmar y Las Gatas), nivel de marea con 3 niveles (alto, medio y bajo) y mes de muestreo con 4 niveles (enero, mayo, julio y noviembre). En los análisis de Permanova se utilizaron 999 permutaciones de los residuos bajo un modelo reducido. Posteriormente, para los factores donde se obtuvieron diferencias significativas, se realizaron comparaciones de pares para identificar los niveles que eran diferentes estadísticamente (Anderson et al., 2008).
Utilizando las variables numéricas, se realizaron regresiones lineales múltiples con el software SPSS Statistics v20, para evaluar el efecto de la cobertura de macroalgas y los sedimentos retenidos (variables independientes) en la abundancia de bivalvos (variable dependiente) en las playas El Palmar y Las Gatas, en los diferentes niveles de marea y en los meses de muestreo.
Por último, se determinó el grupo morfofuncional (GMF): filamentosas (Fil), foliosas (Fol), foliosas corticadas (Foc), filamentosas corticadas (Fic), coriáceas (Cor), calcáreas articuladas (Cal) para cada especie de macroalgas, con base en la propuesta de Steneck y Dethier (1994) y se asociaron con las especies de bivalvos recolectadas.
Resultados
Riqueza y composición de moluscos. En 60 de las 72 muestras recolectadas se encontraron bivalvos, 35 muestras pertenecientes a playa El Palmar y 25 a playa Las Gatas. Se obtuvieron un total de 873 individuos de la clase Bivalvia. Se identificaron 17 especies (fig. 2) agrupadas en 2 subclases, 8 órdenes, 10 familias y 15 géneros. Del total de especies recolectadas, 2 de ellas Parvilucina approximata (Dall, 1901) y Pinna rugosa G. B. Sowerby I, 1835, encontradas en playa El Palmar y playa Las Gatas, respectivamente, no se incluyeron en los análisis de estructura comunitaria, dado que solo se recolectó la concha. Se obtuvieron 13 especies en playa El Palmar y 9 en playa Las Gatas. Las familias Carditidae y Mytilidae fueron las mejor representadas con 3 (18%) y 5 (30%) especies, respectivamente.
En ambas localidades, las especies más abundantes fueron: Brachidontes adamsianus (Dunker, 1857), Leiosolenus aristatus (Dillwyn, 1817) e Isognomon janus Carpenter, 1857, con 376, 205 y 124 individuos, respectivamente, lo cual representó 91% del total de individuos. Brachidontes adamsianus obtuvo la mayor abundancia en El Palmar a lo largo del ciclo anual, con el valor más elevado (161 individuos) en enero. En Las Gatas, Leiosolenus aristatus fue la especie con la mayor abundanciacon 140 individuos en enero; sin embargo, la especie más abundante fue B. adamsianus a lo largo de los otros meses de muestreo. Mientras que las especies menos abundantes fueron: Sphenia fragilis (H. Adams & A. Adams, 1854) con 2 individuos en enero y mayo en El Palmar y 1 individuo en noviembre en Las Gatas, Crassinella ecuadoriana Olsson, 1961 con 2 individuos en mayo y julio en El Palmar, Linucula declivis (Hinds, 1843) con 2 individuos en julio y noviembre en Las Gatas, Mya sp. y Mytilus edulis Linnaeus, 1758 con 1 individuo en enero en El Palmar (tabla 1).
Tabla 1
Abundancia de las especies de bivalvos recolectadas en las localidades de estudio por mes de muestreo. El arreglo sistemático sigue la propuesta de Horton et al. (2024).
Especie
Playa El Palmar
Playa Las Gatas
ene
may
jul
nov
ene
may
jul
nov
Total
Subclase Protobranchia
Orden Nuculida
Familia Nuculidae
Linucula declivis
–
–
–
–
–
–
1
1
2
Subclase Autobranchia
Orden Mytilida
Familia Mytilidae
Brachidontes adamsianus
161
43
18
14
41
9
34
56
376
Brachidontes semilaevis
4
–
–
1
–
–
–
–
5
Leiosolenus aristatus
2
30
1
6
140
7
5
14
205
Modiolus capax
1
2
1
1
–
–
–
–
5
Mytilus edulis
1
–
–
–
–
–
–
–
1
Orden Arcida
Familia Arcidae
Acar rostae
–
–
–
–
–
–
1
3
4
Orden Ostreida
Familia Pteriidae
Isognomon janus
51
32
8
5
2
–
10
16
124
Familia Pinnidae
Pinna rugosa
–
–
–
–
–
–
3
–
3
Orden Lucinida
Familia Lucinidae
Parvilucina approximata
1
–
–
–
–
–
–
–
1
Orden Carditida
Familia Carditidae
Carditamera affinis
5
2
–
–
1
–
–
1
9
Carditamera radiata
–
3
1
–
3
–
3
5
15
Cardites grayi
–
1
3
–
–
–
–
–
4
Familia Crassatellidae
Crassinella ecuadoriana
–
1
1
–
–
–
–
–
2
Orden Venerida
Familia Chamidae
Chama coralloides
3
2
1
–
–
–
–
7
13
Orden Myida
Familia Myidae
Mya sp.
1
–
–
–
–
–
–
–
1
Sphenia fragilis
1
1
–
–
–
–
–
1
3
Total
231
117
34
27
187
16
57
104
773
Figura 2. Especies de bivalvos asociadas a macroalgas recolectadas en Ixtapa-Zihuatanejo, Guerrero, en vista dorsal y ventral. a, Linucula declivis;b, Brachidontes adamsianus;c, Leiosolenus aristatus; d, Mytilus edulis; e, Acar rostae; f, Isognomon janus;g, Pinna rugosa; h, Parvilucina approximata; i, Carditamera radiata; j, Cardites grayi; k, Crassinella ecuadoriana; l, Chama coralloides;m, Sphenia fragilis; n, Modiolus capax;o, Brachidontes semilaevis;p, Carditamera affinis;q, Mya sp.
La riqueza de especies con relación al ciclo anual fue de 13 especies en playa El Palmar, variando entre 5 (noviembre) y 10 especies (enero) a lo largo de los meses, mientras que en playa Las Gatas se obtuvieron 9 especies, con una variación de 2 (mayo) y 9 (noviembre) especies (tabla 1). La curva de acumulación de especies mostró un comportamiento asintótico en El Palmar, mientras que Las Gatas no mostró una tendencia a ser asintótica (fig. 3).
Figura 3. Curva de acumulación de especies de bivalvos registrados en las localidades de estudio. Línea azul: playa El Palmar, línea verde: playa Las Gatas.
En playa El Palmar el nivel intermareal bajo obtuvo el mayor número de especies (13) y el menor número de especies se presentó tanto en el nivel intermareal medio como en el alto (7); mientras que en playa Las Gatas, se presentó el mayor número de especies en el nivel intermareal medio (10) y el nivel alto tuvo el menor número de especies (5). Las especies que se presentaron en ambas localidades en los 3 niveles de la zona intermareal fueron Brachidontes adamsianus, Leiosolenus aristatus e Isognomon janus.
En El Palmar se registraron valores del índice de diversidad de Shannon que fluctuaron entre 1.30 y 2.15 bits/individuo, mientras que en Las Gatas este índice varió entre 0.98 y 2.09 bits/individuo (tabla 2). En general, el índice de equidad de Pielou presentó valores mayores a 0.60 en playa El Palmar, excepto en enero, mientras que en playa Las Gatas se observó que los valores fueron bajos en enero y mayo, y mayores a 0.60 en julio y noviembre. Las pruebas de “t de student”para el índice de diversidad de Shannon entre los meses de muestreo, indicaron diferencias significativas entre todos ellos (p ≤ 0.05) en El Palmar, y Las Gatas (tabla 3).
Tabla 2
Índices comunitarios calculados para las especies recolectadas en playa El Palmar y playa Las Gatas. N = número de individuos, H’= Índice de Shannon, J’= equidad de Pielou y λ = dominancia de Simpson. Los valores máximos y mínimos por localidad para el índice de Shannon (H´) se marcan con negritas.
Playa El Palmar
Playa Las Gatas
Mes
N
H’
J’
λ
N
H’
J’
λ
Enero
230
1.30
0.48
0.54
187
0.99
0.41
0.60
Mayo
117
2.15
0.81
0.27
16
0.98
0.41
0.50
Julio
34
2.03
0.77
0.34
54
1.63
0.68
0.44
noviembre
27
1.77
0.67
0.35
104
2.09
0.87
0.33
En el análisis de escalamiento multidimensional no métrico (nMDS), a partir de los valores de abundancia de las especies de bivalvos, se observó que las muestras se separan en 3 grupos: un primer grupo dominado por estaciones pertenecientes a playa El Palmar (P), un segundo grupo principalmente con estaciones de playa Las Gatas (G) y uno tercero con estaciones de ambas localidades que probablemente se separa por la dominancia de la especie Brachidontes adamsianus (fig. 4). El vector indica la dirección a través del plano de ordenación en la cual aumentan los valores de abundancia de las especies dominantes, la longitud de la línea indica la cantidad de variación total de cada especie, entonces, si toda la variación se explicara, la línea azul alcanzaría el círculo azul.
Tabla 3
Resultado de la prueba de “t de student”para evaluar diferencias significativas del índice de Shannon entre los meses de muestreo por localidad.
Playa El Palmar
Playa Las Gatas
Meses
H´
t
p
gl
H´
t
p
gl
enero vs. mayo
1.30/2.15
-0.009
324
1
0.99/0.98
0.009
169
1
mayo vs. julio
2.15/2.03
0.0004
142
0.98/1.63
-19.84
4
julio vs. noviembre
2.03/1.17
0.023
60
1.63/2.09
-3.18
94
El análisis de Permanova reveló que la composición de bivalvos difiere significativamente entre las diferentes localidades muestreadas (pseudo-F = 5.2373, p = 0.001). No hubo diferencias estadísticamente significativas para los meses de muestreo (pseudo-F=1.431, p = 0.147) y el nivel de marea (pseudo-F = 1.19, p = 0.309) (tabla 4).
Tabla 4
Resultado de los análisis de Permanova para los factores de localidad, mes de muestreo y nivel de marea. GL: Grados de libertad, ms: media suma de cuadrados, perm: permutaciones, * = valores significativos.
Factor
GL
ms
Pseudo-F
p (perm)
Localidad
1
7,844.9
5.2373
0.001*
Res
57
1,497.9
Total
58
Mes
3
2,249.9
1.431
0.147
Res
55
1,572.3
Total
58
Nivel de marea
2
1,915
1.1996
0.309
Res
56
1,596.3
Total
58
Riqueza y composición de macroalgas. Se encontró un total de 3 phyla, 3 clases, 5 subclases, 14 órdenes, 20 familias, 33 géneros y 59 especies de macroalgas; de las cuales, a cada localidad le pertenecen 37 especies. De las 59 especies totales, 11 fueron Chlorophyta (18%), 42 Rhodophyta (70%) y 7 Heterokontophyta-Phaeophyceae (11%). Entre ambas localidades se compartieron 16 especies. Las familias con mayor número de especies fueron Rhodomelaceae con 12 especies, Corallinaceae con 10 y Ceramiaceae y Dictyotaceae con 5 especies cada una (tabla 5).
Asociación entre grupos morfofuncionales y moluscos. Las macroalgas se clasificaron en 6 grupos morfofuncionales: algas filamentosas, foliosas, foliosas corticadas, filamentosas corticadas, coriáceas y calcáreas articuladas (tabla 5). Los grupos dominantes en las 2 localidades fueron las algas filamentosas, filamentosas corticadas y las calcáreas articuladas. En playa El Palmar se presentó una mayor riqueza de especies (14) de bivalvos al interior de las comunidades algales que contenían el grupo de algas calcáreas articuladas. En playa Las Gatas, los grupos morfofuncionales que presentaron mayor cantidad de especies fueron las algas filamentosas, filamentosas corticadas y las calcáreas articuladas con 10 especies cada uno (tabla 6).
Cobertura de macroalgas, abundancia de moluscos y sedimentos retenidos. La regresión lineal por localidad indicó que para playa El Palmar hay una correlación baja con 24% entre los componentes: cobertura de macroalgas, abundancia de bivalvos y peso de sedimento húmedo; sin embargo, se observó una relación moderada entre los sedimentos retenidos por las macroalgas y la abundancia de bivalvos (R = 0.55 y R2 = 0.24). Para playa Las Gatas la regresión lineal múltiple indicó una correlación baja con 10% entre los factores y una relación moderada entre la cobertura de macroalgas y la abundancia de bivalvos (R = 0.42 y R2 = 0.10). La cobertura de macroalgas, abundancia de bivalvos y los sedimentos retenidos varió según la localidad (fig. 5A-C), mes de muestreo (fig. 5D-F) y nivel de marea (fig. 5G-I). En El Palmar la regresión lineal mostró una relación directamente proporcional entre la abundancia de bivalvos y la cobertura de macroalgas. Los valores más altos de cobertura de macroalgas (0.89 m2) se relacionaron con los valores más altos de abundancia de bivalvos.
Con respecto al mes de muestreo, las regresiones lineales múltiples indicaron una correlación alta con 98% (R = 0.99 y R2 = 0.98), entre la cantidad de sedimento retenido y la abundancia de bivalvos. La regresión lineal múltiple sugirió que la abundancia de bivalvos disminuyó a lo largo del ciclo anual, donde enero presentó la mayor abundancia (de 187-230 individuos) y noviembre la menor (27 individuos). La relación de la abundancia de bivalvos con el peso húmedo de sedimento tuvo un comportamiento parecido, disminuyendo conforme al ciclo anual: en las muestras de enero se reportó mayor sedimento retenido (0.56 kg), mientras que en julio se encontraron menos sedimentos retenidos (0.17 kg).
Figura 4. Análisis de escalamiento multidimensional no métrico (nMDS) de las especies de bivalvos recolectadas en las localidades de estudio. Las líneas azules (vectores) indican las especies abundantes y frecuentes (dominantes). El círculo azul representa la variación de las abundancias.
Tabla 5
Especies de macroalgas asociadas a bivalvos, para cada localidad y grupo morfofuncional. Se marca con * la especie de macroalga presente en la localidad. Localidad: playa El Palmar (P), playa Las Gatas (G). Grupos morfofuncionales de macroalgas: filamentosas (Fil), foliosas (Fol), foliosas corticadas (Foc), filamentosas corticadas (Fic), coriáceas (Cor), calcáreas articuladas (Cal). Especies de bivalvos: 1) Linucula declivis, 2) Brachidontes adamsianus, 3) Brachidontes semilaevis, 4) Leiosolenus aristatus, 5) Modiolus capax, 6) Mytilus edulis, 7) Acar rostae, 8) Isognomon janus, 9) Carditamera affinis, 10) Carditamera radiata, 11) Cardites grayi, 12) Crassinella ecuadoriana, 13) Chama coralloides, 14) Mya sp., 15) Sphenia fragilis,16) Pinna rugosa, 17) Parvilucina approximata.
Especies
Grupo morfofuncional
Especies de bivalvos
P
G
Chlorophyta
Ulvales
Ulvaceae
Ulva californica Wille, 1899
Fol
*
–
Ulva intestinalis Linnaeus, 1753
Fol
–
2, 4, 13
Ulva linza Linnaeus, 1753
Fol
–
2, 4, 13
Bryopsidales
Bryopsidaceae
Bryopsis pennata var. minor J. Agardh, 1887
Fil
–
2, 4, 8
Caulerpaceae
Caulerpa chemnitzia (Esper) J.V. Lamouroux, 1809
Fil
2, 4, 8, 10, 17
1, 2, 4, 7, 8, 9, 10, 15
Caulerpa sertularioides (S.G. Gmelin) M. Howe, 1905
Fil
2, 4, 8, 10, 17
1, 2, 4, 7-10, 15
Halimedaceae
Halimeda discoidea Decaisne, 1842
Cal
2, 10, 17
–
Cladophorales
Cladophoraceae
Chaetomorpha antennina (Bory) Kützing, 1847
Fil
2, 3, 7, 8-13
4, 7, 8, 11, 13
Cladophora sp.
Fil
2, 4, 5, 8, 9
–
Cladophora graminea Collins, 1909
Fil
–
16
Cladophora microcladioides Collins, 1909
Fil
2, 4, 5, 8, 9
–
Rhodophyta
Gigartinales
Cystocloniaceae
Hypnea pannosa J. Agardh, 1847
Fic
2-6, 8-15
2, 4, 8-10, 16
Hypnea spinella (C. Agardh) Kützing, 1847
Fic
2-6, 8-15, 17
2, 4, 8-10, 16
Hypnea johnstonii Setchell y N. L. Gardner, 1924
Fic
–
1, 2, 4, 7-10, 16
Phyllophoraceae
Ahnfeltiopsis gigartinoides (J. Agardh) P. C. Silva y DeCew, 1992
Fic
8
–
Gymnogongrus johnstonii (Setchell y N. L. Gardner) E.Y. Dawson, 1961
Fic
2, 8
–
Ceramiales
Ceramiaceae
Centroceras clavulatum (C. Agardh) Montagne, 1846
Fil
2-5, 8
2, 4, 10, 15
Tabla 5. Continúa
Especies
Grupo morfofuncional
Especies de bivalvos
P
G
Ceramium sp.
Fil
–
1, 2, 4, 7, 8, 10, 13, 16
Ceramium camouii E. Y. Dawson, 1944
Fil
–
1, 2, 4, 7, 8, 10, 13, 16
Ceramium zacae Setchell y N. L. Gardner, 1937
Fil
–
1, 2, 4, 7, 8, 10, 13, 16
Gayliella flaccida (HarveyyKützing) T.O. McIvory y L.J. Cho, 2008
Fic
–
2, 4, 7, 8, 11
Delesseriaceae
Taenioma perpusillum (J. Agardh) J. Agardh, 1863
Fil
–
4, 16
Rhodomelaceae
Chondria sp.
Fic
*
*
Herposiphonia secunda (C. Agardh) Ambronn, 1880
Fic
8
2, 4, 8, 10
Herposiphonia tenella (C. Agardh) Ambronn, 1880
Fic
7
–
Melanothamnus simplex (Hollenberg) Díaz-Tapia y Maggs, 2017
Fic
2, 8
–
Melanothamnus sphaerocarpus (Borgesen) Díaz.Tapioa y Maggs, 2017
Fic
–
2, 4, 8
Polysiphonia mollis J. D. Hooker y Harvey, 1847
Fic
*
2, 4, 8, 15
Polysiphonia nathanielii Hollenberg, 1958
Fic
–
2, 4, 8, 15
Polysiphonia subtilissima Montagne, 1840
Fic
–
2, 4, 8, 15
Eutrichosiphonia confusa (Hollenberg) Savoie y G.W. Saunders, 2019
Fic
–
2, 4, 8
Laurencia sp.
Fic
2, 8,
2, 7, 8, 13
Laurencia hancockii E.Y. Dawson, 1944
Fic
2, 8
–
Laurencia subcorymbosa E.Y. Dawson, 1963
Fic
2, 8
–
Rhodymeniales
Rhodymeniaceae
Tayloriella dictyurus (J. Agardh) Kylin, 1956
Fil
2, 8
–
Lomentariaceae
Ceratodictyon tenue (Setchell y N. L. Gardner) J.N. Norris, 2014
Fil
–
2, 4, 8
Corallinales
Corallinaceae
Amphiroa beauvoisii J.V. Lamouroux, 1816
Cal
2-15, 17
1, 2, 4, 7-14, 16
Amphiroa misakiensis Yendo, 1902
Cal
2-15
1, 2, 4, 7-14, 16
Amphiroa rigida J.V. Lamouroux, 1816
Cal
2-15
1, 2, 4, 7-14, 16
Amphiroa subcylindrica E.Y. Dawson, 1953
Cal
2-15
–
Jania capillacea Harvey, 1853
Cal
–
2, 4, 7-10, 15, 16
Jania subpinnata E.Y. Dawson, 1953
Cal
2, 4, 5, 8, 9, 12, 14
2, 4, 7-10, 15, 16
Jania tenella (Kützing) Grunow, 1874
Cal
2, 4, 5, 8, 9, 12, 14
2, 4, 7-10, 15, 16
Jania tenella var. tenella
Cal
2, 4, 5, 8, 9, 12, 14
–
Gelidiales
Gelidiaceae
Gelidiella acerosa (Forsskal) Feldmann y Hamel, 1934
Fic
–
2, 7, 8
Gelidium mcnabbianum (E.Y. Dawson) B. Santelices, 1998
Grateloupia huertana Mateo-Cid, Mendoza-González y Gavio, 2005
Fic
–
4, 8
Grateloupia versicolor (J. Agardh) J. Agardh, 1847
Fic
2, 4, 8
–
Heterokontophyta-Phaeophyceae
Ectocarpales
Scytosiphonaceae
Chnoospora minima Papenfuss, 1956
Fic
*
–
Fucales
Sargassaceae
Sargassum liebmannii Agardh, 1847
Cor
2, 4, 5, 7, 8, 10-13, 15, 17
–
Dictyotales
Dictyotaceae
Dictyota sp.
Foc
–
*
Dictyota dichotoma (Hudson) J.V. Lamouroux, 1809
Foc
2, 4, 5, 8
–
Lobophora variegata (J.V.Lamouroux)Womersley ex E.C.Oliveira, 1977
Fic
8
–
Padina mexicana var. erecta Avila-Ortiz, 2003
Foc
2, 4, 5, 8, 9
–
Padina ramonribae Avila-Ortiz, Pedroche y Díaz-Martínez, 2016
Foc
2, 4, 5, 8, 9
–
La regresión lineal por nivel de marea indicó una correlación significativa con 100% (R = 1 y R2 = 1), es decir, la cobertura de algas y el sedimento retenido están relacionados con la abundancia de bivalvos en los diferentes niveles de marea. El nivel del intermareal bajo presentó los valores más elevados de cobertura de macroalgas con 0.61 m2 (fig. 5G), mayor abundancia de moluscos con 275 individuos (fig. 5H) y mayor cantidad de sedimentos con 0.69 kg (fig. 5I). En el intermareal alto se registraron los valores más bajos de abundancia de bivalvos con 230 individuos (fig. 5H), cobertura de algas con 0.52 m2 (fig. 5G) y peso húmedo de sedimentos con 0.30 kg (fig. 5I), es decir, conforme se asciende en el intermareal, estas variables disminuyen.
Tabla 6
Asociación de las especies de bivalvos con los grupos morfofuncionales propuestos por Steneck y Dethier (1994). Grupos morfofuncionales de macroalgas: filamentosas (Fil), foliosas (Fol), foliosas corticadas (Foc), filamentosas corticadas (Fic), coriáceas (Cor), calcáreas articuladas (Cal). × = Presencia de la especie.
Familia
Especie
Playa El Palmar
Playa Las Gatas
Fil
Fol
Foc
Fic
Cor
Cal
Fil
Fol
Foc
Fic
Cor
Cal
Nuculidae
L. declivis
–
–
–
–
–
–
×
–
–
×
–
×
Mytilidae
B. adamsianus
×
–
×
×
×
×
×
×
–
×
–
×
B. semilaevis
×
–
–
×
–
×
–
–
–
–
–
–
L. aristatus
×
–
×
×
×
×
×
×
–
×
–
×
M. capax
×
–
×
×
×
×
–
–
–
–
–
–
M. edulis
–
–
–
×
–
×
–
–
–
–
–
–
Arcidae
A. rostae
×
–
–
–
×
×
×
–
–
×
–
×
Pteriidae
I. janus
×
–
×
×
×
×
×
–
–
×
–
×
Pinnidae
P. rugosa
–
–
–
–
–
–
×
–
–
×
–
×
Lucinidae
P. approximata
×
–
–
×
×
×
–
–
–
–
–
–
Carditidae
C. affinis
×
–
×
×
–
×
×
–
–
×
–
×
C. radiata
×
–
–
×
×
×
×
–
–
×
–
×
C. grayi
×
–
–
×
×
×
×
–
–
×
–
×
Crassatellidae
C. ecuadoriana
×
–
–
×
×
×
–
–
–
–
–
–
Chamidae
C. coralloides
×
–
–
×
×
×
×
×
–
×
–
×
Myidae
Mya sp.
–
–
–
×
–
×
–
–
–
–
–
–
S. fragilis
–
–
–
×
×
×
×
–
–
×
–
×
Total
12
0
5
14
11
15
11
3
0
11
0
11
Discusión
Riqueza y composición de moluscos. De las 17 especies encontradas en el presente estudio, 16 ya habían sido observadas para el Pacífico tropical mexicano, específicamente para Ixtapa-Zihuatanejo en sustratos rocosos y arenosos (Flores-Rodríguez et al., 2007, 2012; Lesser, 1984; López-Rojas et al.,2017; Lozada, 2010; Sánchez, 2014), excepto Mytilus edulis que es un nuevo registro para Guerrero. Esta especie fue encontrada en playa El Palmar, asociada a los géneros de macroalgas Amphiroa e Hypnea. Dicha especie había sido registrada para el océano Pacífico desde las costas del Ártico, Canadá, EUA, hasta Cabo San Lucas, México, al interior del golfo de California en San Luquitas y Santa Rosalía, y para el Pacífico tropical mexicano en isla Socorro, por lo que su distribución se amplía hacia el sur, a Guerrero en ambientes litorales asociados a las comunidades de macroalgas (Cadena-Cárdenas et al., 2009; Fitch, 1953; Shaw et al., 1988). La ampliación de la distribución de esta especie puede deberse al movimiento de la corriente de California como ha sido reportado para bivalvos (Schulien et al., 2020), gasterópodos y escafópodos que ampliaron su distribución desde la provincia Californiana hacia la Panámica donde se señala la confluencia de especies de moluscos entre ambas provincias (Landa-Jaime y Arciniega-Flores 1998; Ríos-Jara et al., 2003; Gama-Kwick et al., 2021). Otra de las razones por la cual M. edulis amplió su distribución al sur podría ser por el fenómeno de El Niño que proporciona condiciones ambientales para que las especies que habitan en sitios templados y subtropicales se desplacen a sitios tropicales (Díaz y Ortlieb, 1993; Paredes et al., 1998; Velez y Zeballos, 1985).
Figura 5. Variación de cobertura de macroalgas, abundancia de bivalvos y sedimento retenido por localidades (A, B y C), mes de muestreo (D, E y F) y nivel de la zona intermareal (G, H e I). Verde = macroalgas, amarillo = bivalvos, rojo = sedimento retenido.
La familia mejor representada fue Mytilidae con 5 especies: Brachidontes adamsianus, B. semilaevis, Leiosolenus aristatus, Modiolus capax y Mytilus edulis (tabla 1), dichas especies coinciden con lo registrado por Flores-Garza et al.(2014), Galeana-Rebolledo et al.(2012) y Garcés (2011) en Acapulco, así como por López-Rojas et al. (2017) en diferentes localidades en Guerrero. Mytilidae también ha sido la familia mejor representada en otras localidades de Zihuatanejo (muelle municipal), donde se han observado 8 especies (Guzmán, 2022), así como en otras regiones tropicales como Brasil, donde esta familia también fue la mejor representada con 5 especies (Santos et al., 2020).
Una mayor riqueza de especies de la familia Mytilidae en Ixtapa-Zihuatanejo podría explicarse debido a la frecuencia y rápido asentamiento de larvas de bivalvos, ya que son organismos que están en constante reproducción a lo largo del año (Seed, 1969a). Suchanek (1978) mencionó que los miembros de la familia Mytilidae tienden a colonizar rápidamente los espacios disponibles en la zona intermareal rocosa, familia que se ha adaptado a distintos hábitats (Keen, 1971), gracias a su rápido aumento de talla y posterior asentamiento de larvas (Ceccherelli y Rossi, 1984). También se ha mencionado que los organismos de dicha familia pueden producir fibras de biso, por lo que son capaces de anclarse y permanecer en sustratos en los que otras familias de bivalvos no pueden, como rocas, arenas u otros organismos (Keen, 1971; Stella et al.,2010).
En general, se encontró una mayor riqueza de especies en El Palmar, valores que pueden explicarse debido a la heterogeneidad ambiental de una playa expuesta con oleaje intenso como dicho sitio (Morales et al., 2008). Se ha mencionado que la riqueza de especies de bivalvos es mayor en zonas expuestas en donde las olas impactan directamente, por su parte, las zonas protegidas o de menor oleaje presentan menor riqueza específica (Flores-Garza et al., 2014; Flores-Rodríguez et al., 2012; Valdés-González et al., 2004).
En El Palmar, se encontraron valores elevados para la riqueza de especies de bivalvos (17 spp.) respecto de lo encontrado en Las Gatas, lo anterior también se ha observado para otras clases de moluscos como los gasterópodos y poliplacóforos asociados a comunidades de macroalgas en playa El Palmar (Aguilar, 2017). También en esta playa se encontró menor cantidad de sedimentos retenidos por las macroalgas (fig. 5C). El Palmar está conformada por rocas de diferentes tamaños, que pueden moverse conforme se dan los cambios en el oleaje y la marea, lo que le confiere un alto dinamismo que no permite la acumulación de sedimentos (Gibbons, 1988), tal como se observó en el presente estudio. Dichas características ayudan a que se conformen una amplia gama de ambientes que pueden ser colonizados, donde se esperaría una mayor riqueza de especies como se observó para los bivalvos de El Palmar (Benedetti-Cecchi, 2001).
Como se ha mencionado anteriormente, playa Las Gatas es un sitio protegido, por esta razón es un lugar de baja energía, con características poco favorables para el establecimiento de especies de bivalvos de manera muy similar a los estudios realizados en otras zonas de Guerrero (Flores-Garza et al., 2014; Flores-Rodríguez et al., 2012; Valdés-González et al., 2004). Las fluctuaciones observadas entre ambas playas de la riqueza de especies, también podrían deberse a otro tipo de características presentes en algunas familias de bivalvos que no tienen hábitos epifaunales como los que se adhieren a las macroalgas y que presentan hábitos de vida semiinfaunales o infaunales (Garcés, 2011), por lo que estas especies podrían encontrarse en otros sitios, posiblemente en zonas más profundas en la zona submareal, o de igual forma, las especies de bivalvos se pueden ver afectadas por la contaminación al incorporar en sus tejidos bacterias patógenas (Gosling, 2015), que es común de una zona turística como Ixtapa-Zihuatanejo (IMTA, 2010; UNAM, 2013). Lo anterior podría explicar las fluctuaciones en la riqueza de especies de bivalvos observadas en Zihuatanejo.
Los valores de riqueza de especies no variaron considerablemente en las localidades estudiadas a lo largo del año. La riqueza de especies de bivalvos en un ciclo anual puede ser constante, ya que la mayoría de las especies (70%) de bivalvos del presente trabajo son generalmente inmóviles o sedentarias. Lo anterior hace que sus poblaciones se mantengan con pocos cambios con respecto a su riqueza de especies debido a sus patrones de reproducción anuales o bianuales (Baqueiro y Masso, 1988; Flores-Rodríguez et al., 2012; Seed, 1969a).
La especie más abundante en el presente trabajo fue B. adamsianus (Mytilidae). Esta especie ha sido registrada como la más abundante en otras localidades de Zihuatanejo (Guzmán, 2022). En particular, las familias Chamidae y Mytilidae se han registrado como las más abundantes para Guerrero; por su parte, la especie Isognomon janus Carpenter, 1857 (Isognomonidae) también ha presentado un gran número de organismos (Guzmán, 2022; López-Rojas et al. 2017). Las bajas abundancias de los bivalvos encontrados en este trabajo pueden estar dadas por el tipo de hábitos de vida de dichos moluscos. Por ejemplo, el género Sphenia tienen un hábito endolítico (Esqueda-González et al., 2014; Garcés, 2011; Guzmán, 2022), por lo que sería poco probable encontrarlos dentro de las comunidades de macroalgas, ya que es común encontrarlos incrustados en agujeros preexistentes de rocas, fragmentos de madera, incluso otros materiales como las conchas de moluscos, así como entre colonias de briozoos (Coan, 1999). El Permanova mostró que existen diferencias significativas entre localidades, esto puede deberse a que playa Las Gatas es considerada como un sitio protegido y playa El Palmar como un sitio expuesto, esto representó diferencias en abundancia y riqueza de especies, que pueden deberse a los requerimientos medioambientales propios de cada una de las especies de bivalvos como: sustrato, alimento, salinidad o temperatura (Borges et al., 2014; Galeana-Rebolledo et al., 2012; Seed 1969a).
En playa El Palmar, la curva de acumulación de especies fue asintótica, lo que sugiere que se encontró a la mayoría de los bivalvos asociados a macroalgas de esta localidad. Mientras que, en playa Las Gatas, la curva de acumulación de especies no fue asintótica. La cantidad de especies recolectadas en un sitio está relacionada con el esfuerzo de muestreo (Moreno, 2001). Por lo que el menor número de muestras en donde se encontraron bivalvos en playa Las Gatas (25), en comparación con las de playa El Palmar (35), podría explicarse por este hecho. Se ha mencionado que una curva de acumulación de especies en muy raras ocasiones llega a ser asintótica, ya que siempre habrá especies que no se recolecten o durante los muestreos pueden encontrarse especies raras, lo que puede estar determinado por el sitio de muestreo, temporada del año, tipo de sustrato, entre otras variables (Jiménez-Valverde y Hortal, 2003).
Además, otra de las razones para que la curva de acumulación de especies no haya sido asintótica para Las Gatas, podría deberse a que, en ambas localidades, las muestras de bivalvos fueron obtenidas de comunidades de macroalgas, que es un sustrato muy específico y es distinto a las rocas y arena que componen la zona intermareal que corresponden a los sustratos que se han estudiado en la mayoría de los distintos trabajos malacológicos de Guerrero. Por lo anterior, el inventario podría estar incompleto; sin embargo, es una buena aproximación de la biodiversidad de las especies de bivalvos asociadas a comunidades de macroalgas de la zona norte de Guerrero en Ixtapa-Zihuatanejo. Para tener una curva de acumulación de especies asintótica se recomienda muestrear todos los sustratos posibles y de esta forma, obtener casi todas las especies de bivalvos de la zona.
Se encontraron cambios con respecto a la abundancia de bivalvos asociados a macroalgas a lo largo del año, fluctuaciones que pudieran estar relacionadas con sus patrones reproductivos, lo que podría explicar la disminución en la abundancia de bivalvos en los meses cálidos (mayo y julio) y un aumento en la cantidad de individuos en los meses fríos (enero y noviembre). Los cambios en la abundancia de bivalvos se han estudiado en trabajos sobre reproducción de diferentes especies de moluscos (Baqueiro y Aldana, 2000, 2003). Algunas de las especies de bivalvos que han sido utilizadas en estudios reproductivos o sobre ciclos gonádicos son: Chione undatella (G. B. Sowerby I, 1835), Megapitaria aurantiaca (G. B. Sowerby I, 1831) (Veneridae), Brachidontes rodriguezii (d’Orbigny, 1842), Mytilus edulis y Mytilus chilensis Hupé, 1854 (Mytilidae), así como Anadara tuberculosa (G. B. Sowerby I, 1833) (Arcidae), tanto en México como otros países de Sudamérica. Dichos estudios destacan que los bivalvos se reproducen continuamente a lo largo del año y se ha mencionado que el momento del desove comprende desde la primavera hasta el otoño con temperaturas por encima de los 25 °C; se ha observado que las larvas se instalan en octubre y noviembre, con una metamorfosis de 15 a 25 días, dichas larvas permanecen adheridas a las algas de noviembre a mayo, en donde llevan a cabo su metamorfosis (Aguillón, 2011; Baqueiro y Masso, 1988; García-Domínguez et al., 2008; Hernández-Moreno et al., 2020; Oyarzún et al., 2011; Seed, 1969a; Torroglosa, 2015). Solo se encontró a Mytilus edulis como especie de importancia comercial para el presente trabajo. En especies que no se les ha considerado importantes para la industria pesquera, es muy poco lo que se conoce acerca de sus hábitos reproductivos (Aguilar, 2017).
Las 3 especies más abundantes del presente trabajo son epifaunales de hábitos filtradores por suspensión (García-Cubas, 1981), Isognomon janus (Isognomonidae) habita en la zona intermareal hasta profundidades de 20 m, Leiosolenus aristatus (Mytilidae)puede encontrarse desde el litoral hasta los 300 m sobre rocas u otros bivalvos (Coan y Valentich-Scott, 2012) y Brachidontes adamsianus (Mytilidae)habita sobre grietas de rocas grandes en zonas expuestas (Landa-Jaime et al., 2013). Las especies que no pertenecen a las familias Isognomonidae y Mytilidae, en general, son organismos infaunales de hábitos filtradores por suspensión, a excepción de la familia Myidae que son infaunales perforadores de hábitos filtradores por suspensión (García-Cubas, 1981; Coan, 1999).
Los bivalvos están presentes en diferentes lugares al interior de la zona intermareal rocosa y hay especies que tienen preferencia por algún sitio a lo largo de dicha zona (Román-Contreras et al.,1991; Sibaja-Cordero y Vargas-Zamora, 2006; Suchanek, 1978). En el presente estudio se observó un incremento en la abundancia y riqueza en los niveles intermareales medio y bajo. Este mismo patrón se ha observado en otras especies de moluscos como gasterópodos y poliplacóforos de Zihuatanejo (Aguilar, 2017; Gama-Kwick et al., 2021).
El índice de diversidad de Shannon obtenido en este trabajo fue bajo, menor a 2.16 bits/individuo para ambos sitios de muestreo, en comparación con otros trabajos sobre bivalvos asociados a macroalgas para Zihuatanejo, como el de Guzmán (2022), donde se encontraron valores de 3.41 bits/individuo en el muelle municipal. Flores-Garza et al.(2014), en su estudio en Acapulco, presentaron valores de 3.65 bits/individuo y Galeana-Rebolledo et al.(2012) reportaron valores de 3.64 bits/individuo. Las diferencias con respecto al índice de diversidad de Shannon pueden deberse a la baja riqueza de especies encontrada en Ixtapa-Zihuatanejo, respecto de otras localidades del Pacífico tropical mexicano, donde se han registrado valores superiores a 17 especies; Reguero y García-Cubas (1989) encontraron 53 especies de bivalvos para Nayarit, y Esqueda-González et al. (2014) reportaron 89 especies de bivalvos para Sinaloa. Lo anterior también puede relacionarse con los altos valores de abundancia de ciertas especies, como es el caso de Brachidontes adamsianus. Ésto también se ve reflejado en los valores calculados del índice de dominancia de Simpson, el cual dio como resultado valores moderados, ya que la abundancia de las especies de bivalvos no es homogénea a lo largo de la zona intermareal rocosa en ambas localidades del presente estudio.
Garcés (2011), describió valores similares del índice de diversidad de Shannon para especies de bivalvos en sustratos rocosos en Acapulco (2.41 bits/individuo) y mencionó que éstos varían según el tipo de sustrato, ya que la riqueza de especies fue mayor en sustratos arenosos que en sustratos rocosos, lo que ocasiona que en estos sustratos, los valores disminuyan.
En el nMDS se observan 3 agrupaciones: una con muestras de la localidad de playa El Palmar, otra con estaciones de playa Las Gatas y la tercera con estaciones de ambas localidades, donde solamente hubo presencia de la especie Brachidontes adamsianus (tabla 1). Dicha especie es una de las más comunes en varias localidades de Jalisco, Oaxaca y Guerrero, por lo que sus valores de abundancia en Ixtapa-Zihuatanejo concuerdan con lo encontrado en otras contribuciones (Castro-Mondragón et al., 2016; Galeana-Rebolledo et al., 2012; Garcés, 2011; Holguín-Quiñones y González-Pedraza 1989; Landa-Jaime et al., 2013; López-Rojas et al., 2017; Torreblanca-Ramírez et al., 2012).
Asociación entre grupos morfofuncionales y moluscos. Los bivalvos recolectados tuvieron una mayor presencia en el grupo morfofuncional de algas filamentosas en ambas localidades, este mismo resultado se observó en muelle municipal en Zihuatanejo por Guzmán (2022) para los bivalvos asociados a macroalgas. Lo anterior puede verse favorecido porque este grupo de algas proporciona refugio ante el oleaje y la desecación, de igual forma tienen un papel fundamental en el asentamiento de las larvas de bivalvos, ya que son utilizadas como un sitio para evitar la competencia entre estadios juveniles y adultos (Dobretsov, 1999; Seed, 1969a).
Las larvas de la familia Mytilidae son atraídas hacia los filamentos de algas rojas de los géneros Ceramium y Polysiphonia, en donde llevan a cabo su metamorfosis (Seed, 1969a), ambos géneros fueron encontrados en el presente trabajo. Las especies de bivalvos de la familia Mytilidae también se encontraron junto con otros géneros de macroalgas como: Bryopsis, Caulerpa, Chaetomorpha y Cladophora (Chlorophyta), dichos géneros también se han relacionado a comunidades de bivalvos en el Caribe colombiano (Quirós-Rodríguez y Campos, 2013). Los géneros Gayliella, Herposiphonia, Taenioma y Tayloriella (Rhodophyta) también se han observado junto a especies de bivalvos, en particular el género Tayloriella se ha encontrado vinculado a 9 diferentes especies de bivalvos en el muelle municipal en Zihuatanejo (Guzmán, 2022). Aun cuando es poco lo que se conoce sobre las asociaciones de bivalvos con macroalgas, es posible sugerir que las larvas de las otras familias de bivalvos encontradas en el presente trabajo (Arcidae, Carditidae, Crassatellidae, Chamidae, Myidae y Pteriidae) puedan tener un comportamiento similar al de Mytilidae y, por esta razón, se podría reconocer su presencia en las comunidades de macroalgas.
La asociación de las especies de bivalvos (B. adamsianus, B. semilaevis, L. aristatus, M. capax, M. edulis, A. rostae, I. janus, C. affinis, C. radiata, C. grayi, C. ecuadoriana, C. coralloides, Mya sp. y S. fragilis) con el grupo de algas filamentosas corticadas fue explicado para especies del género Gigartina porSeed (1969a), quien afirmó que este tipo de algas proporciona una mayor protección y partículas de sedimento retenido (alimento), comparado con las algas filamentosas. En el presente trabajo se encontraron especies de los géneros Ceratodyction, Gelidiella, Gelidium, Gracilaria e Hypnea, que podrían tener una relación similar a la dada por las especies de Gigartina, ya que comparten el mismo grupo morfofuncional propuesto por Steneck y Dethier (1994).
Todas las especies de bivalvos se asociaron al grupo de algas calcáreas articuladas. Las especies del género Corallina proporcionan condiciones ambientales adecuadas para los bivalvos como: refugio, captación de partículas y cantidad de CaCO3 (Seed, 1969a). De esta forma, los bivalvos son capaces de alcanzar tallas mayores en menor tiempo (Seed, 1969a, b). Probablemente, la cantidad de CaCO3 obtenida del ambiente por las macroalgas, queda disponible para que los bivalvos la aprovechen, una vez que las macroalgas mueren. En el presente trabajo, se observó a las especies de algas calcáreas articuladas de los géneros Amphiroa, Halimeda y Jania asociadas a las especies de bivalvos son muy similares a las del género Corallina y comparten el mismo grupo morfofuncional, por lo que podrían funcionar de la misma forma al ser algas que pueden fijar CaCO3 y ser una fuente de aprovechamiento para otros organismos; estas algas calcáreas son la fuente principal de carbonatos marinos (Feely et al., 2004).
Cobertura de macroalgas, abundancia de moluscos y sedimento retenido. La cobertura de macroalgas, abundancia de bivalvos y sedimento retenido disminuyeron conforme se aumentó el nivel del intermareal, los valores más elevados para dichas variables se observaron en el nivel del intermareal bajo (fig. 5G-I). Los resultados de la regresión lineal múltiple sustentan lo anterior, ya que se encontró una relación entre la cobertura de macroalgas y abundancia de bivalvos. Lo anterior podría explicarse por las características del terreno (emersión), debido a que, en un ambiente cambiante como la zona intermareal rocosa, las macroalgas quedan expuestas a una mayor radiación solar en el nivel alto. Por lo tanto, al haber menos céspedes algales (Huovinen et al., 2006), la retención de sedimento es baja, estos 2 factores influyen en las necesidades de cada especie de bivalvo, lo que determina la supervivencia de estos moluscos en esta zona (Airoldi, 2003; Rosenberg, 1977).
Renaud et al. (1997) encontraron una relación entre la abundancia de macroalgas con respecto al sedimento, donde la cobertura de macroalgas era sistemáticamente mayor en las zonas con baja cantidad de sedimentos y un aumento notable en la abundancia de macroalgas se observó después de la remoción de sedimentos. Esto se relaciona con lo encontrado en el presente trabajo, donde se observó una menor cobertura de macroalgas asociada a valores elevados de sedimento. Con respecto de los bivalvos, Forster y Zettler (2004) observaron que la biomasa de Mya arenaria Linnaeus, 1758 se redujo con la presencia de sedimentos finos, lo que podría sugerir que en playa Las Gatas existe una menor abundancia de bivalvos (Airoldi, 2003).
El sedimento en las comunidades de macroalgas queda retenido ya que se acumula en los espacios que hay entre los talos; las cantidades de sedimento acumuladas pueden estar determinadas por la complejidad estructural del alga y las condiciones del medio (García, 2009). La variación en la tasa de sedimentación podría ser un factor de alteración y estrés sobre las comunidades de macroalgas, lo que ocasiona que la tasa de crecimiento algal se vea reducida por la falta de incidencia de luz. Por ello, la fauna que ahí se establece, como las especies de bivalvos, también es menor, ya que una gran cantidad de sedimentos retenidos provoca que los organismos puedan sofocarse y el reclutamiento larval se vea reducido (Airoldi, 2003; Rosenberg, 1977). La clase Bivalvia tiene una preferencia por sustratos arenosos, rocosos, lodosos y fangosos, sin embargo, cuando los sitios no tienen corrientes fuertes y la acumulación de sedimento es alta, la abundancia de bivalvos baja drásticamente (Gosling, 2015).
Las larvas de bivalvos se instalan sobre las algas, donde permanecen adheridas a ellas y llevan a cabo su metamorfosis entre 15 y 25 días; en este lapso, los céspedes algales les proporcionan alimento por el sedimento que retienen y protección contra depredadores y factores abióticos. Posteriormente, los bivalvos juveniles migran fuera de las algas ya que han alcanzado una talla óptima o las algas ya no les brindan suficientes recursos, y llegan a otro sitio que será el definitivo para llevar a cabo el resto de su ciclo de vida (Seed, 1969a, 1969b; Suchanek, 1978).
Olafsson (1986) encontró una relación significativa entre la abundancia de bivalvos y el sedimento, estos resultados son consistentes con lo encontrado en el presente estudio en Ixtapa-Zihuatanejo. Sin embargo, Vázquez (2009) en playa Las Gatas no encontró relación entre la riqueza de especies y el sedimento retenido. Por su parte, en playa El Palmar, que es una zona expuesta, una posible explicación para la relación entre abundancia de bivalvos y sedimento es debido al tipo de hábitos de vida de la mayoría de las especies encontradas en el presente trabajo, ya que la mayoría de ellas son epifaunales o infaunales con hábitos filtradores suspensívoros. La erosión del sustrato rocoso por la acción de las olas y por el movimiento de los sedimentos puede generar heterogeneidad ambiental (Airoldi, 2003). La mezcla constante del agua provee mayor humedad, así como nutrientes al suspender los sedimentos, lo que ocasiona una mayor supervivencia de los organismos, lo que explicaría la relación entre abundancia y sedimento (Gama-Kwick et al., 2021).
Las condiciones de playa El Palmar con un oleaje intenso (Morales et al., 2008), no permite la retención de sedimentos, su baja cantidad podría influir en la composición de las macroalgas. Por lo tanto, la composición de los invertebrados al interior de éstos puede ser muy particular en esta localidad (Gama-Kwick et al., 2021), lo que puede incrementar su diversidad (Chemello y Milazzo, 2002; Prathep et al., 2003). En contraste, playa Las Gatas es un sitio protegido con un oleaje de baja energía y alta cantidad de sedimentos, por lo que su comunidad de macroalgas y bivalvos puede ser menos diversa, abundante y compleja. Lo anterior es similar a lo encontrado en trabajos de la zona para otras clases de moluscos como gasterópodos y poliplacóforos (Aguilar, 2017).
Las comunidades de macroalgas pueden funcionar como sustrato de captura de las larvas desde la columna de agua (Aguilar, 2017). Dichos sitios constituyen zonas de crianza en parte o en todo el ciclo de vida de los bivalvos, ya que proporcionan protección y alimento (Seed, 1969a, b), que está determinado por los sedimentos que las macroalgas retienen.
El presente trabajo aporta resultados que contribuyen al conocimiento de la biodiversidad marina, en especial de los bivalvos asociados a la ficoflora del Pacífico tropical mexicano. Los esfuerzos posteriores deberían centrarse en explorar la biodiversidad de los invertebrados sobre sustratos diferentes en el litoral rocoso, posiblemente utilizando metodologías similares a las del presente estudio con la finalidad de generar comparaciones. En los estudios futuros se debe profundizar en el conocimiento de la distribución geográfica de Mytilus edulis y así corroborar los resultados de este trabajo. Se debe promover el estudio de las interacciones ecológicas, ya que muchas especies de moluscos son impactadas por diferentes factores tanto bióticos, como la depredación o la epibiosis (Aguilar-Estrada et al., 2022; García-Ibáñez et al., 2014; Quiroz-González et al., 2020), así como por distintas condiciones abióticas como circulación del agua, temperatura, pH, régimen de mareas y sedimento (López et al., 2017, 2023). Por ello, es fundamental realizar más investigaciones enfocadas hacia el estudio de la variación espacio-temporal de los organismos en periodos distintos a un ciclo anual en diferentes localidades de México, con el objetivo de sentar las bases pertinentes para su conservación y posterior manejo ante el incremento del desarrollo de infraestructuras turísticas/urbanas en zonas como Guerrero, que pueden tener un efecto desfavorable sobre las comunidades intermareales a largo plazo (Zamorano y Leyte-Morales, 2009).
Agradecimientos
Al proyecto DGAPA-PAPIIT, UNAM (IN220714), al Registro Nacional de Pesca y Acuacultura por el permiso para la recolección de material biológico (DF00000208). A Norma López por el préstamo de las instalaciones de la UMDI-Zihuatanejo, a Carlos Candelaria por su apoyo técnico en campo y a Isabel Bieler, por su apoyo en la toma de las fotografías de los ejemplares para este estudio.
Referencias
Abbott, I. A. (1999). Marine red algae of the Hawaiian Island. Honolulu, Hawaii: Bishop Museum Press.
Abbott, I. A. y Hollenberg, G. J. (1976). Marine algae of California. Stanford, California: Sanford University Press.
Aguilar, L. G. (2017). Estructura comunitaria de los moluscos (gasterópodos pateliformes y poliplacóforos) asociados a los ensambles macroalgales en el intermareal rocoso de Ixtapa-Zihuatanejo, Guerrero, México (Tesis de maestría). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Aguilar-Estrada, L. G., Ortigosa, D., Urbano, B. y Reguero, M. (2014). Análisis histórico de los gasterópodos de la laguna arrecifal de Isla Verde, Veracruz, México. Revista Mexicana de Biodiversidad, 85, 502–512. https://doi.org/10.7550/rmb. 33802
Aguilar-Estrada, L. G., Ruiz-Boijseauneau, I. y Rodríguez, D. (2017). Estadios juveniles de las especies de gasterópodos pateliformes y de poliplacóforos (Mollusca) asociados a macroalgas intermareales de Guerrero, México. Revista Mexicana de Biodiversidad, 88, 280–299. https://doi.org/10.1016/j.rmb.2017.03.021
Aguilar-Estrada, L. G., Quiroz-González, N., Ruiz-Boijseauneau, I., Álvarez-Castillo, L. y Rodríguez, D. (2022). Algal epibiont species on Chiton articulatus (Mollusca: Polyplacophora) from a rocky intertidal coast from the Mexican Tropical Pacific. Revista Mexicana de Biodiversidad, 93, e934163. https://doi.org/10.22201/ib.20078706e.2022.93.4163
Aguilera, M. A. (2011). The functional roles of herbivores in the rocky intertidal systems in Chile: A review of food preferences and consumptive effects. Revista Chilena de Historia Natural, 84, 241–261.
Aguillón, A. (2011). Variación espacio-temporal del recluta- miento en Mollusca y Echinodermata en bahía de La Paz, Baja California Sur, México (Tesis de maestría). Ciudad de México: Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional.
Airoldi, L. (2003). The effects of sedimentation on rocky coast assemblages. Oceanography and Marine Biology an Annual Review, 41, 167–171.
Anderson, M., Gorley, R. y Clarke, K. (2008). Permanova+ for PRIMER: guide to software and statistical methods, PRIMER-E, Plymouth.
Bakus, G. J. (2007). Quantitative analysis of marine biological communities: field biology and environmental. Hoboken, New Jersey: John Wiley and Sons Inc.
Baqueiro, E. (1979). Sobre la distribución de Megapitaria aurantiaca (Sowerby), M. squalida (Sowerby) y Dosinia ponderosa (Gray) en relación a la granulometría del sedimento (Bivalvia: Veneridae). Anales del Centro de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 6, 25–32.
Baqueiro, E. y Aldana, D. (2000). A review of reproductive patterns of bivalve mollusks from Mexico. Bulletin of Marine Science, 66, 13–27.
Baqueiro, E. y Aldana, D. (2003). Patrones en la biología poblacional de moluscos de importancia comercial en México. Revista de Biología Tropical, 51, 97–107.
Baqueiro, E. y Masso, J. A. (1988). Variaciones poblacionales y reproducción de dos poblaciones de Chione undatella (Sowerby, 1835) bajo diferentes regímenes de pesca en la bahía de La Paz, BCS México. Ciencia Pesquera, 6, 51–67.
Barba-Marino, F., Flores-Rodríguez, P., Flores-Garza, R., García-Ibáñez, S. y Arana-Salvador, D. G. (2010). Biodiversidad y zonificación de la comunidad de moluscos, que habita el sustrato rocoso en dos sitios con distinta acción del oleaje, en la Isla “La Roqueta”, Acapulco, Guerrero, México. En L. J. Rangel, J. Gamboa, S. L. Arriaga y W. M. Contreras (Eds.), Perspectiva en malacología mexicana (pp. 44–56). Villahermosa: Universidad Juárez Autónoma de Tabasco.
Bartlett, M.S. (1937). Properties of sufficiency and statistical test. Proceedings of the Royal Society A, 160, 268–282. https://doi.org/10.1098/rspa.1937.0109
Baumgartner, T. R. y Christensen, N. (1985). Coupling of the Gulf of California to large-scale interannual climatic variability. Journal of Marine Research, 43, 825–848.
Benedetti-Cecchi, L. (2001). Variability in abundance of algae and invertebrates at different spatial scales on rocky sea shores. Marine Ecology Progress Series, 215, 79–92.
Benedetti-Cecchi, L., Rindi, F., Bertocci, I., Bulleri, F. y Cinelli, F. (2001). Spatial variation in development of epibenthic assemblages in a coastal lagoon. Estuarine, Coastal and Shelf Science, 52, 659–668. https://doi.org/10.1006/ecss.20 01.0775
Borges, L., Merckelbach, L. M., Sampaio, Í. y Cragg, S. M. (2014). Diversity, environmental requirements, and biogeography of bivalve wood-borers (Teredinidae) in European coastal waters. Frontiers in Zoology, 11, 1–13. https://doi.org/10.1186/1742-9994-11-13
Bouchet, P., Rocroi, J. P., Bieler, R., Carter, J. G. y Coan, E. V. (2010). Nomenclator of bivalve families with a classification of bivalve families. Malacologia, 52, 1–185. https://doi.org/ 10.4002/040.052.0201
Cadena-Cárdenas, L., Méndez-Rodríguez, L., Zenteno-Savín, T., García-Hernández, J. y Acosta-Vargas, B. (2009). Heavy metal levels in marine mollusks from areas with, or without, mining activities along the Gulf of California, Mexico. Archives of Environmental Contamination and Toxicology, 57, 96–102. https://doi.org/10.1007/s00244-008-9236-0
Castro-Mondragón, H., Flores-Garza, R., Valdez-González, A., Flores-Rodríguez, P., García-Ibáñez, S. y Rosas-Acevedo, J. L. (2016). Diversidad, especies de mayor importancia y composición de tallas de los moluscos en la pesca ribereña en Acapulco, Guerrero, México. Acta Universitaria, 26, 24–34. https://doi.org/10.15174/au.2016.1025.
Ceccherelli, V. U. y Rossi, R. (1984). Settlement, growth and production of the mussel Mytilus galloprovincialis. Marine Ecology Progress Series, 16, 173–184.
Cerros-Cornelio, J. C., Flores-Garza, R., Landa-Jaime, V., García-Ibáñez, S., Rosas-Guerrero, V., Flores-Rodríguez, P. et al. (2021). Composición de especies e ingreso económico por la pesca ribereña de moluscos en la Costa Grande de Guerrero México. Revista Bio Ciencias, 8, e1054. https://doi.org/10.15741/revbio.08.e1054
Chemello, R. y Milazzo, M. (2002). Effect of algal architecture on associated fauna: some evidence from phytal molluscs. Marine Biology, 140, 981–990. https://doi.org/10.1007/s00 227–002–0777–x
Cisneros, F. (2016). Estructura comunitaria de gasterópodos asociados a macroalgas en el litoral rocoso de Ixtapa-Zihuatanejo, Guerrero, México (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Clarke K. R. y Gorley, R. N. (2006) PRIMER v6: User Manual/Tutorial. PRIMER-E Ltd, Plymouth, Reino Unido.
Coan, E. V. (1999). The eastern Pacific species of Sphenia (Bivalvia: Myidae). Nautilus-Sanibel, 113, 103–120. https://doi.org/10.5962/bhl.part.2019
Coan, E. V., Scott, P. V. y Bernard, F. R. (2000). Bivalve seashells of western North America. Santa Bárbara, California: Santa Barbara Museum of Natural History Monographs.
Coan, E. V. y Valentich-Scott, P. (2006). Marine Bivalvia. En C. F. Sturm, T. A. Pearce y A. Valdés (Eds.), The mollusks: a guide to their study, collection, and preservation (pp. 339–349). Pittsburgh, Pennsylvania: American Malacological Society.
Coan E.V. y Valentich-Scott P. (2012). Bivalve seashells of tropical West America. Marine bivalve mollusks from Baja California to Peru. Santa Barbara: Santa Barbara Museum of Natural History Monographs.
Colman, J. (1940). On the faunas inhabiting intertidal seaweeds. Journal of the Marine Biological Association of the United Kingdom, 24, 129–183. https://doi.org/10.1017/S0025315400054503
Dawson, E. Y. (1949). Resultados preliminares de un reco- nocimiento de las algas marinas de la costa del pacífico de México. Revista de la Sociedad Mexicana de Historia Natural, 9, 215–255.
Dawson, E. Y. (1953). Marine red algae of Pacific Mexico. Part 1. Bangiales to Corallinaceae subf. Corallinoidae. Allan Hancock Pacific Expedition, 17, 1–239.
Dawson, E. Y. (1954). Marine red algae of Pacific Mexico. Part 2. Cryptonemiales (cont.). Allan Hancock Pacific Expedition, 17, 241–397.
Dawson, E. Y. (1960). Marine red algae of Pacific Mexico. Part 3. Cryptonemiales, Corallinaceae subf. Melobesioidae. Pacific Naturalist, 2, 3–125.
Dawson, E. Y. (1961). Marine red algae of Pacific Mexico. Part 4. Gigartinales. Pacific Naturalist, 2, 191–343.
Dawson, E. Y. (1963). Marine red algae of Pacific Mexico part 8. Ceramiales: Dasyaceae, Rhodomelaceae. Nova Hedwigia, 6, 401–481.
Dawson, E. Y. y Beaudette, P. T. (1959). Field notes from the 1959 Eastern Pacific Cruise of the Stella Polaris. Pacific Naturalist, 1, 1–24.
Díaz, A. y Ortlieb, L. (1993). El fenómeno “El Niño” y los moluscos de la costa peruana. Bulletin de l’Institut Français d’Études Andines, 22, 159–177.
Dobretsov, S. V. (1999). Effects of macroalgae and biofilm on settlement of blue mussel (Mytilus edulis l.) larvae, Bio- fouling, 14, 153–165. https://doi.org/10.1080/089270199093 78406
Esqueda-González, M., Ríos-Jara, E., Galván-Villa, C. M. y Rodríguez-Zaragoza, F. A. (2014). Species composition, richness, and distribution of marine bivalve molluscs in Bahía de Mazatlán, México. Zookeys, 399, 43–69. https://doi.org/10.3897/zookeys.399.6256
Feely, R. A., Sabine, C. L., Lee, K., Berelson, W., Kleypas, J., Fabry, V. J. et al. (2004). Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science, 305, 362–366. https://doi.org/10.1126/science.1097329
Fitch, J. E. (1953). Common marine bivalves of California. Sacramento, California: California Department of Fish and Game.
Flores, P. (2004). Estructura de la comunidad de moluscos del mesolitoral superior en las playas de facie rocosa del estado de Guerrero, México (Tesis doctoral). Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León. San Nicolás de los Garza.
Flores-Garza, R., Flores-Rodríguez, P., García-Ibáñez, S. y Valdés-González, A. (2007). Demografía del caracol Plicopurpura pansa (Neotaenioglossa: Muricidae) y constitución de la comunidad malacológica asociada en Guerrero, México. Revista de Biología Tropical, 55, 867–878.
Flores-Garza, R., García-Ibáñez, S., Flores-Rodríguez, P., Torreblanca-Ramírez, C., Galeana-Rebolledo, L., Valdés-González, A. et al. (2012). Commercially important marine mollusks for human consumption in Acapulco, México. Natural Resources, 3, 11–17. http://dx.doi.org/10.4236/nr.20 12.31003
Flores-Garza, R., López-Rojas, V., Flores-Rodríguez, P. y Ramírez, C. T. (2014). Diversity, distribution, and composition of the Bivalvia class on the rocky intertidal zone of marine priority region 32, Mexico. Open Journal of Ecology, 4, 961–973 https://doi.org/10.4236/oje.2014.415080
Flores-Garza, R., Torreblanca-Ramírez, C., Flores-Rodríguez, P., García-Ibáñez, S. y Galeana-Rebolledo, L. (2010). Riqueza y análisis de la comunidad malacológica en el mesolitoral rocoso de la playa Tlacopanocha, Acapulco, Guerrero. En L. J. Rangel, J. Gamboa, S. L. Arriaga y W. M. Contreras (Eds.), Perspectiva en malacología mexicana (pp. 125–138). Villahermosa: Universidad Juárez Autónoma de Tabasco.
Flores-Garza, R., Torreblanca-Ramírez, C., Flores-Rodríguez, P., García-Ibáñez, S., Galeana-Rebolledo, L., Valdés-González, A. et al. (2011). Mollusc community from a rocky intertidal zone in Acapulco, Mexico. Biodiversity, 12, 144–153. https://doi.org/10.1080/14888386.2011.625520
Flores-Rodríguez, P., Flores-Garza, R., García-Ibáñez, S. y Valdés-González, A. (2003). Riqueza y diversidad de la malacofauna del mesolitoral rocoso de la Isla la Roqueta, Acapulco, Guerrero, México. Ciencia, Revista de Investigación Científica, 11, 5–14.
Flores-Rodríguez, P., Flores-Garza, R., García-Ibáñez, S. y Valdés-González, A. (2007). Variación en la diversidad malacológica del mesolitoral rocoso en Playa Troncones, La Unión, Guerrero, México. Revista Mexicana de Biodiversidad, 78, 33–40. https://doi.org/10.22201/ib.20078706e.2007.002.298
Flores-Rodríguez, P., Flores-Garza, R., García-Ibáñez, S., Valdés-González, A., Violante-González, J., Santiago, C. E. et al. (2012). Mollusk species richness on the rocky shores of the state of Guerrero, Mexico, as affected by rains and their geographical distribution. Natural Resources, 3, 248–260. http://dx.doi.org/10.4236/nr.2012.34032
Forster, S. y Zettler, M. L. (2004). The capacity of the filter-feeding bivalve Mya arenaria L. to affect water transport in sandy beds. Marine Biology, 144, 1183–1189. https://doi.org/10.1007/s00227-003-1278-2
Galeana-Rebolledo, L., Flores-Garza, R., Torreblanca-Ramírez, C., García-Ibáñez, S., Flores-Rodríguez, P. y López-Rojas, V. I. (2012). Biocenosis de Bivalvia y Polyplacophora del intermareal rocoso en playa Tlacopanocha, Acapulco, Guerrero, México. Latin American Journal of Aquatic Research, 40, 943–954. http://dx.doi.org/10.3856/vol40- issue4-fulltext-11
Galeana-Rebolledo, L., Flores-Garza, R., Violante-González, J., Flores-Rodríguez, P., García-Ibáñez, S., Landa-Jaime, V. et al. (2018). Socioeconomic aspects for coastal mollusk commercial fishing in Costa Chica, Guerrero, México. Natural Resources, 9, 229–241. https://doi.org/10.4236/nr. 2018.96015
García-Domínguez, F. A., Haro-Hernández, D., García-Cuellar, Á., Villalejo-Fuerte, M. y Rodríguez-Astudillo, S. (2008). Ciclo reproductivo de Anadara tuberculosa (Sowerby, 1833) (Arcidae) en Bahía Magdalena, México. Revista de Biología Marina y Oceanografía, 43, 143–152. http://dx.doi.org/10.4067/S0718-19572008000100015.
Gama, N. (2019). Aspectos ecológicos de la comunidad de gasterópodos (Mollusca: Gastropoda) asociados a ensambles macroalgales en el intermareal rocoso de Ixtapa Zihuatanejo, Guerrero, México (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Gama-Kwick, N., Aguilar-Estrada, L. G., Quiroz-González, N. y Ruiz-Boijseuneau, I. (2021). Nuevos registros de gasterópodos (Mollusca) asociados a macroalgas inter- mareales de Guerrero, México. Revista Mexicana de Biodiversidad, 92, 1–18. http://dx.doi.org/10.22201/ib.200 78706e.2021.92.3441
Garcés, J. L. (2011). Micromoluscos bivalvos de la Bahía de Acapulco Guerrero, México: composición específica y diversidad (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
García, M. (2009). Estructura comunitaria de la fauna asociada a algas submareales en tres sitios contrastantes en Zihuatanejo, Guerrero (Tesis de maestría). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
García-Cubas, A. (1981). Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de Términos, Campeche). Publicaciones especiales-Instituto de Ciencias del Mar y Limnología, 5, 1–379.
García-Cubas, A. y Reguero, M. (2007). Catálogo ilustrado de moluscos bivalvos del Golfo de México y Mar Caribe. Ciudad de México: Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México.
García-Ibáñez, S., Flores-Rodríguez, P., Navarro, J. T. N., Garza, R. F. y Moreno, I. E. B. (2014). Respuesta del carnívoro Plicopurpura pansa (Mollusca: Gastropoda) y el herbívoro Chiton articulatus (Mollusca: Polyplacophora) a factores ambientales en Acapulco, México. CienciaUAT, 8, 11–21.
García-Robledo, E., Corzo, A., van Bergeijk, S. y Yúfera, M. (2008). Impacto de las acumulaciones de macroalgas en la comunidad biológica intermareal. Revista de la Sociedad Gaditana de Historia Natural, 8, 115–137.
Gibbons, M. J. (1988). The impact of sediment accumulation, relative habitat complexity and elevation on rocky shore meiofauna. Journal of Experimental Marine Biology Ecology, 122, 225–241. https://doi.org/10.1016/0022-0981(88)90125-6
Gosling, E. (2015). Marine bivalve molluscs. Chichester, West Sussex: John Wiley y Sons.
Guiry, M. D. y Guiry, G. M. (2024). AlgaeBase. National University of Ireland, Galway. Recuperado el 12 enero, 2024 de: http://www.algaebase.org
Guzmán, A. R. (2022). Estructura comunitaria de bivalvos asociados a macroalgas en el litoral rocoso de Muelle Municipal, Zihuatanejo, Guerrero, México (Tesis). Facultad de Ciencias. Universidad Nacional Autónoma de México. Ciudad de México
Hernández-Moreno, E. P., Romo-Piñera, A. K., Fernández-Rivera Melo, F. J., Aguilar-Cruz, C. A., Reyes-Bonilla, H. y López-Vivas, J. M. (2020). Reproductive Study of Megapitaria aurantiaca (Sowerby, 1831) (Bivalvia: Veneridae) in Puerto Libertad, Sonora, Mexico. Journal of Shellfish Research, 39, 441–447.
Holguín-Quiñones, O. E. y González-Pedraza, A. C. (1989). Moluscos de la franja costera del estado de Oaxaca, México. México D.F.: Dirección de Bibliotecas y Publicaciones, Instituto Politécnico Nacional.
Horton, T., Kroh, A., Ahyong, S., Bailly, N., Boyko, C. B. y Brandão, S. N. (2024. World Register of Marine Species. Recuperado el 14 de marzo, 2024: http://www.marinespecies.org
Huovinen, P., Gómez, I. y Lovengreen, C. (2006). A five-year study of solar ultraviolet radiation in Southern Chile (39° S): Potential impact on physiology of coastal marine algae? Photochemistry and Photobiology, 82, 515–522. https://doi.org/10.1562/2005-07-05-RA-601
IMTA (Instituto Mexicano de Tecnología del Agua). (2010). Estudio de clasificación de la Bahía de Ixtapa-Zihuata- nejo. No. FON-CNA-2004-02-016. Informe final. Comisión Nacional del Agua.
Jiménez-Valverde, A. y Hortal, J. (2003). Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8, 151–161.
Jorgensen, C. B. (1996). Bivalve filter feeding revisited. Marine Ecology Progress Series, 142, 287–302.
Jover-Capote, A. y Diez, Y. L. (2017). Abundancia de moluscos en mantos de macroalgas del mesolitoral rocoso en la costa suroriental de Cuba. Amici Molluscarum, 25, 27–43.
Keen, A. M. (1971). Sea shells of Tropical West America Marine mollusks from Baja California to Peru. Standford, California: Stanford University Press.
Kuk-Dzul, J. G., Padilla, J. G., Torreblanca, C., Flores, R., Flores, P. y Muñiz, X. I. (2019). Structure of molluscan communities in shallow subtidal rocky bottoms of Acapulco, Mexico. Turkish Journal of Zoology, 43, 465–479.
Landa-Jaime, V. y Arciniega-Flores, J. (1998). Macromoluscos bentónicos de fondos blandos de la plataforma continental de Jalisco y Colima, México. Ciencias Marinas, 24, 155–l67.
Landa-Jaime, V., Michel-Morfín, E., Arciniega-Flores, J., Castillo-Vargasmachuca, S. y Saucedo-Lozano, M. (2013). Moluscos asociados al arrecife coralino de Tenacatita, Jalisco, en el Pacífico central mexicano. Revista Mexicana de Biodiversidad, 84, 1121–1136. https://doi.org/10.7550/rmb.32994
Lee, R. E. (2008). Basic characteristics of the algae. En R. E. Lee (Ed.), Phycology (pp. 3-29). Nueva York: Cambridge University Press.
Lesser, H. (1984). Prospección sistemática y ecológica de los moluscos bentónicos de la plataforma continental del estado de Guerrero, México (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Levene, H. (1960) Robust tests for equality of variances. En I. Olkin (Ed.), Contributions to probability and statistics (278–292). Palo Alto, California: Standford University Press.
López, N. A., (1993). Caracterización de la ficoflora sublitoral de Acapulco y Zihuatanejo, Gro. (Tesis). Facultad de Cien- cias, Universidad Nacional Autónoma de México. Ciudad de México.
López, N., Candelaria, C. y Ramírez-García, P. (2023). Assessment of macroalgae coverage in a scarcely studied deep rocky reef in the tropical eastern Mexican Pacific. Latin American Journal of Aquatic Research, 51, 23–33. https://doi.org/10.3856/vol51-issue1-fulltext-2920
López, N., Candelaria, C., Ramírez-García, P. y Rodríguez, D. (2017). The structure of tropical turf-forming algae assemblages. Zihuatanejo Bay, México. Latin American Journal of Aquatic Research, 45, 329–340 http://dx.doi.org/10.3856/vol45-issue2-fulltext-9
López-Rojas, V. I., Flores-Garza, R., Flores-Rodríguez, P., Torreblanca-Ramírez, C. y García-Ibáñez, S. (2017). La clase Bivalvia en sitios rocosos de las Regiones Marinas Prioritarias en Guerrero, México: riqueza de especies, abundancia y distribución. Hidrobiológica, 27, 69–86.
Lozada, O. (2010). Actualización sistemática de los bivalvos de la Colección Malacológica Dr. Antonio García-Cubas del Instituto de Ciencias del Mar y Limnología (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Lozada, O. (2015). Presencia de metales pesados en Isogno- mon alatus (Gmelin, 1791) de la laguna Tampamachoco, Veracruz (Tesis de maestría). Facultad de Ciencias Bio- lógicas y Agropecuarias, Universidad Veracruzana. Tuxpan, Veracruz.
Magurran, A. E. (2004). Measuring biological diversity. Malden, Massachusetts: Blackwell Publishing.
Morales, R., Vélez, H., Mejía, A. Ramírez, I., Izurierta, J. y Saldaña, P. (2008). Hidrodinámica de la Bahía de Zihuata- nejo. XXIII Congreso Latinoamericano de Hidráulica, 2 al 6 de septiembre de 2008, Cartagena de Indias, Colombia.
Moreno, C. A. (1995). Macroalgae as a refuge from predation for recruits of the mussel Choromytilus chorus (Molina, 1782) in southern Chile. Journal of Experimental Marine Biology and Ecology, 191, 181–193.
Moreno, C. E. (2001). Métodos para medir la biodiversidad. Zaragoza, España: M&T–Manuales y Tesis SEA.
Oksanen, J., Guillaume-Blanchet, F., Friendly, M., Kindt, R. Legendre, P., McGlinn, D. et al. (2019). Vegan: community ecology package. R package versión 2.5-6. https://CRAN.R-project.org/package=vegan
Olabarria, C. y Chapman, M. G. (2001). Comparison of patterns of spatial variation of microgastropods between two contrasting intertidal habitats. Marine Ecology Progress Series, 220, 201–211. https://doi.org/doi:10.3354/meps220201
Olafsson, E. B. (1986). Density dependence in suspension-feeding and deposit-feeding populations of the bivalve Macoma balthica: a field experiment. Journal of Animal Ecology, 55, 517–526. https://doi.org/10.2307/4735
Oyarzún, P. A., Toro, J. E., Jaramillo, R., Guiñez, R., Briones, C. y Astorga, M. (2011). Ciclo gonadal del chorito Mytilus chilensis (Bivalvia: Mytilidae) en dos localidades del sur de Chile. Latin American Journal of Aquatic Research, 39, 512–525.
Paredes, C., Tarazona, J., Canahuire, E., Romero, L., Cornejo, O. y Cardoso, F. (1998). Presencia de moluscos tropicales de la provincia panameña en la costa central del Perú y su relación con los eventos “El Niño”. Revista Peruana de Biología, 5, 123–128. https://doi.org/10.15381/rpb.v5i2.8330
Pérez, L. M. (2013). Descripción espacio-temporal de la temperatura superficial del mar en el Pacífico Sur Mexicano de 1996-2009 (Tesis). Universidad del Mar. Puerto Ángel, Oaxaca.
Prathep, A., Marrs, R. H. y Norton, T. A. (2003). Spatial and temporal variations in sediment accumulation in an algal turf and their impact on associated fauna. Marine Biology, 142, 381–390. https://doi.org/10.1007/s00227–002–0940–4
Quirós-Rodríguez, J. y Campos, N. H. (2013). Moluscos asociados a ensamblajes macroalgales en el litoral rocoso de Córdoba, Caribe colombiano. Boletín de Investigaciones Marinas y Costeras, 42, 101–120.
Quiroz-González, N., Aguilar-Estrada, L. G., Ruiz-Boijseauneau, I. y Rodríguez, D. (2020). Biodiversidad de algas epizoicas en el Pacífico tropical mexicano. Acta Botanica Mexicana, 127, 1-22. https://doi.org/10.21829/abm127.2020.1645
R Core Team (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reguero, M., y García-Cubas, A. (1989). Moluscos de la plataforma continental de Nayarit: sistemática y ecología (cuatro campañas oceanográficas). Anales del Instituto de Ciencias del Mar y Limnología, 16, 33–58.
Renaud, P. E., Riggs, S. R., Ambrose Jr, W. G., Schmid, K. y Snyder, S. W. (1997). Biological-geological interactions: storm effects on macroalgal communities mediated by sediment characteristics and distribution. Continental Shelf Research, 17, 37–56. https://doi.org/10.1016/0278-43 43(96)00019-2
Ríos-Jara, E., López-Uriarte, E., Pérez-Peña, M. y Juárez-Carrillo, E. (2003). Nuevos registros de escafópodos para las costas de Jalisco y Colima, México. Hidrobiológica, 13, 167–170.
Rodríguez, D., López, N. y González-González, J. (2008). Gelidiales (Rhodophyta) en las costas del Pacífico mexicano con énfasis en las especies tropicales. En A. Sentíes y K. M. Dreckmann (Eds.), Monografías ficológicas, Vol. 3 (pp. 27–74). Ciudad de México: Universidad Autónoma Metropolitana.
Román-Contreras, R., Cruz-Abrego, F. M. e Ibáñez-Aguirre, A. L. (1991). Observaciones ecológicas de los moluscos de la zona intermareal rocosa de la Bahía de Chamela, Jalisco, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 62, 17–31.
Rosenberg, R. (1977). Effects of dredging operations on estuarine benthic macrofauna. Marine Ecology Progress Series, 62, 185–202.
Salazar-Vallejo, S. I. y González, E. (1990). Ecología costera en la región de La Mancha, Veracruz. La Ciencia y el Hombre, 6,101–120.
Salcedo-Martínez, S., Green, G., Gamboa-Contreras, A. y Gómez, P. (1988). Inventario de macroalgas y macroinvertebrados bénticos, presentes en áreas rocosas de la región de Zihuatanejo, Guerrero, México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 15, 73–95.
Sánchez, M. (2014). Base de datos de los moluscos presentes en la colección biológica de la Secretaría de Marina-Armada de México (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Santos, L., Souza, J., Lima, S. y Guimarães, C. (2020). Diversity of bivalve mollusks associated with macroalgae on the continental shelf in the states of Alagoas, Sergipe and Bahia, Northeastern Brazil. Zoological Studies, 59, e58. https://doi.org/10.6620/ZS.2020.59-58
Schulien, J. A., Adams, J. y Felis, J. (2020). Pacific continental shelf environmental assessment (PaCSEA): characterization of seasonal water masses within the Northern California current system using airborne remote sensing off Northern California, Oregon, and Washington, 2011–2012. Camarillo, California: US Department of the Interior, Bureau of Ocean Energy Management/ Pacific OCS Region/ OCS Study BOEM.
Seed, R. (1969a). The ecology of Mytilus edulis L. (Lamellibranchiata) on exposed rocky shores. Oecologia, 3, 277–316. https://doi.org/10.1007/BF00390380
Seed, R. (1969b). The ecology of Mytilus edulis L. (Lamelli- branchiata) on exposed rocky shores: II. Growth and mortality. Oecologia, 3, 317–350. https://www.jstor.org/stable /4214550
Shaw, W. N., Hassler, T. J. y Moran, D. P. (1988). California sea mussel and bay mussel: species profiles. Life histories and environmental requirements of coastal fishes and invertebrates (Pacific Southwest). Washington D.C.: Fish and Wildlife Service.
Sibaja-Cordero, J. A. y Vargas-Zamora, J. A. (2006). Zonación vertical de epifauna y algas en litorales rocosos del Golfo de Nicoya, Costa Rica. Revista de Biología Tropical, 54, 49–67.
Siegel, S. (1990). Estadística no paramétrica: aplicada a las ciencias de la conducta. Ciudad de México: Trillas.
Stella, C., Vijayalakshmi, S. y Siva, J. (2010). Two new records of bivalve species of Mytilidae family from Palk Bay and Mandapam area-south-east coast of India. Global Journal of Environmental Research, 4, 40–42.
Steneck, R. S. y Dethier, M. N. (1994). A functional group approach to the structure of algal-dominated communities. Oikos, 69, 476–498. https://doi.org/10.2307/3545860
Steneck, R. S. y Watling, L. (1982). Feeding capabilities and limitation of herbivorous molluscs: a functional group approach. Marine Biology, 68, 299–319. https://doi.org/10. 1007/BF00409596
Suchanek, T. H. (1978). The ecology of Mytilus edulis L. in exposed rocky intertidal communities. Journal of Experimental MarineBiology and Ecology, 31, 105–120. https://doi.org/10.1016/0022-0981(78)90139-9
Taylor, W. R. (1945). Pacific marine algae of the Allan Hancock Expeditions to the Galapagos Islands. Allan Hancock Pacific Expeditions, 12, 1–528.
Torreblanca, C. (2010). Análisis de la diversidad y estructura de la comunidad de moluscos del mesolitoral rocoso de Acapulco, Guerrero (Tesis).Unidad Académica de Ecología Marina, Universidad Autónoma de Guerrero. Acapulco de Juárez.
Torreblanca-Ramírez, C., Flores-Garza, R., Flores-Rodríguez, P., García-Ibáñez, S. y Galeana-Rebolledo, L. (2012). Riqueza, composición y diversidad de la comunidad de moluscos asociada al sustrato rocoso intermareal de playa Parque de la Reina, Acapulco, México. Revista de Biología Marina y Oceanografía, 47, 283–294. http://dx.doi.org/10.4067/S0718-19572012000200010
Torroglosa, M. E. (2015). Biología reproductiva y crecimiento de Brachidontes rodriguezii (d´Orbign 1846) en sustratos duros artificiales en playas arenosas de la provincia de Buenos Aires (Tesis doctoral). Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Argentina.
Trasviña, A. y Andrade, C. A. (2002). La circulación costera del Pacifico Tropical Oriental, con énfasis en la Alberca Cálida Mexicana (ACM). En Corcas (Eds.), Circulación oceánica y climatología tropical en México y Colombia (pp. 9–37). Bogotá: Consejo Nacional de Acreditación-Ministerio de Educación.
UNAM (Universidad Nacional Autónoma de México) (2013). Laguna Las Salinas Zihuatanejo. Proyecto ejecutivo UNAM 1989-1994 basado en plano base FONATUR-SCT. Universidad Nacional Autónoma de México, México. Recuperado el 14 enero, 2024 de https://mmacreactive.wordpress.com/
Urbano, B. (2004). Estructura comunitaria de gasterópodos de Zihuatanejo, Guerrero (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Valdés-González, A., Flores-Rodríguez, P., Flores-Garza, R. y García-Ibáñez, S. (2004). Molluscan communities of the rocky intertidal zone at two sites with different wave action on Isla La Roqueta, Acapulco, Guerrero, Mexico. Journal of Shellfish Research, 23, 875–880.
Vassallo, A., Dávila, Y., Luviano, N., Deneb-Amozurrutia, S., Vital, X. G., Conejeros, C. A. et al. (2014). Inventario de invertebrados de la zona rocosa intermareal de Montepío, Veracruz, México. Revista Mexicana de Biodiversidad, 85, 349–362. https://doi.org/10.7550/rmb.42628
Vázquez, P. (2009). Efecto del sedimento retenido en la estructura de los ensambles algales submareales (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Vega, C., Olabarria, C. y Carballo, J. L. (2008). Variación espacio-temporal de moluscos y macroalgas en sustratos rocosos intermareales en la bahía de Mazatlán. Ciencia y Mar, 34, 3–16.
Velez, J. y Zeballos, J. (1985). Ampliación de la distribución de algunos peces e invertebrados durante el fenómeno “El Niño” 1982–1983. En W. Arntz, A. Landa y J. Tarazona (Eds.), “El Niño” su impacto en la fauna marina (pp. 173– 180). Callao, Perú: Boletín del Instituto del Mar del Perú.
Villalpando, E. (1986). Diversidad y zonación de moluscos de facie rocosa Isla Roqueta, Acapulco, Gro. (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad de México.
Villegas-Maldonado, S., Neri-García, E., Flores-Garza, R., García-Ibáñez, S., Flores Rodríguez, P. y Arana-Salvador, D. G. (2007). Datos preliminares de la diversidad de moluscos para el consumo humano que se expenden en Acapulco, Guerrero. En E. Ríos-Jara, M. C. Esqueda-González y C. M Galván-Villa (Eds.), Estudios sobre la malacología y conquiliología en México (pp. 57-59). Guadalajara: Universidad de Guadalajara.
Ward, J. E., Sanford, L. P., Newell, R. I. E. y MacDonald, B. A. (1998). A new explanation of particle capture in suspension-feeding bivalve molluscs. Limnology and Oceanography, 43, 741–752.
Wyrtki, K. 1966. Oceanography of the eastern equatorial Pacific Ocean. Oceanography and marine biology: an annual review, 4, 33–68.
Yang, J. L., Satuito, C. G., Bao, W. Y. y Kitamura, H. (2007). Larval settlement and metamorphosis of the mussel Mytilus galloprovincialis on different macroalgae. Marine Biology, 152, 1121–1132. https://doi.org/10.1007/s00227-007-0759-0
Zamorano, P. y Leyte-Morales, G. E. (2009). Equinodermos asociados a formaciones arrecifales en Zihuatanejo y Acapulco, Guerrero, México. Boletín de Investigaciones Marinas y Costeras-INVEMAR, 38, 7–28.
Angélica Rodríguez-Cifuentes a, Jovana M. Jasso-Martínez a, Valeria B. Salinas-Ramos a, Juan José Martínez b, Alejandro Zaldívar-Riverón a, *
a Universidad Nacional Autónoma de México, Instituto de Biología, Colección Nacional de Insectos, Tercer Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
b Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de La Pampa, Uruguay 151, L6300LB, Santa Rosa, La Pampa, Argentina
Received: 17 November 2023; accepted: 15 March 2024
Abstract
Galls are an abnormal growth of plant tissue in response to the presence generally of an inducing insect, which ensures food and protection during specific periods of its life. Besides gall formers, a vast community of arthropods are associated with galls, including inquilines and parasitoids. Few studies have assessed the gall diversity and its associated insect community in Neotropical vascular plants. Here, we characterised the leaf gall diversity of Coccoloba barbadensis Jacq. (Polygonaceae) in a Mexican tropical dry forest, as well as their associated entomofauna based on morphology and DNA barcoding. Five different gall morphotypes were observed during both dry (April-June) and rainy (November) seasons. A total of 34 and 38 species of Diptera, Hymenoptera, Coleoptera, and Lepidoptera were delimited with the 2% divergence criterion and the GMYC model, respectively. Based on our rearing observations and literature, Cecidomyiidae (Diptera) species might induce all leaf gall morphotypes, whereas hymenopterans are represented by parasitoid and probably inquiline species of the families Braconidae, Eulophidae, Eupelmidae, Platygastridae and Torymidae. Our results highlight the importance of performing integrative species delineation studies of arthropods present in galls to have an accurate knowledge of their diversity and trophic interactions.
Keywords: Trophic interactions; Host; Parasitoid; DNA barcode; Gall former
Diversidad de agallas de hojas en la planta neotropical Coccoloba barbadensis (Polygonaceae) y riqueza de especies de sus insectos asociados
Resumen
Las agallas son un crecimiento anormal de tejido de plantas por la presencia de un insecto inductor que le asegura alimento y protección durante periodos específicos. Además de los formadores de agallas, una vasta comunidad de artrópodos está también asociada, incluidos inquilinos y parasitoides. Pocos estudios han evaluado la diversidad de agallas y su comunidad de insectos en plantas vasculares neotropicales. Aquí se caracteriza la diversidad de agallas foliares de Coccoloba barbadensis Jacq. (Polygonaceae) en un bosque seco tropical mexicano, así como su entomofauna asociada basada en morfología y el código de barras del DNA. Se observaron 5 morfotipos de agallas durante las temporadas seca (abril-junio) y lluviosa (noviembre). Se delimitó un total de 34 y 38 especies de Diptera, Hymenoptera, Coleoptera y Lepidoptera con el criterio de divergencia de 2% y el modelo GMYC, respectivamente. Según las observaciones y datos de literatura, especies de Cecidomyiidae (Diptera) inducen todos los morfotipos de agallas, y los himenópteros están representados por especies parasitoides y probablemente inquilinas de las familias Braconidae, Eulophidae, Eupelmidae, Platygastridae y Torymidae. Los resultados resaltan la importancia de estudios integradores para la delimitación de especies de artrópodos de agallas para tener conocimiento preciso de su diversidad e interacciones tróficas.
Palabras clave: Interacciones tróficas; Hospedero; Parasitoide; Código de barras del DNA; Formador de agallas
Introduction
Ecological interactions among species form the basis of ecosystem functioning and underlie evolutionary and ecological principles of conservation biology (Clare et al., 2013). Three types of biological networks have been defined based on interactions and the types of organisms involved (Ings et al., 2009): traditional or antagonistic food webs (e.g., predators and prey/food webs), mutualistic networks (e.g., seed dispersal and pollination), and host-parasitoid networks. The study of these biological networks provides a whole ecosystem outline to examine the effects of biodiversity loss on communities and ecosystem functions (Ings et al., 2009).
Understanding the links of a network relies on the idea that descriptors of interaction structure are unbiased and accurate (Wirta et al., 2014). In practice, however, networks are difficult to create, especially using traditional methods. Taxonomic resolution and the methodology employed to delimit species are crucial to reconstruct interaction structure (Paine, 1980; Kaartinen & Roslin, 2011). If the links are poorly resolved and multiple taxa are inadvertently grouped within the nodes of a web, there is a risk of misunderstanding its composition and thus its functioning system (Kaartinen & Roslin, 2011; Wirta et al., 2014).
An astonishing number of arthropod taxa depend on plants as food resources or closely interact with them. Among these are species that form enclosed structures known as galls (Mani, 1964; Raman, 2011). These structures are defined as abnormal growth of tissues of host plants in response to the activity or presence of an inducing organism (Nieves-Aldrey, 1998; Price, 2005; Redfern et al., 2002). Galls can be found in several plant structures such as flowers, roots, fruits, leaves, thorns, or stems. Arthropods induce galls to ensure food resources and to protect themselves against predators or unfavourable environmental conditions during certain periods of their life cycle (Nieves-Aldrey, 1998; Raman & Withers, 2003).
Gall induction in insects mainly occurs in species of Hymenoptera, but also in species of Diptera, Hemiptera and Thysanoptera, and less frequently in Coleoptera and Lepidoptera (Raman, 2011). Besides gall formers, there is an intricate community of different insect species that also are associated with galls, including inquilines (i.e., species that develop within galls made by other insects and feed on plant tissue), parasitoids of gall formers and inquilines and hyperparasitoids (i.e., parasitoids of other parasitoid species) (Forbes et al., 2015). Despite the great ecological importance of galls in most terrestrial ecosystems due to the extraordinary arthropod diversity that they comprise, to date, most of this species diversity and the interactions that are involved are largely unknown, especially in tropical and subtropical regions.
In recent years, molecular techniques have provided detailed analyses of interaction reconstruction, allowing precise identification of members of natural communities and the structure of networks (Clare et al., 2013; Kaartinen et al., 2010; Wirta et al., 2014). The DNA barcoding locus, a fragment of the cytochrome oxidase I (COI) mitochondrial DNA gene, is the most employed genetic marker for species discrimination of closely related animal species (Hebert et al., 2003; Ratnasingham & Hebert, 2013). This marker is a valuable tool for the rapid identification of megadiverse, poorly known taxa (Hebert et al., 2003). Moreover, it allows the association of morphologically distinct semaphoronts (e.g., insect larvae and adults; Yeo et al., 2018) and sexes of the same species (e.g., Sheffield et al., 2009).
Coccoloba barbadensis is a widely distributed Neotropical vascular plant species that in Mexico occurs in tropical regions from central to southeast Mexico (Howard, 1959). In this study, the diversity of galls on the leaves of the vascular plant C. barbadensis Jacq. (Polygonaceae) in a Mexican tropical dry forest was characterised and their associated entomofauna assessed using both morphological and DNA barcoding data. We highlight the necessity to perform integrative species delimitation studies of arthropods present in galls, particularly in the tropics, to have a more accurate knowledge of their species richness.
Materials and methods
The study was conducted in the Biological Station of Chamela (EBCH), Jalisco, Mexico (19’29” N, 105’01” W; Noguera et al., 2002), owned by the Instituto de Biología, Universidad Nacional Autónoma de México. The Chamela region is mainly composed of tropical dry forest and is characterised by having 85% of the ~ 750 mm of yearly rain from July to November and a mean annual temperature of 24.9 °C (14.8-32 °C) (Méndez-Alonzo et al., 2013). Tropical dry forests frequently show extreme changes in the physiognomy and available resources during the rainy and dry seasons, therefore altering the composition and diversity of their fauna (Razo-González et al., 2014).
We carried out 2 collecting trips at the EBCH, one during the dry (from March to June 2013) and the other during the rainy season (November 2013). Fifteen trees belonging to C. barbadensis were located and marked, all of which were situated near seasonal streams. We collected 1-5 leaves with galls from different parts of the selected trees. By in situ photographs we documented the presence/absence of galls for each tree, as well as general leaf features such as colour and size. The main gall’s features, including shape, size, colour, and pubescence, and the number of all collected galls present on the leaves were classified by morphotypes and recorded.
The levels of infestation, presence, and type of galls were weekly recorded, and the total number of galls per leaf was recorded. Galls were subsequently dissected or maintained in the laboratory to rear their insects. All reared insects were preserved in 96% ethanol and stored at -20 °C.
All collected insects were sorted out into adults, larvae, or pupae, and were counted and discriminated into morphospecies with a Zeiss™ Stemi DV4 (Göttingen, Germany) stereomicroscope. Larvae and adults of Hymenoptera were identified to order and genus level, respectively, using the specialised literature. Larvae and adults of Diptera and larvae of Coleoptera and Lepidoptera could only be identified to family and order level, respectively.
Gall abundance
The relationship between both the total number of collected galls and the average number of each gall morphotype concerning the time of collection during the dry season was evaluated using a simple linear regression with the statistical program R (R Core Team, 2013). Data of rainy season were not statistically analysed, since there were not enough samples to perform statistical tests. The insect family frequency throughout the sampled period was also analysed and then a canonical correspondence analysis to characterise the association between insect species and gall morphotypes with the program Statistica version 10 (StatSoft, Inc., 2011).
Molecular data
It is widely recognised that species misidentifications have negative consequences in ecological studies (Bortolus, 2008; Vink et al., 2012). Species delimitation and identification can be considerably improved using morphology-based taxonomy coupled with DNA sequence data (Dexter et al., 2010). Some representative specimens of most identified morphospecies were molecularly characterised generating for them DNA sequences of a fragment belonging to the barcoding locus COI mitochondrial gene (Hebert et al., 2003). This gene marker has been proved to be a generally reliable tool for the rapid delimitation of animal species, including insects (Hebert et al., 2003, 2004).
The genomic DNA from 1-6 specimens belonging to each discriminated insect morphotype was extracted. DNA extractions were conducted with the kit Tissue and tissue plus SV mini (Gene All®, Seoul, Korea), by placing each individual in 20 μl of proteinase K and 200 μl of TL Buffer, at 56 °C for 8 h. The larvae bodies were completely degraded after digestion, whereas the pupae exoskeletons and the adult individuals were subsequently washed with distilled water, placed back in 96% ethanol, and stored at -20 °C until they were mounted and labelled.
The COI fragment was amplified using the LCO1460/HCO2198 primers (Folmer et al., 1994). The following PCR conditions were used: initial denaturation at 95 ºC for 3 min, followed by 30 cycles of denaturation at 94 °C for 40 s, 40 s annealing at 45 °C, 40 s extension at 72 °C, and a final extension at 72 °C for 7 min. PCRs were prepared in a final volume of 15 μl of reaction mixture, which included 1.5 μl of 10X buffer, 0.75 μl of MgCl 2 (50 mM), 0.3 μl of dNTPs (10 mM), 0.24 μl of each primer (10 μM), 0.12 μl of Taq Platinum polymerase (Invitrogen®, Carlsbad, CA, USA), 8.85 μl of water and 3 μl of DNA template.
Unpurified PCR products were sent for DNA sequencing to the High-Throughput Genomics Unit of the University of Washington, Seattle, USA (http://www.htseq.org/). Sequences were edited and aligned manually based on their translated amino acids and compared individually with the sequences available in GenBank (http://www.ncbi.nlm.nih.gov/genbank) using the BLAST online program (Altschul et al., 1990; http://blast.ncbi.nlm.nih.gov/Blast.cgi).
DNA sequence-based species delimitation of dipterans and hymenopterans was carried out separately with the barcoding locus using 2 approaches, the General Mixed Yule Coalescence (GMYC) model (Pons et al., 2006) and the 2% genetic divergence criterion (Hebert et al., 2003).
The GMYC model requires an ultrametric tree, which was obtained with the program BEAST version 1.7.4 (Drummond et al., 2012), running the analysis for 10 million generations, sampling trees every 1,000 generations, using an uncorrelated lognormal clock and a coalescent tree prior. Only 1 partition was considered, which used the GTR+Γ+I evolutionary model. The duplicated haplotypes from the matrix were removed using the program Collapse 1.2 (Posada, 2004). The first 2,500 trees were eliminated as “burn-in” and the remaining trees were used to reconstruct a maximum clade credibility tree with the program TreeAnnotator version 1.7.4 (part of the BEAST 1.7.4 package). The GMYC model implemented in the SPLITS package (http://r-forge.r-project.org/projects/splits/) was performed with the R program version 2.10.1 (R core Team, 2021).
For the 2% divergence criterion, the allocation of molecular taxonomic units (Hebert et al., 2003) was made depending on the percentage of similarity in the genetic distances of the analysed sequences. If the percentage was less than 2, those MOTUs were considered to belong to the same barcoding species. Uncorrected COI divergences were obtained with the program PAUP* version 4.0 (Swofford, 2003). A Neighbor-Joining (NJ) distance tree for Diptera and Hymenoptera was reconstructed separately with the above program to visualise the genetic distances obtained. The trees obtained from 2 species delineation approaches were visualised with the program Figtree version 1.4.4 (Bouckaert et al., 2014).
Results
Description and abundance of galls
A total of 11,044 and 1,127 galls were dissected from 480 and 50 leaves obtained from the 15 trees that were sampled during the dry and rainy seasons, respectively. Five gall morphotypes from the leaves of C. barbadensis were identified (Fig. 1A-F): 1) capsule-shaped (Fig. 1B), green on both sides, glabrous, with a central inner elongated canal; 2) conical (Fig. 1C), pale green on both sides, ending on a sharp tip on the beam, glabrous, with an internal round chamber; 3) flattened (Fig. 1D), greenish-yellow on the beam and brown on the underside, glabrous, with a horizontal centrally elongated chamber; 4) spherical (Fig. 1E), glabrous, flat, indistinct and brown on the beam, brown and spherical on the underside, distributed irregularly along the leaf; and 5) rounded (Fig. 1F), brown on both sides, with abundant whitish pubescence. All gall morphotypes were recorded in both seasons except gall morphotype 5, which was not recorded during the rainy season.
Figure 1. Leaf gall morphotypes found on Coccoloba barbadensis Jacq. (Polygonaceae). A) Leaves of C. barbadensis; B) capsule-shaped gall; C) conical gall; D) flattened gall; E) spherical gall; F) rounded gall.
A significant relationship (R2 = 0.731, F = 21.796, p < 0.001) between the average number of total galls collected during the sampled weeks was observed, with the presence of galls in the leaves gradually increasing throughout the weeks during the dry season. The number of galls was relatively constant during the first 4 weeks (March 25-29 to April 22-23), but from the fifth week (April 29-30) it gradually increased (Fig. 2A).
Figure 2. A) Simple linear regression showing the average number of collected leaf galls on C. barbadensis during the time of collect in the dry season; B) graphic showing the average number of each leaf gall morphotype during the time of collect in the dry season.
Most galls collected during the dry season corresponded to gall morphotypes 1 and 4 (35% and 32%, respectively) and were present in 87% and 80% of the examined trees, respectively. Cecidomyiid exuviates adhered to the latter 2 gall morphotypes during the first 2 sampling weeks (25-26th of March – 1-2nd of April) and the last week of February, respectively. On the other hand, 13%, 18.5%, and 1.5% of the remaining galls belonged to gall morphotypes 2, 3, and 5 and occurred in 73%, 80%, and 13% of the trees, respectively. During the rainy season, 17.5%, 26.4%, 30.7%, and 25.4% of the collected galls corresponded to morphotype galls 1 to 4, respectively.
The average number of each gall morphotype varied over time during the dry season. There were significant differences between each type of gall (F3,128 = 8.616, p < 0.001) (Fig. 2B). The frequency of gall morphotype 1 gradually increased during the first weeks, though by the last week of April they were no longer registered. In contrast, gall morphotype 2 showed a lower abundance but its number was relatively constant throughout the sampling period, whereas gall morphotypes 3 and 4 had a low frequency but their number increased towards the first week of May. Gall morphotype 5 was only observed during the last 2 sampling weeks.
Integrative taxonomy
A total of 2,008 larvae and 356 adult insects were obtained from the dissected galls during both seasons. The morphospecies including larvae, pupae, and adults were first discriminated, for which were subsequently generated 230 COI sequences (151 sequences of Hymenoptera, 77 of Diptera, 1 of Coleoptera, and 1 of Lepidoptera (GenBank accession numbers in Appendix) resulting in 125 haplotypes. The number of species per family delimited by the 2 DNA sequence-based species delimitation approaches is provided in Table 1. The 2% COI divergence criterion and the GMYC model discriminated between 34 and 38 species, respectively. The NJ distance tree derived from the examined COI sequences of Hymenoptera and Diptera is shown in figures 3 and 4, respectively. There were 2 inconsistencies between both approaches. The 2% COI divergence criterion delimited 1 species of Chrysonotomyia (Entedoninae: Eulophidae) and 2 of Teniupetiolus (Eurytominae: Eurytomidae), whereas the GMYC model divided them into 3 and 2 species, respectively.
DNA sequence data supported most of the delimited species using larvae and adults, except for the only species of Torymus Dalman (Hymenoptera: Torymidae), the 3 species of Cecidomyiidae (Diptera), and the single species of Lepidoptera and Coleoptera, for which we only generated sequences of larvae.
Insect-gall association
Gall morphotype 3 had the highest insect species richness (20 species), which comprised all sampled families except the hymenopteran species of Braconidae (Allorhogas coccolobae Martínez & Zaldívar-Riverón) and Torymidae (Torymus sp.). This gall morphotype also had the highest number of eulophid species (8), from which 5 belong to the subfamily Tetrastichinae and was only present in this gall morphotype. Gall morphotype 1 had 18 associated insect species, with Platygastridae and Eurytomidae (Hymenoptera) being the families with more species (5 and 4 species, respectively; Table 2). Gall morphotypes 2 and 4, on the other hand, registered 9 and 12 species, respectively, whereas gall morphotype 5 only had 4 species, 1 belonging to Eulohpidae (Hymenoptera), 1 to Platygastridae (Hymenoptera) and 2 to Cecidomyiidae (Diptera) (Table 2).
Larvae of Cecidomyiidae were highly abundant throughout the dry season. The abundance of larvae and adults of the families Eurytomidae and Braconidae, on the other hand, considerably increased towards the sixth week of the dry season. In contrast, the presence of immature stages of Eulophidae was more frequent at the beginning of the dry season, though adults were also observed throughout this sampling period. Platygastridae was the hymenopteran family that was most frequently found in both larval and adult stages. Only immature individuals of Cecidomyiidae (Diptera) and Eulophidae (Hymenoptera) were collected during the rainy season.
A statistically significant difference (c2 = 912,989, df = 102, p <0.001) was observed between the 4 morphotypes of galls recorded during the dry season and their associated insect species. Fourteen out of the 37 insect species delimited with 2% barcoding were associated with a single gall morphotype. These included a species of Tenuipetiolus (Eurytomidae) in gall morphotype 1, 5 of Tetrastichinae (Eulophidae) in gall morphotypes 3, 1 species of Chrysonotomyia (Eulophidae) in gall morphotypes 1, 2 and 3, respectively, the single species of Eupelmidae and Torymidae in gall morphotypes 1 and 4, respectively, 2 and 1 species of Platygastridae in gall morphotypes 1 and 3, respectively, and 1 species of Cecidomyiidae in gall morphotype 3. Five delimited species of Cecidomyiidae were recorded in gall morphotypes 3 and 4, whereas 4, 3, and 2 were present in gall morphotypes 2, 1, and 5, respectively.
Discussion
A considerable leaf gall diversity in C. barbadensis is recorded here. Based on the gathered information, the inducers of these 5 types of leaf galls were species of Cecidomyiidae (Diptera) (see below). Other cecidomyiid galls that have been reported on species of Coccoloba include those found on stems of C. mosenii Lindl., on leaves of Coccoloba cf. warmingii Meisn., C. diversifolia Jacq., C. swartzii Kuntze, and C. uvifera Meins., and on inflorescences of C. alnifolia Casar (Mead, 1970; Maia et al., 2008; Ramos-Rodrigues et al., 2014). The number of different types of galls found in C. barbadensis (5 morphotypes) is higher than those reported for the above species (≤ 2 morphotypes). However, based on their general appearance, the gall morphotypes 1 and 2 may be variants made by the same inducer species. Further rearing observations and molecular characterisation of gall inducers will help to confirm the actual gall diversity that is present on the leaves of this plant species.
Figure 3. Neighbour Joining phenogram reconstructed for specimens of Cecidomyiidae that were reared from the 4 leaf gall morphotypes found in C. barbadensis. Bars refer to the 7 species of Cecidomyiidae that were delimited with the 2% barcoding approach.
There is considerable morphological diversity in the leaf gall morphotypes found on C. barbadensis (Fig. 1A-F). All these gall morphotypes were located on the leaf blade, with 4 of them being located on the beam and 1 on the underside. The considerable morphological diversity of these leaf galls can be attributed to several factors, among which include the high synthetic activity, rapid growth, structural and functional features, and characteristic morphogenetic patterns of leaf development (Mani, 1964). Further studies on these leaf galls are therefore necessary for a better understanding of the structural and histological differences among the leaf galls found on C. barbadensis.
Insect species richness
The high morphological diversity found on leaf galls favours the existence of a complex insect community (Mani, 1964); however, few studies have assessed in detail the insect community associated with leaf galls of Neotropical plant species (e.g., Maia, 2012). In this research, the extensive insect rearing, and examination of both molecular and morphological information helped to thoroughly characterise the insect species diversity found in the 5 types of galls present on leaves of C. barbadensis. Despite that, was not possible to confirm at this stage the biology of the reared taxa, and then their probable role is based on the relevant literature and the field observations.
Cecidomyiids, commonly known as gall midges, represent by far the group of insects with the most gall-inducer species (Gagné & Jaschhof, 2021). Species of Cecidomyiidae are mainly gall-formers, though members of the tribe Cecidomyiini are known to have a wide range of biologies, including only simple and complex gall formers, free-living, mycophagous, inquiline phytophagous, predator species of mites, aphids, and coccids, as well as internal parasitoids of aphids and psyllids (Kim et al., 2014; Uechi et al., 2011). The field observations and the insect species diversity found in the 5 leaf gall morphotypes of C. barbadensis strongly suggest that they are induced by cecidomyiid species. Among this evidence was that most of the dissected galls had a single cecidomyiid larva. Moreover, several galls belonging to morphotype 2 had a cecidomyiid pupal exuviae hanging outside of a small opening, which is a common feature of many galls with former cecidomyiids. Since the 5 leaf gall morphotypes were formed by cecidomyiid species, the remaining species of this family delimited in the study probably are inquilines.
Figure 4. Neighbour Joining phenogram reconstructed for specimens of Hymenoptera that were reared from the 4 leaf gall morphotypes found in C. barbadensis. Bars refer to the 25 species of Hymenoptera that were delimited with the 2% barcoding approach.
The cecidomyiid larvae and their emerging adults could not be identified at the genus level; however, a BLAST similarity search of the barcoding locus for the delimited cecidomyiid species suggests that they are closely related to species of the Cecidomyiini genera Contarinia Geer and Macrodiplosis Kieffer. The gall-inducing species of the genus Contarinia are cosmopolitan and can be either monophagous or polyphagous with a wide range of hosts (Uechi et al., 2011). Most species of this genus live gregariously in the floral parts of the plant or in the galls that they induce on the leaves (Gagné & Jaschoff, 2021). Species of Macrodiplosis, on the other hand, are mainly gall inducers on leaves of plant species of the genus Quercus (Kim et al., 2014).
Table 1
Number of insect species discriminated by DNA sequence-based species delineation approaches conducted in this study (2% COI divergence criterion; GMYC method).
Order/Family
Subfamily
Genus
2% DC
GMYC
Diptera
Cecidomyiidae
–
–
7
7
Lepidoptera
–
–
1
–
Coleoptera
–
–
1
–
Hymenoptera
Braconidae
Doryctinae
Allorhogas coccolobae
1
1
Eulophidae
Entedoninae
Chrysonotomyia spp.
6
8
Tetrastichinae
Quadrastichus spp.
5
5
Eupelmidae
Brasema sp.
1
1
Eurytomidae
Eurytominae
Tenuipetiolus spp.
5
7
Platygastridae
Synopeas spp.
3
3
Inostemma spp.
1
1
Undetermined
2
2
Torymidae
Torymus sp.
1
1
Total
34
38
Table 2
Insect species richness by order and family associated with 5 gall morphotypes found on leaves of Coocoloba barbadensis Jacq.
Gall morphotypes (GM)
Insect Order/Family
GM 1
GM 2
GM 3
GM 4
GM 5
TOTAL
Coleoptera
–
–
1
–
–
1
Lepidoptera
–
–
1
–
–
1
Diptera
Cecidomyiidae
3
4
5
5
2
19
Hymenoptera
15
5
13
7
2
42
Eurytomidae
4
2
3
3
–
12
Eulophidae
3
2
8
1
1
15
Eupelmidae
1
–
–
–
–
1
Braconidae
1
–
–
1
–
2
Platygastridae
5
1
2
2
1
11
Torymidae
1
–
–
–
–
1
Total
18
9
20
12
4
63
Currently, 4 gall midge species are known to be associated with species of Coccoloba. The genus Ctenodactylomyia Felt (supertribe Cecidomyiidi, unplaced tribe) has 2 leaf gall inducer species on C. diversifolia Jacq., C. swartzii, and C. uvifera L. from the Caribbean (Gagné & Jaschoff, 2021). Moreover, Marilasioptera tripartite Möhn and Meunieriella magdalenae Wünsch (supertribe Lasiopteridi, tribe Alycaulini) were described as inquilines on galls induced by other insects on species of Coccoloba from El Salvador and Colombia, respectively (Gagné & Jaschoff, 2021). The present study increases the number of gall midge species associated with Coccoloba to 11.
Our study also found a considerable number of hymenopteran species reared from the 5 examined leaf gall morphotypes, most of which probably are parasitoids of the cecidomyiid species. These parasitoid species belong to the wasp families Eulophidae, Eupelmidae, Platygastridae, and Torymidae, whereas the only reared braconid species probably are phytophagous inquiline. Some members of the Platygastridae are known to be koinobiont endoparasitoids of gallery cecidomyiid eggs, and they are known to be closely associated with the parts of the plant where the host gall is found (Masner & Huggert, 1989; Masner, 1993). Eulophidae are also mainly parasitoids of holometabolous insect larvae (though in some cases also of eggs, prepupae, and pupae) of Lepidoptera, Hymenoptera, Diptera, and Coleoptera (Graham, 1991); however, some species have been reported to be phytophagous or predators (Gibson, 1993). Finally, members of the eupelmid subfamily Eupelminae, from which the genus Brasema belongs, mainly are parasitoids of larval stages of various insect hosts (Gibson, 1993).
The wasp family Eurytomidae is represented by entomophagous species that parasitise larval or pupal stages of Coleoptera, Diptera, and Hymenoptera as solitary endoparasitic idiobionts, though some species can be phytophagous feeding on seeds, or inquilines feeding on both their host and gall tissue (Lotfalizadeh et al., 2007; Gates & Delvare, 2008). Species of Tenuipetiolus have been reported to be parasitoids of gall inducer insects, including cecidomyiids and cynipids (Gates & Hanson, 2006; Zhang et al., 2014). Similarly, several species of Torymidae are known to be ectoparasitoids of gall-forming insects of the latter 2 families (Gibson, 1993).
The braconid species A. coccolobae (Doryctinae) was described a decade ago from the Chamela region in Jalisco, Mexico, based on specimens reared from leaf galls of C. barbadensis (Martínez & Zaldívar-Riverón, 2013), that correspond to the morphotype 2. Here, this braconid species was reared not only from the above leaf gall morphotype but also from morphotype 4. Several species of Allorhogas have been confirmed to feed on various plant families either by being gall formers or inquilines of galls made by other insect taxa (Centrella & Shaw, 2010; Chavarría et al., 2009; de Mâcedo et al., 1998; Zaldívar-Riverón et al., 2014). Since none of the Allorhogas species with recorded biology are known to be parasitoids, and due to the low abundance of this species compared with the cecidomyiid species that were reared from the same gall morphotypes, it is presumed that it could be a phytophagous inquiline.
The considerably high, mostly undescribed species diversity of cecidomyiid dipterans and parasitoid hymenopterans that we reared from the 5 leaf gall morphotypes of C. barbadensis remark the necessity to carry out more studies that focus on the species diversity of galls, particularly in tropical regions. Moreover, this study highlights the importance of performing integrative species delineation studies of insects present in galls to have a more accurate knowledge of their actual diversity and trophic interactions.
Acknowledgments
We thank L. Márquez, N. López, and A. Jiménez for their assistance in the laboratory (Lanabio), and Cristina Mayorga for her assistance at the CNIN-IBUNAM. Angélica Rodríguez was supported by a MSc scholarship given by the Consejo Nacional de Humanidades, Ciencias y Tecnologías (Conahcyt, Mexico), and she thanks the Posgrado en Ciencias Biológicas, UNAM, for support during her MSc studies. This study was finished thanks to the grants given by UNAM-DGAPA (PAPIIT Convocatoria 2022, Project IN201622), and Conahcyt, Mexico (Convocatoria Ciencia de Frontera, project number 58548) to AZR.
Appendix. Continued.
Order (Insecta)
Family, Genus
Species
DNA voucher No.
Genbank accession No.
Diptera
Cecidomyiidae
sp.
CeciAN255_L2
PP659768
Diptera
Cecidomyiidae
sp.
CeciAN23_L2
PP659794
Diptera
Cecidomyiidae
sp.
CeciAN193_L2
PP659795
Diptera
Cecidomyiidae
sp.
CeciAN291_A3
PP659802
Diptera
Cecidomyiidae
sp.
CeciAN292_A1
PP659803
Diptera
Cecidomyiidae
sp.
CeciAN257_L2
PP659804
Diptera
Cecidomyiidae
sp.
CeciAN259_L3
PP659805
Diptera
Cecidomyiidae
sp.
CeciAN262_L4
PP659806
Diptera
Cecidomyiidae
sp.
CeciAN207_L4
PP659797
Diptera
Cecidomyiidae
sp.
CeciAN290_A1
PP659801
Diptera
Cecidomyiidae
sp.
CeciAN160_L3
PP659807
Diptera
Cecidomyiidae
sp.
CeciAN02_L4
PP659731
Diptera
Cecidomyiidae
sp.
CeciAN05_L4
PP659732
Diptera
Cecidomyiidae
sp.
CeciAN159_L1
PP659733
Diptera
Cecidomyiidae
sp.
CeciAN165_L2
PP659734
Diptera
Cecidomyiidae
sp.
CeciAN166_L2
PP659735
Diptera
Cecidomyiidae
sp.
CeciAN167_L3
PP659736
Diptera
Cecidomyiidae
sp.
CeciAN168_L4
PP659737
Diptera
Cecidomyiidae
sp.
CeciAN169_L4
PP659738
Diptera
Cecidomyiidae
sp.
CeciAN171_L1
PP659739
Diptera
Cecidomyiidae
sp.
CeciAN173_L1
PP659740
Diptera
Cecidomyiidae
sp.
CeciAN176_L1
PP659741
Diptera
Cecidomyiidae
sp.
CeciAN180_L2
PP659742
Diptera
Cecidomyiidae
sp.
CeciAN183_L1
PP659743
Diptera
Cecidomyiidae
sp.
CeciAN186_L5
PP659744
Diptera
Cecidomyiidae
sp.
CeciAN188_L1
PP659745
Diptera
Cecidomyiidae
sp.
CeciAN191_L1
PP659747
Diptera
Cecidomyiidae
sp.
CeciAN231_L1
PP659750
Diptera
Cecidomyiidae
sp.
CeciAN233_L1
PP659751
Diptera
Cecidomyiidae
sp.
CeciAN234_L2
PP659752
Diptera
Cecidomyiidae
sp.
CeciAN237_L1
PP659753
Diptera
Cecidomyiidae
sp.
CeciAN245_L1
PP659755
Diptera
Cecidomyiidae
sp.
CeciAN267_L1
PP659756
Diptera
Cecidomyiidae
sp.
CeciAN06_A3
PP659791
Diptera
Cecidomyiidae
sp.
CeciAN209_L5
PP659749
Diptera
Cecidomyiidae
sp.
CeciAN43_A2
PP659792
Diptera
Cecidomyiidae
sp.
CeciAN189_L3
PP659746
Diptera
Cecidomyiidae
sp.
CeciAN194_L3
PP659748
Diptera
Cecidomyiidae
sp.
CeciAN239_L4
PP659754
Hymenoptera
Platygaster
sp.
TeleAN08_L3
PP481223
Hymenoptera
Platygaster
sp.
TeleAN19_P3
PP481224
Hymenoptera
Platygaster
sp.
TeleAN46_A1
PP481225
Hymenoptera
Platygaster
sp.
TeleAN45_A3
PP481226
Hymenoptera
Platygaster
sp.
TeleAN50_A3
PP481227
Hymenoptera
Platygaster
sp.
TeleAN30_A3
PP481228
Hymenoptera
Platygaster
sp.
TeleAN132_A3
PP481229
Hymenoptera
Platygaster
sp.
TeleAN133_A1
PP481230
Hymenoptera
Platygaster
sp.
TeleAN134_A3
PP481231
Hymenoptera
Platygaster
sp.
TeleAN135_A1
PP481232
Hymenoptera
Platygaster
sp.
TeleAN137_A1
PP481233
Hymenoptera
Platygaster
sp.
TeleAN138_A3
PP481234
Hymenoptera
Platygaster
sp.
TeleAN141_A3
PP481235
Hymenoptera
Platygaster
sp.
TeleAN151_A3
PP481236
Hymenoptera
Platygaster
sp.
TeleAN152_A3
PP481237
Hymenoptera
Platygaster
sp.
TeleAN153_A3
PP481238
Hymenoptera
Platygaster
sp.
TeleAN154_A3
PP481239
Hymenoptera
Platygaster
sp.
TeleAN156_A3
PP481240
Hymenoptera
Platygaster
sp.
TeleAN215_L3
PP481241
Hymenoptera
Platygaster
sp.
TeleAN216_L3
PP481242
Hymenoptera
Platygaster
sp.
TeleAN217_L3
PP481243
Hymenoptera
Platygaster
sp.
TeleAN218_L3
PP481244
Hymenoptera
Platygaster
sp.
TeleAN224_L1
PP481245
Hymenoptera
Platygaster
sp.
TeleAN225_L1
PP481246
Hymenoptera
Platygaster
sp.
TeleAN275_L3
PP481247
Hymenoptera
Platygaster
sp.
TeleAN276_L3
PP481248
Hymenoptera
Platygaster
sp.
TeleAN281_A3
PP481249
Hymenoptera
Platygaster
sp.
TeleAN158_A1
PP481250
Hymenoptera
Platygaster
sp.
TeleAN110_A1
PP481251
Hymenoptera
Platygaster
sp.
TeleAN111_A3
PP481252
Hymenoptera
Synopeas
sp.
PlatyAN22_A1
PP481253
Hymenoptera
Synopeas
sp.
PlatyAN32_A1
PP481254
Hymenoptera
Synopeas
sp.
PlatyAN52_A1
PP481255
Hymenoptera
Synopeas
sp.
PlatyAN14_A1
PP481256
Hymenoptera
Synopeas
sp.
PlatyAN157_A1
PP481257
Hymenoptera
Synopeas
sp.
PlatyAN53_A1
PP481258
Hymenoptera
Synopeas
sp.
PlatyAN99_A1
PP481259
Hymenoptera
Synopeas
sp.
PlatyAN101_A1
PP481260
Hymenoptera
Synopeas
sp.
PlatyAN102_A1
PP481261
Hymenoptera
Synopeas
sp.
PlatyAN103_A1
PP481262
Hymenoptera
Synopeas
sp.
PlatyAN105_A1
PP481263
Hymenoptera
Synopeas
sp.
PlatyAN190_L2
PP481264
Hymenoptera
Synopeas
sp.
PlatyAN219_L1
PP481265
Hymenoptera
Inostemma
sp.
PlatyAN221_L4
PP481266
Hymenoptera
Synopeas
sp.
PlatyAN223_L2
PP481267
Hymenoptera
Synopeas
sp.
PlatyAN220_L5
PP481268
Hymenoptera
Inostemma
sp.
PlatyAN112_A4
PP481269
Hymenoptera
Synopeas
sp.
PlatyAN116_A1
PP481270
Hymenoptera
Synopeas
sp.
PlatyAN117_A1
PP481271
Hymenoptera
Synopeas
sp.
PlatyAN118_A1
PP481272
Hymenoptera
Inostemma
sp.
PlatyAN126_A1
PP481273
Hymenoptera
Synopeas
sp.
PlatyAN214_L1
PP481274
Hymenoptera
Platygaster
sp.
PlatyAN222_L3
PP481275
Hymenoptera
Synopeas
sp.
PlatyAN272_A1
PP481276
Hymenoptera
Synopeas
sp.
PlatyAN273_L
PP481277
Hymenoptera
Synopeas
sp.
PlatyAN274_L
PP481278
Hymenoptera
Synopeas
sp.
PlatyAN277_L5
PP481279
Hymenoptera
Synopeas
sp.
PlatyAN278_L2
PP481280
Hymenoptera
Synopeas
sp.
PlatyAN279_L1
PP481281
Hymenoptera
Tenuipetiolus
sp.
EurytAN96_A1
PP481282
Hymenoptera
Tenuipetiolus
sp.
EurytAN265_L3
PP481283
Hymenoptera
Chrysonotomyia
sp.
EntedAN01_P1
PP481284
Hymenoptera
Chrysonotomyia
sp.
EntedAN07_A1
PP481285
Hymenoptera
Chrysonotomyia
sp.
EntedAN29_A1
PP481286
Hymenoptera
Chrysonotomyia
sp.
EntedAN11_A1
PP481287
Hymenoptera
Chrysonotomyia
sp.
EntedAN31_A1
PP481288
Hymenoptera
Chrysonotomyia
sp.
EntedAN51_A1
PP481289
Hymenoptera
Chrysonotomyia
sp.
EntedAN59_P1
PP481290
Hymenoptera
Chrysonotomyia
sp.
EntedAN61_P1
PP481291
Hymenoptera
Chrysonotomyia
sp.
EntedAN67_P1
PP481292
Hymenoptera
Chrysonotomyia
sp.
EntedAN69_A1
PP481293
Hymenoptera
Chrysonotomyia
sp.
EntedAN70_A1
PP481294
Hymenoptera
Chrysonotomyia
sp.
EntedAN71_A1
PP481295
Hymenoptera
Chrysonotomyia
sp.
EntedAN72_A1
PP481296
Hymenoptera
Chrysonotomyia
sp.
EntedAN75_A1
PP481297
Hymenoptera
Chrysonotomyia
sp.
EntedAN94_A1
PP481298
Hymenoptera
Chrysonotomyia
sp.
EntedAN95_A1
PP481299
Hymenoptera
Chrysonotomyia
sp.
EntedAN25_A2
PP481300
Hymenoptera
Chrysonotomyia
sp.
EntedAN64_P4
PP481301
Hymenoptera
Chrysonotomyia
sp.
EntedAN73_A3
PP481302
Hymenoptera
Chrysonotomyia
sp.
EntedAN142_A1
PP481303
Hymenoptera
Chrysonotomyia
sp.
EntedAN143_A1
PP481304
Hymenoptera
Chrysonotomyia
sp.
EntedAN144_A1
PP481305
Hymenoptera
Chrysonotomyia
sp.
EntedAN145_A1
PP481306
Hymenoptera
Chrysonotomyia
sp.
EntedAN146_A2
PP481307
Hymenoptera
Chrysonotomyia
sp.
EntedAN198_L3
PP481308
Hymenoptera
Chrysonotomyia
sp.
EntedAN236_L1
PP481309
Hymenoptera
Chrysonotomyia
sp.
EntedAN243_L1
PP481310
Hymenoptera
Chrysonotomyia
sp.
EntedAN244_L1
PP481311
Hymenoptera
Chrysonotomyia
sp.
EntedAN288_A2
PP481312
Hymenoptera
Chrysonotomyia
sp.
EntedAN289_A2
PP481313
Hymenoptera
Chrysonotomyia
sp.
EntedAN58_P1
PP481314
Hymenoptera
Chrysonotomyia
sp.
EntedAN55_A1
PP481315
Hymenoptera
Chrysonotomyia
sp.
EulopAN125_A5
PP481316
Hymenoptera
Tetrastichinae
sp.
TetrasAN84_A3
PP481317
Hymenoptera
Tetrastichinae
sp.
TetrasAN62_P3
PP481318
Hymenoptera
Tetrastichinae
sp.
TetrasAN63_P3
PP481319
Hymenoptera
Tetrastichinae
sp.
TetrasAN66_P3
PP481320
Hymenoptera
Tetrastichinae
sp.
TetrasAN83_A3
PP481321
Hymenoptera
Tetrastichinae
sp.
TetrasAN60_P3
PP481322
Hymenoptera
Tetrastichinae
sp.
TetrasAN269_L3
PP481323
Hymenoptera
Tetrastichinae
sp.
TetrasAN270_L3
PP481324
Hymenoptera
Tenuipetiolus
sp.
EurytAN77_A1
PP481325
Hymenoptera
Tenuipetiolus
sp.
EurytAN81_A3
PP481326
Hymenoptera
Tenuipetiolus
sp.
EurytAN93_A3
PP481327
Hymenoptera
Allorhogas
coccolobae
AllorAN18_L4
PP481328
Hymenoptera
Allorhogas
coccolobae
AllorAN28_A4
PP481329
Hymenoptera
Allorhogas
coccolobae
AllorAN35_A4
PP481330
Hymenoptera
Allorhogas
coccolobae
AllorAN48_A4
PP481331
Hymenoptera
Allorhogas
coccolobae
AllorAN56_A4
PP481332
Hymenoptera
Allorhogas
coccolobae
AllorAN107_A4
PP481333
Hymenoptera
Allorhogas
coccolobae
AllorAN108_A4
PP481334
Hymenoptera
Allorhogas
coccolobae
AllorAN109_A4
PP481335
Hymenoptera
Allorhogas
coccolobae
AllorAN121_A4
PP481336
Hymenoptera
Allorhogas
coccolobae
AllorAN122_A4
PP481337
Hymenoptera
Allorhogas
coccolobae
AllorAN195_L1
PP481338
Hymenoptera
Allorhogas
coccolobae
AllorAN248_L4
PP481339
Hymenoptera
Allorhogas
coccolobae
AllorAN249_L4
PP481340
Hymenoptera
Tenuipetiolus
sp.
EurytAN212_L3
PP481341
Hymenoptera
Tenuipetiolus
sp.
EurytAN228_L3
PP481342
Hymenoptera
Tenuipetiolus
sp.
EurytAN246_L3
PP481343
Hymenoptera
Tenuipetiolus
sp.
EurytAN285_A3
PP481344
Hymenoptera
Tenuipetiolus
sp.
EurytAN286_A3
PP481345
Hymenoptera
Tenuipetiolus
sp.
EurytAN37_A1
PP481346
Hymenoptera
Tenuipetiolus
sp.
EurytAN147_A3
PP481347
Hymenoptera
Tenuipetiolus
sp.
EurytAN148_A4
PP481348
Hymenoptera
Tenuipetiolus
sp.
EurytAN149_A3
PP481349
Hymenoptera
Tenuipetiolus
sp.
EurytAN130_A1
PP481350
Hymenoptera
Tenuipetiolus
sp.
EurytAN87_A2
PP481351
Hymenoptera
Tenuipetiolus
sp.
EurytAN91_A4
PP481352
Hymenoptera
Tenuipetiolus
sp.
EurytAN80_A3
PP481353
Hymenoptera
Tenuipetiolus
sp.
EurytAN82_A4
PP481354
Hymenoptera
Tenuipetiolus
sp.
EurytAN90_A4
PP481355
Hymenoptera
Tenuipetiolus
sp.
EurytAN85_A1
PP481356
Hymenoptera
Torymus
sp.
EurytAN57_A4
PP481357
Hymenoptera
Tenuipetiolus
sp.
EurytAN164_L1
PP481358
Hymenoptera
Tenuipetiolus
sp.
EurytAN227_L1
PP481359
Hymenoptera
Tenuipetiolus
sp.
EurytAN230_L1
PP481360
Hymenoptera
Tenuipetiolus
sp.
EurytAN283_A2
PP481361
Hymenoptera
Tenuipetiolus
sp.
EurytAN33_A4
PP481362
Hymenoptera
Tenuipetiolus
sp.
EurytAN34_A1
PP481363
Hymenoptera
Tenuipetiolus
sp.
EurytAN15_L1
PP481364
Hymenoptera
Tenuipetiolus
sp.
EurytAN114_A3
PP481365
Hymenoptera
Tenuipetiolus
sp.
EurytAN79_A1
PP481366
Hymenoptera
Tenuipetiolus
sp.
EurytAN78_A1
PP481367
Hymenoptera
Tenuipetiolus
sp.
EurytAN24_L1
PP481368
Hymenoptera
Chrysonotomyia
sp.
EurytAN88_A3
PP481369
Hymenoptera
Brasema
sp.
ToryAN170_L1
PP481370
Hymenoptera
Brasema
sp.
ToryAN196_L1
PP481371
Hymenoptera
Torymus
sp.
EurytAN229_L4
PP481372
Hymenoptera
Torymus
sp.
EurytAN266_L4
PP481373
Coleoptera
AN184
PP897664
Lepidoptera
AN250_L3
PP897665
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Bouckaert, R., Heled, J., Kühnert, D., Vaughan, T., Wu, C. H., Xie, D., Suchard, M. A., Rambaut, A., & Drummond, A. J. (2014). BEAST 2: A software platform for Bayesian evolutionary analysis. Plos Computational Biology, 10, e1003537. https://doi.org/10.1371/journal.pcbi.1003537
Centrella, M. L., & Shaw, S. R. (2010). A new species of phytophagous braconid Allorhogas minimus (Hymenoptera: Braconidae: Doryctinae) reared from fruit galls on Miconia longifolia (Melastomataceae) in Costa Rica. International Journal of Tropical Insect Science, 30, 101–107. https://doi.org/10.1017/S1742758410000147
Chavarría, L., Hanson, P., Marsh, P., & Shaw, S. (2009). A phytophagous braconid, Allorhogas conostegia sp. nov.(Hymenoptera: Braconidae), in the fruits of Conostegiaxalapensis (Bonpl.) D. Don (Melastomataceae). Journal of Natural History, 43, 2677–2689. https://doi.org/10.1080/ 00222930903243996
Clare, E. L., Schiestl, F. P., Leitch, A. R., & Chittka, L. (2013). The promise of genomics in the study of plant-pollinator interactions. Genome Biology, 14, 1–11. https://doi.org/10. 1186/gb-2013-14-6-207
de Macêdo, M. V., & Monteiro, R. F. (1989). Seed predation by a braconid wasp, Allorhogas sp. (Hymenoptera). Journal of the New York Entomological Society, 97, 358–362.
Dexter, K. G., Pennington, T. D., & Cunningham, C. W. (2010). Using DNA to assess errors in tropical tree identifications: How often are ecologists wrong and when does it matter? Ecological Monographs, 80, 267–286. https://doi.org/10.1890/09-0267.1
Drummond, A. J., Suchard, M. A., Xie, D., & Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution, 29, 1969–1973. https://doi.org/10.1093/molbev/mss075
Folmer O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
Forbes, A. A., Hall, M. C., Lund, J., Hood, G. R., Izen, R., Egan, S. P. et al. (2016). Parasitoids, hyperparasitoids, and inquilines associated with the sexual and asexual generations of the gall former, Belonocnema treatae (Hymenoptera: Cynipidae). Annals of the Entomological Society of America, 109, 49–63. https://doi.org/10.1093/aesa/sav112
Gates, M. W., & Hanson, P. E. (2006). Familia Eurytomidae, In P. E. Hanson, & I. D. Gauld (Eds.), Hymenoptera de la region tropical (pp. 380–387). Memoirs of the American Entomological Institute, 77, 1–994.
Gates, M., & Delvare, G. (2008). A new species of Eurytoma (Hymenoptera: Eurytomidae) attacking Quadrastichus spp. (Hymenoptera: Eulophidae) galling Erythrina spp. (Fabaceae), with a summary of African Eurytoma biology and species checklist. Zootaxa, 1751, 1–24. https://doi.org/10.11646/zootaxa.1751.1.1
Gibson, L. (1993). Superfamilies Mymarommatoidea and Chalcidoidea. In H. Goulet, & J. T. Huber (Eds.), Hymenoptera of the world: an identification guide to families (pp.570–655). Research Branch, Agriculture, Canada, Publication 1894.
Graham, M. W. R. V. (1991). A reclassification of the European Tetrastichinae (Hymenoptera: Eulophidae). Revision of the remaining genera. Memoirs of the American Entomological Institute, 49, 1–322.
Hebert, P. D., Cywinska, A., Ball, S. L., & DeWaard, J R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 2070, 313–321. https://doi.org/10.1098/rspb.2002.2218
Hebert, P. D., Penton, E. H., Burns, J. M., Janzen, H. D., & Hallwachs, W. (2004). Ten species in one: DNA barcoding reveals cryptic species in the Neotropical skipper butterfly Astrapes fulgerator. Proceeding of the National Academy of Sciences, 101, 14812–14817. https://doi.org/10.1073/pnas. 0406166101
Howard, R. A. (1959). Studies in the genus Coccoloba. VII A synopsis and key to species in Mexico and Central America. Journal of the Arnold Arboretum, 40, 176–203.
Ings, T. C., Montoya, J. M., Bascompte, J., Blüthgen, N., Brown, L., Dormann, C. F. et al. (2009). Ecological networks-beyond food webs. Journal of Animal Ecology, 78, 253–269. https://doi.org/10.1111/j.1365-2656.2008.01460.x
Kaartinen, R., Stone, G. N., Hearn, J., Lohse, K., & Roslin, T. (2010). Revealing secret liaisons: DNA barcoding changes our understanding of food webs. Ecological Entomology, 35, 623–638. https://doi.org/10.1111/j.1365-2311.2010.01224.x
Kaartinen, R., & Roslin, T. (2011). Shrinking by numbers: landscape context affects the species composition but not the quantitative structure of local food webs. Journal of Animal Ecology, 80, 622–631. https://doi.org/10.1111/ j.1365-2656.2011.01811.x
Kim, W., Yukawa, J., Harris, K. M., Minami, T., Matsuo, K., & Skrzypczyńska, M. (2014). Description, host range, and distribution of a new Macrodiplosis species (Diptera: Cecidomyiidae) that induces leaf-margin fold galls on deciduous Quercus (Fagaceae) with comparative notes on Palaearctic congeners. Zootaxa, 3821, 222–238. https://doi.org/10.11646/zootaxa.3821.2.3
Lotfalizadeh, H., Delvare, G., & Rasplus, J. Y. (2007). Phylogenetic analysis of Eurytominae (Chalcidoidea: Eurytomidae) based on morphological characters. Zoological Journal of the Linnean Society, 151, 441–510. https://doi.org/10.1111/j.1096-3642.2007.00308.x
Maia, V. C., Magenta, M. A. G., & Martins, S. E. (2008). Ocorrência e caracterização de galhas de insetos em áreas de restinga de Bertioga (São Paulo, Brasil). Biota Neotropica, 8, 167–197. https://doi.org/10.1590/S1676-06032 008000100020
Maia, V. C. (2012). Richness of hymenopterous galls from South America. Papéis Avulsos de Zoologia, 52, 423–429. https://doi.org/10.1590/S0031-10492012021500001
Mani, M. S. (1964). Ecology of plant galls. The Hague: W. Junk.
Martínez, J. J., & Zaldívar-Riverón, A. (2013). Seven new species of Allorhogas (Hymenoptera: Braconidae: Doryctinae) from Mexico. Revista Mexicana de Biodiversidad, 84, 117–139. https://doi.org/10.7550/rmb.31955
Masner, L., & Huggert, L. (1989). World review and keys to genera of the subfamily Inostemmatinae with the reassignment of the taxa to the Platygastrinae and Sceliotrachelinae (Hymenoptera: Platygastridae). The Memoirs of the Entomological Society of Canada, 121, 3–216.
Masner, L. (1993). Superfamily Platygastroidea. In H. Goulet, & J. T. Huber (Eds.), Hymenoptera of the world: an identification guide to families (pp.558–565). Research Branch, Agriculture, Canada, Publication 1894.
Mead, F. W. (1970). Ctenodactylomyia watsoni Felt, a gall midge of seagrape, Coccoloba uvifera L. Florida (Diptera: Cecidomyiidae). Entomology Circular, 97. Florida Department of Agriculture & Consumer Services Division of Plant Industry. Florida, USA.
Méndez-Alonzo, R., Pineda-García, F., Paz, H., Rosell, J. A., & Olson, M. E. (2013). Leaf phenology is associated with soil water availability and xylem traits in a tropical dry forest. Trees, 27, 745–754. https://doi.org/10.1007/s00468-012-0829-x
Nieves-Aldrey, J. L. (1998). Insectos que inducen la formación de agallas en las plantas: Una fascinante interacción ecológica y evolutiva. Boletín de la Sociedad Entomológica Aragonesa, 23, 3–12.
Noguera, F. A., Vega-Rivera, J. H., García-Aldrete, A. N., Quesada-Avendaño, M. (Eds.). (2002). Historia natural de Chamela. Mexico City: Universidad Nacional Autónoma de México.
Paine, R. T. (1980). Food webs: linkage, interaction strength, and community infrastructure. Journal of Animal Ecology, 49, 667–685. https://doi.org/10.2307/4220
Pons, J., Barraclough, T. G., Gomez-Zurita, J., Cardoso, A., Duran, D. P., Hazell, S., Kamoun, S., Sumlin, W. D., & Vogler, A. P. (2006). Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Systematic Biology, 55, 595-609. https://doi.org/10.1080/10635150600852011
Posada, D. (2004). Collapse: describing haplotypes from sequence alignments. Vigo, Spain: University of Vigo. https://dposada.webs.uvigo.es/
Prosser, S., Martínez-Arce, A., & Elías-Gutiérrez, M. (2013). A new set of primers for COI amplification from freshwater microcrustaceans. Molecular Ecology Resources, 13, 1151–1155. https://doi.org/10.1111/1755-0998.12132
Raman, A., & Withers, T. M. (2003). Oviposition by introduced Ophelimus eucalypti (Hymenoptera: Eulophidae) and morphogenesis of female-induced galls on Eucalyptus saligna (Myrtaceae) in New Zealand. Bulletin of Entomological Research, 93, 55–63. https://doi.org/10.1079/BER2002217
Raman, A. (2011). Morphogenesis of insect-induced plant galls facts and questions. Flora-Morphology, Distribution, Functional Ecology of Plants, 206, 517–533. https://doi.org/10.1016/j.flora.2010.08.004
Ratnasingham, S., & Hebert, P. D. N. (2013). A DNA-based registry for all animal species: the barcode index number (BIN) system. Plos One, 8, e66213. https://doi.org/10.1371/journal.pone.0066213
Razo-González, M., Castano-Meneses, G., Callejas-Chavero, A., Pérez-Velázquez, D., & Palacios-Vargas, J. G. (2014). Temporal variations of soil arthropods community structure in El Pedregal de San Ángel ecological reserve, Mexico City, Mexico. Applied Soil Ecology, 83, 88–94. https://doi.org/10.1016/j.apsoil.2014.02.007
Redfern, M., Shirley, P., & Bloxham, M. (2002). British plant galls: identification of galls on plants and fungi. Shrewsbury, UK: FSC publications.
Rodrigues, A. R., Maia, V. C., & Couri, M. S. (2014). Insect galls of restinga areas of Ilha da Marambaia, Rio de Janeiro, Brazil. Revista Brasileira de Entomologia, 58, 173–197. https://doi.org/10.1590/S0085-56262014000200010
Sheffield, C. S., Hebert, P. D. N., Kevan, P. G., & Packer, L. (2009). DNA barcoding a regional bee (Hymenoptera: Apoidea) fauna and its potential for ecological studies. Molecular Ecology Resources, 9, 196–207. https://doi.org/10.1111/j.1755-0998.2009.02645.x
Statsoft, Inc. (2011). STATISTICA (Data Analysis Software System, version 10. http://www.statsoft.com
Swofford, D. L. (2003). PAUP*. Phylogenetic Analysis using parsimony (*and other methods). Version 4.0. Sinauer Associates, Sunderland, Massachusetts.
Uechi, N., Yukawa, J., Tokuda, M., Ganaha-Kikumura, T., & Taniguchi, M. (2011). New information on host plants and distribution ranges of an invasive gall midge, Contarinia maculipennis (Diptera: Cecidomyiidae), and its congeners in Japan. Applied Entomology and Zoology, 46, 383–389. https://doi.org/10.1007/s13355-011-0050-1
Vink, C. J., Paquin, P., & Cruickshank, R. H. (2012). Taxonomy and irreproducible biological science. Bioscience, 62, 451–452. https://doi.org/10.1525/bio.2012.62.5.3
Wirta, H. K., Hebert, P. D., Kaartinen, R., Prosser, S. W., Várkonyi, G., & Roslin, T. (2014). Complementary molecular information changes our perception of food web structure. Proceedings of the National Academy of Sciences, 111, 1885–1890. https://doi.org/10.1073/pnas.1316990111
Yeo, D., Puniamoorthy, J., Ngiam, R. W. J., & Meier, R. (2018). Towards holomorphology in entomology: rapid and cost-effective adult–larva matching using NGS barcodes. Systematic Entomology, 43, 678–691. https://doi.org/10.1111/syen.12296
Zaldívar-Riverón, A., Martínez, J. J., Belokobylskij, S. A., Pedraza-Lara, C., shaw, S. R., Hanson, P. E., & Varela-Hernández, F. (2014). Systematics and evolution of gall formation in the plant-associated genera of the wasp subfamily Doryctinae (Hymenoptera: Braconidae). Systematic Entomology, 39, 633–659. https://doi.org/10.1111/syen.12078
Zhang, M. Y., Gates, M. W., & Shorthouse, J. D. (2014). Testing species limits of Eurytomidae (Hymenoptera) associated with galls induced by Diplolepis (Hymenoptera: Cynipidae) in Canada using an integrative approach. The Canadian Entomologist, 146, 321–334. https://doi.org/10.4039/tce. 2013.70
Pedro A. Aguilar-Rodríguez a, *, Beatríz del Socorro Bolívar-Cimé a, Miguel Ángel Pérez-Farrera b y M. Cristina MacSwiney G. c
a Universidad Veracruzana, Instituto de Investigaciones Forestales, Parque Ecológico “El Haya”, Carretera Antigua a Coatepec, 91070 Xalapa, Veracruz, México
b Universidad de Ciencias y Artes de Chiapas, Instituto de Ciencias Biologicas, Laboratorio de Ecología, Evolutiva, Herbario Eizi Matuda, Caleras Maciel, 29014 Tuxtla Gutiérrez, Chiapas, México
c Universidad Veracruzana, Centro de Investigaciones Tropicales, José María Morelos Núm. 44 y 45, Col. Centro, 91000 Xalapa, Veracruz, México
Recibido: 6 septiembre 2023; aceptado: 7 marzo 2024
Resumen
La flora de México está en constante actualización y las familias Bromeliaceae y Piperaceae están representadas en el país con numerosas especies, principalmente en los estados del sur. Sin embargo, las exploraciones botánicas han incrementado el número de especies reportadas para cada estado y algunas pueden encontrarse del estudio de material de herbario, de ejemplares mal identificados, o provenir de relictos de bosques cercanos a áreas urbanas. El objetivo de este trabajo es reportar nuevos registros de bromeliáceas y piperáceas en distintos estados de México, y presentar un mapa actualizado de su distribución. Entre 2018 y 2023, como parte de diferentes exploraciones botánicas, incluyendo áreas verdes periurbanas, se colectaron ejemplares y se identificaron con claves de identificación taxonómica y con expertos. Determinamos que eran nuevos aportes para las floras estatales y ampliaciones norteñas para la distribución conocida de esas especies, una de las cuales estaba presente en un remanente de bosque urbano. Los 4 registros nuevos corresponden a Pseudalcantarea macropetala, Tillandsia heterophylla, Werauhia nutans y Piper pseudoasperifolium en bosque mesófilo de montaña y vegetación secundaria, ydemuestran la importancia de continuar las exploraciones botánicas en diferentes tipos de vegetación, considerando también relictos de bosque en áreas urbanas.
Palabras clave: Áreas verdes urbanas; Bosque mesófilo de montaña; Pseudalcantarea; Piper; Tillandsia; Werauhia
New state records of Bromeliaceae and Piperaceae in Mexico
Abstract
The flora of Mexico is constantly updated, and the Bromeliaceae and Piperaceae families are represented in the country with numerous species, mainly in the southern states. However, botanical explorations have increased the species reported for each state, and some can be found by studying herbaria and misidentified specimens, or relict forests close to urban areas. This work aims to report new records of Bromeliaceae and Piperaceae for different Mexican states, an updated distribution map in Mexico. Between 2018 and 2023, as part of different botanical explorations, including peri-urban green areas, specimens were collected and identified using identification keys and experts’ help. We determined new contributions to state floras and northern extensions to the known distribution of these species, one of which was present in urban forest remnants. The four new records correspond to Pseudalcantarea macropetala, Tillandsia heterophylla, Werauhia nutans, and Piper pseudoasperifolium from montane cloud forests and secondary vegetation, demonstrating the importance of carrying out botanical explorations in different types of vegetation, also considering forest remnants in urban areas.
México posee una alta diversidad florística con más de 23,300 especies, distribuidas en 297 familias, con Oaxaca, Chiapas y Veracruz como los estados con el mayor número de especies reportadas (Villaseñor, 2016). Las familias con el mayor número de especies en el país son Asteraceae (3,057 spp.) y Fabaceae (1,903 spp.), mientras que Bromeliaceae (450 spp., 2%) y Piperaceae (245 spp., 1%) albergan un menor número (Villaseñor, 2016). Muchas especies de ambas familias son típicas del bosque mesófilo de montaña (Espejo-Serna et al., 2021; Villaseñor, 2010). Bromeliaceae es casi completamente neotropical, con un solo taxón en África, y más de la mitad de las especies son epífitas, las cuales se distribuyen en distintos hábitats, desde xéricos hasta bosques húmedos, entre los 0 y 3,000 m snm (Benzing, 2000; Espejo-Serna et al., 2005; Zotz, 2013). Dentro de la familia, el género Pseudalcantarea (Mez) Pinzón et Barfuss está representado en México por sus únicas 3 especies reconocidas: Pseudalcantarea grandis (Schltdl.) Pinzón et Barfuss, P. macropetala (Wawra) Pinzón et Barfuss y P. viridiflora (Beer) Pinzón et Barfuss. Estas especies estaban incluidas dentro del género Tillandsia L. (Barfuss et al., 2016), mientras que los géneros Tillandsia y Werauhia J.R. Grant. están representados por 238 y 11 especies, respectivamente. En algunos estados como Hidalgo, San Luis Potosí y Tabasco, el número de Bromeliaceae registradas no superan las 60 especies y algunos géneros, por ejemplo, Tillandsia en San Luis Potosí (19 spp.) y Werauhia en Tabasco (1 sp.), están pobremente representados (Espejo-Serna et al., 2021).
En México, Pseudalcantarea macropetala es registrada para Chiapas, Oaxaca, Puebla y Veracruz (Espejo-Serna et al., 2021), Tillandsia heterophylla É. Morren para Hidalgo, Puebla y Veracruz, mientras que Werauhia nutans (L.B. Sm.) J.R. Grant solo es conocida de Chiapas, Oaxaca y Veracruz (Espejo-Serna et al., 2005, 2021). Por otro lado, de los 2 géneros de Piperaceae registrados en México, Piper posee el mayor número de especies (136 spp.) y Veracruz es el estado con más registros (89 spp.; Villaseñor, 2016). Piper es un género pantropical pero alcanza su mayor diversidad en la región neotropical y se distribuye en una amplia variedad de hábitats, desde los bosques de tierras bajas hasta los bosques montanos, mayormente entre los 0 y 2,500 m snm (Quijano-Abril et al., 2006). En México, Piper pseudoasperifolium C. DC. se distribuye hasta Costa Rica (Tebbs, 1993; Tropicos, 2022), se registra en Chiapas, Guerrero, Jalisco, Nayarit y Oaxaca (Villaseñor, 2016).
En el presente trabajo se reportan por primera vez las Bromeliaceae epífitas Pseudalcantarea macropetala para Hidalgo, Tillandsia heterophylla para San Luis Potosí y Werauhia nutans para Tabasco, y Piper pseudoasperifolium para Veracruz, también se presenta un mapa de la distribución de las 4 especies en México.
Materiales y métodos
Como parte de un taller de divulgación científica en Hidalgo (Tenango de Doria, Centro Ecoturístico “Agua Viva”, 17º21’51” N, 93º37’34” O), exploraciones botánicas en San Luis Potosí (Xilitla, camino a La Silleta, 21º26’20” N, 99º00’30” O) y Tabasco (Villa de Guadalupe, 17º21’51” N, 93º37’34” O), y dentro del proyecto “Funcionalidad socioecológica de áreas verdes urbanas neotropicales en Veracruz” (Jardín Botánico Francisco Javier Clavijero (19º30’53” N, 96º58’36” O), Xalapa y área natural protegida (ANP) La Martinica (19º34’20” N, 96º56’04” O), Banderilla, entre el 2018 y 2023, se colectaron varios especímenes de las familias Bromeliaceae y Piperaceae, entre ellas Pseudalcantarea macropetala, Piper pseudoasperifolium, Tillandsia heterophylla y Werauhia nutans. Si el ejemplar no se encontraba reproductivo, en el caso de las bromelias epífitas, se puso en cultivo en la localidad para su monitoreo hasta el desarrollo completo de la inflorescencia. El material colectado se procesó y se determinó usando claves de identificación taxonómica y monografías de ambos grupos (Espejo-Serna et al., 2005, 2021; Samain y Tebbs, 2020; Tebbs, 1993) y revisión de los herbarios CIB, HEM, MEXU y XAL (acrónimos de acuerdo a Thiers [2016]). Adicionalmente, se revisaron los trabajos de Tebbs (1993); Espejo-Serna et al. (2005, 2021) y para la distribución, se consultaron listados florísticos (Espejo-Serna y López-Ferrari 2004; Espejo-Serna et al., 2005, 2021; Villaseñor, 2016) y bases de datos, Portal de Datos Abiertos UNAM (UNAM, 2022) y Missouri Botanical Garden (TROPICOS, 2022).
Usando la información de un espécimen por estado (tabla 1), se elaboró un mapa de distribución de las 4 especies en México, Pseudalcantarea macropetala: P. A. Aguilar-Rodríguez PA001 (CITRO), P. A. Aguilar-Rodríguez et al. PA0022 (HEM), A. Espejo et al. 6700 (UAMIZ), J. Martínez Meléndez 1785 (HEM), F. Ventura A. 1061 (MEXU); Piper pseudoasperifolium: J. S. Miller et al. 2715 (MO), G. Flores F. y R. Ramírez R. 2514 (MO), R. Torres C. y A. Campos V. 10697 (MO), Calónico-Soto 8878 (MEXU), P. A. Aguilar-Rodríguez PA0025 (HEM); T. heterophylla: J. Ceja et al. 1869 (IEB, UAMIZ), A. Mendoza R. et al. 691 (UAMIZ), T. B. Croat 39637 (MO), M. A. Pérez-Farrera et al. 4019 (HEM); Werauhia nutans: E. Martínez S. y M. A. Soto A. 18654 (MEXU), P. Tenorio y T. Wendt 19335 (UAMIZ), P. A. Aguilar-Rodríguez et al. PA0014 (HEM), P. A. Aguilar-Rodríguez PA0012 (CITRO).
Todo el material colectado se etiquetó y se depositó en el herbario HEM. La identificación de Piper pseudoasperifolium fue corroborada por el especialista en la familia Piperaceae, Ricardo Callejas Posada, del Instituto de Biología, Universidad de Antioquia, Medellín, Colombia.
Resultados
Pseudalcantarea macropetala (Wawra) Pinzón et Barfuss. Fig. 1A.
Hierba epífita, raramente rupícola, arrosetada, de hasta 175 cm de alto, solitaria, con láminas acintadas, de entre 45 y 50 cm de largo, punctulado-lepidotas en el haz y envés, erectas a ascendentes. Inflorescencias terminales, erectas, compuestas, raramente simples, con hasta 7 espigas y un escapo de cerca de 80 cm de largo, cubierto por brácteas imbricadas. Cada espiga contiene hasta 20 flores, con disposición dística y actinomorfas, pétalos color verde claro en antesis, acintados y enrollados en el extremo sobre sí mismos, de hasta 12.3 cm de largo y los estambres subiguales. Cápsulas y semillas fusiformes, estas últimas con un apéndice plumoso blanco. La floración de esta especie ocurre de diciembre a abril, según la localidad, presenta antesis nocturna y sus flores son polinizadas por murciélagos (citada como Tillandsia macropetala; Aguilar-Rodríguez et al., 2014; Krömer et al., 2012). Es posible que se distribuya hasta Centroamérica (Krömer et al., 2012), pero en México, se distribuye en Chiapas, Hidalgo, Oaxaca, Puebla y Veracruz (fig. 2). Crece en bosque tropical subcaducifolio y bosque mesófilo de montaña, entre 1,200 y 2,500 m snm (Espejo-Serna et al., 2021).
Ejemplar examinado: México. Hidalgo, municipio Tenango de Doria, Centro Ecoturístico “Agua Viva”, 20°20’15.93” N, 98°15’29.38” O, 1,395 m, 29.X.2021, P. A. Aguilar-Rodríguez et al. PA0022 (HEM).
Tillandsia heterophylla É. Morren, Belgique Hort. 23: 138. 1873. Tipo: México, Veracruz, aux environs de Córdoba “Cordova”. O. Malzine s. n. (Holotipo: LG). Fig. 1B.
T. virginalis É. Morren, Belgique Hort. 30: 238-240. 1880, nom. illeg.
Hierba epífita arrosetada, glauca, pruinosa de hasta 150 cm de alto, solitaria, con láminas de las hojas acintadas de cerca de 30 a 40 cm de largo, de color verde o teñido de púrpura, punctulado-lepidotas en el haz y envés, acuminadas y erecto-extendidas al ápice. Inflorescencias terminales, erectas, compuestas, a veces simples, con hasta 16 espigas pruinosas y un escapo de hasta 80 cm, cubierto desde la base al ápice por las vainas de las brácteas. Las flores son tubular-campaniformes, con pétalos blancos y estambres más cortos que los pétalos. Cápsulas rostradas y semillas fusiformes, con un apéndice plumoso blanco. Su floración ocurre de febrero a julio, con antesis nocturna a brevemente diurna y sus flores son polinizadas por murciélagos, polillas, colibríes y abejas (Aguilar-Rodríguez et al., 2016; Espejo-Serna et al., 2005). Se le considera endémica de México y se distribuye en Hidalgo, Puebla, San Luis Potosí y Veracruz (fig. 2). Crece en bosque de Quercus y bosque mesófilo de montaña, entre 600 y 1,700 m snm (Espejo-Serna et al., 2005, 2021).
Figura 1. A, Pseudalcantarea macropetala con inflorescencia; B, Tillandsia heterophylla con inflorescencia en desarrollo; C, Werauhia nutans con inflorescencia en desarrollo; y D, Piper pseudoasperifolium con inflorescencia.
Ejemplar examinado: México. San Luis Potosí, municipio Xilitla, camino a La Silleta, 21°25’45” N, 99°00’38” O, 750 m, 31.VII.2021, M. A. Pérez-Farrera et al. 4019 (HEM).
Vriesea nutans L. B. Sm., Phytologia 7(4): 175. 1960. Tipo: Costa Rica, San José, road from Turrialba to Moravia, M. B. Foster 2727 (holotipo: US)
Hierba epífita, arrosetada, de hasta 60 cm de alto, solitaria pero frecuente que forme agrupaciones de varios individuos, con láminas de las hojas acintadas de cerca de 40 cm de largo, punctulado-lepidotas, glabrescentes en haz y envés, agudas a atenuadas en el ápice. Las inflorescencias son terminales con un escapo delgado, de hasta 3 mm de diámetro y de hasta 45 cm de largo, cubierto en toda su longitud por las vainas de las brácteas, las espigas alcanzan hasta 12.5 cm de largo con 5-9 flores con un arreglo dística y zigomorfas, brácteas florales quebradizas cuando están secas. Flores con pétalos blancos y estambres desiguales más corto que los pétalos. Cápsulas pardas oscuras, fusiforme, de hasta 3.3 cm de largo con semillas de hasta 4 mm de largo, con un apéndice plumoso. Su floración ocurre de octubre a diciembre, presenta antesis nocturna y se reproduce por autopolinización y clonación (Aguilar-Rodríguez et al., 2019; Espejo-Serna et al., 2005). Se distribuye desde México hasta Costa Rica. En México ha sido colectada en Oaxaca, Chiapas, Tabasco y Veracruz (fig. 2). La especie crece en bosque tropical perennifolio y bosque mesófilo de montaña, entre 100 y 1,100 m snm (Espejo-Serna et al., 2005, 2007, 2021).
Tabla 1
Sitios de colecta de ejemplares de Pseudalcantarea macropetala, Tillandsia heterophylla, Werauhia nutans y Piper pseudoasperifolium revisadas en el artículo y representadas en la figura 2; se incluyen las localidades de los nuevos registros estatales (en negritas). Abreviaturas de los estados: CHI (Chiapas), GUE (Guerrero), HID (Hidalgo), NAY (Nayarit), OAX (Oaxaca), PUE (Puebla), SLP (San Luis Potosí), TAB (Tabasco), VER (Veracruz).
Estados
Especie (familia)
Pseudalcantarea macropetala (Bromeliaceae)
Tillandsia heterophylla (Bromeliaceae)
Werauhia nutans (Bromeliaceae)
Piper pseudoasperifolium (Piperaceae)
CHI
J. Martínez Meléndez 1785 (HEM), 15°45’39” N 093°04’19” O
E. Martínez S. y M. A. Soto A. 18654 (MEXU), 16°57’15” N 091°35’49” O
J. S. Miller et al. 2715 (MO), 15°06’00” N 092°04’48” O
GUE
Calónico-Soto 8878 (MEXU), 17°35’33” N 099°44’27” O
HID
P. A. Aguilar-Rodríguez et al. PA0022 (HEM), 20º20’15.93” N, 98º15’29.38” O
J. Ceja et al. 1869 (IEB, UAMIZ), 20°45’52” N 098°39’53” O
NAY
G. Flores F. y R. Ramírez R. 2514 (MO), 21°29’N 104°55’W
OAX
A. Campos V. y R. Torres C. 3614 (HEM, MEXU), 16°47’16” N 095°21’59” O
P. Tenorio y T. Wendt 19335 (UAMIZ), 17°08’ N 94°28’ O
R. Torres C. y A. Campos V. 10697 (MO), 16°57’00” N 096°13’00” O
PUE
F. Ventura A. 1061 (MEXU), 19°49›20” N 097°23’51” O
A. Mendoza R. et al. 691 (UAMIZ), 20°01’58” N 097°31’07” O
SLP
M. A. Pérez-Farrera et al. 1056 (HEM), 21°25’45” N, 99°00’38” O
TAB
P. A. Aguilar-Rodríguez et al. PA0015 (HEM), 17º21’51’’ N, 93º37’34’’ W
VER
P. A. Aguilar-Rodríguez PA001 (CITRO), 19°31’12” N 096°59’17” O
T. B. Croat 39637 (MO), 19°09’36” N 096°56’24” O
P. A. Aguilar-Rodríguez PA0012 (CITRO) 18°32’03” N 095°08’18” O
P. A. Aguilar-Rodríguez et al. PA0025 (HEM), 19°35’23” N 096°57’01” O
Ejemplar examinado: México. Tabasco, municipio Huimanguillo, Cerro de las Flores, Villa de Guadalupe, 17°21’51” N, 93°37’34” O, 1,024 m, 7.V.2018, P. A. Aguilar-Rodríguez et al. PA0014 (HEM).
Figura 1. A, Pseudalcantarea macropetala con inflorescencia; B, Tillandsia heterophylla con inflorescencia en desarrollo; C, Werauhia nutans con inflorescencia en desarrollo; y D, Piper pseudoasperifolium con inflorescencia.
Piper pseudoasperifolium C. DC., Prodr. 16(1): 318 (1869). Isotipo: México, Oaxaca, Franco s. n. (G-DC). Fig. 1D.
Arbusto de hasta 5 m de altura, con tallos y peciolos densamente pubescentes. Hojas ovado-lanceoladas a elíptico-lanceoladas, buliformes, glandulosas, con el haz escabroso y el envés densamente pubescente sobre los nervios. Inflorescencias con pedúnculos pubescentes, erectas y tienen menos de 10 cm de largo; los frutos son redondos y glabros, verde claro, de entre 3 y 6 cm de largo. Crece principalmente en bosque mesófilo de montaña, en matorrales y en pinares, entre 0 y 2,010 m snm. Se distribuye de México a Costa Rica (Callejas-Posada, 2020; Tebbs, 1993). Su floración ocurre todo el año (PA Aguilar-R., obs. pers.) y la información sobre su biología reproductiva es desconocida. En México esta especie solo había sido registrada para Chiapas, Guerrero, Jalisco, Nayarit y Oaxaca (fig. 2). Crece en bosque húmedos tropicales, entre 300 y 1,400 m snm (Callejas-Posada, 2020; Villaseñor, 2016).
Figura 2. Presencia en México de algunos ejemplares representativos para la distribución nacional de Pseudalcantarea macropetala cuadros en naranja), Tillandsia heterophylla (círculos verdes), Werauhia nutans (círculos negros) y Piper pseudoasperifolium (cuadros azules), en México. Las coordenadas de los ejemplares colectados se encuentran en la tabla 1.
Ejemplares examinados: México. Veracruz, municipio Banderilla, Área Natural Protegida, La Martinica, 19°35’23” N, 96°57’01” O, 1,543 m, 25.III.2023, P. A. Aguilar-Rodríguez PA0025 (HEM); municipio Xalapa, Jardín Botánico Francisco Javier Clavijero, 19°30’50” N, 96°56’38” O, 1,320 m snm, 25.III.2023, P. A. Aguilar-Rodríguez PA0023 (HEM).
Discusión
El hallazgo de estas 4 especies de plantas en estados con inventarios florísticos recientes (p.e., Tobías-Baeza et al., 2019; Torres-Colín et al., 2017; Vargas-Rueda et al., 2019; Villaseñor et al., 2022) demuestra la importancia de continuar realizando exploraciones y colectas en los diferentes tipos de vegetación de México, pero particularmente en el bosque mesófilo de montaña (solamente T. heterophylla fue colectada en un bosque de encino), uno de los tipos de vegetación en México con una alta diversidad florística (hasta 6,790 spp.; Villaseñor, 2010) y donde la mayoría de las especies reportadas en este estudio fueron encontradas, así como en los fragmentos boscosos urbanos.
Además, las Bromeliaceae Pseudalcantarea macropetala, para Hidalgo, Tillandsia heterophylla para San Luis Potosí, corresponden a los registros más norteños para estas especies con distribución exclusiva en México. Aunque probablemente la primera especie tenga una presencia en Centroamérica, aún no se han analizado ejemplares de herbario de Pseudalcantarea viridiflora, toda vez que hay ejemplares entremezclados de ambas especies en diferentes herbarios (PA Aguilar-R, obs. pers.).
Pseudalcantarea grandis (Schltdl.) Pinzón et Barfuss y P. viridiflora (Beer) Pinzón et Barfuss, son las otras 2 especies del género presentes en México, ambas también se encuentran en Hidalgo (Espejo-Serna et al., 2005, 2021). El registro de P. macropetala representa la tercera especie del género para el estado, mientras que el reporte de Tillandsia heterophylla para San Luis Potosí aumenta el número a 20 spp. dentro del género en el estado (Espejo-Serna et al., 2021). Anteriormente, T. heterophylla se había reportado solo para Hidalgo, Puebla y Veracruz (Espejo-Serna et al., 2005, 2021). El espécimen Reyes-García 6495 (MEXU, MO) colectado en Chiapas y determinado como T. heterophylla debe ser analizado para confirmar su determinación. La especie es abundante en los sitios donde se encuentra y parece seguir una distribución a través de la provincia biogeográfica de la Sierra Madre Oriental (sensu Morrone et al., 2002), por lo que, hipotetizamos, que al menos en el sur de Tamaulipas, pueda estar presente esta especie en hábitats boscosos.
Por otro lado, en este estudio se confirma la presencia de W. nutans para Tabasco, confirmando su presencia en otra localidad de la provincia biogeográfica Veracruzana (sensu Morrone et al., 2002) (tabla 1) y representa la tercera especie del género para el estado, ya que previamente se había reportado W. gladioliflora (H. Wendl.) J.R. Grant (Espejo-Serna et al., 2005, 2021) y W. maculata Espejo-Serna, López-Ferrari, Aguilar-Rodr., Diaz Jim. (Aguilar-Rodríguez et al., 2020).
El reporte de Piper pseudoasperifolium como nuevo registro para Veracruz eleva el número a 90 spp. del género Piper en el estado, lo cual representa 66% de las 136 spp. reportadas en México (Villaseñor, 2016). En la revisión de herbarios y bases de datos se pudo constatar que, a diferencia de otras especies de Piper (p.e., P. aduncum L., P. amalago L., P. hispidum Sw.), P. pseudoasperifolium ha sido poco recolectada en México, con menos de 17 especímenes registrados en Chiapas, 3 en Oaxaca, 2 en Nayarit y 1 en Guerrero, abarcando varias provincias biogeográficas con distintas características, como la del Balsas, Tierras bajas del Pacífico, Veracruzana y de la Sierra Madre del Sur (sensu Morrone et al., 2002). Esto sugiere que esta especie también tendrá presencia en Jalisco, Michoacán y quizá Colima, siendo así una especie con representación en las regiones neártica y neotropical. En Veracruz, esta especie se recolectó en el Jardín Botánico Francisco Javier Clavijero, en el municipio de Xalapa y en el Área Natural Protegida La Martinica, en Banderilla, como parte del proyecto “Funcionalidad socioecológica de áreas verdes urbanas neotropicales en Veracruz”. En ambos sitios, P. pseudoasperifolium crece abundante, tanto en la orilla del bosque, como en el sotobosque y simpátrica con otras especies del mismo género como P. auritum y P. lapathifolium (PA Aguilar-R pers. obs.), lo que sugiere una especie con una gran adaptabilidad a la perturbación antrópica y que crece en simpatría con otras especies morfológicamente muy similares.
Agradecimientos
A Pedro Díaz Jiménez, Melany Aguilar López, Mané Salinas Rodríguez, Diego A. Báez Ruiz, y Rocío Cárcamo Corona por su ayuda en campo y al procesar las muestras; a Ricardo Callejas-Posada por su valiosa ayuda en la identificación Piper. A Adolfo Espejo-Serna por su ayuda para la revisión de especímenes. Y a Mauricio Gerónimo Martínez-Martínez y Melany Aguilar López por la elaboración del mapa. Algunas salidas de campo fueron solventadas por parte del proyecto de Ciencia Básica del Conahcyt “Funcionalidad socioecológica de áreas verdes urbanas neotropicales” (64358).
Referencias
Aguilar-Rodríguez, P. A., Díaz-Jiménez, P., Espejo-Serna, A., López-Ferrari, A. R., Hentrich, H., Yovel, Y. et al. (2020). A new Werauhia (Tillandsioideae, Bromeliaceae) from Mexico with observations about its reproductive biology. Phytotaxa, 446, 128–134. https://doi.org/10.11646/phytotaxa.446.2.6
Aguilar-Rodríguez, P. A., Krömer, T., García-Franco, J. G. y MacSwiney G., M. C. (2016). From dusk till dawn: nocturnal and diurnal pollination in the epiphyte Tillandsia heterophylla (Bromeliaceae). Plant Biology, 18, 37–45. https://doi.org/10.1111/plb.12319
Aguilar-Rodríguez, P. A., MacSwiney G., M. C., Krömer, T., García-Franco, J. G., Knauer, A. y Kessler, M. (2014). First record of bat-pollination in the species-rich genus Tillandsia (Bromeliaceae). Annals of Botany, 113, 1047–1055. https://doi.org/10.1093/aob/mcu031
Aguilar-Rodríguez, P. A., Tschapka, M., García-Franco, J. G., Krömer, T. y MacSwiney G., M. C. (2019). Bromeliads going batty: pollinator partitioning among sympatric chiropterophilous Bromeliaceae. AoB Plants, 11, 1–19. https://doi.org/10.1093/aobpla/plz014
Barfuss, M. H. J., Till, W., Leme, E. M. C., Pinzón, J. P., Manzanares, J. M., Halbritter, H. et al. (2016). Taxonomic revision of Bromeliaceae subfam. Tillandsioideae based on a multi- locus DNA sequence phylogeny and morphology. Phytotaxa, 279, 1–97. https://doi.org/10.11646/phytotaxa.279.1.1
Benzing, D. H. (2000). Bromeliaceae. Profile of an adaptive radiation. Cambridge, RU: Cambridge University Press.
Espejo-Serna, A. y López-Ferrari, A. R. (2004). Notas sobre la familia Bromeliaceae en el Valle de México. ActaBotanica Mexicana, 67, 49–57. https://doi.org/10.21829/abm67.2004.973
Espejo-Serna, A., López-Ferrari, A. R. y Ramírez-Morillo, I. (2005). Bromeliaceae. Flora de Veracruz. Fascículo 136. Xalapa: Instituto de Ecología, A.C.
Espejo-Serna, A., López-Ferrari, A. R., Martínez-Correa, N. y Pulido Esparza, V. A. (2007). Bromeliad flora of Oaxaca, Mexico: Richness and distribution. Acta Botanica Mexicana, 81, 71–147. https://doi.org/10.21829/abm81.2007.1025
Espejo-Serna, A., López-Ferrari, A. R., Mendoza-Ruiz, A., García-Cruz, J., Ceja-Romero, J. y Pérez-García, B. (2021). Mexican vascular epiphytes: richness and distribution. Phytotaxa, 503, 001–124. https://doi.org/10.11646/phyto taxa.503.1.1
Krömer, T., Espejo-Serna, A., López-Ferreri, A. R., Ehlers, R. y Lautner, J. (2012). Taxonomic and nomenclatural status of the Mexican species in the Tillandsia viridiflora complex (Bromeliaceae). Acta Botanica Mexicana, 99, 1–20. https://doi.org/10.21829/abm99.2012.16
Morrone, J., Espinosa-Organista, D. y Llorente-Bousquets, J. (2002). Mexican biogeographic provinces: preliminary scheme, general characterizations, and synonymies. Acta Zoológica Mexicana, 85, 83–108. https://doi.org/10.21829/azm.2002.85851814
Quijano-Abril, M. A., Callejas-Posada, R. y Miranda-Esquivel, D. R. (2006). Areas of endemism and distribution patterns for Neotropical Piper species (Piperaceae). Journal of Biogeography, 33, 1266–1278. https://doi.org/10.1111/j.1365-2699.2006.01501.x
Samain, M. S. y Tebbs, M. C. (2020). Piperaceae. Flora del Bajío y de Regiones Adyacentes, 215, 1–62.
Tebbs, M. C. (1993). Revision of Piper in the new in the New World. 3. The taxonomy of Piper sections Lepiantes and Radula. Bulletin of the Natural History Museum of London, 23, 1-50.
Thiers, B. (2016) Index Herbariorum. A global directory of public herbaria and associated staff. New York Botanical Garden`s virtual herbarium. https://sweetgum.nybg.org/science/ih/
Tobías-Baeza, A., Salvador-Morales, P., Sánchez-Hernández, R., Ruiz-Acosta, S. C., Arrieta-Rivera, A. y Andrade-Prado, H. (2019). Composición florística y carbono en la vegetación arbórea de un área periurbana en Tabasco, México. Ecosistemas y Recursos Agropecuarios, 6, 369–376. https://doi.org/10.19136/era.a6n17.2009
Torres-Colín, R., Parra, J. G., de la Cruz, L. A., Ramírez, M. P., Gómez-Hinostrosa, C., Bárcenas, R. T. et al. (2017). Flora vascular del municipio de Guadalcázar y zonas adyacentes, San Luis Potosí, México. Revista Mexicana de Biodiversidad, 88, 524–554. https://doi.org/10.1016/j.rmb.2017.07.003
TROPICOS. (2022a). Missouri Botanical Garden. Recuperado el 10 de septiembre de 2022 de: https://www.tropicos.org/home
Vargas-Rueda, A. F., Rivera-Hernández, J. E., Cházaro-Basáñez, M. J. y Alcántara-Salinas, G. (2019). Nuevos registros para la flora de Veracruz en el Parque Nacional Cañón del Río Blanco, México. Acta Botanica Mexicana, 126, e1429. https://doi.org/10.21829/abm126.2019.1429
Villaseñor, J. L. (2010). El bosque húmedo de montaña en México y sus plantas vasculares. Catálogo florístico taxonómico. Ciudad de México: Conabio/ UNAM.
Villaseñor, J. L. (2016). Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidad, 87, 559–902. https://doi.org/10.1016/j.rmb.2016.06.017
Villaseñor, J. L., Ortiz, E. y Sánchez-González, A. (2022). Riqueza y distribución de la flora vascular del estado de Hidalgo, México. Revista Mexicana de Biodiversidad, 93, e933920. https://doi.org/10.22201/ib.20078706e.2022.93.3920
Zotz, G. (2013). The systematic distribution of vascular epiphytes —a critical update. Botanical Journal of the Linnean Society, 171, 453–481. https://doi.org/10.1111/boj.12010
Carolina González-Pardo a, Ireri Suazo-Ortuño a, *, Cinthya Mendoza-Almeralla b, David Tafolla-Venegas c, Yurixhi Maldonado-López a y Esperanza Meléndez-Herrera a
a Universidad Michoacana de San Nicolás de Hidalgo, Instituto de Investigaciones sobre los Recursos Naturales, Avenida San Juanito Itzícuaro s/n, Nueva Esperanza, 58330 Morelia, Michoacán, México
b Universidad Autonóma del Estado de Hidalgo, Instituto de Ciencias Básicas e Ingeniería, Centro de Investigaciones Biológicas, Laboratorio de Ecología de Poblaciones, Km 4.5 carretera Pachuca-Tulancingo, 42184 Mineral de La Reforma, Hidalgo, México
c Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Biología, Edificio R, Ciudad Universitaria, 58030 Morelia, Michoacán, México
Recibido: 7 agosto 2023; aceptado: 22 febrero 2024
Resumen
La evaluación del perfil de leucocitos como biomarcador hematológico en las poblaciones de anfibios es cada vez más común en estudios ecológicos en especies amenazadas o en declive. En este estudio evaluamos y comparamos el perfil de leucocitos y el índice neutrófilos/linfocitos (N/L) en frotis de sangre periférica de Ambystoma ordinarium en 3 tipos de hábitats: conservados, urbanizados y agrícolas. Consideramos al perfil leucocitario como un endpoint inmunológico, ya que nos puede proporcionar información sobre la respuesta inmunológica del organismo. De acuerdo con los resultados encontrados, en los individuos de A. ordinarium de los sitios urbanizados y agrícolas se detectaron aumentos en las proporciones de eosinófilos, basófilos y monocitos, y una disminución en las proporciones de linfocitos. Asimismo, en los individuos de los sitios urbanizados y agrícolas se detectaron aumentos en el número de neutrófilos banda, además se reporta por primera vez el hallazgo de células plasmáticas en la sangre de esta especie. En general, los perfiles de leucocitos de los individuos de A. ordinarium en los sitios urbanizados y agrícolas observados en este estudio, podrían interpretarse como respuestas fisiológicas a la perturbación ambiental.
Leukocyte profile as hematologic biomarker in populations of the mountain salamander, Ambystoma ordinarium
Abstract
Assessing the leukocyte profile as a hematological biomarker is now frequently used in ecological studies of threatened or declining species. In this study, we evaluated and compared leukocytes profile and neutrophils/lymphocytes (N/L) ratio in peripheral blood smears of the salamander Ambystoma ordinarium in 3 types of habitats: urbanized, agricultural, and conserved. We considered leukocyte profiles as an immunological endpoint, since it can provide information about the immunological response. Results indicated that A. ordinarium individuals from the urbanized and agricultural sites presented higher proportions of neutrophils, eosinophils, basophils and monocytes and a decrease in the proportions of lymphocytes. Agricultural habitats presented higher N/L ratios. Likewise, in the individuals of urbanized and agricultural sites an increase was registered in the number of neutrophils with a band nucleus, in addition, the finding of plasma cells in the blood of this species is reported for the first time. In general, leukocyte profiles of A. ordinarium individuals in urbanized and agricultural sites observed in this study suggest that these profiles can be interpreted as physiological responses to environmental disturbance.
En la actualidad, una de las preocupaciones más importantes en la conservación de vida silvestre es la pérdida y disminución global de las especies de anfibios (Alvarado, 2021). Los cambios en los hábitats asociados a las actividades antropogénicas y las enfermedades infecciosas representan las principales amenazas (Wake y Vredenburg, 2008). Sin embargo, científicos en todo el mundo, consideran que no existe una sola causa potencial, sino que éstos y otros factores pueden actuar mediante sinergias contribuyendo en la disminución de sus poblaciones (Lips et al., 2005; Stuart et al., 2004).
Los anfibios, pueden ser más vulnerables a los cambios en sus hábitats en comparación con el resto de los vertebrados, por 2 razones principales: poseen una piel delgada y porosa que es permeable al agua, y son organismos ectotermos, por lo que dependen de su entorno para conservar su temperatura (Duellman y Trueb, 1994). Por estos motivos, la evaluación del estado de salud de las poblaciones de anfibios es cada vez más común en estudios ecológicos en especies amenazadas o en declive (Barriga-Vallejo et al., 2015; Das y Mahapatra, 2014; Shutler y Marcogliese, 2011). Los perfiles de leucocitos han sido evaluados con mayor frecuencia porque proporcionan información sobre el estado inmunológico y permiten detectar cambios fisiológicos y patológicos tempranos en los individuos, sobretodo, estudios recientes han comenzado a incorporarlos como biomarcadores para evaluar la salud de los individuos y de su ambiente (Barni et al., 2007; Cabagna et al., 2005; Davis et al., 2010; Salinas et al., 2015, 2019).
Los leucocitos (linfocitos, neutrófilos, eosinófilos, basófilos y monocitos) son células sanguíneas que forman parte del sistema inmunitario, desempeñando funciones cruciales en la defensa contra infecciones y enfermedades (Thrall, 2004). De esta forma, los leucocitos en sangre pueden aumentar rápidamente en una infección, por ejemplo, aumentos en las proporciones de eosinófilos se han asociado con infecciones parasitarias (Davis y Golladay, 2019; Ramírez-Hernández et al., 2019) y en evidencia reciente, se han reportado aumentos en las frecuencias de linfocitos maduros e inmaduros (Salinas et al., 2019). El perfil de leucocitos también ha sido evaluado con éxito como indicador de estrés en poblaciones en ambientes perturbados y alteraciones morfológicas como el aumento de neutrófilos sin segmentación nuclear se han relacionado con ambientes contaminados con desechos urbanos y agrícolas (Barni et al., 2007; Cabagna et al., 2005; Ramírez-Hernández et al., 2019; Romanova y Romanova, 2003).
En México, habitan 14 especies del género Ambystoma (Ramírez-Bautista et al., 2023) y se ha estudiado el perfil de leucocitos en algunas especies como biomarcador de inflamación y estrés asociado a perturbaciones antropogénicas (Barriga-Vallejo et al., 2015; Ramírez-Hernández et al., 2019). La salamandra de arroyo Ambystoma ordinarium se distribuye en el noreste de Michoacán y se encuentra catalogada como en peligro de extinción por la IUCN (2024), y como especie protegida por el gobierno de México (Semarnat, 2010). Particularmente, en varios sitios del área de distribución de esta especie existe un fuerte impacto sobre los arroyos que habita debido a presiones de urbanización, actividades agrícolas y ganaderas (Soto-Rojas, 2012). Considerando el contexto en el que se encuentra esta especie, es importante monitorear sus poblaciones, sobre todo las que están sujetas a la continua perturbación de sus hábitats. Por lo tanto, el objetivo de este estudio fue evaluar y comparar los perfiles de leucocitos como biomarcador hematológico en poblaciones de A. ordinarium de hábitats con diferentes grados de perturbación: conservados, urbanizados y agrícolas.
Materiales y métodos
Se realizaron 2 visitas, la primera en noviembre de 2020 y la segunda en marzo de 2021 a 9 sitios con arroyos habitados por A. ordinarium condiferentes grados de perturbación en Michoacán (fig. 1).Los sitios 1, 2 y 3 se encuentran en zonas conservadas en los municipios de Charo, Morelia y Zinápecuaro. En estos sitios, la vegetación está representada por bosque de pino y pino-encino y no se encuentran afectados por la urbanización, cultivos ni zonas de pastoreo.
Los sitios 4, 5 y 6 son urbanizados y se localizan en la Ciudad de Morelia, la cual tiene una extensión de 1,333 km2 y presenta más de 500,000 habitantes (Magaña-Martínez y Reyes Camacho, 2012; tabla 1, fig. 1). Cada sitio se encuentra a una distancia mínima de 3 km uno con respecto a otro, por lo que corresponden a 3 poblaciones independientes de acuerdo a la poca vagilidad de la especie reportada por Montes-Calderón et al. (2011). Se consideraron los sitios como perturbados debido a: 1) la presencia de construcciones urbanas (López- Granados et al., 2008; Magaña-Martínez y Reyes Camacho, 2012), 2) el vertimiento de aguas contaminadas con fertilizantes y pesticidas (López- Granados et al., 2008) y 3) se encuentran a menos de 1 km de avenidas principales de la ciudad de Morelia (Téllez-Ramírez, 2012).
Los sitios 7, 8 y 9 se encuentran en zonas agrícolas en los municipios de Queréndaro, Indaparapeo y Zinapécuaro (fig. 1). Estos sitios presentan arroyos permanentes con escasa vegetación ribereña y se consideran perturbados porque durante el muestreo de este estudio, estaban rodeados de cultivos de maíz y potreros, y no presentaban vegetación de bosque de pino y pino-encino.
En cada arroyo se realizó una búsqueda intensiva de los ejemplares mediante la técnica de inspección por encuentro visual (VES) (Crump y Scott, 1994). Una vez localizados, se capturaron con red de mano y se colocaron en recipientes con agua de su medio para evitar su desecación. Inmediatamente, en el sitio de colecta, se manipuló a cada ejemplar con guantes estériles y se obtuvo una gota de sangre periférica de un pequeño corte de una de las branquias, llevando a cabo el procedimiento sin sacrificar a los individuos. La gota de sangre se colocó en el extremo de un portaobjetos limpio y con ayuda de un segundo, el cual se colocó en un ángulo de 45° por delante de la gota, se lo hizo retroceder hasta tocar la gota, luego se deslizó ejerciendo una presión suave y firme hacia delante. Cada frotis sanguíneo se secó a temperatura ambiente por 3 min y se fijó con metanol. Al final del procedimiento todos los organismos se liberaron en sus respectivos arroyos de origen. El manejo de las salamandras y las muestras se realizó con el permiso de colecta científica número SGPA/DGVS/13339/19 otorgado por la Semarnat.
Los frotis sanguíneos fueron llevados al laboratorio de Parasitología de la Universidad Michoacana de San Nicolás de Hidalgo (UMSNH) y se cubrieron con el colorante no diluido de Wright, dejándose reposar durante 5 min. Posteriormente, se les agregó solución buffer de Wright gota a gota, hasta que apareció una película metálica sobre la muestra, después se dejaron reposar por 5 min. Finalmente, los frotis fueron lavados con agua destilada, hasta que el colorante se lavó y se dejaron secar.
Figura 1. Mapa de la ubicación de los sitios de estudio de A. ordinarium en algunos municipios de Michoacán. 1. 5.7 km al este de Jaripeo, 2. Agua Zarca, 3. 8.9 km al oeste de Bocaneo, 4. Puente campestre, 5. Filtros viejos, 6. Río Chiquito, 7. 14 km al sur de Queréndaro, 8. 0.75 km al sur de Ziróndaro, 9. 10.86 km al sureste del Municipio de Queréndaro.
Tabla 1
Datos de colecta de Ambystoma ordinarium. Se muestra el nombre, las coordenadas y la categoría de los sitios de colecta y el número y talla de los organismos colectados.
Número y nombre de sitio
Coordenadas
Categoría del sitio
Individuos colectados en invierno 2020
Individuos colectados en primavera 2021
Longitud LHC en mm
1. 5.7 km al este de Jaripeo
19°40’28.3” N, 101°01’44.9” O
Conservado
4
3
74 a 77
2. Agua Zarca
19°34’28.9” N, 101°07’28.2” O
Conservado
3
13
66 a 77
3. 8.9 km al oeste de Bocaneo
19°50’28.6” N, 100°43’55.1” O
Conservado
0
13
67 a 88
4. Puente campestre
19°40’31.3” N, 101°09’27.5′′ O
Urbanizado
3
3
79 a 98
5. Filtros Viejos
19°40’01.0” N, 101°08’36” O
Urbanizado
5
8
77 a 91
6. Río Chiquito
19°36’38.2” N, 101°07’26.8” O
Urbanizado
7
5
81 a 101
7. 14 km al sur de Queréndaro
19°41’05.2” N, 100°52’31.2” O
Agricola
5
11
75 a 94
8. 0.75 km al sur de Ziróndaro
19°42’58.4” N, 100°54’59.6” O
Agricola
9
8
67 a 118
9. 10.86 km al sureste del Municipio de Queréndaro
19°45’25.4” N, 100°50’33.9” O
Agricola
4
5
82 a 111
Cada frotis se observó al microscopio óptico con el aumento 400x y se efectuó el recuento diferencial de leucocitos en movimiento zigzag. Las células fueron contadas por una sola persona para evitar la variabilidad en las observaciones considerando las características morfológicas descritas por Thrall (2004), Hadji-Azimi et al. (1987) y Salinas et al. (2017). Siguiendo a Davis et al. (2008), en cada frotis sanguíneo se contaron 100 células, determinándose las proporciones relativas de los 5 tipos de leucocitos y el índice N/L propuesto como indicador de respuesta al estrés. Para la evaluación morfológica de los leucocitos, durante el recuento diferencial, se contaron los leucocitos con cambios en la coloración y presencia de manifestaciones morfológicas en el citoplasma; además, se evaluó la segmentación nuclear de los neutrófilos (neutrófilos banda, identificados por la falta de segmentación en el núcleo).
Para comparar las proporciones y la morfología de cada tipo de leucocito y los índices N/L de los individuos de A. ordinarium entre los sitios conservados, urbanizados y agrícolas, se utilizaron modelos lineales generalizados mixtos (GLM), con error de distribución Poisson debido a que las variables de respuesta son conteos. Los análisis estadísticos se realizaron en R versión 4.2.0 (R Core Team, 2022) y se usó el paquete ggplot2 versión 3.4.1 (Wickham et al., 2016) para la realización de figuras.
Resultados
Se recolectaron 109 individuos de Ambystoma ordinarium en los 3 sitios, 36 en conservados, 31 en urbanizados y 42 en los agrícolas (tabla 1), 40 ejemplares se obtuvieron en invierno de 2020 y 69 en primavera de 2021. El promedio de la longitud total de los ejemplares fue de 82.44 mm (mínima 66-118 máxima) y de acuerdo con las tallas reportadas por Anderson y Worthington (1971), todos los individuos recolectados fueron adultos metamórficos (tabla 1).
Los promedios en las proporciones de los 5 tipos de leucocitos y el índice N/L de los individuos de A. ordinarium para los sitios conservados, urbanizados y agrícolas se presentan en la tabla 2. Las proporciones de linfocitos mostraron diferencias significativas entre los sitios. Se detectaron disminuciones en las proporciones de estas células en individuos de los sitios urbanizados y agrícolas en comparación con los individuos de los hábitats conservados (fig. 2). No se detectaron diferencias en las proporciones de neutrófilos y los índices N/L entre los sitios (tabla 2). Por último, las proporciones de eosinófilos, basófilos y monocitos mostraron diferencias significativas entre los sitios (tabla 2). Se detectaron aumentos en las proporciones de estas células en individuos de los sitios urbanizados y agrícolas con respecto a los individuos de los sitios conservados (fig. 2).
Figura 2. Gráfica de cajas y alambres que muestra las diferencias en las proporciones de leucocitos entre los hábitats urbanizados, agrícolas y conservados. Las letras representan las diferencias de medias entre grupos de acuerdo al GLM.
Los linfocitos, fueron los leucocitos de menor tamaño, la mayoría de éstos se caracterizaron por ser células redondas con un núcleo central que ocupó la mayor parte del citoplasma basófilo (fig. 3). En los frotis sanguíneos de 2 individuos de los hábitas agrícolas se observaron células plasmáticas, caracterizadas por un núcleo excéntrico, citoplasma abundante con aumento en la basofília y abundantes inclusiones globulares y hialinas (fig. 3).
Tabla 2
Promedios relativos de los leucocitos e índices N/L (±error estándar) en individuos adultos de Ambystoma ordinarium entre hábitats conservados, urbanizados y agrícolas.
Variable de respuesta
Hábitats conservados
Hábitats urbanizados
Hábitats agrícolas
gl
c2
p
Linfocitos
83.1 (± 0.51)
72.8 (± 1.20)
74.5 (± 0.93)
2
27.84
< 0.001
Neutrófilos
8.4 (± 0.69)
8.9 (± 0.95)
9.6 (± 0.97)
2
2.84
0.095
Eosinófilos
3.9 (± 0.38)
8.3 (± 1.53)
9.0 (± 0.84)
2
88.37
< 0.001
Basófilos
2.7 (± 0.42)
4.9 (± 0.68)
4.5 (± 0.52)
2
26.66
< 0.001
Monocitos
1.7 (± 0.43)
4.9 (± 1.24)
2.2 (± 0.39)
2
63.26
< 0.001
Neutrófilos banda
1.5 (± 0.24)
3.1 (± 0.60)
3.8 (± 0.48)
2
42.27
<0.001
Índice N/L
0.10 (± 0.00)
0.10 (± 0.011)
0.13 (± 0.01)
2
0.17
0.698
Los neutrófilos se caracterizaron por ser células redondas irregulares que pueden o no presentar finos gránulos en el citoplasma, su núcleo violeta puede no presentar segmentaciones (neutrófilos banda, fig. 3) ó ser segmentados de 2 a 5 lóbulos (fig. 3). En relación con la comparación de la segmentación nuclear de los neutrófilos entre los sitios, se encontraron variaciones significativas, detectándose incrementos en las proporciones de neutrófilos banda en individuos de los sitios urbanizados y agrícolas con respecto a los individuos de los sitios conservados (fig. 3).
Los eosinófilos fueron células con abundantes gránulos rosados cubriendo el citoplasma, presentan un núcleo violeta generalmente bilobulado (fig. 3) y en pocas ocasiones se observó unilobulado y trilobulado. Con respecto de la morfología de los basófilos, éstos se caracterizaron por ser células redondas u ovaladas con abundantes gránulos púrpuras en el citoplasma que, por lo general, cubren el núcleo redondo o bilobulado (fig. 3). Por último, los monocitos fueron los leucocitos de mayor tamaño, son células redondas con núcleo en forma de riñón o herradura y citoplasma abundante (fig. 3). No se observaron variaciones morfológicas en eosinófilos, basófilos y monocitos.
Discusión
Se evaluaron los perfiles de leucocitos en frotis de sangre de 109 individuos adultos de la salamandra Ambystoma ordinarium, de los cuales 37 se recolectaron en hábitats conservados, 30 en hábitats urbanizados y 42 en hábitats agrícolas. La morfología y coloración de los 5 tipos de leucocitos concuerdan con lo reportado para otras especies de anuros y caudados (Cabagna et al., 2005; Hadji-Azimi et al., 1987; Salinas et al., 2017). Los linfocitos fueron los leucocitos más abundantes, detectándose disminuciones en sus proporciones en individuos de A. ordinarium de los sitios urbanizados y agrícolas considerados como perturbados. En estudios recientes, la disminución de los linfocitos en sangre se ha documentado en especies de anfibios como respuesta a factores estresantes como infecciones y hábitats contaminados con pesticidas, debido a que el aumento en las hormonas del estrés (glucocorticoides) puede inducir la salida de estas células de la sangre a los tejidos linfoides (en anfibios, hígado y bazo) (Davis et al., 2008; Waye et al., 2019). En contraste, los neutrófilos por ser las principales células encargadas del ataque a agentes infecciosos son estimulados a proliferar para migrar a los sitios de inflamación. La disminución en las proporciones de los linfocitos se ha relacionado con el aumento en las proporciones de los neutrófilos (índice N/L) como biomarcador de estrés (Davis et al., 2010). En este estudio, no se detectaron variaciones significativas en las proporciones de neutrófilos y en los índices N/L promedio. En estudios previos en poblaciones silvestres de salamandras del género Ambystoma y otras especies de anfibios, se han reportado índices N/L promedio cercanos a 0.40 (Cabagna et al., 2005; Davis y Durso, 2009; Shutler et al., 2009). Sin embargo, a diferencia de nuestro estudio, Ramírez-Hernández et al. (2019) reportaron para esta especie un índice N/L promedio de 1.5. y 0.9 en hábitats perturbados y conservados, respectivamente. Las diferencias entre ambos resultados podrían deberse a que el tamaño de muestra utilizado por Ramírez-Hernández et al. (2019) fue pequeño en comparación con el tamaño de muestra utilizado en nuestro estudio. Además, nuestros resultados pueden indicar que otros factores no contabilizados están influyendo en las respuestas de estas células en los individuos muestreados. Un factor que aumenta el índice linfocitos y neutrófilos en anfibios es la infección por Batrachochytrium dendrobatidis (Bd; Davis et al., 2010; Savage et al., 2016). Recientemente, se reportó la presencia del hongo quitridio en las mismas poblaciones de A. ordinarium analizadas en este estudio (Mendoza-Almeralla et al., 2023). Los niveles de infección por Bd reportados fueron de entre 112 a 1,856 equivalentes zoosporas genómicas (EZG´s) en 2 sitios conservados; mientras que en los sitios perturbados, el grado de infección fue de 21 a 4,338 EZG´s, ésto sugiere que hay mayor grado de infección en lugares perturbados. Por lo que es importante saber si el número de neutrófilos y linfocitos es afectado por la infección del quitridio.
Figura 3. Leucocitos en frotis de sangre periférica de A. ordinarium. A) Linfocito, B) neutrófilo segmentado, C) neutrófilo banda, D) eosinófilo, E) basófilo, D) monocito, vista a 100X, G) célula plasmática con núcleo excéntrico y citoplasma abundante, vista a 40X.
Por otro lado, se detectaron incrementos de neutrófilos banda en individuos de los sitios perturbados con respecto a los sitios conservados. En procesos inflamatorios, los neutrófilos son impulsados a proliferar y para compensar esta demanda, la médula ósea libera en la sangre células inmaduras (neutrófilos banda) (Davis y Golladay, 2019). En anfibios existe evidencia de incrementos en las proporciones de estas células como respuesta inflamatoria contra fertilizantes y pesticidas (Mann et al., 2009; Romanova et al., 2022). Pese a que en este estudio no evaluamos la presencia de estos contaminantes químicos en el agua, en los sitios urbanizados seleccionados aquí, se han reportado descargas de aguas contaminadas con fertilizantes y pesticidas (López-Granados et al., 2008), y en los sitios agrícolas es probable que el uso de estos agroquímicos sea habitual y su dispersión fuera de estas áreas pueda llegar hasta los arroyos donde habita esta especie. Los incrementos detectados en las proporciones de eosinófilos, basófilos y monocitos en los individuos de los sitios urbanizados y agrícolas también se pueden asociar con la presencia de estos contaminantes químicos en sus hábitats. En algunos estudios en anfibios se han reportado incrementos en las proporciones de eosinófilos en sitios contaminados por pesticidas por su capacidad de reaccionar a antígenos ambientales (Attademo et al., 2014; Romanova y Romanova, 2003). Con respecto a los monocitos, el aumento de estas células podría relacionarse con el incremento de la fagocitosis de desechos tisulares, puesto que, de acuerdo con algunos autores la exposición prolongada a contaminantes químicos aumenta la necrosis tisular (Zhelev, 2007). El papel de los basófilos en las respuestas inmunitarias de los anfibios no es claro (Allender y Fry, 2008), sin embargo, al igual que en otros grupos de vertebrados parecen desempeñar un papel importante en la inflamación (Claver y Quaglia, 2009).
Los estudios realizados por varios investigadores han revelado que los anfibios son capaces de generar una respuesta inmunitaria a antígenos complejos asociados con patógenos y antígenos ambientales (Grogan et al., 2018; Savage y Zamudio, 2011; Zhelev, 2007). Los linfocitos (células T y B), ante la presencia de antígenos son responsables de activar la inmunidad mediada por células específicas de patógenos (células T citotóxicas o auxiliares) y la inmunidad humoral (células plasmáticas secretoras de anticuerpos específicos) (Grogan et al., 2018). El hallazgo de células plasmáticas en la sangre de los individuos de los sitios agrícolas podría atribuirse a la especificidad de la respuesta inmune humoral a antígenos tóxicos presentes en el ambiente. En un estudio previo, Zhelev (2007) reportó aumentos de estas células en individuos de Rana ridibunda en hábitats con contaminación industrial. El hallazgo de estas células es interesante, porque casi no hay datos en la literatura sobre su presencia y apariencia en la sangre de los anfibios. Aunque no podemos asegurar que se trate de este tipo de células, estos datos sin duda sientan las bases para desarrollar estudios bioquímicos e inmunológicos futuros.
En anfibios se ha documentado la función antipara- sitaria de los eosinófilos en infecciones con nemátodos y tremátodos (Belden y Kiesecker, 2005; Davis y Golladay, 2019; Kiesecker, 2002; Rohr et al., 2008). Particularmente, Ramírez-Hernández et al. (2019) reportaron aumentos en las cargas parasitarias en poblaciones de A. ordinarium por 2 especies de tremátodos, Gorgoderina attenuata y Ochetosoma sp.,y 2 especies de nemátodos Cosmocercoides sp.y Hedruris siredonis en hábitats perturbados, siendo uno de estos hábitats correspondiente al sitio Río Chiquito de este estudio. Adicionalmente, Mendoza-Almeralla et al. (2023) reportaron la infección por el nemátodo del género Capillaria en el sitio Filtros viejos. Por lo tanto, los incrementos de eosinófilos detectados en los individuos de estos sitios, podrían relacionarse con la presencia de mayor prevalencia de infecciones parásitarias.
Finalmente, los perfiles de leucocitos de los individuos de A. ordinarium en este estudio proporcionan parámetros hematológicos de comparación entre distintas poblaciones. De acuerdo con nuestros resultados, la evaluación de los perfiles de leucocitos es uno de los métodos más simples y menos invasivos. Los cambios en sus valores, especialmente en las poblaciones de hábitats perturbados, pueden utilizarse con éxito en evaluaciones futuras para detectar cambios fisiológicos y patológicos tempranos en los individuos y puede ser una señal de advertencia de degradación ambiental. Sin embargo, es importante reiterar que interpretar las proporciones de los leucocitos puede ser complicado, debido a que los leucocitos pueden responder a diversos factores (Barni et al., 2007; Cabagna et al., 2005; Romanova y Romanova, 2003; Shutler y Marcogliese, 2011; Shutler et al., 2009). Por ello es necesario llevar a cabo estudios complementarios sobre enfermedades infecciosas, calidad del agua, niveles de hormonas de estrés, presencia de pesticidas o metales pesados, entre otros, que permitan relacionar y estudiar la respuesta de los leucocitos en las poblaciones de esta especie y otras especies de anfibios con los diversos factores o contextos en los que se encuentran estas especies.
Agradecimientos
Este estudio fue parte del proyecto “Descifrando el microbioma de la piel en ajolotes y las consecuencias de la interacción huésped microbioma sobre una enfermedad letal emergente” de la Secretaría de Educación Pública/Consejo Nacional de Humanidades, Ciencias y Tecnologías Ciencias de Frontera. FORDECYT-PRONACES/373914/2020. Los resultados de este estudio forman parte de la tesis de maestría del autor principal, bajo la dirección de ISO y CMA. CGP agradece el apoyo financiero del Programa Nacional de Becas de SEP/Conahcyt.
Referencias
Allender, M. C. y Fry, M. M. (2008). Amphibian hematology. Veterinary Clinics of North America: Exotic Animal Practice, 11, 463–480. https://doi.org/10.1016/j.cvex.2008.03.006
Alvarado, J. G. A. (2021). Anfibios en peligro: amenazas y estrategias efectivas de conservación. Biocenosis, 32, 3–45.https://doi.org/10.22458/rb.v32i1.3552
Anderson, J. D. y Worthington, R. D. (1971). The life history of the Mexican salamander Ambystoma ordinarium Taylor. Herpetologica, 27, 165–176.
Attademo, A. M., Peltzer, P. M., Lajmanovich, R. C., Cabagna-Zenklusen, M. C., Junges, C. M. y Basso, A. (2014). Biological endpoints, enzyme activities, and blood cell parameters in two anuran tadpole species in rice agroecosystems of mid-eastern Argentina. Environmental Monitoring and Assessment, 186, 635–649. https://doi.org/10.1007/s10661-013-3404-z
Barni, S., Boncompagni, E., Grosso, A., Bertone, V., Freitas, I., Fasola, M. et al. (2007). Evaluation of Rana snk esculenta blood cell response to chemical stressors in the environment during the larval and adult phases. Aquatic Toxicology, 81, 45–54. https://doi.org/10.1016/j.aquatox.2006.10.012
Barriga-Vallejo, C., Hernández-Gallegos, O., Von-Herbing, I. H., López-Moreno, A. E., Ruiz-Gómez, M. D. L., Granados-González, G. et al. (2015). Assessing population health of the Toluca Axolotl Ambystoma rivulare (Taylor, 1940) from México using leukocyte profiles. Herpetological Conservation and Biology, 10, 592–601.
Belden, L. K. y Kiesecker, J. M. (2005). Glucocorticosteroid hormone treatment of larval treefrogs increases infection by Alaria sp. trematode cercariae. Journal of Parasitology, 91, 686–688. https://doi.org/10.1645/GE-397R.
Cabagna, M. C., Lajmanovich, R. C., Stringhini, G., Sánchez-Hernández, J. C. y Peltzer, P. M. (2005). Hematological parameters of health status in the common toad Bufo arenarum in agroecosystems of Santa Fe Province, Argentina. Applied Herpetology, 2, 373–380. https://doi.org/10.1163/157075405774483085
Claver, J. A. y Quaglia, A. I. (2009). Comparative morphology, development, and function of blood cells in nonmammalian vertebrates. Journal of Exotic Pet Medicine, 18, 7–97. https://doi.org/10.1053/j.jepm.2009.04.006
Crump, M. L. y Scott, N. Y. (1994). Visual encounter surveys. En W. Heyer, M. A. Donnelley, R. A. Mcdiarmid, L. C. Hayek. y M. C. Foster (Eds.), Measuring and monitoring biological diversity: standard methods for amphibians (pp. 84–92). Washington D.C.: Smithsonian Institution.
Das, M. y Mahapatra, P. K. (2014). Hematology of wild caught Dubois’s tree frog Polypedates teraiensis, Dubois, 1986 (Anura: Rhacophoridae). The Scientific World Journal, 491415, 7. https://doi.org/10.1155/2014/491415
Davis, A. K. y Durso, A. M. (2009). White blood cell differentials of northern cricket frogs (Acris c. crepitans) with a compilation of published values from other amphibians. Herpetologica, 65, 260–267. https://doi.org/10.1655/08-052R1.1
Davis, A. K. y Golladay, C. (2019). A survey of leukocyte profiles of red-backed salamanders from Mountain Lake, Virginia, and associations with host parasite types. Comparative Clinical Pathology, 28, 1743–1750. https://doi.org/10.1007/s00580-019-03015-9
Davis, A. K., Maney, D. L. y Maerz, J. C. (2008). The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists. Functional Ecology, 22, 760–772. https://doi.org/10.1645/GE-397R
Davis, A. K., Keel, M. K., Ferreira, A. y Maerz, J. C. (2010). Effects of chytridiomycosis on circulating white blood cell distribu- tions of bullfrog larvae (Rana catesbeiana). Comparative Clinical Pathology, 19, 49–55. 10.1007/s00580-009-0914-8
Duellman, W. E. y L. Trueb. (1994). The biology of amphibians. Baltimore, Maryland: Johns Hopkins University Press.
Grogan, L. F., Robert, J., Berger, L., Skerratt, L. F., Scheele, B. C., Castley, J. G. et al. (2018). Review of the amphibian immune response to chytridiomycosis, and future directions. Frontiers in Immunology, 9, 2536. https://doi.org/10.3389/fimmu.2018.02536
Hadji-Azimi, I., Coosemans, V. y Canicatti, C. (1987). Atlas of Xenopus laevis laevis hematology. Developmental and Comparative Immunology, 11, 807–874.
Kiesecker, J. M. (2002). Synergism between trematode infection and pesticide exposure: a link to amphibian deformities in nature? Proceedings of the National Academy of Sciences, 99, 9900–9904. https://doi.org/10.1073/pnas.152098899
Lips K. R., Burrowes, P. A., Mendelson, J. R. y Parra-Olea, G. (2005). Amphibian population declines in Latin America: a synthesis. Biotropica, 37, 222–226. https://doi.org/10.1111/j.1744-7429.2005.00029.x
López-Granados, E., Mendoza-Cantú, M., Bocco, G. y Espinosa-Bravo, M. (2008). Patrones de degradacion ambiental en la Cuenca del Lago de Cuitzeo, Michoacán. Una perspectiva espacial. Centro de Investigaciones en Ecosistemas, UNAM/Morelia, Michoacán, Instituto de Geografía, UNAM/ Dirección General de Investigación de Ordenamiento Ecológico y Conservación de los Ecosistemas, INIFAP.
Magaña-Martínez, H. M. y Reyes-Camacho, E. (2012). Parque lineal ecológico los filtros viejos en Morelia, Mich. (Tesina). Facultad Arquitectura, Universidad Michoacana de San Nicolás de Hidalgo. Morelia, Michoacán.
Mann, R. M., Hyne, R. V., Choung, C. B. y Wilson, S. P. (2009). Amphibians and agricultural chemicals: review of the risks in a complex environment. Environmental Pollution, 157, 2903–2927. https://doi.org/10.1016/j.envpol.2009.05.015
Mendoza-Almeralla, C., Tafolla-Venegas, D., González-Pardo, C. y Suazo- Ortuño, I. (2023). Primer registro de infección por Batrachochytrium dendrobatidis y por el nemátodo del género Capillaria y la ausencia de infección por Ribeiroia ondatrae en Ambystoma ordinarium. Revista Latinoamericana de Herpetología, 6, e615-05. https://doi.org/10.22201/fc.25942158e.2023.4.615
Montes-Calderón, A. M., Alvarado-Díaz, J. y Suazo-Ortuño, I. (2011). Abundancia, actividad espacial y crecimiento de Ambystoma ordinarium Taylor 1940 (Caudata: Ambys- tomatidae) en Michoacán, México. Revista Biológicas, 13, 50–53.
Ramírez-Bautista, A., Torres-Hernández, L. A., Cruz-Elizalde, R., Berriozabal-Islas, C., Hernández-Salinas, U., Wilson, L. D. et al. (2023). An updated list of the Mexican herpetofauna: with a summary of historical and contemporary studies. Zookeys, 1166, 287. https://doi.org/10.3897/zookeys.1166.86986
Ramírez-Hernández, G., Suazo-Ortuño, I., Alvarado-Díaz, J., Escalera-Vázquez, L. H., Maldonado-López, Y. y Tafolla-Venegas, D. (2019). Effects of habitat disturbance on parasite infection and stress of the endangered Mexican stream salamander Ambystoma ordinarium. Salamandra, 55, 160–172.
R Core Team. (2022). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Recuperado el 01 junio, 2023 de: https://www.R-project.org/
Rohr, J. R., Schotthoefer, A. M., Raffel, T. R., Carrick, H. J., Halstead, N., Hoverman, J. T. et al. (2008). Agrochemicals in- crease trematode infections in a declining amphibian species. Nature, 455, 1235–1239. https://doi.org/10.1038/nature07281
Romanova, E. B. y Romanova, O. Y. (2003). Peculiarities of leukocytic formula of peripheral blood of green frogs under conditions of anthropogenic load. Journal of Evolutionary Biochemistry and Physiology, 39, 480–484. https://doi.org/10.1023/B:JOEY.0000010246.27310.85
Romanova, E. B., Stolyarova, I. A., Bakiev, A. G. y Gorelov, R. A. (2022). The leukocyte blood composition of Emys orbicularis and Mauremys caspica (Reptilia: Testudines: Emydidae, Geoemydidae) at syntopy. Biology Bulletin, 49, 1923–193. https://doi.org/10.35885/1684-7318-2022-1-79-93
Salinas, Z. A., Salas, N. E., Baraquet, M. y Martino, A. L. (2015). Biomarcadores hematológicos del sapo común Bufo (Rhinella) arenarum en ecosistemas alterados de la provincia de Córdoba. Acta Toxicológica Argentina, 23, 25–35.
Salinas, Z. A., Baraquet, M., Grenat, P. R., Martino, A. L. y Salas, N. E. (2017). Morphology and size of blood cells of Rhinella arenarum (Hensel, 1867) as environmental health assessment in disturbed aquatic ecosystem from central Argentina. Environmental Science and Pollution Research, 24, 24907–24915.
Salinas, Z. A., Babini, M. S., Grenat, P. R., Biolé, F. G., Martino, A. L. y Salas, N. E. (2019). Effect of parasitism of Lernaea cyprinacea on tadpoles of the invasive species Lithobates catesbeianus. Heliyon, 5, 6. https://doi.org/10.1016/j.heliyon. 2019.e01834
Savage, A. E. y Zamudio, K. R. (2011). MHC genotypes associate with resistance to a frog-killing fungus. Proceedings of the National Academy of Sciences, 108, 16705–16710. https://doi.org/10.1073/pnas.1106893108
Savage, A. E., Terrell, K. A., Gratwicke, B., Mattheus, N. M., Augustine, L. y Fleischer, R. C. (2016). Reduced immune function predicts disease susceptibility in frogs infected with a deadly fungal pathogen. Conservation Physiology, 4, cow011. https://doi.org/10.1093/conphys/cow011
Semarnat (Secretaría del Medio Ambiente y Recursos Naturales). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental – Especies nativas de México de flora y fauna silvestres – Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio – Lista de especies en riesgo. Diario Oficial de la Federación. 30 de diciembre de 2010, Segunda Sección, México.
Shutler, D. y Marcogliese, D. J. (2011). Leukocyte profiles of northern leopard frogs, Lithobates pipiens, exposed to pesticides and hematozoa in agricultural wetlands. Copeia, 2, 301–307. https://doi.org/10.1643/CP-10-065
Shutler, D., Smith, T. G. y Robinson, S. R. (2009). Relationships between leukocytes and Hepatozoon spp. in green frogs, Rana clamitans. Journal of Wildlife Diseases, 45, 67–72. https://doi.org/10.7589/0090-3558-45.1.67
Soto Rojas, C. (2012). Uso y selección del microhábitat de la salamandra de montaña Ambystoma ordinarium (Tesis de maestría). Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán.
Stuart, S. N., Chanson, J. S., Cox, N. A., Young, B. E., Rodrigues, A. S., Fischman, D. L. et al. (2004). Status and trends of amphibian declines and extinctions worldwide. Science, 306, 1783-1786. https://doi.org/10.1126/science.1103538
Tellez-Ramirez, K. L. (2012). Programa de revitalización del Área Natural Protegida de los Filtros Viejos. Monografía para obtener el título de arquitecto. Universidad Vasco de Quiroga, Morelia, Michoacán.
Thrall, M. A. (2004). Hematology of reptiles. EnD. B. Baker, T. C. Campbell, D. DeNicola, M. J. Fettman, E. D. Lassen, A. Rebar et al. (Eds.), Veterinary hematology and Clinical Chemistry: text and clinical case presentations. Philadelphia: Lippincott Williams y Wilkins.
Wake, D. B. y Vredenburg, V. T. (2008). Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proceedings of the National Academy of Sciences, 105, 11466–11473. https://doi.org/10.1073/pnas. 0801921105
Waye, H. L., Dolan, P. C. y Hernández, A. (2019). White blood cell profiles in long-term captive and recently captured eastern tiger salamanders (Ambystoma tigrinum). Copeia, 107, 138–143. https://doi.org/10.1643/CP-18-126
Wickham, H., Chang, W. y Wickham, M. H. (2016). Package ‘ggplot2’. Create elegant data visualizations using the grammar of graphics, Version 2, 1–189.
Zhelev, Z. M. (2007). Investigation on the blood differential formula in Rana ridibunda (Anura, Amphibia) from the Area of the Maritsa-Iztok 1 Steam Power Plant. Acta Zoologica Bulgarica, 59, 181–190.