
Molecular identification of composting earthworms using DNA barcoding
Mayela Martínez-Cano a, *, Ana Elena Dorantes-Acosta a, Sergio Martínez-Hernández a, José Antonio García-Pérez b, Huemantzin Balan Ortiz-Oliveros c y Angel Isauro Ortiz-Ceballos a
a Universidad Veracruzana, Instituto de Biotecnología y Ecología Aplicada, Av. de las Culturas Veracruzanas 101, Col. Emiliano Zapata, 91090 Xalapa, Veracruz, México
b Universidad Veracruzana, Facultad de Biología, Circuito Universitario Gonzalo Aguirre Beltrán, Zona Universitaria, 91090 Xalapa, Veracruz, México
c Instituto Nacional de Investigaciones Nucleares, Departamento de Química, La Marquesa, 52750 Ocoyoacac, Estado de México, México
*Autor de correspondencia: maymartinez@uv.mx (M. Martínez-Cano)
Recibido: 30 agosto 2025; aceptado: 4 marzo 2026
Resumen
Las lombrices de tierra composteras desempeñan un papel clave en la descomposición de la materia orgánica, las especies epigeas del género Eisenia son utilizadas ampliamente para efectuar el proceso de lombricompostaje y como bioindicadores en los estudios toxicológicos. Sin embargo, la identidad taxonómica precisa de las especies (semejanza morfológica) representa un desafío y puede afectar la eficacia y función del proceso de lombricompostaje. En este estudio se determinó la identidad de lombrices presentes en 5 instalaciones de vermicompostaje de Veracruz y Quintana Roo, México, empleando análisis molecular basado en la secuenciación del gen mitocondrial COI. El análisis reveló la presencia de 2 especies, Eisenia andrei y Perionyx excavatus, descartando la presencia de Eisenia fetida, la especie asumida previamente por encargados de las plantas de lombricompostaje. Las distancias génicas fueron < 1.3%, mientras que las divergencias con E. fetida superaron 16%, lo que valida la delimitación específica. Nuestros resultados destacan la necesidad de integrar métodos moleculares en la identificación de lombrices de tierra para su uso en los estudios toxicológicos y fortalecer el manejo sostenible de residuos orgánicos.
Palabras clave: Residuos orgánicos; Taxonomía; Especies epigeas
Abstract
Composting earthworms play a key role in the decomposition of organic matter. Epigeic species of the genus Eisenia are widely used both in vermicomposting and as bioindicators in toxicological studies. However, accurate taxonomic identification of these species remains challenging due to their high morphological similarity, which affects the efficiency and function of vermicomposting processes. In this study, the earthworm species present in 5 vermicomposting facilities located in Veracruz and Quintana Roo, México, were identified through molecular analyses of the mitochondrial COI gene. The results revealed the presence of 2 species: Eisenia andrei and Perionyx excavatus, ruling out Eisenia fetida, which had previously been assumed by facility operators. Genetic distances between the identified specimens were below 1.3%, while divergences from E. fetida exceeded 16%, supporting clear species delimitation. Our results highlight the need to integrate molecular methods into earthworm identification for their use in toxicological studies and to strengthen the sustainable management of organic waste.
Keywords: Organic waste; Taxonomy; Epigean species
Introducción
Las lombrices de tierra desempeñan un papel fundamental en los ecosistemas terrestres debido a su contribución a la descomposición de la materia orgánica y a la mejora de las propiedades fisicoquímicas del suelo (Fragoso y Rojas, 2014). Por su relevancia ecológica ha motivado un interés creciente por comprender su diversidad y distribución global. Hasta la fecha, se han descrito alrededor de 5,700 especies de lombrices de tierra, aunque se considera que la diversidad real podría ser mucho mayor debido a la falta de taxonomía y la presencia de especies crípticas (Misirlioğlu et al., 2023). Esta diversidad refleja la adaptación de las lombrices de tierra a distintos hábitats y su uso potencial en diversas aplicaciones biotecnológicas. Las lombrices de tierra epigeas habitan principalmente la hojarasca y se alimentan de materia orgánica en descomposición. Debido a estas características, varias especies han sido empleadas en prácticas sostenibles para el manejo de residuos orgánicos, particularmente en procesos de lombricompostaje (Domínguez y Edwards, 2004). La lombricomposta es el producto de la transformación biológica de residuos orgánicos en biofertilizantes ricos en nutrientes, proceso en el cual las lombrices desempeñan un papel central. No obstante, las especies epigeas utilizadas en lombricompostaje presentan diferencias importantes en su comportamiento, tolerancia ambiental y eficiencia en la degradación de los residuos, lo que genera variabilidad interespecífica en el rendimiento del proceso (Suthar y Singh, 2008). En la región de alta montaña del centro de Veracruz, el lombricompostaje de residuos orgánicos se desarrolló inicialmente a partir del aprovechamiento de la pulpa de café. Durante esta etapa, las lombrices empleadas fueron identificadas morfológicamente como Eisenia fetida (Savigny, 1826) y Metaphire californica (Aranda, 1988). Sin embargo, revisiones taxonómicas posteriores corrigieron dichas identificaciones, demostrando que las poblaciones utilizadas correspondían en realidad a Eisenia andrei (Aranda et al., 1999, 2011). Estos antecedentes evidencian las limitaciones de la identificación basada exclusivamente en criterios morfológicos. De manera paralela, en una instalación de lombricompostaje ubicada en Playa del Carmen, Quintana Roo, se emplean lombrices de tierra para el tratamiento de residuos orgánicos de origen vegetal y animal. En estos sistemas, la identificación de las especies también se ha basado principalmente en la apreciación empírica de los operadores, quienes las reconocen de forma tentativa como E. fetida y asumen que su eficacia en el proceso constituye evidencia suficiente de su identidad taxonómica. Sin embargo, esta práctica puede ser errónea, especialmente cuando se trata de especies morfológicamente similares. Diversas especies de lombrices de tierra han demostrado potencial para la producción de lombricomposta (Domínguez y Edwards, 2004), entre ellas destacan Lumbricus rubellus (Hoffmeister, 1843), Dendrobaena rubida (Savigny, 1826), Eudrilus eugeniae (Kinberg, 1867), Eiseniella tetraedra (Savigny, 1826), Perionyx excavatus (Perrier, 1872), E. fetida y E. andrei (Bouché, 1972). No obstante, no todas las especies aptas para lombricompostaje son esperables en la región; en Veracruz se han registrado principalmente poblaciones de E. fetida y E. andrei en acumulaciones de pulpa de café (Fragoso, 2011). Mientras que especies tropicales como P. excavatus o E. euginae son más habituales en climas cálidos de baja altitud (por ejemplo, Quintana Roo). Especies de origen templado, por ejemplo L. rubellus, D. rubida, E. tetraedra son menos probables en la región, salvo en casos de introducción o microhábitats específicos (Fragoso y Rojas, 2014). Además, la coexistencia de múltiples especies con morfologías similares sugiere que la identificación basada únicamente en criterios visuales puede ser inexacta, lo que resalta la necesidad de emplear herramientas de diagnóstico más precisas. La identificación taxonómica de lombrices de tierra continúa siendo un desafío, especialmente entre especies estrechamente relacionadas. Un caso particularmente relevante es el de E. fetida y E. andrei, 2 especies ampliamente utilizadas en lombricompostaje, ecotoxicología, fisiología y genética. Varias investigaciones han demostrado de manera concluyente que se trata de especies biológicas distintas, aisladas reproductivamente (Domínguez et al., 2005). Asimismo, los análisis filogenéticos basados en los genes 28S y COI (citocromo c oxidasa subunidad I) han corroborado su diferenciación evolutiva (Domínguez y Pérez-Losada, 2010). El reconocimiento de E. fetida y E. andrei como especies distintas tiene implicaciones relevantes. En procesos de lombricompostaje, se ha documentado que E. andrei presenta tasas de crecimiento y reproducción superiores a las de E. fetida (García et al., 2019), lo que la convierte en una opción más eficiente para el tratamiento de residuos orgánicos. Asimismo, en estudios de ecotoxicología no es posible asumir respuestas equivalentes frente a contaminantes, ya que diferentes especies de lombrices pueden diferir en su sensibilidad y fisiología (Zhang et al., 2022). El método tradicional para clasificar las especies se basa en rasgos morfológicos como la pigmentación y las estructuras reproductivas; sin embargo, son características que pueden ser ambiguas y estar afectadas por el ambiente, lo que puede llevar a cometer errores para su identificación (Domínguez et al., 2005). Con base en estas limitaciones de la taxonomía morfológica, se emplean cada vez más técnicas moleculares para distinguir y diferenciar entre especies morfológicamente semejantes. El gen mitocondrial de la subunidad I de la citocromo c oxidasa (COI) se ha utilizado ampliamente como marcador molecular para la identificación de especies debido a su alta divergencia interespecífica y baja variación intraespecífica (James et al., 2010). Los análisis filogenéticos basados en secuencias del gen COI han demostrado ser eficaces para resolver incertidumbres taxonómicas dentro del género Eisenia, ofreciendo un enfoque confiable para la identificación de especies y estudios evolutivos (King et al., 2008). Así, la técnica molecular aporta información relevante para conocer con seguridad la diversidad de lombrices de tierra en sistemas de lombricompostaje y se respalda su uso en prácticas sostenibles de manejo de residuos orgánicos. Dada la importancia de una identificación taxonómica precisa para estudios ecológicos, ecotoxicológicos y aplicaciones biotecnológicas, el presente estudio tuvo como objetivo determinar, mediante un enfoque molecular, la identidad de las lombrices de tierra utilizadas en 4 plantas de vermicompostaje en el centro de Veracruz y una en Quintana Roo, México.
Materiales y métodos
Las lombrices de tierra composteras fueron recolectadas en diversas plantas de lombricompostaje, donde los propietarios y/o responsables las identifican de manera general como E. fetida. Las instalaciones seleccionadas para el estudio incluyen la Unidad de Capacitación para el Desarrollo Rural (MMCA), la Planta de Lombricomposta Teocelo (MMTO), la tienda “Hongos de las Altas Montañas” (MMFT), la Lombricomposta Olmos (MMOL) y Rancho Balam (MMBA). La información correspondiente a la ubicación, altitud, condiciones climáticas, rango de temperatura y sustrato principal de cada instalación se presenta en la tabla 1.
En cada una de las plantas de lombricompostaje (MMCA, MMTO, MMFT, MMOL y MMBA) se recolectó aproximadamente 1 kg de sustrato que contenía lombrices de tierra, constituido como muestra compuesta, obtenida al tomar una muestra de cada una de las camas de lombricompostaje presentes en cada planta. Éste fue transportado en cajas de plástico al Instituto de Biotecnología y Ecología Aplicada (INBIOTECA). En el laboratorio, cada muestra se mantuvo de manera aislada en condiciones controladas utilizando pulpa de café como sustrato (Aranda y Barois, 2000).
A los 60 días de cultivo, en cada uno de los 5 cultivos de lombricomposta se recolectaron 3 lombrices de tierra sanas y sexualmente maduras. Las lombrices de tierra fueron lavadas suavemente con agua corriente para eliminar restos de materia orgánica, después se sumergieron en formol 4%. A las 24 h se realizó la descripción taxonómica con el apoyo de un estereomicroscopio para identificar posibles diferencias morfológicas externas (tamaño corporal, número de segmentos, coloración, posición del clitelo y los tubérculos pubertarios) entre los especímenes recolectados en cada cultivo (Moreno y Borges, 2004). Todas las lombrices de tierra fueron preservadas y depositadas en el INBIOTECA.
Se recolectaron 15 lombrices de tierra (3 por cada unidad de lombricompostaje), enjuagadas suavemente con agua corriente, colocadas en tubos de ensayo con etanol 96% y conservadas a -18 °C hasta su uso. Las lombrices de tierra composteadoras fueron identificadas utilizando la técnica de código de barras genético basada en la secuenciación del gen mitocondrial de la subunidad I del citocromo c oxidasa (650-653 pb). Para ello, se utilizó el método estandarizado por Römbke et al. (2016), ISO (2019) y Prosser et al. (2013). Primero, se cortó una sección de 4 mm del tejido de la cola de 5 lombrices de cada lote utilizando un bisturí. La extracción de DNA se realizó empleando el método de extracción HotShot (Montero-Pau et al., 2008). La concentración de DNA se midió con absorbancia a 260 nm usando un espectrofotómetro GeneQuant II (Pharmacia Biotech Co.).
La reacción en cadena de la polimerasa (PCR) se llevó a cabo siguiendo el protocolo descrito por DeWaard et al. (2008). El gen COI se amplificó utilizando los cebadores ZplankF1_t1 y ZplankR1_t1 (Prosser et al., 2013). La mezcla de reacción de PCR tuvo un volumen total de 10.37 µL, que contenía: 2 µL de agua ultrapura, 6.25 µL de trehalosa al 10%, 1.25 µL de tampón 10X, 0.625 µL de MgCl2 (50 mM), 0.0625 µL de dNTPs (10 mM), 0.125 µL de cada cebador (10 µM) y 0.06 µL de la enzima Platinum Taq DNA polimerasa. Se añadieron 3 µL de DNA de cada muestra a la mezcla de PCR, obteniéndose un volumen final de 13.5 µL por tubo (10.5 µL de mezcla de PCR y 3 µL de DNA). Los tubos para PCR se colocaron en un termociclador Eppendorf con el siguiente programa: desnaturalización inicial a 94 °C durante 1 min; 5 ciclos de: 94 °C por 40 s, 45 °C por 40 s, y 72 °C por 1 min; 35 ciclos de: 94 °C por 40 s, 51 °C por 40 s, y 72 °C por 1 min; extensión final a 72 °C durante 5 min.
Los productos de la PCR se visualizaron en un gel de agarosa precargado 2% (E-Gel, Invitrogen). La secuenciación se realizó utilizando el kit BigDye© Terminator v.3.1, seguido de una secuenciación bidireccional en un secuenciador automático ABI 3730XL. Las secuencias obtenidas (650-653 pb) fueron editadas con el software CodonCode v.3.0.1 (CodonCode Corporation, Dedham, MA, EE.UU.) y posteriormente subidas a la base de datos Barcode of Life Database (BOLD, boldsystems.org), donde están disponibles en el conjunto de datos Taxonomic Identification of Annelids (TIAMD). Todas las secuencias que cumplieron con los estándares mínimos de calidad (> 500 pb, < 1% de bases ambiguas, sin codones de paro ni contaminación) fueron asignadas a un número de índice de código de barras (BIN) en BOLD (Ratnasingham y Hebert, 2013).
Con base en Römbke et al. (2016), se incorporaron secuencias de referencia de Eisenia andrei (JN869997.1 y AY874511.1) y Eisenia fetida 1 y 2 (AY874513.1 y AY874515.1), las cuales son ampliamente utilizadas como referencias en laboratorios de pruebas ecotoxicológicas ISO-21286 (ISO, 2019). Para P. excavatus, se utilizó una secuencia de referencia disponible en BOLD (EWSJA764-08); además, se incluyó una secuencia adicional de E. andrei disponible en BOLD Systems (GBAN0383-06). Posteriormente, se realizó el cálculo de las distancias génicas entre las secuencias, se estimaron utilizando el software Mega v7.0 (Kumar et al., 2016) y se evaluó la confiabilidad de las estimaciones de distancia, se realizaron 1,000 réplicas bootstrap. Las matrices de distancia obtenidas se usaron en los análisis comparativos entre grupos. Las secuencias fueron alineadas previamente utilizando ClustalW incorporado en Mega. Finalmente, la identificación molecular se realizó usando un fenograma Neighbor-Joining (NJ) con 1,000 réplicas bootstrap en Mega v7.0 (Kumar et al., 2016).
| Planta de lombricompostaje | Ubicación | Altitud (m snm) | Clima | Temperatura (°C) | Fuente de alimento |
|---|---|---|---|---|---|
| UNCADER | Coatepec, Ver. (19°27’46.7” N, 96°58’2” O) | 1,200-1,270 | Templado-húmedo | 10 – 29 | Residuos de hojarasca |
| Teocelo | Teocelo, Ver. (19°22’43.3” N, 96°57’55.5” O) | 1,107 | Templado-húmedo | 10 – 30 | Residuos de frutas y verduras |
| Tienda Hongos | Fortín de las Flores, Ver. (18°54’50” N, 96°59’27” O) | 1,000-1,070 | Cálido-subhúmedo | 22 – 31 | Residuos del cultivo de setas y estiércol de ganado |
| Olmos | San Marcos de León, Ver. (19°25’25.2” N, 96°57’50.6” O) | 1,145-1,320 | Cálido-subhúmedo | 20 – 32 | Pulpa de café |
| Rancho Balam | Playa del Carmen, Quintana Roo (20°47’00.8” N, 86°57’27.5” O) | Nivel del mar | Cálido-húmedo | 25.5 – 28 | Estiércol de ganado |
Resultados
De las 15 lombrices de tierra estudiadas, los 3 especímenes procedentes de la planta MMFT mostraron diferencias evidentes en comparación con el resto (MMTO, MMCA, MMBA y MMOL). Las lombrices de tierra MMTO, MMCA, MMBA y MMOL alcanzaron una longitud corporal promedio de 80.4 ± 3.2 mm y de 45.6 ± 2.1 mm las de MMFT. El número promedio de segmentos de las lombrices de tierra en MMTO, MMCA, MMBA y MMOL fue de 100.2 ± 4.5 y en las de MMFT de 88.3 ± 3.8 segmentos. En relación con la coloración, los especímenes de MMTO, MMCA, MMBA y MMOL mostraron un color rojo oscuro homogéneo con franjas intersegmentales amarillas o pálidas; sin embargo, las lombrices de tierra de MMFT mostraron una coloración rosada a marrón claro y sin franjas amarillas. El clitelo de las lombrices de tierra de MMTO, MMCA, MMBA y MMOL se localizó entre los segmentos 25 y 31, mientras que en las de MMFT, entre los segmentos 18 y 24. Los tubérculos pubertarios en las lombrices de tierra de MMTO, MMCA, MMBA y MMOL fueron prominentes y bien definidos entre los segmentos 29 a 31 y en las de MMFT se observaron menos definidos y ubicados entre los segmentos 22 a 24. Las diferencias morfológicas entre los especímenes sugieren que podrían pertenecer a 2 familias distintas de lombrices de tierra: Lumbricidae (MMTO, MMCA, MMBA y MMOL) y Megascolecidae (MMFT).
De las 15 muestras procesadas, se asignaron 14 BINs mediante BOLD. La discrepancia entre el número de muestras procesadas y los BINs asignados se debió a que en un ejemplar de la población MMOL la extracción de DNA no fue exitosa, falló la secuenciación o no cumplió con todos los requisitos para obtener un BIN. Sin embargo, fue posible identificar la especie, ya que la secuenciación se realizó por triplicado. Entre los 14 ejemplares secuenciados con éxito, se identificaron 2 especies de lombrices, cada una con un BIN asignado: E. andrei (BIN AAA8685) y P. excavatus (BIN AAC8637), las secuencias y sus códigos de identificación se detallan en la tabla 2.
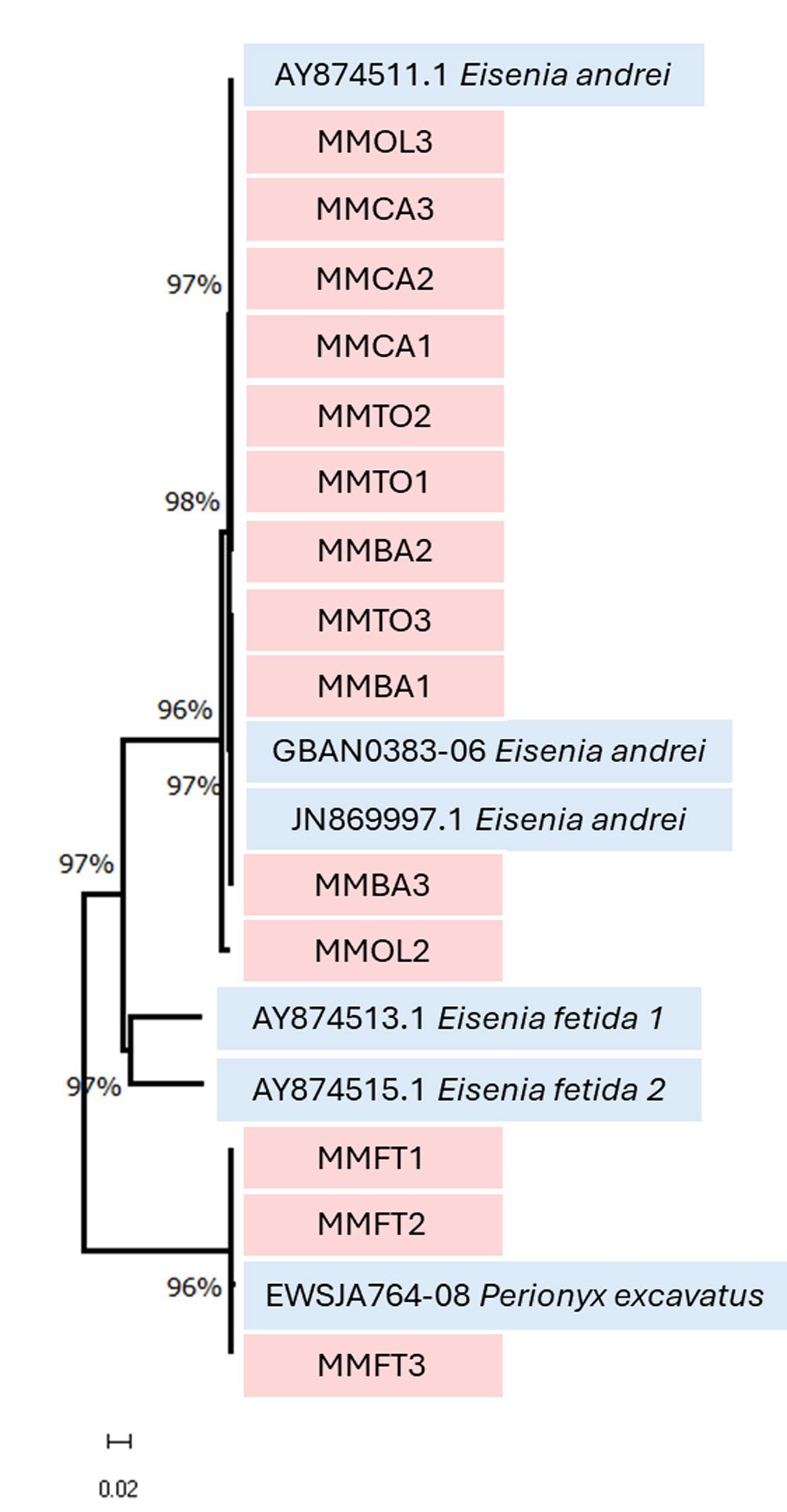
Los resultados de la matriz de distancias genéticas por pares entre las secuencias provenientes de las plantas MMOL, MMTO, MMBA y MMCA y las secuencias de referencia de E. andrei fueron extremadamente bajas, con valores comprendidos entre 0 y 1.3%. En el caso de las secuencias provenientes de la planta MMFT, se registró una divergencia mínima de 0.15% respecto a P. excavatus (tabla 3). El análisis fenético, que permite agrupar organismos en función de su similitud general de caracteres genéticos, se realizó mediante el método de Neighbor-Joining (NJ), el cual construye un árbol fenético basado en distancias entre secuencias. Este análisis reveló 3 clados bien diferenciados: E. andrei, E. fetida y P. excavatus. Las secuencias analizadas se agruparon a los clados de E. andrei y P. excavatus con altos valores de soporte de bootstrap (96-98%), lo que respalda la robustez de estas agrupaciones (fig. 1).
| Código de identificación | Origen | Secuencia 650-653 pb | BIN |
|---|---|---|---|
| TIAMD007-24 (MMCA1) | UNCADER | BOLD Systems | AAA8685 |
| TIAMD008-24 (MMCA42) | UNCADER | BOLD Systems | AAA8685 |
| TIAMD009-24 (MMCA3) | UNCADER | BOLD Systems | AAA8685 |
| TIAMD004-24 (MMTO1) | Teocelo | BOLD Systems | AAA8685 |
| TIAMD005-24 (MMTO3) | Teocelo | BOLD Systems | AAA8685 |
| TIAMD006-24 (MMTO5) | Teocelo | BOLD Systems | AAA8685 |
| TIAMD001-24 (MMFT1) | Tienda Hongos | BOLD Systems | AAC8637 |
| TIAMD002-24 (MMFT2) | Tienda Hongos | BOLD Systems | AAC8637 |
| TIAMD003-24 (MMFT3) | Tienda Hongos | BOLD Systems | AAC8637 |
| TIAMD010-24 (MMOL1) | Olmos | BOLD Systems | AAA8685 |
| TIAMD011-24 (MMOL2) | Olmos | BOLD Systems | AAA8685 |
| TIAMD012-24 (MMOL3) | Olmos | BOLD Systems | AAA8685 |
| TIAMD022-24 (MMBA1) | Rancho Balam | BOLD Systems | AAA8685 |
| TIAMD023-24 (MMBA2) | Rancho Balam | BOLD Systems | AAA8685 |
| TIAMD024-24 (MMBA3) | Rancho Balam | BOLD Systems | AAA8685 |
| AY874513.1 E. fetida | Römbke et al. (2016) | GenBank | — |
| AY874515.1 E. fetida | Römbke et al. (2016) | GenBank | — |
| JN869997.1 E. andrei | Römbke et al. (2016) | GenBank | — |
| GBAN0383-06 E. andrei | Römbke et al. (2016) | BOLD Systems | AAA8685 |
| AY874511.1 E. andrei | Römbke et al. (2016) | GenBank | — |
| EWSJA764-08 P. excavatus | Endemic Species Research Institute | BOLD Systems | AAC8637 |
Discusión
En el presente estudio, se empleó el análisis de código de barras de DNA (secuenciación del gen mitocondrial COI) para identificar las especies de lombrices de tierra presentes en 5 plantas de lombricompostaje. Cuatro plantas localizadas en la región de las altas montañas de Veracruz (MMTO, MMFT, MMCA y MMOL) y una en Playa del Carmen en Quintana Roo (MMBA). La evaluación de caracteres externos permitió identificar al menos 2 grupos morfológicamente distintos entre los ejemplares, posiblemente correspondientes a especies diferentes que, por sus características, podrían ser E. andrei y P. excavatus. De los 15 ejemplares evaluados, 12 provenientes de las plantas de lombricompostaje MMTO, MMCA, MMOL y MMBA compartieron un conjunto homogéneo de características morfológicas, mientras que los 3 individuos de la población MMFT presentaron un fenotipo claramente distinto. Para estudios futuros, se recomienda complementar la investigación con el análisis de caracteres internos y filogenias morfológicas, permitiendo una caracterización completa de las diferencias entre los grupos (Domínguez et al., 2015).
Para la identificación molecular, estudios previos han establecido umbrales de divergencia genética para la delimitación de especies mediante el gen COI. Hebert et al. (2003) encontraron que las divergencias de COI rara vez superan 2% dentro de una especie determinada, mientras que los miembros de especies diferentes suelen presentar divergencias mayores. Chang y James (2011) revisaron la literatura y concluyeron que divergencias inferiores a 9% pueden asignarse a la misma especie. En estudios previos con lombrices de tierra, Römbke et al. (2016) señalaron que valores superiores a 10% podrían indicar diferenciación entre especies. Chang et al. (2009) reportaron que las divergencias intraespecíficas en el gen COI en lombrices de tierra presentan un promedio de 1.3%, mientras que las diferencias interespecíficas alcanzan un promedio de 18.7%.
En nuestro análisis, las distancias genéticas calculadas entre los especímenes muestreados en las plantas de lombricompostaje MMTO, MMCA, MMBA, la secuencia MMOL3 de la planta MMOL y las secuencias de referencia de E. andrei fueron notablemente bajas (< 0.32%). La secuencia MMOL2 presentó una divergencia ligeramente mayor (entre 1.2 y 1.3%) con respecto a las secuencias de referencia de E. andrei; sin embargo, su divergencia genética no excede (2%) los límites típicos observados de variación intraespecífica (Chang et al., 2009). Además, la herramienta de identificación del sistema BOLD asignó a todas estas secuencias, incluida MMOL2, el BIN AAA8685, que coincide con E. andrei (tabla 2). Es importante destacar que no existe un umbral de divergencia genética fijo para determinar cuándo 2 muestras pertenecen a diferentes especies. Sin embargo, tomando en cuenta el patrón observado por Chang et al. (2009) en las divergencias intraespecíficas, indica que los ejemplares recolectados en las plantas de lombricompostaje MMCA, MMBA, MMTO y MMOL pertenecen a la especie E. andrei.
| P. excavatus EWSJA764-08 | E. andrei GBAN0383-06 | E. andrei JN869997.1 | E. andrei AY874511.1 | E. fetida AY874513.1 | E. fetida AY874515.1 | |
|---|---|---|---|---|---|---|
| MMFT1 | 0.0015409276 | 0.2629987796 | 0.2644016040 | 0.2653194932 | 0.2358330274 | 0.2363505591 |
| MMFT2 | 0.0015409276 | 0.2629987796 | 0.2644016040 | 0.2653194932 | 0.2358330274 | 0.2363505591 |
| MMFT3 | 0.0015409276 | 0.2629987796 | 0.2644016040 | 0.2653194932 | 0.2358330274 | 0.2363505591 |
| MMTO1 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMTO2 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMTO3 | 0.2648485363 | 0.0000000000 | 0.0000000000 | 0.0030865172 | 0.1649064730 | 0.1653642510 |
| MMCA1 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMCA2 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMCA3 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMOL2 | 0.2558729431 | 0.0124453913 | 0.0132493333 | 0.0124453913 | 0.1708936388 | 0.1693152390 |
| MMOL3 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMBA1 | 0.2648485363 | 0.0000000000 | 0.0000000000 | 0.0030865172 | 0.1649064730 | 0.1653642510 |
| MMBA2 | 0.2671755709 | 0.0030865172 | 0.0032841858 | 0.0000000000 | 0.1649064730 | 0.1674082417 |
| MMBA3 | 0.2650342592 | 0.0015402451 | 0.0000000000 | 0.0046315330 | 0.1647547930 | 0.1651978963 |
Por otro lado, las secuencias provenientes de la planta MMFT, mostraron una divergencia genética mínima (0.15%) con respecto a la secuencia de referencia de P. excavatus e identificadas como esta especie por BOLD Systems (BIN AAC8637). También, todas las secuencias estudiadas mostraron niveles de divergencia de entre 16 y 23% con respecto a E. fetida, lo que sugiere que ninguna de las secuencias analizadas pertenece a esta especie. Estos hallazgos contradicen las afirmaciones de los propietarios de las plantas de lombricompostaje, quienes indican que las lombrices composteras que cultivan corresponden a E. fetida. Sin embargo, diversas publicaciones sobre el tema señalan la presencia de E. andrei y P. excavatus en instalaciones similares (Aranda et al., 1999, 2011).
Asimismo, el análisis fenético basado en el método de NJ (fig. 1) permitió observar una clara agrupación de las secuencias obtenidas de las plantas MMCA, MMBA, MMTO con las secuencias de referencia de E. andrei. En el caso de la planta MMOL, la secuencia MMOL3, se agrupa consistentemente con el clado de las secuencias de E. andrei. Sin embargo, a pesar de haber sido recolectadas en el mismo sitio y pertenecer a la misma población cultivada, la secuencia MMOL2 se ubicó en una rama separada dentro del árbol fenético, en concordancia con los resultados obtenidos en la matriz de distancias genéticas por pares. Su ubicación en el árbol de NJ sugiere una posible diferenciación intraespecífica. Esta discrepancia puede atribuirse a las limitaciones del método fenético utilizado, el cual se basa únicamente en similitudes de distancia y no en relaciones filogenéticas estadísticamente robustas (Saitou y Nei, 1987). También es posible que MMOL2 represente un haplotipo divergente o una población estructurada genéticamente dentro de E. andrei. Este tipo de variación intraespecífica ha sido documentada previamente en lombrices de tierra cultivadas y no necesariamente implica diferenciación a nivel de especie (Chang et al., 2009). Sin embargo, para esclarecer esta posibilidad, sería recomendable integrar otros datos de secuencias mitocondriales como el 16S (Yadav y Mullah, 2017), secuencias nucleares (H3, ITS2, 18S, 28S) y de genotipado (microsatélites o SNPs) que ofrezcan mayor resolución (Barraux et al., 2024). Este hallazgo, también resalta la importancia de analizar múltiples individuos por sitio y no asumir homogeneidad genética dentro de una misma población cultivada.
Por otro lado, las secuencias de la planta MMFT se agruparon consistentemente con la secuencia de referencia de P. excavatus. Estas agrupaciones presentaron altos valores de soporte bootstrap, lo que respalda la solidez de las asignaciones taxonómicas basadas en el código de barras de DNA.
El análisis fenético basado en el método de NJ es una herramienta útil para la identificación molecular de lombrices de tierra, ya que permite visualizar agrupamientos basados en similitudes genéticas y evaluar la consistencia entre secuencias de prueba y secuencias de referencia. Aunque este enfoque no permite inferencias evolutivas profundas, brinda una delimitación rápida y confiable de especies, como en estudios ecológicos o de lombricompostaje, donde las especies pueden presentar morfologías crípticas (Chang et al., 2009; King et al., 2008). Este enfoque, combinado con el análisis de distancias génicas por pares ha sido validado como una estrategia eficaz para diferenciar especies dentro de géneros complejos como Eisenia, donde la identificación resulta limitada (Römbke et al., 2016). Aunque en este estudio no se secuenció el gen COI completo, la región utilizada (650-653 pb) corresponde al fragmento estandarizado para el código de barras en invertebrados y por su eficacia ha sido ampliamente validado en estudios previos (Hebert et al., 2003; Ratnasingham y Hebert, 2013).
La presencia de P. excavatus en una de las plantas de lombricompostaje resulta particularmente relevante en el ámbito de la ecología aplicada, debido a su elevada tasa de reproducción, rápida colonización del sustrato y amplia tolerancia ambiental (tabla 4) (Suthar y Singh, 2008). Su presencia indica su capacidad de adaptación a condiciones ambientales específicas en las plantas de lombricompostaje, donde los operadores asumían que las lombrices de tierra composteras que cultivan son E. fetida y/o roja californiana.
Por otro lado, E. andrei fue la especie predominante en la mayoría de las plantas analizadas. Su prevalencia se explica por sus características biológicas: distribución cosmopolita, ciclo de vida corto, amplia tolerancia a la variación climática, facilidad de manejo en entornos operativos y experimentales (tabla 4) (Domínguez y Edwards, 2004). Características biológicas que la han convertido en una de las especies más utilizadas en programas de lombricompostaje a nivel global (Domínguez y Edwards, 2004) y utilizada como especie centinela (al igual que E. fetida) en los estudios ecotoxicológicos ISO-17512 (ISO, 2008) y ISO-11268 (ISO, 2012).
En conclusión, el análisis de la región estándar del gen mitocondrial COI (DNA barcoding) permitió identificar con precisión 2 especies de lombrices de tierra composteras E. andrei y P. excavatus, descartando la presencia de E. fetida en las plantas de lombricompostaje evaluadas en las altas montañas del centro de Veracruz y el Caribe mexicano (Quintana Roo). La identificación molecular obtenida concuerda, en gran medida, con revisiones taxonómicas previas basadas en caracteres morfológicos, como las analizadas por Aranda et al. (1999) y Fragoso (2011), las cuales ya habían señalado la presencia predominante de E. andrei en sistemas de lombricompostaje y en tiraderos y cúmulos de pulpa de café del centro de Veracruz. Finalmente, la confirmación de E. andrei y P. excavatus como especies presentes en los sistemas evaluados sugiere la conveniencia de explorar su coexistencia y desempeño conjunto en procesos de lombricompostaje y biorremediación de materiales orgánicos, como el sargazo (Sargassum spp.), con el fin de optimizar la eficiencia biológica y ambiental de estos sistemas (Martínez-Cano et al., 2024).
| Especie | Familia | Origen biogeográfico | Estatus en México | Rango altitudinal (m snm) | Rango térmico (°C) | Presencia en México | Referencias |
|---|---|---|---|---|---|---|---|
| Eisenia andrei | Lumbricidae | Europa | Exótica | 800-2,500 | 10-25 | Sistemas de vermicompostaje o sitios artificiales | (Domínguez y Edwards, 2004; Fragoso, 2011; Fragoso y Rojas, 2014) |
| Perionyx excavatus | Megascolescidae | Asia | Exótica introducida/naturalizada | 0-800 | 20-35 | Suelos de plantaciones y ecosistemas perturbados | (Fragoso, 2011; Fragoso y Rojas, 2014; Suthar y Singh, 2008) |
Agradecimientos
Los autores agradecen a Alma Estrella García Morales por el procesamiento de las muestras de tejido para el análisis de DNA en el Laboratorio de Códigos de Barras de la Vida, Nodo Chetumal. Asimismo, agradecen a los propietarios de las plantas de lombricompostaje por la donación del material biológico. Mayela Martínez Cano agradece al Secihti por la beca de doctorado (1129824).
Referencias
Aranda, E. y Barois, I. (2000). Coffee pulp vermicomposting treatment. En T. Sera, C. R. Soccol, A. Pandey y S. Roussos (Eds.), Coffee biotechnology and quality (pp. 489–506). identificadas en las plantas de lombricompostaje de Veracruz Rango Presencia en Referencias altitudinal térmico México snm) (ºC) 800-2,500 10-25 Sistemas de (Domínguez y vermicompostaje o Edwards, 2004; sitios artificiales Fragoso, 2011; Fragoso y Rojas, 2014) 20-35 Suelos de (Fragoso, 2011; plantaciones Fragoso y Rojas, 2014; y ecosistemas Suthar y Singh, 2008) perturbados Dordrecht: Kluwer Academic Publishers. https://doi.org/10.1007/978-94-017-1068-8_46
Aranda, E., Barois, I., Arellano, P., Irissón, S., Salazar, T., Rodríguez, J. et al. (1999). Vermicomposting in the Tropics. En P. Lavelle, L. Brussaard y P. Hendrix (Eds.), Earthworm management in tropical agroecosystems (pp. 253–287). Wallingford, United Kingdom: CABI Publishing.
Aranda, E., Barois I., Santos, B. M. y Hernandez-Castellanos, B. (2011). Vermicomposting, research and activities in Mexico. En C. A. Edwards, N. Q. Arancon y R. L. Sherman (Eds.), Vermiculture technology: earthworms, organic wastes, and environmental management (pp. 507–32). Boca Raton, Florida: CRC Press.
Barraux, A., Noël, S., Roy, V. y Dupont, L. (2024). Challenges of molecular barcode-based identification of earthworm specimens for biodiversity assessment. Frontiers in Ecology and Evolution, 12, 1358984. https://doi.org/10.3389/fevo.2024.1358984
Chang, C. H. y James, S. (2011). A critique of earthworm molecular phylogenetics. Pedobiologia, 54, S3–S9. https://doi.org/10.1016/j.pedobi.2011.07.015
Chang, C. H., Rougerie, R. y Chen, J. H. (2009). Identifying earthworms through DNA barcodes: pitfalls and promise. Pedobiologia, 52, 171–80. https://doi.org/10.1016/j.pedobi.2008.08.002
de Waard, J., Ivanova, N., Hajibabaei, M. y Hebert, P. (2008). Assembling DNA barcodes: analytical protocols. En C. C. Martin (Ed.), Methods in Molecular Biology: environmental genomics methods (pp. 275–294). Ottawa: Humana Press https://doi.org/10.1007/978-1-59745-548-0_15
Domínguez, J., Aira, M., Breinholt, J. W., Stojanovic, M., James S. W. y Pérez-Losada, M. (2015). Underground evolution: new roots for the old tree of lumbricid earthworms. Molecular Phylogenetics and Evolution, 83, 7–19. https://doi.org/10.1016/j.ympev.2014.10.024
Domínguez, J. y Edwards, C. A. (2004). Vermicomposting organic wastes: a review. Soil Zoology for Sustainable Development in the 21st Century. Cairo: Geocites Publisher.
Domínguez, J. y Pérez-Losada, M. (2010). Eisenia fetida (Savigny, 1826) y Eisenia andrei Bouché, 1972 son dos especies diferentes de lombrices de tierra. Acta Zoológica Mexicana, 26, 321–31. https://doi.org/10.21829/azm.2010.262897
Domínguez, J., Velando A. y Ferreiro, A. (2005). Are Eisenia fetida (Savigny, 1826) and Eisenia andrei Bouché (1972) different biological species? Pedobiologia, 49, 81–87. https://doi.org/10.1016/j.pedobi.2004.08.005
Fragoso, C. (2011). Lombrices de tierra (Annelida: Oligochaeta). La biodiversidad en Veracruz: estudio de estado (pp. 259– 268). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Gobierno del Estado de Veracruz/ Universidad Veracruzana/ Instituto de Ecología A.C.
Fragoso, C. y Rojas, P. (2014). Biodiversidad de lombrices de tierra (Annelida: Oligochaeta: Crassiclitellata) en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), 197–207. https://doi.org/10.7550/rmb.33581
García, S. R., Méndez, G., Velázquez, M. G., Castillo, G. y Retureta, A. (2019). Dinámica poblacional de Eisenia fetida (Savigny, 1826) y Eisenia andrei (Bouché, 1972) en tres sustratos diferentes. Revista Biológico Agropecuaria Tuxpan, 7, 70–78. https://doi.org/10.47808/revistabioagro.v7i2.31
Hebert, P. D. N., Cywinska, A., Ball, S. L. y deWaard, J.R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society B: Biological Sciences, 270, 313–321. https://doi.org/10.1098/rspb.2002.2218
ISO (International Organization for Standardization). (2008). Soil quality: avoidance test for determining the quality of soils and effects of chemicals on behavior. ISO 17512-1. International Organization for Standardization. Disponible en: https://www.iso.org/obp/ui/en/#iso:std:iso:17512:-1:ed-1:v1:en
ISO (International Organization for Standardization). (2012). Soil quality: effects of pollutants on earthworms. ISO 11268-1. International Organization for Standar- dization. Disponible en: https://www.iso.org/obp/ui/#iso:std:iso:11268:-1:ed-2:v1:en
ISO (International Organization for Standardization). (2019). Soil quality: identification of ecotoxicological test species by DNA barcoding. ISO 21286. International Organization for Standardization. Disponible en: https://www.iso.org/obp/ui/en/#iso:std:iso:21286:ed-1:v1:en
James, S. W., Porco D., Decaëns, T., Richard, B., Rougerie, R. y Erséus, C. (2010). DNA Barcoding reveals cryptic diversity in Lumbricus terrestris L., 1758 (Clitellata): resurrection of L. herculeus (Savigny, 1826). Plos One, 5, 1–8. https://doi.org/10.1371/journal.pone.0015629
King, R. A., Tibble, A. L. y Symondson, W. O. C. (2008). Opening a can of worms: unprecedented sympatric cryptic diversity within British lumbricid earthworms. Molecular Ecology, 17, 4684–4698. https://doi.org/10.1111/j.1365-294X.2008.03931.x
Kumar, S., Stecher, G. y Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution, 33, 1870–1874. https://doi.org/10.1093/molbev/msw054
Martínez-Cano, M., Dorantes-Acosta, A. E., Lara-González, R., Salgado-Hernández, E. y Ortiz-Ceballos, A. I. (2024). Effect of sargassum on the behavior and survival of the earthworm Eisenia fetida. Bulletin of Environmental Contamination and Toxicology, 113, 36. https://doi.org/10.1007/s00128-024-03935-5
Montero-Pau, J., Gómez, A. y Muñoz, J. (2008). Application of an inexpensive and high-throughput genomic DNA extraction method for the molecular ecology of zooplanktonic diapausing eggs. Limnology and Oceanography: Methods, 6, 218–222. https://doi.org/10.4319/lom.2008.6.218
Moreno, A. G. y Borges, S. (2004). Advances in earthworm taxonomy. Madrid: Editorial Complutense.
Prosser, S., Martínez-Arce, A. y Elías-Gutiérrez, M. (2013). A new set of primers for COI amplification from freshwater microcrustaceans. Molecular Ecology Resources, 13, 1151– 1155. https://doi.org/10.1111/1755-0998.12132
Ratnasingham, S. y Hebert, P. D. N. (2013). A DNA-based registry for all animal species: The Barcode Index Number (BIN) System. Plos One, 8, e66213. https://doi.org/10.1371/journal.pone.0066213
Römbke, J., Aira, M., Backeljau, T., Breugelmans, K., Domínguez, J., Funke, E. et al. (2016). DNA barcoding of earthworms (Eisenia fetida/andrei complex) from 28 ecotoxicological test laboratories. Applied Soil Ecology, 104, 3–11. https://doi.org/10.1016/j.apsoil.2015.02.010
Saitou, N. y Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4, 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454
Suthar, S. y Singh, S. (2008). Vermicomposting of domestic waste by using two epigeic earthworms (Perionyx excavatus and Perionyx sansibaricus). International Journal of Environmental Science & Technology, 5, 99–106. https://doi.org/10.1007/BF03326002
Yadav, S. y Mullah, M. (2017). A review on molecular markers as tools to study earthworm diversity. International Journal of Pure and Applied Zoology, 5, 62–69.
Zhang, M., Jouquet, P., Dai, J., Xiao, L., Du, Y., Liu, K. et al. (2022). Assessment of bioremediation potential of metal contaminated soils (Cu, Cd, Pb and Zn) by earthworms from their tolerance, accumulation and impact on metal activation and soil quality: a case study in South China. Science of The Total Environment, 820, 152834. https://doi.org/10.1016/j.scitotenv.2021.152834


