
Estructura de la comunidad de Thomisidae (Araneae, Araneomorphae) en Tepoztlán, Morelos, México: estudio estacional de un año
Fernando Pilo-García a, Víctor H. Toledo-Hernández b, *, Alejandro Valdez-Mondragón c
a Universidad Autónoma del Estado de Morelos, Facultad de Ciencias Biológicas, Av. Universidad 1001, Colonia Chamilpa, 62209 Cuernavaca, Morelos, Mexico
b Universidad Autónoma del Estado de Morelos, Centro de Investigación en Biodiversidad y Conservación, Av. Universidad 1001, Colonia Chamilpa, 62209 Cuernavaca, Morelos, Mexico
c Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), Laboratorio de Aracnología, Centro Tlaxcala de Biología de la Conducta (CTBC), Universidad Autónoma de Tlaxcala, Mexico
*Corresponding author: victor.toledo@uaem.mx (V.H. Toledo-Hernández)
Received: 20 August 2025; accepted: 12 February 2026
Abstract
Spiders of the family Thomisidae are scarcely studied in Mexico, and ecological data for this family are practically nonexistent. To fill this gap, we carried out an ecological study of crab spiders in a deciduous tropical forest in San Andrés de la Cal, Tepoztlán, Morelos, central Mexico, evaluating seasonal variation in community structure over the annual cycle. Fieldwork consisted of systematized monthly diurnal collections by 5 people, implementing 3 sampling techniques: beating sheets, sweeping nets, and looking down. In total, we collected 839 spiders representing 9 genera and 16 species, of which 5 genera and 7 species are new state records. The highest richness, abundance, and diversity were recorded in the rainy season; however, unique species were found during each season. According to richness estimators, between 12 and 22% of the estimated species remain to be found in the study area. Evenness factors indicated that the community is composed primarily of rare species, with only a few dominant species. While this study reports a reliable inventory for the region, the species accumulation curve did not reach the asymptote, suggesting that additional ecological studies employing diverse protocols are needed to complement our knowledge of these spiders.
Keywords: Crab spiders; Deciduous tropical forest; Ecology; Diversity; Seasonality
Resumen
En México, la familia Thomisidae está poco estudiada y los datos ecológicos son prácticamente inexistentes. Para subsanar esta deficiencia, realizamos un estudio ecológico de arañas cangrejo en un bosque tropical caducifolio en San Andrés de la Cal, Tepoztlán, Morelos, centro de México, evaluando la variación estacional en la estructura de la comunidad durante un ciclo anual. El trabajo de campo consistió en colectas diurnas mensuales sistematizadas por 5 personas, implementando 3 técnicas de muestreo: paraguas entomológico, red entomológica y búsqueda directa. En total, recolectamos 839 individuos pertenecientes a 9 géneros y 16 especies, de los cuales 5 géneros y 7 especies son nuevos registros estatales. La mayor riqueza, abundancia y diversidad se registraron durante la temporada de lluvias; sin embargo, se registraron especies únicas en cada temporada. Según los estimadores de riqueza, faltan por registrarse en el área de estudio entre 12 y 22% de las especies estimadas. Los factores de uniformidad indicaron que la comunidad está compuesta principalmente por especies raras y pocas especies dominantes. El inventario obtenido es confiable; no obstante, la curva de acumulación no alcanzó la asíntota, sugiriendo la necesidad de realizar estudios ecológicos adicionales empleando diversos protocolos para complementar nuestro conocimiento de estas arañas.
Palabras clave: Arañas cangrejo; Selva baja caducifolia; Ecología; Diversidad; Estacionalidad
Introduction
In general, most studies on spider communities in Mexico have analyzed spatial diversity patterns, with fewer studies focusing on seasonal diversity and abundance (Campuzano & Padilla-Ramírez, 2020; Corcuera et al., 2019; Jiménez et al., 2020, 2024; Menéndez-Acuña et al., 2023; Pérez-Hernández & Zaragoza-Caballero, 2016; Rodríguez-Rodríguez et al., 2015). The study of seasonal diversity variation and its associated processes is crucial for understanding ecosystem dynamics and establishing effective conservation strategies. Moreover, understanding higher-taxon diversity can serve as a useful indicator of overall biodiversity (Williams & Gaston, 1994). Basic ecological information is lacking for many megadiverse biological groups, including spiders (Colwell & Coddington, 1994). In Mexico, 2,345 spider species have been recorded, comprising 455 genera and 70 families (Ponce-Saavedra et al., 2023). Despite this high diversity, only 2 country-level inventories of Mexican spiders have been compiled (Hoffmann, 1976; Jiménez, 1996), and it has been estimated that only ~ 23% of the country’s spider species are known (Francke, 2014; Ponce-Saavedra et al., 2023). Spiders are suitable model organisms for ecological studies due to the ease of obtaining large samples for statistical analyses, their limited dispersal capacity, their presence in almost every ecosystem, and their high abundance (Colwell & Coddington, 1994; Halaj et al., 1998; Jiménez-Valverde & Hortal, 2003; Turnbull, 1973). However, when generating a species inventory for a megadiverse group, encountering every species present at a certain locality takes considerable effort due to the existence of rare species, the wide variety of collecting techniques needed, and the difficulty to systematize sampling protocols (Gotelli & Cowell, 2011; Wolda, 1988). Therefore, it is often beneficial to focus sampling efforts on smaller taxonomic groups (families, subfamilies, tribes), facilitating the standardization of protocols and the increased capacity to detect a higher proportion of the overall species number (Jiménez-Valverde & Lobo, 2004, 2005, 2007). Crab spiders are an abundant and diverse group, one of the dominant spider families in the community. They are an important link in trophic chains as both predators and prey, and due to their proximity to the vegetation, these spiders are susceptible to changes in the environment (Benjamin, 2011; Benjamin et al., 2008; Clausen, 1986; Jocqué & Dippenaar-Schoeman, 2006; Lapp & Dondale, 2017; Morse, 2007). In addition, they are easy to identify in the field, making them a useful model for ecological studies and reliable regional faunistic inventories. In the Mexican state of Morelos, the first record of the family Thomisidae was made by F. O. Pickard-Cambridge (1895), who described Synema madidum from Cuernavaca. In the 128 years since then, only 5 species have been reported in 7 scientific papers (Pickard-Cambridge, 1895, 1900; Gertsch, 1953; Gertsch & Davis, 1940; Hoffmann, 1976; Nieto-Castañeda et al., 2014; Pickard-Cambridge, 1896). Similarly, ecological data for the family has been reported in only a few studies, which mostly focus on higher taxonomic levels (Hernández-Silva, 2016; Menéndez-Acuña, 2017; Nieto-Castañeda et al., 2014; Rivas-Herrera, 2015). To gather basic ecological data and initiate faunistic inventories for the family Thomisidae, we analyzed the community structure of crab spiders throughout an annual cycle. By comparing community structure across the rainy and dry seasons in a deciduous tropical forest, we aim to uncover temporal patterns in thomisid diversity and to contribute to the faunistic knowledge of these spiders in Morelos and Mexico more broadly.
Materials and methods
The study was carried out in the surroundings of San Andrés de la Cal, Tepoztlán, Morelos, in central Mexico (Fig. 1). The study area pertains to the protected areas Corredor Biológico Chichinautzin and Parque Nacional El Tepozteco (Conanp, 1983; INEGI, 2017), located at 18°56’11” N, 99°06’46” W, with an average altitude of 1,507 m asl (Fig. 1). The climate is characterized as (A) Cw2 (W) ig, warm subhumid, with summer rains (García, 2004). The mean annual temperature is 20.5 °C, and the mean annual precipitation is 1,091.8 mm (INEGI, 2017; Ruiz-Rivera, 2001). The dominant primary vegetation is deciduous tropical forest, characterized by trees under 12 meters in height and 2 marked seasons, a rainy season (June to October) and a dry season (November to May) (Balvanera & Maass, 2010; Miranda & Hernández, 1963; Trejo, 2010). Two study sites were selected: “Derrame de Lava” (DL), at 18°56’33.5” N, 99°06’57.4” W, and “Cerro de la Cal” (CC), at 18°56’38” N, 99°06’31” W (Fig. 1). Both sites have specific geomorphological characteristics that generate differences in the composition of woody plants. Sixty species of woody plants are known across both sites. In DL, 24 species are reported only for this site, with Sapium macrocarpum Müll. Arg. (Euphorbiaceae), Ipomea pauciflora M. Martens & Galeotti (Convolvulaceae), and Quercus obtusata Bonpl. (Fagaceae) predominant. In CC, 8 species were observed only at this site, with S. macrocarpum, Bursera fagaroides (Kunth) Engl., B. glabrifolia (Kunth) Engl., and B. copallifera (DC.) Bullock (Burseraceae) predominant (Vergara-Torres et al., 2010).
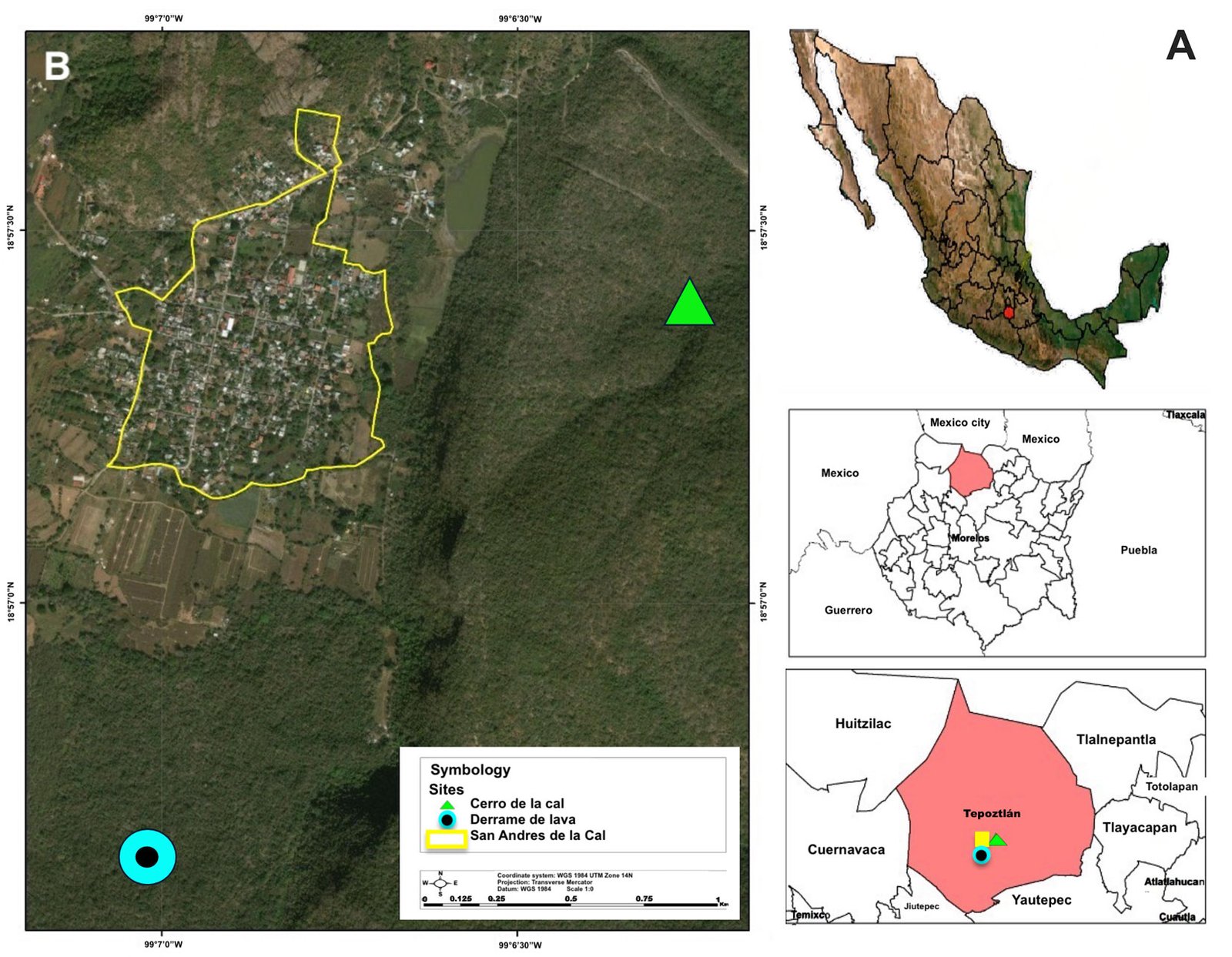
Spiders of the family Thomisidae were collected over a one-year period, from September 2017 to August 2018. A systematic collection protocol was implemented, involving 2 days per month at each site and 5 collectors. Because these spiders are diurnal, collections were made over 6 daylight hours per day, from 10:00 to 16:00 h (Abraham, 1983), with an effort of 6 hours per person. We applied 3 manual sampling techniques: sweeping nets, beating sheets, and looking down (Jiménez-Valverde & Lobo, 2005). Transects were made ad libitum without repeating sampled sections in an area of approximately 2 km². The collected spiders were placed in glass jars containing 96% ethanol and labeled with locality, date, geographical coordinates, altitude, and collectors’ names. The spiders were identified at the Colección de Insectos de la Universidad de Morelos (CIUM), CIByC, UAEM, Cuernavaca, and deposited at the Colección Nacional de Arácnidos (CNAN), IB-UNAM, Mexico City. Identifications were initially conducted at family and genus levels using the keys from Ubick et al. (2017) and Lapp and Dondale (2017), respectively. Only adult specimens were considered for species-level identification (male palps and female epigynum), using specialized literature (Pickard-Cambridge, 1900; Gertsch, 1953; Gertsch & Davis, 1940; Jiménez, 1986, 1992; Pickard-Cambridge, 1896). Adult spiders that could not be determined to genus were assigned as Indeterminate Genus (In. Gen.); those not assignable to species were treated as morphospecies. Species richness and abundance were quantified as the number of species and individuals, respectively. Alpha diversity was calculated using EstimateS v.9.1 (Colwell, 2013), following the true diversity concept of Jost (2006), with Hill numbers for q = 0, 1, 2: order 0 (⁰D) considers only species richness, order 1 (¹D) weights species by proportional frequency, and order 2 (²D) favors abundant species (Jost & González-Oreja, 2012). A dominance-diversity curve was plotted to visually evaluate community structure, and evenness factors were calculated (Jost, 2010). We conducted a χ² test to assess the relationships between adult and immature individuals and between males and females, using R (R Core-Team, 2024). A species accumulation curve was constructed with iNEXT (Chao et al., 2016) following Chao et al. (2014). Finally, 4 non-parametric richness estimators (Chao 1, Chao 2, first- and second-order Jackknife) were computed using EstimateS v.9.1 (Colwell, 2013).
Results
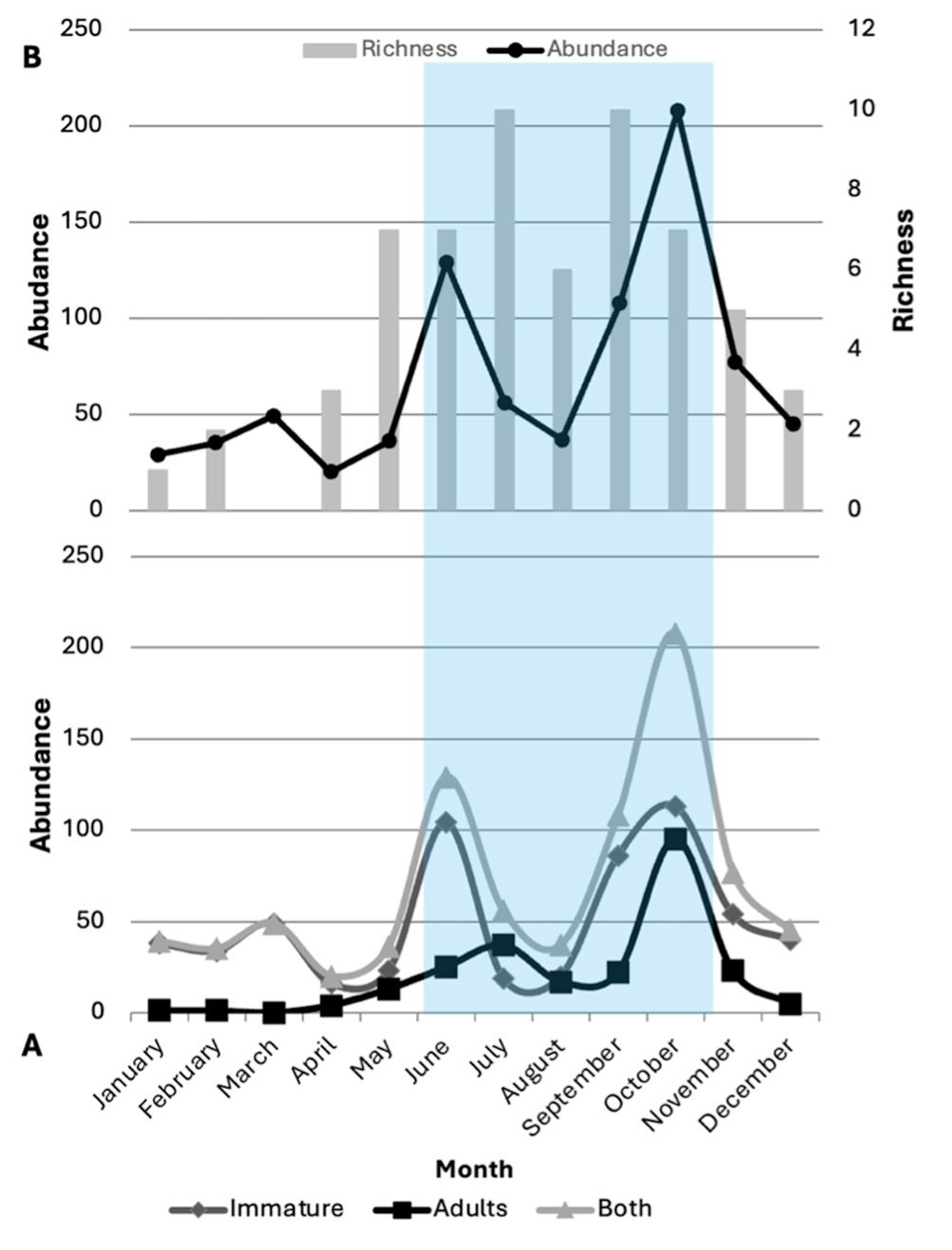
A total of 16 species comprising 11 genera of crab spiders were recorded (Figs. 2-26, Table 1). All species captured represent new records for San Andrés de la Cal, and 9 species and 5 genera are new records for Morelos (Table 1). The genus with the highest richness was Misumenoides F. O. Pickard-Cambridge, 1904, with 4 species (26%), followed by Tmarus Simon, 1875, and Mecaphesa Simon, 1900, each with 2 species. Only 1 species was recorded for all other genera (Table 1). Regarding seasonal richness, a marked seasonality was observed, with higher richness during the rainy season (S = 14). The highest richness was in July and September, with 10 species each (Fig. 26B). In contrast, the dry months showed lower richness (S = 10), with the lowest in January and March, with only 1 species each (Fig. 26B). Eight species were recorded in both seasons, while 6 were found only in the rainy season and 2 only in the dry season (Table 1). Of the 16 species, 31% were found only during 1 or 2 sampled months, while the dominant species was recorded in 9 of the 12 months (Table 1, Fig. 27). A total of 839 individual spiders were collected, with 596 immatures and 243 adults (149 males and 94 females). The highest abundance was recorded in October (N = 208), with 113 immatures and 95 adults, whereas April had the lowest abundance (N = 20), with 16 immatures and 4 adults (Fig. 26A, Table 1). Statistical differences were found between the proportions of adults and immatures over the 12-month sampling period (χ² = 161.11, f. g.= 11, p < 0.0001), as well as between females and males (χ² = 36.768, f. g.= 10, p < 0.0005).
| Month / species | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bucranium aff affinis° | – | 1 | – | 1 | 1 | – | – | – | – | – | – | – | 3 |
| In. Gen. | – | – | – | – | – | – | 13 | 12 | 1 | 1 | – | 1 | 28 |
| In. Gen. 2 | – | – | – | – | – | – | – | 1 | – | – | – | – | 1 |
| Isaloides sp. 1° | – | – | – | – | 1 | 3 | 2 | 1 | – | – | – | – | 7 |
| Mecaphesa dubia* | 1 | – | – | 2 | 2 | 4 | 4 | – | 17 | 78 | 11 | 5 | 124 |
| Mecaphesa rothi* | – | – | – | – | – | – | – | 1 | – | – | – | – | 1 |
| Misumenoides annulipes* | – | – | – | – | – | – | 1 | 2 | 2 | 3 | – | – | 8 |
| Misumenoides formosipes* | – | – | – | 3 | – | 1 | – | 1 | 3 | 1 | – | – | 9 |
| Misumenoides quetzaltocatl* | – | – | – | – | – | – | – | 1 | 1 | – | – | – | 2 |
| Misumenoides sp. 1 | – | – | – | – | – | 7 | 1 | 4 | – | 1 | – | – | 13 |
| Ozyptila sp. 1° | – | 1 | – | – | – | – | – | – | – | – | – | – | 1 |
| Parasynema cirripes | – | – | – | – | – | 2 | – | 3 | 1 | – | – | – | 6 |
| Synema lopezi* | – | – | – | – | 1 | 1 | 2 | 0 | 1 | 3 | 1 | – | 9 |
| Tmarus ehecatltocatl* | – | – | – | – | 4 | 5 | 2 | – | 1 | 4 | 3 | 1 | 20 |
| Tmarus vitusus* | – | – | – | 1 | 1 | 1 | 2 | – | – | – | – | – | 5 |
| Xysticus facetus | – | – | – | – | – | 5 | 1 | – | – | – | – | – | 6 |


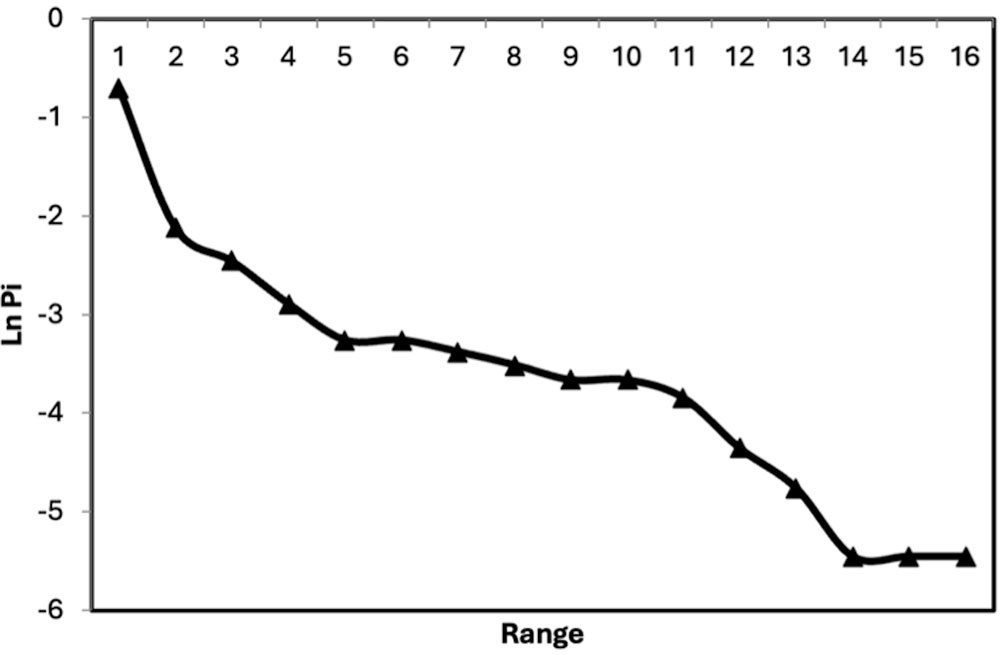
Regarding true diversity, the order-one diversity (¹D) showed that the community was dominated by a few abundant species, with a peak diversity of 5.91 effective species in May, and 3.26 effective dominant species (²D) with a peak diversity of 5.12 effective dominant species in May (Table 2).
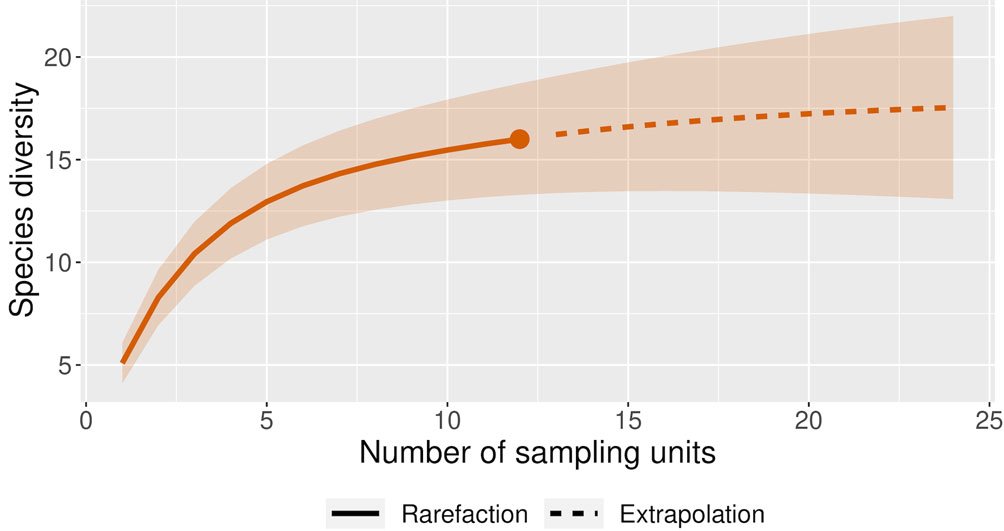
The species accumulation curve showed continuous growth without reaching an asymptote, indicating that the true number of species in the study area is greater than what we recorded (Fig. 28). According to the richness estimators (Chao 1, Chao 2, first-order Jackknife, and second-order Jackknife), a richness of 18-20 species is estimated. This suggests that we obtained between 78-88% of the total richness for the Thomisidae family in the studied locality, while 12-22% of the species were not detected.
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Annual | Dry season | Rainy season | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S | 1 | 2 | – | 7 | 7 | 7 | 10 | 6 | 10 | 7 | 5 | 3 | 16 | 9 | 14 |
| A | 1 | 2 | – | 4 | 13 | 22 | 35 | 20 | 24 | 86 | 18 | 9 | 234 | 47 | 187 |
| ⁰D | 1 | 2 | – | 7 | 7 | 7 | 10 | 6 | 10 | 7 | 5 | 3 | 16 | 9 | 14 |
| ¹D | 1 | 2 | – | 2.83 | 5.91 | 5.93 | 6.66 | 3.56 | 5.99 | 2.12 | 2.85 | 1.98 | 6.56 | 5.08 | 6.29 |
| ²D | 1 | 2 | – | 2.66 | 5.12 | 5.37 | 4.84 | 2.5 | 3.89 | 1.45 | 2.07 | 1.58 | 3.66 | 3.26 | 3.66 |
Discussion
Due to the scarce taxonomic information on the spider family Thomisidae in Mexico, the lack of reference collections and specialists, it is not uncommon for faunistic inventories or ecological studies to report a high percentage of unknown or undescribed crab spider species, often with more than 80% of records identified only as morphospecies (Álvarez-Padilla et al., 2020; Argañaraz & Gleiser, 2017; Desales-Lara, 2014; Nieto-Castañeda et al., 2014; Rivera-Quiroz et al., 2016; Rodríguez-Rodríguez et al., 2015). This project reported only 31% of crab spiders as morphospecies. The records presented herein constitute 19% and 61% of the Thomisidae species and genera reported for the country, respectively. Currently, the spider family Thomisidae in Mexico comprises 18 genera and 83 species (World Spider Catalog, 2025); however, more than one-third of Mexican states have no records, or only a few (Pilo-García, 2022). With these results, Morelos now has the fifth highest number of thomisid species in Mexico (16 species), after Veracruz (19), Baja California (19), Chihuahua (19), and Chiapas (16). Previous studies in Mexican deciduous tropical forests recorded the genera Mecaphesa and Synema Simon, 1864 as the most diverse, but herein we found Misumenoides to have the highest species richness, followed by Mecaphesa and Tmarus. We found that Thomisidae individuals are present year-round and are predominantly immature, a common pattern in Araneae. An interesting observation is that the abundance of males was statistically higher than that of adult females; this is uncommon among spiders but has been recorded in the family, and is consistent with the Gravity Hypothesis of extreme sexual size dimorphism in sit-and-wait predators, where smaller males disperse more actively (Corcobado et al., 2010; Hormiga et al., 2000). The marked seasonality, with greater abundance in the rainy than the dry season, aligns with previous thomisid studies (Cardoso et al., 2007; Hernández-Silva, 2016) and tracks the seasonality of the main prey (butterflies and beetles). Rare species were more abundant in the rainy season, whereas more common species were recorded in the dry season, likely reflecting resource (prey) availability. Most species recorded in spider inventories are rare (singletons or doubletons), making it difficult for the accumulation curve to reach the asymptote; evenness factors confirmed the community is predominantly composed of rare species (Jost, 2010), with a singleton proportion of 19%. In conclusion, the family Thomisidae in Morelos is more diverse than previously considered, highlighting the importance of reliable regional faunistic inventories. Using at least 3 techniques with monthly sampling is sufficient to obtain a reliable inventory, even in deciduous tropical forests, although we recommend additional techniques and combining sampling across both seasons to approach the asymptote and obtain comparable data. We consider Thomisidae to be good ecological models given their close association with vegetative strata and their predatory traits; further studies from other localities will broaden understanding of their community structure and its spatial and temporal variation.
Acknowledgements
We thank the following people who helped with fieldwork: García-Márquez V. D., Guzmán-Martínez U. A., Menéndez-Acuña M., Valle-Parra T. A., Uriostegui-Nava E., García-García T. D., Silvestre R., Rodríguez-Herrera B., and Valenzuela C. We also thank A. Hk. Gómez-Calderón for creating the map. We are grateful to Ma. Ventura Echeverría, Ricardo Paredes León, Alejandro Flores Palacios, and Angélica María Corona López for their comments and suggestions. This work would not have been possible without the permission of the community of San Andrés de la Cal. The Facultad de Ciencias Biológicas (UAEM) and the Colección de Insectos de la Universidad de Morelos (CIUM, CIByC) supported this project.
References
Abraham, B. J. (1983). Spatial and temporal patterns in a sagebrush steppe spider community (Arachnida, Araneae). The Journal of Arachnology, 11, 31‒50.
Álvarez-Padilla, F., Galán-Sánchez, M. A., & Salgueiro-Sepúlveda, J. (2020). A protocol for online documentation of spider biodiversity inventories applied to a Mexican tropical wet forest (Araneae: Araneomorphae). Zootaxa, 4722, 241‒269. https://doi.org/10.11646/zootaxa.4722.3.2
Arango, A. M., Rico-Gray, V., & Parra-Tabla, V. (2000). Population structure, seasonality, and habitat use by the green lynx spider Peucetia viridans (Oxyopidae) inhabiting Cnidoscolus aconitifolius (Euphorbiaceae). The Journal of Arachnology, 28, 185‒194.
Argañaraz, C. I., & Gleiser, R. M. (2017). Does urbanization have positive or negative effects on crab spider (Araneae: Thomisidae) diversity? Zoologia, 34, 1‒8. https://doi.org/10.3897/zoologia.34.e19987
Ávalos, G., Rubios, G. D., Bar. M. E., & González, A. (2007). Arañas (Arachnida: Araneae) asociadas a dos bosques degradados del Chaco húmedo en Corrientes, Argentina. Revista de Biología Tropical, 55, 899‒999. https://doi.org/10.15517/rbt.v55i3-4.5965
Balvanera, P., & Maass, M. (2010). Los servicios ecosistémicos que proveen las selvas secas. In G. Ceballos et al. (Eds.), Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México (pp. 251‒271). México, D.F.: Fondo de Cultura Económica.
Benjamin, S. P. (2011). Phylogenetics and comparative morphology of crab spiders (Araneae: Dionycha, Thomisidae). Zootaxa, 3080, 1‒108. https://doi.org/10.11646/zootaxa.3080.1.1
Benjamin, S. P., Dimitrov, D., Gillispie, R. G., & Hormiga, G. (2008). Family ties: molecular phylogeny of crab spiders (Araneae: Thomisidae). Cladistics, 24, 708‒722. https://doi.org/10.1111/j.1096-0031.2008.00202.x
Campuzano, E. F., & Padilla-Ramírez, J. R. (2020). Spatial and seasonal patterns of ground-dwelling spider assemblages belonging to the retrolateral tibial apophysis clade (Araneae: Araneomorphae) in two remnants of pine-oak forest from Mexico. Studies on Neotropical Fauna and Environment, 56, 220‒233. https://doi.org/10.1080/01650521.2020.1806008
Cardoso, P., Henriques, S. S., Gaspar, C., Crespo, L. C., Carvalho, R., Schmidt, J. B. et al. (2007). Species richness and composition assessment of spiders in a Mediterranean scrubland. Journal of Insect Conservation, 13, 45‒55. https://doi.org/10.1007/s10841-007-9116-3
Chao, A., Gotelli, N. J., Hsieh, T. C., Sander, E. L., Ma, K. H., Colwell, R. K. et al. (2014). Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecological Monographs, 84, 45–67. https://doi.org/10.1890/13-0133.1
Chao, A., Ma, K. H., & Hsieh, T. C. (2016). iNEXT Online: software for interpolation and extrapolation of species diversity. http://chao.stat.nthu.edu.tw/wordpress/software_download/inext-online/
Clausen, I. H. S. (1986). The use of spiders (Araneae) as ecological indicators. Bulletin of the British Arachnological Society, 7, 83‒86.
Coddington, J. A., Agnarsson, I., Miller, J. A., Kuntner, M., & Hormiga, G. (2009). Undersampling bias: the null hypothesis for singleton species in tropical arthropod surveys. Journal of Animal Ecology, 78, 573‒584. https://doi.org/10.1111/j.1365-2656.2009.01525.x
Coddington, J. A., Young, L. H., & Coley, F. A. (1996). Estimating spider species richness in a southern Appalachian cove hardwood forest. The Journal of Arachnology, 24, 111‒128.
Colwell, R. K. (2013). EstimateS: statistical estimation of species richness and shared species from samples. Version 9. http://purl.oclc.org/estimates
Colwell, R. K., & Coddington, J. A. (1994). Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London B, 345, 101‒118.
Connell, J. H. & Orias, E. (1964). The ecological regulation of species diversity. The American Naturalist, 98, 399‒414. https://doi.org/10.1086/282335
Corcobado, G., Rodríguez-Gironés, M. A., de Mas, E., & Moya-Laraño, J. (2010). Introducing the refined gravity hypothesis of extreme sexual size dimorphism. BMC Evolutionary Biology, 10, 236. https://doi.org/10.1186/1471-2148-10-236
Corcuera, P., Jiménez, M. L., & Desales-Lara, M. A. (2019). The spiders of the Churince Region, Cuatro Ciénegas Basin. In F. Alvarez, & M. Ojeda (Eds.), Animal diversity and biogeography of Cuatro Ciénegas Basin (pp. 61‒75). Springer Nature. https://doi.org/10.1007/978-3-030-11262-2_5
Corona-López, A. M., Reza-Pérez, E. V., Toledo-Hernández, V. H., Flores-Palacios, A., Macrae, T. C., Westcott, R. L. et al. (2017). Diversity of Buprestidae (Coleoptera) from El Limón de Cuauchichinola, Tepalcingo, Morelos, Mexico. The Pan-Pacific Entomologist, 93, 71‒83. https://doi.org/10.3956/2017-93.2.71
Cortez-Hernández, A. G. (2019). Diversidad y distribución de la familia Thomisidae (Arachnida: Araneae) de la sierra de Zongolica, Veracruz (Bachelor’s Thesis). FES Zaragoza, UNAM, CDMX.
Desales-Lara, M. A. (2014). Araneofauna (Arachnida: Araneae) del Estado de México, México. Acta Zoológica Mexicana, 30, 298‒320. https://doi.org/10.21829/azm.2014.302104
Desales-Lara, M. A., Francke, O. F., & Sánchez-Nava, P. (2013). Diversidad de arañas (Arachnida: Araneae) en hábitats antropogénicos. Revista Mexicana de Biodiversidad, 84, 291‒305. https://doi.org/10.7550/rmb.31708
Durán-Barrón, C. G., Francke, O. F., & Pérez-Ortiz, T. M. (2009). Diversidad de arañas (Arachnida: Araneae) asociadas con viviendas de la Ciudad de México (Zona Metropolitana). https://doi.org/10.22201/ib.20078706e.2009.001.584
Esquivel-Gómez, L., Abdala-Robles, L., Pinkus-Rendón, M., & Parra-Tabla, V. (2016). Effects of tree species diversity on a community of weaver spiders in a tropical forest plantation. Biotropica, 1‒18. https://doi.org/10.1111/btp.12352
Francke, O. F. (2014). Biodiversidad de Arthropoda (Chelicerata: Arachnida ex Acari) en México. Revista Mexicana de Biodiversidad, 85 (Suppl.), 408‒418. https://doi.org/10.7550/rmb.31914
García-García, M. A., Ibarra-Núñez, G., Martínez-Matínez, L., Santos-Moreno, A., & Sánchez-García, J. A. (2023). Spider diversity (Arachnida: Araneae) in two different coffee management systems and surrounding tropical forest during two contrasting seasons in Oaxaca, Mexico. Acta Zoológica Mexicana, 39, 1‒25. https://doi.org/10.21829/azm.2023.3912584
García, E. (2004). Modificaciones al sistema de clasificación climática de Köppen. Instituto de Geografía. Ciudad de México: UNAM.
Gertsch, W. J. (1953). The spider genera Xysticus, Choriarachne and Oxyptila (Thomisidae: Misumeninae) in North America. The American Museum of Natural History, 102, 413‒482.
Gertsch, W. J., & Davis, L. I. (1940). Report on a collection of spiders from Mexico III. The American Museum of Natural History, 1069, 1‒22.
González-Castillo, M. P., Pérez-Santiago, G., & Correa-Ramírez, M. M. (2014). Inventario preliminar sobre el orden Araneae (Arachnida) en comunidades semiáridas de los valles de Durango, Dgo. México. Entomología Mexicana, 1, 26‒31.
Gotelli, N. J., & Cowell, R. K. (2011). Estimating species richness. In A. E. Magurran, & B. J. McGill (Eds.), Biological diversity: frontiers in measurement and assessment (pp. 34‒335). New York: Oxford Press.
Halaj, J., Ross, D. W., & Moldenke A. R. (1998). Habitat structure and prey availability as predictors of the abundance and community organization of spiders in Western Oregon forest canopies. Journal of Arachnology, 26, 203‒220.
Hernández-Silva, Y. G. (2016). Araneofauna de vegetación en tratamientos de restauración ecológica en selva estacional de Sierra de Huautla, Morelos (Bachelor’s Thesis). Facultad de Ciencias Biológicas, UAEM, Cuernavaca.
Hoffmann, A. (1976). Relación bibliográfica preliminar de las arañas de México (Arachnida, Araneae). Publicaciones Especiales Núm. 3. México D.F.: Instituto de Biología, UNAM.
Hormiga, G., Scharff, N., & Coddington, J. A. (2000). The phylogenetic basis of sexual size dimorphism in orb-weaving spiders (Araneae, Orbiculariae). Systematic Biology, 49, 435‒462.
Jiménez, M. L. (1986). Descripción de una nueva especie de la familia Thomisidae (Arachnida: Araneae) de México. Anales del Instituto de Biología, UNAM, Serie Zoología, 56, 11‒14.
Jiménez, M. L. (1992). New species of crab spider from Baja California Sur (Araneae: Thomisidae). The Journal of Arachnology, 20, 50‒57.
Jiménez, M. L. (1996). Araneae. In J. Llorente-Bousquets et al. (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México (pp. 83‒101). México, D.F.: Conabio.
Jiménez, M. L., Chamé-Vázquez, D., Palacios-Cardiel, C., & Van-Devender, T. R. (2024). Spiders (Arachnida: Araneae) from the Sky Island Mountains of Sonora. The Pan-Pacific Entomologist, 100, 175‒211. https://doi.org/10.3956/2024-100.3.175
Jiménez, M. L., Palacios-Cardiel, C., & Chávez-Lara, I. D. (2020). Las arañas (Arachnida: Araneae) de los Médanos de Samalayuca, Chihuahua y nuevos registros de especies para México. Acta Zoológica Mexicana, 36, 1‒23. https://doi.org/10.21829/azm.2020.3612205
Jiménez-Valverde, A., & Hortal, J. (2003). Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8, 151‒161.
Jiménez-Valverde, A., & Lobo, J. M. (2004). Un método sencillo para seleccionar puntos de muestreo con el objetivo de inventariar taxones hiperdiversos: el caso práctico de las familias Araneidae y Thomisidae (Araneae) en la comunidad de Madrid, España. Ecología, 18, 297‒308.
Jiménez-Valverde, A., & Lobo, J. M. (2005). Determining a combined sampling procedure for a reliable estimation of Araneidae and Thomisidae assemblages (Arachnida: Araneae). The Journal of Arachnology, 33, 33‒42. https://doi.org/10.1636/M03-10
Jiménez-Valverde, A., & Lobo, J. M. (2007). Determinants of local spider (Araneidae and Thomisidae) species richness on a regional scale: climate and altitude vs. habitat structure. Ecological Entomology, 32, 113‒122. https://doi.org/10.1111/j.1365-2311.2006.00848.x
Jocqué, R., & Dippenaar-Schoeman, A. S. (2006). Spider families of the world. Belgium: Royal Museum of Central Africa.
Jost, L. (2006). Entropy and diversity. Oikos, 113, 363‒375. https://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost, L. (2010). The relation between evenness and diversity. Diversity, 2, 207‒232. https://doi.org/10.3390/d2020207
Jost, L., & González-Oreja, J. A. (2012). Midiendo la diversidad biológica: más allá del índice de Shannon. Acta Zoológica Lilloana, 56, 3‒14.
Lapp, T. J., & Dondale, C. D. (2017). Thomisidae. In D. Ubick, P. Paquin, P. E. Cushing, & V. Roth (Eds.), Spiders of North America: an identification manual (pp. 272‒273). Keene, New Hampshire: American Arachnological Society.
Llinas-Gutiérrez, J., & Jiménez, M. L. (2004). Arañas de humedales del sur de Baja California, México. Anales del Instituto de Biología, UNAM, Serie Zoología, 75, 283‒302.
Lopes-Rodrigues, E. N., Marques-Pires, M., & de Souza-Mendonça, M. J. (2023). Seasonal changes in spider diversity in subtropical riparian forest. The Journal of Arachnology, 51, 19‒26. https://doi.org/10.1636/JoA-S-21-049
Luna-Reyes, M., Llorente-Bousquets, J., & Luis-Martínez, A. (2008). Papilionidae de la Sierra de Huautla, Morelos y Puebla, México (Insecta: Lepidoptera). Revista de Biología Tropical, 56, 1677‒1716.
Maldonado-Carrizales, J., Ponce-Saavedra, J., & Valdez-Mondragón, A. (2021). Riqueza y abundancia de arañas (Arachnida: Araneae) en ambientes urbanos y su vegetación aledaña al poniente de la ciudad de Morelia, Michoacán, México. Revista Mexicana de Biodiversidad, 92, 1‒18. https://doi.org/10.22201/ib.20078706e.2021.92.3650
Martínez-Hernández, J. G., Corona-López, A. M., Flores-Palacios, A., Rös, M., & Toledo-Hernández, V. H. (2019). Seasonal diversity of Cerambycidae (Coleoptera) is more complex than thought. PeerJ, 7, e7866. https://doi.org/10.7717/peerj.7866
Martínez-Hernández, J. G., Corona-López, A. M., Flores-Palacios, A., & Toledo-Hernández, V. H. (2022). Structure of Cerambycidae (Coleoptera) community in a tropical deciduous forest in northern Morelos, Mexico. Annals of the Entomological Society of America, 20, 1‒8. https://doi.org/10.1093/aesa/saac005
Maya-Morales, J., Ibarra-Núñez, G., León-Cortés, J. L., & Infante, F. (2012). Understory spider diversity in two remnants of tropical montane cloud forest in Chiapas, Mexico. Journal of Insect Conservation, 16, 25‒38. https://doi.org/10.1007/s10841-011-9391-x
Menéndez-Acuña, M. (2017). Efecto de la diversidad genética de una especie fundadora sobre la comunidad de plantas y arañas asociadas (M.Sc. Thesis). CIByC, UAEM, México.
Menéndez-Acuña, M., Salas-Rodríguez, M., Montiel-Parra, G., Sotuyo, S., & Rosas-Echeverría, M. V. (2023). Seasonality and long-term effect of environmental variables on the orb weaver spider community of a tropical dry forest in the Balsas Basin, Mexico. Diversity, 15, 466. https://doi.org/10.3390/d15030466
Miranda, F., & Hernández, X. E. (1963). Los tipos de vegetación en México y su clasificación. Boletín de la Sociedad Botánica de México, 28, 29‒179. https://doi.org/10.17129/botsci.1084
Morse, D. H. (2007). Predator upon a flower: life history and fitness in a crab spider. Cambridge, MA: Harvard University Press.
Nadal, M. F., Achitte-Schmutzler, H. C., Zanone, I., González, P. I., & Ávalos, G. (2018). Diversidad estacional de arañas en una reserva natural del Espinal en Corrientes, Argentina. Caldasia, 40, 129‒143.
Nieto-Castañeda, I. G., Pérez-Miguel, L., & García-Cano, A. A. (2014). New records of spiders (Arachnida: Araneae) from the Balsas Basin in central Mexico. The Southwestern Naturalist, 59, 426‒430. https://doi.org/10.1894/JC-77.1
Orozco-Gil, M., & Desales-Lara, M. A. (2021). Las arañas (Arachnida: Araneae) del estado de Hidalgo, México. Acta Zoológica Mexicana (N.S.), 31, 1‒23. https://doi.org/10.21829/azm.2023.3912583
Pérez-Hernández, C. X., & Zaragoza-Caballero, S. (2016). Temporal variation in the diversity of Cantharidae (Coleoptera) in seven assemblages in tropical dry forest in Mexico. Tropical Conservation Science, 9, 439‒464. https://doi.org/10.1177/194008291600900124
Pickard-Cambridge, F. O. (1895-1903). Arachnida: Araneidea and Opiliones. Biologia Centrali Americana, 2, 89‒214.
Pickard-Cambridge, O. (1896). Arachnida-Araneidea. Biologia Centrali Americana, 1, 57‒288.
Pilo-García, F. (2022). Estructura de la comunidad de la familia Thomisidae (Arachnida: Araneae) en San Andrés de la Cal, Tepoztlán, Morelos (Bachelor’s Thesis). Facultad de Ciencias Biológicas, UAEM, Cuernavaca.
Piña-Rodríguez, O. P., García-Cano, A. A., & Espinosa-Organista, D. H. (2015). Inventario de arañas (Arachnida: Araneae) de la zona noroeste de Guerrero, México. Boletín de la Sociedad Mexicana de Entomología, 1, 26‒32.
Ponce-Saavedra, J. et al. (2023). The fauna of Arachnids in the Anthropocene of Mexico. In R. W. Jones et al. (Eds.), Mexican Fauna in the Anthropocene (pp. 17–46). Cham: Springer. https://doi.org/10.1007/978-3-031-17277-9_2
R Core-Team (2024). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Reyes-González, R., Toledo-Hernández, V. H., Flores-Palacios, A., Rös, M., Bueno-Villegas, J., & Corona-López, A. M. (2024). Exploring the proportion of rarity in tropical insects. Écoscience, 1‒10. https://doi.org/10.1080/11956860.2024.2435133
Rivas-Herrera, C. I. (2015). Araneofauna errante (Arachnida: Araneae) en tratamientos de restauración ecológica en la selva estacional de Sierra de Huautla, México (Bachelor’s Thesis). Facultad de Ciencias Biológicas, UAEM, Cuernavaca.
Rivera-Quiroz, F. A., Garcilazo-Cruz, U., & Álvarez-Padilla, F. (2016). Spider cyberdiversity (Araneae: Araneomorphae) in an ecotouristic tropical forest fragment in Xilitla, Mexico. https://doi.org/10.1016/j.rmb.2016.07.011
Rodríguez-Rodríguez, S. E., Solís-Catalán, K. P., & Valdez-Mondragón, A. (2015). Diversity and seasonal abundance of anthropogenic spiders (Arachnida: Araneae) in different urban zones of the city of Chilpancingo, Guerrero, México. https://doi.org/10.1016/j.rmb.2015.09.002
Rosas-Echeverría, M. V., Coyote-Ávila, C. A., Aguilar-Dorantes, K. M., & Martínez-Peralta, C. (2019). Diversity of butterflies (Lepidoptera) in Sierra de Huautla, Morelos, México. Annals of the Entomological Society of America, 112, 409‒417. https://doi.org/10.1093/aesa/saz022
Ruiz-Rivera, C. A. (2001). San Andrés de la Cal: culto a los señores del tiempo en rituales agrarios. Cuernavaca, Morelos: UAEM.
Samu, F., & Lövei, G. L. (1995). Species richness of a spider community (Araneae). European Journal of Entomology, 92, 633‒638.
Silva, D., & Coddington, J. A. (1996). Spiders of Pakitza (Madre de Dios, Perú). In D. E. Wilson, & A. Sandoval (Eds.), The biodiversity of southeastern Perú (pp. 253‒308). Washington, D.C.: Smithsonian Institute.
Sørensen, L. L. (2002). Stratification of the spider fauna in a Tanzanian forest. In Y. Basset et al. (Eds.), Arthropods of tropical forest (pp. 92‒101). Cambridge, UK: Cambridge University Press.
Trejo, I. (2010). Las selvas secas del Pacífico mexicano. In G. Ceballos et al. (Eds.), Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México (pp. 41‒51). México, D.F.: Fondo de Cultura Económica.
Turnbull, A. L. (1973). Ecology of the true spiders (Araneomorphae). Annual Review of Entomology, 18, 305‒348. https://doi.org/10.1146/annurev.en.18.010173.001513
Ubick, D., Paquin, P., Cushing, P. E., & Roth, V. (2017). Spiders of North America: an identification manual, 2nd Edition. New Hampshire: American Arachnological Society.
Vergara-Torres, C. A., Pacheco-Álvarez, M. C., & Flores-Palacios, A. (2010). Host preference and host limitation of vascular epiphytes in a tropical dry forest of central Mexico. Journal of Tropical Ecology, 26, 563‒570. https://doi.org/10.1017/S0266467410000349
Williams, P. H., & Gaston, K. J. (1994). Measuring more of biodiversity: can higher-taxon richness predict wholesale species richness? Biological Conservation, 67, 211‒217. https://doi.org/10.1016/0006-3207(94)90612-2
Wolda, H. (1988). Insect seasonality: why? Annual Review of Ecology and Systematics, 19, 1‒18. https://doi.org/10.1146/annurev.es.19.110188.000245


