
Four new records of the genus Septoglomus (Glomeromycota) in Mexico
Reyna P. Hipólito-Piedras a, Heriberto Méndez-Cortés b, *, Hugo M. Ramírez-Tobias b, Victor Olalde-Portugal c, Carlos Arturo Aguirre-Salado d, Ángel Natanael Rojas-Velázquez b y Aracely Mena-Echevarríae
a Universidad Autónoma de San Luis Potosí, Facultad de Agronomía y Veterinaria, Doctorado en Ciencias Agropecuarias, Carretera San Luis Potosí-Matehuala Km. 14.5, Ejido Palma de la Cruz, 78321 Soledad de Graciano Sánchez, San Luis Potosí, México
b Universidad Autónoma de San Luis Potosí, Facultad de Agronomía y Veterinaria, Carretera San Luis Potosí- Matehuala Km. 14.5, Ejido Palma de la Cruz, 78321 Soledad de Graciano Sánchez, San Luis Potosí, México
c Instituto Politécnico Nacional, Centro de Investigación y de Estudios Avanzados-Unidad Irapuato, Departamento de Biotecnología y Bioquímica, Libramiento Norte Carretera Irapuato-León Km 9.6, 36824 Irapuato, Guanajuato, México
d Universidad Autónoma de San Luis Potosí, Facultad de Ingeniería, Av. Dr. Manuel Nava 4, 78290 San Luis Potosí, San Luis Potosí, México
e Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Campo Experimental General Terán-INIFAP, Km 31 Carretera Montemorelos-China, 67400 General Terán, Nuevo León, México
*Autor para correspondencia: heriberto.mendez@uaslp.mx (H. Méndez-Cortés)
Recibido: 13 febrero 2024; aceptado: 23 julio 2025
Resumen
Los hongos micorrizógenos arbusculares (HMA) son organismos esenciales y de suma importancia para las plantas terrestres. Se seleccionaron distintas comunidades vegetales de México en los estados de Coahuila, Guanajuato, San Luis Potosí, Tamaulipas y Veracruz, en donde se recolectó suelo rizosférico con el objetivo de aislar esporas de HMA. La identificación taxonómica se llevó a cabo mediante el análisis de las características morfológicas de las esporas, comparándolas con los artículos originales de cada especie descrita. Se identificaron 4 especies de HMA pertenecientes al género Septoglomus (S. altomontanum, S. fuscum, S. furcatum y S. turnauae), que constituyen su primer registro en México. Con estas 4 especies, la riqueza total de HMA en México se eleva a 171, lo cual evidencia una diversidad significativa de HMA en las comunidades vegetales del país.
Palabras clave: Glomeromycota; Micorriza arbuscular; Glomeraceae
Abstract
Arbuscular mycorrhizal fungi (AMF) are essential and extremely important organisms for terrestrial plants. Different plant communities of Mexico were selected in the states of Coahuila, Guanajuato, San Luis Potosí, Tamaulipas and Veracruz, where rhizospheric soil was collected with the objective of isolating AMF spores. Taxonomic identification was carried out by analyzing the morphological characteristics of the spores, comparing them with the original articles of each described species. Four species of AMF belonging to the genus Septoglomus (S. altomontanum, S. fuscum, S. furcatum, and S. turnauae) were identified, constituting their first record in Mexico. With the record of these 4 species, the total richness of AMF in Mexico rises to 171, evidencing a significant diversity of AMF in the plant communities of the country.
Keywords: Glomeromycota; Arbuscular mycorrhizae; Glomeraceae
Introducción
Los hongos micorrizógenos arbusculares (HMA) son organismos cruciales y de suma importancia en la ecología y fisiología de las plantas terrestres. Fueron considerados inicialmente en el filo Zygomycota; sin embargo, a través de sus características moleculares, morfológicas y ecológicas, fueron reorganizados en un nuevo filo monofilético denominado Glomeromycota (Schüßler et al., 2001). Actualmente, la clasificación taxonómica de los HMA incluye 3 clases, 4 órdenes y 43 géneros (Schüβler, 2025; Wijayawardene et al., 2020).
Uno de los géneros incluidos en Glomeromycota es Septoglomus. Originalmente, las especies que ahora se ubican en este género se agruparon dentro del género Glomus. Sin embargo, Schüßler y Walker (2010) las reubicaron en el género Funneliformis, basándose en criterios morfológicos. Posteriormente, Oehl et al. (2011) dividieron Glomus en 2 grupos taxonómicos, proponiendo los géneros Simiglomus y Septoglomus con base en análisis de secuencias ribosomales y en la morfología. Sin embargo, Redecker et al. (2013) validaron únicamente al género Septoglomus y recomendaron conservar su estatus en espera de mayor evidencia, particularmente a partir de estudios filogenéticos del DNAr.
La especie de referencia taxonómica para la clasificación del género Septoglomus fue Glomus constrictum (Trappe) Sieverd., GA Silva et Oehl (Glomeraceae) (actualmente, S. constrictum). A partir de ésta, se establecieron los criterios taxonómicos del género, que se caracterizan por la producción de esporas de manera individual o en pequeños grupos. Dichas esporas presentan una o múltiples capas y están conectadas a hifas que se extienden de forma continua hasta la pared de la espora, adoptan una forma cilíndrica o de embudo y presentan un poro cerrado por un septo ubicado en la base o debajo de la pared de la espora. Generalmente, estas esporas no reaccionan a la aplicación del reactivo de Melzer; sin embargo, sus estructuras adquieren una coloración azul oscuro al ser tratadas con azul de tripano (Oehl et al., 2011).
Actualmente, el género Septoglomus comprende 13 especies, lo que representa aproximadamente 4% de la diversidad total de HMA. Estas especies han sido documentadas en diversos estudios, como los de Błaszkowski et al. (2004, 2014, 2013, 2023), Goto et al. (2013), Symanczik et al. (2014), Chimal-Sánchez et al. (2020), Oehl et al. (2011, 2019), Palenzuela et al. (2013) y Guillén et al. (2020). Sin embargo, la clasificación de algunas de estas especies continúa en revisión. Por ejemplo, recientemente S. deserticola G.A. Silva, Oehl et Sieverd. y S. viscosum C. Walker, D. Redecker, D. Stille et A. Schüßler (Glomeraceae) fueron reclasificados en el nuevo género Blaszkowskia, con base en análisis filogenéticos y en su morfología distintiva (da Silva et al., 2023).
Las especies del género Septoglomus han sido registradas en diferentes continentes; en Europa, por Palenzuela et al. (2013), Blaszkowski et al. (2014), Oehl et al. (2019) y Guillén et al. (2020); en América, por Błaszkowski et al. (2013), Goto et al. (2013) y Chimal-Sánchez et al. (2020); en Asia, por Symanczik et al. (2014) y Oehl et al. (2019) y en África por Błaszkowski et al. (2010, 2013). El registro de estas especies fue en dunas marítimas, bosques lluviosos y tropicales, así como en matorrales xerófilos. No obstante, se ha observado que este género demuestra una destacada capacidad de adaptación y una dominancia notable en entornos áridos, en comparación con otros géneros de HMA. Además, muestra una eficiencia notable en la colonización de las raíces de cultivos agrícolas y forestales (Guardiola-Márquez et al., 2022).
En México se han identificado 4 especies del género Septoglomus. Sin embargo, es crucial llevar a cabo investigaciones a mayor escala, que abarquen tanto el ámbito ecológico como el morfológico para comprender mejor su distribución en el país. Asimismo, intensificar el muestreo para aumentar el conocimiento sobre la riqueza de especies de HMA en el territorio nacional (Polo-Marcial et al., 2021). El objetivo de esta investigación fue contribuir al conocimiento sobre la diversidad de Septoglomus mediante la documentación de 4 nuevos registros obtenidos en distintos tipos de vegetación de México.
Materiales y métodos
El muestreo del suelo rizosférico se realizó durante la época de lluvias (agosto, 2020-2021), en 6 sitios ubicados en 4 estados de la República Mexicana (fig. 1, tabla 1). En cada uno de los sitios se estableció un transecto de 1 km, donde cada 200 m se tomó una submuestra a una profundidad de 0 a 20 cm, la cual fue trasladada al laboratorio de Fitopatología, de la Facultad de Agronomía y Veterinaria, de la Universidad Autónoma de San Luis Potosí.
La extracción de esporas de HMA se realizó por el método de tamizado húmedo y decantación (Gerdemann y Nicolson, 1963), utilizando 500 g de suelo seco, seguido de una centrifugación en sacarosa al 70% a 2,000 rpm (Daniels y Skipper, 1982). Las esporas se montaron en preparaciones permanentes con alcohol polivinílico-lacto-glicerol (PVLG) y PVLG con reactivo de Melzer (INVAM, 2023).
La identificación se llevó a cabo de acuerdo con las características morfológicas de la espora, como lo sugieren da Silva et al. (2023). Así, se registraron el tamaño, las características de la pared de la espora y de la hifa de sostén, y la reacción al reactivo de Melzer. La observación de tales características fue en un microscopio óptico Zeiss Primo Star (Jena, Alemania) con iluminación fija Koehler.
Las mediciones de las características de las esporas de HMA se realizaron en fotografías tomadas con el software de microscopía ZEN blue (versión 3.4) (Jena, Alemania), con objetivos 40x y 100x. Tales características se contrastaron con lo descrito en las claves dicotómicas de Palenzuela et al. (2013) y Oehl et al. (2019); así como con los artículos de referencia (Blaszkowski et al., 2013, 2014; Palenzuela et al., 2013) y se elaboraron claves para la identificación de las especies. Los ejemplares de referencia fueron depositados en el herbario Isidro Palacios, perteneciente al Instituto de Investigación de Zonas Desérticas, de la Universidad Autónoma de San Luis Potosí (SLPM).
Descripciones
Se identificaron un total de 62 especies de HMA, en los 9 sitios de muestreo. De éstas, 21% pertenecen al género Acaulospora, 15% a Glomus, 11% a Septoglomus, 10% a Ambispora, 10% a Rhizophagus, 5% a Entrophospora, 5% a Funneliformis, 5% a Gigaspora, 5% a Scutellospora, 3% a Diversipora y 10% restante se distribuye en igual proporción entre los géneros Blazskowskia, Dominikia, Pacispora y Sieverdingia.
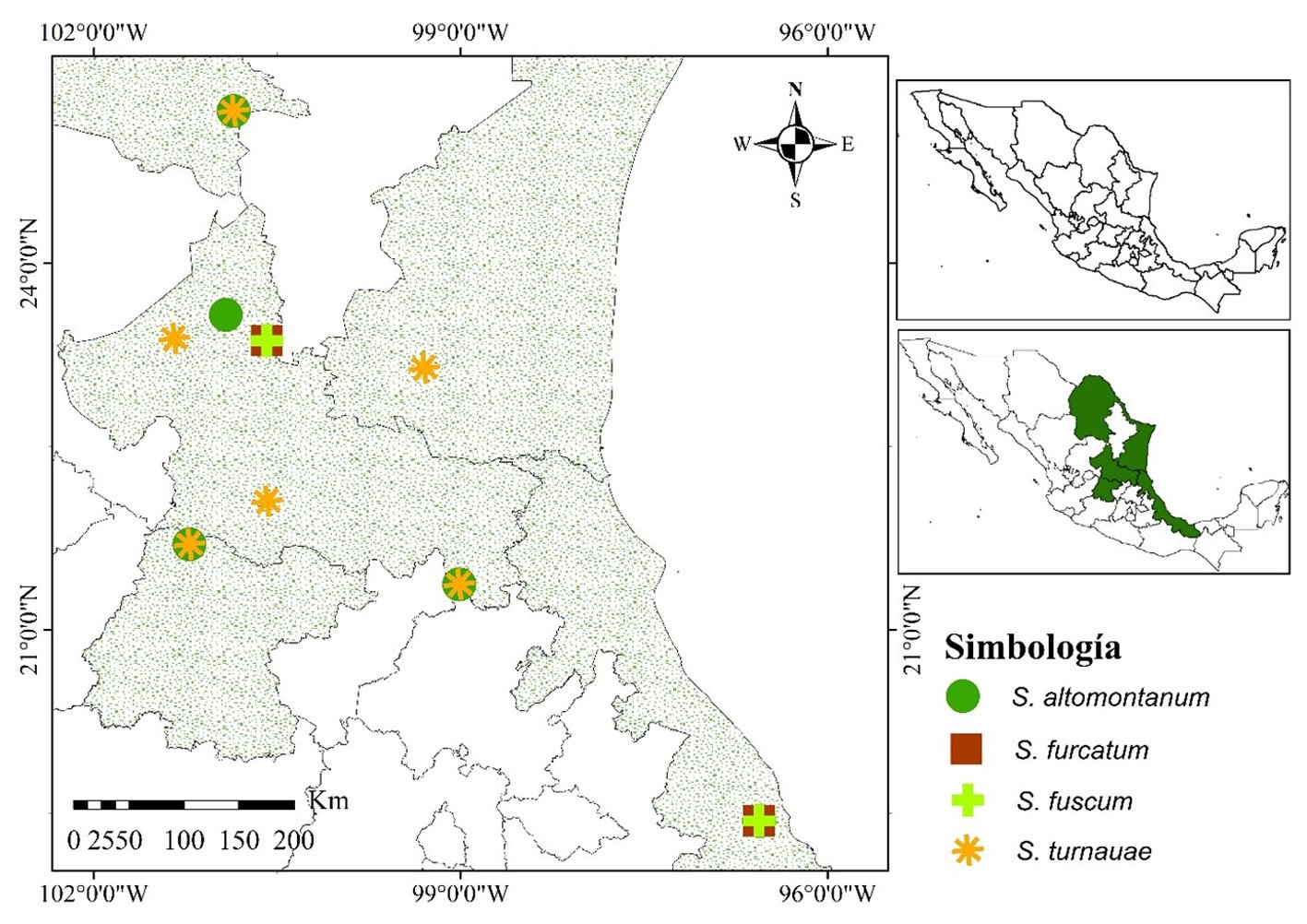
Figura 1. Distribución geográfica de sitios de muestreo y 4 especies del género Septoglomus en México.
Las especies registradas del género Septoglomus fueron: S. constrictum, S. mexicanum E. Chimal-Sánchez, C. Senés-Guerrero, NM Montaño, L. Varela, R. García-Sánchez, A. Pacheco, S. Montaño-Arias y SL Camargo-Ricalde, S. xanthium (Błaszk., Blanke, Renker y Buscot) GA Silva, Oehl y Sieverd., S. altomontanum Palenz., Oehl, Azcón-Aguilar y GA Silva, S. fuscum, S. furcatum Błaszk., Chwat, Kovács y Ryszka, y S. turnauae Błaszk., Chwat y Ryszka(Glomeraceae). Las últimas 4 son registro nacional, por lo cual se presenta la descripción morfológica y las ilustraciones correspondientes.
Tabla 1
Caracterización de los sitios de muestreo de Septoglomus en México.
| Estado | Sitio | Coordenadas geográficas (Datum WGS84) | Vegetación | Suelo | Clima |
| San Luis Potosí | 1. San José de Coronados | 23°34’35.4” N 100°55’09.5” O | Bosque de pino | Xerosol | Árido templado |
| 2. Xilitla | 21°22’21.4” N 99°00’39.0” O | Bosque mesófilo de montaña | Litosol | Semicálido húmedo | |
| 3. Matehuala | 23°21’51.1” N 100°35’01.7” O | Matorral xerófilo | Xerosol | Árido semicálido | |
| 4. Sierra de Álvarez | 22°03’06.3” N 100°34’50.8” O | Bosque de encino | Litosol | Semicálido | |
| 5. Charcas | 23°22’58.5” N 101°20’18.5” O | Pastizal | Xerosol | Semiárido | |
| Guanajuato | 6. San Felipe | 21°41’56.4” N 101°12’57.6” O | Bosque de encino | Feozem | Semiárido templado |
| Veracruz | 7. Plan de Higuera | 19°26’17.0” N 96°33’35.6” O | Bosque tropical caducifolio | Vertisol | Cálido subhúmedo |
| Coahuila | 8. Sierra Hermosa | 25°14’42” N 100°51’18” O | Bosque de pino | Litosol | Semiárido templado |
| Tamaulipas | 9. Jaumave | 23°08’26.0” N 99°17’48.4” O | Bosque de encino | Litosol | Semiárido templado |
Fuente: datos generados a partir de las capas disponibles en Geoportal y Enciclovida, México, Conabio (2018).
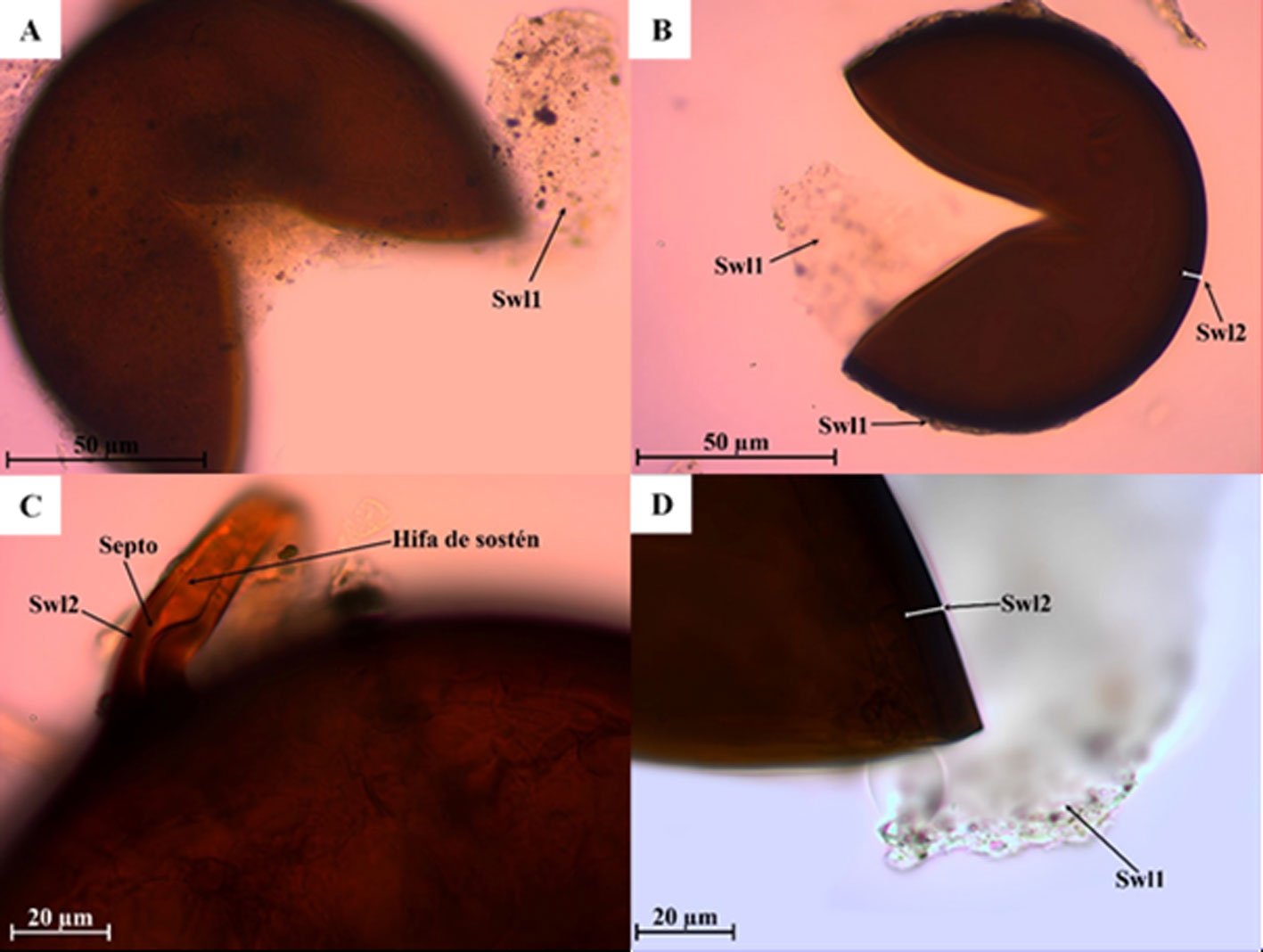
Septoglomus altomontanum Palenz., Oehl, Azcón-Aguilar y GA Silva, 2013.
Fig. 2a-d
Las esporas se forman individualmente en el suelo. Tienen forma ovalada y globosa, de 128-152 × 121-160 µm, de color marrón rojizo tornándose a negro rojizo. Las esporas están compuestas de 2 capas (Swl1-2). Swl1 es subhialina a amarillo oscuro, acompañada de partículas del suelo, de 2.1 a 3.2 µm de espesor, con la presión al oprimir el cubreobjetos esta capa se desprende. Swl2 es de marrón rojizo a negro rojizo, lisa, laminada, de 3.8 a 8.2 µm de espesor. Ninguna de las capas reacciona al reactivo de Melzer. La hifa de sostén es de forma curveada, continua a la pared de la espora, de un color marrón rojizo, es más ancha en la base de la espora y llega a medir 22 µm de largo. El poro de la espora está cerrado por un septo concoloro o una tonalidad más clara que la espora y surge de la capa Swl2.
Material examinado: México, Guanajuato, San Felipe (21°41’56.4” N, 101°12’57.6” O, 2,445 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-Piedras, 08-2021. México, San Luis Potosí, Catorce, San Antonio de Coronados (23°35’11.8” N, 100°54’01.2” O, 2,455 m snm) en un suelo rizosférico de un bosque de pino, R. P. Hipólito-Piedras, 08-2021. México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. (CH-SLPM-228).
Distribución y hábitat. Septoglomus altomontanum se registró por primera vez en el Parque Nacional Sierra Nevada en Andalucía, España, donde se encontró asociada a Pinguicola grandiflora Lam. y P. nevadensis H. Lindb(Lentibulariaceae), así como a otras especies vegetales presentes en el suelo circundante (Palenzuela et al., 2013). Posteriormente, se identificó en muestras de suelo rizosférico de eucaliptos en el noreste de Tailandia (Khaekhum et al., 2017), así como en un bosque enano de cerezo de montaña en el monte Fanjing, que forma parte de las montañas Wuling, ubicadas en la provincia de Guizhou, China (He et al., 2021). Adicionalmente, se ha documentado la presencia de S. altomontanum en un bosque de Cryptomeria japonica (Miq.) P.D.Sell, (Taxodiaceae) en estas mismas montañas (Ming et al., 2023). En México, se aisló de suelo rizosférico de un bosque de encino, en Guanajuato, bosque de pino en San Luis Potosí y bosque tropical caducifolio en Veracruz.
Septoglomus furcatum Błaszk., Chwat, Kovács y Ryszka, 2013.
Fig. 3a-d
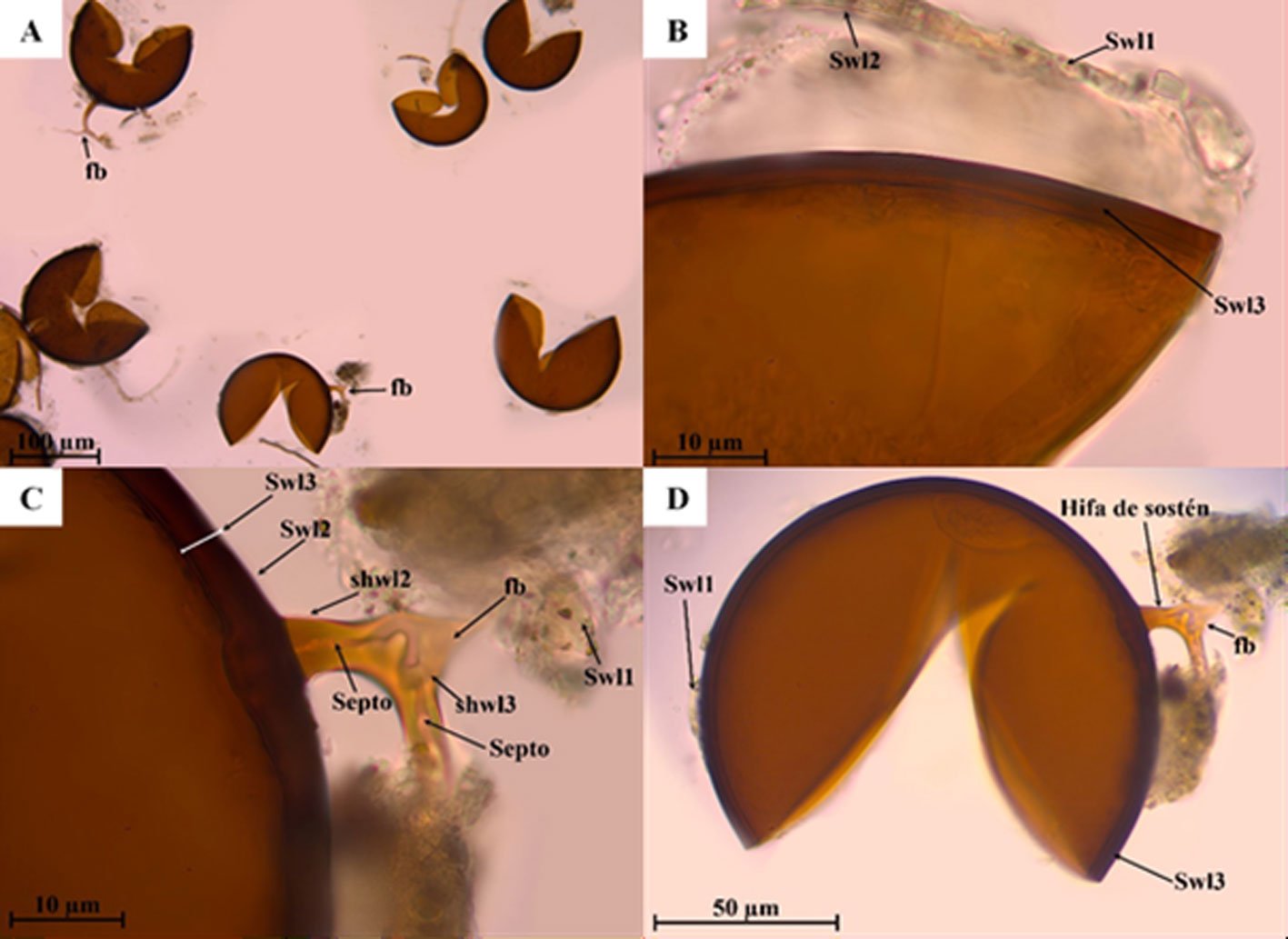
Figura 3. Estructura subcelular de Septoglomus furcatum A) Grupo de esporas, bifurcación de hifa (fb), característica de la especie en PVLG (10x); B) capas externas Swl1-3 en reactivo de Melzer (100x); C) capas externas Swl1-3 de la pared de la espora, capas Shwl1-2, septo y bifurcación de la hifa de sostén en reactivo de Melzer (100x); D) espora e hifa de sostén en PVLG (40x).
Esporas de color marrón rojizo a marrón oscuro; globosas de (106-) 138 (-167) µm de diámetro. La pared de la espora consta de 3 capas (Swl1-3). Capa Swl1, formando la superficie de la espora, semipermanente, hialina con restos de suelo de 1.6 µm de espesor y se desprende con la presión al oprimir el cubreobjetos. Swl2 semipermanente, hialina de 1.3 a 3.1 µm de espesor. Swl3 es laminada, lisa, marrón rojizo de 5.3 a 10.9 µm de espesor. Ninguna de las capas de la pared de la espora se tiñe con el reactivo de Melzer. La hifa de sostén es de color marrón parduzco; de forma recta de 10.5 a 30.3 µm, frecuentemente presenta una bifurcación (fb) a distancia de la pared de la espora. La pared de la hifa está compuesta de 3 capas (shwl1-3), las cuales son continuas a las paredes de la espora (Swl1-3). El poro de la espora está cerrado. El septo se encuentra en la base de la espora y pueden encontrarse numerosos septos a lo largo de la hifa.
Material examinado: México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. México, San Luis Potosí, Matehuala (23°21’51.1” N, 100°35’01.7” O, 1,373 m snm) en un suelo rizosférico de matorral xerófilo, R. P. Hipólito-Piedras, 08-2021 (CH-SLPM-229).
Distribución y hábitat. Septoglomus furcatum, se registró por primera vez asociado a raíces de Cordia oncocalyx Allemann (Cordiaceae) en Brasil. Posteriormente se realizó un cultivo monoespecífico utilizando a Plantago lanceolata L. (Plantaginaceae) como planta huésped, en donde S. furcatum formó micorrizas con numerosos arbúsculos, vesículas e hifas poco frecuentes (Blaszkowski et al., 2013). El segundo reporte en Brasil, se evidenció a través de una investigación en torno a la retroalimentación de suelo-planta, donde la mencionan como una especie clave, propagada en Senna uniflora (Mill.) HSIrwin y Barneby (Caesalpiniaceae), con suelo de áreas degradadas que han sido restauradas en ese país (Medeiros et al., 2022). La presencia de S. furcatum en el continente asiático seregistró asociado a las raíces de Rosa acicularis Lindl. (Rosaceae) junto a una baja diversidad de 3 especies de HMA (Zu et al., 2019), mientras que Ming et al. (2023), la registraron en un bosque de Pinus damingshanensis W.C.Cheng y L.K.Fu (Pinaceae) , Cryptomeria japonica y en un bosque lanceolado de Cunninghamia R.Br. (Cupressaceae), junto con 27 especies más. En México, se aisló de suelo rizosférico de un bosque tropical caducifolio en Veracruz y matorral xerófilo en San Luis Potosí.
Septoglomus fuscum Błaszk., Chwat, Kovács y Ryszka, 2013.
Fig. 4a-d
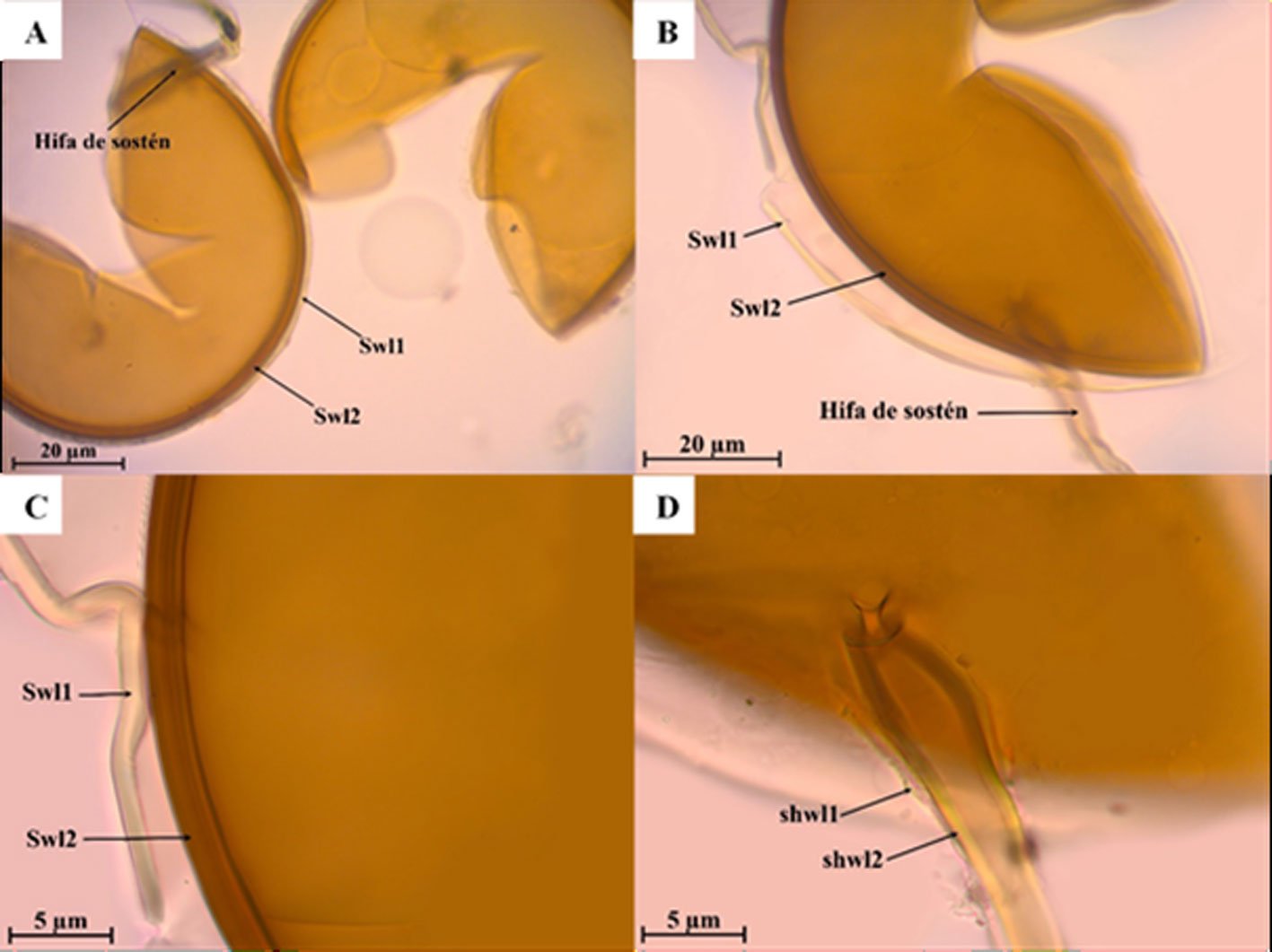
Las esporas se forman en el suelo en racimos sueltos. Grupos de 89-194 × 141-248 µm con 2 a 7 esporas. Esporas de color blanco amarillento en la juventud, anaranjado parduzco a marrón oscuro en la madurez; globosas a subglobosas; (32-) 51 (-102) µm de diámetro. La pared de la espora consta de 2 capas (Swl1-2). Swl1, es semipersistente, semiflexible, de color amarillo dorado, de 1.8 a 2.4 µm de grosor, al oprimir el cubreobjetos se separa de la capa Swl2. Swl2 es laminada, lisa, de amarillo a anaranjado parduzco de (1.8-) 3.5 (-6.7) µm de grosor. Ninguna de las capas de la pared de la espora se tiñe con el reactivo de Melzer. La hifa de sostén es de color anaranjado parduzco a marrón oscuro en esporas maduras; recta o recurvada, cilíndrica en forma de embudo, a veces ligeramente constreñida en la base de la espora; (5.8-) 7.4 (-12.3) µm de anchura en la base de la espora. Pared de la hifa de color amarillo a anaranjado parduzco de (0.8-) 2.7 (-5.8) µm de grosor. El poro es abierto de 1.5 a 2.7 µm de diámetro.
Material examinado: México, Veracruz, Actopan, Plan de Higuera (19°26’17.0” N, 96°33’35.6” O, 140 m snm) en un suelo rizosférico de un bosque tropical caducifolio, H. Méndez-Cortés, 08-2020. México, San Luis Potosí, Matehuala (23°21’51.1” N, 100°35’01.7” O, 1,373 m snm) en un suelo rizosférico de matorral xerófilo, R.P. Hipólito-Piedras, 08-2021 (CH-SLP-230).
Distribución y hábitat. Septoglomus fuscum se registró por primera vez asociado con raíces de Arctotheca populifolia (P.J.Bergius) Norl. (Asteraceae) en Sudáfrica. Posteriormente, se realizaron cultivos monoespecíficos con Plantago lanceolata como cultivo huésped, en donde se observaron estructuras como arbúsculos, hifas intra y extrarradicales (Blaszkowski et al., 2013). En México, se aisló de suelo rizosférico de bosque tropical caducifolio en Veracruz y un matorral xerófilo en San Luis Potosí.
Septoglomus turnauae Błaszk., Chwat, Ryszka y Orfanoudakis, 2014.
Fig. 5a-d
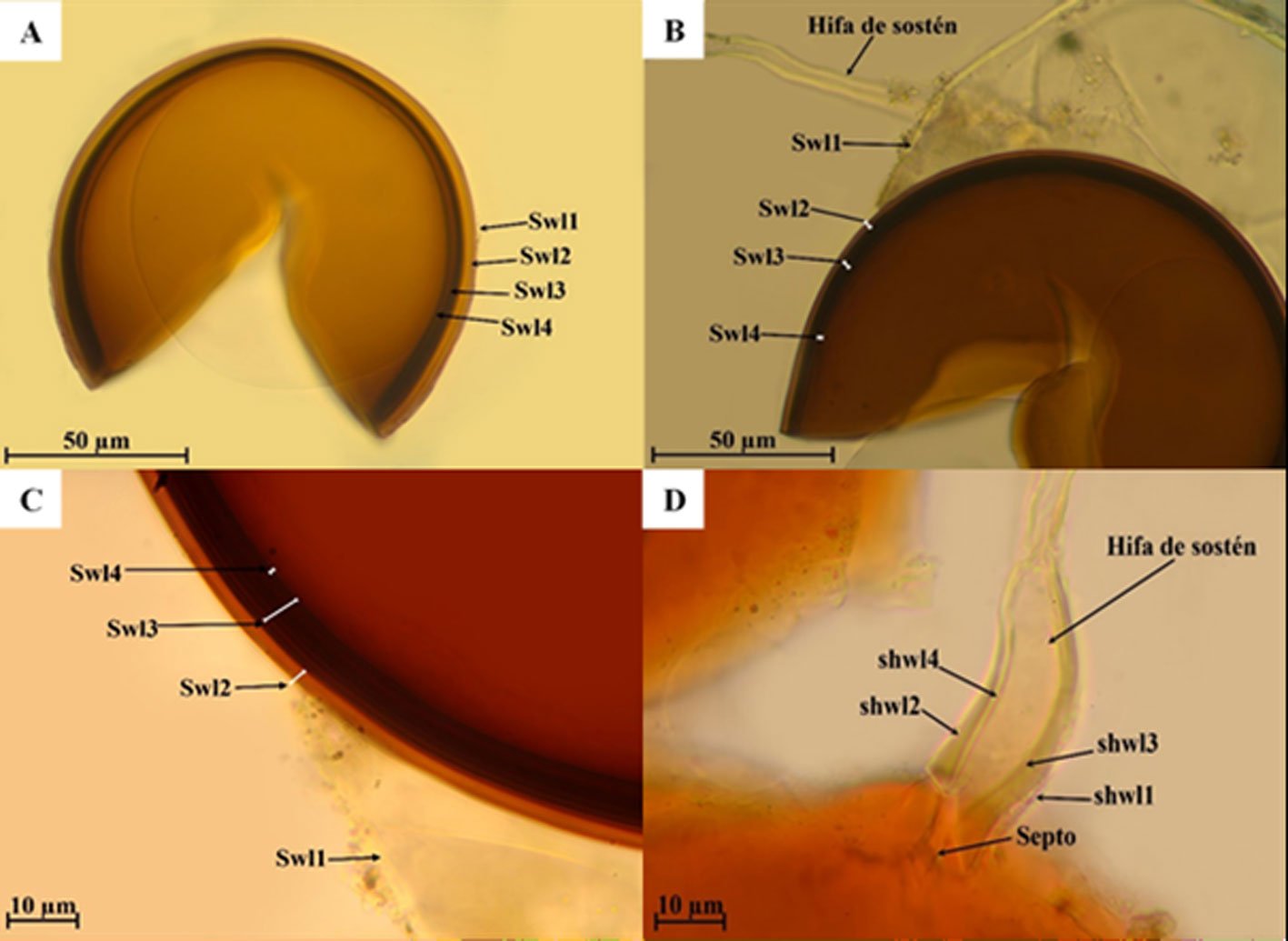
Las esporas se forman de manera solitaria en el suelo. Esporas de color anaranjado amarronado a marrón oscuro, globosas de (45) 64 (-102) µm de diámetro. La pared de la espora está compuesta de 4 capas (Swl1-4). Swl1, es evanescente, hialina, de 3.1 µm de diámetro. Swl2 es permanente, laminada, lisa, de anaranjado claro de 1.8 a 9.2 µm de espesor. Swl3 es laminada, lisa, de color anaranjado claro a anaranjado parduzco, de 3.5 a 9.3 µm de espesor. Swl4 semiflexible, lisa, de color anaranjado parduzco de 1.0 a 1.7 µm de espesor. Ninguna de las capas se tiñe con el reactivo de Melzer. La hifa es de color anaranjado parduzco de (4.5-) 8.2 (-10.7) µm de espesor en la base de la espora, la hifa es continua con las capas Swl1-3. El poro es de (1.3-) 2.7 (-3.2) µm de diámetro, estrechándose con la edad.
Material examinado: Coahuila, Sierra Hermosa (25°14’42” N, 100°51’18” O, 2,255 m snm) en suelo rizosférico de un bosque de pino, H. Méndez-Cortés, 08-2020. Guanajuato, San Felipe (21°41’56.4” N, 101°12’57.6” O, 2,445 m snm), en un suelo rizosférico de un bosque de encino, R.P. Hipólito-Piedras. San Luis Potosí, Xilitla (21°22’21.4” N, 99°00’39.0” O, 857 m snm) en un suelo rizosférico de un bosque mesófilo de montaña, R. P. Hipólito-Piedras, 08-2021. México, San Luis Potosí, Charcas (23°22’58.5” N, 101°20’18.5” O, 2,471 m snm) en un suelo rizosférico de un pastizal, R. P. Hipólito-Piedras, 08-2021. San Luis Potosí, Sierra de Álvarez (22°03’06.3” N, 100°34’50.8” O, 2,700 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-Piedras, 08-2021. Tamaulipas, Jaumave (23°08’26.0” N, 99°17’48.4” O, 742 m snm) en un suelo rizosférico de un bosque de encino, R. P. Hipólito-piedras, 08-2021. (CH-SLPM-230).
Distribución y hábitat. S. turnuauae se registró en cultivos trampa inoculados con el suelo de la rizósfera y raíces de Cistus sp. L. (Cistaceae), en Italia. Posteriormente, se realizaron cultivos monoespecíficos con Plantago lanceolata, donde se observaron arbúsculos e hifas (Blaszkowski et al., 2014). En México, se aisló de suelo rizosférico de bosques de encino en Guanajuato y Tamaulipas, bosques de pino en Coahuila, bosque mesófilo de montaña y pastizal en San Luis Potosí.
Clave de las especies del género Septoglomus presentes en México
1. Esporas mayores a 200 micras 4
1. Esporas menores a 200 micras 2
2. Esporas con 2 capas en la pared de la espora 3
2. Esporas con 3 capas en la pared de la espora 5
2. Esporas con 4 capas en la pared de la espora 6
3 Esporas amarillas a marrones 7
3. Esporas rojas a negras 8
4. Esporas solitarias; de color anaranjado rojizo a marrón rojizo; globosa; (154.5-)202.8 (-228.9) µm de diámetro. La pared de la espora costa de 4 capas (Swl1-4). Capa Swl1 es evanescente hialina, lisa, con la edad se torna rugosa o se desprende. Swl2 permanente, laminada de color anaranjado, Swl3 es permanente, lisa y laminada de color anaranjado a marrón. Swl4 semiflexible, hialina, lisa, raramente se separa de swl3. Hifa de sostén amarilla a marrón, recurvada de 4 capas (shwl1-4), con poro cerrado por un septo situado en la base de la espora Septoglomus mexicanum
5. Esporas solitarias; de color marrón rojizo a marrón oscuro; globosas a subglobosas; (106-) 138 (-167) µm de diámetro. La pared de la espora consta de 3 capas (Swl1-3). Capa Swl1, semipermanente, hialina a anaranjado claro, lisa con gránulos de suelo. Swl2 semipermanente, hialina a amarillo dorado. Swl3 es laminada, lisa, marrón rojizo a marrón oscuro. La hifa de sostén es de color marrón parduzco a marrón oscuro; rectas o curvadas, cilíndricas, bifurcada, compuestas por 3 capas (shwl1-3), poro cerrado por un septo en la base de la espora, además, a lo largo de la hifa se encontrarán septos transversales Septoglomus furcatum
6. Esporas solitarias; de color anaranjado parduzco a marrón oscuro; globosas a subglobosas; (110-) 133(-165) μm de diámetro. La pared de la espora consta de 4 capas (Swl1-4). Swl1, es evanescente, hialina y lisa. Swl2 permanente, laminada, lisa, de color anaranjado claro a anaranjado parduzco. Swl3 es laminada, lisa, de color anaranjado claro a anaranjado parduzco. Swl4 semiflexible, lisa, de color anaranjado claro a anaranjado parduzco. La hifa de sostén es de color anaranjado parduzco a marrón oscuro, constituida de 3 capas (shwl1-3) con un septo en la base de la espora que no invade el lumen de la hifa de sostén Septoglomus turnauae
7. Esporas en racimos o solitarias, de color amarillento en la juventud, anaranjado parduzco a marrón oscuro en la madurez; globosas a subglobosas; (20- ) 47 (-90) µm de diámetro. La pared de la espora consta de 2 capas (Swl1-2). Swl1, semi persistente, semiflexible, de color blanco anaranjado a amarillo dorado. Swl2 laminada, lisa, blanco amarillento en esporas juveniles, anaranjado parduzco a marrón oscuro en esporas maduras. La hifa de sostén de color anaranjado parduzco a marrón oscuro; recta o recurvada, cilíndrica a forma de embudo, con poro abierto, sin septo, estrechamiento gradual hasta llegar al centro del lumen de la hifa Septoglomus fuscum
8. Esporas solitarias; de color marrón rojizo oscuro a negro rojizo; ovoides, elípticas y globosas, de 137-175 (-208) × 125-170 (-204) µm de diámetro. La pared de la espora está compuesta por 2 capas (Swl1-2). Swl1 es subhialina, lisa, de color amarillo oscuro. Swl2 es de marrón rojizo oscuro a negro rojizo, lisa, laminada. La hifa de sostén de color amarillo oscuro a un marrón rojizo, cilíndrica, curveada, ancha en la base de la espora estrechándose debajo de esta, poro cerrado, septo concoloro a la base de la espora y tonalidad clara cuando se encuentra debajo de la pared de la espora Septoglomus altomontanum
Discusión
En México, el género Septoglomus está representado por 33% de las especies descritas en el mundo, lo que equivale a 5 de las 13 especies conocidas. Entre éstas, destaca S. constrictum debido a su amplia distribución y frecuentes registros en diversas comunidades vegetales. Por otro lado, S. xanthium ha sido principalmente registrada en agroecosistemas (Polo-Marcial et al., 2021). Septoglomus mexicanum se ha registrado en asociación con Prosopis laevigata (Humb. y Bonpl. ex Willd.) M.C.Johnst. (Mimosaceae) (Chimal-Sánchez et al., 2020), similar a S. titan BT Goto y GA Silva (Glomeraceae) que se ha registrado en zonas áridas del país (Guardiola-Márquez et al., 2022).
De acuerdo con los registros de las especies de HMA descritas en esta investigación, Septoglomus altomontanum fue la única especie cuya distribución coincidió con la descripción original al ser aislada en sitios con vegetación de coníferas y encinos, a altitudes superiores a 2,000 m, lo cual es similar a lo reportado por Palenzuela et al. (2013), quienes documentaron su presencia en zonas con vegetación similar, localizadas entre 1,800 y 3,100 m de altitud.
Septoglomus altomontanum se diferencia de las especies de HMA descritas hasta la fecha, incluso de las del mismo género, en términos de tamaño, color, estructura, forma e hifas de las esporas. La especie que podría generar algún debate conforme a su morfología es S. furcatum; no obstante, ésta presenta la particularidad de tener una hifa que se adelgaza a medida que se aleja de la base de la espora (Blaszkowski et al., 2013), mientras que S. altomontanum mantiene un grosor uniforme a lo largo de la pared de la espora y la hifa (Palenzuela et al., 2013).
Septoglomus furcatum se destaca por sus esporas grandes de tonalidad oscura que presentan una pared de 3 capas y, con frecuencia, hifas ramificadas, incluso llegando a tener 2 hifas cercanas entre sí, característica que no se encuentra en otras especies del género Septoglomus (Blaszkowski et al., 2013).A primera vista, las esporas de S. furcatum pueden parecer similares a las de S. constrictum (Trappe) Sieverd., GA Silva et Oehl; sin embargo, se diferencian por la estructura de la pared de la espora, ya que S. furcatum tiene 3 capas, mientras que S. constrictum tiene solo 2. Además, la espora de S. constrictum tiende a ser de un tono anaranjado parduzco, mientras que la de S. furcatum es de color marrón oscuro. La característica de S. furcatum de poseer una hifa ramificada, es similar a la de las esporas de G. brohultii Sieverd. y R.A. Herrera (Glomeraceae). Sin embargo, G. brohultii tiene una tonalidad más clara tanto en la pared de la espora como en la hifa, a diferencia de S. furcatum, que presenta una tonalidad oscura uniforme tanto en la pared como en la hifa (Blaszkowski et al., 2013; Oehl et al., 2011).
Septoglomus fuscum es similar a S. xanthium, tanto en características moleculares como en características morfológicas. No obstante, se ha observado que S. fuscum forma racimos sueltos de esporas, mientras que S. xanthium se compone de esporas solitarias que se adhieren y se desarrollan dentro de las raíces (Blaszkowski et al., 2004). Al analizar las capas, se nota que S. xanthium posee una capa adicional permanente que S. fuscum no presenta; sin embargo, S. fuscum tiene un grosor de capa laminada de 2 a 3 veces mayor que el de S. xanthium (Blaszkowski et al., 2013).
Desde una perspectiva filogenética, S. turnauae presenta una estrecha relación con S. fuscum, lo que sugiere un origen común entre ambas especies, a través de análisis ribosomales que las agrupan en un mismo clado. Sin embargo, se destaca que la mayoría de las esporas de S. fuscum se agrupan en racimos sueltos (Blaszkowski et al., 2013), en contraste con las esporas individuales de S. turnauae. Al comparar el número de capas, S. turnauae muestra similitudes con S. mexicanum, ambas compuestas por 4 capas en su estructura, aunque, difieren en el color de las esporas. Mientras que S. mexicanum presenta tonalidades que van desde el rojizo hasta el marrón (Chimal-Sánchez et al., 2020), las esporas de S. turnauae tienen un tono anaranjado parduzco y tonalidades diferentes, conforme empieza otra capa (Blaszkowski et al., 2014).
Previo a esta investigación, se consideraba que la distribución del género Septoglomus era limitada, sin embargo, donde se localiza expresa un comportamiento dominante respecto a los demás géneros de HMA (Guardiola-Márquez et al., 2022). No obstante, la presente investigación, que ha registrado 4 especies pertenecientes a este género, revela una riqueza de HMA previamente desconocida en las comunidades vegetales de México. Además, sugiere que este género no se encuentra restringido a una distribución específica, ya que se ha documentado en 5 comunidades vegetales distintas en el país. El reciente hallazgo de S. altomontanum, S. fuscum, S. furcatum y S. turnauae eleva el número de especies registradas en México a 171, ésto constituye aproximadamente 48% de la riqueza global de HMA y 62% de las especies se han reportado del género Septoglomus en el país.
Agradecimientos
A la Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti) por la beca otorgada (775694) a la primera autora para sus estudios de posgrado.
Referencias
Błaszkowski, J., Blanke, V., Renker, C., y Buscot, F. (2004). Glomus aurantium and G. anthium, new species in Glomeromycota. Mycotaxon, 90, 447–467. https://doi.org/10.5962/p.431247
Błaszkowski, J., Chwat, G., Góralska, A., Ryszka, P. y Orfanoudakis, M. (2014). Septoglomus jasnowskae and Septoglomus turnauae, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycology Progress, 13, 985. https://doi.org/10.1007/s11557-014-0985-z
Blaszkowski, J., Chwat, G., Kovacs, G. M., Gaspar, B. K., Ryszka, P., Orowska, E. et al. (2013). Septoglomus fuscum and S. furcatum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia, 105, 670–680. https://doi.org/10.3852/12-127
Błaszkowski, J., Kovács, G. M., Balazs, T. K., Orlowska, E., Sadravi, M., Wubet, T. et al. (2010). Glomus africanum and G. iranicum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia, 102, 1450–1462. https://doi.org/10.3852/09-302
Błaszkowski, J., Yamato, M., Niezgoda, P., Zubek, S., Milczarski, P., Malinowski, R. et al. (2023). A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycological Progress, 22, 1–15. https://doi.org/10.1007/s11557-023-01882-9
Chimal-Sánchez, E., Senés-Guerrero, C., Varela, L., Montaño, N. M., García-Sánchez, R., Pacheco, A. et al. L. (2020). Septoglomus mexicanum, a new species of arbuscular mycorrhizal fungi from semiarid regions in Mexico. Mycologia, 112, 121–132. https://doi.org/10.1080/00275514.2019.1671147
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2018). Sistema Nacional de Información sobre Biodiversidad (SNIB). Registros de ejemplares, versión 2017-12. Geoportal y Enciclovida, México. Recuperado el 28 julio, 2023 de: http://www.conabio.gob.mx/informacion/gis/
da Silva, G. A., Corazon-Guivin, M. A., de Assis, D. M. A. y Oehl, F. (2023). Blaszkowskia, a new genus in Glomeraceae. Mycological Progress, 22, 1–14. http://dx.doi.org/10.1007/s11557-023-01919-z
Daniels, H. B. A. y Skipper, H. D. (1982). Methods for the recovery and quantitative estimation of propagules from soil. En N. C. Schenck (Ed.), Methods and principles of mycorrhizal research (pp. 29–37). St. Paul, Minn.: American Society for Phytopathology,
Gerdemann, J. W. y Nicolson, T. J. (1963). Spores of mycorrhizal Endogonespecies extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society, 46, 235–244. http://dx.doi.org/10.1016/S0007-1536(63)80079-0
Goto, B. T., Araújo, A. F., Soares, A. C. F., de Almeida-Ferreira, A. C., Maia, L. C., da Silva-Sousa, C. et al. (2013). Septoglomus titan, a new fungus in the Glomeraceae (Glomeromycetes) from Bahia, Brazil. Mycotaxon, 124, 101–109. http://dx.doi.org/10.5248/124.101
Guardiola-Márquez, C. E., Pacheco, A., Mora-Godinez, S., Schüßler, A., Gradilla-Hernández, M. S. y Senés-Guerrero, C. (2022). Septoglomus species dominate the arbuscular mycorrhiza of five crop plants in an arid region of northern Mexico. Symbiosis, 87, 93–106. http://dx.doi.org/10.1007/s13199-022-00851-2
Guillén, A., Serrano-Tamay, F. J., Peris, J. B. y Arrillaga, I. (2020). Glomus ibericum, Septoglomus mediterraneum, and Funneliformis pilosus, three new species of arbuscular mycorrhizal fungi. Mycologia, 112, 819–828. https://doi.org/10.1080/00275514.2020.1771992
He, R. J., Ming, Y., Yao, L. M., Zhu, Q. Q. y Jiang, L. (2021). Diversity of arbuscular mycorrhizal fungi in mossy dwarf forest of Fanjing Mountain, Guizhou Province, Southwest China. Mycosystema, 40, 2843–2853. https://doi.org/10.13346/j.mycosystema.210188
INVAM (International Culture Collection of Vesicular Arbuscular Mycorrhizal Fungi). (2023). Recuperado el 28 agosto, 2023 de: http://invam.caf.wvu.edu/
Khaekhum, S., Lumyong, S., Kuyper, T. W. y Boonlue, S. (2017). Species richness and composition of arbuscular mycorrhizal fungi occurring on eucalypt trees (Eucalyptus camaldulensis Dehnh.) in rainy and dry season. Current Research in Environmental and Applied Mycology, 7, 282–292. http://dx.doi.org/10.5943/cream/7/4/5
Medeiros, A. S., Scaloppi, J. C., Damasceno, E. S., Goto, B. T., Vieira, D. C. M., Socolowski, F. et al. (2022). Arbuscular mycorrhizal fungi communities shaped by host-plant affect the outcome of plant-soil feedback in dryland restoration. Journal of Applied Ecology, 60, 507–518. https://doi.org/10.1111/1365-2664.14330
Ming, Y., He, R., Yao, L., Zhu, Q., Long, C. y Jiang, L. (2023). Diversity of soil arbuscular mycorrhizal fungi in different coniferous forests in Fanjing Mountain. Journal of Yunnan Agricultural University (Natural Science), 38, 149–157. https://doi.org.10.12101/j.issn.1004-390X(n).202108033
Niezgoda, P., Błaszkowski, J., Błaszkowski, T., Stanisławczyk, A., Zubek, S., Milczarski, P. et al. (2024). Three new species of arbuscular mycorrhizal fungi (Glomeromycota) and Acaulospora gedanensis revised. Frontiers in Microbiology, 15, 1320014. https://doi.org/10.3389/fmicb.2024.1320014
Oehl, F., Sánchez-Castro, I., da Silva, D. K. A., Santos, V. M., Palenzuela Jiménez, E. J. y da Silva, G. A. (2019). Septoglomus nigrum, a new arbuscular mycorrhizal fungus from France, Germany and Switzerland. Nova Hedwigia, 109, 121–134. http://dx.doi.org/10.1127/nova_hedwigia/2019/0523
Oehl, F., Silva, G. A. D., Goto, B. T. y Sieverding, E. (2011). Glomeromycota: three new genera and glomoid species reorganized. Mycotaxon, 116, 75–120. http://dx.doi.org/10.5248/116.75
Palenzuela, J., Azcón-Aguilar, C., Barea, J. M., da Silva, G. A. y Oehl, F. (2013). Septoglomus altomontanum, a new arbuscular mycorrhizal fungus from mountainous and alpine areas in Andalucía (southern Spain). IMA Fungus, 4, 243–249. https://doi.org/10.5598/imafungus.2013.04.02.09
Polo-Marcial, M. H., Lara-Pérez, L. A., Goto, B. T., Margarito-Vista, X. y Andrade-Torres A. (2021). Glomeromycota in Mexico, a country with very high richness. Sydowia, 74, 33–63. http://dx.doi.org/10.12905/0380.sydowia74-2021-0033
Redecker, D., Schüßler, A., Stockinger, H., Stürmer, S. L., Morton, J. B. y Walker, C. (2013). An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza, 23, 515–531. https://doi.org/10.1007/s00572-013-0486-y
Schüßler, A., Schwarzott, D. y Walker, C. (2001). A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycological Research, 105, 1413–1421. https://doi.org/10.1017/S0953756201005196
Schüßler, A. y Walker, C. (2010). The Glomeromycota. A species list with new families and new genera. Recuperado el 28 agosto, 2023 de: http://schuessler.userweb.mwn.de/amphylo/
Symanczik, S., Błaszkowski, J., Chwat, G., Boller, T., Wiemken, A. y Al-Yahya’ei, M. N. (2014). Three new species of arbuscular mycorrhizal fungi discovered at one location in a desert of Oman: Diversispora omaniana, Septoglomus nakheelum and Rhizophagus arabicus. Mycologia, 106, 243–259. https://doi.org/10.3852/106.2.243
Zu, Y., Ping, Y., Mu, L. y Yang, T. (2019). The diversity of arbuscular mycorrhizal fungi of Rosa acicularis ‘Luhe’in saline areas. Journal of Forestry Research, 30, 1507–1512. https://doi.org/10.1007/s11676-018-0748-9
Wijayawardene, N. N., Hyde, K. D., Al-Ani, L. K. T., Tedersoo, L., Haelewaters, D., Rajeshkumar, K. C. et al. (2020). Outline of fungi and fungus-like taxa. Mycosphere Online. Journal of Fungal Biology, 11, 1060–1456. https://doi.org/10.5943/mycosphere/13/1/2
Schüβler, A. 2025. Glomeromycota: species list. Recuperado en marzo, 2025 de: http://www.amf-phylogeny.com/


