
The diversity of vertebrates visiting the strobili of the Dioon edule complex and their potential for pollination
Abraham Isaías Galavíz-Hernández a, Laura Yáñez-Espinosa a, b, *, Alberto Prado c y Joel Flores d
a Universidad Autónoma de San Luis Potosí, Facultad de Ciencias, Av. Chapultepec Núm. 1570, Priv. del Pedregal, 78295 San Luis Potosí, San Luis Potosí, México
b Universidad Autónoma de San Luis Potosí, Instituto de Investigación de Zonas Desérticas, Altair Núm. 200, Col. Del Llano, 78377 San Luis Potosí, San Luis Potosí, México
c Universidad Nacional Autónoma de México, Escuela Nacional de Estudios Superiores, Campus Juriquilla, Boulevard Juriquilla Núm. 3001, 76230 Querétaro, Querétaro, México
d Instituto Potosino de Investigación Científica y Tecnológica, División de Ciencias Ambientales, Camino a la Presa San José Núm. 2055, Col. Lomas 4 sección, 78216 San Luis Potosí, San Luis Potosí, México
*Autor para correspondencia: lyaneze@uaslp.mx (L. Yáñez-Espinosa)
Recibido: 13 octubre 2025; aceptado: 23 febrero 2026
Resumen
La presencia de toxinas en Zamiaceae sugiere interacciones limitadas con vertebrados, aunque existe registro de depredación y dispersión de semillas. El estróbilo masculino, poco considerado para el análisis de interacciones, podría representar un recurso para la fauna. Las especies Dioon edule Lindl. y Dioon angustifolium Miq. constituyen un complejo de cícadas endémicas de México con riesgo de extinción, por lo que es esencial explorar sus relaciones ecológicas ya que sus visitantes podrían tener la capacidad de acercar el polen de un estróbilo a otro. Monitoreamos estróbilos masculinos y femeninos de las especies del complejo con cámaras en la sierra Madre Oriental. El objetivo fue analizar la diversidad de visitantes y su relevancia respecto de su potencial polinizador. Concluimos el registro de una elevada diversidad de fauna nativa que interactúa con los estróbilos de diversas formas y es casi totalmente distinta para cada especie del complejo. Identificamos a las aves como un posible grupo con mayor potencial de transferencia de polen. Los vertebrados que visitan ambos estróbilos son agentes con potencial polinizador de Zamiaceae. Sin embargo, los visitantes de alta frecuencia que interactúan solo con el estróbilo masculino también podrían estar robando el polen.
Palabras clave: Biodiversidad; Interacción planta-animal; Ecología reproductiva
Abstract
The presence of toxins in the Zamiaceae family suggests limited interaction with vertebrates, despite records of predation and seed dispersal. The male strobilus has received little attention in studies of these interactions, but it could represent a valuable wildlife resource. The Dioon edule Lindl. and Dioon angustifolium Miq. species constitute an endemic Mexican cycad complex at risk of extinction; therefore, exploring their ecological relationships is essential, as their visitors may be able to transfer pollen from one strobilus to another. We monitored the male and female strobili of these species in the Sierra Madre Oriental using cameras. Our aim was to analyze visitor diversity and its relevance to pollination potential. We recorded a high diversity of native fauna that interacts with the strobili in various ways, which are almost entirely distinct for each species in the complex. We identified birds as the group with the greatest potential for pollen transfer. Vertebrates that visit both strobili could potentially pollinate the Zamiaceae family. However, visitors that interact only with the male strobilus at high frequency may also be stealing pollen.
Keywords: Biodiversity; Plant-animal interaction; Reproductive ecology
Introducción
Las interacciones ecológicas son factores clave para el funcionamiento de los ecosistemas y para la generación de la biodiversidad en el planeta (Bascompte, 2009). El transporte de polen y la dispersión de semillas representan interacciones positivas que las plantas utilizan mediante una variedad de animales alados como insectos, aves y murciélagos (Herrera y Pellmyr, 2002; Ratto et al., 2018). Asimismo, se ha propuesto que en el pasado estos procesos pudieron haber sido facilitados por grandes vertebrados ahora extintos (Crowley et al., 2011; Hall y Walter, 2013). En las últimas 4 décadas, las investigaciones sobre la interacción planta-polinizador han señalado que los visitantes a las especies vegetales están relacionados filogenéticamente como resultado de un proceso de selección, es decir, dicho sistema ha evolucionado hacia una alta especialización (Campbell, 1989; Herrera, 1987).
Las zamiáceas son un grupo de gimnospermas que incluye a los géneros Bowenia Hook, Ceratozamia Brongniart, Dioon Lindley, Encephalartos Lehm, Lepidozamia Regel, Macrozamia Miquel, Microcycas (Miq.) A. DC., Stangeria T. Moore y Zamia L. (Nicolalde-Morejón et al., 2014; Stevenson, 1992). El género Dioon incluye a las especies D. edule y D. angustifolium, ambas endémicas de México. Estas especies presentan una altura de hasta 1.5 m, con tronco erecto y copa rígida de hojas largas de color verde claro. La estructura básica de sus estróbilos es similar en ambos sexos, con un eje central y esporófilas o escamas (Vovides, 1990). No obstante, los masculinos son cilíndricos y más largos que los femeninos, que son globulares.
El género Dioon se distribuye principalmente a lo largo de la sierra Madre Oriental de México y en algunas regiones de Centroamérica (Lázaro-Zermeño et al., 2012). Por su parte, D. angustifolium se encuentra principalmente en el extremo boreal de la sierra Madre Oriental, en Nuevo León y Tamaulipas (Rubio et al., 2020), mientras que D. edule en Tamaulipas, San Luis Potosí, Hidalgo, Querétaro y Veracruz (Whitelock, 2004; Yáñez-Espinosa, 2006). Ambas especies se encuentran catalogadas como en peligro de extinción, debido al cambio en el uso de suelo y la extracción ilegal de ejemplares (Semarnat, 2010).
Dioon edule y D. angustifolium son consideradas por Dorsey et al. (2018) como parte de un complejo, ya que presentan morfología muy similar sin límites definidos, lo que sugiere una posible continuidad entre ambas especies, además de la ausencia de una separación genética clara (Dorsey et al., 2018; González-Astorga et al., 2005; Rubio-Méndez et al., 2020).
Se ha argumentado que las cícadas se adaptaron a interacciones con grandes vertebrados ahora extintos (Hall y Walter, 2013; Janzen y Martin, 1982). A pesar de la extinción de la megafauna, las cícadas han logrado prevalecer gracias a estrategias ecológicas. Una de ellas es la formación de rodales, los cuales generan grupos de alta densidad poblacional, que favorecen la cercanía entre estróbilos masculinos y femeninos al ser organismos dioicos (Hall y Walter, 2013). Esta proximidad, pudo haber favorecido la polinización anemófila, mecanismo que se consideró predominante hasta principios del siglo XXI (Chamberlain, 1919, 1937).
Durante las últimas décadas se ha demostrado la contribución de insectos en la polinización de las cícadas (Terry et al., 2005, 2008). Estas interacciones parecen haber existido desde el Mesozoico, de las cuales las cícadas dependen actualmente. Los principales polinizadores corresponden a escarabajos de las familias Curculionidae y Erotylidae, los cuales habitan los estróbilos y poseen adaptaciones diseñadas para la polinivoría (Tang et al., 2018). Sin embargo, los reportes actuales de vertebrados visitantes a estróbilos de gimnospermas y especialmente de cícadas, demuestran su posible participación en el proceso de polinización (Fuster y Traveset, 2019; Yáñez-Espinosa, Barragán-Torres et al., 2021).
Se sabe que las zamiáceas presentan compuestos activos neurotóxicos y hepatotóxicos como metilazoximetanol, β-metilamino-L-alanina, β-oxalilamino-L-alanina y cicasina (Rivadeneyra y Rodriguez, 2014). Sin embargo, se han reportado alrededor del mundo interacciones ecológicas entre estructuras reproductivas de organismos vegetales, incluidas las cícadas y una diversidad de vertebrados. Estas interacciones incluyen reptiles (Fuster y Traveset, 2019; Nyhagen et al., 2001; Varela y Bucher, 2002), mamíferos (Cárdenas et al., 2017; González-Christen, 1990; Jones et al., 2024; Pérez-Farrera y Vovides, 2004; Pérez-Farrera et al., 2006; Vovides, 1990; Wester y Stanway, 2009; Yáñez-Espinosa, Barragán-Torres et al., 2021) y aves (David et al., 2015; Jones et al., 2024; Viana et al., 2016; Yáñez-Espinosa, Balderas-González et al., 2021), donde se les sugiere como potenciales polinizadores o dispersores de la planta a la cual visitan. Al buscar en sus estructuras reproductivas un recurso, estos trepan para lamerlas, picotearlas o morderlas. Estas conductas favorecen la adherencia de polen accidental a sus cuerpos, distribuyéndolo entre plantas masculinas y femeninas. En las interacciones ecológicas, la alta variedad de rasgos funcionales podría sugerir que no se trata solo de fauna especialista.
El análisis planta-vertebrado, en cícadas, se ha centrado esencialmente en la dispersión y depredación de semillas, monitoreando solo el estróbilo femenino, este hecho podría deberse a que el estróbilo masculino, independientemente de los insectos consumidores de polen, que son atraídos por sus compuestos volátiles (Suinyuy et al., 2012), no parece representar un recurso directo para los vertebrados como ocurre en otras gimnospermas de la familia Ephedraceae Dumort. (Celedón et al., 2016) o Ginkgoaceae Engl. (Gin et al., 2012). Asimismo, el tiempo en el que está disponible el polen suele ser corto (de 2 a 3 semanas). Sin embargo, indirectamente el estróbilo masculino podría ser una fuente importante de otros recursos, como los insectos que forman parte de la dieta de los vertebrados (Lovette y Fitzpatrick, 2016; Pianka, 1970), principalmente los escarabajos que obtienen del estróbilo masculino un refugio (Tang et al., 2018) y una fuente de alimento (Aguilar et al., 2017).
Considerando lo anterior, el estudio y monitoreo de los estróbilos masculinos y femeninos durante la etapa de polinización es esencial para comprender los procesos reproductivos en Zamiaceae. Si bien, la exclusión de sus insectos polinizadores reduce drásticamente el éxito reproductivo (Terry et al., 2008), los vertebrados también podrían desempeñar un papel relevante como transportadores potenciales de polen, contribuyendo al proceso de reproducción. En este contexto, es crucial explorar la composición de la comunidad de vertebrados que interactúan con las plantas del complejo D. edule, interacciones que podrían repercutir en la polinización. El estudio de estas interacciones es fundamental para desarrollar estrategias efectivas que promuevan la conservación íntegra y prioritaria de Zamiaceae y sus interacciones bióticas (Terry, 2005), especialmente en un grupo con alto riesgo de extinción. Nos planteamos el objetivo de determinar la diversidad de vertebrados visitantes y las interacciones que estos mantienen con los estróbilos masculinos y femeninos de las plantas del complejo D. edule Lindl. Zamiaceae (D. edule y D. angustifolium). El estudio se llevó a cabo durante el periodo de reproducción en 2 localidades de la sierra Madre Oriental.
Materiales y métodos
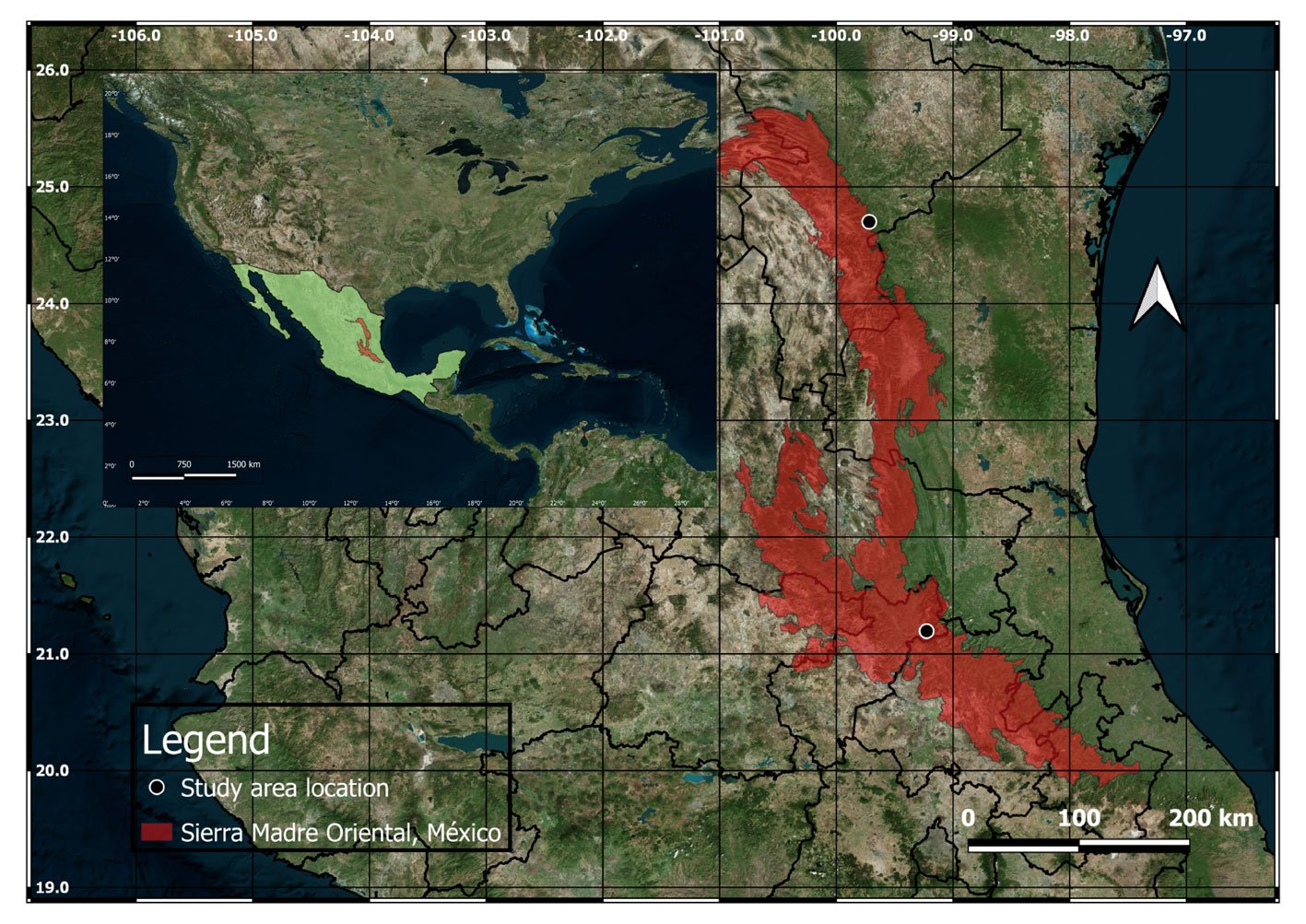
Para D. edule se seleccionó el municipio de Pisaflores, Hidalgo, que presenta un clima predominantemente tropical lluvioso, con una temperatura media anual de 26 °C. La vegetación está compuesta principalmente por bosque mesófilo de montaña, bosque de pino-encino, selva baja caducifolia y matorral xerófilo (Luna-Vega, 1997). Para D. angustifolium se seleccionó el municipio de Linares, Nuevo León, cuyo clima es principalmente semicálido subhúmedo con lluvias en verano. Con una temperatura media anual de 22.4 °C. Su vegetación está compuesta, principalmente, por matorral espinoso tamaulipeco, bosque de encino, selva baja caducifolia y pastizal (INEGI Marco Geoestadístico 2010, Versión 4.3). La localización geográfica exacta se reserva, para proteger las especies de amenazas como el tráfico ilegal o la actividad humana destructiva.
En los municipios antes mencionados se realizaron recorridos durante la etapa de polinización de las plantas del complejo D. edule en las áreas donde está reportada su presencia. Para este estudio, un sitio se definió como aquel punto que contara con la presencia de un par de plantas con el estróbilo en su periodo de polinización, se seleccionaron 10 sitios totales. Las coordenadas de las localidades de estudio fueron representadas en un mapa de la sierra Madre Oriental, el cual fue elaborado empleando el software QGIS 3.32.3 (QGIS Development Team, 2023) (fig. 1).
Una vez definidos y seleccionados los sitios para la observación de visitantes, se colocó una cámara trampa UOVision, modelo UV557 (fig. 2). Cada cámara estuvo enfocando y cubriendo la totalidad del estróbilo hacia cada una de las plantas (10 cámaras por sitio). El muestreo se realizó entre septiembre y noviembre de 2023 para D. angustifolium y entre agosto y octubre de 2023 para D. edule. Las cámaras fueron configuradas con modo video en formato VGA, resolución alta (8 megapixeles), sensibilidad normal y tiempo de filmación de 1 min. Analizados los videos, que representan casi 3 h de filmación en archivos de 1 min de duración, se realizó la identificación manual de éstos, utilizando los parámetros estándar y caracteres específicos descritos en las claves de identificación para campo y laboratorio de Álvarez-Castañeda (2017), Balderas y González-Hernández (2021) y Perlo (2006).
Para determinar a categoría de especie los ejemplares de roedores observados, fue necesario capturarlos y con base en los protocolos establecidos por Mills et al. (1998) y la guía técnica de Álvarez-Castañeda (2017), se colocó un transecto horizontal de 60 trampas Sherman plegables, no letales (HB Sherman traps, Tallahassee, FL, EE. UU.) con dimensiones de 23 x 8 x 9 cm, 60 en cada localidad, para un total de 120. Las trampas fueron cebadas con avena integral y esencia de vainilla, orientadas hacia la zona que, de acuerdo con lo observado en los videos, corresponde a la zona con mayor actividad de roedores, cubriendo los sitios de estudio y con una separación de 5 m entre ellas. Las trampas fueron colocadas por la noche y revisadas a la mañana siguiente, durante 3 días. Para su posterior identificación, cada uno de los ejemplares fue medido, fotografiado y liberado en el mismo punto de recolecta.

Análisis de datos
Primeramente, se analizaron las variables para evitar la colinealidad, excluyendo las que la presentaban, para posteriormente construir una matriz de datos de las observaciones de las especies registradas en cada uno de los sitios de estudio. La matriz fue sometida a un análisis con el software Xlstat 2020.3.1.1005 y con la finalidad de reducir el número de variables mediante la transformación de las variables originales en nuevas variables que son combinaciones lineales de las originales. Se tomaron como etiquetas de las variables los nombres de las especies y los sitios de estudio. Al ser variables discretas, se seleccionó una matriz de covarianza, para finalmente obtener los gráficos biplot de covarianza.
Para comparar la similitud de las comunidades de estudio respecto a sus especies visitantes y utilizando como criterio el coeficiente de Jaccard (Jaccard, 1901), se analizó la similitud de las comunidades respecto a las especies visitantes en cada uno de sus sitios, a través de un análisis de agrupamiento aglomerativo jerárquico (CAJ) con el software Xlstat (Addinsoft, 3.1.1005, 2020). Se seleccionaron como etiquetas de las variables a los sitios de estudio y las abundancias de cada especie registrada bajo un método de aglomeración de vinculación completa, generando así un dendrograma para la observación gráfica.
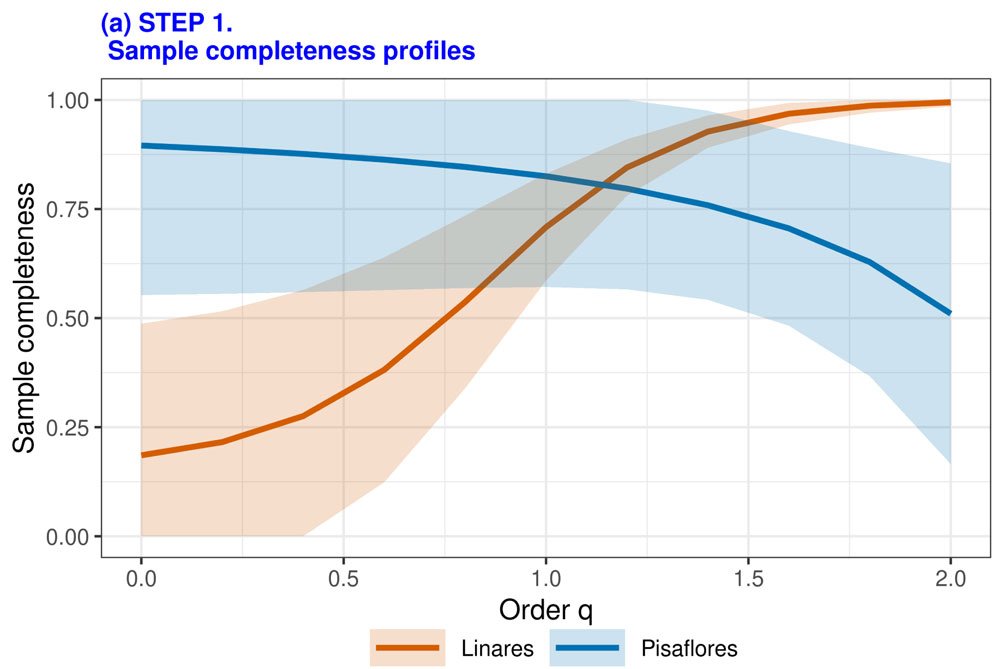
Para evaluar y comparar la biodiversidad y completitud de muestras de las localidades y los sitios de estudio, se emplearon los números de Hill. Las abundancias en las localidades fueron analizadas bajo el software iNext (Hsieh et al., 2016) utilizando el procedimiento de 4 pasos: 1) cálculo de los perfiles de completitud de la muestra; 2) rarificación/extrapolación basada en el tamaño de muestra; 3) aplicación de rarificación/extrapolación basada en cobertura, y 4) generación de perfiles de equidad.
Asimismo, para comparar la similitud de especies visitantes entre las localidades de estudio, se utilizó el índice de diversidad beta de Sorensen (Sorensen, 1948), evaluado mediante el software Past versión 4.03 (Hammer et al., 2001).
La clasificación de los tipos de interacción se realizó mediante la observación de las conductas de cada uno de los visitantes a ambos estróbilos, modificando y aumentando las categorías de Acevedo-Quintero et al. (2016), y se obtuvieron las siguientes clasificaciones. Neutro: visitar cualquier parte de la planta, pero no el estróbilo; trepar: subir al ápice del estróbilo; vuelo: volar alrededor del estróbilo; recorrido: recorrer o pasar por la base del estróbilo; salto: saltar sobre el ápice del estróbilo; percha: posarse sobre el ápice del estróbilo; picoteo: picotear entre las escamas de los estróbilos; depredación: morder el estróbilo parcial o totalmente.
Resultados
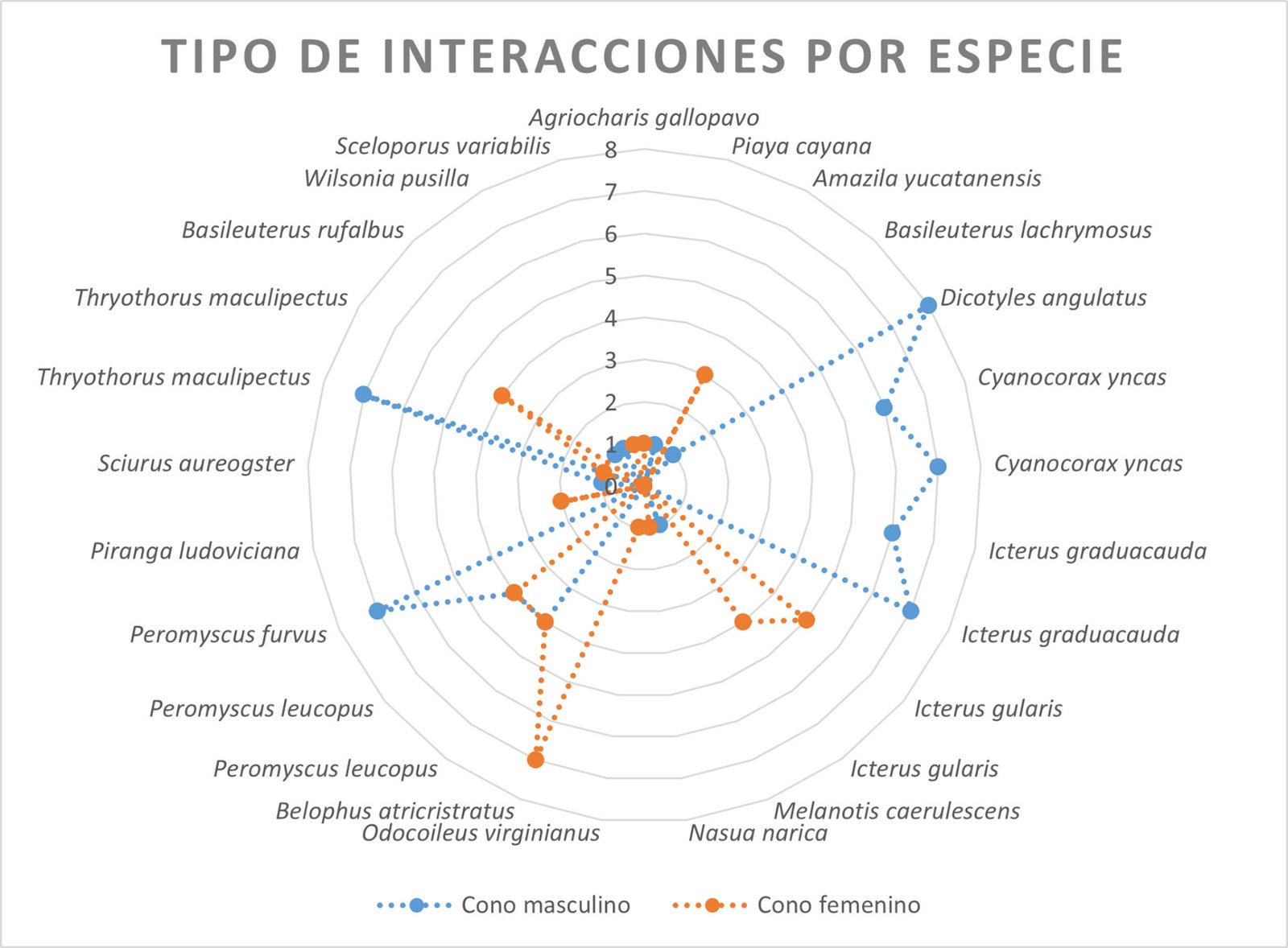
Se obtuvo el registro de 20 especies (figs. 3, 4) sumando las 2 localidades de estudio ubicadas en la sierra Madre Oriental; 13 aves, 6 mamíferos y 1 reptil. Se tuvieron un total de 49 visitas a los estróbilos (tabla 1), donde el ratón Peromyscus leucopus fue la especie con mayor frecuencia (38.78%), seguido del ave Thryothorus maculipectus (14.29%). Asimismo, se observó la presencia de invertebrados en ambos estróbilos que incluyen miembros de los órdenes Coleoptera, Hymenoptera y Orthoptera.
Tras el análisis de interacciones de las especies registradas, el cono masculino presentó un mayor número de interacciones (15) en comparación con el femenino (11); donde Peromyscus furvus y T. maculipectus fueron las únicas especies que presentaron actividad con ambos tipos de cono (fig. 5). Las interacciones de tipo neutro y picoteo fueron las más frecuentes en el cono masculino y las de tipo neutro y recorrido para el cono femenino. De esta forma, los grupos con mayor número de interacción con los estróbilos de D. angustifolium y D. edule fueron las aves y los mamíferos, respectivamente.


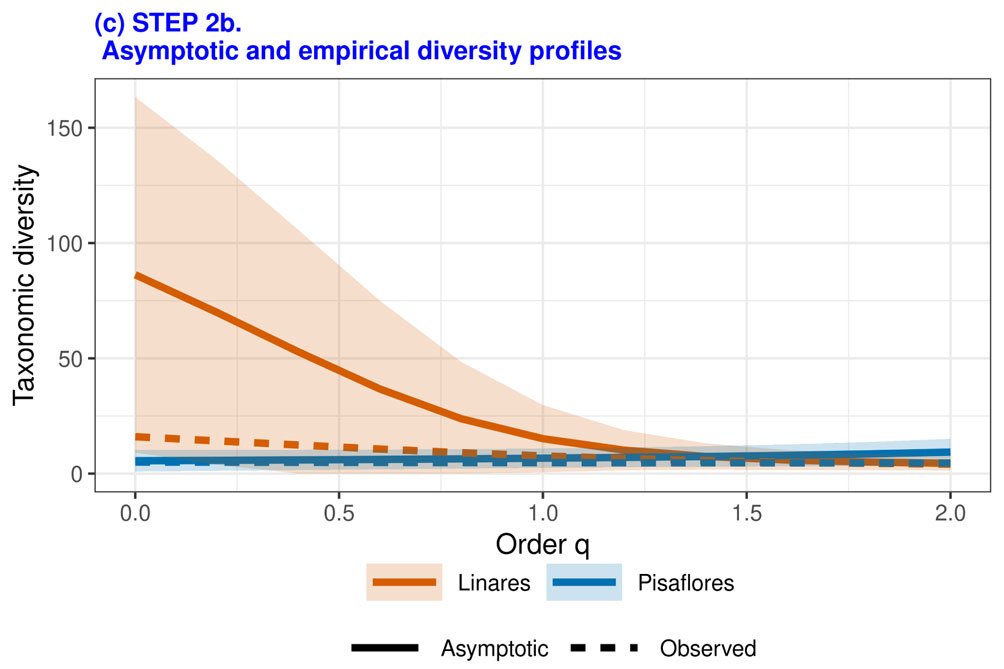
Aunque los índices Shannon de diversidad alfa de ambas localidades mostraron valores similares, el índice de diversidad en Linares (2.03) resultó superior al de Pisaflores (1.55). El índice de diversidad beta de Sorensen resultó alto (0.90). La completitud de las muestras varió con base en el orden de sensibilidad para especies raras (q), resultando Pisaflores con una menor completitud respecto a Linares, especialmente cuando q presenta valores bajos (fig. 6).
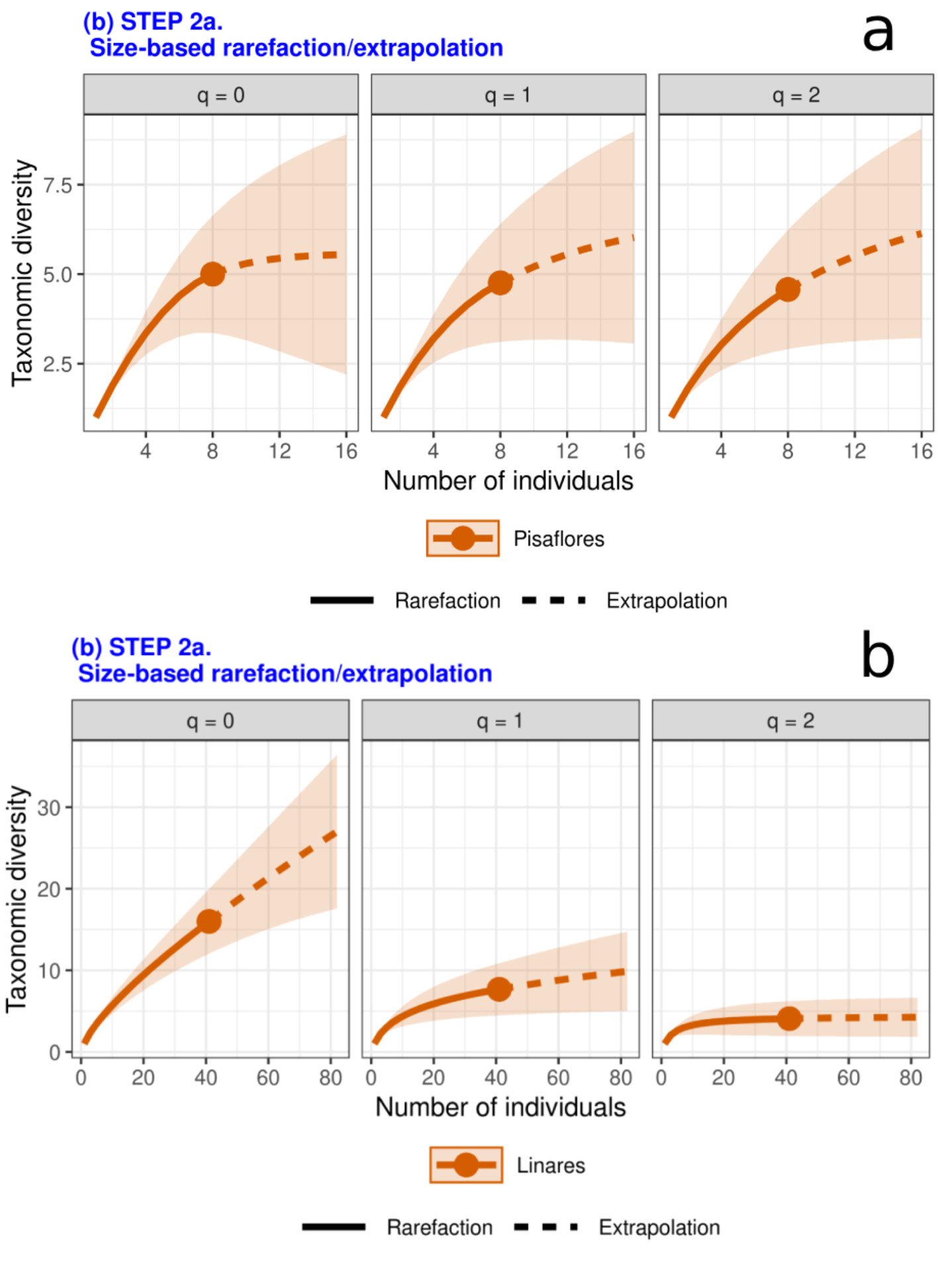
Rarificación/extrapolación con base en tamaño de la muestra (paso 2). Se presenta un incremento de la diversidad taxonómica cuando el tamaño muestral aumenta, Linares presentó mayores valores de biodiversidad, respecto a todos los niveles de q en comparación a Pisaflores (fig. 7a, b). La diversidad taxonómica presentó una diferencia entre las localidades, resultando Linares con una mayor diversidad estimada asintótica (fig. 8).
Rarificación/extrapolación basada en cobertura. Se observa un ajuste en la diversidad taxonómica respecto a la cobertura de la muestra. La localidad de Linares presenta una mayor diversidad para una misma cobertura en comparación a Pisaflores. En cuanto a la uniformidad en la distribución o equidad de especies, ésta presenta una disminución cuando el orden q incrementa. La localidad de Pisaflores muestra una menor equidad en comparación con Linares.
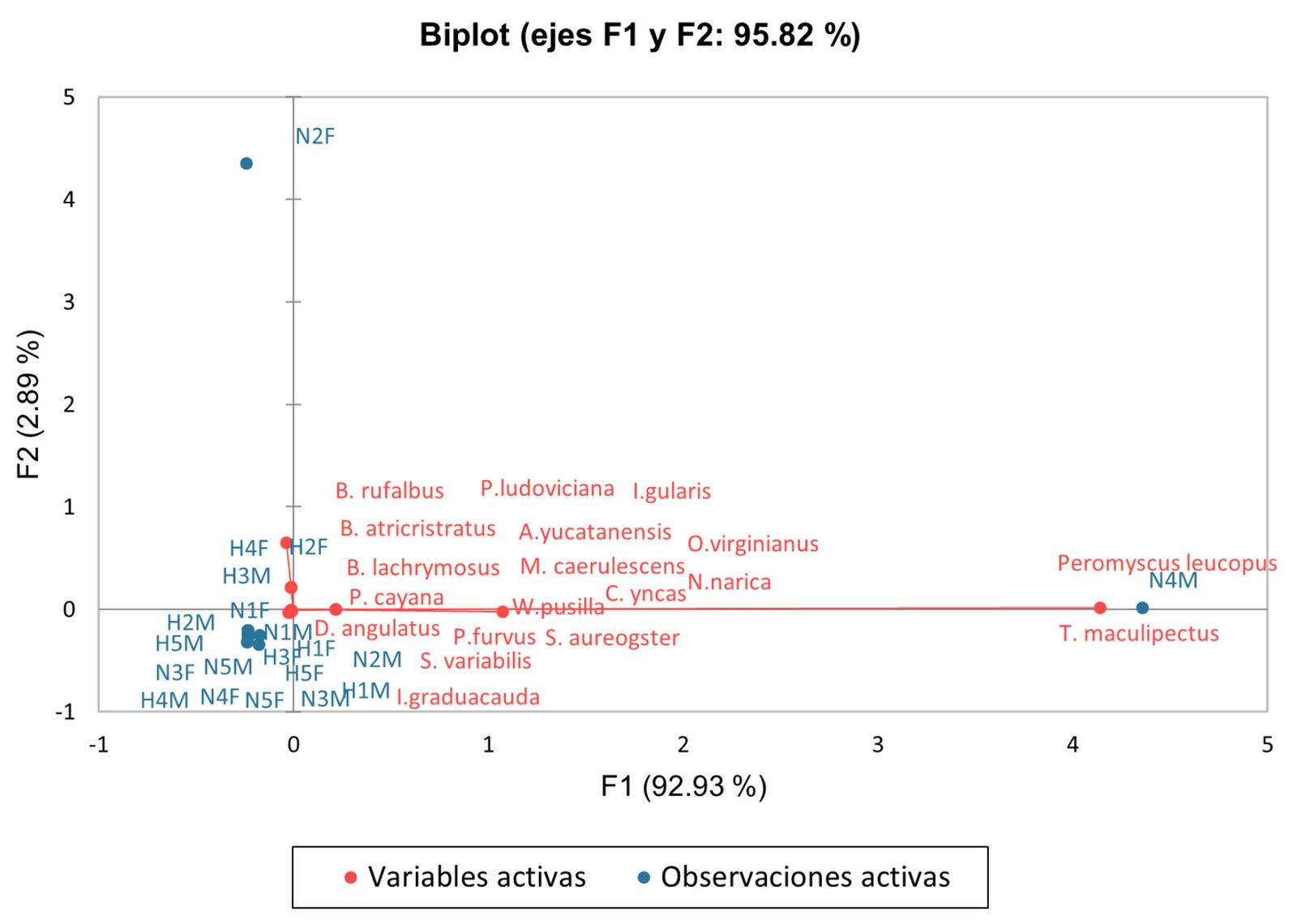
Valores resultantes de los números de Hill. Con valores para q,0 = 27.01, q,1 = 9.88 y q,2 = 4.25. La localidad de Linares presentó una mayor riqueza de especies y diversidad en general, mientras que Pisaflores con q,0 = 4.76, q,1 = 4.50, q,2 = 4.28 muestra una abundancia relativa ligeramente mayor en la distribución de las especies más abundantes. Los gráficos biplot del PCA muestran la distribución de las variables a lo largo de los ejes F1 (92.93%) y F2 (2.88%), los cuales explican, conjuntamente, 98.82% de la varianza total. Dado el carácter exploratorio del PCA y el tamaño reducido de muestra, los resultados se interpretan como un resumen de los patrones generales de asociación entre variables y sitios, por lo que continuamos realizando estudios incluyendo más sitios (fig. 9).
Discusión
La alta diversidad de visitantes e interacciones que se registraron en el presente estudio, establecen un precedente significativo al destacar el papel de los vertebrados en el proceso de polinización de plantas como las cícadas. Este proceso es considerado principalmente por invertebrados en diversas regiones del mundo (Hall y Walter, 2013). Esto se debe a que los estróbilos masculinos y femeninos representan un recurso alimenticio. Sin embargo, nuestros hallazgos coinciden con lo reportado en otras investigaciones que señalan el rol de vertebrados como transportadores de polen, el cual podría estar siendo subestimado (Hooks y Espindola, 2021).
Los resultados del presente estudio sugieren que el proceso de polinización en las plantas del complejo D. edule es más complicado que lo previamente pensado. El proceso involucra tanto factores abióticos (viento) como bióticos (vertebrados e invertebrados). Las interacciones de fauna con estróbilos representan redes complejas (Sounders, 2017). Con base en nuestros resultados y observaciones, se puede indicar que la finalidad de los vertebrados podría ser la búsqueda de una fuente de alimento en los estróbilos. En particular, los estróbilos albergan la variedad de invertebrados que forman parte de la dieta de vertebrados. El posarse, frotarse o acercarse a los estróbilos masculinos llenos de polen, podría provocar la adherencia al pelo, escamas o plumas. Posteriormente, este polen puede ser transportado durante la visita al cono femenino.
La posición de las variables N4M y P. leucopus en el espacio del PCA es consistente con la alta frecuencia de visita del ratón P. leucopus únicamente al estróbilo masculino de D. angustifolium, lo cual coincide con los patrones observados en los registros de interacción. Esto sugiere que P. leucopus podría contribuir al alejamiento del polen, en lugar de facilitar la transferencia a los estróbilos femeninos.
| Nombre científico | Familia | Frec # | Frec % | |
|---|---|---|---|---|
| Aves | Agriocharis gallopavo | Phasianidae | 1 | 2.04 |
| Amazila yucatanensis | Trochilidae | 1 | 2.04 | |
| Basileuterus lachrymosus | Parulidae | 1 | 2.04 | |
| Basileuterus rufalbus | Parulidae | 3 | 6.12 | |
| Baeolophus atricristratus | Trogonidae | 1 | 2.04 | |
| Cyanocorax yncas | Corvidae | 1 | 2.04 | |
| Icterus graduacauda | Icteridae | 2 | 4.08 | |
| Icterus gularis | Icteridae | 1 | 2.04 | |
| Melanotis caerulescens | Mimidae | 1 | 2.04 | |
| Piaya cayana | Cuculidae | 1 | 2.04 | |
| Piranga ludoviciana | Cardinalidae | 1 | 2.04 | |
| Thryothorus maculipectus | Troglodytidae | 7 | 14.29 | |
| Wilsonia pusilla | Parulidae | 1 | 2.04 | |
| Mamíferos | Dicotyles angulatus | Tayassuidae | 1 | 2.04 |
| Nasua narica | Procyonidae | 1 | 2.04 | |
| Odocoileus virginianus | Cervidae | 1 | 2.04 | |
| Peromyscus furvus | Cricetidae | 2 | 4.08 | |
| Peromyscus leucopus | Cricetidae | 19 | 38.78 | |
| Sciurus aureogaster | Sciuridae | 1 | 2.04 | |
| Reptiles | Sceloporus variabilis | Phrynosomatidae | 2 | 4.08 |
| Total | 49 | 100.0 |
Visitantes con potencial polinizador, adaptación ecológica y diversidad
Las cícadas mantienen una relación con los vertebrados actuales, además de emplear estrategias de densidad que favorecen la cercanía entre estróbilos masculinos y femeninos (Hall y Walter, 2013). De este modo, dada la alta frecuencia de visitantes como T. maculipectus a las plantas del complejo D. edule, podemos identificarlos como visitantes con potencial polinizador, al transportar granos de polen entre estróbilos, similar al papel que desempeñan las lagartijas Podarcis lilfordi, que transportan polen entre los estróbilos de la gimnosperma Ephedra fragilis (Ephedraceae) (Fuster y Traveset, 2019).
Los granos de polen transportados son posteriormente introducidos por los invertebrados que habitan los estróbilos de la cícada, aprovechando sus estructuras especializadas y completando así el proceso de polinización (Tang et al., 2018). Al considerar la cercanía de los estróbilos y los visitantes observados, se puede argumentar que tras perder sus relaciones con la megafauna, las cícadas han ido adoptando estrategias para aprovechar a la fauna actual, asegurando así la obtención de éste y otros beneficios ecológicos que les han permitido prevalecer.
Los valores de Hill muestran que la localidad de Nuevo León presenta una mayor riqueza y diversidad general, así como una mayor equidad en la distribución de especies. Además, al considerar el análisis de 4 pasos, Nuevo León exhibe niveles superiores de equidad y diversidad en la mayoría de los análisis, lo que sugiere que esta localidad tiene una comunidad más diversa y uniforme. En Nuevo León, una mayor variedad de especies interactúa con los estróbilos de D. angustifolium. Esto sugiere que los recursos que estos ofrecen durante la temporada de polinización atraen a una gran variedad de especies, independientemente de su abundancia.
La alta diversidad en la comunidad faunística de la localidad de Nuevo León incrementa la complejidad de las interacciones bióticas y la probabilidad de que algunos vertebrados actúen como visitantes con potencial polinizador. Asimismo, la elevada riqueza de especies respalda la idea de que las plantas del complejo D. edule dependen de faunas locales generalistas, lo cual podría conferir una mayor resiliencia frente a cambios ambientales. En contraste, en la localidad de Hidalgo, los niveles de equidad indican que los estróbilos de la especie D. edule son visitados por menos especies, pero de forma más consistente. Esto implica la posible dependencia de un número reducido de visitantes frecuentes, así como una mayor vulnerabilidad ante la pérdida de especies clave.
Thryothorus maculipectus surge como la única especie presente en los estróbilos de ambas especies. Esto indica que, aunque las localidades de estudio comparten 12 de las 20 especies registradas por las cámaras, las especies del complejo D. edule tienen visitantes mayormente distintos. Esta diferencia en la composición de fauna visitante se corrobora con el alto índice de diversidad beta, que evidencia una clara disimilitud entre las localidades de estudio y sugiere un remplazo de especies a lo largo de un gradiente espacial (Koleff et al., 2003). Los resultados obtenidos indican que, para sus interacciones, el complejo D. edule depende de faunas locales residentes, al contar con agentes bióticos con potencial de polinización de un carácter generalista, con respecto al hábitat.
La diferencia entre las especies de visitantes a cada especie del complejo D. edule, si bien parece ser distinta en cuanto a su diversidad, no lo es en cuanto a la variación de rasgos funcionales, la mayoría de ellos se limitaron a pequeñas aves y mamíferos. Esta diferencia no se podría sugerir como una asociación especialista como la observada en casos de angiospermas como Salvia plurispicata (Lamiaceae) con abejas o Salvia elegans (Lamiaceae) con colibríes (Espino et al., 2014), sino más bien un simple reemplazamiento por fauna residente con rasgos más generales que aprovecha los recursos de la planta como en el caso de la lagartija P. lilfordi con la gimnosperma E. fragilis (Fuster y Traveset, 2019). El fenómeno de reemplazamiento para las especies del complejo D. edule podría ser consecuencia de la separación biogeográfica que generó su diferenciación, propuesta por Gonzalez et al. (2005).
Los hallazgos aportan evidencia de que los estróbilos masculinos de cícadas también pueden ser consumidos por Dicotyles angulatus (fig. 4b), un fenómeno anecdótico que hasta ahora solo había sido reportado con evidencia para las semillas del estróbilo femenino, por lo cual estamos realizando más monitoreos que refuercen esta evidencia. Este hecho resulta relevante, ya que podría repercutir en el proceso de polinización. La depredación del estróbilo masculino podría limitar la interacción de otros visitantes con potencial polinizador.
Estos resultados proporcionan un punto de partida para investigaciones futuras sobre el valor nutricional del estróbilo. Asimismo, amplían el conocimiento sobre las interacciones ecológicas entre estas gimnospermas y vertebrados, y subrayan la importancia de considerar a los conos masculinos en los análisis de dinámicas de consumo de las cícadas.
Efectividad de los visitantes como vectores de polinización
Los roedores P. furvus y P. leucopus actúan como un visitante con potencial polinizador y el visitante más frecuente, respectivamente, del cono masculino de D. angustifolium. Este hallazgo coincide con estudios previos, como los de Balderas (2021), en los que los roedores fueron también identificados como los principales visitantes y dispersores de semillas de D. edule. Se ha descrito que los roedores pueden recoger polen en su pelaje mientras se alimentan de materia vegetal y actúan como polinizadores accidentales (Wester et al., 2024), lo que también se observó en esta investigación con los estróbilos de ambas especies de plantas del complejo D. edule. Sin embargo, la alta frecuencia de visita de P. leucopus únicamente al estróbilo masculino, representa un impacto que podría ser ambivalente, entre la polinización y el robo de polen.
Al resultar las aves el grupo taxonómico más diverso que visitó las plantas, interactuando a través de recorridos, perchas y picoteos dentro de ambos estróbilos y considerando que son reconocidas como importantes dispersoras de semillas en cícadas (Symes, 2018), surge una hipótesis que no solo las sugiere como dispersoras, sino también como los visitantes con potencial polinizador más probables para el complejo D. edule. Al ser organismos alados, las aves podrían transportar el polen a distancias y altitudes significativamente mayores en comparación con los organismos terrestres.
Ante los resultados y observaciones anteriores, surge la necesidad de ampliar el número de investigaciones centradas en la capacidad de los vertebrados para transportar efectivamente el polen de las plantas del complejo D. edule, así como comparar la contribución significativa al proceso de polinización entre los grupos de fauna que visitan las especies del complejo. Dado el alarmante índice de extinción entre los mamíferos y los vertebrados en general (Ceballos y Ehrlich, 2002; Ceballos et al., 2017), la disminución de estas especies y sus interacciones representa una amenaza significativa para la supervivencia de las poblaciones del complejo D. edule.
Una alta diversidad de vertebrados, incluyendo aves (13 especies), mamíferos (6 especies) y reptiles (1 especie), interactúan con los conos femeninos y masculinos de D. edule y D. angustifolium durante su fase de polinización, en esta etapa, los estróbilos representan una fuente indirecta de alimento que atrae un mayor número de interacciones, sobre todo en el estróbilo masculino. Las interacciones bióticas entre plantas y fauna son esenciales para la persistencia de las cícadas, estas plantas aprovechan la fauna actual para mantener interacciones que, en combinación con estrategias como la formación de rodales de alta densidad, favorecen la polinización debido a la proximidad entre estróbilos.
Este estudio genera antecedentes importantes para considerar la probable interferencia de vertebrados en el proceso de polinización de plantas del complejo D. edule.
Es fundamental ampliar las investigaciones sobre interacciones entre fauna y gimnospermas para comprender, a una escala mayor, las relaciones ecológicas entre estos organismos y contribuir a su preservación, especialmente en grupos en riesgo de extinción como las Zamiaceae.
Agradecimientos
Agradecemos al Conahcyt-México por financiar el proyecto CBF2023-2024-2780, Ciencia Básica y de Frontera 2023-2024. A Leslie Lugo, Ulises Casarubia, Luis Porras, Uriel Martínez, Sebastián Salazar, Alondra Alvarado y David Balderas por su apoyo en el trabajo técnico de campo. A Romeo Tinajero por el asesoramiento en la identificación de aves. A Rafael, Jesús y José Ángel Briones por permitirnos el acceso a los sitios. A los dos revisores anónimos cuyas observaciones contribuyeron a mejorar el manuscrito.
Referencias
Acevedo-Quintero, J. F. y Zamora-Abrego, J. G. (2016). Role of mammals on seed dispersal and predation processes of Mauritia flexuosa (Arecaceae) in the Colombian Amazon. Revista de Biología Tropical, 64, 5–15. https://doi.org/10.15517/rbt.v64i1.18157
Aguilar, P. O., Rivera-Fernández, A., Iglesias-Andreu, L. G., Vovides, P. A. y Núñez de Cáceres-González, F. F. (2017). Extinction risk of Zamia inermis: a demographic study in its single natural population. Biodiversity and Conservation, 26, 787–800. https://doi.org/10.1007/s10531-016-1270-z
Álvarez-Castañeda, S. T., Álvarez, T. y González-Ruiz, N. (2017). Keys for identifying Mexican mammals. Baltimore: Johns Hopkins University Press. https://doi.org/10.1353/book.50028
Balderas-González, D. (2021). Depredación y dispersión de semillas de Dioon edule por fauna nativa en tres localidades de la Sierra Madre Oriental (Nuevo León, Tamaulipas, San Luis Potosí) (Tesis de maestría). Universidad Autónoma de San Luis Potosí. San Luis Potosí, México.
Balderas-Valdivia, C. J. y González-Hernández, A. (2021). Inventario de la herpetofauna de México 2021. Herpetología Mexicana, 2, 10–71.
Bascompte, J. (2009). Disentangling the web of life. Science, 325, 416–419. https://doi.org/10.1126/science.1170749
Cai, C., Escalona, H. E., Li, L., Yin, Z., Huang, D. y Engel, M. S. (2018). Beetle pollination of cycads in the Mesozoic. Current Biology, 28, 2806–2812. https://doi.org/10.1016/j.cub.2018.06.036
Campbell, D. R. (1989). Measurements of selection in a hermaphroditic plant: variation in male and female pollination success. Evolution, 43, 318–334. https://doi.org/10.2307/2409210
Cárdenas-Cajamarca, J. D. (2017). Contribución de aves y mamíferos en la polinización de Oreocallis grandiflora (Lam.) R. Br. (Proteacea) en un matorral montano andino del sur de Ecuador (Tesis). Facultad de Ciencias y Tecnología, Universidad del Azuay. Cuenca, Ecuador.
Ceballos, G. y Ehrlich, P. R. (2002). Mammal population losses and the extinction crisis. Science, 296, 904–907. https://doi.org/10.1126/science.1069349
Ceballos, G., Ehrlich, P. R. y Dirzo, R. (2017). Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proceedings of the National Academy of Sciences, 114, E6089–E6096. https://doi.org/10.1073/pnas.1704949114
Celedón-Neghme, C., Santamaría, L. y González-Teuber, M. (2016). The role of pollination drops in animal pollination in the Mediterranean gymnosperm Ephedra fragilis (Gnetales). Plant Ecology, 217, 1545–1552. https://doi.org/10.1007/s11258-016-0667-9
Chamberlain, C. J. (1919). The living cycads. Chicago, Illinois: The University of Chicago Press.
Chamberlain, C. J. (1937). Gymnosperms: structure and evolution. Chicago, Illinois: The University of Chicago Press.
Chemnick, J. y Gregory, T. (2022). Dioon angustifolium, the IUCN red list of threatened species 2022. https://dx.doi.org/10.2305/IUCN.UK.2022-1.RLTS.T178868A243384900.en
Chemnick, J. y Gregory, T. (2022). Dioon edule, the IUCN red list of threatened species 2022. https://dx.doi.org/10.2305/IUCN.UK.2022-1.RLTS.T42158A243385825.en
Crowley, B. E., Godfrey, L. R. y Irwin, M. T. (2011). A glance to the past: subfossils, stable isotopes, seed dispersal, and lemur species loss in southern Madagascar. American Journal of Primatology, 73, 25–37. https://doi.org/10.1002/ajp.20817
David, J. P., Manakadan, R. y Ganesh, T. (2015). Frugivoría y dispersión de semillas por aves y mamíferos en los bosques siempreverdes secos tropicales costeros del sur de la India: una revisión. Ecología Tropical, 56, 41–55.
Dorsey, B. L., Gregory, T. J., Sass, C. y Specht, C. D. (2018). Pleistocene diversification in an ancient lineage: a role for glacial cycles in the evolutionary history of Dioon Lindl. (Zamiaceae). American Journal of Botany, 105, 1512–1530. https://doi.org/10.1002/ajb2.1149
Espino-Espino, J., Rosas, F. y Cuevas-García, E. (2014). Variación temporal de visitantes florales en dos especies simpátricas de Salvia con floración simultánea y síndrome de polinización contrastante. Revista Mexicana de Biodiversidad, 85, 161– 166. https://doi.org/10.7550/rmb.35248
Fuster, F. y Traveset, A. (2019). Evidence for a double mutualistic interaction between a lizard and a Mediterranean gymnosperm, Ephedra fragilis. AoB Plants, 11, plz001. https://doi.org/10.1093/aobpla/plz001
González-Astorga, J., Vovides, A. P., Cruz-Angon, A., Octavio- Aguilar, P. y Iglesias, C. (2005). Allozyme variation in the three extant populations of the narrowly endemic cycad Dioon angustifolium Miq. (Zamiaceae) from North- eastern Mexico. Annals of Botany, 95, 999–1007. https://doi.org/10.1093/aob/mci106
González-Astorga, J., Vovides, A. P., Ferrer, M. M. y Iglesias, C. (2003). Population genetics of Dioon edule Lindl. (Zamiaceae, Cycadales): biogeographical and evolutionary implications. Biological Journal of the Linnean Society, 80, 457–467. https://doi.org/10.1046/j.1095-8312.2003.00257.x
González-Christen, A. (1990). Algunas interacciones entre Dioon edule (Zamiaceae) y Peromyscus mexicanus (Rodentia: Cricetidae). La Ciencia y el Hombre, 5, 77–92.
González, D., Vovides, A. P. y Bárcenas, C. (2008). Phylogenetic relationships of the Neotropical genus Dioon (Cycadales, Zamiaceae) based on nuclear and chloroplast DNA sequence data. Systematic Botany, 33, 229–236. https://doi.org/10.1600/036364408784571699
Gutiérrez-Ortega, J. S., Salinas-Rodríguez, M. M., Martínez, J. F., Molina-Freaner, F., Pérez-Farrera, M. A., Vovides, A. P. et al. (2018). The phylogeography of the cycad genus Dioon (Zamiaceae) clarifies its Cenozoic expansion and diversification in the Mexican transition zone. Annals of Botany, 121, 535–548. https://doi.org/10.1093/aob/mcx165
Hall, J. A. y Walter, G. H. (2013). Seed dispersal of the Australian cycad Macrozamia miquelii (Zamiaceae): Are cycads megafauna-dispersed “grove forming” plants? American Journal of Botany, 100, 1127–1136. https://doi.org/10.3732/ajb.1200115
Hammer, O., Harper, D. A. T. y Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Paleontologia Electrónica, 4, 1–9.
Herrera, C. M. y Pellmyr, O. (2009). Plant animal interactions: an evolutionary approach. Oxford, UK: Blackwell Science.
Herrera, J. (1987). Biología reproductiva de algunas especies del matorral de Doñana. Anales del Jardín Botánico de Madrid, 44, 483–497.
Hill, M. O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology, 54, 427–432. https://doi.org/10.2307/1934352
Hooks, C. y Espíndola, A. (2021). Rodents and other non- flying mammal pollinators. Maryland agronomy news. The University of Maryland Extension. Recuperado el 14 enero, 2025, de: https://blog.umd.edu/agronomynews/2021/05/
Hsieh, T. C., Ma, K. H. y Chao, A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods in Ecology and Evolution, 7, 1451–1456. https://doi.org/10.1111/2041-210X.12613
Jaccard, P. (1901). Estudio comparativo de la distribución floral en una porción de los Alpes y el Jura. Bulletin de la Societe Vaudoise des Sciences Naturelles, 37, 547–579.
Janzen, D. H. y Martin, P. S. (1982). Neotropical anachronisms: the fruits the gomphotheres ate. Science, 215, 19–27. https://doi.org/10.1126/science.215.4528.19
Jin, B., Zhang, L., Lu, Y., Wang, D., Jiang, X. X., Zhang, M. et al. (2012). The mechanism of pollination drop withdrawal in Ginkgo biloba L. BMC Plant Biology, 12, 1–9. https://doi.org/10.1186/1471-2229-12-59
Jones, M. M., Liu, J. y Nagalingum, N. S. (2024). Evidence of a historical metapopulation: conservation genomics and management strategies of Cycas candida (Cycadaceae). International Journal of Plant Sciences, 185, 361–369. https://doi.org/10.1086/729720
Jolliffe, I. T. (2002). Principal component analysis for special types of data. Principal component analysis (pp. 338–372). Nueva York: Springer New York. https://doi.org/10.1007/0-387-22440-8_13
Koleff, P., Gaston, K. J. y Lennon, J. J. (2003). Measuring beta diversity for presence-absence data. Journal of Animal Ecology, 72, 367–382. https://doi.org/10.1046/j.1365-2656.2003.00710.x
Lázaro-Zermeño, J. M., González-Espinosa, M., Mendoza, A. y Martínez-Ramos, M. (2012). Natural history of Dioon merolae (Zamiaceae) in Chiapas, Mexico. Botanical Sciences, 90, 73–87. https://doi.org/10.17129/botsci.387
Lovette, I. J. y Fitzpatrick, J. W. (2016). The Cornell Lab. of Ornithology handbook of bird biology, Third edition. Oxford, UK: John Wiley & Sons.
Luna-Vega, M. I. (1997). Florística y biogeografía de algunos bosques mesófilos de la Huasteca Hidalguense: Fase I (Tenango de Doria y Tlanchinol). Comisión Nacional Para el Conocimiento y uso de la Biodiversidad. Occurrence dataset https://doi.org/10.15468/sbkl18
Mills, J. N., Childs, J. E., Ksiazek, T. G., Peters, C. J. y Velleca, W. M. (1998). Métodos para trampeo y muestreo de mamíferos pequeños para estudios virológicos. Atlanta, Georgia: U.S. Department of Health & Human Services, Public Health Service, Centers for disease control and prevention. Recuperado el 15 marzo, 2025, de: https://stacks.cdc.gov/view/cdc/11583
Mora, R., Yáñez-Espinosa, L., Flores, J. y Nava-Zárate, N. (2013). Strobilus and seed production of Dioon edule (Zamiaceae) in a population with low seedling density in San Luis Potosí, México. Tropical Conservation Science, 6, 268–282. https://doi.org/10.1177/194008291300600208
Moretti, A., Caputo, P., Cozzolino, S., De Luca, P., Gaudio, L., Siniscalco Gigliano, G. y Stevenson, D. W. (1993). A phylogenetic analysis of Dioon (Zamiaceae). American Journal of Botany, 80, 204–214. https://doi.org/10.1002/j.1537-2197.1993.tb13790.x
Nicolalde-Morejón, F., González-Astorga, J., Vergara-Silva, F., Stevenson, D. W., Rojas-Soto, O. y Medina-Villarreal, A. (2014). Biodiversidad de Zamiaceae en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), S114–S125. https://doi.org/10.7550/rmb.38114
Norstog, K. y Nicholls, T. J. (2019). The biology of the cycads. Ithaca, NY: Cornell University Press. https://doi.org/10.7591/9781501737329
Nyhagen, D. F., Kragelund, C., Olesen, J. M. y Jones, C. G. (2001). Insular interactions between lizards and flowers: flower visitation by an endemic Mauritian gecko. Journal of Tropical Ecology, 17, 755–761. https://doi.org/10.1017/S0266467401001560
Osborne, R., Calonje, M. A., Hill, K. D., Stanberg, L. y Stevenson, D. W. (2012). The world list of cycads. Memoirs of the New York Botanical Garden, Vol 106 (pp. 480–510). Nueva York: New York Botanical Garden Press.
Owens, J. N., Takaso, T. y Runions, C. J. (1998). Pollination in conifers. Trends in Plant Science, 3, 479–485. https://doi.org/10.1016/S1360-1385(98)01337-5
Pérez-Farrera, M. A., Quintana-Ascencio, P. F., Salvatierra- Izaba, B. y Vovides, A. P. (2000). Population dynamics of Ceratozamia matudai Lundell (Zamiaceae) in El Triunfo Biosphere Reserve, Chiapas, Mexico. Journal of The Torrey Botanical Society, 127, 291–299. https://doi.org/10.2307/3088647
Pérez-Farrera, M. A., Vovides, A. P., Hernández-Sandoval, L., González, D. y Martínez, M. (2004). A morphometric analysis of the Ceratozamia norstogii complex (Zamiaceae). En T. Walters y R. Osborne (Eds.), Cycad classification: concepts and recommendations (pp. 127– 136). Wallingford, UK: CABI Publishing. Https://Doi. Org/10.1079/9780851997414.0127
Pérez-Farrera, M. A., Vovides, A. P., Octavio-Aguilar, P., González-Astorga, J., De la Cruz-Rodríguez, J., Hernández- Jonapá, R. et al. (2006). Demography of the cycad Ceratozamia mirandae (Zamiaceae) under disturbed and undisturbed conditions in a biosphere reserve of Mexico. Plant Ecology, 187, 97–108. https://doi.org/10.1007/s11258-006-9135-2
Perlo, B. V. (2006). Field guide to the birds of Mexico and Central America. Princeton, NJ: Princeton University Press.
Pianka, E. R. (1970). On r-and k-selection. The American Naturalist, 104, 592–597. https://doi.org/10.1086/282697
QGIS Development Team. (2023). QGIS Geographic Information System (Versión 3.32.3 [Software]. Open Source Geospatial Foundation. https://qgis.org
Ratto, F., Simmons, B. I., Spake, R., Zamora-Gutiérrez, V., MacDonald, M. A., Merriman, J. C. et al. (2018). Global importance of vertebrate pollinators for plant reproductive success: a meta-analysis. Frontiers in Ecology and the Environment, 16, 82–90.
Rivadeneyra-Domínguez, E. y Rodríguez-Landa, J. F. (2014). Las cícadas y su relación con algunas enfermedades neurodegenerativas. Neurología, 29, 517–522. https://doi.org/10.1016/j.nrl.2013.03.005
Rojas-Soto, O. R., Nicolalde-Morejón, F. y Yáñez-Arenas, C. (2014). Dioon edule (chamal). Registros de presencia usados para elaborar el mapa de distribución potencial. Proyecto: JM057. Distribución geográfica y ecológica potencial de las Cícadas de México. Xalapa: Instituto de Ecología, A.C. Recuperado el 13 de septiembre, 2024, de: http://geoportal.conabio.gob.mx/metadatos/doc/html/ded057rpgw.html
Rubio-Méndez, G., Yáñez-Espinosa, L. y Leija, E. G. (2020). Are the Dioon edule (Zamiaceae) forms from San Luis Potosí proposed by Whitelock (2004) recognizable? Morphological evidence. Revista Mexicana de Biodiversidad, 91, e913167. https://doi.org/10.22201/ib.20078706e.2020.91.3167
Rubio-Méndez, G., Yáñez-Espinosa, L., Salinas-Rodríguez, M. M., Hernández-Hernández, K. A. y Balderas-González, D. (2019). Demografía y estructura poblacional de Dioon angustifolium (Zamiaceae) en el noreste de México. Botanical Sciences, 97, 685–690. https://doi.org/10.17129/botsci.2262
Saunders, M. E. (2017). Insect pollinators collect pollen from wind-pollinated plants: implications for pollination ecology and sustainable agriculture. Insect Conservation and Diversity, 11, 13–31. https://doi.org/10.1111/icad.12243
Shannon, C. E. (1948). A mathematical theory of communication. The Bell System Technical Journal, 27, 379–423. https://doi.org/10.1002/j.1538-7305.1948.tb01338.x
Sorensen, T. (1948). A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biologiske Skrifter, 5, 1–34.
Stevenson, D. W. (1992). A formal classification of the extant cycads. Brittonia, 44, 220–223. https://doi.org/10.2307/2806837
Suinyuy, T. N., Donaldson, J. S., Johnson, S. D. y DeWet Bosenberg, J. (2012). Role of cycad cone volatile emissions and thermogenesis in the pollination of Encephalartos villosus Lem.: preliminary findings from studies of plant traits and insect responses. Memoirs of the New York Botanical Garden, Vol 106 (pp. 318–334). Nueva York: New York Botanical Garden Press.
Symes, C. T. (2018). Cycad seed dispersal: the importance of large frugivorous birds. Biodiversity Observations, 9, 9–4. https://doi.org/10.15641/bo.v9i0.503
Tang, W., Xu, G., O’Brien, C. W., Calonje, M., Franz, N. M., Johnston, M. A. et al. (2018). Molecular and morphological phylogenetic analyses of New World cycad beetles: what they reveal about cycad evolution in the New World. Diversity, 10, 38. https://doi.org/10.3390/d10020038
Terry, I., Forster, P. I., Moore, C. J., Roemer, R. B., & Machin, P. J. (2008). Demographics, pollination syndrome and conservation status of Macrozamia platyrhachis (Zamiaceae), a geographically restricted Queensland cycad. Australian Journal of Botany, 56, 321–332. https://doi.org/10.1071/BT06202
Terry, L. I., Walter, G. H., Donaldson, J. S., Snow, E., Forster, P. I. y Machin, P. J. (2005). Pollination of Australian Macrozamia cycads (Zamiaceae): effectiveness and behavior of specialist vectors in a dependent mutualism. American Journal of Botany, 92, 931–940. https://doi.org/10.3732/ajb.92.6.931
Varela, R. O. y Bucher, E. H. (2002). Seed dispersal by Chelonoidis chilensis in the Chalco Dry Woodland of Argentina. Journal of Herpetology, 36, 137–140. https://doi.org/10.1670/0022-1511(2002)036[0137:SDBCCI]2.0.CO;2
Viana, D. S., Santamaría, L. y Figuerola, J. (2016). Optimal methods for fitting probability distributions to propagule retention time in studies of zoochorous dispersal. BMC Ecology, 16, 3. https://doi.org/10.1186/s12898-016-0057-0
Vovides, A. P. (1990). Spatial distribution, survival, and fecundity of Dioon edule (Zamiaceae) in a tropical deciduous forest in Veracruz, Mexico, with notes on its habitat. American Journal of Botany, 77, 1532–1543. https://doi.org/10.1002/j.1537-2197.1990.tb11394.x
Vovides, A. P., Ogata, N., Sosa, V. y Peña-García, E. (1997). Pollination of endangered Cuban cycad Microcycas calocoma (Miq.) A. DC. Botanical Journal of the Linnean Society, 125, 201–210. https://doi.org/10.1111/j.1095-8339.1997.tb02254.x
Wester, P., Stanway, R. y Pauw, A. (2009). Mice pollinate the Pagoda Lily, Whiteheadia bifolia (Hyacinthaceae)— First field observations with photographic documentation of rodent pollination in South Africa. South African Journal of Botany, 75, 713–719. https://doi.org/10.1016/j.sajb.2009.07.005
Whitelock, L. M. (2004). Variation in the Mexican cycad Dioon edule (Zamiaceae). Botanical Review, 70, 240–249. https://www.jstor.org/stable/4354476
Yáñez-Espinosa, L. (2006). Las cycadas: biología y conservación en México. Ciudad de México: Universidad Autónoma Chapingo.
Yáñez-Espinosa, L., Balderas-González, D., Rubio-Méndez, G., Barragán-Torres, F. y Espinosa-Reyes, G. (2021). Dispersión de semillas de Dioon edule por aves. En H. Reyes-Hernández, J. A. Reyes-Agüero y A. Durán-Fernández (Eds.), Desafíos para la conservación de los ecosistemas y su biodiversidad en la Reserva de la Biosfera Sierra del Abra Tanchipa (pp. 38–48). San Luis Potosí, México: Universidad Autónoma de San Luis Potosí/ Conanp.
Yáñez-Espinosa, L., Barragán-Torres, F., Ibarra-Hernández, A. B. y Morales, J. I. (2021). Dispersal of Dioon edule cycad seeds by rodents in a tropical oak forest in Mexico. Tropical Zoology, 34, 57–71. https://doi.org/10.4081/tz.2021.105


