José Alan Herrera-García a, Mahinda Martinez a, b, *, Pilar Zamora-Tavares c, Ofelia Vargas c, Luis Hernández-Sandoval a, b
a Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales, Av. de las Ciencias, s/n, 76230 Juriquilla, Querétaro, Mexico
b Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales, Biología, Laboratorio Nacional de Identificación y Caracterización Vegetal, Av. de las Ciencias, s/n, 76230 Juriquilla, Querétaro, Mexico
c Universidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias, Instituto de Botánica, Departamento de Botánica y Zoología, Laboratorio Nacional de Identificación y Caracterización Vegetal, Av. Ing. Ramón Padilla Sánchez, 45200 Zapopan, Jalisco, Mexico
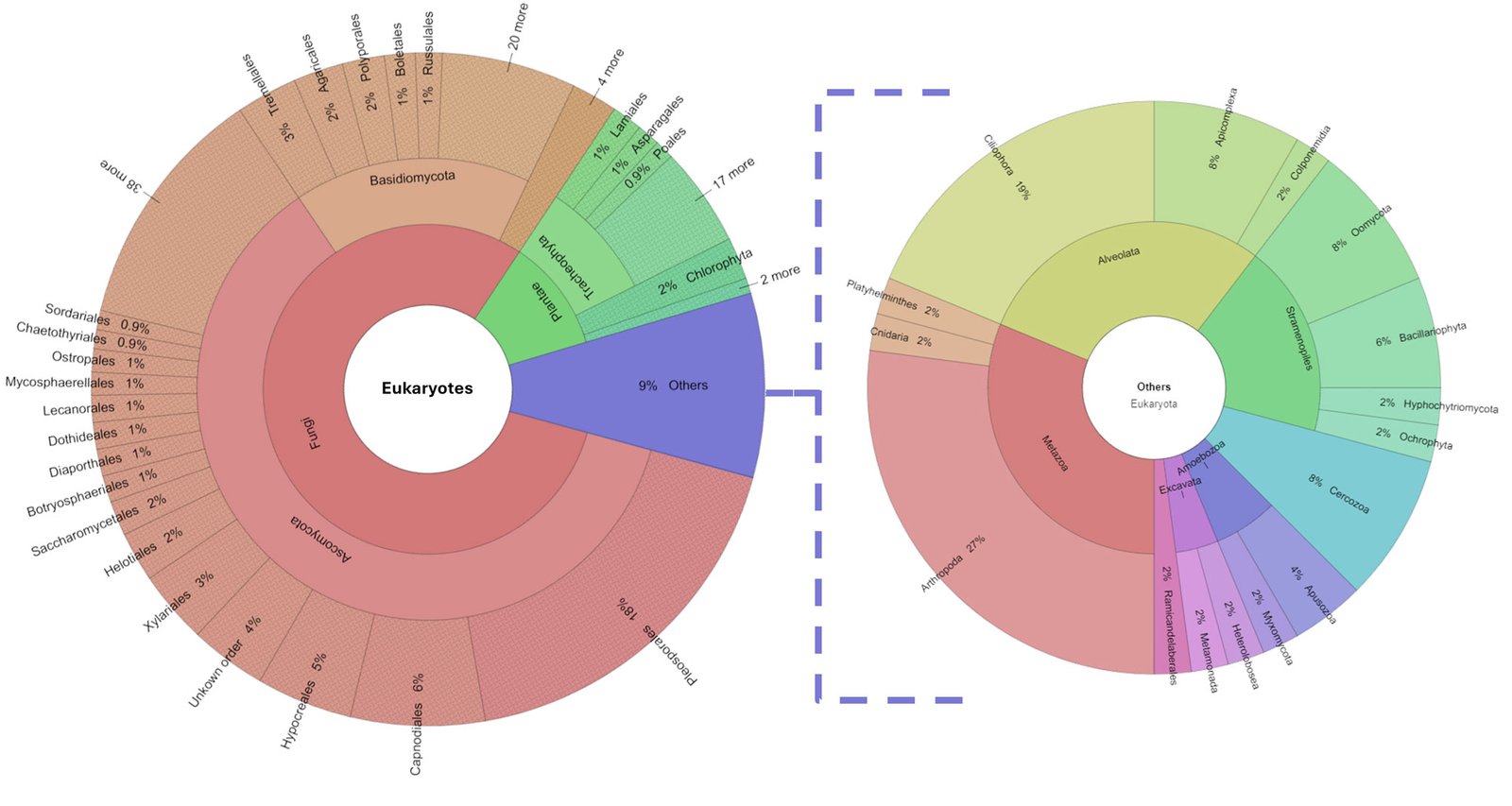
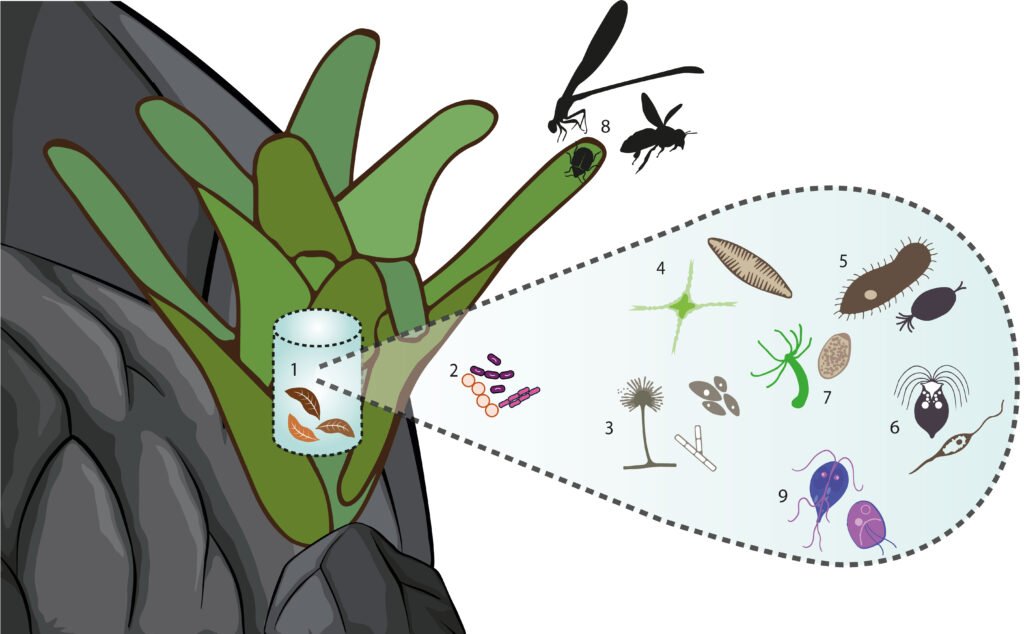
Some bromeliads form a compact rosette that accumulates detritus and water, known as phytotelma. The phytotelma is a lentic ephemeral aquatic environment that forms diverse communities with complex trophic levels. Pseudalcantarea grandis, a saxicolous plant, forms a phytotelma. To understand the importance of P. grandis as a eukaryotic diversity reservoir in arid zones, we collected water samples from 5 plants growing in a dry canyon in Zimapán, Hidalgo, Mexico. We analyzed them through metabarcoding of the ITS1 (Internal Transcribed Spacer) and the partial 5.8S gene. We used the Ion Torrent PGM platform for the sequencing, and the taxonomic assignation for the amplicons was made with BLAST in Genbank at NCBI. We found 26 phyla and 543 genera, 80% of which belonged to Ascomycota, Basidiomycota, Blastocladiomycota Chytridiomycota, Glomeromycota, Mucoromycota, and Zoopagomycota phyla. The remaining 20% was composed of 19 phyla belonging to other kingdoms. Photosynthetic organisms were represented by the phyla Bacillariophyta, Charophyta, Chlorophyta, and Ochrophyta. The vascular plants do not live in the tank but constitute the debris sustaining the large number of decomposers. The trophic levels in the tank were detritus, micro- and macro-decomposers, filter feeders, photosynthesizers, micro-predators, aquatic volume predators, surface predators, and parasites.
Diversidad de eucariotes y niveles tróficos dentro de la bromelia tanque Pseudalcantarea grandis en una zona árida detectados por metabarcoding de ADN ambiental
Resumen
Algunas bromelias forman rosetas compactas que acumulan detritus y agua. Esta acumulación se conoce como fitotelma, un hábitat acuático léntico y efímero con comunidades diversas y niveles tróficos complejos. Psedalcantarea grandis es una planta saxícola y forma un fitotelma. Para entender la importancia de P. grandis como reservorio de diversidad acuática en una zona árida, colectamos muestras de agua de 5 plantas en un cañón de Zimapán, Hidalgo, México y las analizamos por metabarcoding del ITS1 (Internal Transcribed Spacer) y una región parcial del gen 5.8S. La secuenciación se hizo en la plataforma Ion Torrent PGM. Asignamos la identidad taxonómica de los amplicones utilizando BLAST de Genbank. Encontramos 26 phyla y 543 géneros, 80% pertenecen a los phyla fúngicos Ascomycota, Basidiomycota, Blastocladiomycota Chytridiomycota, Glomeromycota, Mucoromycota, y Zoopagomycota. El 20% restante está compuesto por 19 phyla de otros reinos. Los organismos fotosintéticos estuvieron representados por los phyla Bacillariophyta, Charophyta, Chlorophyta y Ochrophyta. Otros organismos fotosintéticos que corresponden a plantas vasculares no viven dentro del tanque, pero forman la hojarasca que mantiene a los descomponedores. Los niveles tróficos en el tanque fueron detritus, micro y macrodescomponedores, filtradores, fotosintetizadores, microdepredadores, depredadores del volumen de agua, depredadores de superficie y parásitos.
Palabras clave: Acuático; Efímero; Zona árida; Fitotelma
Introduction
The Bromeliaceae family is comprised of almost 3,700 species distributed mostly in tropical areas of the Americas (Gouda et al., 2024). They are herbaceous perennial monocots with leaves arranged in rosettes, many of which are epiphytes (Ramírez-Murillo et al., 2004; Rzedowski, 2006). There are 442 species in Mexico (Espejo-Serna & López-Ferrari, 2018) that frequently grow in nutrient and mineral-deficient environments (Bernal et al., 2006; Ramírez-Murillo et al., 2004). Some species form compact rosettes with absorbent trichomes in their interior that allow the accumulation of solids rich in nutrients and water, known as phytotelma (Benzing, 2000; Goffredi et al., 2011).
Phytotelmata are lentic aquatic environments, mostly ephemeral that last less than 3 months (Mogi, 2004). They are freshwater habitats for diverse communities including viruses, Archaea, and bacteria (Brouard et al., 2013, Goffredi et al., 2011). Among the eukaryotes, aquatic mosses, green algae, diatoms, protists, fungi, insects, amphibians, and crustaceans have been documented (Benzing, 2000; Brandt et al., 2017; Kitching, 2001; Ramos & do Nascimento Moura, 2019; Rodríguez-Núñez et al., 2018; Simão et al., 2020).
Bromeliads that form phytotelma accumulate essential mineral elements, which are the main nitrogen source for the plant (Kitching, 2001). The presence of detritivores and predators is related to the nitrogen concentration in the leaves. Predators increase nutrient flux from the leaf litter of nearby plants to the bromeliad (Benzing & Renfrow, 1974; Ngai & Srivastava, 2006; Nievola et al., 2001; Takahashi & Mercier, 2011).
Unlike terrestrial and aquatic communities in which plants and algae are the main nutrient resources, in the phytotelma leaf litter and invertebrate remains play that role. In trophic networks inside the phytotelma, protists and rotifers are considered as micro-predators that also consume organic particles. Macroinvertebrates can consume the detritus, filter feeders, aquatic predators, and surface predators, whereas bacteria and fungi are the main decomposers that obtain energy directly from the detritus (Brouard et al., 2012; Mogi, 2004).
Gomes et al. (2015) characterized the enzymatic activity of the fungal community inside the bromeliad tank of Vriesea minarum in Brazil. Using cultivation techniques they identified 36 species, 22 of which were Basidiomycota and 14 were Ascomycota. The most relevant genera were Cryptococcus, Candida, and Aureobasidium. These organisms contain enzymes that degrade vegetal material.
Nutrient intake for the plant is further facilitated by insects (Ngai & Srivastava, 2006). For example, odonatan larvae ingest detritivores, contributing to the nitrogen cycle within the bromeliad by defecating. Leachates from defecation release nitrogen in a form available to the bromeliad and create a suitable niche for other microorganisms providing substrata (Benzing & Renfrow, 1974).
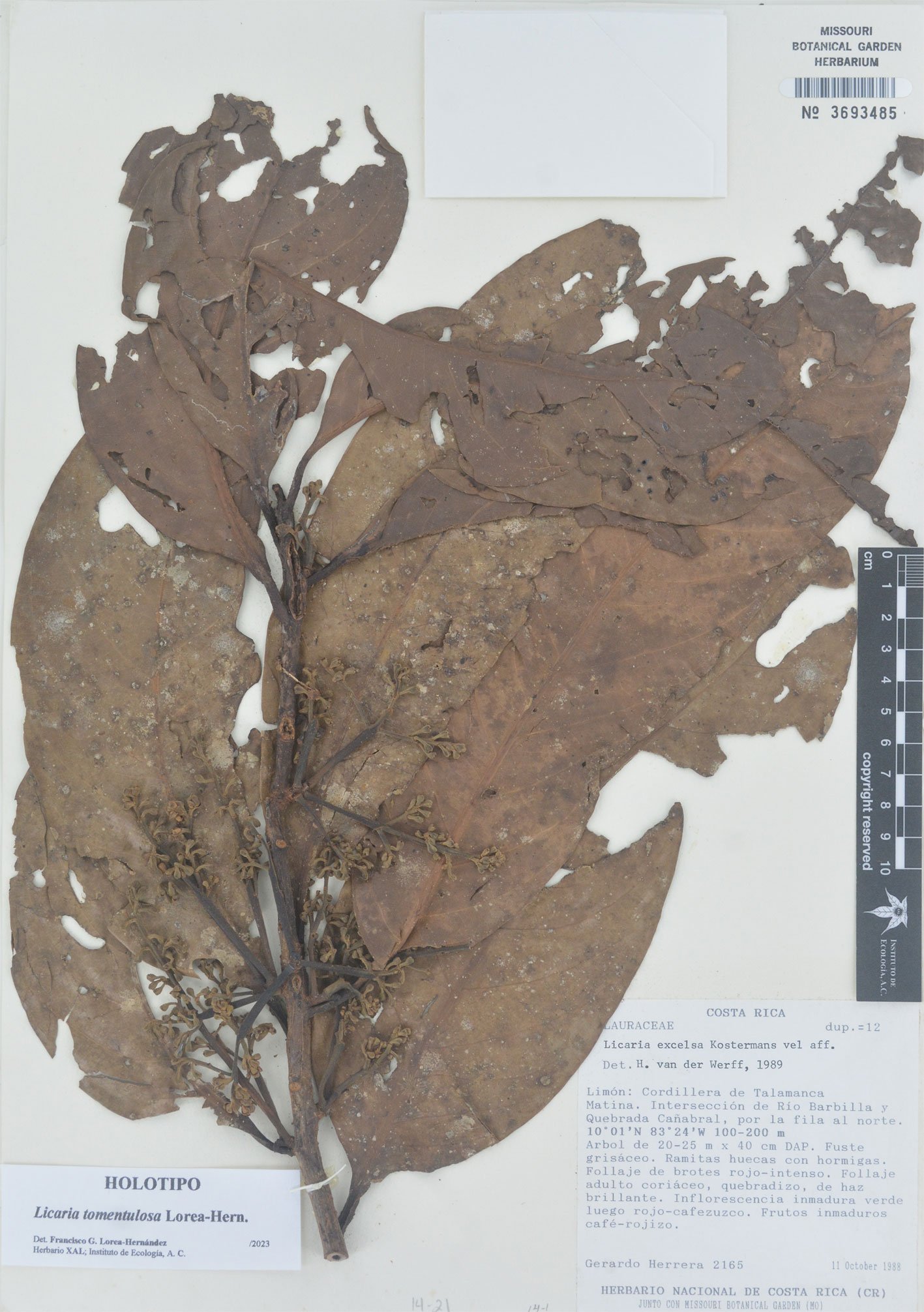
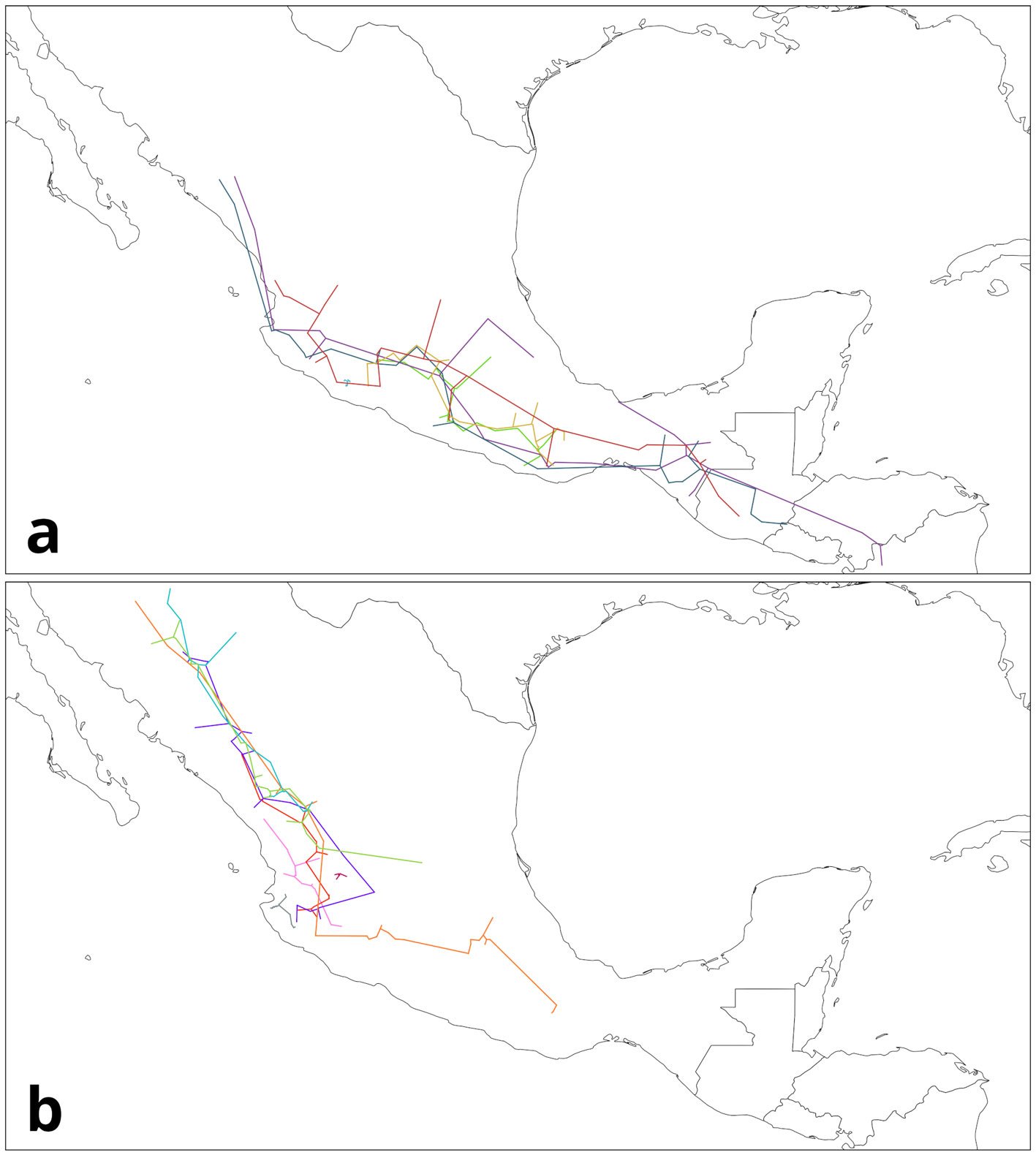
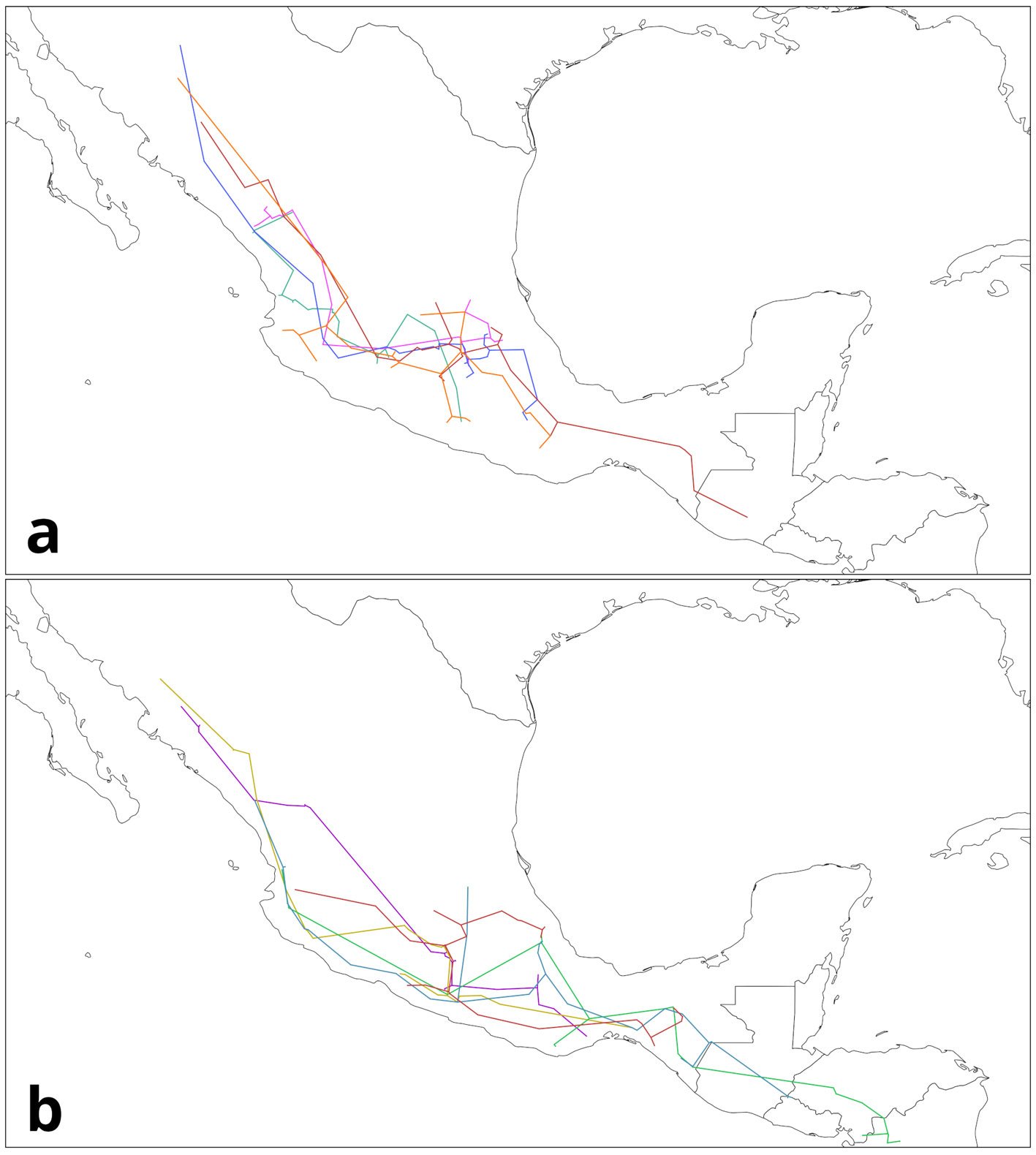
Pseudalcantarea grandis (Schltdl.) Pinzón & Barfuss (Fig. 1A) is a saxicolous tank bromeliad that reaches 2.5 m in height and forms a highly ramified inflorescence in March and April. It is distributed from Central Mexico (Guanajuato, Querétaro, Hidalgo) toward the south (Puebla, Oaxaca, and Chiapas) and into Honduras. It is considered a MegaMexico II endemic species (Espejo-Serna et al., 2010). At the locality of Las Adjuntas, in Hidalgo, the plant is known as “tinaja”, “jarilla”, and “soluche de agua” because its tank can store water. The species grows in crags of the main rivers in the northeastern portion of the region known as Bajío. It grows at elevations ranging from 400 to 1,600 m asl (Espejo-Serna et al., 2010; Rzedowski, 2006).
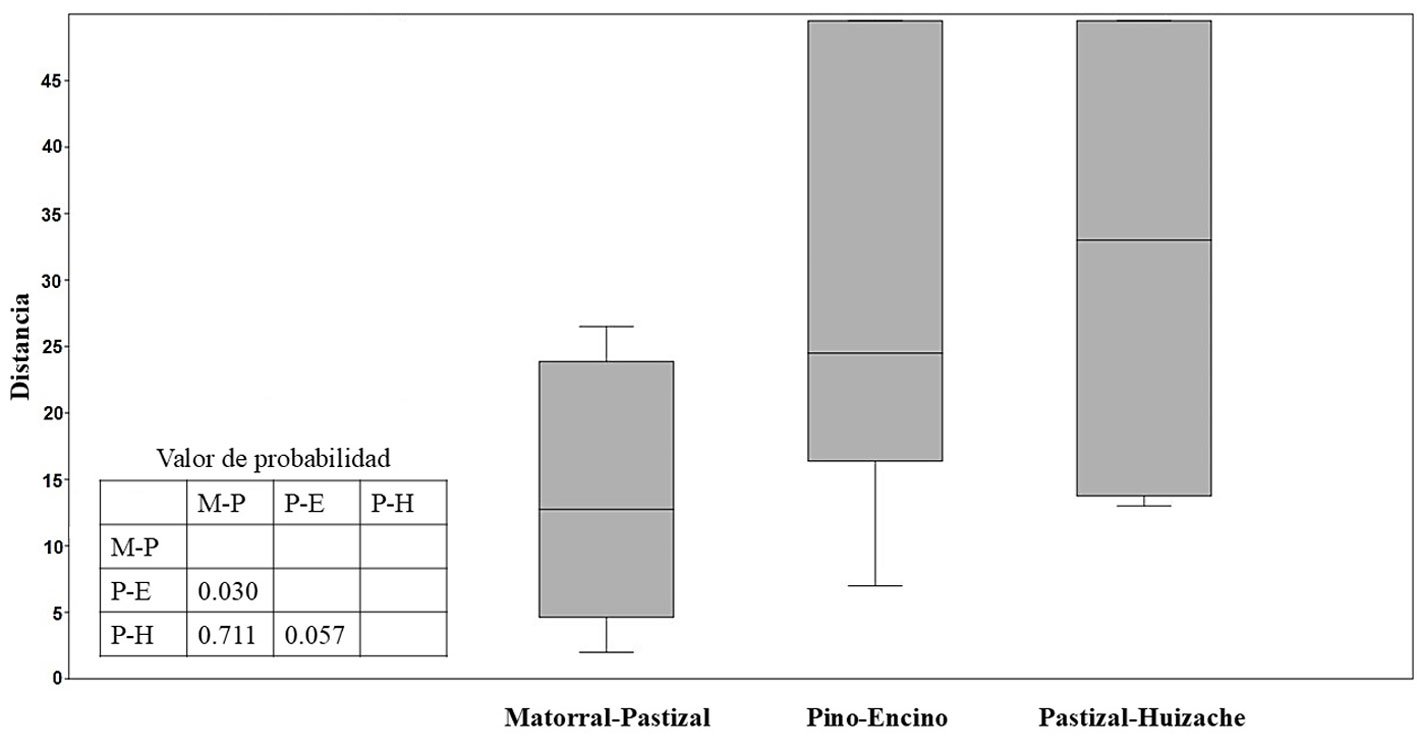
Figure 1. Life form and habitat of the tank bromeliad Pseudalcantarea grandis. A, Plant and inflorescence of Pseudalcantarea grandis; B, Las Angosturas Canyon crags where the tank bromeliad grows.
Metabarcoding studies describing the communities associated with phytotelma have mostly focused on specific groups such as vertebrates (Brozio et al., 2017), ciliates (Simão et al., 2017), or bacteria (Louca et al., 2017; Rodríguez-Núñez et al., 2018). In Pseudalcantarea grandis, 297 bacteria genera were found, with Proteobacteria (37%), Actinobacteria (19%), and Firmicutes (15%) comprising the highest percentage (71%). The main metabolic functions were aerobic chemoheterotrophy and fermentation. However, rare biosphere bacteria were also found, which could favor micro-ecosystem resilience and resistance (Herrera-García et al., 2022). Comprehensive sequencing of the eukaryotic diversity inside tank bromeliads has been performed in tropical zones (Simão et al., 2020), but not arid areas. The objectives of our study were to describe the eukaryotic diversity in Pseudalcantarea grandis phytotelma to understand which vascular plants form the litter, and to infer the putative trophic levels of the phytotelmata in an arid zone.
Materials and methods
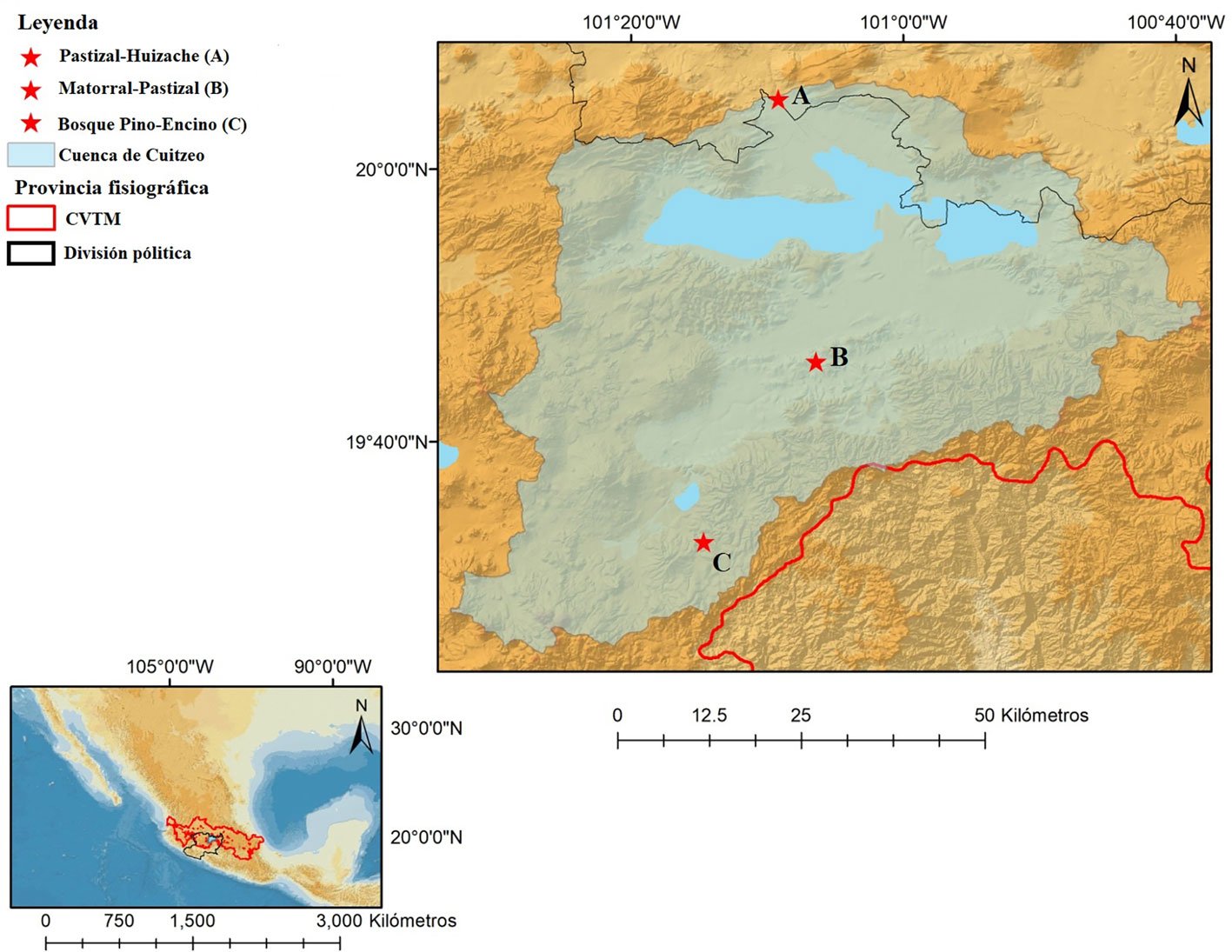
Water samples were collected during the 2018 rainy season at Las Angosturas Canyon, Zimapán, Hidalgo, Mexico (20°50.933’ N, 99°26.7’ W, 900 m asl, see Herrera-García et al. [2022] for a location map) within the Queretano-Hidalguense arid zone, which has been described as a high-diversity and endemicity area (Hernández-Magaña et al., 2017; Hernández & Bárcenas, 1995; Rojas et al., 2013). This arid zone is considered the southernmost portion of the Chihuahuan Desert floristic province. It consists mostly of arid valleys and depressions surrounded by mountains (Hernández & Gómez-Hinostrosa, 2005).
Las Angosturas Canyon is approximately 12 km long with a mixture of xerophytic scrub and tropical deciduous forest (Fig. 1B). Bromeliads grow on vertical crags with different amounts of the surrounding vegetation. Therefore, the tanks are frequently filled with leaf litter. We selected individuals that were accessible enough to be collected by a rappel and were more than 50 cm in diameter. The associated plants are listed in Table 1. To collect the water inside the bromeliads we used Nest® cell scrapers to scratch the inside of each tank, and the water in the bromeliads was vigorously shaken to obtain a homogeneous sample. Water volumes of 50 to 100 ml were collected using 10 ml sterile serological pipettes. The samples were stored in 50 ml conical Falcon tubes, transported on dry ice, and stored at -79 °C until processing. Physicochemical water parameters were not determined.
Table 1
Las Angosturas Canyon floristic inventory. Plants directly above the sampled individuals are noted in the second column. Vouchers and photographs are noted in the third column.
Cylindropuntia imbricata (Haw.) F. M. Knuth ssp. cardenche (Griffiths) U. Guzmán
A. Herrera 12
Cactaceae
Coryphanta sp.
Photographic record
Cactaceae
Echinocereus pentalophus Lem.
Photographic record
Cactaceae
Echinocactus platyacanthus Link & Otto
M. Figueroa 12
Cactaceae
Ferocacutus histrix (DC.) G.E.Linds.
Photographic record
Cactaceae
Mammilaria elongata DC.
surrounding
Photographic record
Cactaceae
Mammillaria longimamma DC.
Photographic record
Cactaceae
Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console
surrounding
Photographic record
Cactaceae
Neobuxbaumia polylopha (DC.) Backeberg
Photographic record
Cactaceae
Opuntia imbricata (Haw.) F. M. Kunth
Photographic record
Cactaceae
Opuntia microdasys (Lehm.) Pfeiff.
Photographic record
Cactaceae
Opuntia rastrera F.A.C. Weber
surrounding
Photographic record
Cactaceae
Stenocereusqueretaroensis (F.A.C.Weber ex Mathes.) Buxb.
Photographic record
Cactaceae
Strombocactus disciformis (DC.) Britton & Rose
Photographic record
Cannabaceae
Celtis pallida Torr.
A. Herrera 2
Capparaceae
Capparis incana Kunth
A. Herrera 5
Convolvulaceae
Ipomoea rzedowskii E. Carranza
A. Herrera 17
Crassulaceae
Echeveria secunda Booth
surrounding
A. Herrera 26
Crassulaceae
Pachyphytum sp.
A. Herrera ND
Crassulaceae
Sedum sp.
surrounding
A. Herrera ND
Euphorbiaceae
Cnidoscolus tubulosus (Muell. Arg.) I.M. Johnst.
A. Herrera 9
Euphorbiaceae
Acalypha monostachya Cav.
A. Herrera 15
Euphorbiaceae
Croton ciliato-glandulifer Ort.
L. Hernández 5029
Euphorbiaceae
Jatropha dioica Sessé ex Cerv.
A. Herrera ND
Euphorbiaceae
Ricinus communis L.
A. Herrera ND
Fabaceae
Acacia berlandieri Benth.
surrounding
Photographic record
Fabaceae
Albizia occidentalis Brandegee
Photographic record
Fabaceae
Bauhinia sp.
Photographic record
Fabaceae
Lysiloma microphylla Bentham
A. Herrera 36
Fabaceae
Mimosa leucaenoides Bentham
surrounding
Photographic record
Fabaceae
Mimosa martindelcampoi F. G. Medrano
Photographic record
Fabaceae
Mimosa puberula Bentham
A. Herrera 3
Fabaceae
Pithecellobium dulce (Roxb.) Bentham
Photographic record
Fabaceae
Neltuma laevigata (Humb. & Bonpl. ex Willd.) Britton & Rose
M. Martínez ND
Fabaceae
Vachellia farnesiana (L.) Willd. & Arn.
A. Herrera 10
Table 1. Continued
Family
Species
Association
Reference at QMEX
Fouquieriaceae
Fouquieria splendens Engelm.
surrounding
Photographic record
Lentibulariaceae
Pinguicula aff. moctezumae Zamudio & R.Z. Ortega
Photographic record
Lythraceae
Heimia salicifolia (Kunth) Link
A. Herrera 13
Onagraceae
Hauya elegans DC.
surrounding
A. Herrera 14
Malpighiaceae
Mascagnia macroptera (Moc. & Sessé ex DC.) Nied.
A. Herrera 8
Malvaceae
Pseudobombax ellipticum (Kunth) Dugand
A. Herrera ND
Malvaceae
Malvaviscus arboreus Cav.
M. Martínez 5317
Myrtaceae
Psidium guajava L.
surrounding
Photographic record
Papaveraceae
Argemone ochroleuca Sweet
M. Martínez 6713
Plantaginaceae
Rusellia polyedra Zucc.
A. Herrera 27
Platanaceae
Platanus mexicana Moric.
A. Herrera 16
Poaceae
Arundinaria sp.
A. Herrera ND
Poaceae
Cenchrus sp.
A. Herrera ND
Poaceae
Cynodon dactylon (L.) Pers.
M. Martínez 3197
Poaceae
Eragrostis sp.
E. Carranza 5251
Primulaceae
Samolus ebracteatus Kunth
A. Herrera 11
Pteridaceae
Argyrochosma formosa (Liebm.) Windham
A. Herrera 23
Pteridaceae
Notholaena affinis (Mett.) T. Moore
A. Herrera 24
Pteridaceae
Notholaena jacalensis Pray
A. Herrera 25
Pteridaceae
Pellaea sp.
A. Herrera 34
Ranunculaceae
Clematis drummondii Torr. & A.Gray
M. Martínez 4434
Rhamnaceae
Karwinskia subcordata Schlecht.
A. Herrera 22
Rubiaceae
Nernstia mexicana (Zucc. & Mart. ex DC.) Urb.
A. Herrera 32
Salicaceae
Neopringlea integrifolia (Hemsl.) S. Watson
A. Herrera 19
Salicaceae
Salix humboldtiana Willd.
Photographic record
Sapindaceae
Dodonaea viscosa (L.) Jacq.
A. Herrera ND
Sapindaceae
Sapindus saponaria L.
A. Herrera ND
Sapindaceae
Serjania sp.
A. Herrera ND
Selaginellaceae
Selaginella lepidophylla (Hook. & Grev.) Spring.
surrounding
Photographic record
Selaginellaceae
Selaginella ribae Valdespino
A. Herrera 29
Selaginellaceae
Selaginella selowii Hieron.
A. Herrera 28
Solanaceae
Datura inoxia Miller
V. Martínez 1
Solanaceae
Nicotiana glauca Graham
Photographic record
Solanaceae
Nicotiana trigonophylla Dunal
O. García 434
Solanaceae
Physalis cinerascens (Dunal) Hitch.
L. Hernández 3769
Solanaceae
Physalisphiladelphica Lam.
A. Herrera 38
Solanaceae
Solanum lycopersicon L.
Photographic record
Taxodiaceae
Taxodium mucronatum Ten.
M. Martínez 3237
Urticaceae
Urera sp.
H. Rubio 302
Zygophyllaceae
Morkillia acuminata Rose & Painter
surrounding
A. Herrera 18
DNA extraction
Water samples were homogenized and 100 ml was filtered through a 0.22 µm Millipore® nitrocellulose membrane. The membrane was frozen and macerated in liquid nitrogen. We extracted total DNA using the QIAmp DNA Extraction® kit following the manufacturer’s instructions by duplicate to obtain pseudoreplicates and verify reproducibility. DNA quality and concentration were evaluated using NanoDrop® spectrophotometry.
Amplicon sequencing
To characterize eukaryotic diversity, we amplified a portion of the 5.8 S Internal Transcibed Spacer (ITS) with the ITS1 and ITS2 primers designated by White et al. (1990). The PCR reaction consisted of a final volume of 25 µl, that contained 2 mM of dNTP´s, 2mM µl of each primer, 0.4% DMSO, 0.4 % BSA, 2.5 mM MgCl2, 1.2 mM Buffer, 1.25 U Platinum Taq, 60ng/µl DNA and H2O. Thermocycler conditions were an initial step at 95 °C for 3 min, followed by 30 cycles at 95 °C for 1 min; 52 °C, 45 s, and 72 °C, 2 min, with a final extension step at 72 °C for 5 min. Amplicons were purified with Agencourt® AMPure® XP.
To construct the libraries, we used the Ion Plus Fragment Library kit. The presence, size, and concentration of the fragment were analyzed using Bioanalyzer 2100 with high-sensitivity DNA assay (Agilent). Libraries were quantified using real-time PCR to obtain an equimolar dilution factor to mix the 3 libraries. The template was prepared using a PCR emulsion in the Ion One Touch 2 System (Life Technologies) and quantified by fluorometry in Qubit® 3.0 (Thermo Fisher Scientific). Finally, the template was loaded onto the PGM 318TM chip using the 400-pair base fragment sequencing kit, according to the Ion PGM™ Hi‑Q™ View Sequencing Kit protocol.
We sampled the vascular plants growing at the canyon, directly above the bromeliad, and also the surrounding vegetation. Voucher specimens were deposited at QMEX herbarium. We compared the similarity of the plant inventory obtained by sequencing against the floristic list obtained by field sampling through a Sorensen coefficient analysis at the family level.
Sequencing quality was evaluated using the FastQC program. Sequences were filtered by the quality value of Phred > 20. We selected sequences larger than 100 bp in the CLC Genomic Workbench v.11.01 (QIAGEN Bioinformatics, Aarhus, Denmark) platform. In the Microbial Genomics module application, we performed an analysis based on the amplicons to aggregate the sequences in operational taxonomic units (OTUs) considering only 99% similarity among them. We deleted unique reads and chimeras. Taxonomic assignation of the amplicons was performed with BLAST in 2023 via the Genbank at NCBI database (Altschul et al., 1990). Over 90% of the OTUs had identity percentages higher than 95% and only 54 OTUs had lower percentages, ranging from 75 to 80%. We manually reviewed them and corroborated the genus of each one. Since BLAST provides determinations at the genus and species levels, we used MEGAN Community Edition V. 6.24.4 to assign kingdom, phylum, class, order, and family. We loaded the BLAST results using the lowest basal common ancestor assignation algorithm (LCA). The analysis is based on the taxonomic hierarchies recognized by NCBI and the results are displayed as a phylogenetic tree that allows simple observation of taxonomic diversity (Huson et al., 2016). We manually reviewed the classifications and to corroborate the taxonomic assignations we used the classification proposed by Simpson (2006) for plants, and Tree of Life (2022) for the other eukaryotic marine taxa (such as Cnidaria). The OTUs at the genus level were used to define the total eukaryotic group diversity present in our sample.
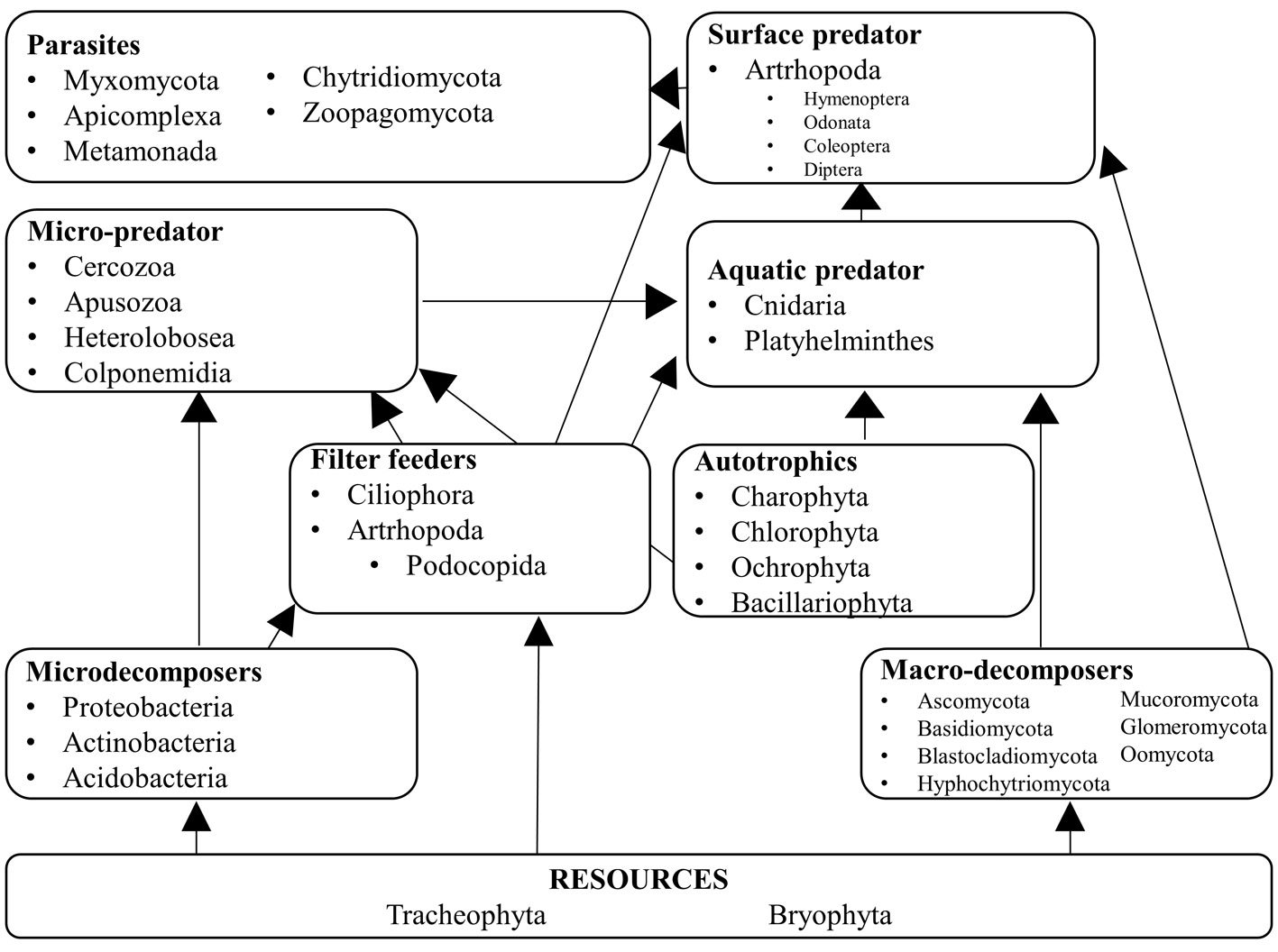
We assigned the ecological function of each genus following the criteria of Mogi (2004) and Brouard et al. (2012). We considered 9 categories: 1) detritus formed by leaf litter and vegetal matter that serves as the main resource for the trophic network; 2) micro decomposers integrated by bacteria (Herrera-García et al., 2022); 3) macro decomposers formed by saprobiotic fungi; 4) filter feeders that use small particles including microorganisms that are in turn consumed by aquatic and surface predators; 5) photosynthetic organisms that require sunlight and serve as food for predators; 6) micro predators that feed on photosynthetic organisms, filter feeders, and micro-decomposers; 7) aquatic predators which are macroinvertebrates that live in the water column and feed mostly on algae and bacteria; 8) surface predators which are restricted to the uppermost portion of the water column and feed on protists, bacteria, and algae; and 9) parasites which are obligate vertebrate parasites that use arthropods as vectors.
Results
The 5 sampled plants had volumes that varied from 50 to 100 ml. The 2 pseudoreplicates were compared and considered as a single pool due to the similarity of the resulting OTUs. We obtained a total of 3,276,538 lectures. After quality and size filtration, the number of useful lectures was reduced to 1,284,998 representing a total of 23,948 OTUs, 762 of which could not be assigned to the species or genus taxonomic category provided by BLAST.
The diversity of organisms living in the tank consisted of 26 phyla and 543 genera. See Supplementary material T1 for a list of assigned taxa. Fungi were dominant, as 80% of the genera belonged to Ascomycota, Basidiomycota, Blastocladiomycota Chytridiomycota, Glomeromycota, Mucoromycota, and Zoopagomycota phyla. The remaining 20% was composed of 19 phyla: Apicomplexa, Apusozoa, Arthropoda, Bacillariophyta, Bryophyta, Cercozoa, Charophyta, Chlorophyta, Ciliophora, Cnidaria, Colponemidia, Heterolobosea, Hyphochytriomycota, Metamonada, Myxomycota, Ochrophyta, Oomycota, Platyhelminthes, and Tracheophyta (Fig. 2). We identified 25 genera of photosynthetic algae from the phyla Bacillariophyta (with the genera Pseudo-nitzschia, Navicula, and Stephanodiscus), Charophyta (Staurastrum), Chlorophyta (Leskea, Didymogenes, Meyerella, Dolichomastix, Bathycoccus, Mychonastes, Trebouxia, Chamaetrichon, Hazenia, Chloroidium, and Pleurastrum), and Ochrophyta (Nannochloropsis). The other photosynthetic organisms were Bryophyta (Fontinalis, Leskea, and Thuidium).
Protists were represented by 44 genera, 4 of which are relevant to human health: Plasmodium (Apicomplexa) which causes paludism, Giardia (Metamonada) which is responsible for giardiasis, Neobalantidium (Cilliophora) that causes balantidiosis, and Spirometra (Plathelmyntes) which is responsible for sparganosis.
Tracheophyta (vascular plants) do not live in the phytotelma but do constitute the debris that accumulates in the bromeliad. They comprised 11% of the identified genera. We found a Sorensen coefficient of 45% similarity among the methods in which 14 taxa at the family level were shared (Supplementary material F1). Arthropoda were represented by 13 genera of the Coleoptera, Diptera, Hymenoptera, Lepidoptera, Odonata, and Pocopodia orders.
Of the eukaryotic organisms, 79% were classified as decomposers, and 7% were classified as micro-predators. In the tank, 3% were photosynthetic organisms, 2% were parasites, and the remaining 9% corresponded to the vascular plants that constitute the detritus. Tracheophyta and Bryophyta constituted the vegetal resources available to micro- and macro-decomposers, and filter feeders. Cercozoa, Apusozoa, Heterolobosea, and Colponemidia were considered micro-predators because they consume some photosynthetic organisms, filter feeders, and micro-decomposers. Ciliophora and 2 Arthropoda genera (Cyprideis and Notodromas) were some of the filter feeders. Apicomplexa, Chytridiomycota, Metamonada, Myxomycota, and Zoopagomycota were the parasites. Aquatic predators were mostly metazoans (Cnidaria and Platyhelminthes) that use filter feeders, photosynthesizers, and macro decomposers as resources. The arthropods Coleoptera, Diptera, Hymenoptera, Lepidoptera, and Odonata were part of the uppermost categories of the trophic network (surface predators). However, their exoskeletons and/or excretions become part of the tank resource or contribute to the nutrient cycling of the microecosystem (Figs. 3, 4).
Figure 2. Percentage of eukaryotic diversity present in the Pseudalcantarea grandis tank.
Discussion
Eukaryotic composition of the Pseudalcantarea grandis community
The abundance of fungi in the Pseudalcantarea tank appeared to correlate with their function in the trophic network. Fungi are the most important degrading group, responsible for organic decomposition and nutrient recycling in forests, aquatic ecosystems, and the phytotelma (Costa & Gusmão, 2015; Grossart et al., 2019; Grothjan et al., 2019). Fungi also have multiple functions in aquatic environment interactions that favor antagonistic and symbiotic members of the community. They can be predators, parasites, or food for heterotrophic protists. Some can use organic matter, pollen, or zooplankton exoskeletons (Zoopagomycota, Chytridiomycota) (Grossart et al., 2019). Vegetal matter decomposition enhances detritus quality for detritivores degrading vegetal polysaccharides into monosaccharides through enzymatic reactions which are then easily digested by microorganisms (Krauss et al., 2011). Fungi and protist interactions for vegetal matter transformation are poorly documented. A symbiotic relationship between them is unknown and difficult to study because of the microscopic scale at which they occur (Grossart et al., 2019).
The fungal diversity found in the Pseudalcantarea grandis tank was high compared to that reported in previous studies of other Bromeliaceae species. We found 436 genera, whereas Gomes et al. (2015) identified 36 genera using cultivation techniques. Other papers already pointed out that metagenomic studies detect higher diversity levels than other techniques (Simão et al., 2020). The 36 genera found by Gomes et al. (2015) were also found in P. grandis. Cryptococcus, Candida, and Aureobasidium have specific enzymatic activity in plant material degradation, which suggests that degradation reactions by these organisms are frequent in the phytotelma. The primers used in our study were developed for fungi (White et al., 1990), therefore they might be overrepresented.
We found 44 protist genera in the phytotelma. Some have mixotrophic nutrition, in that they obtain their energy through photo- and heterotrophy depending on the environmental conditions in which they grow (Jones, 2000). In aquatic environments where light is available but dissolved organic carbon (DOC) is scarce, photosynthetic organisms are better represented. Low light and high DOC favor heterotrophic organisms (Jones, 2000). The latter condition is what was present in the sampled tanks. Therefore, the development of photosynthetic protists is not favored because of the large and abundant bacterial community that competes for elements such as phosphorus (Brouard et al., 2012; Herrera-García et al., 2022). In the rainy season, the tank of P. grandis is surrounded by vegetation that intercepts light and deposits leaf litter, therefore favoring the conditions for fungi and decomposers (Grossart et al., 2019; Herrera-García et al., 2022, Kitching, 2000).
Direct observations and sampling of the phytotelma of tropical zones have revealed Diptera, Odonata, Oligochaeta, Ostracoda, beetles, copepods, pseudoscorpions, scorpions, isopods, Lepidoptera, hemipterans, homopterans, orthopterans, and arachnids in tank bromeliads (Cutz-Pool et al., 2016; Marino et al., 2013). Using environmental DNA with specific primers, amphibians (Brozio et al., 2017) and ciliates (Simão et al., 2017) have been found in tank bromeliads with high water availability. These results suggest that aridity and strong water seasonality in our study area were responsible for the lack of amphibians and the low arthropod and ciliate diversity we found.
Phytotelma seasonality is an important factor. In a rainy forest, the annual precipitation is 3,000 mm and rain is present for 280 days, therefore the water in the tank lasts longer (Brouard et al., 2012). In contrast, in our location, the annual precipitation is 391 mm and the phytotelma is available only through the rainy season from May to June (INFAED, 2012). The presence of Plasmodium is relevant since its most common vector is Aedes, suggesting that at some point during the phytotelma duration, the mosquito is in contact with the water, completing the parasite life cycle (Williams, 2007).
The absence of vertebrates is characteristic of phytotelma communities (Mogi, 2004). One exception is in rainforest bromeliad, where bromeliad tadpoles can be found. We did not find vertebrates. Strong seasonality was probably the reason for their absence. Not all OTUs could be assigned to the species or genus taxonomic category at 99% identity level we used. We did not find an identity for 762 of the 23,984 sequences, possibly because the sequences of these organisms are not available in the NCBI database, or because the organisms have not yet been described.
The tracheophytes found in the phytotelma could not be identified at the generic level. There are 2 possible explanations: the reads were short (150 bp) and therefore insufficient, or the genera growing at the canyon are not in the GenBank NCBI (National Center for Biotechnology Information) database. However, 30 families of vascular plants were detected, 16 of which correspond to the families found by field collection.
Trophic structure of the Pseudalcantarea grandis tank
We propose 9 trophic levels for tank bromeliads in arid zones, —2 more than those suggested by Mogi (2004), and 3 more than those suggested by Brouard et al. (2012). Detritus is the main nutrient source. Macrodecomposers process leaf litter into small organic matter particles, including their waste. The particles are then stored in the phytotelma where filterers and invertebrates process them. Dead organisms, feces, and leaf litter stored at the bottom of the tank are used by bacteria and other microorganisms such as fungi to assimilate nutrients (Brouard et al., 2012). We found that 48 plant genera (45 Tracheophyta, and 3 Bryophyta) constitute the detritus, and although Brouard et al. (2012) recognized organic litter as a resource, they did not identify the organisms that provided it. The large amount of detritus was due to the type of surrounding vegetation, which was a tropical deciduous forest in this study. In P. grandis macro decomposers are fungi of the Ascomycota, Basidiomycota, and Myxomycota phyla. We concur with Brouard et al. (2012) that ciliates are filterers. Mogi (2004) did not consider microorganisms to be autotrophs, but Brouard et al. (2012) included that category, and in P. grandis algae constitute this level. Insects were categorized as surface predators in their diagram; they include the odonate family Coenagrionidae, which includes the 2 genera we found in P. grandis, Ischnura, and Agriocnemis (Brouard et al., 2012). Neither Mogi (2004) nor Brouard et al. (2012) included parasites at a trophic level, but some of the genera found in P. grandis are obligate parasites, mostly from the Alveolata kingdom, which has birds and mammals as hosts but uses arthropods as vectors.
We found 26 phyla with 543 genera in the phytotelma of P. grandis growing in an arid zone. The identified diversity suggested that the organisms that inhabit these small ephemeral water bodies are adapted to prolonged dry spells and develop quickly when the phytotelma has water. The biota was mostly composed of fungi (over 80% of the diversity) that specialize in plant detritus degradation. Water bodies shelter aquatic groups that cannot exist in areas outside the P. grandis phytotelma. It is easier to assess the diversity of organisms within a tank than to comprehend their interactions. The trophic network proposed for eukaryotes indicates that they fulfill different functions.
As final considerations, we conclude that the analyzed phytotelma had a large detritus accumulation, and water was present briefly. Most of the diversity belonged to fungi (80%) because of the large amount of plant detritus in the tank. Photosynthesizers were scarce but included 25 algal genera and 3 Bryophyta. We found 45% Sorensen coefficient similarity between the plant detritus and the specimens collected with herbarium specimens. We also found a low arthropod and ciliate diversity, and the tank also harbors protist genera, some of which have medical implications. We found 9 trophic levels in the tank. Unlike tropical areas, in which algal production can support non-detrital food webs, in our arid zone system, detritus degradation was the main energy source.
Acknowledgments
Funding was provided by Conahcyt through grant 293833 for the Laboratorio Nacional de Identificación y Caracterización Vegetal. Diana Velázquez designed and executed Figure 3. Two anonymous reviewers helped to improve the manuscript with their comments.
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Benzing, D. H., & Renfrow, A. (1974). The mineral nutrition of Bromeliaceae. Botanical Gazette, 135, 281–288. https://doi.org/10.1086/336762
Benzing, D. H. (2000). Bromeliaceae: profile of an adaptive radiation. Cambridge, U.K.: Cambridge University Press.
Bernal, R., Valverde, T., & Hernández-Rosas, L. (2006). Habitat preference of the epiphyte Tillandsia recurvata (Bromeliaceae) in a semi-desert environment in Central Mexico. Canadian Journal of Botany, 83, 1238–1247. https://doi.org/10.1139/b05-076
Brandt, F. B., Martinson, G. O., & Conrad, R. (2017). Bromeliad tanks are unique habitats for microbial communities involved in methane turnover. Plant and Soil, 410, 167–179. https://doi.org/10.1007/s11104-016-2988-9
Brouard, O., Céréghino, R., Corbara, B., Leroy, C., Pelozuelo, L., Dejean, A. et al. (2012). Understorey environments influence functional diversity in tank-bromeliad ecosystems. Freshwater Biology, 57, 815–823. https://doi.org/10.1111/j.1365-2427.2012.02749.x
Brozio, S., Manson, C., Gourevitch, E., Burns, T. J., Greener, M. S., Downie, J. R. et al. (2017). Development and application of an eDNA method to detect the critically endangered Trinidad golden tree frog (Phytotriades auratus) in bromeliad phytotelmata. Plos One, 12, 1–8. https://doi.org/10.1371/journal.pone.0170619
Costa, L. A., & Gusmão, L. F. P. (2015). Characterization saprobic fungi on leaf litter of two species of trees in the Atlantic Forest, Brazil. Brazilian Journal of Microbiology, 46, 1027–1035. https://doi.org/10.1590/S1517-838246420140548
Cutz-Pool, L. Q., Ramírez-Vázquez, U. Y, Castro-Pérez, J. M, Puc-Paz, W. A., & Ortiz-León, H. J. (2016). La artropo- dofauna asociada a Tillandsia fasciculata en bajos inundados de tres sitios de Quintana Roo, México. Entomología Mexicana, 3, 576–581.
Espejo-Serna, A., & López-Ferrari, A. R. (2018). La familia Bromeliaceae en México. Botanical Sciences, 96, 533–554. https://doi.org/10.17129/botsci.1918
Espejo-Serna, A., López-Ferrari, A., & Ramírez-Murillo, I. (2010). Bromeliaceae. Flora del Bajío y Regiones Adyacentes, 165, 1–82.
Goffredi, S. K., Kantor, A. H., & Woodside, W. T. (2011). Aquatic Microbial Habitats Within a Neotropical Rain- forest: Bromeliads and pH-Associated Trends in Bacterial Diversity and Composition. Microbial Ecology, 61, 529–542. https://doi.org/10.1007/s00248-010-9781-8
Gomes, F. C. O., Safar, S. V. B., Marques, A. R., Medeiros, A. O., Santos, A. R. O., Carvalho, C. et al. (2015). The diversity and extracellular enzymatic activities of yeasts isolated from water tanks of Vriesea minarum, an endangered bromeliad species in Brazil, and the description of Occultifur brasiliensis f.a., sp. nov. Antonie van Leeuwenhoek, International Journal of General and Molecular Microbiology, 107, 597–611. https://doi.org/10.1007/s10482-014-0356-4
Gouda, E. J., Butcher, D., & Dijkgraaf, L. (2024) Encyclopaedia of Bromeliads, Version 5. Utrecht University Botanic Gardens, online (accessed: 02-10-2024): http://bromeliad.nl/encyclopedia/
Grossart, H. P., Van den Wyngaert, S., Kagami, M., Wurzbacher, C., Cunliffe, M., & Rojas-Jimenez, K. (2019). Fungi in aquatic ecosystems. Nature Reviews Microbiology, 17, 339–354. https://doi.org/10.1038/s41579-019-0175-8
Grothjan, J. J., & Young, E. B. (2019). Diverse microbial communities hosted by the model carnivorous pitcher plant Sarracenia purpurea: Analysis of both bacterial and eukaryotic composition across distinct host plant populations. PeerJ, 2019, e6392. https://doi.org/10.7717/peerj.6392
Hernández, H. M., & Bárcenas, R. T. (1995). Endangered Cacti in the Chihuahuan Desert: I. Distribution Patterns. Conservation Biology, 9, 1176–1188. https://doi.org/10.1046/j.1523-1739.1995.9051169.x-i1
Hernández, H. M., & Gómez-Hinostrosa, C. (2005). Cactus diversity and endemism in the Chihuahuan Desert Region. EnJ. L. Cartron, G. Ceballos, & R. Felger (Eds.), Biodiversity, ecosystems and conservation in Northern Mexico (pp264–275). New York: Oxford University Press.
Hernández-Magaña, R., Hernández-Oria, J. G., & Chávez, R. (2017). Datos para la conservación florística en función de la amplitud geográfica de las especies en el Semidesierto Queretano, México. Acta Botanica Mexicana, 140, 105. https://doi.org/10.21829/abm99.2012.22
Herrera-García, J. A., Martínez, M., Zamora-Tavares, P., Vargas-Ponce, O., Hernández-Sandoval, L., & Rodríguez-Zaragoza, F. A. (2022). Metabarcoding of the phytotelmata of Pseudalcantarea grandis (Bromeliaceae) from an arid zone. PeerJ, 10, e12706. https://doi.org/10.7717/peerj.12706
Huson, D. H., Beier, S., Flade, I., Górska, A., El-Hadidi, M., Mitra, S. et al. (2016). MEGAN Community Edition – Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. Plos Computational Biology, 12, 1–12. https://doi.org/10.1371/journal.pcbi.1004957
INAFED (Instituto Nacional para el Federalismo y el Desarrollo Municipal). (2012). Zimapán. Enciclopedia de los municipios y Delegaciones de México. http://www.inafed.gob.mx/work/enciclopedia/EMM13hidalgo/municipios/13084a.html
Jones, R. I. (2000). Mixotrophy in planktonic protists: an overview. Freshwater Biology, 45, 219–226. https://doi.org/10.1046/j.1365-2427.2000.00672.x
Kitching, R. (2000). Food webs and container habitats: the natural history and ecology of phytotelma. Cambridge, U.K.: Cambridge University Press. https://doi.org/10.1017/CBO9780511542107
Kitching, R. L. (2001). Food webs in phytotelmata: “Bottom-Up” and “Top- Down’” Explanations for Community Structure. Annual Review of Entomology, 46, 729–760. https://doi.org/10.1146/annurev.ento.51.110104.151120
Krauss, G. J., Solé, M., Krauss, G., Schlosser, D., Wesenberg, D., & Bärlocher, F. (2011). Fungi in freshwaters: Ecology, physiology and biochemical potential. FEMS Microbiology Reviews, 35, 620–651. https://doi.org/10.1111/j.1574-6976.20 11.00266.x
Louca, S., Jacques, S. M. S., Pires, A. P. F., Leal, J. S., González, A. L., Doebeli, M. et al. (2017). Functional structure of the bromeliad tank microbiome is strongly shaped by local geochemical conditions. Environmental Microbiology, 19, 3132–3151. https://doi.org/10.1111/1462-2920.13788
Marino, N. A. C., Srivastava, D. S., & Farjalla, V. F. (2013). Aquatic macroinvertebrate community composition in tank-bromeliads is determined by bromeliad species and its constrained characteristics. Insect Conservation and Diversity, 6, 372–380. https://doi.org/10.1111/j.1752-4598.2012.00224.x
Mogi, M. (2004). Phytotelmata: hidden freshwater habitats supporting unique faunas. In C. M. Yule, & H. S. Yong (Eds.), Freshwater invertebrates of the Malaysian region (pp. 13–22). Kuala Lumpur, Malaysia: Academy of Sciences Malaysia.
Ngai, J. T., & Srivastava, D. S. (2006). Predators accelerate nutrient cycling in a bromeliad ecosystem. Science, 314, 963. https://doi.org/10.1126/science.1132598
Nievola, C. C., Mercier, H., & Majerowicz, N. (2001). Levels of Nitrogen assimilation in bromeliads. Journal of Plant Nutrition, 24, 1387–1398. https://doi.org/10.1081/PLN-100106989
Ramírez-Murillo, I., Carnevali-Fernández, G., & Chi-May, F. (2004). Guía ilustrada de las Bromeliaceae de la porción mexicana de la península de Yucatán. Yucatán: Centro de Investigación Científica de Yucatán.
Ramos, G. J. P., & do Nascimento Moura, C. W. (2019). Algae and cyanobacteria in phytotelmata: diversity, ecological aspects, and conservation. Biodiversity and Conservation, 28, 1667–1697. https://doi.org/10.1007/s10531-019-01771-2
Rodríguez-Núñez, K. M., Rullan-Cardec, J. M., & Ríos-Velázquez, C. (2018). The metagenome of bromeliads phytotelma in Puerto Rico. Data in Brief, 16, 19–22. https://doi.org/10.1016/j.dib.2017.10.065
Rojas, S., Castillejos-Cruz, C., & Solano, E. (2013). Florística y relaciones fitogeográficas del matorral xerófilo en el valle de Tecozautla, Hidalgo, México. Botanical Sciences, 91, 273–294. https://doi.org/10.17129/botsci.8
Rzedowski, J. (2006). Vegetación de México. México D.F.: Conabio.
Simão, T. L. L., Borges, A. G., Gano, K. A., Davis-Richardson, A. G., Brown, C. T., Fagen, J. R. et al. (2017). Characterization of ciliate diversity in bromeliad tank waters from the Brazilian Atlantic Forest. European Journal of Protistology, 61, 359–365. https://doi.org/10.1016/j.ejop.2017.05.005
Simão, T. L. L., Utz, L. R. P., Dias, R., Giongo, A., Triplett, E. W., & Eizirik, E. (2020). Remarkably complex microbial community composition in bromeliad tank waters revealed by eDNA metabarcoding. Journal of Eucaryotic Microbiology, 67, 593–607. https://doi.org/10.1111/jeu.12814
Simpson, M. G. (2006). Plant systematics. Amsterdam: Elsevier/Academic Press.
Takahashi, C. A., & Mercier, H. (2011). Nitrogen metabolism in leaves of a tank epiphytic bromeliad: characterization of a spatial and functional division. Journal of Plant Physiology, 168, 1208–1216. https://doi.org/10.1016/j.jplph.2011.01.008
Tree of Life Web Project, (2019). Tree of life. Website. http://tolweb.org/tree/
White, T. J., Bruns, T., Lee, S., & Taylor, J. (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for Phylogenetics. In M. A. Innis, D. H. Gelfand, J. J. Sninsky, T. J. White (Eds.), PCR Protocols(January) (pp. 315–322). San Diego, CA: Academic Press. https://doi.org/10.1016/b978-0-12-372180-8.50042-1
Williams, D. D. (2007). Other temporary water habitats. The Biology of temporary waters. Oxford, UK: Oxford University Press. https://doi.org/10.1093/acprof:oso/9780198528128.001.0001
Received: 28 February 2024; accepted: 14 February 2025
Abstract
The nematode Metaparasitylenchus hypothenemi (Poinar) (Tylenchida: Allantonematidae) is a parasite of the coffee berry borer (Hypothenemus hampei), one of the most significant pests affecting coffee crops. This study aimed to analyze the genetic variation of M. hypothenemi using the mitochondrial cytochrome oxidase subunit I (COI) gene. Reproductive females of M. hypothenemi were extracted from parasitized wild coffee berry borers for DNA extraction, amplification, and sequencing. Phylogenetic analyses revealed 2 well-differentiated lineages and 6 haplotypes distributed across 18 populations along a 100 km transect. Global genetic diversity was moderate (Hd = 0.52 ± 0.06), with a dominant haplotype present in 16 populations and others being representative or geographically isolated. Populations showed high genetic differentiation and restricted gene flow. These findings confirm the utility of the COI gene for analyzing the genetic variation of M. hypothenemi and provide a molecular basis for future studies on its biology and management.
Variabilidad genética del nemátodo Metaparasitylenchus hypothenemi (Tylenchida:Allantonematidae), parásito natural de la broca del café (Coleoptera: Curculionidae: Scolytinae) usando secuencias de COI
Resumen
El nemátodo Metaparasitylenchus hypothenemi (Poinar) (Tylenchida: Allantonematidae) es un parásito de la broca del café (Hypothenemus hampei), una de las principales plagas del cultivo de café. Este estudio tuvo como objetivo analizar la variación genética de M. hypothenemi mediante el gen mitocondrial citocromo oxidasa subunidad I (COI). Se obtuvieron hembras reproductoras de M. hypothenemi de brocas silvestres parasitadas para la extracción, amplificación y secuenciación de DNA. Los análisis filogenéticos revelaron 2 linajes bien diferenciados y 6 haplotipos distribuidos en 18 poblaciones a lo largo de un transecto de 100 km. La diversidad genética global fue moderada (Hd = 0.52 ± 0.06), con un haplotipo dominante en 16 poblaciones y otros representativos o aislados geográficamente. Las poblaciones mostraron una alta diferenciación genética y flujo genético restringido. Estos resultados confirman la utilidad del gen COI para analizar la variación genética de M. hypothenemi y proporcionan bases moleculares para futuros estudios sobre su biología y manejo.
Palabras clave: Endoparásito obligado; Plaga de insectos; Variación genética; Gen COI; Árbol filogenético; Estructura genética
Introduction
The nematode, Metaparasitylenchus hypothenemi (Poinar) (Tylenchida: Allantonematidae), is an obligate endoparasite of the coffee berry borer (CBB), Hypo- thenemus hampei (Ferrari) (Coleoptera: Curculionidae, Scolytinae), the most important insect pest of coffee worldwide (Le Pelley, 1968). Metaparasitylenchus hypothenemi was discovered in a commercial coffee plantation in southeastern Mexico attacking H. hampei adults (Castillo et al., 2002). M. hypothenemi reduce the fecundity and longevity of the females H. hampei (Castillo et al., 2019). This parasite-host relationship highlights the importance of studying its genetic variability and understanding its ecological role. The study of genetic variability in parasitic nematodes like M. hypothenemi requires molecular tools that enable precise population characterization. Among these, the mitochondrial gene cytochrome oxidase subunit I (COI) has proven to be an efficient marker for identifying specimens, clarifying evolutionary relationships among species, and analyzing population divergence (Hebert et al., 2003; Marsjan & Oldenbroek, 2007). This marker has been successfully used to analyze population structure and infer phylogenetic relationships in related nematodes, including parasites and entomopathogens such as Heterorhabditis marelatus and Deladenus proximus (Blouin et al., 1999; Fitza et al., 2019; Hartshorn et al., 2017; Saeb & David, 2014). However, there are few nematode sequences in both the BOLD Systems database and GenBank, and those corresponding to insect-parasitic nematodes are even scarcer, with M. hypothenemi being a species without previous genetic studies. The objective of this study was to analyze the genetic variability, in terms of the phylogeny, frequency of haplotypes and genetic differentiation between popula- tions of the nematode Metaparasitylenchus hypothenemi collected from Soconusco, the only region in the world where this nematode has been systematically recorded.
Materials and methods
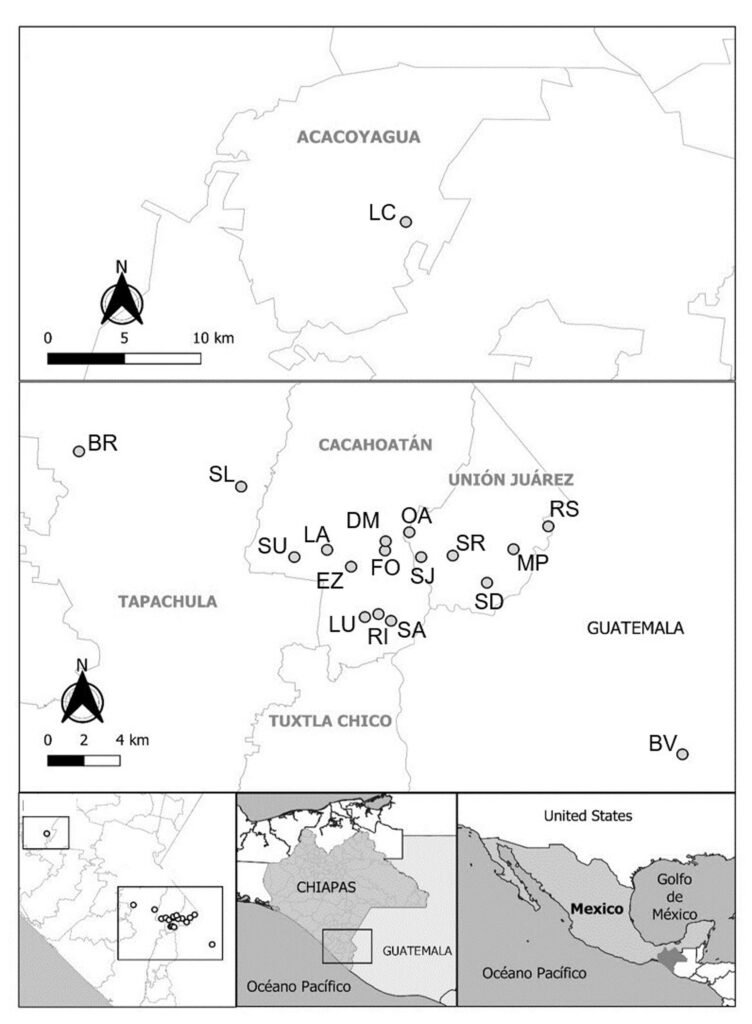
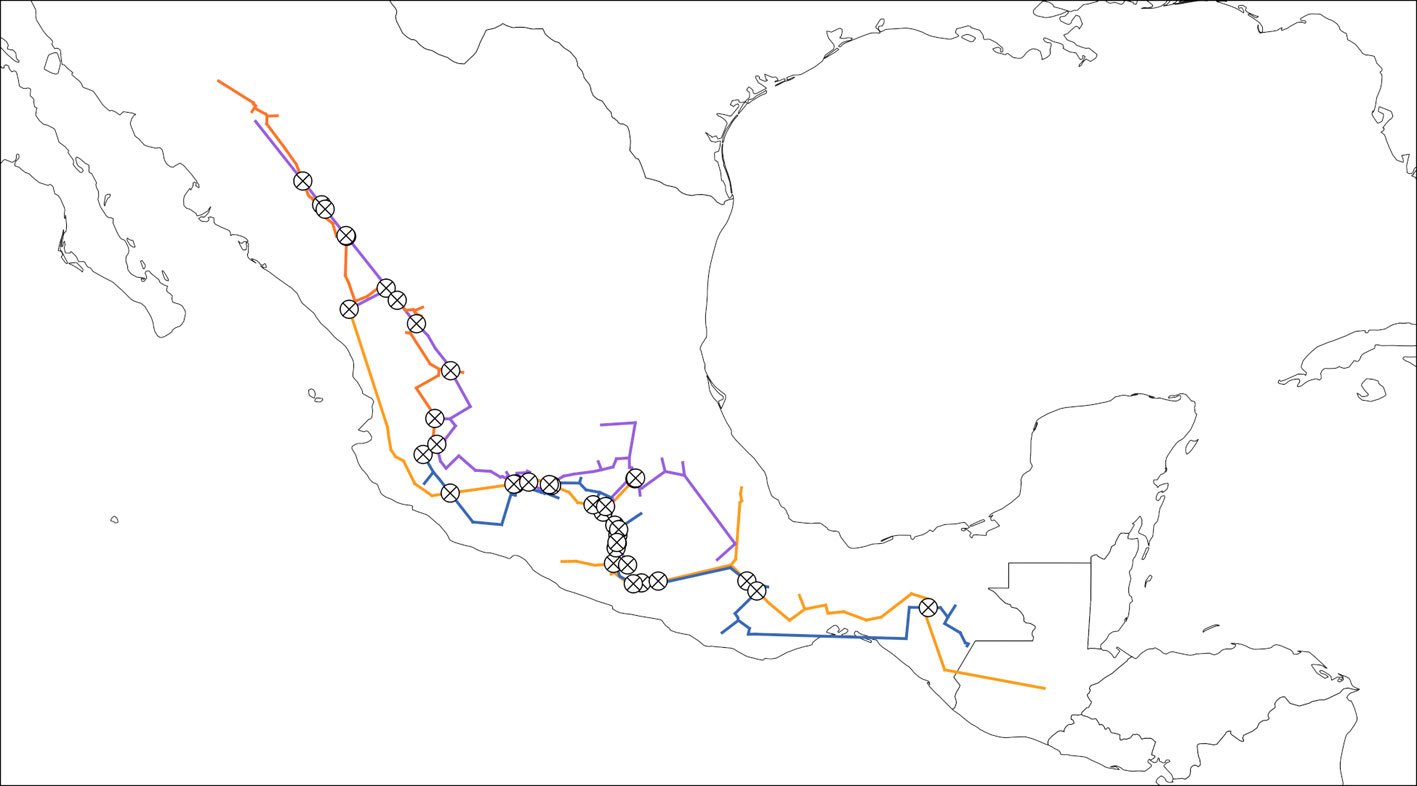
Five reproductive females of M. hypothenemi were collected in each of 18 localities from the Soconusco region, Chiapas, Mexico (Table 1, Fig. 1). Reproductive females of M. hypothenemi were extracted from the abdominal cavity of H. hampei, which were obtained from 100 coffee berries collected from coffee plants per locality. Reproductive females were selected for sampling due to their larger size compared to males, which facilitated DNA extraction and ensured the viability of the samples. Moreover, their genetic material plays a key role in transmitting genetic diversity to subsequent generations (Stewart & Larsson, 2014). The nematodes were stored in 2.5 ml microtubes containing 96% alcohol at -20 °C, until molecular analysis. The samples obtained in each sampled locality were considered as a population.
DNA extraction, amplification, and sequencing. A total of 90 nematodes of M. hypothenemi were used for DNA extraction. Total DNA from each specimen was extracted from the body complete of the nematode using a standard glass fiber method (Ivanova et al., 2006). A barcode region of approximately 658 base pairs (bp) of the mitochondrial COI gene was amplified using the primers ZplankF1_t1 (5’-TGTAAAACGACGGCCAGTTCTASWAATCATAARGATATTGG-3’) and ZplankR1_t1 (5’-CAGGAAACAGCTATGACTTCAGGRTGRCCRAARAATCA-3’) (Prosser et al., 2013). The final volume of the PCR mix was 12.5 µl, containing 0.12 µl of each primer (0.01 µM), 0.6 µl of MgCl2 (50 mM), 2 µl of ultrapure water, 0.06 µl of each dNTP (0.05 mM), 1.25 µl of 10X PCR buffer, 6.25 µl of trehalose 10%, 0.06 µl of Taq DNA polymerase (5U/µl) (Platinum® Taq, Invitrogen), and 3.5 µl of DNA template (Hajibabaei et al., 2005). The PCR amplification was carried out using the following conditions: 1 min at 94°C, followed by 5 cycles of 94˚C for 40 sec, 45˚C for 40 sec and 72˚C for 1 min, followed by 35 cycles of 94˚C for 40 sec, 51˚C for 40 sec and 72˚C for 1 min, with a final extension of 72˚C for 5 minutes. PCR products were checked for quality and length using electrophoresis on ethidium bromide stained, 2% agarose gels (E-Gel 96 Invitrogen, Carlsbad, CA) and were bidirectionally sequenced by Eurofins Genomics (USA). The sequences were edited using Codon Code Aligner v. 8.0.1 (Codon Code Corporation) and uploaded to both the Barcode of Life Data System (BOLD, www.boldsystems.org) and GenBank. In BOLD, they were labeled as Parasitic Nematode of coffee berry borer (PNBC), and in GenBank, they were assigned the accession numbers MT520707 to MT520790.
Figure 1. Geographic location of coffee plantations sampled to determine the presence of coffee berry borers infected with the nematode M. hypothenemi.
Genetic diversity and population structure. The sequences were aligned using the ClustalW algorithm in the MEGA X v. 7.0.26 (Kumar et al., 2018) software, optimizing global similarity to ensure accuracy in phylogenetic inferences. Genetic diversity was estimated by calculating the number of segregation sites (S), number of haplotypes (h), haplotype diversity (Hd), nucleotide diversity (Π) and average number of nucleotide differences (K) using the software DnaSP v. 6.12.03 (Rozas & Librado, 2009). To test the hypothesis of neutral evolution and to understand potential demographic events, such as expansion trends, Tajima’s D (Tajima, 1989) and Fu and Li’s D and F indices were estimated (Fu & Li, 1993). Genetic differentiation among populations was analyzed using the pairwise fixation index (FST). An analysis of molecular variance (AMOVA) was also performed to assess the statistical significance of FST values (Excoffier & Lischer, 2010). Two separate AMOVA analyses were conducted to determine whether genetic differentiation between these lineages was statistically significant: i) considering the populations as a panmictic group and ii) the populations were grouped based on the 2 phylogenetic lineages identified in the maximum likelihood tree. Furthermore, the number of migrants per generation (Nm), an indirect measure of gene flow between populations, was estimated. All analyses were performed using Arlequin v. 3.5 (Excoffier & Lischer, 2010). The relationship between genetic differences (FST) and geographic distances among populations was analyzed using the Mantel test (Mantel, 1967). Two matrices were compared: genetic differences between population pairs and geographic distances (in kilometers). Geographic distances were calculated using QGIS v3.10.3 software (https://qgis.org). These distances were used to perform a Mantel test to assess the potential relationship between genetic differentiation and geographic separation in the context of an isolation-by-distance pattern. The statistical analysis was conducted in R (R Core Team, 2020) using the Vegan package (Oksanen et al., 2010), with 1,000 permutations to assess significance.
Table 1
Geographical locations where CBB samples infected with M. hypothenemi were collected in Mexico and Guatemala.
Municipality
Locality
Country
Code
Geographic coordinates
Latitude
Longitude
Tapachula
Santa Lucía
Mexico
SL
15°04’42.5’’
92°13’42.5
Finca Brasil
Mexico
BR
15°05’50.3’’
92°18’43.1’’
Cacahoatán
Salvador Urbina
Mexico
SU
15°02’26.9’’
92°12’03.9
La Alianza
Mexico
LA
15°02’40.8’’
92°11’03.2’’
El Zapote
Mexico
EZ
15°02’08.4’’
92°10’18.7’’
La Unidad
Mexico
LU
15°00’31.4’’
92°09’53.2
Dos de Mayo
Mexico
DM
15°02’55.0’’
92°09’14.5’’
Faja de Oro
Mexico
FO
15°02’56.4’’
92°09’14.4’’
Rosario Ixtal
Mexico
RI
15°00’37.2’’
92°09’28.0’’
San Antonio
Mexico
SA
15°00’24.2’’
92°09’04.7’’
Unión Juárez
Santo Domingo
Mexico
SD
15°01’37.5’’
92°06’06.9’’
San Rafael
Mexico
SR
15°02’29.8’’
92°07’10.3’’
San Jerónimo
Mexico
SJ
15°02’26.9’’
92°08’08.9’’
Monte Perla
Mexico
MP
15°02’42.1’’
92°05’17.8’’
Río Suchiate
Mexico
RS
15°03’26.1’’
92°04’13.2’’
Once de Abril
Mexico
OA
15°03’14.8’
92°08’30.9’’
Acacoyagua
Los Cacaos
Mexico
LC
15°23’23’’
92°39’13.0’’
San Pablo
BuenaVista
Guatemala
BV
14°57’53.1
91°59’48.7’’
The phylogeny of M. hypothenemi was inferred using RAxML v. 2.0. (Stamatakis, 2006). The General Time Reversible model with a proportion of invariant sites (GTR+I) was selected as the best approximation to the (TIM3+I) (-ln L 1,588.857; Delta-AIC = 0.0; AIC = 3,573.631) model obtained from the Akaike information criterion (AIC) (Akaike, 1974) analysis performed in JModelTest v. 0.1.1 (Posada, 2008). The analysis included 10,000 bootstrap replicates to ensure robust nodal support. Sequences of the nematode Howardula aoronymphium (Welch) (Tylenchida: Allantonematidae, GenBank: AY589466) was used as external group. The relationship between the identified haplotypes was analysed using a haplotype network generated in the PopART software v. 1.7 (Leigh & Bryant, 2015), applying the Median-Joining criterion.
Results
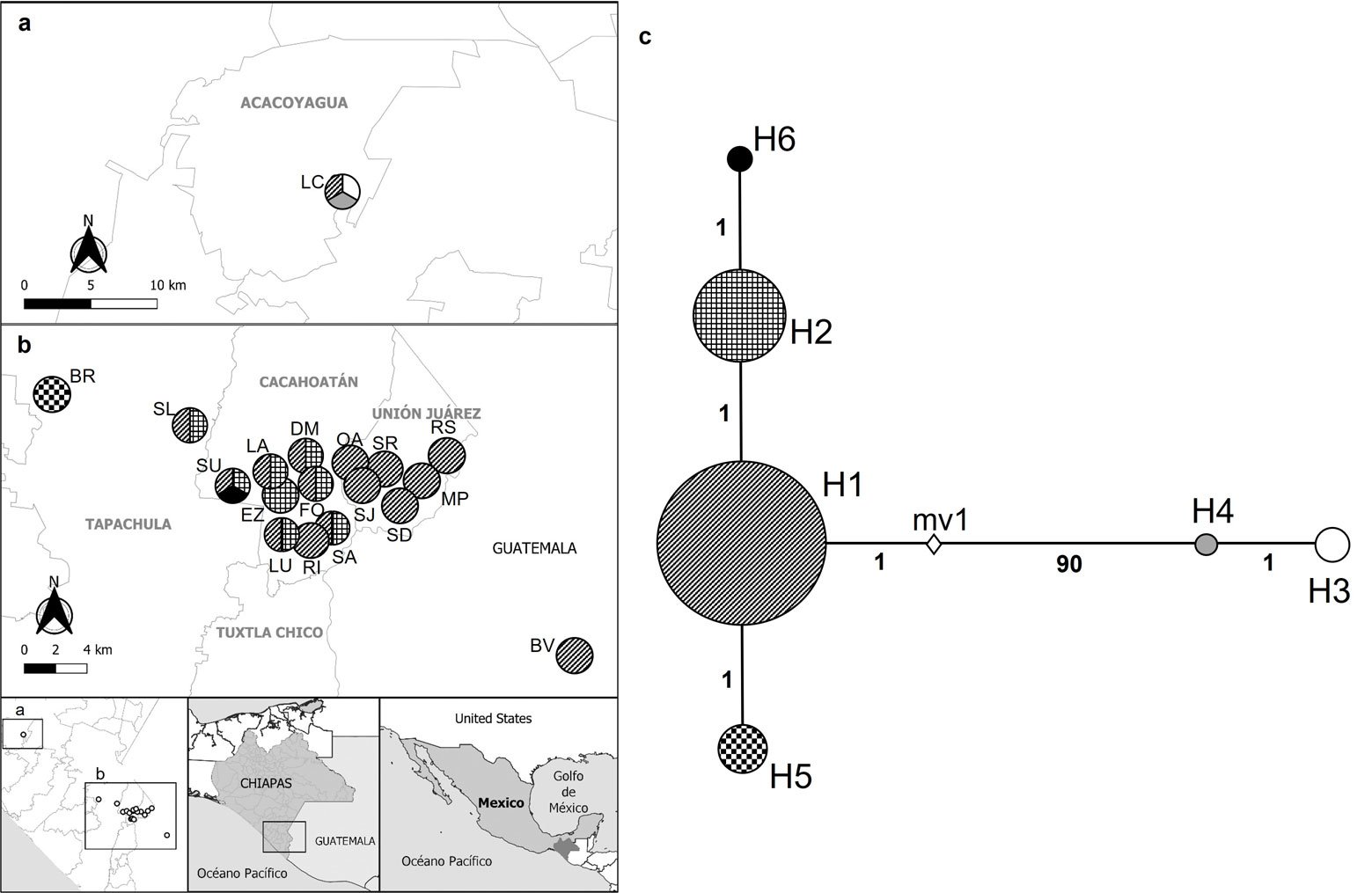
A total of 76 sequences of 560 bp from M. hypothenemi were analysed revealing 93 polymorphic sites (16.6%), whose A-T content (67.6%) was higher than the G-C content (32.4%). Global genetic diversity was moderate (Hd = 0.52 ± 0.06) (± SD) (Table 2). Ten populations presented zero diversity, while the SU and LC populations presented the highest genetic diversity from 18 populations (Table 2). The global neutrality tests were statistically significant according to Tajima values (D = -2.062, p < 0.05) and Fu and Li (D = 2.167, p < 0.02) (Table 2). Tajima and Fu and Li values were not significant inside of each population. A total of 6 haplotypes were identified, including one dominant haplotype (H1 = 50) and 4 unique ones (H3 = 2, H4 = 1, H5 = 5, and H6 = 1). The dominant haplotype was present in 16 from 18 populations sampled, while the second most common haplotype (H2 = 17) was recorded in 8 populations (Table 3).
High genetic differentiation was observed in M. hypothenemi, with a global value FST of 0.66 (p < 0.05) and a low number of migrants per generation (Nm = 0.50). The EZ, BR, and LC populations showed greater genetic differentiation than the rest of the populations, with an average distance between these and the other populations of 1.7-66.5 km, 9.5-50 km, and 50.5-87.5 km, respectively (Table 4). The smallest geographical distance was between the FO and DM populations (0.04 km), with no genetic differentiation between them. The AMOVA analysis conducted under panmictic indicate that genetic variation was greater between populations (66.61%) than within populations (33.39%). A second AMOVA, grouping populations according to the 2 phylogenetic lineages identified in the maximum likelihood tree, revealed a higher level of genetic differentiation between the lineages, with an FST of 0.99 (p < 0.01), indicating that the differentiation between the 2 lineages was statistically significant. In this second AMOVA, genetic variation was mainly distributed between lineages (99.42%) rather than within them (0.58%). The Mantel test revealed a significant correlation between the genetic pairwise distances and the geographic distances (km) among M. hypothenemi populations (r = 0.4728, p < 0.0001), consistent with a pattern of isolation by distance.
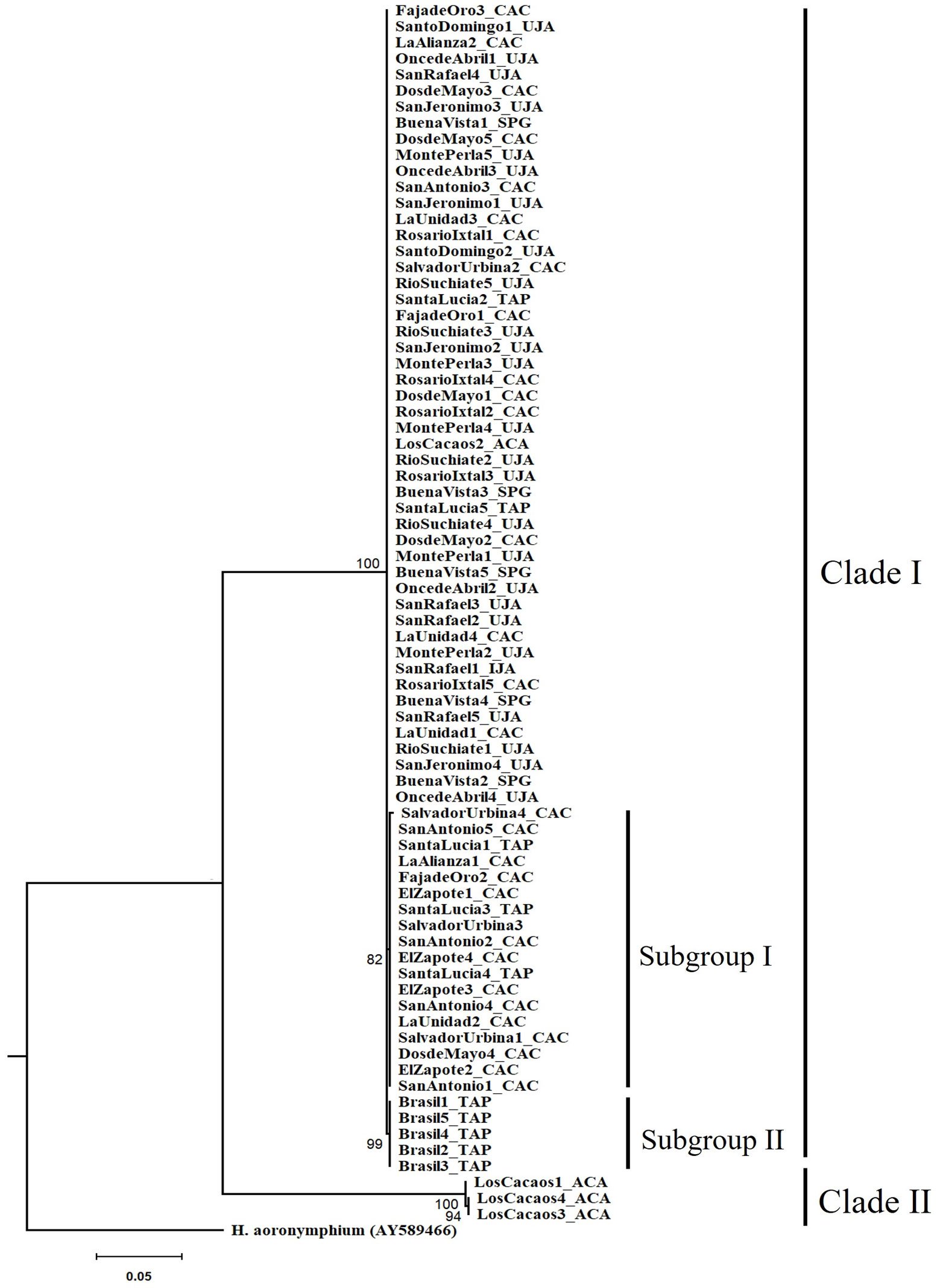
The tree topology revealed 2 independent clades with high support (> 90%). Clade I formed by 3 monophyletic groups, which corresponds to 96% of the specimens distributed in the municipalities of Tapachula, Cacahoatán, Unión Juárez, and San Pablo (Guatemala). Inside clade I, 2 subgroups were observed. The first subgroup had a bootstrap support value of 82%, while the second subgroup showed a higher support, with a bootstrap value of 99%. Clade II formed by 3 specimens belonging to the LC (Acacoyagua) population. A phylogenetic separation (16.5%) was observed between clade I and clade II specimens (Fig. 2).
The most frequent haplotype was H1, followed by H2. Both haplotypes were found in most of the studied sites, being particularly frequent in the municipalities of Tapachula, Cacahoatán, Unión Juárez, Acacoyagua, and San Pablo. On the other hand, H6 was exclusively detected in SU, in the municipality of Cacahoatán. H3 and H4 were restricted to LC, in Acacoyagua, while H5 was only found in BR, in Tapachula. The distribution of haplotypes by municipality and their frequencies are shown in Table 3. The inferred haplotype network identified H1 as the central, while the other haplotypes appeared as its derivatives. H4 diverged from H1 by 90 mutational steps, whereas H2, H5, and H6 diverged from H1 by only 1 mutational step (Fig. 3C).
Discussion
This study presents the first genetic description of M. hypothenemi populations, a nematode endoparasite of H. hampei, whose presence has only been naturally recorded in several coffee plantations in Mexico (Pérez et al., 2014). The genetic variability of M. hypothenemi suggests significant differentiation among its populations. Although not all populations exhibit the same degree of differentiation, the results support the hypothesis that this parasite is endemic to southeastern Mexico (Poinar et al., 2004). The genetic complexity of the populations of this parasite is similar to that observed in populations of endemic species (Blouin et al., 1999; Stock 2009).
This study also includes the first genetic sequences of M. hypothenemi in the BOLD System database. The COI gene has not yet been widely adopted as a tool for identifying parasitic nematode species, a high genetic structure in M. hypothenemi, an obligate parasite of H. hampei.
Table 2
Estimation of the genetic diversity in 18 populations of the nematode M. hypothenemi collected in the Soconusco region, Chiapas, Mexico.
Population
N
S
H
Su
Hd (± SD)
Π (± SD)
K
Tajima
Fu and Li
D
D
F
SL
5
1
2
0
0.600 (0.175)
0.0010 (0.0003)
0.600
1.224
1.224
1.157
BR
5
0
1
0
0
0
0
0
0
0
SU
4
2
3
2
0.833 (0.222)
0.0017 (0.0006)
1.000
-0.709
-0.709
-0.604
LA
2
1
2
1
1 (0.500)
0.0017 (0.0008)
1
–
–
–
EZ
4
0
1
0
0
0
0
0
0
0
LU
4
1
2
1
0.500 (0.265)
0.0008 (0.0004)
0.500
-0.612
-0.612
-0.478
DM
5
1
2
1
0.400 (0.237)
0.0007 (0.0004)
0.400
-0.816
-0.816
-0.771
FO
3
1
2
1
0.666 (0.314)
0.0011 (0.0000)
0.666
–
–
–
RI
5
0
1
0
0
0
0
0
0
0
SA
5
1
2
1
0.400 (0.237)
0.0007 (0.0000)
0.400
-0.816
-0.816
-0.771
SD
2
0
1
0
0
0
0
–
–
–
SR
5
0
1
0
0
0
0
0
0
0
SJ
4
0
1
0
0
0
0
0
0
0
MP
5
0
1
0
0
0
0
0
0
0
RS
5
0
1
0
0
0
0
0
0
0
OA
4
0
1
0
0
0
0
0
0
0
LC
4
92
3
91
0.833 (0.222)
0.0824 (0.0431)
46.166
-0.836
-0.836
-0.895
BV
5
0
1
0
0
0
0
0
0
0
Total
76
93
6
0
0.519 (0.056)
0.0134 (0.0067)
7.6291
-2.0622*
2.1674**
0.5539
N, Number of sequences; S, number of segregating sites; H, number of haplotypes; Su, number of unique sites; Hd, haplotype diversity; Π, nucleotide diversity; K, average number of nucleotide differences. ±SD: Standard deviation. The D Tajima index (Tajima, 1989), D and F of Fu and Li (Fu & Li, 1993). The hyphen represents an estimate not performed due to a limited number of sequences used. * p < 0.05 ** p < 0.02.
Identifying and detecting M. hypothenemi using classical taxonomy remains challenging due to its small size and the subtlety of its morphological characteristics, requiring a fast and reliable method such as DNA barcoding (Gonçalves et al., 2021). For example, 11 new marine nematode species from the Mexican Caribbean have been identified using this technique (Martínez et al., 2020). However, the COI gene has not yet been widely used to identify parasitic nematode species. To date, only 2 species have been reported naturally parasitizing adult coffee berry borers: Panagrolaimus sp. in India (Varaprasad, 1994) and M. hypothenemi in Mexico (Castillo et al., 2002). Nevertheless, the presence of M. hypothenemi has only been detected in this region of the world, and its identification through DNA barcoding could expand our understanding of this species’ geographic range.
Molecular analyses using the COI gene identified 6 haplotypes distributed among the sampled populations, with moderate global genetic diversity (Hd = 0.52) (Wang, 2020). This level of genetic diversity, although lower than that reported for other nematodes such as Heterorhabditis bacteriophora (Hd = 0.8; Saeb & David, 2014), is notable considering the limited geographic distribution of M. hypothenemi and the environmental pressures it faces in Soconusco coffee plantations (Simota et al., 2024). However, this genetic diversity was higher compared to other studies that used 40-120 specimens per population (Klimpel et al., 2007; Powers et al., 2018), possibly due to the number of samples used per population or the high evolutionary rate of mitochondrial DNA (Avise et al., 1987; Brown et al., 1979). Therefore, our sample size may have influenced the estimation of population parameters, and a larger number of specimens per population is recommended in future studies. The observed genetic diversity reflects the adaptive capacity of populations to environmental changes or selective pressure (Clarke, 1979), key aspects for the survival of M. hypothenemi in a fragmented environment. In this sense, understanding the genetic variation of this nematode is essential for designing management and conservation strategies aimed at preserving its role as a potential biological regulator of H. hampei.
Table 3
Frequency of female haplotypes for 18 populations of the nematode Metaparasitylenchus hypothenemi collected in the Soconusco region, Chiapas, Mexico and San Pablo, Guatemala.
Municipality
Population
*N
Haplotipe frequency
Tapachula
SL
5
H1(2), H2(3)
BR
5
H5(5)
Cacahoatán
SU
4
H1(1), H2(2), H6(1)
LA
2
H1(1), H2(1)
EZ
4
H2(4)
LU
4
H1(3), H2(1)
DM
5
H1(4), H2(1)
FO
3
H1(2), H2(1)
RI
5
H1(5)
SA
5
H1(1), H2(4)
Unión Juárez
SD
2
H1(2)
SR
5
H1(5)
SJ
4
H1(4)
MP
5
H1(5)
RS
5
H1(5)
OA
4
H1(4)
Acacoyagua
LC
4
H1(1), H3(2), H4(1)
San Pablo
BV
5
H1(5)
* Number of sequences used in the analysis; the number in parenthesis is the female specimens observed for each haplotype.
Selective neutrality analyses showed significant negative values at a global level, which commonly occur in populations undergoing demographic expansion (Schmidt, 2002), a process that likely began when this pest invaded Mexico in 1978 (Baker, 1984). Selective neutrality analyses revealed negative values for the DM, SA, and LC populations, while the SL population presented a positive value. Neutrality values within populations were not significant, indicating that the studied populations are not under the effects of natural selection (Schmidt, 2002). Therefore, it is feasible that the parasitism of H. hampei by M. hypothenemi represents a novel ecological interaction, as often occurs when a parasite is introduced into a new environment (Bush et al., 2001).
The determination of the origin of the parasite and the age of a parasite-host interaction is complex, due to the variety of mechanisms involved in the historical evolution of parasitism, including the possibility of an initial accidental association (Rico, 2011). The diversity of scolytines associated with coffee plantations in the Soconusco region is very high (Equihua, 1992), and H. hampei is endemic to Africa (Le Pelley, 1968), where populations can reach up to 11 million adults per hectare in mixed plantations of Robusta and Arabica coffee (Baker & Barrera, 1993). Under these conditions, it is likely that an endemic parasitic nematode from this region formed one or more new host-parasite associations with H. hampei (Bickford et al., 2007). Indeed, Poinar et al. (2004) hypothesized that M. hypothenemi first infected the insect in the New World.
Genetic differentiation analyses (FST) revealed high genetic differentiation among M. hypothenemi populations (FST = 0.66, p < 0.05), with gene flow that, in general, is very limited (Nm = 0.50), possibly due to the nematode’s dependence on host movement and the physical barriers imposed by a fragmented landscape (Ali et al., 2016; Fonseca & Netto, 2006; McGaughran et al., 2014). AMOVA confirmed that most genetic variation is found between populations (66.61%), indicating significant genetic structure and rejecting the hypothesis of panmixia. The observed level of genetic differentiation (FST = 0.66) suggests significant population structure, making panmixia unlikely. Additionally, the grouping of populations according to the 2 identified phylogenetic lineages revealed even greater genetic differentiation (FST = 0.99, p < 0.01). This high level of differentiation suggests strong genetic structuring, further reinforcing the rejection of panmixia. These findings highlight the need for further biological studies on M. hypothenemi to explore the existence of 2 lineages or even distinct species.
Table 4
Genetic (FST) and geographic distances (km, above the diagonal in bold) calculated for 18 populations of the nematode M. hypothenemi collected in the Soconusco region, Chiapas, Mexico and San Pablo, Guatemala.
LA
SD
EZ
LU
DM
FO
SR
SJ
MP
RI
RS
OA
SL
*BV
LC
BR
SU
SA
LA
0
9.36
1.70
4.54
3.38
3.39
7.20
5.40
10.68
4.82
12.75
4.82
6.19
22.67
64.85
15.38
1.92
5.59
SD
0
0
7.84
7.29
6.27
6.29
2.54
4.07
2.50
6.49
4.86
5.37
15.20
13.59
73.49
24.65
11.14
5.94
EZ
0.38
1
0
3.10
2.45
2.48
5.86
4.05
9.36
3.22
11.55
3.91
7.90
21.01
66.55
17.03
3.30
3.95
LU
0
0
0.66
0
4.59
4.64
6.2
4.81
9.42
0.79
11.82
5.65
10.51
19.32
68.99
19.12
5.39
1.51
DM
0
0
0.72
0
0
0.04
3.91
2.20
7.33
4.28
9.36
1.48
8.92
19.83
67.34
18.39
5.30
4.67
FO
0
0
0.57
0
0
0
3.92
2.22
7.32
4.32
9.35
1.46
8.91
19.84
67.31
18.38
5.32
4.71
SR
0.47
0
1
0.06
0
0.18
0
1.81
3.49
5.50
5.74
2.85
12.80
16.11
70.96
22.30
9.07
5.25
SJ
0.38
0
1
0
0
0.11
0
0
5.31
4.18
7.51
1.62
11.13
17.63
69.50
20.59
7.26
4.16
MP
0.47
0
1
0.06
0
0.18
0
0
0
8.64
2.41
6.05
16.04
13.54
73.70
25.57
12.56
8.21
RI
0.47
0
1
0.06
0
0.18
0
0
0
0
11.04
5.18
10.93
18.61
69.50
19.70
5.89
0.82
RS
0.47
0
1
0.06
0
0.18
0
0
0
0
0
7.97
17.76
13.15
74.73
27.26
14.66
10.62
OA
0.38
0
1
0
0
0.11
0
0
0
0
0
0
10.00
18.96
68.12
19.53
6.75
5.37
SL
0
0.28
0.19
0
0.10
0
0.5
0.45
0.5
0.5
0.5
0.45
0
28.72
58.65
9.52
5.18
11.72
*BV
0.47
0
1
0.06
0
0.18
0
0
0
0
0
0
0.5
0
87.08
38.05
24.25
17.81
LC
0.52
0.52
0.66
0.66
0.70
0.60
0.70
0.66
0.70
0.70
0.70
0.66
0.70
0.70
0
50.05
63.61
70.31
BR
0.85
1
1
0.82
0.83
0.82
1
1
1
1
1
1
0.81
1
0.70
0
13.85
20.53
SU
0
0.31
0
0.14
0.24
0
0.55
0.5
0.55
0.55
0.55
0.5
0
0.55
0.66
0.78
0
6.71
SA
0
0.62
0
0.31
0.41
0.15
0.75
0.72
0.75
0.75
0.75
0.72
0
0.75
0.70
0.88
0
0
* Site located in San Pablo, Guatemala.
Pairwise FST values showed that some populations, such as EZ, BR, and LC, exhibit high genetic differentiation compared to other populations, despite moderate geographic distances (average of 1.7-87.5 km). This suggests that these geographic distances might be sufficient to induce isolation by distance, although the presence of specific environmental characteristics or geographic barriers could also play a role. However, the presence of one common haplotype alongside 3 highly divergent ones could indicate the existence of cryptic lineages, migration from other populations, or the persistence of ancestral genetic diversity (Blouin et al., 1995; Chávez-González et al., 2022; Nieberding et al., 2005). Further studies with a larger sample size and broader geographic coverage are needed to clarify these patterns. On the other hand, the FO and DM populations, separated by only 0.04 km, showed no genetic differentiation, suggesting a high degree of gene flow between them, likely facilitated by their geographic proximity and a more homogeneous landscape.
Figure 2. Phylogenetic relationships of Metaparasitylenchus hypothenemi (76 sequences) using maximum likelihood. A sequence of the parasitic nematode Howardula aoronymphium (Tylenchidae: Allantonematidae) was used as outgroup (GenBank AY589466). The scale bar represents the number of expected nucleotide substitutions per site. Municipalities: TAP, Tapachula; UJA, Unión Juárez; CAC, Cacahoatán; ACA, Acacoyagua y SPG, San Pablo Guatemala. The numbers between the nodes in the tree correspond to bootstrap values.
The high genetic differentiation detected in this study could have important implications for the conservation of M. hypothenemi. On one hand, it suggests that populations are undergoing local adaptation processes, which could result in genetically distinct lineages that might be lost if their specific habitats are not protected. The significant correlation between genetic and geographic distances (r = 0.4728, p < 0.0001) obtained through the Mantel test supports the hypothesis of a pattern of isolation by distance. However, the moderate correlation value suggests that, in addition to geographic distance, other factors such as local environmental conditions, coffee plantation management practices, or landscape fragmentation may also contribute to the observed genetic differentiation (Diniz-Filho et al., 2013).
Figure 3. a-b, Distribution of the Metaparasitylenchus hypothenemi haplotypes in the 5 municipalities from the Soconusco region, Chiapas, Mexico and the municipality of San Pablo, Guatemala; c, haplotype network development using PopART v. 1.7. Wefts of the circles represent each haplotype. The size of the circles is proportional to the frequency of the haplotype inside the populations. The numbers on the lines connecting the haplotypes represent the mutational steps. The white rhombus (mv1) represents an extinct haplotype or an unsampled point. LC, Ejido Los Cacaos; LA, La Alianza; EZ, El Zapote; LU, La Unidad; DM, Dos de Mayo; FO, Faja de Oro; RI, Rosario Ixtal, SA, San Antonio: BV, Buena Vista; SU, Salvador Urbina; SL, Santa Lucía; BR, Brasil; SD, Santo Domingo; SR, San Rafael; SJ, San Jerónimo; MP, Monte Perla; RS, Río Suchiate; OA, Once de Abril.
The presence of 2 well-differentiated clades with high phylogenetic support (> 90%) suggests complex evolutionary processes. Clade I, which includes 96% of specimens distributed in the municipalities of Tapachula, Cacahoatán, Unión Juárez, and San Pablo, also showed internal substructures, indicating genetic differentiation within these populations. The formation of these subgroups could be interpreted as a possible response to geographic isolation or ecological barriers that have favored genetic divergence among populations (Wright, 1943). On the other hand, Clade II, composed only of 3 specimens from the LC population, showed a 16.4% phylogenetic separation from Clade I. This differentiation is considerably high and exceeds the intraspecific threshold generally accepted for genetic barcoding (1-2%; Lanteri, 2007). However, studies on nematodes have reported higher genetic divergences between species, as observed in Rhabdochona spp., where genetic differentiation range from 11% to 15% (Caspeta-Mandujano et al., 2021; Santacruz et al., 2020). This suggests that further comparisons with closely related taxa are needed to determine whether this divergence corresponds to an ongoing speciation process or represents intraspecific variation. The existence of 2 well-differentiated clades raises interesting questions about the evolutionary and ecological processes shaping M. hypothenemi populations. One possibility is that these differences reflect a conserved ancestral lineage in certain geographically isolated populations, such as LC. Another possibility is that the exclusive haplotypes are the result of recent local adaptation events or genetic drift in response to landscape fragmentation (Cheptou et al., 2017; Safran & Nosil, 2012). It is also plausible that these differences reflect the presence of a different species or a cryptic species with conserved morphology but genetic divergence. To confirm the occurrence of this genetic divergence process, further genetic studies with greater representation of the isolated population, as well as additional morphological studies, would be necessary. The use of nuclear markers would be particularly useful to determine whether the observed genetic differentiation is restricted to mitochondrial DNA or also occurs at the genome-wide level.
The haplotype analysis supports the hypothesis of genetic differentiation among populations. The high frequency and wide distribution of haplotype H1 found in most of the studied sites, suggest that it may represent the ancestral haplotype and the origin of other derived haplotypes. Nevertheless, the high divergence observed between haplogroups, particularly the separation of H4 by 90 mutational steps, indicates significant genetic structuring and highlights the complexity of inferring ancestral states (Avise, 2000). Therefore, it is not possible to confirm H1 as the ancestral haplotype without additional phylogenetic or coalescent analyses. In contrast, the exclusivity of H3 and H4 in the LC population, as well as H6 in SU and H5 in BR, reflects possible geographic isolation, local adaptation, and genetic drift processes (Forster, 2004).
These patterns are consistent with the theory that M. hypothenemi populations are influenced by geographic barriers that restrict gene flow and promote differentiation among localities. The search for this parasitism in the entire study region is required to determine the possibility of a greater number of M. hypothenemi haplotypes. The greater genetic difference of M. hypothenemi was observed in the population with greater geographical isolation (100 km), suggesting that more haplotypes could be located as more sites with the presence of the parasite are detected. Consequently, a complete genetic description of the M. hypothenemi populations require of the location of new sites with presence of this parasitism, whether in the study region or other coffee-growing regions of Chiapas and Central America.
This study added 6 new geographic records for M. hypothenemi, in addition to those previously known (Pérez et al., 2015). However, we believe that the identification of new sites might be influenced by the result of random interactions caused by the transport of infested fruits with the pest and the adaptation of the parasite to local climatic conditions.
This study provides the first evidence of the American origin of this nematode. Our study also opens possibilities for future research on the genetic and geographic diversity of this species, its relationship with environmental variables. These aspects are essential for designing conservation strategies for this species and its management for the control of H. hampei, an important global pest.
Acknowledgements
Special thanks to Guadalupe Eugenia Zarza Franco for her assistance in interpreting the results, to Fernando E. Vega for the comments on the manuscript, and to Jose Higinio López Urbina for making the maps.
References
Akaike, H. (1974). A new look at the statistical model identification. IEEE Transactions on Automatic Control, 19, 716–723. https://doi.org/10.1109/TAC.1974.1100705
Ali, N., Tavoillot, J., Chapuis, E., & Mateille, T. (2016). Trend to explain the distribution of root–knot nematodes Meloidogyne spp. associated with olive trees in Morocco. Agriculture, Ecosystems and Environment, 225, 22–32. https://doi.org/10.1016/j.agee.2016.03.042
Avise, J. C. (2000). Phylogeography: the history and formation of species(Vol. 447). Cambridge, Massachusetts, London England: Harvard University Press.
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neige, J. E., Reeb, C. A., & Saunders, N. C. (1987). Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics. Annual Review of Ecology and Systematics, 18, 489–522. https://doi.org/10.1146/annurev.es.18.110187.002421
Baker, P. S. (1984). Some aspects of the behaviour of the coffee berry borer in relation to its control in southern Mexico (Coleoptera: Scolytidae). Folia Entomológica Mexicana, 61, 9–24.
Baker, P. S., & Barrera, J. F. (1993). A field study of a population of coffee berry borer, Hypothenemus hampei (Coleoptera; Scolytidae), in Chiapas, Mexico. Tropical Agriculture, 70, 351–355.
Bickford, D., Lohman, D. J., Sodhi, N. S., Ng, P. K., Meier, R., Winker, K., Ingram, K. K., & Das, I. (2007). Cryptic species as a window on diversity and conservation. Trends in Ecology and Evolution, 22, 148–155. https://doi.org/10.1016/j.tree.2006.11.004
Blouin, M. S., Liu, J., & Berry, R. E. (1999). Life cycle variation and the genetic structure of nematode populations. Heredity, 83, 253–259. https://doi.org/10.1038/sj.hdy.6885420
Blouin, M. S., Yowell, C. A., Courtney, C. H., & Dame, J. B. (1995). Host movement and the genetic structure of populations of parasitic nematodes. Genetics, 141, 1007–1014. https://doi.org/10.1093/genetics/141.3.1007
Brown, W. M., George, M., & Wilson, A.C. (1979). Rapid evolution of animal mitochondrial DNA. Proceedings of the National Academy of Sciences, 76, 1967–1971. https://doi.org/10.1073/pnas.76.4.1967
Bush, A. O., Fernández, J. C., Esch, G. W., Seed, J. R., & Ndez, J. C. F. (2001). Parasitism: the diversity and ecology of animal parasites. Cambridge, UK: Cambridge University Press.
Castillo, A., Infante, F., Barrera, J. F., Carta, L., & Vega, F. E. (2002). First field report of a nematode (Tylenchida: Sphaerularioidea) attacking the coffee berry borer, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) in the Americas. Journal of Invertebrate Pathology, 79, 99–202. https://doi.org/10.1016/S00222011(02)00024-1
Castillo, A., Martínez, F., Gómez, J., Cisneros, J., & Vega, F. E. (2019). Sterility of the coffee berry borer, Hypothenemus hampei (Coleoptera: Curculionidae), caused by the nematode Metaparasitylenchus hypothenemi (Tylenchidae: Allantonematidae). Biocontrol Science and Technology, 29, 786–795. https://doi.org/10.1080/09583157.2019.1597336
Caspeta-Mandujano, J. M., Salinas-Ocampo, J. C., Suárez-Rodríguez, R., Martínez-Ramírez, C., & Matamoros, W. A. (2021). Morphological and molecular evidence for a new rhabdochonid species, Rhabdochona (Nematoda: Rhabdochonidae), parasitizing Eugerres mexicanus (Perciformes: Gerreidae), from the Lacantún River in the Biosphere Reserve of Montes Azules, Chiapas, Mexico. Revista Mexicana de Biodiversidad, 92, 01–11. https://doi.org/10.22201/ib.20078706e.2021.92.3266
Cháves-González, L. E., Morales-Calvo, F., Mora, J., Solano-Barquero, A., Verocai, G. G., & Rojas, A. (2022). What lies behind the curtain: Cryptic diversity in helminth parasites of human and veterinary importance. Current Research in Parasitology & Vector-Borne Diseases, 2, 100094. https://doi.org/10.1016/j.crpvbd.2022.100094
Cheptou, P. O., Hargreaves, A. L., Bonte, D., & Jacquemyn, H. (2017). Adaptation to fragmentation: evolutionary dynamics driven by human influences. Philosophical Transactions of the Royal Society B: Biological Sciences, 372, 20160037. https://doi.org/10.1098/rstb.2016.0037
Clarke, B. C. (1979). The evolution of genetic diversity. Proceedings of the Royal Society of London. Series B. Biological Sciences, 205, 453–474. https://doi.org/10.1098/rspb.1979.0079
Diniz-Filho, J. A. F., Soares, T. N., Lima, J. S., Dobrovolski, R., Landeiro, V. L., Telles, M. P. D. C., & Bini, L. M. (2013). Mantel test in population genetics. Genetics and Molecular Biology, 36, 475–485. https://doi.org/10.1590/S141547572013000400002
Equihua, M. A. (1992). Coleópteros Scolytidae atraídos a trampas NTP–80 en el Soconusco, Chiapas, México. Folia Entomológica Mexicana, 84, 55–66.
Excoffier, L., & Lischer, H. E. L. (2010). Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources, 10, 564–567. https://doi.org/10.1111/j. 1755-0998.2010.02847.x
Fitza, K. N., Garnas, J. R., Lombardero, M. J., Ayres, M. P., Krivak–Tetley, F. E., Ahumada, R., & Slippers, B. (2019). The global diversity of Deladenus siricidicola in native and non–native populations. Biological Control, 132, 57–65. https://doi.org/10.1016/j.biocontrol.2019.01.014
Fonseca, G., & Netto, S. A. (2006). Shallow sublittoral benthic communities of the Laguna estuarine system, Brazilian Journal of Oceanography, 54, 41–54.
Forster, P. (2004). Ice ages and the mitochondrial DNA chronology of human dispersals: a review. Philosophical Transactions of the Royal Society, 359, 255–264. https://doi.org/10.1098/rstb.2003.1394
Fu, Y. X., & Li, W. H. (1993). Statistical tests of neutrality of mutations. Genetics, 133, 693–709. https://doi.org/10.1093/genetics/133.3.693
Gonçalves, L. T., Bianchi, F. M., Deprá, M., & Calegaro-Marques, C. (2021). Barcoding a can of worms: testing cox1 performance as a DNA barcode of Nematoda. Genome, 64, 705–717. https://doi.org/10.1139/gen-2020-0140
Hajibabaei, M., DeWaard, J. R., Ivanova, N. V., Ratnasingham, S., Dooh, R. T., Kirk, S. L. et al. (2005). Critical factors for assembling a high volume of DNA barcodes. Philosophical Transactions of the Royal Society B: Biological Sciences, 360, 1959–1967. https://doi.org/10.1098/rstb.2005.1727
Hartshorn, J. A., Fisher, J. R., Riggins, J. J., & Stephen, F. M. (2017). Molecular identification of Deladenus proximus Bedding, 1974 (Tylenchida: Neotylenchidae), a parasite of Sirex nigricornis (Hymenoptera: Siricidae). Nematology, 19, 15–20. https://doi.org/10.1163/15685411-00003029
Hebert, P. D., Cywinska, A., Ball, S. L., & Dewaard, J. R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270, 313–321. https://doi.org/10.1098/rspb.2002.2218
Ivanova, N. V., Dewaard, J. R., & Hebert, P. D. (2006). An inexpensive, automation-friendly protocol for recovering high-quality DNA. Molecular Ecology Notes, 6, 998–1002. https://doi.org/10.1111/j.1471-8286.2006.01428.x
Klimpel, S., Kleinertz, S., Hanel, R., & Rückert, S. (2007). Genetic variability in Hysterothylacium aduncum, a raphidascarid nematode isolated from sprat (Sprattus sprattus) of different geographical areas of the northeastern Atlantic. Jounal Parasitology Research, 101, 1425–1430. https://doi.org/10.1007/s00436-007-06620
Kumar S., Stecher G., Li M., Knyaz C., & Tamura K. (2018). MEGA X: molecular Evolutionary Genetics Analysis across computing platforms. Molecular Biology Evolution, 35, 1547–1549. https://doi.org/10.1093/molbev/msy096
Lanteri, A. A. (2007). Código de barras del ADN y sus posibles aplicaciones en el campo de la entomología. Revista de la Sociedad Entomológica Argentina, 66, 15–25.
Le Pelley, R. H. (1968). Pests of coffee. London: Longmans, Green and Co.
Leigh, J. W., Bryant, D., & Nakagawa, S. (2015). POPART: full–feature software for haplotype network construction. Methods in Ecology & Evolution, 6, 1110–1116. https://doi.org/10.1111/2041-210X.12410
Mantel, N. (1967). The detection of disease clustering and a generalized regression approach. Cancer Research, 27, 209–220.
Marsjan, P. A., & Oldenbroek, J. K. (2007). Molecular markers, a tool for exploring genetic diversity. In B. Rischkowsky, & D.Pilling (Eds.), The state of the world’s animal genetic resources for food and agriculture. Rome: Food and Agriculture Organization.
Martínez, A. A., Jesús, N. D., & Leasi, F. (2020). DNA Barcoding for delimitation of putative Mexican marine nematodes species. Diversity, 12, 107. https://doi.org/10.3390/d12030107
McGaughran, A., Morgan, K., & Sommer, R. J. (2014). Environmental variables explain genetic structure in a beetle–associated nematode. Plos One, 9, 87317. https://doi.org/10.1371/journal.pone.0087317
Nieberding, C., Libois, R., Douady, C. J., Morand, S., & Michaux, J. R. (2005). Phylogeography of a nematode (Heligmosomoides polygyrus) in the western Palearctic region: persistence of northern cryptic populations during ice ages? Molecular Ecology, 14, 765–779. https://doi.org/10.1111/j.1365294X.2005.02440.x
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., O’hara, R. B., Simpson, G. L., & Wagner, H. (2010). Vegan: community ecology package. R package version 1.17–4. http://CRAN.R–project.org/package=vegan
Pérez, J., Infante, F., Poinar, G. Jr., Castillo, A., & Vega, F. E. (2015). Natural parasitism of Metaparasitylenchus hypothenemi (Tylenchida: Allantonematidae) on the coffee berry borer in Chiapas, Mexico. Biocontrol Science and Technology, 25, 608–612. https://doi.org/10.1080/09583157.2014.991910
Poinar, G. Jr., Vega, F. E., Castillo, A., Chavéz, I. E., & Infante, F. (2004). Metaparasitylenchus hypothenemi Sp. (Nematoda: Allantonematidae), a parasite of the co-ee berry borer, Hypothenemus hampei (Curculionidae: Scolytinae). Journal of Parasitology, 90, 1106–1110. https://doi.org/10.1645/GE-3369
Posada, D. (2008). Model Test: phylogenetic model averaging. Molecular Biology and Evolution, 25, 1253–1256. https://doi.org/10.1093/molbev/msn083
Powers, T., Harris, T., Higgins, R., Mullin, P., & Powers, K. (2018). Discovery and identification of Meloidogyne species using COI DNA barcoding. Journal of Nematology, 50, 399–412. https://doi.org/10.21307/jofnem-2018-029
Prosser, S., Martínez, A. A., & Elías, G. M. (2013). A new set of primers for COI amplification from freshwater microcrustaceans. Molecular Ecology Resources, 13, 151–1155. https://doi.org/10.1111/1755-0998.12132
Rico, G. (2011). The evolution of host-parasite interactions: coevolution, sexual selection and other suggested theories. Revista UDCA Actualidad and Divulgación Científica, 14, 119–130.
R Core Team (2020). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R–project.org/
Rozas, J., & Librado, P. (2009). DnaSP v. 5. A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25, 1451–1452. https://doi.org/10.1093/bioinformatics/btp187
Saeb, A. T., & David, S. K. (2014). Comparative estimation of genetic diversity in population studies using molecular sampling and traditional sampling methods. Biomedical Informatics, 10, 347–352. https://doi.org/10.6026/9732063 0010347
Safran, R. J., & Nosil, P. (2012). Speciation: the origin of new species. Nature Education Knowledge, 3, 17.
Santacruz, A., Ornelas-García, C. P., & De León, G. P. P. (2020). Diversity of Rhabdochona mexicana (Nematoda: Rhabdochonidae), a parasite of Astyanax spp. (Characidae) in Mexico and Guatemala, using mitochondrial and nuclear genes, with the description of a new species. Journal of Helminthology, 94, 1–11. https://doi.org/10.1017/S0022149X19000014
Simota-Ruiz, M., Castillo, A., Cisneros-Hernández, J., & Carmona-Castro, O. (2024). Predicted distribution of Metaparasitylenchus hypothenemi (Tylenchida: Allantonematidae), parasite of the coffee berry borer. Journal of Nematology, 56, 1–10. https://doi.org/10.2478/jofnem-2024-0031
Schmidt, D., & Pool, J. (2002). The effect of population history on the distribution of the Tajima’s D statistic. Population English Edition, 1, 1–8.
Stamatakis A. (2006). RAxML-VI-HPC: maximum likelihood–based phylogenetic analyses with thousands of taxa and mixed models, Bioinformatics, 22, 2688–2690. https://doi.org/10.1093/bioinformatics/btl446
Stewart, J. B., & Larsson, N. G. (2014). Keeping mtDNA in shape between generations. Plos Genetics, 10, 1–7. https://doi.org/10.1371/journal.pgen.1004670
Stock, S. P. (2009). Molecular approaches and the taxonomy of insect–parasitic and pathogenic nematodes. In S. P. Stock, J. Vandenberg, I. Glazer, & N. Boemare (Eds.), Insect pathogens: molecular approaches and techniques (pp. 71–100). Wallingford UK: CABI. https://doi.org/10.1079/9781845934781.0071
Tajima, F. (1989). Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics, 123, 585–595. https://doi.org/10.1093/genetics/123.3.585
Varaprasad, K. S., Balasubramanian, S., Diwakar, B. J., & Ramarao, C. V. (1994). First report of an entomogenous nematode, Panagrolaimus sp. from coffee berry borer, Hypothenemus hampei (Ferrari) from Karnataka, India. Plant Protection Bulletin (Faridabad), 46, 34–35.
Wang, S. Q. (2020). Genetic diversity and population structure of the endangered species Paeonia decomposita endemic to China and implications for its conservation. BMC Plant Biology, 20, 1–14. https://doi.org/10.1186/s12870-020-02682-z
Wright, S. (1943). Isolation by distance. Genetics, 28, 114–138. https://doi.org/10.1093/genetics/28.2.114
César Marco Aurelio Jurado-Vargas a, *, José Cruz-de León b, José Tulio Mendez-Montiel c
a Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Biología, Laboratorio de Investigación en Invertebrados, Ciudad Universitaria, Av. Fco. J. Mujica s/n, 58030 Morelia, Michoacán, México
b Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Ingeniería en Tecnología de la Madera, Laboratorio de Conservación y Preservación de la Madera, Ciudad Universitaria, Av. Fco. J. Mujica s/n, 58030 Morelia, Michoacán, México
c Universidad Autónoma Chapingo, Dirección de Ciencias Forestales, Carretera Federal México-Texcoco Km 38.5, 56230 Texcoco, Estado de México, México
*Autor para correspondencia: cjurado@umich.mx (C.M.A. Jurado-Vargas)
A partir de un análisis morfológico y taxonómico de adultos del género Calymmaderus Solier, 1849 (Ptinidae: Dorcatominae), se describen y proponen 2 nuevas especies para el Neotrópico en México: C. robustus sp. nov. y C. semioblongus sp. nov. Ellas se distinguen de las 7 especies reportadas y reconocidas actualmente para México con base en su coloración, pubescencia y estrías elitrales laterales. Se describe la estructura genital de machos.
Two new Neotropical species of the genus Calymmaderus (Coleoptera: Ptinidae), associated to structural wood in Taretan, Michoacán, Mexico
Abstract
Through morphological and taxonomic analysis of adults of the genus Calymmaderus Solier, 1849 (Ptinidae: Dorcatominae), 2 new Neotropical species in Mexico are described and proposed: C. robustus sp. nov. and C. semioblongus sp. nov. They are distinguished from the 7 species currently reported and recognized for Mexico based on their coloration, pubescence and lateral elitral striae. The genital structure of males is described.
El género Calymmaderus incluye especies de escarabajos xilófagos de distribución mundial, para el cual se reconocen 89 especies americanas (Blackwelder, 1945; Lüer y Honour, 2017; Toskina, 2000; White, 1974, 1982, 1983, 1984), de las que 7 especies se encuentran en México: C. dejeani (Pic, 1905), C. donckieri (Pic, 1904), C. oblongus (Gorham, 1883), C. pupatus (Gorham, 1883), C. semirufus (Champion, 1913), C. sharpi (Gorham, 1886) y C. subvestitus (Champion, 1913) (White, 1983; Zaragoza et al., 2016). El estudio de las especies de este género es complicado debido a la falta de revisiones taxonómicas, lo limitado de sus descripciones diagnósticas y la carencia, en la mayoría de los casos de ilustraciones de los órganos genitales, estudio que le da mayor robustez a la determinación de especies (Bercedo et al., 2008; Lüer y Honour, 2017; Viñolas, 2018). Los miembros de Calymmaderus se distinguen de las demás especies de Ptinidae por su cuerpo alargado y oblongo, antenas con los 2 últimos segmentos de la clava estrechamente unidos, coxas protorácicas expuestas ventralmente, un lóbulo metasternal bifurcado que aloja al último segmento antenal cuando el cuerpo está en reposo y el número de suturas elitrales laterales (Arango, 2012; Español, 1992; White, 1971). Las especies de este género son de importancia económica al dañar la madera estructural de edificios históricos y bienes culturales. La especie más conocida en México es C. oblongus (Gorham, 1883), a la que varios autores señalan como asociada al deterioro de la madera (Bercedo et al., 2008; Cibrián et al., 1995; Jurado-Vargas, 2020; Jurado-Vargas y Cruz, 2010, 2020; Jurado-Vargas et al., 2003; Pichardo et al., 2017; White, 1974).
El presente estudio tiene como objetivo describir 2 especies nuevas de Calymmaderus, que fueron encontradas en la localidad de Taretan, Michoacán, México, ubicada en la Zona de Transición Mexicana, que es un área de alta diversidad y endemismos de artrópodos y presenta la mayor mezcla biótica entre elementos neárticos y neotropicales (Halffter, 2017). Ambas especies representan taxones morfológicamente diferentes a las especies descritas en la literatura conocida para este grupo. White fue el último investigador que publicó datos sobre especies de Calymmaderus y otros géneros de escarabajos de la familia Ptinidae americanas entre 1981 y 1984, dejando pendientes algunos ejemplares con estatus incierto aún por resolver. En este trabajo presentamos la descripción y propuesta de las 2 nuevas especies de Calymmaderus referidas.
Materiales y métodos
El área de estudio corresponde a la localidad de Taretan, Michoacán, México (1,140 m snm, 19°20’3.32” N, 101°55’3.02” O) (fig. 1), que presenta un clima cálido A (w), con precipitación media anual de 1,240 mm y temperatura media anual 25° C (INEGI, 2010).
Se recolectaron 463 ejemplares del género Calymmaderus de la techumbre de madera del templo de San Ildefonso por medio de 2 trampas de luz blanca, durante el periodo de emergencia de los adultos en los meses de junio-agosto del 2017 y 2018. Los ejemplares se fijaron en una solución de alcohol al 80%. La determinación de los ejemplares se realizó usando descripciones y claves de identificación para el género y especies de Calymmaderus (Arango, 2012; White,1971, 1974, 1982, 1983, 1984). En el proceso, la preparación de los órganos se realizó mediante la técnica de montaje permanente en laminillas con resina sintética (Gaviño et al., 2004).
La descripción de los caracteres morfológicos de los ejemplares adultos se realizó mediante un microscopio estereoscópico Nikon a 45 aumentos. Se obtuvieron imágenes de los ejemplares en posición dorsal, lateral y ventral, además de metasterno, abdomen y antenas, mediante un microscopio estereoscópico Carl Zeiss (modelo Axio Zoom V16). Las imágenes de las preparaciones de la estructura genital se lograron a través de un microscopio compuesto Marca Leica a 100 aumentos y una cámara Panasonic adaptada a la distancia focal del ocular. Para obtener detalles adicionales de la morfología externa de los ejemplares, se tomaron imágenes por medio del microscopio electrónico de barrido modelo JSM-6400.
Se determinaron las medidas de largo y ancho de 30 ejemplares de cada una de las especies que se describen con un vernier digital, con las que se realizó un análisis estadístico descriptivo básico de los parámetros utilizados para su descripción. La fenología de emergencia de adultos se determinó mediante gráficos de los registros mensuales de captura de los ejemplares adultos en los periodos de trabajo. Los holotipos y paratipos de las 2 especies se depositaron en la Colección de Invertebrados (Insectos Xilófagos) de la Facultad de Biología de la Universidad Michoacana (CIFBUM-XIL).
Figura 1. Ubicación de Taretan, localidad tipo de las 2 nuevas especies de Calymmaderus (INEGI, 2016).
Diagnosis. Cuerpo robusto en vista lateral, 1.9 veces más largo que ancho, color negro uniforme mate, pubescencia blanquecina decumbente y abundante, separada menos de su longitud, élitros paralelos después de la base hasta 3/4 partes; antenas desde la base de café rojizo a café negruzco en los últimos segmentos; metasterno con menos pubescencia que resto del cuerpo. Lóbulo mestasternal con muesca profunda en forma característica de U.
Holotipo ♂, largo 3.8 mm, ancho 2.1 mm. Cuerpo robusto en vista lateral, 1.9 veces más largo que ancho (figs. 2, 3), color negro mate, excepto en palpos maxilares y labiales, que presentan el segmento basal café oscuro y los otros segmentos café rojizo. Tarsos de color café rojizo. Pubescencia decumbente blanquecina, abundante, uniforme en todo el cuerpo. Puntuaciones elitrales de la superficie ampliamente distribuidas, no alineadas e intercaladas con otras puntuaciones diminutas en todo el cuerpo y separadas 1.0 veces su diámetro. Cabeza: ojos grandes y abultados, distancia interocular 1.1 veces el diámetro vertical de un ojo. Clava antenal ligeramente mayor (1.1 veces) que el resto de los segmentos; 9º segmento de la clava antenal ligeramente más largo que los segmentos 10º y 11º (fig. 6). Pronoto: acampanado, 1.9 veces más ancho que largo; borde anterior curvo y márgenes laterales redondeados. Superficie densamente punteada y pilosa, con las puntuaciones laterales equidistantes y separadas en general el equivalente a su diámetro; en el disco las puntuaciones son más separadas (1-2 veces su diámetro). Élitro: puntuaciones del disco y laterales separadas de 1 a 2 veces su diámetro (algunas veces un poco más separadas y alargadas); puntuaciones más cercanas a la base elitral más aglutinadas y puntuaciones laterales hacia la parte distal del élitro más alargadas; ranura lateral visible después de la base del tercer esternito hasta el ápice (fig. 4), abarcando menos de la mitad del élitro. Metasterno: convexo, más ancho que largo, carinas bien delimitadas; puntuaciones cercanas al proceso metasternal más pequeñas, separadas 1.0 veces su diámetro; puntuaciones se agrandan más hacia la parte media y a los lados, separadas de 1.0 a 1.5 veces su diámetro; en la parte media y hasta el borde posterior son más pequeñas y menos densas; surco longitudinal corto y no muy marcado (fig. 5). Abdomen: primero, segundo y quinto esternitos más o menos del mismo largo; segundo con borde posterior ligeramente curvo en centro; tercero un poco más corto que anteriores, y cuarto más corto que el resto (fig. 3). Edeago: longitud: 1.1 mm; anchura 0.48 mm. Parámeros bifurcados apicalmente, con borde apical redondeado; piezas accesorias de los parámeros 4.2 veces más largos que su ancho, con pilosidad larga desde la mitad hasta su extremo. Endófalo: ápice más esbelto que la base, terminando en punta con un diente curvo en forma de gancho, saco interno provisto de por lo menos doce espinas transversales a lo largo; las 2 más cercanas a la base 2 veces más largas que las demás y de posición paralela; 2 espinas al centro más pequeñas y entrecruzadas con las espinas basales más largas (fig. 14).
Paratipo ♀. Aspecto general del cuerpo similar al macho. Largo 4.1 ancho 2.3 mm, ojos evidentemente más grandes y abultados que el macho, distancia interocular 1.3 veces el diámetro vertical de un ojo. Tercera y cuarta sutura de los esternitos curvada a los lados.
Resumen taxonómico
Etimología. El epíteto de la especie robustus deriva de la forma robusta del tórax en vista lateral (mucho más alta que el abdomen), que caracteriza y diferencia a los ejemplares de esta especie en comparación con las especies conocidas.
Material examinado. Holotipo ♂: CIFBUM-XIL Núm. 224. México: Michoacán. Taretan. 19.VII.2017. C. Jurado Col. Trampa de luz blanca, madera estructural de pino. 19°20’3.32” N. 101°55’3.02” O. 1,140 m snm. Paratipos: 28; 13 ♂, 17 ♀. CIFBUM-XIL Núms. 226-252, con los mismos datos del holotipo.
Comentarios taxonómicos
Variabilidad: largo 3.1-4.2 mm (media 3.72) ancho 1.7-2.3 mm (media 2.08). Macho más pequeño que la hembra (de 3.1 a 3.7 mm; tabla 1). Ojos más pequeños, distancia interocular de 1.0 a 1.1 veces del diámetro vertical del ojo; último esternito normal, con sutura recta. Hembra: 3.8 a 4.2 mm, ojos más grandes y abultados que el macho, distancia interocular de 1.2 a 1.3 veces el diámetro vertical de un ojo; último esternito con sutura ligeramente curvada en el centro. Especie con morfología poco variable; color general negruzco mate, pubescencia blanquecina uniforme, algunas veces con pubescencia menos abundante en el metasterno.
Figuras 2-6. Calymmaderus robustus sp. nov. 2) Vista dorsal; 3) vista ventral; 4) vista lateral; 5) metasterno; 6) antena.
Especie de hábitos nocturnos asociada con madera estructural de Pinus sp.; los ejemplares se capturaron en el interior de la techumbre de madera del templo de “San Ildefonso” Taretan, con trampa de luz. La emergencia de adultos inicia la última semana de junio, con máximo a mediados de julio y disminuye la última semana de julio; la emergencia es rara en agosto. Cohabita con otras especies de Ptinidae y termes, reinfestando durante muchos años la madera del mismo sitio (fig. 7).
Figura 7. Emergencia adultos de Calymmaderus robustus sp. nov., en 2 ciclos anuales.
Tabla 1
Medidas y variación de largo y ancho de machos y hembras adultos de Calymmaderus robustus sp. nov.
Medidas
Promedio (mm)
Error estándar
Rango (mm)
N (hembras) = 13 N (machos) = 17
Machos
Hembras
Machos
Hembras
Machos
Hembras
Longitud
3.438
3.947
0.055
0.032
3.1 – 3.7
3.8 – 4.2
Ancho
1.931
2.194
0.036
0.020
1.2 – 2.1
2.1 – 2.3
De acuerdo con la clave de identificación usada para especies de Calymmaderus (White, 1983), los ejemplares son definitivamente diferentes a las descripciones de las especies mexicanas descritas, considerando la variación en el conjunto de los siguientes caracteres. Todas las especies citadas para México presentan cuerpo oblongo o semiesférico (C. sharpi) en vista lateral; en cuanto al color, los ejemplares descritos de todas las especies van de café rojizo o café oscuro a casi negro, mientras que la coloración del integumento de la especie que se describe es negro mate uniforme. En cuanto a la pubescencia, a diferencia de las especies conocidas en las que tiene tonalidades amarillentas a grisáceas, la especie propuesta presenta pubescencia blanquecina, que contrasta fuertemente con el color del cuerpo. La mayoría de las poblaciones de las especies comparten un tamaño de longitud y ancho similares, excepto C. donckieri y C. subvestitus, que son notablemente más pequeñas. La carencia de información acerca de la armadura genital de las especies no permite hacer comparaciones al respecto. Sin embargo, se presenta la comparación de la armadura genital del macho de las 2 especies propuestas en este documento y C. oblongus, especie mejor conocida de amplia distribución en México (figs. 14-16), lo que respalda la diferenciación entre ellas.
Diagnosis. Forma oblonga, esbelto en vista dorsal, 2.1 veces más largo que ancho, élitros paralelos después de la base hasta 3/4 partes de su extensión; superficie del cuerpo de coloración homogénea de café oscuro a casi negro; pubescencia amarillenta densa y decumbente, separada menos del largo de su longitud; puntuaciones laterales de élitros más agrandadas en la base que en el resto del élitro.
Holotipo ♂, largo 3.8 mm, ancho 1.8 mm. Coloración de café oscuro a casi negro; superficie brillante; antenas y tarsos con la misma coloración del cuerpo; palpos labiales y palpos maxilares café rojizo. Cuerpo oblongo y alargado, 2.1 veces más largo que ancho (figs. 8, 9). Vestidura: pubescencia amarillenta, densa, decumbente y abundante en todo el cuerpo, puntuaciones uniformes en todo el cuerpo, excepto las puntuaciones laterales del élitro que se presentan de forma aglutinada e irregular (fig. 10). Cabeza: ojos pequeños separados 1.4 veces el diámetro vertical de un ojo. Clava antenal 1.3 veces más larga que el resto de los segmentos, noveno segmento de la clava antenal del mismo largo que el décimo y undécimo (fig. 12). Pronoto: acampanado 1.6 veces más largo que ancho, convexo dorsalmente; margen lateralmente redondeado, con un reborde lateral y borde anterior regularmente curvo, no proyectado; disco no prominente; pubescencia uniforme, superficie densamente punteada. Élitro: puntuaciones elitrales irregulares, no alineadas en hileras, separadas de 1 a 2 veces su diámetro, en el disco elitral también a la misma distancia; puntuaciones laterales de la base un poco más grandes y algunas están separadas el equivalente a su diámetro; en el tercio posterior del élitro las puntuaciones son más pequeñas; puntuaciones basales laterales aglutinadas formando una estría a la altura del segundo esternito, que continua hasta el ápice ya bien marcada como ranura basal; por encima otra línea no muy marcada forma otra estría también a nivel del segundo esternito, la ranura ocupa ligeramente más de la mitad del élitro hasta el ápice. Metasterno: ancho del lóbulo metasternal, escasamente mayor que su longitud, muesca profunda, ápice del lóbulo muy arqueado en forma de V; puntuaciones más grandes cerca del lóbulo y a lo largo del borde anterior del metasterno, separadas el equivalente a su diámetro; a los lados las puntuaciones son más espaciadas de 1.0 a 1.5 veces su diámetro, algunas separadas hasta 2 veces su diámetro; del centro hasta el borde posterior, las puntuaciones son más pequeñas y escasas, separadas de 1.0 a 2.0 veces su diámetro; surco longitudinal ligeramente marcado y extendido hasta la mitad del metasterno (fig. 11). Abdomen: segundo ventrito más largo que el resto; el primero y tercero de largo similar, el cuarto es el más estrecho, el quinto presenta una concavidad paralela al ventrito cerca del borde posterior (fig. 9). Edeago: longitud 0.65 mm; anchura 0.41 mm, parámeros bifurcados apicalmente, el más grande de apariencia bilobulada en el extremo; piezas accesorias de los parámeros 4.5 veces más largos que anchos, con pilosidades desde la mitad hasta el ápice. Endofalo: ápice redondeado, saco interno con un par de dientes paralelos al endofalo en la base; sobre estos dientes, hay 2 dientes adicionales curvos con aspecto de media luna (fig. 15).
Figuras 8-12. Calymmaderus semioblongus sp. nov. 8) Vista dorsal; 9) vista ventral; 10) vista lateral; 11) metasterno; 12) antena.
Paratipo ♀. Aspecto general del cuerpo similar al macho. Largo 4.1 mm. Ancho 1.9 mm. Distancia interocular 1.4 veces el diámetro vertical del ojo, segunda y tercera suturas abdominales curvadas en el centro, quinta sutura ligeramente curvada a los lados.
Resumen taxonómico
Etimología. El epíteto semioblongus es aplicado por la similitud morfológica externa con la especie oblongus, conocida para varias localidades de México.
Material examinado. Holotipo ♂: CIFBUM-XIL No. 253, Michoacán, Taretan. 18.VIII.2017. C. Jurado Col. Trampa de luz blanca, madera estructural de pino. Col. C. Jurado. 19°20’3.32” N. 101°55’3.02” 0. 1,150 m.snm. Paratipos: 29 individuos CIFBUM-XIL Núms. 255 a 282 (14 ♂ y 14 ♀), mismos datos que el holotipo.
Comentarios taxonómicos
Variabilidad: largo: 3.0-4.8 mm; ancho 1.4- 2.1 mm. Especie muy homogénea. Los machos notablemente menores que las hembras, con tamaño que varía de 3.0 a 3.7 mm (a veces hasta 3.8 mm), con ojos más pequeños y menos separados que en las hembras (distancia interocular de 0.8 a 1.2 el diámetro vertical de un ojo); área frontal algo más aplanada que la hembra, surco longitudinal del metasterno más largo que en la hembra.
Figura 13. Emergencia de Calymmaderus semioblongus sp. nov., en 2 ciclos anuales.
Tabla 2
Medidas y variación de largo y ancho de machos y hembras adultos de Calymmaderus semioblongus sp. nov.
Medidas
Promedio (mm)
Error estándar
Rango (mm)
N (machos) = 15 N (hembras) = 15
Machos
Hembras
Machos
Hembras
Machos
Hembras
Longitud
3.507
4.133
0.059
0.070
3.0-3.8
3.8-4.8
Ancho
1.627
1.886
0.025
0.026
1.4-1.7
1.7-2.1
Tabla 3. Medidas y variación de largo y ancho de machos y hembras adultos de Calymmaderus oblongus de la localidad de Tacícuaro, Michoacán.
Medidas
Medidas
Promedio (mm)
Error estándar
Rango (mm)
N (machos) = 15 N (hembras) = 15
n♂= 15 n♀= 15
♂
♀
♂
♀
♂
♀
Longitud
Longitud
3.507
4.133
0.060
0.070
3.0-3.8
3.8-4.8
Ancho
Ancho
1.627
1.887
0.025
0.026
1.4-1.7
1.7-2.1
Figura 14-16. Estructura genital de machos de las 3 especies de Calymmaderus vistas al microscopio compuesto 100X: 14) C. robustus sp. nov; 15) C.semioblongus sp. nov; 16) C.oblongus.
Hembra: cuerpo más grande que el macho, de 3.9 a 4.8 mm, ojos más grandes y separados que el macho (distancia interocular de 1.4 a 1.6 veces el diámetro vertical del ojo), última sutura esternal más curva a los lados que en el macho.
Especie de hábitos nocturnos asociada con madera estructural de Pinus sp. Se capturó en el interior de la techumbre de madera del templo de San Ildefonso en Taretan, Michoacán, con trampa de luz. La emergencia de los adultos inicia la última semana del mes de junio, en el mes de julio se registra la máxima emergencia con el inicio del verano, hasta disminuir en agosto con pocos ejemplares y en septiembre su emergencia es rara. Cohabita con otras especies xilófagas de Ptinidae y de termitas. Reinfesta las estructuras de madera por muchos años en el mismo sitio (fig. 13).
Calymmaderussemioblongus sp. nov. es una especie diferente a las otras especies mexicanas del género (White, 1983). Muestra similitud morfológica con la especie oblongus, pero con diferencias evidentes como la coloración del integumento (café oscuro en la especie descrita, a diferencia de oblongus que tiende más a café rojizo; sus medidas de longitud y ancho del cuerpo son menores (3.0 a 4.5 mm de longitud y ancho 1.4 a 2.1 mm, tabla 2), en comparación con C. oblongus (de 3.3 a 5.2 mm de largo y ancho de 1.6 a 2.4 mm, tabla 3). Ambas especies presentan de 2 a 3 hileras de puntuaciones laterales desde la base elitral que forman estrías; en oblongus, la estría más externa y cercana a la ranura elitral es más corta y las otras 2 estrías son menos marcadas que en la especie que se describe. La comparación de la estructura genital de machos de C. oblongus y C. semioblongus sp. nov. también mostró diferencias claras en el número y posición de espinas en el endofalo de estas 2 especies; C.oblongus presenta 2 espinas en forma de cornamenta en la parte más alejada de la base (figs. 14-16).
Agradecimientos
Al Laboratorio de Investigación en Invertebrados de la Facultad de Biología y a la División de Estudios de Posgrado de la Facultad de Ingeniería en Tecnología de la Madera de la UMSNH, por apoyar el proyecto de insectos xilófagos en madera estructural en Michoacán. A Mauricio Quesada, jefe del laboratorio Nacional de Síntesis Ecológica, ENES, UNAM, Unidad Morelia, por facilitar el uso del microscopio Axiostar Zoom V16, para la toma de imágenes de ejemplares. A las autoridades eclesiásticas del Templo de la “Asunción” de Taretan (en especial al párroco Francisco Javier Valencia Durón), por permitir el trabajo de campo en el inmueble. A Roberto Sibaja por la elaboración y ubicación del mapa del sitio. Finalmente agradecer a José Fernando Villaseñor Gómez, de la Facultad de Biología, quien colaboró en la última revisión del texto, con sugerencias acertadas y positivas.
Referencias
Arango, R. A. y Young, D. K. (2012). Death-watch and spider beetles of Wisconsin—Coleoptera: Ptinidae. USDA Forest Service, Forest Products Laboratory, General Technical Report FPL-GTR-209. https://doi.org/10.2737/FPL-GTR-209
Bercedo, P., García, B. R. y Arnaiz, L. (2008). El género Calymmaderus Solier, 1849 nuevo para Canarias y descripción de una nueva especie (Coleoptera: Ptinidae: Dorcatominae). Boletín de la Sociedad Entomológica Aragonesa, 42, 33–35.
Blackwelder, R. E. (1945). Checklist of the coleopterous insects of México, Central America, The West Indies, and South America (Ptinidae, Gnostidae and Anobiidae). Bulletin of the United States National Museum, 185, 401–406. https://doi.org/10.5479/si.03629236.185.3
Cibrián, T. D., Méndez M. J. T., Campos, B. R., Harry, O. Y. y Flores, L. J. (1995). Insectos forestales de México. Texcoco, Estado de México: Universidad Autónoma Chapingo.
Español, C. F. (1992). Coleoptera: Anobiidae. Fauna Ibérica, Vol. 2. Museo Nacional de Ciencias Naturales. Madrid: CSIC.
Gaviño, G., Juárez, L. C. y Figueroa, T. H. (2004). Técnicas biológicas selectas de laboratorio y campo. México D.F.: Limusa.
Halffter, G. (2017). La Zona de Transición Mexicana y la megadiversidad de México: del marco histórico a la riqueza actual. Dugesiana, 2, 77–89. https://doi.org/10.32870/dugesiana.v24i2.6572
INEGI (Instituto Nacional de Geografía y Estadística). (2010). Compendio de información geográfica municipal. Taretan, Michoacán de Ocampo. Clave geoestadística 16087. Instituto Nacional de Geografía y Estadística. Recuperado el 17 diciembre, 2024 de: https://www.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/16/16087.pdf
Jurado-Vargas, C. M. A. (2020). Incidencia y daño de insectos en la techumbre de madera de cinco edificios históricos (Templos) de Michoacán (Tesis doctoral). Facultad de Ingeniería en Tecnología de la madera, Universidad Michoacana de San Nicolás de Hidalgo. Morelia, Michoacán.
Jurado-Vargas, C. M. A., Campos, B. R. y Cruz-de León, J. (2003). Anóbidos asociados a la madera en uso en dos monumentos históricos de Michoacán. Entomología Mexicana, 2, 803–806.
Jurado-Vargas, C. M. A. y Cruz-de León, J. (2010). Insectos que dañan la madera de edificios históricos de Morelia, Michoacán. Entomología Mexicana, 9, 103–110.
Jurado-Vargas, C. M. A. y Cruz-de León, J. (2020). Evaluación de la incidencia de insectos xilófagos en la madera estructural del Templo de Ucareo en Michoacán. Entomología Mexicana, 7, 389–395.
Lüher, A. H. y Honour, R. (2017). Descripción de dos especies nuevas de Calymmaderus Solier, 1849 (Coleoptera: Ptinidae) de Chile. Boletín de la Sociedad Entomológica Aragonesa, 60, 105–110.
Pichardo, S. L., San Marino, C. A., Jiménez, Q. E. y Torres, H. B. (2017). Familia Ptinidae. En D. Cibrian-Tovar (Ed.), Fundamentos de entomología forestal (pp. 268–272). Texcoco, Estado de México: Universidad Autónoma Chapingo.
Toskina, I. N. (2000). New wood-boring beetles (Coleoptera: Anobiidae) from Paraguay. Russian Entomological Journal, 9, 199–240.
Viñolas, A. (2018). Nuevos datos sobre la validez específica de Ernobius vinolasi Novoa y Baselga, 2000 (Coleoptera: Ptinidae: Ernobiinae). Arquivos Entomoloxicos, 19, 75–80. https://doi.org/10.5281/zenodo.12767756
White, R. E. (1971). Key to North American genera of Anobiidae, with phylogenetic and synonymic notes (Coleoptera). Annals of the Entomological Society of America, 64, 179–191. https://doi.org/10.1093/aesa/64.1.179
White, R. E. (1974). The Dorcatominae and Tricoryninae of Chile (Coleoptera: Anobiidae). Transactions of the American Entomological Society, 100, 191–253.
White, R. E. (1982). A catalog of the Coleoptera of America North of Mexico. Family Anobiidae. Systematic Entomology Laboratory, Agricultural Research Service, U.S. Dept. of Agriculture, U.S.A.
White, R. E. (1983). Keys to Neotropical species of Calymmaderus Solier and species of Calytheca White, taxonomic notes (Coleoptera: Anobiidae). Proceedings of the Entomological Society of Washington, 85, 229–250.
White, R. E. (1984). Eight new species of Anobiidae (Coleoptera) from Jamaica. The Coleopterists Bulletin, 38, 240–248.
Instituto de Ecología, A.C., Carretera antigua a Coatepec 351, Col. El Haya, 91073 Xalapa, Veracruz, Mexico
*Corresponding author: francisco.lorea@inecol.mx (F. Lorea-Hernández)
Received: 27 February 2024; accepted: 21 January 2025
Abstract
Following a detailed morphological survey of the Licaria collections in several herbaria, various taxonomic entities not recognized before were detected. Here, 11 new species of Licaria from Mesoamerica are described and illustrated. Possible relations to other species in the genus are commented.
Keywords: Lauraceae of Central America; Lauraceae of Mexico; Licaria of Central America; Licaria of Mexico
Nuevas especies de Licaria (Lauraceae) de Mesoamérica
Resumen
Como resultado de un análisis morfológico detallado de las colecciones del género Licaria en diferentes herbarios, se detectaron varias entidades taxonómicas no reconocidas previamente. Aquí se describen e ilustran 11 especies nuevas del género Licaria de la región mesoamericana. Se comentan además las posibles relaciones con otras especies del género.
Palabras clave: Lauraceae de Centroamérica; Lauraceae de México; Licaria de Centroamérica; Licaria de México
Introduction
Licaria is an endemic genus to the Americas represented mostly by medium-sized to large tree species that grow principally in the intertropical region of this part of the world. The hermaphrodite, perigynous flowers with only the third whorl of stamens fertile, whose anthers are bisporangiate, as well as the fruit often seated on a double rimmed cupule, constitute the combination of characters that distinguishes the genus among the Lauraceae. Licaria was last revised by Kurz (2000), who recognized 38 species, including 12 in the Mesoamerican area. That work is an adaptation with minor changes of Kurz doctoral dissertation (Kurz, 1983). Since then, in the lapse of 40 years, subtracting new combinations and synonyms, 31 new species have been described, adding 9 to the flora of Mesoamerica (Burger & van der Werff, 1990; Gómez-Laurito & Cascante, 1999; Gómez-Laurito & Estrada, 2002; Hammel, 1986; van der Werff, 1988, 2009). The impulse given to field work with the onset of the Flora Mesoamericana project resulted in a significant increment of herbarium specimens during the last 2 decades of the past century, but Kurz did not see most of them; those specimens have been frequently the source of new species described afterwards in Licaria. Furthermore, the expanded collections have improved our knowledge about variation of morphology within species and consequently have led to better circumscriptions of them. On the other hand, it is important to say that Kurz missed part of the diversity of Licaria because he relied mainly on what the big museums of Europe and the USA have, but did not search in several of the smaller regional herbaria; some of which preserve important specimens that support the recognition of several species that have elsewhere been synonymized or validate larger geographic distributions of taxa.
Contrary to what is found in most American genera of the Lauracae, Licaria presents a rather wide variation in flower morphology, e.g., in orientation of tepals, extent of stamens fusion, presence and extent of fusion of glands, presence of staminodes, and shape and position of anther openings. These features, together with the presence and type of hairiness on flower parts, as well as position and structure of inflorescence, constitute the basis for the recognition of species in the genus (Kostermans, 1937; Kurz, 2000; Mez, 1889). In the course of the revision of Licaria for the Flora Mesoamericana project, a number of herbarium specimens with distinctive character combinations were found, which had not been recorded previously in the genus. Differences are mainly in flower morphology, but complementary vegetative features or other data emphasize their singularity. In several cases, there are no additional specimens, but the collection on which the description is based. This situation has not been considered a drawback since, as is discussed in every case, the peculiarity of the plants is so patent that there is little chance to think they represent part of the variability of any other known species. The case strengthens the idea that we have as yet an incomplete picture of the extent of diversity and distribution in many taxa. Field work is still important and necessary.
Materials and methods
Specimens identified as Licaria (as well as unidentified, putative lauraceous material) collected in the Mesoamerican area from different herbaria (A, CAS, DS, ENCB, F, GH, HEM, K, MEXU, MICH, MO, NY, P, TEX, US, XAL) were carefully analyzed, both for vegetative and reproductive morphology. Particular attention was paid to floral characters; flowers were dissected using a Zeiss, Stemi DV4 stereomicroscope. Information of vegetative characters was collected directly from dry specimens, while floral morphology was surveyed in rehydrated material. Groups of specimens sharing similar morphology were matched with keys and descriptions of currently accepted species, in order to apply the correct names to them. To prevent misinterpretations, all available type specimens (either from herbaria cited above or accesible on Global Plants database (http://plants.jstor.org/search?plantNam) were also considered during the process of identification. Specimens that did not fit any of the known species were evaluated to determine their singularity; those whose floral and vegetative features combined do not overlap with that of accepted taxa are here proposed as new species. Following the methodology mentioned above, description of vegetative characters is based on dry specimens, while floral characters are described as they look in rehydrated material. In order to apply a standardized nomenclature to characters, applicable information in Radford et al. (1974) was used; particularly, base and apex of leaves, pubescence, tridimensional form of flowers, and shape of tepals were described according to that text.
Diagnosis. Trees similar to L. alata Miranda, but different because of the convex flanks of the base of the leaves, sparsely sericeous lower leaf surface, obloid flowers, densely tomentose tepals abaxially, partially pubescent staminal filaments abaxially and adaxially, densely tomentose hypanthium outside, densely hirsute-tomentose inside, and sparsely puberulent style.
Trees up to 15 m tall; twigs sericeous, soon glabrate, brownish gray or dark brown, lenticellate, conspicuously ridged, the ridges narrow, wing-like (at least when dry), each ridge originates at one extreme of petiole insertion, acropetal. Buds densely sericeous-tomentose. Leaves alternate, petioles 9-10.5(-12) mm long, bi-marginate above, glabrous, round below, sericeous, glabrescent, blades (17.5-)23-28 × (3.5-)4.5-7 cm, narrowly elliptic, pinninerved, secondary veins 11-13 pairs, upper leaf surface glabrous, lower surface sparsely sericeous, glabrescent, leaf apex acuminate, leaf base obtuse, somewhat conduplicate, the basal flanks of the blade convex and projected above the midvein. Inflorescences (7.5-)10-16.5 cm long, axillary to tiny, deciduous bracts, on the proximal section of new twigs, paniculate, sericeous-tomentose, peduncle (2.5-)4-6.5 cm long, sometimes glabrescent, pedicels 0.5-1.2(-2.5) mm long, densely tomentose, hairs yellowish-gray to yellowish-brown. Flowers obloid, tepals conspicuously inflexed, outer tepals 0.8-1 × 1.7-2 mm, very widely ovate, densely tomentose abaxially, slightly papillose toward the apex, glabrous or sometimes with scattered sericeous hairs at the base adaxially, inner tepals 0.6 × 1-1.1 mm, ovate or elliptic, tomentose abaxially, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 1-1.1 mm long, fused throughout, filaments pubescent at the base abaxially, glabrous adaxially or pubescent on the upper section, anthers 0.2-0.3 mm long, glabrous, with apical openings, glands ca. 0.2 mm, free, compressed, glabrous, sometimes reduced or absent, hypanthium ca. 0.8 mm deep, densely tomentose outside, hirsute-tomentose inside, pistil 1.3-1.4 mm long, ovary 0.8-1 mm long, glabrous, style sparsely puberulent. Fruits unknown.
Taxonomic summary
Type. Mexico. Chiapas: municipality of Petalcingo (actually municipality of Tila), steep slope of Ahk’ulbal Nab above Petalcingo, 1,700 m, 28 March 1981, D. E. Breedlove 50394 (holotype CAS; isotype MO 6485834).
Etymology. This species is dedicated to Dr. Dennis E. Breedlove, who spent so much effort in field work documenting the flora of Chiapas, pursuing the aim of elaborating a plant species compendium for that region of Mexico.
Distribution and habitat. So far the species is known only from the type collection, in an area covered by montane rain forest which, according to Breedlove (data from label of type specimen), had a canopy layer reaching 25 – 35 m, and species of the genera Alfaroa, Brunellia, Calatola, Hedyosmum, Matudea, Meliosma, Nectandra, Oreopanax, Quercus, and Turpinia, among others.
Phenology. Regarding the date when the plant was collected, it must have flowers around the end of winter and early days of spring; fruit season is not known, but possibly occurs during winter, for it has been observed that maturation of fruits in most Lauraceae takes around a year after flowering. It seems that the species is not deciduous.
Conservation status. Given the deep environmental degradation that currently prevails in the region where the species is only known, it is suspected that it is critically endangered.
Remarks
The presence of erect, concave tepals, and stamens with extrorse sporangia places L. breedlovei in subgenus Licaria. It is one element in the group (here called Licaria excelsa species group) constituted by L. alata, L. excelsa Kosterm., L. minutiflora (here described), L. pergamentacea W. C. Burger, L. sarukhanii (here described), and L. tomentulosa (here described), as it shares with them morphological features like ridged twigs, large narrow-elliptic leaves (frequently reaching 28-30 cm long), and anthers with sporangia opening apically. Within the group, L. breedlovei is distinguished from the other species by the combination of somewhat conduplicate leaf-base, lower leaf surface persistently sericeous, stamens fused throughout, and sparsely pubescent style. In addition, it differs particularly from L. alata for the obloid flowers (vs. ellipsoid), tepals densely tomentose abaxially (vs. tepals glabrous abaxially), filaments pubescent abaxially and adaxially (vs. filaments glabrous), and hypanthium densely tomentose outside and inside (vs. hypanthium glabrous on both faces).
Diagnosis. Trees, similar to L. misantlae (Brandegee) Kosterm., glabrous throughout, leaves mostly caudate, inflorescences botryoid, flowers long-pedicellate, without staminodes, and fruit cupule thick, deeply crateriform, its outer rim slightly lobed.
Trees up to 25 m; twigs glabrous, smooth or slightly ribbed, reddish-brown or grayish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (3.5-)6.5-9(-11) mm long, slightly sulcate above, rounded and smooth or slightly ribbed below, glabrous, blades (4.5-)7-11(-13) × (1.5-)2.5-4(-5) cm, elliptic or narrowly elliptic, pinninerved, secondary veins 6-11 pairs, both upper and lower leaf surfaces glabrous, leaf apex caudate, sometimes acuminate, base cuneate to obtuse. Inflorescences 2.5-5 cm long, axillary to tiny, decidous bracts, disposed on very short shoots axillary to leaves, botryoid, or apparently paniculate, due to suppression of terminal bud of the floriferous branchlet, glabrous throughout, peduncle 0.2-1.7 cm long, pedicels (6-)10-13(-16) mm long, glabrous. Flowers widely obovoid to turbinate, greenish-yellow, tepals concave, conspicuously inflexed, outer tepals 0.9-1.3 × 1.1-1.7 mm, widely ovate, blunt cuspidate, glabrous abaxially and adaxially, short-ciliate, inner tepals 0.75-1.1 × 1.1-1.2 mm, ovate to widely ovate, glabrous abaxially and adaxially, short-ciliate, staminodes of whorls I and II absent, stamens of whorl III 0.6-1 mm long, fused throughout, filaments glabrous abaxially and adaxially, sometimes with a few hairs at the base abaxially, anthers ca. 0.1 mm, glabrous, with apical openings, glands ca. 0.4 mm, free, oblong, sometimes almost square, obtuse or sub-acute, glabrous, hypanthium 0.6-0.9 mm deep, glabrous outside and inside, pistil ca. 1.5 mm long, glabrous throughout, ovary 0.7-0.9 mm long. Fruits ca. 27 × 21 mm, ovoid or ellipsoid, cupule ca. 15 × 20 mm, crateriform, clearly bimarginate, inner margin ca. 2 mm tall, erect, outer margin ca. 3.5 mm tall, oblique to slightly reflexed, shallowly lobed, thick, pedicel ca. 5.5 × 4.5 (at the base) and 7 (at the apex) mm, obconic, continuous with the cupule.
Taxonomic summary
Type. Costa Rica. Puntarenas: Reserva Forestal Golfo Dulce, Osa Peninsula, Rancho Quemado, 8°44’ N, 83°36’ W, 200-300 m, 2 May 1988, B. Hammel et al. 16790 (holotype MO; isotypes MEXU, TEX).
Figure 1. Licaria breedlovei Lorea-Hern. sp. nov., general view.
Paratypes. Costa Rica. Guanacaste: Cantón de Tilarán, San Gerardo Abajo, río Caño Negro, Fincas Quesada y Arce, 10°18’40’’ N, 84°50’02’’ W, 1,100-1,200 m, 5 December 1991, E. Bello & E. Cruz 4262 (XAL). Puntarenas: Cantón de Osa, Rincón, filas al margen izquierdo de Quebrada Vaquedano, 8°38’45’’ N, 83°35’25’’ W, 400 m, 21 July 1990, G. Herrera 4000 (XAL); Cantón de Osa, Aguabuena, cuenca media y superior de Quebrada Orito, Rincón, 8°42’40’’ N, 83°31’40’’ W, 400 m, 25 October 1990, G. Herrera 4510 (MEXU); Cantón de Osa, Rancho Quemado, sector oeste, Sierpe, 8°41’00’’ N, 83°35’40’’ W, 350 m, 25 August 1982, J. Marín & G. Marín 499 (MO, XAL).
Figure 2. Licaria dolichopoda Lorea-Hern. sp. nov., general view.
Etymology. The name of this species alludes to the distinctive long pedicels that bear the flowers.
Distribution and habitat. Currently the species is known from 2 rather distant areas in Costa Rica that differ in ecological conditions. One of them, in the province of Guanacaste, is located in the Pacific foothills of the southern end of the Sierra de Guanacaste, while the other, in the province of Puntarenas, is in the lowlands of the Peninsula de Osa toward the southeastern extreme of the country. The first one is covered by pre-montane humid forest, and the second by humid tropical forest.
Phenology. Flowering is apparently distributed in 2 peaks: May-July and October-December. Fruits are only known from October.
Conservation status. There is no information about the abundance of the species in the places where it has been collected, neither about the ecological conditions of the vegetation there. However, the distribution range in the southeast is embedded in the Reserva Forestal Golfo Dulce, by the boundaries with the Parque Nacional Corcovado. Thus, there might not be problems with the persistence of the species in that region. The other point of the species distribution is, according to satellite images, within a region with deeply transformed vegetation, although not too far from the Parque Nacional Volcán Arenal, which could hold the species in its flora.
Because of the erect, concave tepals, and extrorse sporangia of stamens in the flowers, L. dolichopoda must be also considered in the subgenus Licaria. The glabrous condition found in every structure of the plant body (stem, leaves, and flowers), as well as the apical openings of stamens, and the lack of staminodes found in this species places it beside L. eurypaniculata (here described), from which it differs in the botryoid architecture of the inflorescence, and the conspicuous caudate leaves. Collection duplicates of L. dolichopoda have been previously distributed as L. cufodontisii Kosterm. (Herrera 4000), and as L. misantlae (Herrera 4510).
Diagnosis. Among the species with glabrous leaves, inflexed tepals, lacking staminodes, as well as apical anther openings, this species is distinguished by paniculate inflorescences where the flowers are not aggregate on the floriferous axes, orbicular flowers, tepal surface glabrous abaxially and adaxially, fused stamens throughout, glabrous hypanthium outside and inside, and glabrous pistil.
Trees 6-7 m tall; twigs glabrous, smooth or slightly ribbed, reddish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (5.5-)7-9(-10.5) mm long, slightly sulcate above, glabrous, blades (7-)11-16(-19.5) × (2-)3-4.5(-6) cm, narrowly elliptic to lanceolate or elliptic, pinninerved, secondary veins 7-9 pairs, both upper and lower leaf surfaces glabrous, leaf apex acuminate to short caudate, base obtuse to cuneate. Inflorescences 10-11 cm long, apparently terminal, if it is axillary to a bract, this one is inconspicuous, paniculate, basal axes longer than the peduncle, glabrous throughout or sparsely puberulent toward the end of the secondary axes, flowers spaced, not aggregate on the axes, peduncle ca. 3 cm long, pedicels 2-3.2 mm long, glabrous or sparsely puberulent. Flowers spheroidal, greenish, tepals conspicuously inflexed, outer tepals ca. 0.5 × 0.9-1.0 mm, widely ovate to depressed ovate, glabrous abaxially and adaxially, inner tepals ca. 0.4 × 0.6-0.7 mm, ovate or almost orbicular, glabrous abaxially and adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.5-0.6 mm long, fused throughout, filaments glabrous outside, sparsely tomentose inside, anthers ca. 0.1 mm, glabrous, with apical openings, glands ca. 0.1 mm, free, widely elliptic or orbicular, glabrous, hypanthium ca. 0.7 mm deep, glabrous outside, glabrous or sparsely pubescent on the distal section inside, pistil ca. 1.1-1.2 mm long, glabrous throughout, ovary ca. 0.6-0.7 mm long. Fruits (not wholly ripe) 15-17 × 12-12.5 mm, ovoid or ellipsoid, cupule 10-11 × 14-15 mm, crateriform, clearly bimarginate, inner margin 1.1-1.3 mm tall, erect, outer margin 0.5-0.7 mm tall, perpendicular to the inner one, pedicel 3-3.6 × 2-2.2 (at the base) and 3.6-4 (at the apex) mm, obconic.
Taxonomic summary
Type. Panama. Bocas del Toro: along road to Chiriquí Grande, c. 10 road miles from continental divide and about 2 miles along road east of highway, 8°45’ N, 82°15’ W, 300 m, 15 April 1987, G. McPherson 10830 (holotype MO; isotype XAL 124596).
Paratype. Panama. Bocas del Toro: along road to Chiriquí Grande, 10 road-miles from continental divide, ca. 2 road-miles along road east of highway, 8°55’04’’ N, 82°10’04’’ W, 300 m, 9 February 1987, G. McPherson 10453 (MO 5048286).
Etymology. The name of this species alludes to the very long basal secondary axes of the inflorescence, which give a broad triangular profile to the panicle.
Distribution and habitat. The only 2 known collections of the species come from the same area in the Atlantic lowlands of western Panama. No information about the vegetation found at the site was recorded, but considering the geographical factors of the place, it is expected to be tropical rain forest.
Phenology. Flowers in spring, and fruits mature in winter or early spring. The plant is not deciduous.
Conservation status. It seems that the species is not frequent in the area where it was found, for it has been collected only twice. On the other hand, the view of the area from satellite images shows that most of the original vegetation has been cleared; therefore, the species might be endangered.
Remarks
There are no species that come close morphologically to L. eurypaniculata. The other species with a general glabrous condition almost throughout the plant body, L. dolichopoda, is very different, in the shape of the leaves and the structure of the inflorescence. Given the erect, concave tepals, and stamens with extrorse sporangia that L. eurypaniculata presents, it is also a member of the subgenus Licaria.
Diagnosis. Trees similar to L. excelsa, but distinguished by the obovate to oblanceolate leaves, densely yellowish to orange-brown puberulent inflorescences, tepals with basal half conspicuously swollen, puberulent abaxial surface, sericeous adaxial surface at the base, apically pubescent ovary, and pubescent style.
Trees 15-20 m tall, trunk ca. 25 cm DBH, bark smooth, fragrant; twigs hollow, inhabited by ants, glabrous, sparsely lenticellate, slightly ridged, each ridge originating at one extreme of petiole insertion, acropetal. Buds glabrous or partially pubescent. Leaves alternate, petioles 10-12(-14) mm long, bimarginate above, glabrous, blades (20-)24-30 × (9-)11-13 cm, obovate or oblanceolate, sometimes elliptic, pinninerved, secondary veins (12)14-16 pairs, leaf surface glabrous above and below, leaf apex apiculate, sometimes apiculate-acuminate, leaf base obtuse or rounded. Inflorescences 9-15 cm, seemingly terminal, but actually axillary to tiny, deciduous bracts, on the proximal section of new twigs, paniculate, conspicuously puberulent along all axes, hairs yellowish to orange-brown, peduncle (1-)3.5-4.5 cm, sparsely puberulent, glabrescent, pedicels 2.5-5.5(-7) mm, densely puberulent. Flowers widely obovoid, yellowish-green, fragrant, perianth thick, coriaceous, tepals clearly inflexed, their base conspicuously swollen, outer tepals 1.2-1.4 × 1.6-1.9 mm, widely ovate, puberulent abaxially, sericeous-tomentose at the base and toward the margins adaxially, hairs orange-brown, inner tepals 0.8-1 × 1-1.2 mm, ovate, puberulent abaxially, sericeous-tomentose at the base and tomentose at the middle adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.7-0.8 mm long, fused along their filaments, filaments tomentose at the base outside, tomentose throughout inside, hairs orange-brown, anthers ca. 0.2 mm, free or fused just at the base, glabrous, with apical openings, glands ca. 0.3 mm, only 3 given the fusion of adjoining glands, widely oblong, glabrous, hypanthium 1-1.2 mm deep, obconic, densely puberulent outside, hairs yellowish to orange-brown, sericeous-tomentose inside, hairs reddish-brown, pistil 1.6-1.8 mm long, top of the ovary and style pubescent, ovary 0.8-1.2 mm long. Fruits (nearly ripe) 19-23 × 15.5-16.5 mm, ellipsoid, cupule ca. 16.5 × 18.5 mm, cotyliform, sparsely lenticellate, seemingly tri-margined, the 2 regular margins plus the swollen projections of the tepals, inner margin ca. 1.6 mm tall, erect, outer margin 2.2-2.5 mm tall, erect, pedicel 4.5-7 mm long, continuous with the base of the cupule, 3.5 mm diam. at the base.
Taxonomic summary
Type. Costa Rica. Limón: Reserva Biológica Hitoy Cerere, 300 m aguas abajo de la confluencia del río Hitoy con el río Cerere, margen izquierda por la fila que lleva al cerro Bobócora, 9°39’00’’ N, 83°02’45’’ W, 200 m, 20 February 1989, G. Herrera & A. Chacón 2427 (holotype MO, isotype XAL 124445).
Etymology. The name is derived from the conspicuous swollen condition of the base of the tepals; the feature is distinctive.
Distribution and habitat. So far, this species is known only from the southwestern end of Costa Rica. Although no information about the habitat was recorded, there is no doubt that the place lies within the tropical rain forest territory.
Figure 4. Licaria eurypaniculata Lorea-Hern. sp. nov., general view.
Phenology. With flowers and ripe fruits around the end of winter. The species has perennial leaves.
Conservation status. There is no information about the abundance or extent of distribution of the species, but as it is known to grow in the grounds of a nature reserve, it can be expected that it is not under high risk of extinction but certainly endangered.
Figure 5. Licaria gibbitepala Lorea-Hern. sp. nov., general view.
Remarks
Licaria gibbitepala seems to be closely related to L. tomentulosa and, at the same time, to the group of species around L. excelsa. All of them have ridged twigs, large leaves, flowers with erect, concave tepals, no staminodes, and sporangia with apical openings. The hollow twigs, along with the obovate to oblanceolate leaves, and the conspicuous gibbous base of the tepals distinguish L. gibbitepala.
Trees to 6 m tall; twigs smooth, dark brown to reddish brown, pruinose, puberulent, glabrescent, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (4-)8-11 mm long, puberulent, soon glabrous, canaliculate above, blades 8.5-13 × 3.5-5.5 cm, elliptic or narrowly elliptic, pinninerved, secondary veins 5-7 pairs, leaf surface glabrous above and below, but the lower surface puberulent in the beginning, leaf apex caudate, sometimes just acuminate, base obtuse to cuneate. Inflorescences 2.5-3.5 cm long, axillary to leaves and to tiny, deciduous bracts, on the proximal section of new twigs, botryoid, racemiform, few-flowered (less than 10 flowers), glabrous throughout or with some hairs toward the end of the peduncle, peduncle 1.8-2 cm long, pedicels 3-4.5 mm long, glabrous. Flowers obovoid, pale yellow, tepals erect to slightly inflexed, concave, outer tepals ca. 0.8 × 1.3 mm, very widely ovate to depressed ovate, glabrous abaxially, with some long, appressed hairs ascending from the base adaxially, inner tepals ca. 0.5 × 0.8 mm, ovate or widely ovate, glabrous abaxially, adaxially like the outer tepals, staminodes of whorls I and II absent, stamens of whorl III ca. 0.7 mm long, free, but very close one to each other, widely ovate in outline, filaments sparsely tomentulose outside and inside, anthers ca. 0.4 mm, tomentulose at the base outside, tomentulose inside along the central line, openings lateral, oblique, glands ca. 0.4 mm, rounded, glabrous, hypanthium ca. 0.7 mm deep, obconic, glabrous inside and outside, pistil ca. 1.7 mm long, glabrous, ovary ca. 0.8 mm long. Fruit unknown.
Figure 7. Licaria gracilis Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Panama. Chiriquí: Punta Burica, El Chorogo, alrededores de la finca de Fernando Chavarría, adyacente al límite fronterizo, cabecera del río San Bartolo, 8°17’07’’ N, 82°58’56’’ W, 395 m, 15 May 2007, J. E. Aranda et al. 3912 (holotype MO 6456198).
Etymology. The name of the species is derived from the attractive view that the slender inflorescences give to the plant.
Distribution and habitat. The species is known only from the location where it was first and last collected. No information about the habitat is mentioned in the data-label of the specimen, except that was collected nearby a ranch. The original vegetation must have been tropical evergreen forest or semi-evergreen forest.
Phenology. Flowers during spring; ripe fruits expected during winter or early spring, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. The species might be (critically) endangered, for most of the land in the area where it was collected has been transformed for diverse agricultural purposes.
Remarks
The few-flowered, botryoid inflorescence, tiny flowers, and very oblique, lateral openings of the sporangia distinguish this species straightaway. There is no species whose general morphology indicates association with L. gracilis. The singular way that the openings of the stamens are displayed resemble that found in Licaria cogolloi van der Werff, and L. caribaea Gómez-Lau. & Cascante, but besides this, there is no other feature that might suggest a relationship to those species. For the features of its flowers, Licaria gracilis belongs to subgenus Licaria.
Figure 8. Licaria minutiflora Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. pergamentacea, but distinct by the presence of hollow twigs, glabrous buds, abaxially densely tomentulose tepals, internally pubescent hypanthium, apically pubescent ovary, and pubescent style.
Trees up to 20 m tall, trunk ca. 30 cm DBH; twigs hollow, inhabited by ants, glabrous, smooth or slightly ridged, each ridge originates at one extreme of petiole insertion, acropetal, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (7-)15-20(-30) mm, glabrous, bimarginate above, blades (11-)20-28(-34.5) × (3-)5-9(-13) cm, narrowly elliptic or narrowly ovate, pinninerved, secondary veins 9-12 pairs, leaf surface glabrous on both sides, leaf apex acute, slightly apiculate, base acute or obtuse. Inflorescences (6-)9-12(-15) cm long, axillary to tiny, deciduous bracts, on the proximal section of new twigs or axillary to leaves, paniculate, tomentulose, peduncle (0.5-)1.5-3(-4) cm long, pedicels (1.2-)2.5-4(-6) mm long, densely tomentulose. Flowers obovoid or ellipsoid, yellowish-green, tepals inflexed, outer tepals 0.5-0.8 × 0.8-1 mm, widely ovate, densely tomentulose abaxially, tepal surface concealed or almost so by hairs, glabrous adaxially, inner tepals 0.5-0.7 × 0.5-0.6 mm, ovate, densely tomentulose abaxially, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.5-0.6 mm long, free or barely united by filament base, filaments tomentose at base outside, glabrous or sparsely pubescent along the medial line inside, anthers ca. 0.2 mm, glabrous, openings apical, glands ca. 0.2 mm, free, rounded, hypanthium 0.6-0.8 mm deep, obconic, densely tomentulose outside, sericeous on upper half inside, hairs yellowish or reddish, pistil ca. 1.4 mm long, top of the ovary and style pubescent, ovary 0.8-1 mm long. Fruits ca. 19.5 × 15-16.5 mm, ellipsoid, cupule 15-16 × 18-20 mm, urceolate, lenticellate, conspicuously bimarginate, inner margin 1.2-1.5 mm tall, outer margin 1-1.2 mm, extended, pedicel 6.5-9 mm long, continuous with the cupule, 2.8-3.6 mm diameter at base.
Taxonomic summary
Type. Costa Rica. Alajuela: camino entre la estación de la Reserva Forestal de San Ramón y el camino a la colonia Palmareña, finca de don Bolívar Ruiz, margen derecha río San Lorencito, 10°12’53’’ N, 84°36’28’’ W, February 1987, G. Herrera 500 (holotype MO 3587621; isotypes MEXU 638804, TEX).
Paratypes. Costa Rica. Alajuela: Reserva Biológica Monteverde, Poco Sol, La Cutacha de San Bosco, 10°22’ N, 84°40’ W, 900 m, 1 April 1989, E. Bello 784 (MEXU 1304162; MO 6142952; XAL 124442); Bosque Eterno de los Niños, Reserva de Arenal, río Peñas Blancas, Quebrada Agua Gata, Finca Villalobos, 10°23’ N, 84°42’ W, 1,000 m, 20 April 1990, E. Bello 2208 (MO 6130824; XAL 124446); Cantón de Upala, Colonia La Libertad, 10°52’ N, 85°17’ W, 300 m, 3 August 1991, Q. Jiménez & G. Rivera 1011 (MO 6117397; XAL 124597). Guanacaste: Parque Nacional Guanacaste, Estación Pitilla, 10°00’15’’ N, 85°25.2’ W, 500 m, 27 May 1989, G. Herrera et al. 2942 (MO; XAL 124428).
Figure 9. Inflorescence detail of Licaria gracilis Lorea-Hern. sp. nov. (above), and Licaria minutiflora Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Etymology. The very small flowers, whose tepals are less than 1 mm long, is the feature on which the species name is based.
Distribution and habitat. This species is known from the northern hills of the Cordillera de Guanacaste (Guanacaste Mountain Range) and the western part of the Cordillera Central (Central Mountain Range), between 200 and1,000 m asl. Prevailing vegetation in the region is tropical evergreen forest and montane rain forest. The species has been collected also in pasture fields. There is a fruiting specimen from the Osa Peninsula (Hammel et al. 16984) that seems to belong to L. minutiflora but until confirmed with a flowering specimen, the presence of the species in this part of the country remains uncertain.
Phenology. Flowers toward the end of winter and early spring; fruits must be ripe around the end of autumn or early winter.
Conservation status. Most specimens of this species have been collected within nature reserves. Therefore, it is considered not threatened, even though its abundance is still unknown.
Remarks
As has been mentioned elsewhere in this paper, Licaria minutiflora is part of the L. excelsa species group. Its general appearance resembles that of L. pergamentacea, for the size and shape of the leaves, structure and hairiness of the inflorescence, and for having small oblong-ellipsoid, not coriaceous flowers. However, besides the presence of hollow twigs, it differs by having flowers with the upper part of the hypanthium homogeneously pubescent inside, and a pubescent pistil.
Figure 10. Licaria ochracea Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. multinervis H. W. Kurz, but differing by lower surface indument of leaves composed by 2 types of hairs, the most numerous tomentulose, the fewer sericeous, long inflorescences, terminal or axillary to leaves, with rachis up to 15 cm, fully exserted anthers, clearly stalked glands, conspicuous hypanthium tube projected beyond the insertion point of stamens.
Trees up to 22 m tall, twigs smooth, densely tomentose, hairs initially yellowish-brown, then greysh, sparsely and inconspicuously lenticellate. Buds tomentose. Leaves alternate, petioles (10-)15-20 mm long, tomentose, glabrescent, channeled above, blades 12.5-18.5 × (2.5)3.5-4.5 cm, narrowly elliptic, sometimes narrowly oblanceolate, pinninerved, secondary veins (9)10-13 pairs, leaf surface glabrous above, tomentose below, most hairs sinuous, rather appressed, or patent, fewer hairs straight, appressed, leaf apex acuminate, base narrowly cuneate. Inflorescences (7-)10-18 cm long, axillary to leaves, less frequently terminal, paniculate, densely tomentose, hairs like on twigs, flowers agglomerate at the end of terminal axes, peduncle 0.2-1.5(-3.5) cm long, pedicels (1-)1.5-2 mm long, tomentose. Flowers narrowly oblong, tepals erect, concave, outer tepals, 1-1.3(-1.5) × (1-)1.1-1.3 mm, widely ovate, tomentose abaxially, glabrous adaxially, except for a few long, appressed hairs, coming from the base, inner tepals 1-1.1(-1.4) × 0.8-0.9 mm, ovate, with a pattern of pubescence similar to that of outer tepals, staminodes of whorls I and II absent, stamens of whorl III 3-3.3(-3.8) mm long, coherent along their filaments or even at the base of the anthers, easily separable, filaments tomentose on both faces, anthers 0.9-1.1 mm long, free or coherent at the base, glabrous, completely exserted, openings dorsolateral, glands 0.7-0.8 mm, free, oblanceolate, glabrous, clearly stalked, stalk pubescent, hypanthium 1.2-1.3(-1.6) mm deep, extended 0.3 mm beyond the insertion point of stamens, tomentose outside, densely hirsute-tomentose inside, hairs golden-brown, pistil 3.6-3.9 mm long, sparsely pubescent, at least some hairs on the upper half of the ovary and lower half of the style, ovary 0.9-1.1 mm long. Fruits unknown.
Figure 11. Licaria rufotricha Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Nicaragua. Matagalpa (according to current maps it should be Jinotega): pasture and small woods, tropical premontane forest, Hacienda Santa María del Ostuma, 10 km N of Matagalpa, 1,300 m, 17 July 1978, P. C. Vincelli 756 (holotype MO 2984723; isotypes LL, MEXU 691292).
Etymology. The name refers to the conspicuous orange-brown pubescence of the species on young twigs and main axes of inflorescences.
Distribution and habitat. Only known from the place of type collection, where the Cordillera Isabelia and Cordillera Dariense meet.
Phenology. Flowers in summer; fruit season is not known, but since it has been observed that maturation of fruits in most Lauraceae takes around a year after flowering, possibly ripe fruits are present during spring or early summer.
Conservation status. There is no certainty on the conditions that might be affecting the survival of the species. The type was collected in a private property that used to be preserved for ecotourism activities, but its current situation is unknown. However, since the species has not been collected again, it is suspected to be endangered.
Remarks
The collection on which the description of this species is based was initially identified by Kurz (2000) as L. multinervis; actually it is cited as a paratype of this species. But the impression of being conspecific with this taxon disappears with a detailed survey of the morphology. Important differences are the pubescence on the lower surface of the leaves (simple in L. multinervis vs. made of 2 types of hairs in L. ochracea), position of the inflorescences (axillary to small bracts in the proximal section of new branches in L. multinervis vs. axillary to leaves or terminal in L. ochracea), length of inflorescences (up to 3.5 cm long in L. multinervis vs. 5-7 times longer in L. ochracea), protrusion of anthers (partially exserted in L. multinervis vs. fully exerted in L. ochracea), and projection of hypanthium tube beyond the insertion point of stamens (short in L. multinervis vs. conspicuous in L. ochracea). However, L. multinervis and L. ochracea seem to be more closely related to each other than to the rest of species in the area that present fully exserted stamens, namely L. agglomerata van der Werff, L. capitata (Schltdl. et Cham.) Kosterm., L. nitida van der Werff, and L. vanderwerffii (here described).
Diagnosis. Small trees, distinct for the conspicuously obovoid flowers, aggregate toward the end of the secondary floriferous axes, with inflexed tepals, glabrous abaxially, reddish-brown hirsute-sericeous adaxially, lacking staminodes, stamens fused throughout, reddish-brown hirsute, anther openings apical, hypanthium mostly glabrous outside, reddish-brown hirsute inside, and ovary distally pubescent.
Trees to 12 m tall; twigs smooth or inconspicuously ridged, each ridge originating at one extreme of petiole insertion, acropetal, lenticellate, grayish, densely puberulent, glabrescent. Buds glabrous. Leaves alternate, petioles 10-13 (-17) mm long, slightly channeled above, puberulent, blades (14.5-)16.5-20(-23) × 4-5.5(-9) cm, elliptic or narrowly elliptic, pinninerved, secondary veins 6-7 pairs, leaf surface densely puberulent above when young, glabrescent, rather persistently puberulent below, leaf apex acuminate, base cuneate, sometimes obtuse or narrowly cuneate to indistinctly attenuate. Inflorescences (3-)5-8(-10.5) cm, axillary to tiny, deciduous bracts, on the proximal section of new twigs, or axillary to new leaves, paniculate, flowers aggregate at the end of secondary axes, densely puberulent on peduncle and rachis, glabrate toward the ultimate axes, peduncle (2-)3.5-5.5(-7.5) cm long, pedicels 2-3.5(-4.5) mm long, glabrous or sparsely puberulent. Flowers obovoid or obconic, pale yellow, tepals conspicuously inflexed, outer tepals 1-1.2 × 2-2.4 mm, widely ovate, glabrous abaxially, hirsute-sericeous adaxially, hairs reddish-brown, inner tepals 0.8-1 × 0.8-0.9 mm, ovate or elliptic, glabrous abaxially, generally mostly hirsute adaxially, hairs reddish-brown, surpassing the apical margin of the tepals, making them appear fimbriate, staminodes of whorls I and II absent, stamens of whorl III, 0.6-0.7 mm long, fused throughout, filaments hirsute on both faces, hairs reddish-brown, anthers ca. 0.2 mm, pubescent, except around the margin of the openings, hairs like those of the filaments, glands ca. 0.4 mm, free, rounded or oblate, glabrous, hypanthium 0.8-0.9 mm deep, glabrous or sparsely puberulent outside, hirsute inside, hairs like those of the filaments, pistil 1.7-2 mm long, ovary ca. 0.8 mm long, sparsely pubescent distally, hairs whitish. Fruits unknown.
Figure 12. Inflorescence detail of Licaria ochracea Lorea-Hern. sp. nov. (above), and Licaria rufotricha Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Taxonomic summary
Type. Panama. Panama: 5-10 km NE of Altos de Pacora, on trail at end of road, ca. 750 m, 7 March 1975, S. Mori & J. Kallunki 4977 (holotype MO 2992664).
Etymology. The name given to this species refers to the dense red-brown pubescence in the interior of the flowers, covering the adaxial surface of tepals, stamen filaments, anthers, and hypanthium inside.
Distribution and habitat. Just known from the place where the type was collected (around 25-30 km NE of Panama City), with tropical evergreen forest or semi-evergreen forest as original vegetation.
Phenology. Flowers around the end of winter or early spring; ripe fruits expected during winter, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. Probably the species is endangered. Most of the vegetation in the surroundings of the place where the species was collected has been cut down for cattle raising or agricultural purposes. But the southern boundary of the Parque Nacional Chagres (Chagres National Park) is very close to that place, and the Nature Reserve Cocobolo is a few kilometers ENE of it as well. So, even it has not been collected again, it is hoped the species is still around there.
Remarks
There is no other species in the Mesoamerican area with the distinctive reddish-brown pubescence found in the interior of the Licaria rufotricha flowers. Yellowish to reddish-brown pubescence is present on the filaments of stamens and/or inside the hypanthium of several species, like L. agglomerata, L. excelsa, L. gibbitepala, and L. tomentulosa, but it is never found on the tepals surface. In fact, its general morphology does not suggest a relation to any species in the region.
Diagnosis. Trees similar to L. alata, but differing by terminal, subsessile inflorescences, longer pedicels (up to 10.5 mm), flowers widely ellipsoid, abaxial tepal surface tomentose at the base and papillose toward the apex, hypanthium densely tomentose outside, sericeous-tomentose inside, and style pubescent.
Trees to 20 m tall, twigs glabrous, grayish to grayish-brown, slightly ridged, each ridge originating at one extreme of petiole insertion, acropetal, sparsely lenticellate. Buds glabrous or pubescent at the apex of the bud scales. Leaves alternate, petioles (11-)13-17(-20) mm long, slightly bimarginate, glabrous, blades (12.5-)20-32(-37) × (5-)7-10(11.5) cm, narrowly elliptic, occasionally oblanceolate, pinninerved, secondary veins (12)14-18 pairs, leaf surface glabrous on both sides, leaf apex acuminate, sometimes acute, base obtuse or acute. Inflorescences 12-22 cm long, terminal, paniculate, tomentose, soon glabrescent, hairs patent or oblique, grey or yellowish-grey, peduncle 0.2-0.4 cm long, pedicels (3-)5-8(-10.5) mm long, glabrous or sparsely tomentose. Flowers broadly ellipsoid, pale green, tepals inflexed, outer tepals 0.8-0.9 × 1.8-2 mm, very widely ovate, concave, tomentose over the basal half, and papillose-pubescent over the distal half abaxially, glabrous or with few straight hairs at base adaxially, inner tepals 0.7-0.9 × 0.4-0.8 mm, ovate, pubescence pattern abaxially like that of outer tepals, but less papillose, glabrous adaxially, staminodes of whorls I and II absent, stamens of whorl III 0.7-0.9 mm long, fused along their filaments, filaments tomentose along the central line on both faces, anthers 0.2-0.4 mm long, glabrous, openings apical, glands ca. 0.4 mm, oblong, flattened, frequently reduced or absent, glabrous or tomentose on the base abaxially, hypanthium ca. 1 mm deep, densely tomentose outside, hairs sinuous, sericeous-tomentose inside, pistil 1.3-1.6 mm long, ovary 0.8-1.2 mm long, glabrous, style rather densely pubescent. Fruits 27-30 × 17-18 mm, ellipsoid, cupule 15-17.5 × 18-19.5 mm, urceolate, bimarginate, but the margins barely discernible, ca. 0.5 mm, erect, pedicel 4-5 × 3.5-4.5 mm.
Figure 13. Licaria sarukhanii Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Mexico. Chiapas: municipio de Ángel Albino Corzo, Reserva de la Biosfera El Triunfo, aprox. 1.5 km al E de Campamento El Triunfo, sobre el Sendero Bandera, 15°36’ N, 92°50’ W, 1,850 m, 27 April 1993, F. Lorea 5522 (holotype XAL 151063; isotypes [to be distributed]).
Paratypes. Mexico. Chiapas: municipio de Ángel Albino Corzo, Polígono I de la Reserva de la Biosfera El Triunfo, 15°39’ N, 92°48’ W, 1,900 m, 21 February 1993, S. Solórzano 67 (MEXU 754117); Reserva El Triunfo, camino a Cerro de la Bandera, 25 March 1986, M. L. Ávila and V. H. Hernández s/n (MEXU 877857, TEX 146383); Reserva de la Biosfera El Triunfo, sendero Palo Gordo, 15°40’10’’ N, 92°48’42’’ W, 1,990 m, 22 March 2006, F. González-García s/n (XAL 113841).
Etymology. The species is dedicated to José Sarukhán, a Mexican plant ecologist who has played an important role in making a huge amount of information about Mexico’s biodiversity, ecosystem conservation, and sustainable development accessible to the public.
Distribution and habitat. So far, the species is known only from the central region of the Sierra Madre de Chiapas, from 1,800 to 2,000 m asl where the montane rain forest is the dominant type of vegetation.
Phenology. Flowers toward the end of winter and early spring; ripe fruits were found in the same interval of time.
Conservation status. All the collections of the species come from the nature reserve El Triunfo, located in the Sierra Madre de Chiapas, which has persisted largely undisturbed for its almost forty years of existence. Thus, considering the size of the area covered with montane rain forest in the reserve, the species might be regarded as vulnerable.
Remarks
Licaria sarukhanii is part of the L. excelsa species group, with a closer relation to L. breedlovei, because they share tomentose inflorescences and flowers rather widely ellipsoid to obloid. They differ by the conduplicate leaf bases, sparsely sericeous lower leaf surface, conspicuously pedunculate inflorescences axillary to small bracts or leaves, fused anthers, and style sparsely pubescent present in L. breedlovei, in contrast to flat leaf base, glabrous lower leaf surface, terminal, sub-sessile inflorescences, free anthers, and conspicuously pubescent style in L. sarukhanii.
Figure 14. Licaria tomentulosa Lorea-Hern. sp. nov., general view.
Diagnosis. Trees similar to L. excelsa, but distinguished by presenting hollow twigs, inflorescences yellowish-brown tomentulose, flowers conspicuously obovoid, tepals densely tomentulose abaxially, and apex of the ovary and style conspicuously pubescent.
Trees up to 25 m tall, and trunk 50 cm DBH, twigs hollow, inhabited by ants, smooth or slightly ridged, glabrous, grayish-brown, sparsely lenticellate. Buds glabrous. Leaves alternate, petioles (14-)17-22(-24) mm long, bimarginate above, glabrous, blades (22-)25-30(-34) × (6-)8-10(-12) cm, narrowly elliptic or elliptic, pinninerved, secondary veins (9)11-13 pairs, leaf surface glabrous on both sides, leaf apex acute or short acuminate, base obtuse, rounded or shortly attenuate, particularly in leaves close to the twig tips. Inflorescences 6.5-9.5(13.5) cm long, axillary to tiny, deciduous bracts, on very short branches axillary to leaves, paniculate, tomentulose, hairs yellowish-brown, peduncle 2.5-4 cm long, pedicels (1-)2-3.5 mm long, densely tomentulose. Flowers obovoid, thick, coriaceous, greenish-yellow, tepals inflexed, concave, outer tepals 0.7-1 × 1.4-1.7 mm, widely ovate, densely tomentulose abaxially, hairs almost concealing the surface, marginal tomentose bands and some long, appressed hairs at the base adaxially, inner tepals 0.6-0.9 × 0.8-0.9 mm, ovate, sometimes slightly conduplicate, densely tomentulose abaxially, pattern of pubescence adaxially like in the outer tepals, staminodes of whorls I and II absent, stamens of whorl III 0.8-1.1 mm long, fused throughout, filaments tomentose outside on lower half, densely tomentose all over the inner face, hairs reddish-brown, anthers ca. 0.2 mm, glabrous outside, densely tomentose inside, openings apical, glands 0.3-0.4 mm, frequently fused between adyacent stamens, transversely oblong, oblong when free, hypanthium 1-1.2 mm deep, densely tomentulose outside, tomentulose inside, sometimes only in the upper half, hairs mainly reddish-brown, pistil 2-2.3 mm long, ovary 1.1-1.4 mm long, apex of the ovary and style conspicuously pubescent. Fruits (not fully ripe) ca. 13 × 13 mm, spheroidal, cupule 13-14 × 17-18 mm, crateriform, lenticellate, bimarginate, inner margin 0.7-1 mm tall, erect, outer margin 0.5-0.7 mm, erect to spreading, pedicel 4-6 × 3-4 (apex) and 2 mm (base).
Taxonomic summary
Type. Costa Rica. Limón: Cordillera de Talamanca, Matina, intersección de río Barbilla y quebrada Cañabral, por la fila al norte, 10°01’ N, 83°24’ W, 100-200 m, 11 October 1988, G. Herrera 2165 (holotype MO 3693485; isotypes MEXU 529888, 529889, 717774, and 718334; TEX).
Paratypes. Costa Rica. Alajuela: Upala, Bijagua, El Pilón, Cerro La Carmela, entre río Celeste y cabeceras del río Chimurria, 10°43’15’’ N, 84°59’45’’ W, 1,000 m, 11 July 1988, G. Herrera 2056 (MEXU 529875, 717781; MO 3693486; US 3655515). Limón: Reserva Biológica Hitoy Cerere, Valle de la Estrella, sendero a Cerro Bobócara, 9°41’00’’ N, 83°04’20’’ W, 798 m, 17 August 1990, G. Herrera 4115 (MEXU 1304090; MO 6142953; XAL 124444); Reserva Indígena Talamanca, camino a Soki entre la Quebrada Amubri, margen izquierda de río Lari, 9°29’40’’ N, 82°89’40’’ W, 200 m, 29 June 1989, A. Chacón 25 (MO; XAL 124437).
Etymology. The name is for the abundant tomentulose pubescence that covers the axes of the inflorescence, as well as the abaxial surface of tepals and hypanthium.
Distribution and habitat. Known only from Costa Rica, along the Atlantic lowlands adjacent to the Sierra de Talamanca, and the Sierra de Guanacaste. Tropical rain forest used to be the predominant vegetation there, but currently a big amount of land has been changed to agricultural activities or other affairs.
Phenology. Flowers in summer and early autumn; ripe fruits in summer.
Conservation status. Given the deterioration of the habitat, the species might be considered vulnerable or endangered, in spite of its wide range of distribution.
Remarks
Licaria excelsa and L. tomentulosa are much alike. Considering the available material, distinguishing characters seem to work well to separate both species; hollow twigs, as well as type and distribution of pubescence on vegetative and floral parts, are consistently present in all the specimens considered as L. tomentulosa, making it a clear morphological unit, distinct from L. excelsa.
Figure 15. Inflorescence detail of Licaria sarukhanii Lorea-Hern. sp. nov. (above), and Licaria tomentulosa Lorea-Hern. sp. nov. (below). Scale bars 1 mm.
Diagnosis. Similar to L. agglomerata, but different by the free stamens, white pubescence of stamens, hypanthium densely white tomentose inside, and pistil glabrous.
Shrubs or small trees 1-3 m tall, twigs hollow, smooth, sparsely lenticellate, glabrous, reddish-brown. Buds glabrous. Leaves alternate, petioles ca. 12.5-15 mm long, slightly channeled and bimarginate above, glabrous, blades ca. 15-23 × 6-7.5 cm, narrowly elliptic, pinninerved, secondary veins 9-11 pairs, leaf surface glabrous on both sides, leaf apex acuminate, base obtuse or cuneate. Inflorescences ca. 1.5-2 cm long, axillary to leaves, axillary to tiny, deciduous bracts, on the proximal sections of new twigs too, paniculate, but strongly condensed, with very short rachis and secondary axes, peduncle ca. 0.5 cm long, glabrous, pedicels (1.2-)1.5-2.5(-3) mm long, glabrous. Flowers ellipsoid, slightly swollen in the middle of the hypanthium, greenish, tepals erect, outer tepals 0.5-0.6 × 0.5-0.6 mm, ovate, glabrous abaxially, glabrous adaxially except for some sericeous hairs at the base, inner tepals 0.4-0.5 × 0.2-0.3 mm, ovate, glabrous abaxially, glabrous adaxially or with some sericeous hairs at the base, staminodes of whorls I and II absent, stamens of whorl III 1-1.2 mm long, free, filaments white-tomentose throughout, denser on inside, anthers 0.3-0.4 mm, wholly exserted, glabrous or sparsely tomentose at the base outside, tomentose on lower half inside, openings dorsal, glands absent, hypanthium 1.1-1.3 mm deep, narrowly rhomboid, extending a little beyond the insertion point of stamens, glabrous outside, densely white-tomentose inside, pistil 2-2.3 mm long, glabrous, ovary 1-1.2 mm. Fruit unknown.
Figure 16. Licaria vanderwerffii Lorea-Hern. sp. nov., general view.
Taxonomic summary
Type. Panama. Darién: Cerro Tacarcuna S slope, 1,250-1,450 m, 26 January 1975, A. Gentry & S. Mori 13920 (holotype MO 2300651; isotype F 1763407).
Etymology. The species is named after Hendrik (Henk) Hessel van der Werff, a prolific scholar of the family Lauraceae in the Neotropics.
Distribution and habitat. So far, only known from the type collection, in the mountains of the southeastern edge of Panama, where premontane rain forest prevails.
Phenology. Flowers in winter; probably ripe fruits expected during autumn or early winter, since maturation of fruits in most Lauraceae takes around a year after flowering.
Conservation status. The place of the type collection is within the boundaries of the nature reserve Parque Nacional del Darién, but the altitudinal range within which the specimen was collected stretches barely for 40 km along the reserve. So, the species should be considered endangered.
Remarks
The collection on which the description of L. vanderwerffii is based was cited by Kurz (2000) as L. triandra, but there are several features that separate the former from this species. Whereas L. triandra presents branchlets solid, paniculate inflorescences (up to 6 cm long), stamens fused throughout (or almost so), glands at the base of filaments, anthers partially exserted, and hypanthium glabrous inside, L. vanderwerffii has hollow branchlets, sub-capitate inflorescences (up to 2 cm long), stamens free, no glands at the base of filaments, anthers wholly exserted, and hypanthium densely white tomentose inside.
Figure 17. Inflorescence detail of Licaria vanderwerffii Lorea-Hern. sp. nov. Scale bar 1 mm.
This species is closely related to L. agglomerata; both present small, glomerate inflorescences, ellipsoid to narrowly rhomboid flowers with erect tepals, no staminodes, fully exserted stamens, no glands on filaments, pubescent stamens, and hypanthium tomentose within. However, L. vanderwerffii differs by having stamens free throughout, white tomentum on stamens and (densely on) inner face of hypanthium, and glabrous pistil, against stamens fused by their filaments, brownish-orange tomentum on stamens and upper part of hypanthium inside (sometimes completely glabrous), and puberulent style in L. agglomerata. It is worth to mention that they differ in ecological conditions as well; while L. agglomerata grows in lowland habitats (200-700 m), L. vanderwerffii is a mountain dweller above 1,200 m.
Acknowledgements
I would like to thank the curators of the herbaria cited in the Materials and methods section for allowing the study of specimens of Licaria in their collections. I also thank Jens Rohwer and an anonymous reviewer for their comments and calling my attention to several slips and wrong phrasing in some paragraphs. Eva Piedra assisted me with the preparation of figures. Phil Brewster kindly reviewed the final english version of the manuscript. I thank one of the reviewers who suggested to include the following exsiccata as part of the studied material for several of the species described here, however, it was not possible to see the specimens. So even they correspond to duplicates of the studied material, they are cited here just as a reference for researchers who might need to study them.
L. breedlovei; D. Breedlove 50394, isotype TEX 472525.
L. dolichopoda; B. Hammel et al. 16790, isotypes CR 147811 and CR 2918355; E. Bello & E. Cruz 4262, paratype CR 1596155; G. Herrera 4000, paratype CR 1596509; G. Herrera 4510, paratypes CR 159602, CR 176525 and CR 1561583; J. Marín & G. Marín 499, paratype CR 1596363.
L. gibbitepala; G. Herrera & A. Chacón 2427, isotype CR 1596536.
L. minutiflora; G. Herrera 500, isotype CR 151205; E. Bello 784, paratype CR 1513414; E. Bello 2208, paratype CR 1596135; Q. Jiménez & G Rivera 1011, paratypes CR 1561584 and CR 157780; G. Herrera 2942, paratype CR 1596539.
L. tomentulosa; G. Herrera 2165, isotypes CR 146925 and CR 2921047; G. Herrera 2056, paratypes CR 147034, and CR 2920969; G. Herrera 4115, paratypes CR 1596519, and CR 207025; A. Chacón 25, paratype CR 1596195.
References
Burger, W., & van der Werff, H. (1990). Flora Costaricensis. Family 80 Lauraceae. Fieldiana, Botany, n. s., 23, 1–129.
Gómez-Laurito, J., & Cascante, A. (1999). Licaria caribaea (Lauraceae): a new species from the Caribbean lowlands of Costa Rica. Novon, 9, 199–201. https://doi.org/10.2307/3391798
Gómez-Laurito, J., & Estrada, A. (2002). Licaria leonis (Lauraceae), una nueva especie del Pacífico costarricense, y algunas notas sobre Licaria multinervis H. Kurz. Lankesteriana, 3, 5–9. https://doi.org/10.15517/lank.v2i1.23136
Hammel, B. E. (1986). New species and notes on Lauraceae from the Caribbean lowlands of Costa Rica. Journal of the Arnold Arboretum, 67, 123–136. https://doi.org/10.5962/bhl.part.27389
Kostermans, A. J. G. H. (1937). Revision of the Lauraceae II. The genera Endlicheria, Cryptocarya (American species) and Licaria. Recueil des Travaux Botaniques Néerlandais, 34, 500–609.
Kurz, H. W. (1983). Fortpflanzungsbiologie einiger Gattungen neotropischer Lauraceen und Revision der Gattung Licaria (Lauraceae) (Ph.D. Thesis). University of Hamburg, Germany.
Kurz, H. W. (2000). Revision der Gattung Licaria (Lauraceae). Mitteilungen aus dem Institut für Allgemeine Botanik in Hamburg, 28/29, 89–221.
Mez, C. (1889). Lauraceae americanae. Jahrbuch des Königlichen Botanischen Gartens und des Botanischen Museums zu Berlin, 5, 1–556.
Radford, A. E., Dickison, W. C., Massey, J. R., & Bell, C. R. (1974). Vascular plant systematics. New York: Harper & Row.
Van der Werff, H. (1988). Eight new species and one new combination of Neotropical Lauraceae. Annals of the Missouri Botanical Garden, 75, 402–419. https://doi.org/10.2307/2399431
Van der Werff, H. (2009). Nine new species of Licaria (Lauraceae) from tropical America. Harvard Papers in Botany, 14, 145–159. https://doi.org/10.3100/025.014.0206
Raymundo Cervantes-Barriga a, Joaquín Arroyo-Cabrales b, Alejandro H. Marín-Leyva a, Javier Ponce-Saavedra c, Florent Rivals d, e y Tiberio C. Monterrubio-Ricof, *
a Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Biología, Laboratorio de Paleontología, Edif. R Planta baja, Ciudad Universitaria, Francisco J. Múgica s/n, Col. Felícitas del Río, 58030 Morelia, Michoacán, México
b Instituto Nacional de Antropología e Historia, Subdirección de laboratorio y Apoyo Académico, Laboratorio de Arqueozoología “M. en C. Ticul Álvarez Solórzano”, Moneda Núm. 16, Col. Centro, 06060 Ciudad de México, México
c Universidad Michoacana de San Nicolás de Hidalgo, Laboratorio de Entomología “Biol. Sócrates Cisneros Paz”, Edificio B4 2° piso, Ciudad Universitaria, Francisco J. Múgica s/n, Col. Felícitas del Río, 58030 Morelia, Michoacán, México
d Institut Català de Paleoecologia Humana i Evolució Social, Zona Educacional 4, Campus Sescelades URV (Edifici W3), 43007 Tarragona, España
e Universitat Rovira i Virgili, Departamentd’Història i Història de l’Art, Avinguda 35 de Catalunya 35, 43002 Tarragona, España
f Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Biología, Laboratorio de Vertebrados Terrestres Prioritarios, Edificio “R” 2° Piso, Ciudad Universitaria, Francisco J. Múgica s/n, Col. Felícitas del Río, 58030 Morelia, Michoacán, México
*Autor para correspondencia: tmonter2002@yahoo.com.mx (T.C. Monterrubio-Rico)
Recibido: 22 enero 2024; aceptado: 30 enero 2025
Resumen
Los roedores en México representan 49% de los mamíferos y aunque se asume que las perturbaciones no los afectan en comparación con los mamíferos de mayor tamaño, los estudios al respecto son escasos. Se analizó la comunidad de pequeños roedores en los hábitats terrestres de la cuenca de Cuitzeo, los cuales experimentan distintos grados de antropización. Se recolectaron roedores en pastizal-huizache, matorral-pastizal y bosque de pino-encino durante un año, y se caracterizó la estructura de la comunidad vegetal de cada sitio. Se registraron 17 especies de las familias Cricetidae (15), Heteromyidae (1) y Muridae (1). La riqueza y diversidad especifica fue mayor en el pastizal (H´ = 2.43) y menor en el matorral (H´ = 1.02). Sin embargo, la abundancia fue mayor en el matorral (N = 47) y la menor en el pastizal (N = 21). En época invernal se registró mayor riqueza específica (12) y abundancia (N = 62). Los resultados permiten comprender la dinámica estacional de las comunidades de pequeños roedores en escenarios de perturbación de los hábitats naturales. La cuenca de Cuitzeo todavía alberga una riqueza destacable y se constituye en una región para la conservación de pequeños mamíferos.
Palabras clave: Ensamble de roedores; Diversidad; Hábitat; Estacionalidad
Spatial and seasonal analysis of small rodent communities in the Cuitzeo basin
Abstract
Rodents in México represent 49% of the mammals, and although it’s assumed that they are less affected by disturbance in comparison to the larger-sized mammals, studies on the topic are scarce. We analyzed the small rodent communities in the terrestrial habitats of the Cuitzeo basin, which are experiencing different degrees of anthropogenic disturbance. Rodents were collected in the grassland-huizache, scrub-grassland, and pine-oak forest over a year, and the vegetation community structure was characterized at each sampling site. Seventeen species were identified from the following families: Cricetidae (15), Heteromyidae (1), and Muridae (1). Specific richness and diversity were highest in the grassland (H´ = 2.43) and lowest in the scrub (H´ = 1.02). However, the abundance was highest in the scrub (N = 47) and lowest in the grassland (N = 21). The highest specific richness was recorded during the winter (12) as well as abundance (N = 62). The results increase our understanding of the seasonal dynamic of the small rodent communities in scenarios of habitat disturbance. The Cuitzeo basin still harbors noteworthy richness and constitutes a region for small mammal community conservation.
Las cuencas, como parte de las regiones naturales y libres de la división política del estado, permiten una adecuada comprensión de la diversidad que se presenta (Aguilar et al., 2010). La cuenca de Cuitzeo al noroeste de Michoacán presenta el segundo lago más grande en el interior de México y junto con sus humedales crea un mosaico de hábitats en la región. Históricamente, esta cuenca presentó cobertura de 12 asociaciones vegetales: 6 categorías en sistemas boscosos (bosque de encino, bosque de pino, bosque de pino-encino, bosque de cedro, bosque de oyamel y bosque mesófilo de montaña), 3 tipos de vegetación no boscoso (mezquital, matorral subtropical y pastizal) y 3 asociados a humedales y cuerpos de aguas (bosque de galería, tular y carrizal) (Madrigal y Guridi, 2009). Sin embargo, la pérdida de vegetación en la cuenca ha aumentado y en la actualidad 55% de la superficie presenta zonas agropecuarias y asentamientos humanos (Bravo et al., 2008; Correa et al., 2014; INEGI, 2023). Con respecto a la fauna, se ha registrado riqueza importante en diversos grupos como insectos y arácnidos (Ponce y Quijano, 2010), peces (Medina y Ortega, 2010), anfibios y reptiles (García y Flores, 2010), aves (Pérez et al., 2002; Villaseñor y Villaseñor, 2010), y más recientemente de mamíferos (Monterrubio et al., 2019); destacando en este último grupo los roedores por su alta riqueza específica.
Por su relevancia ecológica al formar la base de redes tróficas y por sus servicios ambientales como la depredación de semillas, así como rasgos de sus historias de vida como sus altas tasas de natalidad, variación y tolerancia en sus distribuciones y, en general, una notable adaptación a modificaciones al ambiente (Ceballos y Oliva, 2005), hace de los roedores un taxón que permite estudiar cómo varía la estructura local de sus comunidades en función de las características estructurales de la vegetación donde habitan. El entendimiento de estos procesos es fundamental para examinar la respuesta en los ensambles y comunidades de mamíferos ante el escenario cambiante de la cuenca de Cuitzeo, región considerada prioritaria para la conservación de biodiversidad, que experimenta alto impacto humano y efectos del cambio climático (Aguilar et al., 2010; Hernández et al., 2023).
En algunos estados de México como Durango, por ejemplo, Villanueva et al. (2017) evidenciaron cómo 6 especies se asocian a un gradiente de vegetación que va desde zonas boscosas hasta áreas de matorral y pastizal. Peromyscus difficilis y Sigmodon ochrognathus se asociaron con mayor frecuencia a bosques, en cambio P. boylii y P. pectoralis a zonas áridas. Por su parte, P. hooperi y Neotoma leucodon resultaron más tolerantes y se presentan a lo largo del gradiente. En Veracruz se estudió el gradiente de borde al interior de un bosque mesófilo de montaña para observar la zonación por parte de 9 especies de roedores y 1 marsupial. Aunque la riqueza específica fue similar a lo largo de los sitios de estudio, se observó que la distribución es significativa con relación a las tallas, roedores de tallas similares se evitan y roedores de diferentes tallas coexistían en los mismos sitios muestreados (González et al., 2012).
Por otro lado, la distribución y el uso del hábitat por los pequeños roedores también ocurre a nivel específico, ya sea por edad o por sexo. Neotomodon alstoni, por ejemplo, se ha estudiado en 5 hábitats del Ajusco (zacatonal, pradera, bosque, ecotono bosque-zacatonal y ecotono bosque-pradera) y se demostró que las hembras prefieren el zacatonal, y los machos los bosques y ecotonos; las hembras buscan sitios de mejor calidad que los machos y estos últimos muestran patrones de distribución más en relación con la densidad de hembras que con la calidad del microhábitat (Rojas et al., 2012).
Para Michoacán, un análisis sobre la distribución de 48 especies de roedores en las regiones fisiográficas del estado registró que cada región muestra diferencias significativas en relación con la abundancia de roedores, siendo el Cinturón Volcánico Transmexicano el de mayor diversidad, seguida del Altiplano Mexicano, destacándose que las 2 provincias son las de mayor diversidad, a pesar del alto número de asentamientos humanos en ellas (Sánchez et al., 2005).
El objetivo general del presente estudio fue evaluar la riqueza y diversidad de roedores en 3 hábitats: pastizal, matorral subtropical y bosque de pino-encino en la cuenca de Cuitzeo, examinando su variación entre épocas del año, además de caracterizar la vegetación de los hábitats.
Materiales y métodos
La cuenca de Cuitzeo se ubica principalmente en el centro-norte de Michoacán, compartiendo una porción menor del extremo norte de la cuenca con el estado de Guanajuato. Se sitúa en un intervalo de elevación de 1,750 a 2,359 m snm y presenta 3,675 km² de superficie (Bravo et al., 2008). Actualmente, 6% de su superficie es ocupada por asentamientos humanos y 8% por cuerpos de agua. Los campos de cultivo, pastos inducidos y plantaciones forestales representan 51%. Los tipos de vegetación de la cuenca seleccionados en este estudio representan 1% para pastizales naturales, 14% matorrales y 20% bosques abiertos y cerrados (Correa et al., 2014). La cuenca presente clima templado en 3 subgrupos: semifrío (en partes altas de las montañas del sur), semicálido (a los alrededores del lago de Cuitzeo) y subhúmedo (en la mayor parte de la cuenca). La temperatura media anual va de los 14 a los 17 °C y en las partes altas de la cuenca puede ser de 10 °C. La precipitación en las partes altas oscila entre 1,200 a 1,500 mm; en la parte media y baja entre 800 y 1,000 mm (INEGI, 2023).
Los sitios seleccionados para el muestreo fueron: al norte, el área a menor elevación, donde predomina el pastizal, el sitio se conoce como “La Cinta” (1,838 m snm). A elevación intermedia y en el centro de la cuenca, el sitio “Misión del Valle”, localizado en una zona suburbana con vegetación que presenta ecotonos de extensos matorrales y pastizales (1,896 m snm), y al sur, un sitio de bosque con vegetación de pino-encino conocido como “La Planta” (2,132 m snm) (fig. 1).
De acuerdo con información de temperatura y precipitación de localidades cercanas a las de este estudio, las temperaturas más altas en la región se presentan principalmente en los meses de abril y mayo (temporada de secas); mientras que los meses con mayores precipitaciones (temporada de húmedas) se concentran en julio, agosto y septiembre. El resto del año presenta bajas temperaturas y precipitaciones (temporada de secas frías) (Carlón y Mendoza, 2007).
Se efectuaron 4 salidas de campo durante un ciclo anual, las 3 primeras en 2021 y la última en 2022. Los roedores se capturaron con trampas Sherman. El primer muestreo (secas) inició del 30 de abril al 11 de mayo con esfuerzo de 60 trampas (540 noches/trampa), sin embargo, dada la baja captura se decidió aumentar un día más de recolecta en las siguientes temporadas. El segundo muestreo (lluvias) fue del 30 de agosto al 13 de septiembre con un esfuerzo 720 noches/trampa. Durante la temporada de secas frías se llevaron a cabo 2 salidas; una posterior a la temporada de lluvias (post-lluvias) durante el 26 de noviembre a 10 de diciembre con 70 trampas (840 noches/trampa), no obstante, las capturas aun seguían siendo pocas por lo que se hizo un último muestreo (secas frías), del 27 de enero al 10 de febrero de 2022 con 90 trampas (1,068 noches/trampa) con el fin de lograr tener un inventario más completo. El esfuerzo total acumulado en el estudio fue de 3,168 noches/trampa. La dispersión espacial del sistema de trampeo incluyó líneas de trampas con separación de 10 m, las cuales se colocaron antes de la puesta del sol y se revisaron al día siguiente por la mañana. El cebo consistió de una mezcla de maíz triturado y esencia de vainilla, el cual se encapsuló en una rejilla metálica de 3 cm³ colocada dentro de la trampa. Los roedores capturados se sacrificaron con pentobarbital sódico, siguiendo los lineamientos de la Sociedad Americana de Mamíferos (Sikes, 2016) y las recomendaciones convencionales de recolección de datos sugeridas por Romero et al. (2007).
Los resultados presentados en este artículo forman parte de un proyecto en el que también se analiza la alimentación y su efecto sobre la dentición de los roedores, por lo que fue necesario sacrificar a los ejemplares. Los roedores recolectados se depositaron para resguardo en el Laboratorio de Mastozoología de la Universidad Michoacana de San Nicolás de Hidalgo (UMSNH), donde como parte de su identificación se comparó con ejemplares existentes en la colección, además del uso de claves (Godinez y Guerrero, 2014; Mammal Diversity Database, 2024; Núñez y Pastrana, 1990). Se utilizó el catálogo nomenclatural de Ramírez et al. (2014) y se atendieron las modificaciones recientes para Peromyscus maniculatus (ahora P. labecula), P. boylii (ahora P. kilpatricki) y P. melanophrys (ahora P. zamorae) (Bradley et al., 2017, 2019; López et al., 2019). La recolecta científica se efectuó al amparo del permiso especial clave SGPA/DGVS/02243/22.
Figura 1. Localización de la cuenca de Cuitzeo, los límites se muestran en tono claro azul claro. En azul se indica el lago de Cuitzeo. Hábitats de estudio, A: pastizal-huizache, B: matorral-pastizal, C: bosque de pino-encino. CVTM: Cinturón Volcánico Transmexicano. Mapa elaborado por Alejandro H. Marín-Leyva.
La caracterización de la vegetación se hizo en el estrato herbáceo mediante 5 cuadrantes de 5 m², cada cuadrante con 5 unidades de muestreo de 1 m² (25 unidades en total, por cada sitio y para cada estación), distribuidas en arreglo “5 de oros”, una en el centro y las demás en las esquinas (Senasica, 2024). Se identificaron familias y géneros, y se registró su abundancia y cobertura horizontal en porcentaje. La determinación taxonómica de las plantas se limitó a género debido al estado fenológico de las plantas en algunas estaciones, se utilizó el término morfotipo para las plantas solo identificadas hasta familia.
Para la vegetación se describió la estructura general en los niveles taxonómicos de familia y género para cada sitio y época del año incluyendo composición y abundancias.
Para los roedores se analizó la riqueza y abundancia para cada localidad, época de recolecta y para todo el año. Para evaluar la relación entre la riqueza de especies en función del esfuerzo de muestreo, se elaboraron curvas de rarefacción para cada sitio que permitieron estimar la eficiencia del muestreo utilizando el acumulado por individuos (Chao et al., 2014, 2016). Posteriormente, se estimó la completitud del muestreo a través de las estimaciones no paramétricas de riqueza de Chao 2 y bootstrap. Se elaboraron curvas de rango-abundancia para describir las relaciones de abundancia entre las especies y determinar las dominancias para cada localidad y época del año.
Se calcularon los índices de diversidad de Shannon-Wiener (H´) y dominancia Simpson (D), tanto por sitio de estudio como por temporada. Se utilizó la prueba de t modificada por Hutchenson (1970) para probar las diferencias estadísticas, se incluyó además el cálculo de los límites de confianza (LC) a 95% por el método de percentiles. Para calcular la equitatividad de las especies se usó el índice de equitatividad de Pielou (J). Para conocer el grado de semejanza entre comunidades se usó el índice de similitud de Jaccard para composición de especies y el índice de Bray-Curtis para considerar la abundancia en la comparación (Magurran, 2004).
Con el fin de evaluar posibles diferencias en la composición de especies en relación con los hábitats, se aplicó un análisis de las similitudes entre comunidades (Anosim), con 10,000 permutaciones y disimilitud de Bray-Curtis. Todos los análisis antes mencionados se hicieron con el Software Past 4.12 (Hamer et al., 2001).
Resultados
La comunidad vegetal durante un ciclo anual se caracterizó por presentar 31 familias y 105 géneros de plantas en una muestra de 15,060 individuos del estrato herbáceo, de los que 51% correspondió a la asociación pastizal-huizache. El 39% se registró para matorral-pastizal y 10% en pino-encino.
En la asociación pastizal-huizache predominó la familia Poaceae, con abundancia relativa (AR) de 80% y cobertura horizontal relativa (CH) de 86%. Destacan los géneros Chloris (AR = 44%; CH = 51%), Paspalum (AR = 12%; CH = 3%) y Distichlis (AR = 9%; CH = 22%), esas proporciones fueron constantes durante todo el año. En el estrato arbóreo destaca el huizache (Vachellia sp.).
Tabla 1
Especies registradas y sus abundancias (absoluta y relativa = tasas de captura) en 3 asociaciones vegetales de la cuenca de Cuitzeo en un ciclo de muestreo anual. S = Secas; H = húmedas; PLL = posterior a lluvias; SF = secas frías.
Especie
Pastizal-huizache (1,838 m snm)
Matorral-pastizal (1,896 m snm)
Pino-encino (2,132 m snm)
Subtotales (tasa de captura)
Heteromys irroratus
0
2
0
0
0
1
0
7
0
0
0
0
10 (0.31)
Baiomys taylori
0
1
0
3
1
1
3
28
0
0
0
0
37 (1.16)
Baiomys musculus
1
0
0
0
1
0
0
0
0
0
0
0
2 (0.063)
Sigmodon hispidus
0
0
0
2
0
0
0
0
0
0
0
0
2 (0.063)
Sigmodon mascotensis
0
0
0
1
0
0
0
0
0
0
0
0
1(0.031)
Oligoryzomys fulvescens
0
1
0
0
0
0
0
0
1
0
0
0
2 (0.063)
Peromyscus labecula
0
0
0
0
0
0
0
0
0
0
1
0
1(0.031)
Peromyscus melanotis
0
0
0
0
0
0
0
0
0
0
0
1
1(0.031)
Peromyscus difficilis
0
1
0
0
0
0
0
1
0
1
4
5
12 (0.37)
Peromyscus hylocetes
0
0
0
1
0
0
0
0
0
0
0
1
2 (0.063)
Peromyscus kilpatricki
0
0
0
1
0
0
0
0
0
0
0
3
4 (0.12)
Peromyscus pectoralis
0
2
0
0
0
0
0
0
0
0
0
0
2 (0.063)
Peromyscus gratus
0
0
0
0
0
0
0
0
0
0
3
1
4 (0.12)
Peromyscus zamorae
0
0
0
0
0
0
0
0
0
0
2
0
2 (0.063)
Reithrodontomys sumichrasti
0
0
0
0
0
0
0
0
1
0
0
5
6 (0.18)
Reithrodontomys fulvescens
0
0
0
0
0
0
0
1
0
0
1
0
2 (0.063)
Mus musculus
0
3
1
1
2
0
1
0
0
0
0
0
8 (0.25)
Submuestra por sitio (%)
21 (21.4)
47 (47.9)
30 (30.6)
98 (100%)
Riqueza de especies por sitio
11
6
10
17
Tasa de captura/área
1.98
4.45
2.84
3.09
Especies exclusivas por sitio
3
0
5
8
Tabla 2
Parámetros de riqueza, abundancia (en paréntesis) y tasa de captura de la comunidad de roedores en 3 asociaciones vegetales en función a la época.
Secas
Húmedas
Posterior a lluvias
Secas frías
Pastizal-huizache
1-(1)-0.56
6-(10)-4.1
1-(1)-0.3
6-(9)-2.5
Matorral-pastizal
3-(4)-2.2
2-(2)-0.83
2-(4)-1.4
4-(37)-10.4
Pino-encino
2-(2)-1.1
1-(1)-0.42
5-(11)-3.9
6-(16)-4.5
Esfuerzo de muestreo/trampas-noche
540
720
840
1,068
Especies
5
6
7
12
Especies exclusivas
1
1
2
6
Sub muestra estacional (%)
7 (7.1)
13 (13.2)
16 (16.3)
62 (63.2)
Tasas de captura general/estación
1.30
1.81
1.90
5.81
El matorral-pastizal está integrado proporcionalmente por las familias Asteraceae (18%), Poaceae (16%), Fabaceae (14%) y Euphorbiaceae (11%). Difiere del pastizal al presentar variación estacional en la estructura y composición de las plantas en el año. Por ejemplo, en secas Lolium presentó la mayor abundancia y cobertura horizontal (AR = 43%; CH = 32%); en temporada húmeda Marina (AR = 29%; CH = 21%), Euphorbia (AR = 22%; CH = 13%) y Adenophyllum (AR = 20%; CH = 19%); en la temporada posterior a lluvias Simsia (AR = 20%; CH = 20%) y Eragrostis (AR = 16%; CH = 16%), y en secas frías, una especie de gramínea (AR = 42%; CH = 55%), que por su fenología no pudo ser identificada. También se presentaron elementos aislados de Opuntia.
Figura 2. A, Curvas de rarefacción de riqueza de especies en función de las abundancias de roedores en sitios de colecta; B, curva de rarefacción con riqueza de especies en función abundancias de roedores colectadas durante el ciclo anual en los 3 sitios.
La vegetación en el área de pino-encino se caracterizó en el estrato arbóreo por la presencia de Pinus y Quercus. El estrato herbáceo se caracterizó por el predominio de las familias Poaceae, Rosaceae, Asteraceae, Oxiladaceae, Fabaceae, Anacardiaceae, Pteridaceae y Cyperaceae con 78% de la muestra total. El género Rubus (AR = 29%; CH = 13%) predominó en secas, mientras que en húmedas fue Oxalis (AR = 31%; CH = 9%) y Centella (AR = 19%; CH = 17%); en la temporada post-lluvias (AR = 19%; CH = 11%) y secas frías (AR = 27%; CH = 13%) fueron macollos de gramíneas que carecían de espiga por lo que no se identificaron, posiblemente del género Microstegium (apéndice 1).
La muestra de roedores capturados incluyó 98 individuos. Los ejemplares se identificaron como 16 especies nativas y Mus musculus (introducida), pertenecientes a 7 géneros en las familias Cricetidae (15), Heteromyidae (1) y Muridae (1) (tabla 1).
Con base en la curva de rarefacción, se interpreta el esfuerzo de muestreo como suficiente en registrar la riqueza esperada de los 3 sitios; con una cobertura de la muestra de 93% en matorral-pastizal, 83% en el bosque de pino-encino y 72% en pastizal-huizache (fig. 2A). A nivel regional, según los estimadores Chao 2 y bootstrap, la riqueza se espera entre 18-19 especies, solo 2 por encima de la riqueza observada, lo que permite asumir un muestreo de inventario de especies de roedores para la zona suficientemente completo, y que se corrobora con una cobertura de muestra total de 97% en la curva de rarefacción (fig. 2B). Siendo en el pastizal-huizache y en el bosque de pino-encino donde existe mayor oportunidad de registrar otras especies.
La mayor riqueza de especies se observó en el pastizal-huizache, seguido de la asociación de pino-encino y la asociación matorral-pastizal, y en este mismo orden las abundancias se comportaron de menor a mayor (tabla 1; fig. 3A). En relación con la temporada de recolecta, la riqueza y abundancia en secas es en la que se obtuvo la menor cantidad de ejemplares. Posteriormente, se incrementó el éxito de captura para las temporadas siguientes hasta llegar a secas frías (tabla 2); sumando los datos de riqueza y abundancia de las 3 comunidades se observa un incremento en la complejidad (fig. 3B). En las temporadas invernales (secas frías) se destaca la mayor abundancia y tasas de captura de todo el estudio, particularmente en el matorral-pastizal. También se registró la mayor riqueza de especies (n = 12), la mayor abundancia absoluta (62 individuos) y la mayor tasa de captura de 5.8/100 trampas-noche (tabla 2).
Figura 3. A, Curvas de rango-abundancia entre los sitios de estudio; B, curvas de rango-abundancia entre las temporadas del año de estudio sumando los 3 hábitats.
Baiomys taylori fue la especie más abundante, representó 33% del total de individuos recolectados, con los valores más altos en la temporada de secas frías, se presentó durante todo el año principalmente en matorral-pastizal (n = 33), seguido de pastizal-huizache (n = 4). La especie que resultó con mayor abundancia en el hábitat de pastizal-huizache fue Mus musculus, que también se registró en matorral-pastizal y presente durante todo el año. Para el bosque de pino-encino las especies con más individuos fueron Peromyscus difficilis y Reithrodontomys sumichrasti, la primera se encontró todo el año exceptuando en secas, además destaca por ser la única registrada para las 3 localidades. Por el contrario, las especies Peromyscus labecula, Peromyscus melanotis, y Sigmodon mascotensis, estuvieron representadas por solo 1 individuo; mientras que Baiomys musculus, Oligoryzomys fulvescens, Peromyscus hylocetes, Peromyscus zamorae, Peromyscus pectoralis, Reithrodontomys fulvescens y Sigmodon hispidus contaron con 2 ejemplares, estas especies en conjunto representan 59% de la riqueza total del estudio (fig. 3; tabla 1).
Tabla 3
Valores con el índice de diversidad de Shannon-Wiener (H´), índice dominancia de Simpson (D) e índice de equidad de Pielou (J) para la comunidad de roedores de la cuenca de Cuitzeo. Prueba de t modificada por Hutchenson H´/D para diferencias significativas (*).
Prueba de t modificada por Hutchenson
H´
D
J
Matorral-pastizal
Pastizal-huizache
Pino-encino
Matorral-pastizal
0.97
0.52
0.54
D = 3.8822E-05*
D = 0.00042*
Pastizal-huizache
2.19
0.13
0.91
H´ = 3.035E-06*
D = 0.3657
Pino-encino
1.93
0.18
0.84
H´ = 6.886E-05*
H´ = 0.276
Mediante el índice de diversidad de Shannon-Wiener (H´) y la equitatividad de Pielou (J), con límites de confianza (LC) de 95%, se concluye que el hábitat que presentó mayor diversidad y equitatividad en la comunidad de pequeños roedores fue el pastizal-huizache con H´ = 2.19 (LC = 1.69-2.29) y J = 0.91 (LC = 0.81-0.96), en cambio matorral-pastizal fue poco diverso con H´ = 0.97 (LC = 0.66-1.22) y su equitatividad fue las más baja con J = 0.54 (LC = 0.42-0.70); el bosque de pino-encino presento una diversidad de H´ = 1.93 (LC = 1.68-2.13) y J = 0.84 (LC = 0.74-0.92). La prueba de t modificada por Hutchenson no muestra diferencias significativas entre pastizal-huizache y pino-encino, pero sí entre matorral-pastizal con respecto a las otras 2 comunidades. Este efecto también se observa con el cálculo de los límites de confianza en donde solamente en pastizal-huizache y pino-encino existe traslape tanto en H´ como en J. En relación con la temporada de recolecta y como se observa en la tabla 1, en algunos casos solo se capturaron 1 o 2 ejemplares por sitio y temporada, por tanto, conocer la diversidad de los sitios en función de las temporadas no es posible. Aun así, si se toma la sumatoria de los 3 hábitats por época de recolecta se observa poca variación de la diversidad: secas, H´ = 1.55 (LC = 0.79-1.55); húmedas H´ = 1.73 (LC = 1.26-1.73); post lluvias, H´ = 1.84 (LC = 1.35-1.87) y finalmente secas frías, H´ = 1.75 (LC = 1.41-1.99), sin diferencias significativas entre temporadas del año y traslape de los intervalos de confianza en todos los casos.
Por su parte, con el índice de dominancia de Simpson (D) el valor más alto se registró en matorral-pastizal con D = 0.52 y su dominancia con la especie B. taylori es diferente a los otros 2 sitios, en donde no se observaron especies dominantes (tabla 3).
De acuerdo con el índice de similitud de Jaccard para la composición de la comunidad de roedores, el matorral-pastizal y pastizal-huizaches presentan mayor similitud, a diferencia de la comparación entre matorral-pastizal y bosque pino-encino con menor similitud; considerando las abundancias, este patrón se repite según el índice de Bray-Curtis (tabla 4). Con el análisis de similitud entre comunidades (Anosim) se estima diferencias significativas (R = 0.35; p = 0.008) en la composición y abundancia entre los hábitats matorral-pastizal y bosque de pino-encino (p = 0.030) (fig. 4).
Tabla 4
Índice de similitud de Jaccard (ISJ) y Bray-Curtis (BC) para la comunidad de roedores en las 3 asociaciones vegetales examinadas de la cuenca de Cuitzeo.
Matorral-pastizal
Pastizal-huizache
Pino-encino
Matorral-pastizal
1
BC = 0.32
BC = 0.05
Pastizal-huizache
ISJ = 0.41
1
BC = 0.15
Pino-encino
ISJ = 0.14
ISJ = 0.23
1
Figura 4. Boxplot y valores de probabilidad del análisis de similitudes (Anosim) entre las 3 asociaciones vegetales para la comunidad de roedores en de la cuenca de Cuitzeo (R = 0.35; p = 0.008). M-P = Matorral-pastizal, P-E = bosque de pino-encino y P-H = pastizal-huizache.
Discusión
Los resultados obtenidos ofrecen 4 aspectos emergentes: riqueza de especies, baja abundancia general, mayores abundancias invernales y baja similitud relativa entre los ambientes muestreados. Recientemente, se evaluó la mastofauna que habita en la cuenca de Cuitzeo. En total se registraron 68 especies de mamíferos, con 28 del orden Rodentia, que representaron 54% de las especies registradas en Michoacán (Monterrubio et al., 2019). Con el presente trabajo se suman al registro de Rodentia para la región a B. musculus, P. pectoralis y O. fulvescens, especies que están presentes en otras áreas del eje Neovolcánico y el bajío (Sánchez et al., 2005), pero que no habían sido registradas para la cuenca. Al incluir estos nuevos registros para la cuenca, en esta zona se presenta 60% de las especies de roedores documentadas para Michoacán (Monterrubio et al., 2014). Otro aspecto notable, es que el esfuerzo de muestreo desarrollado proporcionó el registro del mayor número de especies (16) en cualquier estudio efectuado para la cuenca (Apéndice 2). Esta riqueza sobresale también en el contexto nacional, ya que los intervalos de riqueza específica reportados suelen estar entre 5 y 18 especies (Apéndice 3), generalmente se observan valores bajos en áreas pequeñas y aumentan cuando el estudio es a escala regional.
En contraste, fue sorpresiva la baja abundancia general, con solo 98 registros, en comparación con otros estudios de diferentes regiones del país, en los que con esfuerzos entre 455 y 28,242 noches/trampa se obtienen capturas superiores a 100 individuos (Apéndice 3). Una razón probable de la baja abundancia para la mayoría de especies puede ser la disminución de las precipitaciones en la región. Con base en los datos históricos y comparando con los últimos años, se ha experimentado sequía en algunas partes de la cuenca (Hernández et al., 2023), para lo que probablemente hayan respondido las poblaciones de roedores con menor reproducción o migración. Otra evidencia es la cifra de capturas del muestreo de la temporada de secas frías (fig. 3B) y posterior a la época de lluvia de la cuenca, dado que se registró un incremento notable en el número de capturas, tanto en riqueza específica como en abundancia. Este fenómeno es similar al observado en otros estudios (Flores y Vázquez, 2016; Zaragoza et al., 2022). El aumento poblacional en esta época ocurrió para B. taylori, P. difficilis y H. irroratus; en las primeras 2 especies ya se ha registrado aumento en densidades poblacionales en temporadas frías del año (Eshelman y Cameron, 1987; Fernández et al., 2010), mientras que el aumento de H. irroratus, según antecedentes, se reporta de agosto a noviembre (Dowler y Genoways, 1978).
El índice de diversidad no muestra variación significativa entre épocas del año, esto podría indicar que la diversidad es explicada en mayor medida por el hábitat y que genera niveles importantes de estabilidad reflejada en los valores de equitatividad obtenidos (entre 0.57 y 1), lo cual también sugieren Luévano et al. (2008), particularmente en zonas de matorrales.
Las 3 comunidades de roedores examinadas muestran en general baja similitud, y parece observarse una posible influencia de la estratificación de la vegetación. En general, a mayor complejidad estructural de la vegetación con presencia del estrato arbóreo, se observa mayor riqueza de especies. Este patrón se presentó para el pastizal-huizache y el bosque de pino-encino en donde no existió diferencia significativa de diversidad y tampoco hubo traslape de intervalos de confianza. En cambio, la abundancia de roedores fue mayor en el matorral-pastizal y en el pino-encino, en ambos hábitats se observa mayor riqueza de la vegetación a nivel herbáceo. El patrón de aumento en las abundancias para ambientes de mayor heterogeneidad también lo encontraron Morales et al. (2019) y contrasta con la homogeneidad del estrato herbáceo y baja densidad de roedores registrada para el pastizal-huizache de este estudio. Todavía no es clara la relación entre la estructura de la comunidad de roedores y la conectividad de los hábitats. La dinámica de la cuenca de Cuitzeo implica un aumento de la colonización humana y más actividades agropecuarias en las últimas décadas (Escamilla y Aguilar, 2010). Dicha expansión ha degradado y fragmentado los hábitats originarios, hasta llegar a tener 6% con asentamientos humanos y 50% de la cuenca destinado a zonas de cultivo y pastoreo (INEGI, 2023). Con este panorama, es posible que la estructura y composición de las comunidades de pequeños roedores observada puede deberse al grado de aislamiento diferenciado entre los distintos hábitats. Los pastizales presentan conectividad con los extensos cultivos, permitiendo la cohesión espacial de algunas especies, mientras que las zonas de matorral y arbóreas presentan índices muy bajos de conectividad (Correa et al., 2014). El aislamiento en estos hábitats, principalmente matorrales, puede disminuir a los depredadores, lo que se ve reflejado en el aumento de poblaciones de roedores más generalistas y menor riqueza especifica (Young et al., 2015); este proceso puede estar presente en el sitio de matorral-pastizal. Aunque en el caso de las zonas montañosas de la cuenca con bosques, todavía parece existir mayor conectividad y esta se ve reflejada en la riqueza de la mastofauna (Monterrubio et al., 2019).
El pastizal-huizache se extendía originalmente en forma extensa (López et al., 2010), ahora es un ambiente con elevada fragmentación y es quizás uno de los hábitats más amenazados por el elevado establecimiento de asentamientos humanos y campos de cultivo. Sin embargo, la diversidad de roedores observada en este hábitat parece todavía estar explicada por la vegetación más que por la antropización vecina. Los 3 géneros de plantas herbáceas más representativos de la zona (Chloris, Distichlis y Panicum) son gramíneas no invasivas y componentes de la vegetación original (Herrera y Pámanes, 2010). Peromyscus pectoralis, Sigmodon hispidus y S. mascotensis resultaron exclusivas a esta localidad entre los 3 hábitats estudiados, y parecen coincidir con ambientes en que se han registrado regularmente, que incluyen aridez relativa, pastizales y ambientes rocosos (Baccus et al., 2009; Fleharty y Olson, 1969; Martínez et al., 2017). En hábitats similares la exclusividad reportada suele ser de 1 a 2 especies (Cimé et al., 2010; Elizalde et al., 2014; Hernández et al., 2012), lo cual coincide con lo registrado en este trabajo. Además, la cercanía con sembradíos parece proveer de alimento, lo que facilita la coexistencia entre especies, ya que los roedores también consumen materiales de los cultivos, como se ha documentado en plantaciones de caña (Peña et al., 2009).
Para el matorral-pastizal, la baja diversidad de roedores podría explicarse en parte por la poca conectividad y por la biología de la comunidad vegetal, ya que los géneros más abundantes están asociados con ambientes alterados. En temporada seca el género herbáceo Lolium (Poaceae), especie introducida en México (Herrera y Pámanes, 2010), fue la más abundante y fue en esta temporada cuando se recolectaron pocos roedores (n = 4), 2 de los cuales fueron M. musculus, especie que puede considerarse indicadora de deterioro del hábitat. En relación con las plantas de temporadas restantes del año, son géneros nativos con distribución principalmente en matorrales como Marina (M. cf. nutans) y Adenophyllum, y junto con Euphorbia (E. cf. dentata), Simsia (S. cf. foetida) y Eragrostis (E. cf. intermedia), pero crecen en afinidad a lugares ruderales con tendencia a disturbios (Rzedowski y Rzedowski, 2005). En los recorridos de campo se observaron asentamientos humanos e infraestructura inconclusa, indicativa de la expansión de la mancha urbana, así como fauna doméstica, perros principalmente, caminando en búsqueda de alimento entre la vegetación.
El registro de B. taylori, especie que se conoce con una mayor densidad de individuos distribuyéndose en matorrales y zonas áridas (Packard, 1960), coincidió con el hábitat de recolecta. Este cricétido junto con H. irroratus podrían ilustrar su resiliencia a los ambientes perturbados, tendencia que se ha reportado para mamíferos de tallas pequeñas, particularmente en roedores y murciélagos (Dirzo et al., 2014), aunque también pudiesen ser evidencia de que el hábitat de matorral-pastizal en la región experimenta un proceso de homogeneización de la comunidad de roedores.
Por su parte, el bosque de pino-encino (La Planta) solo se diferencia con el pastizal-huizache por tener una especie menos (la invasora M. musculus); pero en índices cuantitativos la similitud fue baja. Este hábitat, donde cobertura y dominancia de biomasa vegetal corresponden a Pinus y Quercus (Gómez-Tagle et al., 2015), no se recolectaron especies de roedores introducidas. O. fulvescens, R. sumichrasti y las especies del género Peromyscus se reportan en bosques conservados y áreas con corrientes de agua cercanas (Fernández et al., 2010; Spencer y Cameron, 1982), lo cual coincide en lo general con este estudio. Sin embargo, aunque parece un espacio conservado por la estructura de su comunidad de roedores y su flora, también presenta cambio en el uso de suelo en los alrededores, principalmente por expansión del cultivo del aguacate y arándano. Aquí se presentaron 5 especies exclusivas, y coincide con lo registrado en otros estudios en ambientes semejantes en los que la exclusividad de especies va de 1-4 (Flores y Vázquez, 2016; Mendoza y Horváth, 2013; Villanueva, et al., 2017; Zalapa et al., 2012).
Es importante continuar con muestreos con el fin de conocer mejor la dinámica estacional y espacial de las poblaciones de roedores en los distintos ambientes, ya que cada hábitat difiere de nivel de fragmentación y extensión en la cuenca, sobre todo en esta región considerada de importancia para la conservación de la biodiversidad en todos los niveles (Aguilar et al., 2010). Finalmente, en el centro de México desaparecen rápidamente los hábitats naturales, es urgente utilizar la información generada por múltiples inventarios para la priorización y establecimiento de áreas a restaurar y para conservar espacios representativos de todos los hábitats, especialmente donde habitan especies endémicas como lo es Peromyscus ensinki (Bradley et al., 2021). En la cuenca debe estudiarse el grado de tolerancia que especies listadas en categorías de riesgo tienen a la perturbación y como coexisten con especies introducidas, anticipando y examinando los riesgos potenciales de enfermedades zoonóticas, sin olvidar que el bienestar económico no puede lograrse en un entorno sin servicios ambientales y biodiversidad (Villafán et al., 2021; Young et al., 2014).
Referencias
Aguilar, V., Kolb, M., Koleff, P. y Urquiza-Haas, T. (2010). Las cuencas de México y su biodiversidad: una visión integral de las prioridades de conservación. En H. Cotler, A. Garrido, N. Luna, C. Enríquez y M. L. Cuevas (Eds.), Las cuencas hidrográficas de México. Diagnóstico y priorización (pp. 142–151). Ciudad de México: Instituto Nacional de Ecología.
Aragón, E. E., Garza, A. y Cervantes, F. A. (2009). Estructura y organización de los ensambles de roedores de un bosque de la Sierra Madre Occidental, Durango, México. Revista Chilena de Historia Natural, 82, 523–542. http://dx.doi.org/10.4067/S0716-078X2009000400007
Baccus, J. T., Hardwick, J. M., Huffman, D. G. y Kainer, M. A. (2009). Seasonal trophic ecology of the white-ankled mouse, Peromyscus pectoralis (Rodentia: Muridae) in central Texas. Texas Journal of Science, 61, 97–118.
Barragán, F., Lorenzo, C., Morón, A., Briones, M. A. y López, S. (2010). Bat and rodent diversity in a fragmented landscape on the Isthmus of Tehuantepec, Oaxaca, México. Tropical Conservation Science, 3, 1–16. https://doi.org/10. 1177/194008291000300101
Bradley, R. D., Francis, J. Q., Platt, R. N., Soniat, T. J., Álvarez, D. y Lindsey, L. L. (2019). Mitochondrial DNA sequence data indicate evidence for multiple species within Peromyscus maniculatus. Lubbock, TX: Museum of Texas Tech University.
Bradley, R. D., Ordóñez, N., Ceballos, G., Rogers, D. S. y Schmidly, D. J. (2017). A new species in the Peromyscus boylii species group (Cricetidae: Neotominae) from Michoacán, México. Journal of Mammalogy, 98, 154–165. https://doi.org/10.1093/jmammal/gyw160
Bradley, R. D., Ordóñez, N., Thompson, C. W., Wright, E. A., Ceballos, G., Kilpatrick, C. W. et al. (2021). Two new species of Peromyscus (Cricetidae: Neotominae) from the Transverse Volcanic Belt of México. Journal of Mammalogy, 103, 255–274. https://doi.org/10.1093/jmammal/gyab128
Bravo-Espinosa, M., García-Oliva, F., Ríos-Patrón, E., Mendoza-Cantú, M. E., Barrera-Camacho, G., López-Granados, E. et al. (2008). La cuenca del lago de Cuitzeo: problemática, perspectivas y retos hacia su desarrollo sostenible. Morelia Michoacán, México: Consejo Estatal de Ciencia y Tecnología de Michoacán (COECYT). https://doi.org/10.22201/ciga.9789707035782p.2008
Carlón, T. y Mendoza, M. (2007). Análisis hidrometeorológico de las estaciones de la cuenca del lago de Cuitzeo. Investigaciones Geográficas, Boletín del Instituto de Geografía, UNAM, 63, 56–76. https://doi.org/10.14350/rig.29910
Castro, A., Salame, A., Vergara, J., Castillo, A. y Ramírez, J. (2008). Fluctuaciones de micromamíferos terrestres en bosques templados aledaños a la Ciudad de México, Distrito Federal. En C. Lorenzo, E. Espinoza y J. Ortega (Eds.), Avances en el estudio de los mamíferos de México II (pp. 391–410). San Cristóbal de las Casas, Chiapas: Asociación Mexicana de Mastozoología.
Ceballos, G. y Oliva, G. (2005). Los mamíferos silvestres de México. México D.F.: Conabio/ Fondo de Cultura Económica.
Chao, A., Gotelli, N. J., Hsieh, T. C., Sander, E. L., Ma, K. H., Colwell, R. K. et al. (2014). Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecological Monographs, 84, 45–67. https://doi.org/10.1890/13-0133.1al/gyw160
Chao, A., Ma, K. H. y Hsieh, T. C. (2016) iNEXT (iNterpolation and EXTrapolation) Online: Software for interpolation and extrapolation of species diversity. Program and user’s guide. http://chao.stat.nthu.edu.tw/wordpress/software_download/inext-online/
Cimé, J. A., Hernández, S. F., Barrientos, R. C. y Castro, A. A. (2010). Diversidad de pequeños roedores en una selva baja caducifolia del noreste de Yucatán, México. Therya, 1, 23–39. https://doi.org/10.12933/therya-10-2
Correa, C. A., Mendoza, M. E. y López, E. (2014). Análisis del cambio en la conectividad estructural del paisaje (1975-2008) de la cuenca de Cuitzeo, Michoacán, México. Revista de Geografía Norte Grande, 59, 7–23. http://dx.doi.org/10.4067/S0718-34022014000300002
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J. B. y Collen, B. (2014). Defaunation in the Anthropocene. Science, 345, 401–406. https://www.science.org/doi/10.1126/science.1251817
Dowler, R. C. y Genoways, H. H. (1978). Liomys irroratus. Mammalian Species, 82, 1–6. https://doi.org/10.2307/3503813
Elizalde, C., López, J. C., Hernández, L., Laundré, J. W., Cervantes, F. A., Morales-Mejía, F. M. et al. (2014). Registro de presencia y actividades de algunos mamíferos en el Desierto Chihuahuense, México. Therya, 5, 793–816. https://doi.org/10.12933/therya-14-252
Escamilla, I. y Aguilar, G. (2010). Urbanización. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 160–163). México D.F.: UNAM/ UMSNH.
Eshelman, B. D. y Cameron, G. N. (1987). Baiomys taylori. Mammalian Species, 285, 1–7. https://doi.org/10.2307/3503776
Fernández, J. A., García-Campusano, F. y Hafner, M. S. (2010). Peromyscus difficilis (Rodentia: Cricetidae). Mammalian Species, 42, 220–229. https://doi.org/10.1644/867.1
Fleharty, E. D. y Olson, L. E. (1969). Summer food habits of Microtus ochrogaster and Sigmodon hispidus. Journal of Mammalogy, 50, 475–486. https://doi.org/10.2307/1378774
Flores, R. y Vázquez, G. (2016). Influence of vegetation type and season on rodent assemblage in a mexican temperate forest mosaic. Therya, 7, 357–369. https://doi.org/10.12933/therya-16-390
García, U. O. y Flores, O. A. (2010). Biodiversidad: anfibios y reptiles. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 94–95). México D.F.: UNAM/ UMSNH.
García, C., Romero, M. L. y Sánchez C. (2002). Comparison of rodent communities in sites with different degrees of disturbance in deciduous forest of southeastern Morelos, México. Acta Zoológica Mexicana, 85, 153–168. https://doi.org/10.21829/azm.2002.85851819
Gloria-Tapia, G., López, C., González, A. y Hernández, S. F. (2012). Diversidad de roedores y su relación con la heterogeneidad ambiental en la cuenca del río Nazas, Durango, México. En F. A. Cervantes y C. Ballesteros (Eds.), Estudio sobre la biología de roedores silvestres mexicanos (pp. 59–70). México D.F.: Instituto de Biología, UNAM.
Godinez, E. y Guerrero, S. (2014). Los roedores de Jalisco, México: clave de determinación. Therya, 5, 633–678. https://doi.org/10.12933/therya-14-212
Gómez-Tagle, A., Gómez-Tagle, A. F., Ávila, J. A. y Bruijnzeel, L. (2015). Partición de la precipitación en un bosque tropical montano de pino-encino en el centro de México. Bosque, 36, 505–518. https://doi.org/10.4067/S0717-92002015000300017
González, A., Rodríguez, N. V. y Marín, G. (2012). Composición del ensamble de pequeños mamíferos del borde de un bosque mesófilo de montaña en Veracruz, México. En F. A. Cervantes y C. Ballesteros (Eds.), Estudio sobre la biología de roedores silvestres mexicanos (pp. 85–96). México D.F.: Instituto de Biología, UNAM.
Hamer, Q., Harper, D. A. T. y Ryan, P. D. (2001). PasT: Paleontological Statistics Software Package for Education and Data Analysis. Paleontologia Electronica, 4, 1–9.
Hernández, S. F., Cimé, J. A., Medina, S. y Durán C. M. (2012). Parámetros poblacionales del ratón yucateco Peromyscus yucatanicus de una selva baja caducifolia del norte de Yucatán, México. En F. A. Cervantes y C. Ballesteros. (Eds.), Estudio sobre la biología de roedores silvestres mexicanos (pp. 151–164). México D.F.: Instituto de Biología, UNAM
Hernández, L., Laundré, J. W., González, A., López, J. y Grajales, K. M. (2011). Tale of two metrics: density and biomass in a desert rodent community. Journal of Mammalogy, 94, 840–851. https://doi.org/10.1644/10-MAMM-A-175.1
Hernández, A., Ortiz, C. F. y Alcaraz, J. V. (2023). El riesgo agrícola ante el cambio climático en la región de Cuitzeo del estado de Michoacán, México. Acta Universitaria Multidisciplinary Scientific Journal, 33, 1–19. https://doi.org/10.15174/au.2023.3763
Herrera, Y. y Pámanes, D. S. (2010). Guía de pastos de Zacatecas. Durango, Durango, México: IPN/ Conabio.
Hutchenson, K. (1970). A test for comparing diversities base on the Shannon formula. Journal of Theoretical Biology, 29, 151–154.
INEGI (Instituto Nacional de Estadística, Geografía e Informática). (2023). Anuario de estadística por entidad federativa. https://www.inegi.org.mx/contenidos/productos/prod_serv/contenidos/espanol/bvinegi/productos/nueva_estruc/889463906353.pdf
López, C., García, D. F., López, J. C. y Elizalde, C. (2019). Multiple lines of evidence reveal a composite of species in the plateau mouse, Peromyscus melanophrys (Rodentia Cricetidae). Journal of Mammalogy, 100, 1583–1598. https://doi.org/10.1093/jmammal/gyz106
López, E., Mendoza, M. E. y Bocco, G. (2010). Cobertura vegetal y uso del terreno. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 52–57). México D.F.: UNAM/ UMSNH.
Luévano, J., Mellink, E., Riojas, M. E. y Flores, J. L. (2008). Comunidades de roedores nocturnos en un ecotono de matorrales micrófilos y zacatal gipsófilo en San Luis Potosí, México. Revista Mexicana de Biodiversidad, 79, 197–203. https://doi.org/10.22201/ib.20078706e.2008.001.528
Madrigal, X. y Guridi, L. L. (2009). Especies arbóreas en riesgo de la cuenca de Cuitzeo, Michoacán y Guanajuato, México. Morelia: Universidad Michoacana de San Nicolás de Hidalgo.
Magurran, A. E. (2004). Measuring biological diversity. Reino Unido: Blackwell Science Ltd.
Mammal Diversity Database. (2024). Explore current mammalian taxonomy. Recuperado el 16 de junio, 2024 de: https://www.mammaldiversity.org/taxa.html
Martínez, S. T., Schnell, G. D., Sánchez, C. y Romero, M L. (2017). Sigmodon mascotensis (Rodentia: Cricetidae). Mammalian Species, 49, 109–118. https://doi.org/10.1093/mspecies/sex013
Medina, M. y Ortega, J. M. (2010). Biodiversidad: peces. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 90–93). México D.F.: UNAM/ UMSNH.
Mendoza, V. H. y Horváth, A. (2013). Roedores y murciélagos en la zona cafetalera del volcán Tacaná, Chiapas, México. Therya, 4, 409–423. https://doi.org/10.12933/therya-13-116
Monroy, O., Zarco, M. M., Ramírez, J. y Ulises Aguilera, U. (2011). Diversidad de mamíferos de la Reserva Natural Sierra Nanchititla, México. Revista Mexicana de Biodiversidad, 82, 237–248. https://doi.org/10.22201/ib.20078706e.2011.1.377
Monterrubio, T. C., Charre, J. F., Colín, C. Z. y León, L. (2014). Los mamíferos del estado de Michoacán. Revista Mexicana de Mastozoología Nueva Época, 4, 1–17. https://doi.org/10.22201/ie.20074484e.2014.4.2.193
Monterrubio, T. C., Guido, D., Alanís, L., León, L. y Medellín, J. F. (2019). Mamíferos silvestres de la cuenca de Cuitzeo, México. Acta Universitaria Multidisciplinary Scientific Journal, 29, 1–15. https://doi.org/10.15174/au.2019.1892
Morales, S. P., Álvarez, M. Y., Zamora, M. E., Dirzo, R., Oyama, K. y Ávila, L. D. (2019). Rodent community responses to vegetation and landscape changes in early successional stages of tropical dry forest. Forest Ecology and Management, 433, 633–644. https://doi.org/10.1016/j.foreco.2018.11.037
Núñez, A. y Pastrana, G. (1990). Los roedores michoacanos: manual de identificación. Morelia, Michoacán: Coordinación de la investigación científica, Universidad Michoacana de San Nicolás de Hidalgo.
Packard, R. L. (1960). Speciation an evolution of pygmy mice, genus Baiomys. University of Kansas Publications, Museum of Natural History, 9, 579–670. https://doi.org/10.5962/bhl.part.6498
Peña, J. L., López, J., Alarcón, B., Vargas, M., Vázquez, I. y Landeros, C. (2009). Composición de la dieta de Sigmodon hispidus (Rodentia: Cricetidae) en caña de azúcar. Mastozoología Neotropical, 16, 365–378.
Pérez-Arteaga, A., Gaston, K. J. y Kershaw, M. (2002). Undesignated sites in Mexico qualifying as wetlands of international importance. Biological Conservation, 107, 47–57. https://doi.org/10.1016/S0006-3207(02)00043-5
Pérez, M. y Santos, A. (2010). Movements and capture-recapture data analysis of the vesper rat (Nyctomys sumichrasti: Rodentia, Muridae) in a tropical forest in northeastern Oaxaca, México. Acta Zoológica Mexicana, 26, 627–638. https://doi.org/10.21829/azm.2010.263804
Ponce, J. y Quijano, A. F. (2010). Biodiversidad: insectos y arácnidos. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 82–84). México D.F.: UNAM/ UMSNH.
Ramírez, J., González, N., Gardner, A. L. y Arroyo, J. (2014). List of recent land mammals of México. Lubbock, TX: Museum of Texas Tech University.
Rojas, A., Aguilar, M., Sánchez, C. y Noguera, O. (2012). Uso del espacio y dinámica poblacional del ratón de los volcanes (Neotomodon alstoni) en el Cerro del Ajusco, Distrito Federal, México. En F. A. Cervantes y C. Ballesteros (Eds.), Estudio sobre la biología de roedores silvestres mexicanos (pp. 211–224). México D.F.: Instituto de Biología, UNAM.
Romero, M. L., Sánchez, C., García, C. y Owen, R. (2007). Mamíferos pequeños. Manual de técnicas de captura, preparación preservación y estudio. México D.F.: UNAM.
Ruán, I., Manson, R. H. e Íñiguez, L. I. (2008). Respuesta al borde en poblaciones de pequeños mamíferos en remanentes de bosque mesófilo de montaña del centro de Veracruz. En C. Lorenzo, E. Espinoza y J. Ortega (Eds.), Avances en el estudio de los mamíferos de México II (pp. 511–530). San Cristóbal de las Casas, Chiapas: Asociación Mexicana de Mastozoología.
Rzedowski, G. y Rzedowski, J. (2005). Flora fanerogámica del valle de México. Pátzcuaro, Michoacán: Instituto de Ecología, A.C./ Conabio.
Sánchez, C., García, C. y Romero, M. L. (2005). Distribución y diversidad de roedores del estado de Michoacán, y su asociación con las provincias fisiográficas. En V. Sánchez y R. A. Medellín (Eds). Contribuciones mastozoológicas en homenaje a Bernardo Villa (pp. 451–460). México D.F.: Instituto de Biología; Instituto de Ecología, UNAM/ Conabio.
Semarnat (Secretaría del Medio Ambiente y Recursos Naturales). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección ambiental – Especies nativas de México de flora y fauna silvestres – Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio – Lista de especies en riesgo. Diario Oficial de la Federación. 30 de diciembre de 2010, Segunda Sección, México.
Senasica (Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria). (2024). Manual de muestreo programa de vigilancia epidemiológica fitosanitaria. https://www.gob.mx/cms/uploads/attachment/file/895962/MANUAL_DE_MUESTREO_2024.pdf
Sikes, R. S. (2016). 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. Journal of Mammalogy, 97, 663–688. https://doi.org/10.1093/jmammal/gyw078
Spencer, S. R. y Cameron, G. N. (1982). Reithrodontomys fulvescens. Mammalian Species, 174, 1–7. https://doi.org/10.2307/3503795
Vázquez, L. B., Medellín, R. A. y Camerón, G. N. (2000). Population and community ecology of small rodents in montane forest of western México. Journal of Mammalogy, 81, 77–85. https://doi.org/10.1644/1545-1542(2000)081<0077:PACEOS>2.0.CO;2
Villafán, K. B., Arellanes, Y., López, R. y Ayala, D. A. (2021). Situación socioambiental en el lago de Cuitzeo, Michoacán (México), desde la responsabilidad social. Economía, Sociedad y Territorio, 21, 599–629. https://doi.org/10.22136/est20211694
Villanueva, A. I., Delgado, D. A., Heynes, S. A., Ruacho, L. y López, C. (2017). Habitat selection by rodents at the transition between the Sierra Madre Occidental and the Mexican Plateau, México. Journal of Mammalogy, 98, 293–301. https://doi.org/10.1093/jmammal/gyw173
Villaseñor-Gómez, L. E. y Villaseñor-Gómez, F. J. (2010). Biodiversidad: avifauna. En S. Cram, L. Galicia, I. Israde, C. López y B. Ávila (Eds.), Atlas de la cuenca del lago de Cuitzeo: análisis de su geografía y entorno socioambiental (pp. 96–99). México D.F.: UNAM/ UMSNH.
Young, H. S., Dirzo, R., Helgen, K. M., McCauley, D. J., Billeter, S. A., Kosoy, M. Y. et al. (2014). Declines in large wildlife increase landscape-level prevalence of rodent-borne disease in Africa. Proceedings of the National Academy of Sciences, 111, 7036–7041. https://doi.org/10.1073/pnas.1404958111
Young, H. S., McCauley, D. J., Dirzo, R., Goheen, J. R., Agwanda, B., Brook, C. et al. (2015). Context-dependent effects of large-wildlife declines on small-mammal communities in central Kenya. Ecological Applications, 25, 348–360. https://doi.org/10.1890/14-0995.1
Zalapa, S. S, Guerrero, S., Badii, M. H. y Cervantes F. A. (2012). Variación espacial del ensamble de pequeños mamíferos de tres áreas de bosque tropical subcaducifolio en la costa norte de Jalisco, México. En F. A. Cervantes y C. Ballesteros (Eds.), Estudio sobre la biología de roedores silvestres mexicanos (pp. 117–126). México D.F.: Instituto de Biología, UNAM.
Zaragoza, E. P., Rodríguez, N. V., Hernández, S. F., León, L. S. y González, M. C. (2022). Abundance, microhabitat and feeding of Peromyscus yucatanicus and Peromyscus mexicanus in the Mexican tropics. Therya, 12, 129–142. https://doi.org/10.12933/therya-22-1189
Carlos Fernando Castillo-García a, b, Juan J. Morrone c, Isaías Hazarmabeth Salgado-Ugarte a, David Espinosa a, *
a Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza, Batalla del 5 de mayo s/n, Ejército de Oriente Zona Peñón, Iztapalapa, 09230 Ciudad de México, Mexico
b Universidad Nacional Autónoma de México, Posgrado en Ciencias Biológicas, Unidad de Posgrado, Edificio D, 1er. Piso, Circuito de Posgrados, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
c Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Biología Evolutiva, Museo de Zoología “Alfonso L. Herrera”, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
*Corresponding author: despinos@unam.mx (D. Espinosa)
Received: 22 January 2024; accepted: 25 February 2025
Abstract
A software program for undertaking track analysis named Panbiotracks is introduced. It aims to solve various issues that currently are present in similar software packages. Panbiotracks is intended: 1) to be a fast, accurate, and reliable tool to generate individual tracks, generalized tracks and panbiogeographical nodes; 2) to solve the dependency on old and obsolete software that other packages have; and 3) to be a free and open source program that can be used in a variety of environments and that can be updated, modified, and improved continuously.
Se presenta un programa para llevar a cabo el análisis de trazos llamado Panbiotracks. Su objetivo es resolver varios problemas que están presentes en paquetes de software similares. Panbiotracks intenta: 1) ser una herramienta rápida, precisa y confiable para generar trazos individuales, trazos generalizados y nodos panbiogeográficos; 2) resolver la dependencia en software anticuado y obsoleto que tienen otros paquetes; y 3) ser un programa libre y de código abierto que pueda ser utilizado en una variedad de entornos y que puede ser actualizado, modificado y mejorado continuamente.
Palabras clave: Panbiogeografía; Análisis de trazos; Python; Software; Sistemática
Introduction
Biological evolution is closely linked to geological history. In the second half of the past century, León Croizat developed a new theoretical approach that he called “panbiogeography”. According to panbiogeography, the geographical distribution of taxa is mainly influenced by geological changes that lead to vicariance processes, where geographical barriers are the main cause of biota fragmentation, which in turn leads to speciation (Morrone, 2015).
Track analysis is panbiogeography’s primary methodological tool (Craw, 1988; Morrone, 2015). It is based on the spatial congruence between tracks, which are geometrical representations of taxon distribution. The 3 components of track analysis are individual tracks, generalized tracks, and panbiogeographical nodes (Morrone, 2015; Page, 1987). An individual track (Fig. 1a-d) is the basic unit of a panbiogeographical analysis, and is defined as an open, non-cyclic graph, also known as a minimum spanning tree (MST), whose vertices represent the geographical locations of the taxon, and the edges are the shortest paths that connect each of them. Mathematically speaking, it is a graph with n localities that are connected through n-1 edges, and whose total length is the shortest possible (Page, 1987). It can be thought of as a graphical representation of the main coordinates of a taxon in space. The basic method to draw an individual track consists of selecting a random locality and linking it to the nearest one using a straight line, then connecting both to another one, also the nearest, and repeating these steps until all the localities are connected, but without any loop between any of them. The most straightforward way to automate this is by building an adjacency matrix to find the edges, then saving the track to a file. However, some difficulty arises when measuring the distances between points, because this needs to be done considering the shape of the earth. There are multiple methods to do this, which are explained with more detail in the Algorithms subsection.
A generalized track (Fig. 1e) is a graph built from the statistically significant superposition between 2 or more individual tracks from different taxa, and it represents the history of ancestral biotas that were fragmented in the past (Craw, 1988; Morrone, 2015). However, there isn’t a formalized implementation of this concept, resulting in multiple methods that can be used to quantify it, like Parsimony Analysis of Endemicity (PAE) (Morrone, 2014; Rosen, 1988), Track Compatibility Analysis (Craw, 1988, 1989), or geometrical methods. A major issue is the difficulty of automating the process to find the generalized tracks, since the superposition between individual tracks cannot be calculated easily. Some programs, like MartiTracks (Echeverría-Londoño & Miranda-Esquivel, 2011), have approached the problem from a purely geometrical standpoint, using distances between segments to decide whether they are congruent with each other or not, but the results have not been satisfactory (Ferrari et al., 2013). The software Trazos2004 (Rojas-Parra, 2007) also uses a geometrical method, where the program finds the intersections between individual tracks and builds a generalized track from those intersections. This method does not follow the formal definition of a generalized track, since it considers only the superpositions between tracks and ignores those cases where 2 segments may be very close to each other, but without overlapping, which could also be considered as spatial congruence (Zunino & Zullini, 2003; Morrone, 2015). Even then, this method is fast and can be used to get an overall congruence approximation, since in many cases the generalized tracks obtained will reflect a general degree of similarity between individual tracks. To differentiate this approach from others, we opted to refer to the generalized tracks obtained by this method as “internal generalized tracks” (IGT).
A panbiogeographical node (Fig. 1f), marked by the superposition between the terminal segments of 2 or more generalized tracks, indicates the location of a complex area of tectonic or biotic convergence (Craw, 1988; Morrone, 2015); however, this concept may have multiple interpretations. Heads (2004) mentions that a node might represent the location of endemism, high diversity, distribution boundaries, areas of disjunction and “anomalous” absences, but also states that there are more meanings to it than these. One problem arises from the existing ways to quantify a node. As mentioned, a node is located at the ends of a generalized track, or more specifically, where 2 terminal vertices (those that are connected only to another vertex) from different generalized tracks overlap. However, other authors had considered the intersections between individual tracks, or the intersections between non-terminal segments of 2 or more generalized tracks, as nodes (Grehan, 2001; Henderson, 1989). Morrone (2015) mentions that, to avoid confusion, the former might be called “individual nodes” and the latter “generalized nodes”. In this regard, panbiogeographical nodes can be considered as a subset of generalized nodes, since the latter accounts for all the intersections between generalized tracks, while the former takes into account only those that occur at the terminal segments of generalized tracks. This means that a tool that finds generalized nodes can be used to find panbiogeographical nodes, if only those that fulfill the proper definition will be counted. Out of the 3 concepts, nodes have received the least attention from a computational point of view, possibly because the process to limit the search of the intersections between tracks to only those that happen at the terminal segments has not been properly explored.
Figure 1. Panbiogeography’s main components. a-d) Individual tracks; e) generalized tracks; f) panbiogeographical node. Maps by CF Castillo-García.
The usefulness of panbiogeography and track analysis has been questioned many times, mainly regarding its ability to generate meaningful results about the evolution of species through space and time (Seberg, 1986; Waters et al., 2013). Others have criticized how track analysis is done, like the criteria used for track orientation (Seberg, 1986), or the lack of a true quantitative process to obtain generalized tracks and nodes (Ferrari et al., 2013). However, many studies have used the panbiogeographical method to identify distributional patterns of many taxa, like mammals (Escalante et al., 2018; Florentin et al., 2016), mollusks (Aguilar-Estrada & Morrone, 2022; López et al., 2022), plants (García-Díaz et al., 2023; Puga-Jiménez et al., 2013), birds (Beauchamp, 1989), fungi (González-Ávila et al., 2017), arthropods (Maya-Martínez et al., 2011), and fossil taxa (Gallo et al., 2013; Hernández-Cisneros & Vélez-Juarbe, 2021). The ability to detect ancestral biotas through generalized tracks and nodes has been mentioned by other researchers and panbiogeography has also been considered useful from other points of view, such as searching for areas of biological complexity like areas of endemism and priority conservation areas (Craw et al., 1999; Escalante et al., 2018; Miguel-Talonia & Escalante, 2013; Morrone, 2015). Because of this, we consider that the development and improvement of new methods and tools for track analysis are important and should be continued.
In recent times, several software packages have been published to automate track analysis using different algorithms and programming languages to calculate both the MSTs and the superposition between tracks and nodes. Some of them, like Trazos2004 (Rojas-Parra, 2007), Croizat (Cavalcanti, 2009), and fossil (Vavrek, 2011), use Prim’s algorithm (Sedgewick & Wayne, 2011). PASSaGE (Rosenberg & Anderson, 2011) uses its own algorithm, while SAM (Rangel et al., 2010) and MartiTracks (Echeverría-Londoño & Miranda-Esquivel, 2011) do not specify which algorithm they use. A list of the aforementioned software packages and the algorithms used by each is presented in Table 1. These programs, however, suffer from one or various of several issues: lack of documentation about what internal methods or algorithms they use, lack of accuracy in their results (Escalante et al., 2017, 2018), dependency on obsolete and unsupported software, difficulty in obtaining or reviewing their source code to improve or modify it, and lack of support for different operating systems other than Microsoft Windows and, sometimes, Apple macOS.
Table 1
Comparison between different software packages for performing track analysis and their algorithms.
Program
Algorithms used
Measuring distance method
Platform
Croizat
Prim (1957), Bron and Kerbosch (1973), Wormwald (1984)
Not specified
Windows, macOS, Linux
Fossil
Not specified
Not specified
Independent
MartiTracks
Proprietary
Not specified
Windows, Linux
PASSaGE
Proprietary
Proprietary
Windows, macOS, Linux
SAM
Not specified
Not specified
Windows
Trazos2004
Prim (1957)
Bessel formulae
Windows
Panbiotracks
Panbiotracks is a Python program for track analysis that aims to overcome some of the issues mentioned above, namely the accuracy of results, dependency on obsolete software, analysis speed, and portability between operating systems. Panbiotracks addresses these issues in the following manner: 1) Accuracy and speed, by using an improved set of functions, Panbiotracks generates results in a shorter time and in more accurately than its predecessors. Moreover, since its code is smaller and is based on modern tool kits, it can be updated and improved faster than other solutions. 2) Dependency on obsolete software, since Panbiotracks uses modern releases of the Python programming language and its code is constantly reviewed to ensure that it can be used with newer versions, it is guaranteed that it will not be made obsolete in the short term. Additionally, its code can be ported to future versions of Python more easily. 3) Portability and compatibility, releases of Panbiotracks are published currently for the Windows and Linux operating systems, and a version for macOS is planned. Moreover, its source code is available publicly, which allows anyone to download it and build their own executable.
Technical information
Panbiotracks is programmed in the Python programming language version 3. It consists of one main command-line executable that can be configured at runtime to do different analyses by passing a flag that enables a particular function. Current functions are: -m I flag, generates individual tracks from a list of taxon names and localities, established by their geographical coordinates (latitude and longitude), and saves them to an ESRI shape file (SHP); -m P flag, generates an internal generalized track (IGT) from a set of individual tracks. It identifies the intersections between individual tracks and it marks the associated coordinates, then builds a new track based on those coordinates and saves it to an ESRI shape file (SHP); -m N flag, locates and identifies “generalized nodes”, which result from the intersections between 2 or more IGTs and saves them to an ESRI shape file (SHP).
Panbiotracks can be downloaded from its GitHub repository and its source code is freely available and licensed under the GNU General Public License version 3: https://github.com/cfnnmcg/panbiotracks
Algorithms
The first component of a track analysis is the individual track, which, according to Page (1987), is defined as a minimum spanning tree (MST) with n vertices and n-1 edges, that does not contain any loop, and whose total length (the sum of all of the lengths of its edges) is not greater than the length of any other graph built from the same vertices (Sedgewick & Wayne, 2011). To build an individual track, the starting point are presence records from the taxa to be evaluated, which will be treated as the vertices of the MST. To create the MST, Panbiotracks builds an adjacency matrix to save the list of the nodes and the connections between them, with as many rows and columns as there are localities in the input file. In an adjacency matrix, each row and column crossing represents a node, and the cells or intersections between them are the graph’s edges. In each cell there can be a numeric value indicating whether there is a connection between nodes or not. Since an MST is a weighted graph, that is, a graph whose edges have a length value, presences are coded with the length of each edge, while absences are coded with a zero (Table 2).
Table 2
Example of an adjacency matrix used by Panbiotracks to represent a MST.
1
2
3
4
5
1
0
66.827884
450.588278
467.852057
67.569065
2
66.827884
0
486.027102
499.716705
1.152371
3
450.588278
486.027102
0
33.259944
487.176345
4
467.852057
499.716705
33.259944
0
500.858559
5
67.569065
1.152371
487.176345
500.858559
0
Since Earth is not a 2-dimensional plane but a spheroid, it is necessary to devise a method to calculate the correct minimal distances between localities. Several approximations have been used by many authors. Rojas-Parra (2007) used Bessel’s trigonometric formulae, which are somewhat similar to the haversine formula (Brummelen, 2013) in that they both calculate the distance between 2 points located over a sphere. In the software PASSaGE (Rosenberg & Anderson, 2011), the authors used a formula to measure the great circle distance over the earth’s surface. However, since the shape of our planet is not a perfect sphere, these methods can lead to inaccurate results. To address this issue, Panbiotracks makes use of Vincenty’s formulae (Vincenty, 1975), which take into account Earth’s spheroid shape, providing a more accurate distance measurement between points. Using this method, Panbiotracks calculates the distance in kilometers between each pair of localities and stores the result in the corresponding cell. To build properly the MST there are numerous algorithms; Prim’s (Prim, 1957) and Kruskal’s (Kruskal, 1956) algorithms are the most used (Graham & Hell, 1985). Panbiotracks uses Prim’s algorithm to build the MST from the adjacency matrix, where a random vertex is selected, then the program checks the distance between it and the others. The shortest distance is selected and the associated vertex added to a temporal matrix. The software then repeats the process until there are no remaining vertices. Finally, a function is used to draw the track from this matrix of vertices and distances and save it to a SHP file.
To build an internal generalized track (IGT), Panbiotracks starts from a set of individual tracks belonging to different taxa and, upon loading the files, the software removes any rows with duplicate data, based on the combination of the taxon name and its coordinates. This is done to ensure that no duplicate points are present. Then, it uses a function where each individual track is compared with all the others to assess if there is any overlapping point between them. An overlap is considered positive when any segment or segments from the first individual track superimpose with any other segment from the second individual track. When this happens, the program stores the point or points where the overlap take place, as well as their coordinates, in a list, then repeats the process for each of the other individual tracks until no more overlapping points are found. If a segment from one track coincides in its totality with another segment from another track, then their starting and ending vertices and the associated coordinates are added to the list. This list is then processed with the same algorithm as with the individual tracks, where the shortest distance between vertices is calculated, then a MST is built from that data and the IGT is saved to a SHP file.
To detect generalized nodes, Panbiotracks uses a modified version of the method used for the IGTs. First, it loads a set of 2 or more IGTs or generalized tracks into memory. A function to compare each track with all the others to find any intersecting point between them is used. This function will detect any overlapping points between segments and will store them and their coordinates in a list. Then another function is used to transform this set of points into a multi-point geometry. This geometry is then saved to a SHP file.
Input and output data
Panbiotracks is programmed with ease of use in mind, so its operation is straightforward. The initial input data needed to do a track analysis is a list of taxa and localities contained within a comma-separated (CSV) file that is loaded to Panbiotracks using the individual tracks flag. This file must have 3 columns: species, lat, and lon, with the taxon name, its latitude, and its longitude, respectively:
species,lat,lon
taxon_A,19.432637,-99.133205
taxon_A,19.1498,-99.0275
taxon_B,20.676215,-103.346979
taxon_B,21.00498,-102.9752
taxon_C,25.671798,-100.309384
taxon_C,24.61789,-98.762498
For IGT generation and node identification, the input data are SHP files containing tracks. The output data for all 3 functions are SHP files as well, containing MultiLineString data in the case of individual tracks and IGTs, and MultiPoint data in the case of nodes. Panbiotracks has been tested with CSV files containing thousands of records, with minimal memory impact and generating the corresponding individual tracks in less than 60 seconds. When computing IGTs or nodes, running time escalated according to the number of input files being evaluated, since the program needs to assess each possible combination of tracks.
Example analysis
To demonstrate the capabilities and use of Panbiotracks, an example analysis is presented. Detailed instructions and explanations about running and using the software can be found in the project’s documentation, available at the GitHub repository. For this example, occurrence data from the Global Biodiversity Information System (GBIF) were used, corresponding to several species of pine (genus Pinus) and oak (genus Quercus), mainly from the Mexican mountain ranges (GBIF.org, 2016a, 2016b), and totaling over 3 thousand records. These data were saved to a file named pinus_quercus_simplified.csv and formatted according to the example specified in the previous section. The following code block shows a sample of this file and its format:
The CSV file is loaded into Panbiotracks using the following command, assuming that it is in the same directory as the program:
panbiotracks -m I -i ./pinus_quercus_simplified.csv -o ./pinus_quercus/
Where panbiotracks is the name of the executable program. Option -m I configures the individual tracks function. The -i flag defines the name of the input file and where it is located, and the -o flag defines the path where the individual tracks file or files will be saved. Panbiotracks generated a separate SHP file for each of the taxon names contained in the CSV file (Fig. 2). These files can be projected by any GIS software, like QGIS (Fig. 3).
Internal Generalized Tracks
To generate the IGTs, the individual tracks were first grouped based on their geographical proximity and record density (where the majority of records from a given species was located). For this example, we focused on 4 groups (Table 3), 2 from each genus, mainly located in the Sierra Madre Occidental (SMOcc), the Sierra Madre del Sur (SMS), and the Eje Volcánico Transversal (EVT). Pinus groups covered mainly the SMS and the SMOcc (Fig. 4), while Quercus groups spanned across the SMS, the EVT, and the SMOcc (Fig. 5). For each group, IGTs were built using a command similar to the following:
Option -m P configures the program to build IGTs. The individual tracks files needed are defined after the -i flag and must be separated by spaces. After analyzing the individual tracks, Panbiotracks will save the IGT to the output file name defined after the -o flag. Please note that, unlike the command for individual tracks, the name defined here is not the name of the directory where you want to save the IGT, but the name of the file itself. This process is repeated for each IGT that needs to be done. As with the individual tracks, the SHP files of the IGTs can be projected in QGIS or other GIS software (Fig. 6).
Table 3
Table of the species used in the example analysis and their grouping.
Genus
Group
Species
Pinus
SMS
P. devoniana, P. lawsonii, P. maximinoi, P. oocarpa, P. pringlei, P. rzedowskii
SMOcc
P. durangensis, P. engelmannii, P. jaliscana, P. leiophylla, P. lumholtzii, P. luzmariae, P. maximartinezii, P. praetermissa
The 4 sets of individual tracks gave 4 IGTs, which were then loaded into Panbiotracks to find the generalized nodes between them. The syntax follows the same logic as the previous cases:
panbiotracks -m N -i ./pinus_quercus/igt/pinus_30_pac_smocc-sms.shp ./pinus_quercus/igt/quercus_30_pac_smocc-evt.shp ./pinus_quercus/igt/quercus_30_pac_smocc-sms-evt.shp -o ./pinus_quercus/gn/pinus30-smocc-sms_quercus30-smocc-evt-smocc-sms-evt
Where -m N configures the program to find generalized nodes. The rules to add the input and output files are the same as with the IGTs. In this case, each input path and file name (pinus_30_pac_smocc-sms.shp, quercus_30_pac_smocc-evt.shp, quercus_30_pac_smocc-sms-evt.shp) corresponds to an IGT and is separated from the others by a space, whereas the output file name is defined after the -o flag. As with the IGTs, the name corresponds to the actual file and not a directory. Panbiotracks will generate a single SHP file with all the generalized nodes in it (Fig. 7).
Figure 2. Individual tracks files generated by Panbiotracks.
Conclusions and future developments
Though many software tools exist for assisting panbiogeography’s track analysis, most of them are difficult to use, difficult to distribute, rely on outdated software or are outdated themselves, lack precision in their algorithms or results, their results are difficult to reproduce, or their development has been halted for more than a decade in some cases. Panbiotracks aims to solve those problems by using efficient algorithms in constant optimization, by developing methods that are fast and precise, and by using a modern software stack that is extensible, easy to implement, and potent.
Panbiotracks’ algorithm for building individual tracks is faster and, thanks to its use of Vincenty’s formulae, more precise. In tests with CSV files with more than 3 thousand records, Panbiotracks generated the corresponding individual tracks in less than thirty seconds. When compared with other programs, like Trazos2004 (Rojas-Parra, 2007), the tracks generated with Panbiotracks were more precise, meaning that the distances between their vertices were more accurately calculated, and they did not have issues in their construction of loops (closed segments within a track), problems that have been observed with Trazos2004.
Figure 3. Individual tracks generated by Panbiotracks. Top, individual tracks from Pinus data; bottom, individual tracks from Quercus data. Maps by CF Castillo-García.
As mentioned, the algorithm for generating IGTs finds any intersection between pair of tracks to get a list of vertices from which the IGT will be built. This same algorithm is used to find the generalized nodes within IGTs. This method has various advantages, like its speed and straightforward approach. An IGT is a type of generalized track that only takes into account “true” connections, that is, intersections between individual tracks. From this point of view, it can be said that an IGT is a form of generalized track with the least ambiguity, since an overlap can be identified without any uncertainty. Considering this, a tool that can identify and generate these features is very valuable and useful.
There are, however, some pending issues regarding the IGTs and the method to identify nodes. If we consider the formal definition of a generalized track, it is necessary to add improvements to the method used and allow the program to consider other degrees of congruence between individual tracks besides the direct superposition. For example, with the current method, segments of individual tracks that are very close to each other but that do not intersect, will not be marked as part of the generalized track. This can lead to the dismissal of areas where 2 or more taxa share a similar evolutionary path, which can be represented by a generalized track, but since they do not overlap, the program does not consider their congruence. An example of this can be seen in Figure 4a, where the 2 westernmost segments are almost parallel, but the algorithm does not count them for the IGT (Fig. 6a). A potential solution is to add an option to define a buffer or area of influence around the individual tracks, and use that to compute the generalized track, but this method requires more testing to be implemented properly. There are other methods that are currently being researched for their inclusion in Panbiotracks. One is the Fréchet distance, which measures the degree of similarity between curves (Alt & Godau, 1995; Aronov et al., 2006). Another is the Hausdorff distance, which measures the distance of 2 subsets of a metric space (Bai et al., 2011; van Kreveld et al., 2022). Both concepts can potentially be used to detect and build generalized tracks with more accuracy than with present methods.
Figure 4. Groups of individual tracks from Pinus data: a) SMS group; b) SMOcc group. Maps by CF Castillo-García.
It also should be noted that Panbiotracks currently does not automatically separate the ITs that will be used for the IGTs. Because of this, it is strongly advised to first use methods like PAE-PCE (Luna-Vega et al., 1999, 2000) or NDM/NVDM (Goloboff, 2005) to segregate those ITs that are useful for building an IGT. These methods have been used to identify areas of endemism (García-Barros et al., 2002; Santiago-Alvarado et al., 2022), but they can also be used to define sets of species whose ITs are related enough to form an IGT.
The formal definition of a panbiogeographical node indicates that it is present only at the intersection of 2 or more endpoint vertices from 2 or more generalized tracks, that is, those vertices located at the periphery of the MST that only have one edge connecting them to another node (Henderson, 1989; Morrone, 2015). Since Panbiotracks takes into account all the intersections between generalized tracks, it does not quite follow this definition. However, as mentioned before, the generalized nodes can be seen as a set of which the panbiogeographical nodes are a part. As such, it is necessary to develop an improved algorithm that can filter and locate only the desired localities. This also applies for those cases where 2 terminal segments from different IGTs do not overlap. Under certain circumstances, a panbiogeographical node can be said to be present at a given location, but the IGTs might not overlap and, consequently, the software will not consider this area as a node. In Fig. 6, for example, the area towards the northwest of the map, where there are terminal ends of the IGTs, might be considered a node, but the program does not mark it. To solve this, a similar solution to that of the IGTs may be useful: define a buffer and evaluate the degree of overlap not between tracks, but between these buffers.
Figure 5. Groups of individual tracks from Quercus data: a) SMOcc-EVT; b) SMOcc-SMS-EVT. Maps by CF Castillo-García.
Automating track analysis and other methods in biogeography is essential and of utmost importance to the future of this field of knowledge. With the introduction of faster computer systems and programming languages specialized in data manipulation and analysis, like Python and Julia, it is necessary to develop new algorithms and improve existing ones. Moreover, another issue of great importance is to ensure that the software can be updated and improved and not let it become obsolete or dependent on outdated systems and software.
Figure 6. Generalized tracks identified from the ITs: a) IGT from the Pinus SMS group; b) IGT from the Pinus SMOcc group; c) IGT from the Quercus SMOcc-EVT group; d) IGT from the Quercus SMOcc-SMS-EVT group. Maps by CF Castillo-García. Figure 7. Generalized nodes identified from the intersections from all 4 groups. Map by CF Castillo-García.
Acknowledgments
This paper serves as a fulfillment of CFC-G for obtaining a M.Sc. degree in the Posgrado en Ciencias Biológicas, UNAM. We thank to the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (Secihti) for the support of this research through a graduate scholarship (No. 1178796) to CFC-G. This work was funded by the project IN215321 (DGAPA-PAPIIT).
References
Aguilar-Estrada, L. G., & Morrone, J. J. (2022). Distributional patterns of Vetigastropoda (Mollusca) all over the world: a track analysis. Zoological Journal of the Linnean Society, 196, 442–452. https://doi.org/10.1093/zoolinnean/zlac004
Alt, H., & Godau, M. (1995). Computing the Fréchet distance between two polygonal curves. International Journal of Computational Geometry & Applications, 5, 75–91. https://doi.org/10.1142/S0218195995000064
Aronov, B., Har-Peled, S., Knauer, C., Wang, Y., & Wenk, C. (2006). Fréchet distance for curves, revisited. In Y. Azar, & T. Erlebach (Eds.), Algorithms – ESA 2006 (pp. 52–63). Springer. https://doi.org/10.1007/11841036_8
Bai, Y. B., Yong, J. H., Liu, C. Y., Liu, X. M., & Meng, Y. (2011). Polyline approach for approximating Hausdorff distance between planar free-form curves. Computer-Aided Design, 43, 687–698. https://doi.org/10.1016/j.cad.2011.02.008
Beauchamp, A. J. (1989). Panbiogeography and rails of the genus Gallirallus. New Zealand Journal of Zoology, 16, 763–772. https://doi.org/10.1080/03014223.1989.10422933
Bron, C., & Kerbosch, J. (1973). Algorithm 457: Finding all cliques of an undirected graph. Communications of the ACM, 16, 575–577. https://doi.org/10.1145/362342.362367
Brummelen, G. V. (2013). Heavenly mathematics: the forgotten art of spherical trigonometry. Princeton, NJ: Princeton University Press.
Cavalcanti, M. J. (2009). Croizat: a software package for quantitative analysis in panbiogeography. Biogeografía, 4, 4–6.
Craw, R. (1988). Continuing the synthesis between panbiogeography, phylogenetic systematics and geology as illustrated by empirical studies on the biogeography of New Zealand and the Chatham Islands. Systematic Biology, 37, 291–310. https://doi.org/10.1093/sysbio/37.3.291
Craw, R. (1989). New Zealand biogeography: a panbiogeographic approach. New Zealand Journal of Zoology, 16, 527–547. https://doi.org/10.1080/03014223.1989.10422921
Craw, R., Grehan, J., & Heads, M. (1999). Panbiogeography: tracking the history of life. Oxford, UK: Oxford University Press.
Echeverría-Londoño, S., & Miranda-Esquivel, D. R. (2011). MartiTracks: a geometrical approach for identifying geographical patterns of distribution. Plos One, 6, e18460. https://doi.org/10.1371/journal.pone.0018460
Escalante, T., Noguera-Urbano, E. A., & Corona, W. (2018). Track analysis of the Nearctic region: identifying complex areas with mammals. Journal of Zoological Systematics and Evolutionary Research, 56, 466–477. https://doi.org/ 10.1111/jzs.12211
Escalante, T., Noguera-Urbano, E. A., Pimentel, B., & Aguado-Bautista, O. (2017). Methodological issues in modern track analysis. Evolutionary Biology, 44, 284–293. https://doi.org/10.1007/s11692-016-9401-8
Ferrari, A., Barão, K. R., & Simões, F. L. (2013). Quantitative panbiogeography: Was the congruence problem solved? Systematics and Biodiversity, 11, 285–302. https://doi.org/10.1080/14772000.2013.834488
Florentin, J. E., Arana, M. D., & Salas, R. M. (2016). Panbiogeographic analysis of Galianthe subgenus Ebelia (Rubiaceae). Rodriguésia, 67, 437–444. https://doi.org/10. 1590/2175-7860201667214
Gallo, V., Avilla, L. S., Pereira, R. C. L., & Absolon, B. A. (2013). Distributional patterns of herbivore megamammals during the Late Pleistocene of South America. Anais Da Academia Brasileira de Ciências, 85, 533–546. https://doi.org/10.1590/S0001-37652013000200005
García-Barros, E., Gurrea, P., Luciáñez, M. J., Cano, J. M., Munguira, M. L., Moreno, J. C. et al. (2002). Parsimony analysis of endemicity and its application to animal and plant geographical distributions in the Ibero-Balearic region (western Mediterranean). Journal of Biogeography, 29, 109–124. https://doi.org/10.1046/j.1365-2699.2002.00653.x
García-Díaz, R. F., Valdez-Hernández, E. F., Martínez-Cárdenas, L., Díaz-Nájera, F., & Ayvar-Serna, S. (2023). Diversity and distribution of Andean tubers: an agrogeographic analysis. Investigaciones y Estudios – UNA, 14, 59–70. https://doi.org/10.57201/IEUNA2313312
GBIF.org. (2016a). GBIF occurrence download. The Global Biodiversity Information Facility. https://doi.org/10.15468/DL.DGUZHT
GBIF.org. (2016b). GBIF occurrence download. The Global Biodiversity Information Facility. https://doi.org/10.15468/DL.ORDDD1
Goloboff, P. (2005). NDM/VNDM (Versión 2.5). Programs for identification of areas of endemism. Programs and documentation available at: http://www.zmuc.dk/public/phylogeny/endemism
González-Ávila, A., Contreras-Medina, R., Espinosa, D., & Luna-Vega, I. (2017). Track analysis of the order Gomphales (Fungi: Basidiomycota) in Mexico. Phytotaxa, 316, 22. https://doi.org/10.11646/phytotaxa.316.1.2
Graham, R. L., & Hell, P. (1985). On the history of the minimum spanning tree problem. Annals of the History of Computing, 7, 43–57. https://doi.org/10.1109/MAHC.1985.10011
Grehan, J. (2001). Biogeography and evolution of the Galapagos: Integration of the biological and geological evidence. Biological Journal of the Linnean Society, 74, 267–287. https://doi.org/10.1111/j.1095-8312.2001.tb01392.x
Heads, M. (2004). What is a node? Journal of Biogeography, 31, 1883–1891. https://doi.org/10.1111/j.1365-2699.2004.01201.x
Henderson, I. M. (1989). Quantitative panbiogeography: an investigation into concepts and methods. New Zealand Journal of Zoology, 16, 495–510. https://doi.org/10.1080/03014223.1989.10422918
Hernández-Cisneros, A. E., & Vélez-Juarbe, J. (2021). Palaeobiogeography of the North Pacific toothed mysticetes (Cetacea, Aetiocetidae): a key to Oligocene cetacean distributional patterns. Palaeontology, 64, 51–61. https://doi.org/10.1111/pala.12507
Kruskal, J. B. (1956). On the shortest spanning subtree of a graph and the traveling salesman problem. Proceedings of the American Mathematical Society, 7, 48–50. https://community.ams.org/journals/proc/1956-007-01/S0002-9939-1956-0078686-7/S0002-9939-1956-0078686-7.pdf
López, B., Naranjo-García, E., & Mejía, O. (2022). Diversity patterns of Mexican land and freshwater snails: a spatiotemporal approach. Revista Mexicana de Biodiversidad, 93, e933966. https://doi.org/10.22201/ib.20078706e.2022.93.3966
Luna-Vega, I., Alcántara-Ayala, O., Espinosa-Organista, D., & Morrone, J. J. (1999). Historical relationships of the Mexican cloud forests: A preliminary vicariance model applying Parsimony Analysis of Endemicity to vascular plant taxa. Journal of Biogeography, 26, 1299–1305. https://doi.org/10.1046/j.1365-2699.1999.00361.x
Luna-Vega, I., Alcántara-Ayala, O., Morrone, J. J., & Espinosa, D. (2000). Track analysis and conservation priorities in the cloud forests of Hidalgo, Mexico. Diversity and Distributions, 6, 137–143. https://doi.org/10.1046/j.1472-4642.2000.00079.x
Maya-Martínez, A., Schmitter-Soto, J. J., & Pozo, C. (2011). Panbiogeography of the Yucatán Peninsula based on Charaxinae (Lepidoptera: Nymphalidae). Florida Entomologist, 94, 527–533. https://doi.org/10.1653/024.094.0317
Miguel-Talonia, C., & Escalante, T. (2013). Los nodos: el aporte de la panbiogeografía al entendimiento de la biodiversidad. Biogeografía, 6, 30–42.
Morrone, J. J. (2014). Parsimony analysis of endemicity (PAE) revisited. Journal of Biogeography, 41, 842–854. https:// doi.org/10.1111/jbi.12251
Morrone, J. J. (2015). Track analysis beyond panbiogeography. Journal of Biogeography, 42, 413–425. https://doi.org/10. 1111/jbi.12467
Page, R. D. M. (1987). Graphs and generalized tracks: quantifying Croizat’s panbiogeography. Systematic Zoology, 36, 1. https://doi.org/10.2307/2413304
Prim, R. C. (1957). Shortest connection networks and some generalizations. The Bell System Technical Journal, 36, 1389–1401. https://doi.org/10.1002/j.1538-7305.1957.tb01515.x
Puga-Jiménez, A. L., Andrés-Hernández, A. R., Carrillo-Ruiz, H., Espinosa, D., & Rivas-Arancibia, S. P. (2013). Patrones de distribución del género Zanthoxylum L. (Rutaceae) en México. Revista Mexicana de Biodiversidad, 84, 1179–1188. https://doi.org/10.7550/rmb.32047
Rangel, T. F., Diniz-Filho, J. A. F., & Bini, L. M. (2010). SAM: A comprehensive application for Spatial Analysis in Macroecology. Ecography, 33, 46–50. https://doi.org/10.1111/j.1600-0587.2009.06299.x
Rojas-Parra, C. A. (2007). Una herramienta automatizada para realizar análisis panbiogeográficos. Biogeografía, 1, 31–33.
Rosen, B. R. (1988). From fossils to earth history: applied historical biogeography. In A. A. Myers, & P. S. Giller (Eds.), Analytical biogeography: an integrated approach to the study of animal and plant distributions (pp. 437–481). Netherlands: Springer. https://doi.org/10.1007/978-94-009-1199-4_17
Rosenberg, M. S., & Anderson, C. D. (2011). PASSaGE: Pattern Analysis, Spatial Statistics and Geographic Exegesis. Version 2. Methods in Ecology and Evolution, 2, 229–232. https://doi.org/10.1111/j.2041-210X.2010.00081.x
Santiago-Alvarado, M., Luna-Vega, I., Rivas, G., & Espinosa, D. (2022). Effect of cell size and thresholds in NDM/NVDM methods on recognizing areas of endemism. Zootaxa, 5134, 1–33. https://doi.org/10.11646/zootaxa.5134.1.1
Seberg, O. (1986). A critique of the theory and methods of panbiogeography. Systematic Zoology, 35, 369–380. https://doi.org/10.2307/2413388
Sedgewick, R., & Wayne, K. (2011). Algorithms (4th ed). Boston, Massachusetts: Addison-Wesley Professional.
van Kreveld, M., Miltzow, T., Ophelders, T., Sonke, W., & Vermeulen, J. L. (2022). Between shapes, using the Hausdorff distance. Computational Geometry, 100, 101817. https://doi.org/10.1016/j.comgeo.2021.101817
Vavrek, M. J. (2011). Fossil: palaeoecological and palaeogeographical analysis tools. Palaeontologia Electronica, 14, 16.
Vincenty, T. (1975). Direct and inverse solutions of geodesics on the ellipsoid with application of nested equations. Survey Review, 23, 88–93. https://doi.org/10.1179/sre.1975.23.176.88
Waters, J. M., Trewick, S. A., Paterson, A. M., Spencer, H. G., Kennedy, M., Craw, D. et al. (2013). Biogeography off the tracks. Systematic Biology, 62, 494–498. https://doi.org/10.1093/sysbio/syt013
Wormald, N. C. (1984). Generating random regular graphs. Journal of Algorithms, 5, 247–280. https://doi.org/10.1016/ 0196-6774(84)90030-0
Zunino, M., & Zullini, A. (2003). Biogeografía: la dimensión espacial de la evolución (Vol. 259). Ciudad de México: Fondo de Cultura Económica.
José Ángel Zamarripa-Fernández a, b, *, Ismael Hinojosa-Díaz c, Moisés Armando Luis-Martínez d, Ricardo Ayala a
a Universidad Nacional Autónoma de México, Instituto de Biología, Estación de Biología Chamela (Sede Colima), Km 59, Carr. Fed. 200, Melaque – Pto. Vallarta, La Huerta, 48895 Chamela, Jalisco, Mexico
b Universidad Nacional Autónoma de México, Posgrado en Ciencias Biológicas, Edificio D, 1°Piso, Circuito de Posgrados, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
c Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, 3er. Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
d Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Biología Evolutiva, Museo de Zoología “Alonso L. Herrera”, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
A taxonomic revision of the cleptoparasitic bee genus Mesoplia Lepeletier, 1841 from Mexico and Central America is presented. We recognize 4 new species, resulting in a total of 10 species for the study area: Mesoplia (Mesoplia) insignis (Smith, 1879), M. (M.) sapphirina Melo & Rocha-Filho, 2011, M. (M.) regalis (Smith, 1854), M. (M.) dugesi (Cockerell, 1917), M. (M.) rufipes (Perty, 1933), M. (M.) veracruzana sp. nov., M. (M.) oaxacana sp. nov., M. (M.) tica sp. nov., M.(Eumelissa) ornata (Spinola, 1841)and M.(E.) carmelitae sp. nov. We propose the following synonymies: M. imperatrix (Friese, 1913) = M. (M.) insignis, and M. guatemalensis (Cockerell, 1912) = M. (M.) rufipes. We describe the previously unknown female of M. (M.) dugesi and provide diagnoses for both sexes based on the holotype. An identification key for the species, illustrations, distributional records (available as supplementary material), and lists of hosts and floral associations are provided.
Revisión taxonómica y distribución de las abejas cleptoparásitas del género Mesoplia (Apidae: Ericrocidini) de México y Centroamérica
Resumen
Se presenta una revisión taxonómica del género de abejas cleptoparásitas Mesoplia Lepeletier de México y Centroamérica. Se reconocen 4 especies nuevas, dando un total de 10 especies para el área en estudio: Mesoplia (Mesoplia) insignis (Smith, 1879), M. (M.) sapphirina Melo & Rocha-Filho, 2011, M. (M.) regalis (Smith, 1854), M. (M.) dugesi (Cockerell, 1917), M. (M.) rufipes (Perty, 1933), M. (M.) veracruzana sp. nov., M. (M.) oaxacana sp. nov., M. (M.) tica sp. nov., M. (Eumelissa) ornata (Spinola, 1841) y M. (E.) carmelitae sp. nov. Se encontraron las siguientes sinonimias: M. imperatrix (Friese, 1913) = M. (M.) insignis y M. guatemalensis (Cockerell, 1912) = M. (M.) rufipes. Se describe la hembra de M. (M.) dugesi, previamente desconocida, y se proporcionan las diagnosis para ambos sexos basados en el tipo. Se aportan una clave de identificación de las especies, ilustraciones, registros de distribución (disponible como material complementario), una lista de huéspedes y asociaciones florales.
The tribe Ericrocidini is composed mainly of cleptoparasitic bees that have Neotropical affinity, with the exception of the genus Ericrocis Cresson, 1887, which is Nearctic (Martins et al., 2018). The greatest diversity occurs in the Amazon region (Silveira et al., 2002; Snelling & Brooks, 1985). Their known hosts are species of the tribe Centridini (Rocha-Filho et al., 2009). The most easily recognizable morphological character of the tribe is the large, bifurcate or multidentate apex of the middle tibial spur (Michener, 2007).
The genus Mesoplia Lepeletier, 1841 is the richest in the tribe with 19 described species (Table 1), although it is estimated that it could include more species (Rozen et al., 2011). It is distributed from northern Mexico (Sinaloa and Tamaulipas) to Argentina (Snelling & Brooks, 1985). Mesoplia is recognized by the following characters: internal orbits more or less parallel; presence of 2 tubercles on the scutellum of variable development; meso and metabasitarsus without lateral patches of short, flattened setae; tiny metallic scales blue or green, mainly in the dorsal area of the metasoma, frequently also with patches of yellowish or whitish pubescence; S5 of male evidently emarginated and hidden under S4 (Genaro & Breto, 2022; Michener, 2007; Snelling & Brooks, 1985; Vivallo, 2010).
The subgeneric classification was proposed by Snelling and Brooks (1985). They proposed 2 subgenera based on morphological characters, Mesoplia and Eumelissa. Males of the former have posterior tibial spurs that do not extend beyond the basal third of the basitarsus, the posterior tibia has an internal distal patch of setae and S5 evidently emarginate and hidden under S4. Females of the same subgenus have a broad pygidial plate, which occupies the exposed dorsal surface of T6, covered with setae or scales and dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces. In contrast, males of the subgenus Eumelissa have long posterior tibial spurs reaching to or beyond middle of basitarsus. Females of the same subgenus have narrow pygidial plate, does not completely occupy the exposed dorsal surface of T6, without scales or setae and dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces (Michener, 2007). Furthermore, Moure (1960a, b) recognizes 2 groups: bifrons and azurea, both of which are part of the subgenus Mesoplia s. str.Males of the bifrons grouphave conspicuous hind tibia setal brush, 2 spurs on the posterior tibia, ventral basal surface of posterior femur without a robust posterior projection. Females have pronotum without a carina extending laterally from the pronotal collar to the pronotal lobe.On the other hand, males of the azurea grouphave inconspicuous hind tibia setal brush, 1 spur on the posterior tibia and ventral basal surface of posterior femur with a robust posterior projection. Females of this group have pronotum with a carina extending laterally from the pronotal collar to the pronotal lobe.
The known hosts include several species of Centridini (Rocha-Filho et al., 2009). For Mesoplia, as in the rest of the tribe, the biology of most species is practically unknown and their geographical distribution uncertain, since some of the information available in the literature is based on mistaken identifications (Vivallo, 2014).
In this work we update the knowledge of the species of Mesoplia forMexico and Central America, providing diagnoses, descriptions, distributional data, identification keys, and a summary of floral and host records. We describe 4 new species and establish 2 new synonyms.
Materials and methods
We reviewed 363 specimens from the following collections where studied specimens are deposited (names of curators and/or collection managers are in parentheses): EBCh, Estación de Biología Chamela, IBUNAM, Jalisco, Mexico (R. Ayala Barajas); CNIN, Colección Nacional de Insectos, Instituto de Biología, UNAM, Mexico City (I.A. Hinojosa Díaz); MZFC, Museo de Zoología “Alfonso L. Herrera”, Facultad de Ciencias, UNAM, Mexico City (M.A. Luis Martínez); CZUG, Centro de Estudios en Zoología, CUCBA, Universidad de Guadalajara, UDG, Jalisco, Mexico (José Luis Navarrete, Hugo Fierros); HFL, Colección Particular de Hugo Eduardo Fierros López, Jalisco, Mexico (Hugo Fierros); IEXA, Colección Entomológica del Instituto de Ecología, A.C., INECOL, Veracruz, Mexico (Jorge E. Valenzuela, Viridiana Vega); ECOAB, Colección de Abejas de El Colegio de la Frontera Sur, Chiapas, Mexico (Rémy Vandame, Jorge A. Mérida Rivas, Philippe Sagot); CER-UADY, Colección Regional Entomológica de la Universidad Autónoma de Yucatán, UADY, Yucatán, Mexico, (Virginia Meléndez Ramírez, María José López Gómez); SEMC, Snow Entomological Collection, Division of Entomology, University of Kansas Natural History Museum, KU, Lawrence, Kansas, USA (Zachary Falin, Michael Engel, Rachel Osborn); MNCR, Museo Nacional de Costa Rica, Heredia, Costa Rica (Maricelle Méndez, Marcela Sánchez); MZUCR, Museo de Zoología, Universidad de Costa Rica, San José, Costa Rica (Mauricio Fernández, Paul Hanson); CINAT, Centro de Investigaciones Apícolas Tropicales, Universidad Nacional de Costa Rica, UNA, Heredia, Costa Rica (Mario Gallardo, Eduardo Herrera).
Table 1
Recognized species of Mesoplia Lepeletier, 1841. * New synonyms.
Dominica, Guadeloupe, Martinique and Saint Vincent
Mesoplia chalybaea (Friese, 1912)
Both
Brazil
Mesoplia dugesi (Cockerell, 1917)
Both
Mexico
Mesoplia guatemalensis (Cockerell, 1912)*
–
–
Mesoplia oaxacana sp. nov.
Female
Mexico
Mesoplia rufipes (Perty, 1833)
Both
Mexico to South America
Eumelissa
Mesoplia alboguttata (Ducke, 1905)
Both
Bolivia and Brazil
Mesoplia carmelitae sp. nov.
Both
México and Costa Rica
Mesoplia decorata (Smith, 1854)
Both
Brazil, Ecuador, Guatemala, Panama and Venezuela
Mesoplia friesei (Ducke, 1902)
Both
Brazil, Colombia and Paraguay
Mesoplia guedesii (Ducke, 1902)
Female
Brazil
Mesoplia ornata (Spinola, 1841)
Both
Mexico to French Guiana
Type specimens of M. sapphirina were studied. For the rest of the species high-resolution images of holotypes were available, which allowed verification of the species that were delimited and that had been previously described: for M. insignis (both sexes)and M. regalis on the website of the Natural History Museum (London, England); the holotype of M. dugesi on the website of the National Museum of Natural History, Smithsonian Institution (Washington D.C., USA); for M. imperatrix images of holotype from the Museum für Naturkunde (Berlin, Germany) were provided by Stefanie Krause; and the holotype images of M. guatemalensis from the American Museum of Natural History (New York, USA) were provided by Corey Smith. Delimitation of other species followed original descriptions and specimens previously identified by taxonomists who worked with the genus and compared their specimens with types deposited in the mentioned collections. Due to the morphological characteristics of the species of this genus, it is possible to be certain of their identification.
Morphological terminology follows Michener (2007); for integument sculpture we follow Harris (1979). Metasomal terga and sterna are designated as T and S respectively, numbered from anterior to posterior. We used F for antennal flagellum, with F1 following the pedicel. Male genitalia were dissected along with S7 and S8. The subgeneric classification follows Snelling and Brooks (1985) and the classification of groups follows Moure (1960a). All measurements are given in millimeters (mm). Images for most of the structures and habitus were taken with a Leica MC 170 HD camera adapted to a Leica M205C stereoscopic microscope except some of the habitus photographs were taken with a Cannon EOS 70D DSLR camera and a Canon EF-S 60 mm macro lens. For Mesoplia tica sp. nov., images were taken using an Olympus DP74 camera attached to an Olympus SZX16 stereoscopic microscope. Images were processed with Adobe Photoshop CS5®, Photopea.com, HeliconSoft and Leica Application Suite. The scale bar indicates 0.5 mm in the genitalia images. Distribution maps were produced from specimen locality data (available as supplementary material) using QGis software and the base map of Esri Physical and were prepared by the first author. Geospatial coordinates are presented in DMS. Plant associations were based on the information present in the specimen label. For the cleptoparasite-host associations we reviewed and compiled the published information categorized following Rocha-Filho et al. (2009).
Description
We recognize 10 species of Mesoplia for Mesoamerica; 4 are new to science herein described and illustrated. In addition, 2 synonyms are recognized, Mesoplia imperatrix (Friese, 1913) = M. insignis (Smith, 1854) and M. guatemalensis (Cockerell, 1912) = M. rufipes (Perty, 1833).
Genus Mesoplia Lepeletier, 1841
Mesoplia Lepeletier, 1841: 457. Type species: Mesocheira azurea Lepeletier & Serville, 1825: Museo Regionale di Scienze Naturale, Torino, Italy (♀♂), monobasic.
Melissa Smith, 1854: 279. Type species: Mesocheira azurea Lepeletier & Serville, 1825, designated by Sandhouse, 1943: 570.
Diagnosis. Maxillary palps elongated, with 3 or 4 segments; internal orbits more or less parallel; scutellum bituberculate (in some species not very evident); medial basitarsus with a flattened distal process on the posterior margin, more evident in females (Figs. 1, 2); dorsal region of the metasoma covered with tiny scales that give it a metallic blue or green color, regularly with small patches of yellow or white pubescence; S5 of the male evidently emarginate and hidden under S4; body length from 10 to 25 mm.
Subgenus Mesoplia Lepeletier, 1841
Diagnosis. Male: distal inner surface of the posterior tibia with a setal brush, not very evident in some species (Fig. 3A-E); hind tibial spurs not reaching beyond basal third of basitarsus, some species with only 1 spur present. Female: pygidial plate broad, occupying the exposed dorsal surface of T6; pygidial plate covered with scales or setae (Fig. 4A-G); dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces.
M. bifrons group(sensu Moure, 1960a, b)
Diagnosis. Male: hind tibia with setal brush conspicuous (Fig. 3A-C) and 2 spurs; posterior femur with ventral basal margin without a robust posterior projection; posterior basitarsus with inner surface without a longitudinal carina. Female: without a carina between pronotal collar and the pronotal lobe (Fig. 5A).
Mesoplia (Mesoplia) insignis (Smith, 1879)
(Figs. 3A, 4A, 5A, 6, 7A, 8-10, 35)
Melissa insignis Smith, 1879: 108. Syntypes: British Museum Natural History, London, England (♀♂). Type locality: Mexico, Veracruz, Orizaba.
Melissa imperialis Friese, 1912: 205. Holotype: Museum für Naturkunde, Berlin, Germany (♀). Type locality: Mexico, Morelos. New junior synonym.
Melissa imperatrix Friese, 1913: 89. nom. nov. for Melissa imperialis Friese, 1912. New junior synonym.
Melissa imperatrix Friese, 1921: 266. nom. nov. for Melissa imperialis Friese, 1912. New junior synonym.
Diagnosis. Male: metallic greenish body, with pubescence on head and mesosoma mostly bright yellow; integument of antenna, pronotal lobe, tegula and legs black; scutellar tubercles conical, inconspicuous due to dense pubescence (Fig. 6D), with perceptible cleft between them; robust forked medial tibial spur, with dark integument, wide anterior projection and with between 2 and 4 internal teeth; pubescence of the terga mainly whitish with a spot of bright yellow pubescence on both lateral margins of T1, at least also present in T2 (Fig. 6A); surface of T7 without scales but with black pubescence; apex of T7 slightly forked, both apices truncated and bare at tip (Fig. 7A); S4 shallowly emarginate, with several rows of compound pubescence on the apical margin, dark in color in the center and becoming light at the laterally; digitiform lobe of the gonostylus triangular and broad, projecting upwards (Fig. 8C, D).
Figure 1. Middle leg basitarsus, male: A, Mesoplia (M.) sapphirina. B, Mesoplia (M.) dugesi. C, Mesoplia (M.) rufipes. D, Mesoplia (E.) carmelitae sp. nov. E, Mesoplia (E.) ornata.
Female: body dark metallic blue and black pubescence, but with some small patches of whitish pubescence behind the antennal area, preoccipital area, lateral margins of the scutum to the level of tegula, and lateral margins of S2-S4 (Fig. 9A-C); black integument on antenna pronotal lobe, tegula and legs; scape with dark metallic blue; integument of scutum and tegula with scales that give a dark metallic blue appearance; scutellar tubercles mamilliform; tergal pubescence mainly dark; ventral surface of mesosoma and metasoma with metallic blue scales; middle tibial spur robust with black integument, forked, anterior projection with 1 to 4 internal teeth; pygidial plate with a broad and quadrangular apex, without scales on the surface, with blackish pubescence that does not reach the apex, black integument (Fig. 4A).
Variation. Some females from Morelos with white setae on lateral edges of the labrum and posterolateral edges of the propodeum. Pubescence and the presence of metallic scales in some body areas of males seem to be relatively variable, labral setae can be yellow, whitish or dark; the clypeus may have yellow setae or metallic blue scales, as well as the dorsal surface of the scutellar tubercles.
Material examined. Mexico: Jalisco: 22.5 km al NE de Magdalena, 20°58’46.17” N, 104°6’52.33” W, 1,066.8 m, 19-VII-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1254912); Tala, Bosque Escuela, 20°35’44.9988” N, 103°38’15” W, 1,499 m, 7-IX-2012, H. E. Fierros-López, 1♀ (CZUG); ANP Bosque la Primavera, S. Nopales, 20°36’3.4” N, 103°38’20.4” W, 1,500 m, 26-IX-2013, O. Serrano-Pineda, 1♀ (CZUG); Tequila, Volcán de Tequila, 20°51’52” N, 103°50’36” W, 1,300 m, 29-VII-1995, H. E. Fierros-López, 1♀ (CZUG 580); 20°49’36” N, 103°50’45” W, 1,300 m, 29-VIII-1995, H. E. Fierros-López, 1♀ 1♂ (HFL), 20°51’36” N, 103°51’0” W, 1,370 m, 12-VI-2022, K. Sanchez, 1♂ (ECOAB 87968); Tlajomulco de Zúñiga, Fraccionamiento Nueva Galicia coto Valencia, 20°34’39” N, 103°26’40” W, 1,618 m, 24-IX-2022, J. A. Zamarripa-Fernández, C. Sánchez-Bastidas e I. I. Alfaro-Ríos, 5♀ (CZUG); Zapopan, Barranca la Experiencia, 20°57’42.804” N, 103°24’5.292” W, 1,300 m, 14-IX-1996, G. Mercado, 1♀ (CZUG 1112), 1♂ (CZUG 1113), 22-VIII-2009, G. Mercado, 1♀ (CZUG EXP0440), 1♂ (CZUG EXP0446), 5-IX-2009, G. Mercado, 1♂ (CZUG EXP0445), 1,350 m, 29-VII-2009, G. Mercado, 1♂ (HFL), 29-VII-2009, G. Mercado, 1♂ (CZUG); Parque ecológico Huilotán, 20°57’42.804” N, 103°24’5.292” W, 1,121 m, 7-IX-2023, J. A. Zamarripa-Fernández, C. Morales & J. M. Sandoval, 3♀ 4♂ (CZUG). Morelos: Jojutla de Juárez, Cerro del Higuerón, 18°34’0.3876” N, 99°9’55.2816” W, 1,060 m, 26-VII-1981, C. Deloya, 1♀ (MZFC 28612); Yautepec de Zaragoza, 18°50’55.6116” N, 99°4’42.0888” W, 1,249 m, 15-XI-1980, J. M. Labougle, 4♀ (CNIN 3042-3045) 2♂ (CNIN 3046; 3047). Oaxaca: Santo Domingo Tehuantepec, 17 km W of Tehuantepec, 16°21’59.328” N, 95°22’59.088” W, 8-IX-1965, D. Janzen, 2♂ (SEMC 1254910; 1254911).
Distribution. This species is endemic to Mexico and distributed from the center of country to the north of the Isthmus of Tehuantepec (Fig. 10) between 1,060-1,616 m of altitude, a distribution that corresponds to the Trans-Mexican Volcanic Belt and Sierra Madre del Sur.
Figure 3. Hind leg, male: A, Mesoplia (M.) insignis. B, Mesoplia (M.) tica sp. nov. C, Mesoplia (M.) sapphirina. D, Mesoplia (M.) dugesi. E, Mesoplia (M.) rufipes. F, Mesoplia (E.) carmelitae sp. nov. G. Mesoplia (E.) ornata.
Taxonomic comments. This species is morphologically similar to Mesoplia tica sp. nov. Males differ in the apex of T7 slightly forked with both apices truncated and bare at tip (Fig. 7A), and the pubescence of the mesosoma almost entirely bright yellow (Fig. 6). The female pygidial plate has a broadly quadrangular apex, with blackish pubescence on the surface (Fig. 4A), and the scutellar tubercles are less projected upward than in Mesoplia tica sp. nov.
Diagnosis. Male: metallic greenish body, with bright yellow pubescence on the clypeus, anterior surface of the mesepisternum, pronotal lobe, anterior region of the scutellar disc, posterior region of the scutellum, scutellum and lateral areas of the propodeum (Fig. 11); tegula with black setae on inner anterior and posterior edge; scutellar tubercles conical and strongly projecting upwards, with a perceptible cleft between them; robust forked medial tibial spur, with blackish integument, wide anterior projection and with between 3 and 4 internal teeth; tergal pubescence mainly whitish, with a spot of bright yellowpubescence on both lateral margins of T1 (Fig. 11B); tergal scales metallic greenish-copper; apex of T7 noticeably forked, both apices rounded (Fig. 7B); dorsal lobe of gonostyle triangular and broad projected upwards (Fig. 12C, D).
Female: body dark metallic and blue pubescence, with scales blue-greenish on head and mesosoma, and metallic blue to the metasoma; with patches of whitish pubescence on labrum, anterior lateral edges of the clypeus, behind the antennal insertion, preoccipital region, upper region of the gena, lateral surface of the mesepisternum, lateral margins of the scutum at the level of the tegula, posterior region of the scutellum, lateral apex of T5 and lateral apices of S2-S4 (Fig. 13); black integument on antenna, pronotal lobe, tegula and legs; scape with blue-green scales; tegula with metallic blue scales; scutellar tubercles subconical, strongly projecting upwards (Figs. 13A, D, 35C); middle tibial spur robust with black integument, forked, anterior projection with 2 to 3 internal teeth; pygidial plate with apex more or less rounded, with scales on the surface that do not reach the apex, and black integument (Fig. 4B).
Figure 6. Mesoplia (M.) insignis, male. A, Lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: male. Body length 17.5 mm, forewing length 13.4 mm, head length 4.2 mm, intertegular distance 4.3 mm, T2 length 2.5 mm. Head: mandible with an internal tooth. Labrum subrectangular, concave surface, with a very evident preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles conical, strongly projecting upwards, cleft between them evident. Vertical metanotum and propodeum. Middle tibial spine rounded and short. Medial tibial spur robust forked, anteriorly projecting with 3 to 4 internal teeth. Middle basitarsus with posterior margin smooth and with short and straight flattened distal process. Hind tibia with 2 short spurs at the inner apex. Metasoma: T7 with a flattened surface, noticeably forked apex, with a moderately deep central cleft, medium distance between the apices, both ends rounded. Color: in general, with black integument. Head: labrum, rostrum, scape, pedicel and f1 black, remainder of flagellum greyish. Mandible with apical third yellowish brown, the rest black. Mesosoma: pronotal lobe, tegula, wing veins, legs, middle tibial spur, black. With smoked wings. Forewings without dark spots evident. Metasoma: with black sternum. Surface sculpture: head, labrum with most of surface rugose, by basal edge polished. Mesosoma: tegula with stippling. The propodeum strigulate. Pubescence: black and bright yellow. Head: labrum central surface with white plumose setae and scattered simple black setae, lower edge with a pair of long black tufts, lateral edges with simple black setae, upper edge bare. Clypeus densely covered with bright yellow plumose setae. Supraclypeal area, forehead close to the lower edge of the ocelli and vertex with metallic bluish-green scales. Supraclypeal area with black setae. With a spot of bright yellow setae in the upper part of the paraocular area. Preoccipital region and upper gena region with bright yellow setae. Lower region of the gena with black setae. Mesosoma: anterior surface of the mesepisternum, pronotal lobe, region anterior to the central disc of the scutum, posterior region of the scutellum, metascutum and lateral areas of the propodeum covered almost exclusively by bright yellow setae. The rest covered by black setae. Central disc of the scutum and most of the surface of the tegula with metallic green scales. Both the anterior and posterior inner edges of the tegula with black setae. Naked propodeal triangle. Internal apex of posterior tibia with very evident short blackish setal brush (Fig. 3B). Metasoma: terga covered with metallic greenish-copper scales. Anterior edge of T1 with scattered simple black and white setae, with a spot of yellow pubescence on both lateral margins. Surfaces of T3 to T6 with several series of rows of simple black setae. S2 to S4 with a short white patch of pubescence on the lateral edges. Scattered black sternum setae. S4 with long, branched black setae near the posterior edge and with several rows of simple black setae on the surface. S2 to S4 with a more or less triangular white spot on the lateral margins; dorsal lobe of gonostyle triangular and broad projected upwards.
Allotype: female. Body length 16.8 mm, forewing length 12.6 mm, head length 4 mm, intertegular distance 4.5 mm, T2 length 2.3 mm. Head: mandible with an internal tooth; labrum subrectangular with flat surface, and with a very evident preapical margin; ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles with the conical shape strongly projecting upwards, an evident cleft between them. Vertical metanotum and propodeum. Middle tibial spine pointed and short. Medial tibial spur robust forked, anteriorly projecting with 2 to 3 internal teeth. Medium basitarsus with slightly wavy posterior margin and a flattened curved distal process. Metasoma: pygidial plate with apex more or less rounded. Color: integument black. Head: labrum, rostrum, scape, pedicel and F1 black, remainder of flagellum greyish. Apical third of mandible dark reddish brown with nearby yellowish brown spot, basal half black. Mesosoma: pronotal lobe, tegula, wing veins, legs, middle tibial spur, black. Apex of distal process of middle basitarsus dark reddish brown. Evenly smoked wings. Forewings without obvious dark spots. Metasoma: black sternum. Surface sculpture: head. Rugulous labrum. Mesosoma: finely punctuate tegula. Dotted propodeum. Metasoma: dotted pigidial plate. Pubescence: mainly black, but head covered mainly with white setae. Head: labrum surface with white setae branched and few scattered simple black setae, with a pair of tufts of black setae on the anterior margin and with setae on the lateral edges. Scape almost entire surface of anterior area with metallic blue-green scales. Mesosoma: covered mainly with black setae, except for patches of white setae on the collar of the pronotum, lateral region of the mesepisternum, lateral edges of the scutum, near the tegula, posterior region of the scutellum. Scales of the central disc of the scutum and the pronotal lobe metallic blue-green, of the same tone as those of the face. Scales of the tegula, mesepisternum and scutellum metallic blue. Metasoma: virtually the entire surface covered with metallic blue scales and the setae present are almost exclusively black, except for a spot of branched white setae on the lateral edges of T1, several rows of simple white pubescence on the surface of T4 and T5, a triangular spot of short branched white pubescence on the lateral ends of S3 to S5 and another small spot on the lateral edge of T5. Sterna also with metallic blue scales. Pygidial plate with metallic scales on the surface.
Variation. Some males have bright yellow setae on the external surface of anterior tibia and little pale-yellow pubescence on the lateral edge of S4.
Figure 8. Mesoplia (M.) insignis, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Taxonomic summary
Type material. Holotype: ♂ MNCR INB0004333459, CR. CART (Costa Rica, Cartago), Ochomongo, San Nicolás, Finca Kirqua, 9°54’41.29” N, 83°56’1.81” W, 14-II-2010/1-III-2010, W. Porras (MNCR). Allotype: ♀ MNCR INB0004333299, CR. HER [Costa Rica, Heredia], Sarapiquí, La Isla, 10°24’50.79” N, 84°7’42.24” W, 100-200 m, 22-II-2017/17-IV-2017, I. A. Chacón (MNCR). Paratypes: same data as the holotype ♀ MNCR INB0004333460 (MNCR), ♀ MNCR A5002767; CR. ALAJ [Costa Rica, Alajuela], Virgen del Socorro, 10°17’0.20” N, 84°10’38.22” W, 750 m, 15-IX-1982, I. A. Chacón (MNCR); ♂ MNCR A5057872 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, Estación Biológica La Selva, 10°25’50.61” N, 84°10’38.21” W, 61 m, 24-VI-1988, I. A. Chacón (MNCR), ♀ SEMC 1254906, 10°25’35.81” N, 84°1’1.16” W, 80 m, 19-V-1993, J. Ashe & A. Ashe (SEMC); ♀ MZUCR HY6823 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, La Isla, 10°14’7.70” N, 83°54’43.45” W, 221 m, VI-2011/VII-2011, I. A. Chacón (MZUCR); ♀ CINAT JZ344 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, La Tirimbina, 10°23’52.20” N, 84°8’9.16” W, 186 m, 1997, Sergio (CINAT), ♀ MNCR JZ357, same location as the previous one, 25-VI-1997, Posada (MNCR); ♀ MZUCR HY6827 CR. PUNT [Costa Rica, Puntarenas], Reserva Forestal Golfo Dulce, 24 km W. Piedras Blancas, 8°45’17.43” N, 83°29’5.51” W, 200 m, IV-1992/V-1992, P. Hanson (CINAT).
Etymology. The specific epithet tica refers to the demonym with which the inhabitants of Costa Rica recognize themselves.
Distribution. Registered only for Costa Rica and it is possible that this species be endemic to this country (Fig. 10), present between 61 to 750 m altitude.
Taxonomic comments. This species is morphologically like M. insignis, the main differences are that the female has the scutellar tubercles strongly projecting upwards (Fig. 6A), and the pygidial plate covered by metallic scales and its apex is rounded (Fig. 4B); and the male has the apex of T7 noticeably forked, both apices rounded (Fig. 7B).
Figure 9. Mesoplia (M.) insignis, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Mesoplia (Mesoplia) regalis (Smith, 1854)
Melissa regalis Smith, 1854: 280. Syntypes: British Museum Natural History, London, England (♀♂). Type locality: Brazil, Pará, Santarém.
Diagnosis. Male: body metallic green and pubescence white and brown; reddish brown on antennal integument, pronotal lobe, tegula and legs; scutum and tegula with metallic green scales; scutellar tubercles with a flattened surface, median slit between them; bifurcated middle tibial spur, with integument brown; anterior projection with 2 to 4 internal teeth; anterior edge of T1 with scattered white setae, lateral edges with a brown and white spot; T7 with the flat tips, with setae below it that are noticeable in dorsal view.
Figure 10. Map with the known distribution of Mesoplia (Eumelissa) carmelitae sp. nov., M. (E.) ornata, M. (Mesoplia) dugesi, M. (M.) insignis, M. (M.) oaxacana sp. nov., M. (M.) regalis, M. (M.) tica nov. sp., and M. (M.) veracruzana sp. nov.
Female: body metallic green, with white pubescence; antennal integument, pronotal lobe, tegula and legs reddish brown; scutum and tegulas with scales metallic green; scutellar tubercles more developed than in M. sapphirina with a deep depression between them and tubercles conspicuously pointed the delimiting carina is more developed only along the tubercles gradually fading to the sides; bifurcated middle tibia spur, integument brown; anterior projection with 2 to 4 internal teeth; pygidial plate with rounded apex and metallic scales that do not reach the apex (Melo & Rocha-Filho, 2011).
Taxonomic summary
Material examined. 1♂: Panama: Islas de las Perlas, Isla del Rey, 8°23’34.70” N, 78°54’24.11” W, 139 m, 13-IV-1981, R. W. Brooks, 1♂ (SEMC 1254921).
Distribution. Mainly South American, with the northern limit in Panama (Moure & Melo, 2023).
Taxonomic comments. According with Melo and Rocha-Filho (2011), this is a species similar morphologically to M. sapphirina, the main difference is that in this species the tubercles of the scutellum have the distal edge projecting upwards, more evident in females; the dorsal surface of the T7 in males bulging, with flat apices and with setae inserted underneath, distinguishable even in dorsal view. Specimens from Brazil deposited in SEMC were analyzed, whose distribution data were not included because they are from a distribution area not included in this study.
Mesoplia sapphirina Melo & Rocha-Filho, 2011: 30. Holotype: Department of Zoology, Universidade Federal do Pará, Curitiba, Brazil (♀). Type locality: Panamá, ‘Old Panama’.
Figure 11. Mesoplia (M.) tica sp. nov. Holotype, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Diagnosis. Male: body metallic blue, with whitish setae (Fig. 14); F1 light reddish brown; remainder of flagellum, pronotal lobe, tegula, most of legs and sterna dark reddish brown; scutum and tegula with metallic blue scales; scutellar tubercles poorly developed, posterior carina conspicuous; middle tibial spur forked, both apical branches with robust teeth, outer branch with 2 small internal teeth (Fig. 16A); dorsal surface of T7 bulging, its projected ventral portion conspicuously excavated; with a pair of apical projections of T7 with rounded apices, emargination between them relatively deep (Fig. 7C); dorsal and digitiform lobes of the gonostylus well developed (Fig. 15C, D).
Female: body metallic blue, with whitish setae (Fig. 17); F1 light reddish brown; remainder of flagellum, pronotal lobe, tegula, most of legs and sterna dark reddish brown; scutellar tubercles weakly developed, its surface only slightly convex and its tubercles low and broadly rounded; the carina that delimits the tubercles posteriorly well developed and runs continuously from one side of the scutellum to the other; middle tibial spur forked, both apical branches with robust teeth, outer branch with 2 small internal teeth (Fig. 18A); pygidial plate with oval apex and covered with decumbent bluish metallic scaly hairs, except for the apex which is naked (Fig. 4C).
Figure 12. Mesoplia (M.) tica sp. nov. Holotype male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Variation. Some males have yellow setae on the posterior surface of the scutellar tubercles.
Taxonomic summary
Material examined. Costa Rica: Cartago: Cachí, 9°49’52.06” N, 83°48’18.12” W, 1,016 m, 28-V-1974, G. Rivera, 1♀ (MZUCR HY6608). Guanacaste: Bagaces: Barrio el Brasil, 10°32’57.36” N, 85°15’54.11” W, 114 m, 5-V-2022, K. Borbón, 1♀ (CINAT JZ340), 3♂ (CINAT JZ341 – CINAT JZ343). 10°32’58.60” N, 85°15’52.42” W, 110 m, 8-IV-2023, K. Borbón, 1♀ (MZUCR HY6131), 1♂ (MZUCR HY6131). Filadelfia, 10°26’51.27” N, 85°32’59.86” W, 22 m, 28-IX-1959, M. Leiva, 1♂ (MZUCR HY6816); La Cruz: Cuajiniquil, Bahía Tomás, Salinitas, in mangrove, 10°55’15.60” N, 85°42’57.96” W, 0 m, 28-I-2022, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2249), 7-II-2022/7-III-2022, M. M. Chavarría & P. Hanson, 2♀ (MZUCR HY5067), (MZUCR HY5068), 25-III-2021, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2345), V-2022, M. M. Chavarría & P. Hanson, 1♀ (MZUCR HY6838); Nicoya: Sámara, 9°52’51.60” N, 85°31’36.30” W, 17-I-2009, C. Reichie, 1♀ (MZUCR HY5051). San José: San Pedro de Montes de Oca, 9°56’11.41” N, 84°2’58.50” W, 1,201 m, 20-VI-1959, Y. Gómez, 1♀ (MZUCR HY5061). Mexico: Chiapas: La Concordia, 16°7’12.00” N, 92°39’0.00” W, 616 m, 1-IV-2016, P. Sagot, 1♀ (ECOAB 58710), 16°3’36.00” N, 92°33’36.00” W, 550 m, 5-III-2020, P. Sagot, 1♂ (ECOAB 134687); Mazatan, 14°55’12.00” N, 92°37’48.00” W, 6 m, 2-II-2019, P. Sagot, 1♂ (ECOAB 130647), 14°55’48.00” N, 92°37’12.00” W, 8 m, 2-II-2019, I. Pérez, 1♂ (ECOAB 130665); Tonalá, 15°30’0.00” N, 92°24’0.00” W, 3 m, 21-IV-2011, 1♀ (ECOAB 56616). Jalisco: La Huerta: Chamela, 19°29’54.77” N, 105°2’41.22” W, 10-IX-1981, S. Bullock, 1♀ (EBCH SB736), 19-X-1985, S. Bullock, 1♀ (SEMC 1451101), 27-IX-1985, R. Ayala, 1♀ (SEMC 1451080); Estación Biológica Chamela, 19°29’54.77” N, 105°2’41.22” W, 25-IX- 1985, C. D. Michener, 1♀ (EBCH JZ65); La Manzanilla, 19°17’0.03” N, 104°47’13.06” W, 1 m, 16-XI-1994, A. Moreno, 1♀ (HFL 302), 1♀ (CZUG 302), 16-XI-1994, H. E. Fierros-López, 1♀ (HFL JZ7), 1♂ (HFL JZ8), 1♀ (CZUG 309); Playa Careyitos, 19°29’36.18” N, 105°1’29.56” W, 27-X-1985, R. Ayala, 1♀ (EBCH RA219), 27-IX-1985, R. Ayala, 1♀ (EBCH RA218), 30-IX-1985, C. D. Michener, 1♀ (SEMC 1451078); Playa Careyitos 6 Km. South of Chamela, 19°29’36.18” N, 105°1’29.56” W, 27-IX-1985, A. Rodríguez P., 1♀ (EBCH JZ63), R. B. Roberts, 1♀ (EBCH JZ64); Playa Teopa, 19°23’48.69” N, 105°1’9.27” W, 4-X-1985, J. G. Rozen 1♀ (EBCH JZ62), R. Ayala, 1♀ (EBCH JZ69). Oaxaca: San Pedro Huamelula, 15°55’48.00” N, 95°46’48.00” W, 22 m, 21-XII-2019, P. Sagot, 1♂ (ECOAB 133378). Sinaloa: San Ignacio, Autopista 4 Río-Piaxtla, 23°55’59.28” N, 104°25’59.88” W, 1,184 m, 18-III-1990, W. Bell, D. Conlon & R. L. Mickley, 1♀ (SEMC 1451099). Tabasco: Tenosique, 17°18’0.00” N, 91°10’12.00” W, 126 m, 23-V-2018, A. Tapia, 1♀ (ECOAB 120153). Tamaulipas: Altamira, Playa Altamira, 22°29’59.53” N, 97°51’41.04” W, 3-VI-1985, M. S. Wasbauer & J. E. Slansky, 1♀ (SEMC 1442315); Llera de Canales, 23°19’24.79” N, 99°1’19.66” W, 267 m, 19-VII-1954, Univ. Of Kans. Mex. Exped., 1♂ (SEMC 1451076); Soto la Marina, La Pesca, 23°46’31.53” N, 97°44’21.63” W, 4 m, 1-VII-1981, B. Miller, B. Porter & L. Stage, 6♀ (SEMC 1451070 – SEMC1451075); Tampico, 22°13’57.47” N, 97°53’30.53” W, 2 m, 10-VI-1951, H.E. Evans, 6♀ (SEMC 1451089 – SEMC 1451094), 1♂ (SEMC 1451088). Veracruz: without coordinates, 25-VII- 1995, 1♀ (CERUADY 239ver). 25-VII-1995, 1♂ (CERUADY 230ver); Actopan: Cofre de Perote, 19°35’45.96” N, 96°22’47.28” W, 10 m, VI-2019, P. Luna & W. Dáttilo, 1♀ (IEXA JZ38); La Mancha, 19°21’12.60” N, 96°13’27.48” W, 3 m, VI-2022, B. Ratoni & W. Dáttilo, 1♂ (EBCH 622 1), 19°21’16.92” N, 96°13’29.64” W, 15 m, VI-2022, B. Ratoni & W. Dáttilo, 1♀ (EBCH 623 7), 1♂ (EBCH 623 6), V-2023, C. Pinilla, 2♂ (EBCH JZ355, EBCH JZ356). Boca del Río: 8 km SE of Boca del Río, 19°3’13.58” N, 96°2’42.53” W, 9 m, 22-VII-1990, W. Bell, D. Conlon & R. L. Mickley, 1♂ (SEMC 1451100); San Andrés Tuxtla: Estación biológica Los Tuxtlas, 18°31’0.00” N, 95°7’59.59” W, 136 m, 16-IX-2018, I. Huerta, 1♀ (CZUG JZ 13); Tecolutla: 20°28’47.00” N, 97°0’36.00” W, H. E. Evans, 1♀ (SEMC 1451095), 2.41402 km N of Tecolutla, 20°29’33.25” N, 97°1’34.21” W, 4.572 m, 12-VI-1985, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451077). Yucatán: Tizimín: El Cuyo, 21°31’5.99” N, 87°40’59.00” W, 25-VII-1995, H. Moo, 2♀ (CERUADY 009406(02450)), (009411(02477)), 1♂ (009414(02451)), R. Tut, 7♀ (CERUADY 009574(02453), (009546(02454)), (009543(02455)), (009539(02456)), (009541(02474)), (009545(02475)), (009536(02476)), 1♂ (009544(02461)), 14-VI-1995, R. Tut, 8♀ (CERUADY 007998(02452)), (007987(02462)), (007992(02457)), (007986(02465)), (007980(02466)), (007982(02467)), (008004(02469)), (008000(02473)), 5♂ (007996(02458)), (007979(02460)), (007994(02463)), (007983(02472)), (008006(02479)), 24-V-1995, R. Tut, 1♀ (006649(02468)), 1♂ (006651(02478)), H. Moo, 1♂ (006844(02464)), 4-IV-1995, R. Tut, 1♀ (005166(02470)), 1♂ (005175(02459)). Nicaragua: León: Campus Agropecuário UNAN-León, 12°25’22.14” N, 86°57’8.68” W, 16-V-2022, R. Varela, 1♀ (LEECAV JZ292). Panama: Chimán: Brujas: Punta Bruja, Zona del Canal, 8°35’6.51” N, 78°31’56.69” W, 4 m, 25-I-1929, C. H. Curran, 1♂ (SEMC 1451102). Panamá: Panamá Viejo, 9°0’22.09” N, 79°29’6.29” W, 5 m, 16-IV-1945, C. D. Michener, 1♂ (SEMC 1451079). Panamá Oeste: Fort Kobbe, 8°55’0.30” N, 79°34’59.59” W, 8 m, 11-I-1960, W. J. Hanson, 2♀ (SEMC 14510972), (SEMC 1451098), 2♂ (SEMC 1451096), (SEMC 1427034).
Figure 13. Mesoplia (M.) tica sp. nov. Allotype, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Distribution. The northernmost distribution has been recorded in Sonora, along the Pacific Ocean slope and in Tamaulipas on the Gulf of Mexico slope, to the south the limit of its distribution it is in the center of Panama (Fig. 19).
Taxonomic comments. It is a species morphologically similar to M. regalis, the main difference is that the apices of the scutellar tubercles are rather flattened (Fig. 16A, D), not projected upwards (this character can be confused if the scales in this area of the body have been lost), while males do not present setae that protrude in dorsal view below T7 and the apices of this are rounded (Fig. 7C).
Diagnosis. Female: body metallic blue, with pubescence mostly blackish, but bright yellow setae on head and mesosoma, especially on the pronotum, mesepisternum, posterior region of the scutellum, metanotum and lateral areas of the propodeum (Fig. 20); integument black in antenna, pronotal lobe, tegula and legs; posterior apex of scutellar tubercles projecting upwards (Fig. 20C); middle tibial spur forked, with black integument, the anterior projection with 3 inner teeth; tergal pubescence mainly dark with a bright yellow spot on both lateral margins of T1 and T2 (Fig. 20A); pygidial plate with rounded apex, but with a notch in the medial part of the apex, with scales that does not reach the apex, and with black integument (Fig. 4D).
Figure 14. Mesoplia (M.) sapphirina, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: female. Body length 17.9 mm, forewing length 12.6 mm, head length 5 mm, intertegular distance 4.2 mm, T2 length 2.5 mm. Head: mandible with an internal tooth. Labrum subtriangular, surface slightly concave, with a raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: scutum convex in lateral view. Scutellar tubercles not very evident due to the large amount of pubescence, a noticeable cleft between them. Metanotum and propodeum vertical. Spine of the middle tibia long pointed. Medial tibial spur robust and forked, anterior projection with 3 internal teeth. Medial basitarsus with smooth posterior margin and a flattened distal process that is slightly curved externally. Metasoma: broad pygidial plate at the base, apex subtruncate with a slight central notch. Color: integument blackish. Head: black on face, labrum and antenna; the flagellum with integument opaque. Apical third of mandible yellowish brown, except for black teeth. Mesosoma: almost all black. Apex of distal process of middle basitarsus reddish brown. Wings overall evenly smoked (Fig. 20A, B); forewings with the anterior edge of the marginal cell darker. Veins dark. Metasoma: between the terga and sterna reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Reticulated propodeum. Metasoma: dotted pigidial plate. Pubescence: mostly bright yellow and black. Head: labrum discal area with white plumose setae and scattered simple black setae, lower edge with a pair of long dark tufts, lateral edges with simple black setae, upper edge bare. Clypeus with lower margin with white plumose setae, sides with pale yellow plumose setae, central surface with light metallic blue scales. Suplaclypeal area with few long and scattered black simple setae and with scales equal to those of the clypeus. Frons with black setae and light metallic green scales near the lower edge of the ocelli. Paraocular area with pale yellow plumose setae. Vertex and preoccipital region with bright yellowcompound setae. Gena with setae bright yellow, except for the lower region that has black. Mesosoma: pronotal lobe with the anterior and external lateral margin with bright yellow setae, posterior margin with interspersed white and black setae, inner margin with mainly black setae, surface with dark metallic blue scales (Fig. 20). Mesepisternum upper region with bright yellow setae, lower region black. Scutum covered mainly by black setae, except for the lateral edges near the posterior two-thirds of the tegula that have interspersed bright yellow setae, central disc with light metallic green scales. Tegula black on the anterior and inner edge and with dark metallic blue scales on the surface. Upper of scutellum with few black setae and light metallic blue scales and bright yellow on posterior region, that projects a little between the tubercles. On the metanotum bright yellow. Propodeum upper region naked, posterolateral region bright yellow. Legs are mainly dark and with dark metallic blue scales on the external surfaces, with a tuft of white setae on the posterior margin of the femurs of the forelegs and on the external apices of the middle femurs. Metasoma: terga covered with metallic bluish-green scales. Anterior edge of T1 with scattered white setae, lateral edges with a bright yellow spot; T2 with a smaller pale-yellow spot in the lateral marginal area. T4 and T5 with few white simple setae in lateral view. T6 with thick black setae on the sides of the pygidial plate. Pygidial plate with light metallic green scales that do not reach the apex. Sterna with scattered black setae. S2 to S4 with a sub triangular white spot on the lateral margins.
Figure 15. Mesoplia (M.) sapphirina, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Male: unknown.
Taxonomic summary
Type material. Holotype: ♀ IEXA JZ39 MEX. VER [Mexico, Veracruz], Coatepec, 19°26’42.36” N, 96°57’52.13” W, 1,200 m, 6-VI-2020, G. Quintos-Andrade (IEXA). Paratypes: 3♀: UNAM-LT JZ352 MEX. VER [Mexico, Veracruz], San Andrés Tuxtla, Estación Biológica Los Tuxtlas, 18°35’5.50” N, 95°4’26.58” W, 160 m, 1985-VII-6, A. Ibarra (UNAM-LT). Same location as the previous one, 2♀ UNAM-LT JZ353, 1985-VI-2, P. Sinica. UNAM-LT JZ354, 1986-VII-20, P. Sinica.
Etymology. The name veracruzana refers to the known distribution of this species, the state of Veracruz, Mexico.
Distribution. Only known for the localities of Coatepec and San Andrés Tuxtla (Fig. 10), although there are photographic records in iNaturalist near Xalapa and Orizaba, localities with cloud forest and tropical forest.
Taxonomic comments. The female of this species has a pubescent color similar to that in the male of M. insignis, so they could be confused. In this species, the bright yellow pubescence on the metasoma is only located in the pronotum, mesepisternum, posterior region of the scutellum, metanotum and lateral areas of the propodeum (Fig. 20), unlike M. insignis whose metasoma is almost completely covered by yellow setae (Fig. 6).
M. azurea group(sensu Moure, 1960a, 1960b)
Diagnosis. Male: hind tibia with an inconspicuous setal brush and with 1 spur; ventral basal surface of posterior femur with a robust posterior projection (Fig. 3D, E); inner surface of basitarsus with a longitudinal carina. Female: with carina extending laterally from the pronotal collar, to the pronotal lobe (Fig. 5B).
Mesoplia (Mesoplia) dugesi (Cockerell, 1917)
(Figs. 1B, 3D, 4E, 5B, 7D, 10, 16B, 21-23, 35E)
Mesonychium dugesi Cockerell, 1917: 477. Holotype: National Museum of Natural History, Smithsonian Institution, Washington DC, United States of America (♂). Type locality: Mexico, Guanajuato.
Diagnosis. Male: body dark metallic blue, pubescence mainly black (Fig. 21), except for some white spots on the central surface of the labrum, anterior edge of the superior gena and clypeus; integument blackish on antenna, pronotal lobe, tegula and legs, except for the F1 which is usually dark reddish brown; tegula with metallic blue scales; wing with black veins; scutellar tubercles practically flattened and without a cleft between them; middle tibial spur forked with dark reddish-brown integument, anterior projection with 1 to 3 internal teeth (Fig. 16B); femur posterior with a robust posterior projection on the ventral basal surface (Fig. 3D); apex of T7 bifurcated, both with an acute angle and close to each other (Fig. 7D); dorsal lobe of gonostylus reduced (Fig. 22C, D).
Female: body dark metallic blue, with dark pubescence at first glance (Fig. 23), in a deeper examination some areas with whitish pubescence can be seen on labrum, superior area of the gena, internal margin of the pronotal lobe and a small spot on the posterior surface of the scutellum and between the tubercles; integument of antenna, pronotal lobe, tegula and legs blackish (dark reddish); labrum, vertex, preoccipital area and tegula with metallic blue scales; with a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5B); wings with black veins (Fig. 23A, B); scutellar tubercles conical, slightly projecting with a shallow cleft between them (Fig. 23D); middle tibial spur forked with dark reddish-brown integument, anterior projection with 3 internal teeth; T1 with black setae; terga with metallic blue scales; pygidial plate with a narrow apex and completely covered by metallic bluish-green scales (Fig. 4E).
Description. Female: body length, 16.4 mm, forewing length 12.7 mm, head length 4.1 mm, intertegular distance 3.9 mm, T2 length 1.7 mm. Head: mandible with an internal tooth. Labrum subrectangular, surface slightly concave, with a raised preapical margin. Ocelli aligned with the upper margin of the eyes, protruding slightly above the head in lateral view. Mesosoma: with carina that runs laterally from the collar of the pronotum to the pronotal lobe. Convex scutum in lateral view. Scutellar tubercles rounded, without posterior carina, with a median slit between them. Metanotum and propodeum vertical. Medial tibial spine short rounded. Medial tibial spur bifurcated, anterior projection with 3 internal teeth. Medial basitarsus with pentadentate external margin and with a flattened distal process that is slightly curved externally. Metasoma: broad pygidial plate at the base, narrow rounded apex. Color: integument blackish to dark reddish brown. Head: labrum with reddish brown lateral and anterior edges, blackish surface. Apical region of mandible blackish (yellowish brown) reddish brown in basal region. Scape and pedicel blackish to reddish brown. F1 reddish brown, remainder of flagellum brown. Mesosoma: pronotal lobe and tegula dark reddish brown. Almost transparent wings. Forewings with the anterior edge and the distal apex of the marginal cell darker, also presenting a darker spot that goes from the external edge of the 2r-m to the wing apex (Fig. 23B). Dark veins. Inner surfaces of legs dark reddish brown, outer surfaces blackish. Apex of the distal process of the medial basitarsus ferruginous and medial tibial spur reddish brown. Metasoma: blackish sterna, except for S6 which is reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Strigulate propodeum. Metasoma: dotted pigidial plate. Pubescence: mostly blackish. Head: labrum central surface with white feathery setae and scattered simple black setae, lower edge with a pair of long dark tufts, lateral edges with simple black setae, upper edge bare. Clypeus and supraclypeal area with metallic blue scales and long black simple setae. Scape with short black setae. Frons with black setae and metallic blue scales on the lower edge of the ocelli. Paraocular area mainly with simple black setae, with a tuft of setae near the antennal alveolus. Vertex and preoccipital region with simple black setae and metallic blue scales. Gena with black setae, except for the middle region near the eyes which has white setae. Mesosoma: with black setae except for the inner margin of the pronotal lobe and a small spot on the posterior surface of the scutellum and between the tubercles. Pronotal lobe with metallic blue scales. Anterior region of the scutum with dense setae coverage. Central disc with scattered setae and light metallic blue scales. Tegula with black setae on the anterior edge and metallic blue scales on the inner edge. Scutellum upper region with few black setae and metallic blue scales, posterior region also with metallic blue scales. Naked propodeal triangle. Legs are mainly black, with only a patch of white setae on the external apex of the anterior tibia and on the external apex of the middle femur. External surface of the tibiae of all legs with metallic blue scales and on the posterior basitarsus. Metasoma: with black setae except for the lateral margins of S2 to S4 which have a triangular spot of short white setae. Sterna with scattered black setae.
Variation. Some males have a greater proportion of white setae in some body regions, on all margins of the pronotal lobe, in the anterior region of the scutum, anterior and posterior apex of the tegula, between the tubercles and in the posterior region of the scutellum, metanotum and on the supralateral margins of the propodeum.
Taxonomic summary
Material exanimated. Mexico: Jalisco: Mazamitla, Puerta del Zapatero, 19°50’29.00” N, 103°5’52.00” W, 1,615 m, 27-VIII-2005, H. E. Fierros-López, 1♂ (HFL); Teocuitatlán: Laguna de Sayula, 19°56’55.00” N, 103°27’56.00” W, 1,350 m, 28-VIII-2011, A. Aguilar, 1♂ (HFL 1111); Zapopan: Barranca la Experiencia, 20°44’6.13” N, 103°19’38.38” W, 1,200 m, 23-VIII-1997, G. Mercado, 1♀ (HFL 1107), 23-VIII-1996, G. Mercado, 1♂ (HFL), 14-IX-1996, G. Mercado, 1♂ (CZUG 1108), 1,350 m, 27-IX-2009, G. Mercado, 1♀ (CZUG). Michoacán: Nahuatzen: Zacapu, 19°45’22.46” N, 101°46’52.61” W, 2,152 m, 10-IX-2014, G. Mérida, 1♂ (ECOAB 25661).
Distribution. Only known from Jalisco, Guanajuato and Michoacán in México, although there is at least one record in iNaturalist for Querétaro (Fig. 35E). Its distribution seems to be restricted to the center of the country (Fig. 10).
Taxonomic comments. Is the darkest species distributed in the region, both the general pubescence of the body and the integument of the legs are mainly blackish.
Mesoplia guatemalensis Cockerell, 1912: 567. Holotype: National Museum of Natural History, Smithsonian Institution, Washington D.C., United States of America (♀). Type locality: Guatemala, Gualan. New junior synonym.
Diagnosis. Male: body metallic blue-greenish coloration and with whitish pubescence (Fig. 24); integument of the scape, pedicel and f1 ferruginous (in some specimens inner margin of the scape darker); integument ferruginous on apical flagellomere in posterior view, pronotal lobe and tegula; integument of legs dark brown, on external surface and ferruginous on internal; tegula without scales (Fig. 24B); wing veins ferruginous; with rounded scutellar tubercles, with posterior carina and no cleft between them; femur posterior with a robust posterior projection on the ventral basal surface (Fig. 3E, 35F); terga with blue-green scales; apex of T7 bifurcated, both with an acute angle and close to each other (Fig. 7E); dorsal lobe of gonostylus reduced (Fig. 25C, D).
Female: body metallic blue-greenish coloration and with whitish pubescence (Fig. 26); integument ferruginous on scape, pedicel and f1, pronotal lobe, tegula and legs (some populations with dark brown integument); with a carina extending laterally from the pronotal collar to the pronotal lobe; tegula without metallic scales (Fig. 26B); wing veins ferruginous; conical tubercles slightly projected with a shallow cleft between them; terga with dorsal pubescence whitish; middle tibial spur bifurcated with ferruginous integument, anterior projection with 2 or 3 internal teeth (Fig. 18B); pubescence of the anterior edge of T1 whitish; terga with green scales; pygidial plate with a narrow apex and covered by metallic bluish-green scales (Fig. 4F).
Notes made by Moure (1960a) from the holotype: “The pubescence of the clypeus and paraocular area, especially in the upper part, tends to very pale yellow. With few dark hairs in the supraclipeal area, next to the ocelli and on the frons. The pubescence of the thorax is pale on the pronotum, with dark setae on the lobes. Dark setae in the anterior third of the mesoscutum, but divided in the middle by a line of white setae and bordered posteriorly by white setae. Mesoscutum disc with fine bronze-green scales and few setae; The punctuation is very evident there in two sizes, the small denser dots and the thick ones very dispersed. On the mesepisternum there is white pubescence above and below an irregular disc area of dark setae and ventrally the setae are also dark. The terga are covered with bronze-green scales and in certain light give the impression of being dark at the base of each tergum, each side of the terga with a small longitudinal line of white setae on the border with the ventral surface which is covered with setae quite short white ones. The long setae of T1 toT5 are white, in the last two black. The integument of mandibles, labrum and legs is rusty; the tegulae and pronotal lobes of a lighter rust color.
Figure 20. Mesoplia (M.) veracruzana sp. nov. Holotype, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
The spur of the middle tibia widened towards the apex and bifurcated, with the inner ramus having a single tooth and the outer ramus having a toothed inner edge. The middle basitarsus with the posterior angle between the external and internal surfaces only edged, not laminated. Hind femurs thickened near base projecting backwards at a large finger-like angle. The tibia moderately thickened at the apex, internally more or less flattened, distally on the internal surface a short imperfect tuft and with a relatively short spur. The posterior basitarsus with the inner side flattened and smooth, largely glabrous and separated from the posterior surface by a thin carina. T7 with the apex narrowly emarginated in a paraboloid shape, the tips relatively close together and strongly rounded. The S4 widely emarginated, procurved, with a wide short band ending at the level of the edge. S5 with deeper emargination, very pubescent, hairs curved towards the center. Approximate length 13.2 mm, forewings including tegula 13 mm; head width 4.28 mm, abdomen 5.45 mm.”
Variation.The shape of the S8 seems to be higly variable; and the shape of the basal projection of the posterior femur on males, in some individuals it is wider, while in others it is narrower.
Material examined. Costa Rica: no location data, 1♂ (MZUCR HY6836). Guanacaste: Bagaces, 8 km al NE de Bagaces, Pan Am Hwy, 10°32’17.67” N, 85°19’53.58” W, 116 m, 23-II-2000, P. Ronchi & G. W. Frankie, 1♂ (MZUCR HY5770), 24-II-2000, P. Ronchi & G. W. Frankie, 2♂ (MZUCR HY3446), (MZUCR HY3448), 12-II-2000, P. Ronchi & G. W. Frankie, 2♂ (MZUCR HY3450), (MZUCR HY3447), 9-III-2000, P. Ronchi & G. W. Frankie, 1♂ (MZUCR HY6835); Urban Bagaces, 10°31’39.84” N, 85°15’25.26” W, 83 m, 12-III-2004, L. Sandoval & G. W. Frankie, 2♂ (MZUCR HY6825), (MZUCR HY6824); Hacienda Monteverde 9 km NW Bagaces, 10°33’17.91” N, 85°16’12.50” W, 91 m, 24-II-1996, G. W. Frankie, 1♀ (MZUCR HY5697), 18-III- 2022, J. Lobo, 1♀ (MZUCR HY6828); Carrillo: Filadelfia, 11.2654 SW of Filadelfia, 10°22’21.18” N, 85°37’29.14” W, 85 m, 3-III-1954, A. Wille & H. Daly, 1♀ (SEMC 1451159); Hojancha: Lajas, 9°59’34.55” N, 85°27’5.06” W, 200 m, 18-V-2014, S. Galbraith, 2♀ (INB0004444317 – INB0004444319); La Cruz, Cuajiniquil, Bahía Tomás, Salinitas, in mangrove, 10°55’17.40” N, 85°42’57.96” W, 0 m, 25-III-2021, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2792), 7-II-2022/7-III-2022, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY5064), 25-III-2021, M. F. Otárola & J. Lobo, 2♀ (MZUCR HY2348), (MZUCR HY2349), 18-III-2022/-IV-2022, M. M. Chavarría & P. Hanson, 1♀ (MZUCR HY5065), 9-IV-2022/7-V-2022, M. M. Chavarría & P. Hanson, 3♀ (MZUCR HY5063), (MZUCR HY5066), (MZUCR HY5066); Liberia, 10°38’55.46” N, 85°26’39.36” W, 149 m, 21-II-2013, 1♀ (MZUCR HY3449); Barrio La Arena, 10°36’21.96” N, 85°25’30.23” W, 142 m, 15-V-1980, E. Ponce, 1♂ (MZUCR HY6817); Pan Am Hwy, 10°38’32.38” N, 85°27’13.36” W, 132 m, 22-II-2004, G. W. Frankie & S. B. Vinson, 1♂ (MZUCR HY5696); Santa Cruz, Playa Junquillal, 10°10’7.04” N, 85°48’47.87” W, 3 m, 3-III-1976, R. M. Bohart, 1♀ (SEMC 1442317). Heredia: CINAT Lagunilla, 9°58’23.58” N, 84°7’45.55” W, 1,038 m, 30-VII-1997, A. Ortiz, 1♂ (CINAT JZ337), 27-III-2017, R. Díaz, 1♀ (CINAT JZ339); Sarapiquí: Puerto Viejo, Estación Biológica La Selva (OET), 10°25’49.70” N, 84°0’24.96” W, 50 m, 17-VII-1995, P. Hanson, 1♀ (MZUCR HY6826), 2-IV-1993, P. Hanson & C. Godoy, 1♀ (MZUCR HY6818). Puntarenas: Golfo Dulce, 8°42’13.42” N, 83°29’8.86” W, 10 m, XII-1989/III-1990, P. Hanson, 1♀ (MZUCR HY6821); Gromaco, 34 km S of Potrero Grande en río, 8°50’49.49” N, 83°2’41.43” W, 836 m, 21-VII-1963, C. D. Michener & W. Kerfoot, 1♀ (SEMC 1451161); Rincon, 5 km S of Rincon, 8°40’57.72” N, 83°28’24.94” W, 21 m, 15-III-1973, E. Borrows, 1♂ (SEMC 1451162). San José, 9°40’0.01” N, 84°0’0.00” W, C. D. Michener, 1♀ (SEMC 1451160); Pérez Zeledón, Esperanza, 9°16’30.64” N, 83°42’9.86” W, 807 m, II-1997, N. Siles, 1♂ (CINAT JZ336), 13-IX-1999, A. Ortiz, 1♂ (CINAT JZ338); San Antonio de Escazú, 9°53’23.88” N, 84°8’17.66” W, 1,454 m, W. Edward, 1♂ (MZUCR HY6834). Guatemala: without locality, 28-III-1923, 1♀ (SEMC 1451087). Huehuetenango: La Libertad, 15°34’12.00” N, 91°51’0.00” W, 915 m, 27-IV-2010, M. Hurtado, 1♂ (ECOAB 25637), 28-IV-2010, C. Balboa, 2♂ (ECOAB 25639), (ECOAB 38876), L. Aguilar, 1♂ (ECOAB 25642), 26-II-2010, C. Balboa 2♂ (ECOAB 25643), (ECOAB 25646); San Antonio Huista, 15°36’36.00” N, 91°44’24.00” W, 1,192 m, 23-II-2010, D. Sánchez, 1♂ (ECOAB 25645). Honduras: Ocotepeque: Lucerna, Autopista 4, 9 km N of Lucerna, 14°37’36.00” N, 89°3’42.00” W, 792.48 m, 26-V-2003, D. Brzoska, 1♀ (SMO 418386). Mexico: No location data, J. Mérida, 1♀ (ECOAB 25656). Campeche: Champotón, 18°33’0.00” N, 89°56’24.00” W, 155 m, 8-VI-2018, J. Mérida, 3♀ (ECOAB 121736 – ECOAB 121738), 18°27’0.00” N, 89°53’24.00” W, 168 m, 6-VI-2018, 7♀ (ECOAB 121803 – ECOAB 121806), (ECOAB 121813 – ECOAB 121816). Chiapas: Acacoyagua, 15°24’0.00” N, 92°39’0.00” W, 862 m, 15-V-2007, R. Ayala, 1♂ (ECOAB 56633); Arriaga: 16°19’12.00” N, 93°58’48.00” W, 367.5 m, 10-III-2009, M. Guzmán, 1♂ (ECOAB 25641), 19-V-2009, C. Balboa, 1♀ (ECOAB 25657); Ej. López Mateos, 16°20’46.60” N, 93°58’26.80” W, 255 m, 13-III-2009, C. Balboa, M. Guzmán y M. Cigarroa, 2♂ (EBCH ECO-TAB57347), (EBCH ECO-TAB57276); Cacahoatán, 5 km. sur de Cacahoatán, 14°57’0.00” N, 92°9’0.00” W, 340 m, 18-IV-1993, A. Rodríguez P., 2♀ (EBCH JZ70), (SMO 317151), 2♂ (EBCH JZ77), (SMO 317154); La Concordia, 15°45’0.00” N, 92°58’12.00” W, 921 m, 23-IV-2021, P. Sagot, 1♂ (ECOAB 125000), J. Mérida, 2♂ (ECOAB 125229), (ECOAB 125230), 5-III-2020, P. Sagot, 2♂ (ECOAB 134690), (ECOAB 134691), E. Gómez, 2♂ (ECOAB 134756), (ECOAB 134757), 16°2’24.00” N, 92°33’36.00” W, 580 m, 5-III-2020, P. Sagot, 1♂ (ECOAB 134785); Mapastepec, 15°31’12.00” N, 92°48’0.00” W, 316 m, 22-II-2006, J. Esponda, 1♂ (ECOAB 25644), 1-IV-2005, M. Rincón, 1♀ (ECOAB 25660); Motozintla de Mendoza, 15°22’48.00” N, 92°19’12.00” W, 1,230 m, 7-II-2011, P. Sagot, 1♂ (ECOAB 25638), 15°19’12.00” N, 92°20’24.00” W, 1,000 m, 3-II-2019, P. Sagot, 1♂ (ECOAB 131078); Ocosingo, 16°54’36.00” N, 93°40’48.00” W, 950 m, 15-IV-2010, J. Mérida, 1♂ (ECOAB 56637); San Juan Cancuc, 16°54’0.00” N, 92°21’36.00” W, 1,170 m, 6-IV-2010, J. Mérida, 1♂ (ECOAB 25654); Siltepec, 15°34’48.00” N, 92°39’0.00” W, 1,709 m, 9-III-2022, L. Arroyo, 1♀ (ECOAB 71706), 1♂ (ECOAB 71705); Tapachula, 14°53’24.00” N, 92°17’24.00” W, 127 m, 9-VI-1989, A. Aquino, 1♂ (ECOAB 25648), 14°54’0.00” N, 92°18’36.00” W, 102 m, 14-XII-1988, A. Aquino, 2♂ (ECOAB 25649), (ECOAB 25651), 18-I-1989, A. Aquino, 1♂ (ECOAB 25650), 1♀ (ECOAB 25658), M. Cigarroa, 2♂ (ECOAB 25652), (ECOAB 25653), 1♀ (ECOAB 25659); Tonalá, 16°8’24.00” N, 93°40’48.00” W, 407.5 m, 21-V-2009, C. Balboa, 1♂ (ECOAB 56635), 12-VII-2009, C. Balboa, 1♀ (ECOAB 56636), 9-III-2009, M. Guzmán, 1♀ (ECOAB 25655), 16°30’0.00” N, 93°40’48.00” W, 407.5 m, 12-II-2009, C. Balboa, 1♀ (ECOAB 61187), 15°56’24.00” N, 93°49’12.00” W, 6 m, 31-I-2019, I. Pérez, 1♂ (ECOAB 130806), 25-I-1989, M. Cigarroa, 1♂ (ECOAB:66072); Tuxtla Chico, 14°58’12.00” N, 92°10’48.00” W, 395 m, 18-II-1992, M. Medina, 1♀ (ECOAB 56638), 1♂ (ECOAB:25647); Venustiano Carranza, 16°18’60.00” N, 92°33’59.76” W, 570 m, 21-IV-1993, J. L. Neff, 1♀ (EBCH JZ71). Guerrero: Iguala, 18°20’37.00” N, 99°32’21.01” W, 731.52 m, 4-II-1954, R. R. Dreisbach, 1♂ (SEMC 1451084); Tlapa de Comonfort, 17°33’0.00” N, 98°34’48.00” W, 1,063 m, 17-IV-1989, A. Aquino, 1♂ (ECOAB 56634). Jalisco: Cabo Corrientes, 19°39’0.00” N, 105°10’48.00” W, 49 m, 8-IX-2018, J. Mérida 1♀ (ECOAB 121569); El Tuito, 20°21’20.50” N, 105°18’59.96” W, 18-VI-1989, A. Borquez, 1♂ (EBCH JZ79); Jardín Botánico, 20°27’57.64” N, 105°17’30.89” W, 360 m, 26-VII-2014, C. E. Alatorre-Bracamontes y M. Vásquez-Bolaños, 1♂ (CZUG JZ12); La Huerta: Chamela, 19°29’54.77” N, 105°2’41.22” W, 20-VII-1985, R. Ayala, 2♂ (EBCH RA178bis JZ73), 1♂ (EBCH RA178bis JZ76), 19-XI-1989, G. Rodríguez, 2♂ (EBCH JZ74), (EBCH JZ75); Tequila: Volcán de Tequila, 20°51’50.99” N, 103°50’35.00” W, 1,300 m, 15-III-1996, H. E. Fierros-López, 1♂ (HFL JZ11), 15-III-1995, H. E. Fierros-López, 1♂ (CZUG 344); Zapopan: Barranca la Experiencia, 20°44’6.13” N, 103°19’38.38” W, 30-I-1997, G. Mercado, 1♂ (CZUG 1110), 31-VII-1996, G. Mercado, 1♂ (CZUG 1109). Morelos: Temixo: Cuentepec, 18°51’38.16” N, 99°19’30.72” W, 1,485 m, 31-X-2019, D. A. Hernández-Márquez, 1♀ (MZFC 33199), 18°51’27.72” N, 99°19’40.80” W, 1,455 m, 22-XII-2018, D. A. Hernández-Márquez, 1♀ (MZFC 32554). Querétaro: Jalpan de Serra, 21°13’0.01” N, 99°28’21.00” W, 770 m, 24-V-1989, D. Yanega, 2♀ (SEMC 1451144), (SEMC 1451145). Nayarit: San Blas, 20 km al sur de San Blas, 21°23’25.00” N, 105°6’14.00” W, 10 m, 7-IV-1994, F. Noguera, 1♂ (SMO 328474). Oaxaca: San Pedro Totolapa, 8.04672 km al noroeste de San Pedro Totolapa, 16°43’23.12” N, 96°19’14.27” W, 1,158.24 m, 6-VII-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451086); Putla Villa de Guerrero, 17°0’36.00” N, 97°57’0.00” W, 849 m, 24-V-2020, Wilebaldo, V., 1♀ (ECOAB 58886). Puebla: Cuetzalan del Progreso, 20°3’36.00” N, 97°28’12.00” W, 413 m, 26-III-2020, L. Arroyo, 1♂ (ECOAB 137985). Sonora: Álamos, La Aduana, 27°2’27.16” N, 109°0’57.91” W, 567 m, 15-III-1962, L. A. Stange, 1♀ (SEMC 1442316). Tabasco: Huimanguillo, 17°49’48.00” N, 93°55’12.00” W, 21 m, 3-VI-2015, P. Sagot, 1♂ (ECOAB 77134). Veracruz: without locality, 1♀ (CERUADY 037ver); Acayucan, 48.2803 km al sur de Acayucan, 17°38’6.41” N, 94°54’14.40” W, 56 m, 21-IV-1962, L. A. Stage, 1♀ (SEMC 1442318); Coatepec, 19°26’42.36” N, 96°57’52.13” W, 1,200 m, 1-VI- 2020, G. Quintos-Andrade, 3♀ (IEXA JZ40 – IEXA JZ42), 3-VI- 2020, G. Quintos-Andrade, 1♀ (IEXA JZ43). 4-VI- 2020, G. Quintos-Andrade, 1♀ (IEXA JZ44), 7-VI-2020, G. Quintos-Andrade, 1♀ (IEXA JZ45), 6-VI- 2020, G. Quintos-Andrade, 3♀ (IEXA JZ46 – IEXA JZ48); Medellín: Paso del Toro, 4.82803 km SW of Paso del Toro, 19°0’46.43” N, 96°8’39.30” W, 15.24 m, 23-VI-1961, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451081); Teocelo, Llano Gande, 19°22’13.01” N, 96°52’50.99” W, 833 m, 17-VI-1990, G. Rodríguez, 1♀ (SEMC1451143); Xalapa, 35.4056 km SW of Xalapa, 19°24’20.92” N, 96°38’46.46” W, 335.28 m, 29-VI-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451176); Veracruz, 18°54’29.30” N, 96°8’16.01” W, 27-III-1951, W. P. Stephen, 1♂ (SEMC 1451083). Yucatán: Izamal, Plaza Izamal, 20°56’1.00” N, 89°1’5.00” W, 15 m, 24-V-1995, W. May, 1♀ (CERUADY 22841(02486)), 1♂ CERUADY 22838(02482), A. Canto, 2♂ (CERUADY 22488(02482)), (CERUADY:22481(02484)); Kinchil, Parque Central Kinchil, 20°55’1.23” N, 89°56’50.18” W, 8 m, 24-V-1995, F. León, 1♂ (CERUADY 019671(02480)), 8-V-1995, R. Rodríguez, 1♂ (CERUADY 021772(02481)); Xmatkuil, CCBA, 20°52’2.86” N, 89°37’28.10” W, 11 m, 18-V-2022, A. Sansores, 1♂ (CERUADY JZ206), 16-V-2022, 1♀ (CERUADY JZ211), 1♂ (CERUADY JZ207). Nicaragua: Rivas: Cárdenas, 11°14’24.00” N, 85°42’36.00” W, 90 m, 26-V-2015, E. Dorgay, 1♀ (ECOAB 66669); San Juan del Sur, 11°6’36.00” N, 85°45’36.00” W, 1 m, 6-IV-2017, M. Calero, 1♀ (ECOAB 80382), 0 m, 6-IV-2017, M. Calero, 1♂ (ECOAB 80393). Panama: Colón, 3.21869 km NW of Gamboa, 9°7’42.46” N, 79°42’55.28” W, 65 m, 21-I-1958, W. J. Hanson, 1♀ (SEMC 1451152); 8 km NW of Gamboa, pipeline road, 9°8’38.63” N, 79°43’38.22” W, 61 m, 12-I-1981, C. D. Michener, 1♀ (SEMC 1451153). Panamá: Zona del Canal, Juan Mina, 9°13’39.67” N, 79°57’2.59” W, 106 m, 22-VI-1945, C. D. Michener, 1♂ (SEMC 1451082); Curundú, 8°59’26.73” N, 79°32’27.29” W, 34 m, 31-III-1981, R. W. Brooks, 1♂ (SMO 725157), 19-V-1981, R. W. Brooks, 4♂ (SMO 725158), (SEMC 1254922 – SEMC1254924). Panamá Oeste: Laguna, Chame, 8°39’53.99” N, 80°6’10.90” W, 792.48 m, 29-IV-1945, C. D. Michener, 1♂ (SEMC1451085); Zona del Canal, Río Corona, 8°28’55.22” N, 80°1’42.15” W, 609.6 m, 3-V-1952, C. W. Rettenmeyer, 1♀ (SEMC 1451146); Isla Barro Colorado, 9°11’0.00” N, 79°51’0.00” W, 21-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1451147), 16-IV-1956, C. W. Rettenmeyer, 1♀ (SEMC 1451148), 15-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1254915), 19-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1254916), 7-III-1967, M. Naumann, 1♀ (SEMC 1451149); Fort Kobbe, 8°55’0.30” N, 79°34’59.59” W, 8 m, 11-I-1960, W. J. Hanson, 1♀ (SEMC 1451150), 1♂ (SEMC 1451151).
Figure 22. Mesoplia (M.) dugesi, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Distribution. Registered from south Sonora in northern Mexico, to Brazil, according to opinion of Melo and Genaro (Genaro & Franz, 2008), its distribution is restricted to continental America, so its presence on the islands of the continent would be ruled out (Fig. 27).
Taxonomic comments. Within the genus, it is the species with the widest distribution (Fig. 27), largest number of registered hosts (Table 2). This is possibly a species complex; we did not find morphological characters that allow us to delimit it in a group of species.
Diagnosis. Female: body metallic blue but black in face and mesosoma and metasoma (Fig. 28); scape with dark integument on the base and reddish brown on apex; pedicel with dark integument; integument of f1 ferruginous, remainder of flagellum dark brown; labrum, without metallic blue scales on vertex, occiput and tegula (or not very evident on tegula (Fig. 28B)); tegula integument dark brown; legs with integument on outer surface of blackish and inner surface dark reddish brown; with a carina extending laterally from the pronotal collar to the pronotal lobe; conical scutellar tubercles moderately projected, with a median cleft between them (Fig. 28D); tergal pubescence blackish; middle tibial spur forked with dark reddish-brown integument, anterior projection with 4 internal teeth; pygidial plate with a broad apex and almost completely covered by metallic bluish-green scales (Fig. 4G).
Figure 23. Mesoplia (M.) dugesi, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: female. Body length 13.5 mm, forewing length 12.5 mm, head length 3.5 mm, intertegular distance 4.4 mm, T2 length 2.8 mm. Head: mandible with an internal tooth. Labrum subrectangular, surface slightly concave, with a raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: with a carina extending laterally from the pronotal collar to the pronotal lobe. Scutum convex in lateral view. Scutellar tubercles conical, without posterior carina, with an evident cleft between them. Vertical metanotum and propodeum. Short pointed middle tibial spine. Medial tibial spur bifurcated, anterior projection with 4 internal teeth. Medium basitarsus with tridentate external margin and with a thin flattened distal process slightly curved externally. Metasoma: broad pygidial plate at the base, and with a broad rounded apex (Fig. 4G). Color: integument blackish to dark reddish brown. Head: scape mostly blackish, with dark reddish brown upper outer edge. Pedicel also bicolor. F1 ferruginous, remainder of flagellum brown. Basal and apical region of the mandible dark, with a yellowish brown area in the base of the teeth and reddish brown near the base. Mesosoma: pronotal lobe and tegula blackish. Wings almost transparent. Forewings with the anterior edge and the distal apex of the marginal cell darker, also presenting a darker spot that goes from the external edge of the 2r-m to the wing apex (Fig. 28B). Very dark veins. Inner surfaces of legs dark reddish brown, outer surfaces blackish. Apex of distal process of middle basitarsus blackish. Middle tibial spur reddish-brown. Metasoma: sterna blackish, except for S6 which is reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: dotted pigidial plate. Pubescence: approximately the same proportion of blackish and whitish. Head: labrum discal area with white plumose setae and scattered simple black setae; lower edge with a pair of long dark tufts; lateral edges with simple black setae and upper edge bare. Clypeus and supraclypeal area, with lateral with white plumose setae, discal surface with little pubescence, some elongated black simple setae evident in lateral view. Frons with brown setae. Paraocular area with white plumose setae. Vertex and preoccipital region with white setae. Gena with white setae, except for the darks on lower area. Mesosoma: pronotal lobe, with the inner and posterior margins with white setae, rest with blackish setae, with light metallic green scales, only slightly evident in frontal view. Mesepisternum with blackish setae, with a thick longitudinal white line. Scutum covered mainly by black setae, except for the lateral edges near the posterior two-thirds of the tegula that have white setae, a central disc with light metallic green scales. Tegula with black setae on the anterior margin and white on the posterior, and with few light metallic green scales on the inner margin, only slightly evident in the opposite lateral view. Scutellum, upper area with few black setae and light metallic blue scales, posterior part with white setae project a little between the tubercles, also with light metallic blue scales in posterior view. Metanotum blackish. Propodeum upper region naked, posterolateral region blackish. Mainly black legs. Outer surface of anterior tibia with a few light metallic green scales, some white plumose setae and simple blackish elongated setae and also present on the basitarsus. With light metallic blue scales evident on the outer surface of the tibia and basitarsus of the middle and hind legs. Metasoma: terga covered with metallic blue-green scales and setae almost black and simple, except on the side of T5, S2 to S4. T6, with thick black setae on the sides; pygidial plate with light metallic green scales that do not reach the apex (Fig. 4G); sterna with scattered black setae.
Figure 24. Mesoplia (M.) rufipes, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Male: unknown.
Taxonomic summary
Type material. Holotype: ♀ ECOAB 74052 MEX. OAX [Mexico, Oaxaca], Santiago Nacaltepec, 17°28’12.00” N, 96°49’12.00” W, 1,377 m, 4-XII-2014, L. M. Reyes (ECOAB).
Etymology. The name oaxacana makes reference to the known distribution of this species, the state of Oaxaca.
Distribution. It is only known from the type locality (Fig. 10).
Taxonomic comments. This species is similar to M. dugesi since the majority of the pubescence is dark. But this species has few metallic scales on the face (Fig. 28C) and tegula (Fig. 28B), and the apex of the pygidial plate is broad (Fig. 4G).
Subgenus Eumelissa Snelling & Brooks, 1985
Eumelissa Snelling & Brooks, 1985: 21. Type species: Melissa decorata Smith, 1854: British Museum Natural History, London, England (♀), by original designation.
Diagnosis. Male: posterior tibia, distal inner surface without a conspicuous setal brush (Fig. 3F, G); hind tibial spurs reaching to or beyond middle of basitarsus, both present. Female: pygidial plate narrow, does not completely occupy the exposed dorsal surface of T6; pygidial plate glabrous (Figs. 4H, 23D); dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces.
Diagnosis. Male: body metallic blue, with mostly whitish pubescence; whitish-yellowish pubescence on the face, except for the frons, which has brown pubescence; scape and pedicel with integument dark; basal and apical flagellomere, pronotal lobe, tegula and legs with integument ferruginous; anterior region of the scutellar disc pubescence with a color pattern that goes from blackish in the anterior region, brownish-yellowish to whitish (Fig. 29B); scutellar tubercles flattened; forked spur of middle tibia ferruginous, becoming dark at apex, both branches short, external apex with 3 or 4 internal teeth (Fig. 16D). Terga pubescence whitish, with a pair of white spots with short pubescence in the marginal area of T2 to T6 (Fig. 29D); forked apex of T7, both tips truncated and bare, median cleft shallow (Fig. 7F); S4 bilobed, with a band of very short, dark hairs on the distal margin; dorsal lobe of the gonostylus elongated and projected towards the penis valvae (Fig. 30C, D).
Figure 25. Mesoplia (M.) rufipes, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Female: body metallic blue, with mostly whitish pubescence; F1 shorter than F2; scape and pedicel integument reddish brown; F1, pronotal lobe, tegula and legs integument ferruginous; pubescence of the anterior region of the scutum short and branched, with a characteristic pattern that transversely intersperses spots of whitish and blackish setae (Fig 20B); scutellar tubercles flattened with a superficial cleft between them and with short, bushy black pubescence on the apical edge; terga pubescence whitish; with a pair of white spots with short pubescence in the marginal area of T2 to T4 (Fig. 31A); mesosoma and metasoma, ventral surface with metallic blue scales; middle tibial spur forked with ferruginous integument, becoming dark at apex, both branches short, external apex with a series of 3 or 4 short internal teeth (Fig. 18D); pygidial plate narrow at the base, does not cover the entire posterior margin of T5, glabrous, without scales or hairs (Fig. 4H).
Description. Holotype: male. Body length, 11.3 mm (10.8-11.9; n = 4), forewing length 9.4 mm (9.1-10; n = 4), head length 3.4 mm (3.4-3.5; n = 4), intertegular distance 3.3 mm (2.8-3.5; n = 4), T2 length 1.729 mm (1.6-1.8; n = 4). Head: mandible with an internal tooth. Labrum subrectangular, flat surface, with a slightly elevated preapical margin. Ocelli aligned to the upper margin of the eyes, protruding only a little above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles with a flattened surface, a superficial cleft between them. Vertical metanotum and propodeum. Medial tibial sine short pointed (Fig. 1D). Medial tibial spur bifurcated near the apex, noticeably elongated, internal tip with a single thin elongated tooth, external tip with a series of 3 or 4 internal teeth (Fig. 16D). Medial basitarsus with posterior margin smooth and with short and straight flattened distal process (Fig. 1D). Posterior femur thin. Posterior tibia with 2 spurs at the apex, the inner one longer than the outer one (Fig. 3F). Metasoma: T7 with a flattened surface, bifurcated, with a shallow central cleft, short distance between the apices, both ends truncated (Fig. 7F). Color: blackish to ferruginous integument. Head: ferruginous labrum. Blackish face. Ferruginous scape (some specimens with a reddish spot at the apex). Dark brown pedicel. Ferruginous F1 and F11, remainder of flagellum brown. Apical third of the mandible with apical third blackish (some specimens with yellowish-brown tints), rest dark reddish brown. Mesosoma: ferruginous pronotal lobe and tegula. Blackish mesepisternum. Apex of distal process of middle basitarsus reddish brown. Spur of the middle tibia mainly ferruginous, with blackish apices. Wings almost transparent. Forewings with the anterior edge of the marginal cell darker, also presenting a subtly darkened spot that goes from the outer apex of the marginal cell to the wing apex. Brown veins. Ferruginous legs. Blackish propodeum. Metasoma: ferruginous sterna. Surface sculpture: head. Labrum with most of the surface rugose, the basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: polished T7 apical surface. Pubescence: mainly whitish. Head: labrum discal area with pale yellowish plumose setae; lower edge with a pair of long brown tufts; lateral edges with simple brown setae and upper edge bare. Clypeus and suplaclypeal area, with pale yellowish plumose setae. Frons with brown setae. Paraocular area with pale yellowish plumose and simple setae, some specimens with bright yellow setae in the upper region. Vertex and preoccipital region with white compound setae. Gena with white compound setae, except for the brown on lower area. Mesosoma: pronotal lobe, with yellowish anterior margins, white lateral and posterior margins, practically naked posterior surface. Mesepisternum anterior margin with bright yellow and brown setae, some specimens with the lower portion with a black spot, rest of the surface white except for a spot of black setae below the tegula. Scutum anterior area with a characteristic pattern that goes from blackish, passing through yellowish-brown, until reaching whitish in the region of the discal surface, the latter with metallic blue scales and scattered setae (Fig. 29B). Axila lateral edges with black setae. Tegula with white setae on the inner anterior edge and at the posterior apex, and black on the inner and outer anterior edge, rest of the surface bare. Scutellum upper region with few scattered short black setae and metallic blue scales, scutellar tubercles posterior apices with long black setae and with a line of short, branched white setae between them, posterior area with long white, branched setae (Fig. 29B). Metanotum with white setae. Propodeum upper region naked, posterolateral region whitish-light brown. Legs mainly dark. Anterior tibia outer surface scaleless, with a tuft of short plumose white setae on the outer basal surface, with long simple dark setae running from the outer apex of the tibia and extending along the basitarsus. External surface of middle and posterior tibia and basitarsus with metallic blue scales evident on the external surface. Coxa and trochanter with simple long white setae scattered. Metasoma: terga and sterna with metallic blue scales. Sternum disc with scattered long thin white setae. Anterior edge of T1 with thin white moderately bushy setae, lateral edges with a white patch of short plumose setae. Marginal zone of T2 to T4 with a pair of white spots with short pubescence, from T4 onwards they tend to merge with a lateral spot (Fig. 29D). T7 with simple black setae on the sides. T1 to T6 with a lateral stripe of short, branched setae. Sterna setae scattered black. S4 with short, branched brown setae near the apex and with several rows of simple black setae. S2 to S4 with a more or less triangular white spot on the lateral margins.
Figure 26. Mesoplia (M.) rufipes, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Allotype: female. Body length 12.3 mm (11-14.3; n = 3), forewing length 12.3 mm (9.3-17.5; n = 3), head length 3.5 mm (n = 3), intertegular distance 3.5 mm (3.4-3.6; n = 3), T2 length 1.8 mm (1.6-1.9; n=3). Head: mandible with an internal tooth. Labrum subrectangular, surface flattened, with a subtly raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding only a little above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles flattened, with a superficial cleft between them (Fig. 31A, D). Vertical metanotum and propodeum. Medial tibial spine short pointed. Medial tibial spur notably elongated bifurcated near apex, inner tip with a single slender tooth elongated and curved posteriorly, outer apex has a series of 3 or 4 short inner teeth curved anteriorly (Fig. 18D). Basitarsus medium with smooth posterior margin and with flattened distal process slightly curved externally (Fig. 2D). Metasoma: narrow pygidial plate at the base, forming an angle of approximately 30°, apex rounded (Fig. 4H). Color: blackish to ferruginous integument. Head: scape with dark brown inner edge, rest ferruginous. Pedicel also bicolor, dark brown and ferruginous. Flagellum goes from ferrugous in the basal region to dark brown in the distal region. Apical third of mandible blackish, base dark reddish brown, with a yellowish-brown region at base of teeth extending approximately to end of first third. Mesosoma: pronotal lobe, tegula, upper region of the mesepisternum ferruginous. Wings almost transparents; forewings with the anterior edge of the marginal cell darker, also presenting a subtly darkened spot that goes from the outer apex of the marginal cell to the wing apex. Brown veins. Ferruginous legs. Blackish propodeum. Metasoma: dark pygidial plate. Ferruginous sterna. Surface sculpture: head. Labrum with most of the surface rugose, basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: polished pygidial plate. Pubescence: mostly whitish with some blackish areas. Head: labrum with a thick transverse central band of branched white setae and with few scattered brown simple setae; lower edge with a pair of long brown tufts and some short simple brown setae; lateral edges with simple short brown setae and upper edge bare. Clypeus and supraclypeal area with short white plumose setae, discal area with little pubescence, some simple elongated white setae evident in lateral view. Scape with a tuft of short white plumose setae at the outer edge. Frons with brown setae. Paraocular area with short white plumose setae and some long simple. Vertex and preoccipital region with white compound setae. Gena with white compound setae, except for the lower region that has brown. Mesosoma: pronotal lobe with lateral and posterior margins with short white compound setae, with scattered simple blackish setae (Fig. 31C); anterior surface with metallic blue scales, evident in frontal view; discal surface practically naked. Surface surrounding the pronotal lobe with a thick strip of light brown branched short pubescence. Mesepisternum upper surface mainly with white branched short setae, lower surface with metallic blue scales. Scutum anterior area with short and branched setae, with a characteristic pattern, central area with a thin whitish stripe, followed on both sides by a more or less rectangular blackish spot, these followed by a more or less rectangular whitish spot, after them, a quadrangular blackish spot bordered by whitish setae (the whitish areas have interspersed dark setae); discal area with scattered short simple white setae and metallic blue scales, lateral edges with branched short white setae. Tegula with white setae on the inner anterior edge and at the posterior apex, and black on the inner and outer anterior edge, rest of the surface bare. Scutellum upper region with few scattered short black setae and metallic blue scales; bushy black setae at the posterior apices of the tubercles and with a line of short, branched white setae between them, posterior region with long white, branched setae (Fig. 31B, D). Metanotum with white setae. Propodeum upper region naked, posterolateral region whitish-light brown. Legs mainly dark. Anterior tibia outer surface scaleless, with a tuft of short plumose white setae on the outer basal surface, with long simple dark setae running from the outer apex of the tibia and extending along the basitarsus. External surface of middle and posterior tibia and basitarsus with metallic blue scales evident on the external surface. Coxa and trochanter with simple long white setae scattered. Metasoma: terga and sterna with metallic blue scales. Anterior edge of T1 with white simple long setae, lateral apices with a whitish light brown spot. Marginal zone of T2 to T4 with a pair of white spots with short pubescence, in T4 they tend to merge with a lateral spot. Lateral margins of T2 toT5 and S2 to S4with a strip of short white setae (Fig. 31A). T6 with thick black setae on the sides of the naked pygidial plate. Sterna with scattered black setae.
Variation. Some males from Costa Rica also present yellow setae on the white spots of T4 to T6, the white pubescence spots appear from T1 and they all tend to be fused together.
Figure 27. Map with the known distribution of Mesoplia (M.) rufipes.
Taxonomic summary
Type material. Holotype: ♂ ECOAB 566228, MEX. CHIS [Mexico, Chiapas], Arriaga, 16°13’48.00” N, 93°58’48.00” W, 367.5 m, 10-III-2009, R. Vargas (ECOAB). Allotype: ♀ ECOAB 134689, MEX. CHIS [Mexico, Chiapas] La Concordia, 16°3’36.00” N, 92°34’48.00” W, 550 m, 5-III-2020 P. Sagot (ECOAB). Paratypes: ♀ MZUCR HY6837, CR. GUA [Costa Rica, Guanacaste], La Cruz, Cuajiniquil Bahía Tomás, Salinitas, in mangrove, 10°55’17.40” N, 85°42’57.96” W, 7-III-2022/18-III-2022, M. M. Chavarría & P. Hanson (MZUCR); ♂ MZUCR HY10213, CR. GUA [Costa Rica, Guanacaste], La Cruz, Cuajiniquil, Parque Nacional Santa Rosa, 10°54’19.51” N, 85°46’11.78” W, 80 m, 20-II-2024, M. F. Otárola, M. Solano & M. M. Chavarría (MZUCR), ♂ MZUCR HY10221, 10°54’49.68” N, 85°48’15.41” W, 33 m, 21-II-2024, M. F. Otárola, M. Solano & M. M. Chavarría (MZUCR), ♂ MZUCR HY10257, 10°50’20.40” N, 85°37’4.80” W, 295 m, 7-II-2019, J. Lobo (MZUCR), same data as the previous one, ♀ MZUCR HY10257; ♂ ECOAB 566230, GU. HUE [Guatemala, Huehuetenango], San Antonio Huista, 15°40’12.00” N, 91°57’36.00” W, 1,007 m, 20-II-2010, J. Mérida (ECOAB). Same locality as the holotype, 2♀ ECOAB 566231, 566232, 13-III-2009, C. Balboa (ECOAB); same data as the allotype, ♀ ECOAB 134688 (ECOAB); ♂ ECOAB 566229, MEX. CHIS [Mexico, Chiapas], Tonalá, 16°8’24.00” N, 93°40’48.00” W, 407.5 m, 19-I-2009, C. Balboa (ECOAB); ♂ ECOAB 67237, MEX. CHIS [Mexico, Chiapas], Chiapilla, 16°33’0.00” N, 92°0’0.00” W, 561 m, 1-I-2017, P. Sagot; ♂ ECOAB 67320, MEX. CHIS [Mexico, Chiapas], Venustiano Carranza, 16°21’0.00” N, 92°37’12.00” W, 597 m, 1-II-2017, P. Sagot (ECOAB); ♂ ECOAB 67390 MEX. CHIS [Mexico, Chiapas], Solcotenango, 16°9’0.00” N, 92°22’48.00” W, 723 m, 14-I-2017, P. Sagot. (ECOAB); ♂ ECOAB 125155, MEX. CHIS [Mexico, Chiapas], Nicolás Ruiz, 16°28’12.00” N, 92°36’0.00” W, 890 m, 8-III-2020, P. Sagot (ECOAB); ♀ ECO-TAE57096, MEX. CHIS [Mexico, Chiapas], Arriaga, ej. López Mateos, 16°20’46.60” N, 93°58’26.80” W, 255 m, 10-III-2009, C. Balboa, M. Guzmán & M. Cigarroa (EBCH). Same data as the previous one, 2♀ ECO-TAE57326, ECO-TAE57327, 3♂ ECO-TAE67405, ECO-TAE57406, ECO-TAE57407. ♀ HFL JZ5, MEX. JAL [Mexico, Jalisco], La Huerta, Chamela, Estación biológica, vereda Chachalaca, 19°29’37.20” N, 105°2’44.94” W, 300 m, 9-IV-2014/11-IV-2014, E. Ramírez & A. Estrada (CZUG); ♀ CNIN-AP 009852, MEX, OAX [Mexico, Oaxaca], Santa Maria Huatulco, Parque Nacional Huatulco, 15°45’59.40” N, 96°12’11.00” W, 14-IV-2022, A. Saldivar & P. Benítez (CNIN).
Other material examined. ♂ MZUCR HY5700, no collection data (MZUCR).
Etymology. This species is dedicated to Luz del Carmen Fernández Rodríguez, mother of the first author. The noun is feminine and declined in the genitive case.
Distribution. It is only known from Jalisco, Oaxaca and Chiapas in Mexico, Huehuetenango in Guatemala and Guanacaste in Costa Rica, it is highly probable that it is also found in Belize, El Salvador, Honduras and Nicaragua (Fig. 10).
Taxonomic comments. This species is easily recognizable by the middle tibial spur with 2 short arms in respect to the rest of the spur (Figs. 29D, 33D) and with spots of whitish setae on the sublateral area of T2 to T6 in males (Fig. 29D) and T2 to T4 in females (Fig. 31A).
Table 2
Known host associations for species of Mesoplia Lepeletier, 1841.
Cleptoparasite
Hosts
Relationship record
Type of evidence
Reference
Mesoplia (M.) bifrons (Fabricius, 1804)
Centris (Centris) caxiensis Ducke, 1907
Brazil
Confirmed
Rocha-Filho et al. (2009)
Mesoplia (M.) cubensis Genaro & Breto, 2022
Centris (Centris) poecila Lepeletier, 1841
Cuba
Probable
Genaro and Breto (2022)
Mesoplia (E.) decorata (Smith, 1854)
C. (Centris) flavofasciata Friese, 1900
Costa Rica, Guanacaste
Presumed
Vinson et al. (1987)
Mesoplia (M.) dugesi (Cockerell, 1917)
C. (Exallocentris) aterrima Smith, 1854
Mexico
Suspected
Snelling (1984)
Mesoplia (M.) insignis (Smith, 1879)
C. (Acritocentris) agilis Smith, 1874
Mexico, Jalisco
Probable
This work
Mesoplia (M.) regalis (Smith, 1854)
C. (Centris) flavofasciata Friese, 1900
Costa Rica, Guanacaste
Confirmed
Vinson et al. (1987)
C. (Centris) flavifrons (Fabricius 1775)
Brazil, Paraiba
Confirmed
Martins et al.(2014)
Mesoplia (M.) rufipes (Perty, 1833)
C. (Centris) aenea Lepeletier, 1841
Brazil, Cerrado
Probable
Aguiar and Gaglianone (2003); Rocha-Filho et al. (2009)
Mesocheira ornata Spinola, 1841: 145. Holotype: Museo Regionale di Scienze Naturale, Torino, Italy (♀). Type locality: French Guiana, Cayenne.
Diagnosis. Male: body metallic green, bright yellow and black pubescence (Fig. 32); integument of the antenna, pronotal lobe, tegula and legs black, except for some internal regions of the hind leg which are dark reddish; pubescence of the head and mesosoma mostly bright yellow; conical scutellar tubercles, with a noticeable cleft between them; short and wide forked medial tibial spur, with dark reddish integument that becomes blackish at the apex, wide anterior projection, with 2 to 3 internal teeth (Fig. 3G); pubescence of the terga mainly whitish with a bright yellow maculation on both lateral margins of T1 and T2 (Fig. 32A, B); apex of T7 forked, both pointed and bare at tip, median cleft shallow but fairly wide, separating apices considerably (Fig. 7G); S4 bilobed, with several rows of compound pubescence on the apical margin, dark in color in the center and becomes light in the middle of the apices and continues that color towards the lateral edges of the lobes; digitiform and dorsal lobe of the gonostylus of similar size (Fig. 33C, D).
Female: body metallic blue, black and pale yellow pubescence (Fig. 34); integument of antenna, pronotal lobe, tegula and legs black; F1 a little longer than F2; yellowish pubescence in preoccipital region, gena, collar of the pronotum, a broad band in the center of the mesepisternum, the posterior region of the scutellum and metascutum; conical scutellar tubercles, projecting upwards with an evident cleft between them; robust medial tibial spur, bifurcated practically in half, internal tip curved posteriorly, has an internal tooth that in some cases is located on the bifurcation, external apex wider, has 2 internal teeth; pygidial plate, narrow at the base, rounded apex, forming an angle of approximately 35°, glabrous, without scales or hairs (Fig. 34D).
Figure 29. Mesoplia (E.) carmelitae sp. nov. Holotype, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, detail of the metasoma.
Taxonomic summary
Material examined. Costa Rica: Guanacaste: Estación Biológica Maritza, 10°57’25.06” N, 85°29’42.01” W, 573 m, 1988/1999, 1♀ (MZUCR HY6820), 1990, 1♀ (MZUCR HY6822). Mexico: Chiapas: La Concordia, 15°58’48.00” N, 92°48’36.00” W, 581 m, 2-III-2016, P. Sagot, 1♂ (ECOAB 76697).
Distribution. Previously it had only been recorded in South America. If the identity of this species is confirmed, its distribution would expand to southern Mexico in Chiapas (Fig. 10).
Taxonomic comments. The male is similar to M. insignis due to the yellow coloration of the mesosoma. It is differentiated among other characters by the lack of pubescence brush on the inner margin of the posterior tibia (Fig. 3G) and by the shape of the T7 whose internal notch resembles a semicircle (Fig. 7G).
Key for species of Mesoplia Lepeletier, 1841 of Mexico and Central America.
Figure 30. Mesoplia (E.) carmelitae sp. nov. Holotype, male: A, S7. B, S8.C and D, genital capsule, dorsal and lateral view, respectively.
Males
1. Distal inner surface of the posterior tibia, with a setal brush (Fig. 3A-E); hind tibial spurs not reaching beyond basal third of basitarsus, some species with only 1 spur present [M. (Mesoplia)] ………………………………………………………………… 2
– Distal inner surface of the posterior tibia, without a setal brush (Fig. 3F, G); hind tibial spurs reaching to or beyond middle of basitarsus (Fig. 3F), both present M. (Eumelissa) …………………………………………………………………7
2(1). Inconspicuous hind tibia setal brush (Fig. 3D, E); 1 spur on the posterior tibia; ventral basal surface of posterior femur with a robust posterior projection; inner surface of posterior basitarsus with a longitudinal carina (Group azurea) ………………………………………………………………… 3
– Conspicuous hind tibia setal brush (Fig. 3 A-C); 2 spurs on the posterior tibia; ventral basal surface of posterior femur without a robust posterior projection; inner surface of posterior basitarsus without a longitudinal carina (Group bifrons) ………………………………………………………………… 4
3(2). Setae of most of the body mainly dark; metallic scales on the tegula present (Fig. 21B); integument of legs blackish (Mexico) Mesoplia dugesi (Cockerell, 1917)
– Setae of most of the body mainly whitish; metallic scales on the tegula ausent (Fig. 24B); integument of legs redish brown (Mexico to South America) Mesoplia rufipes (Perty, 1833)
– Pubescence of the mesosoma mainly blackish and whitish, if it presents yellowish setae they are restricted to the posterior region of the scutellum ………………………………………………………………… 6
Figure 31. Mesoplia (E.) carmelitae sp. nov. Allotype, female: A, lateral habitus; B, frontal habitus; C, dorsal habitus; D, posterior view of the scutellar tubercles.
5(4). Pubescence of the mesosoma almost exclusively bright yellow; central disc of scutum and dorsal surface of scutellar tubercles with yellow setae; metasoma with a spot of bright yellow pubescence on lateral margins of T1 and T2 (repeated in some specimens until T4) (Fig. 6A); T7 concavity shallow; T7 apices broadly rounded (Fig. 7A) (México) ………………………………………………………………… Mesoplia insignis (Smith, 1879)
– Pubescence of the mesosoma bright yellow interrupted by a broad transverse band in the anterior region; central disc of scutum and dorsal surface of scutellar tubercles with scattered black setae; metasoma with a bright yellow pubescent spot on both lateral margins of T1 (some specimens also with a very small spot on T2) (Fig. 11A); T7 concavity deep; T7 apices narrowly rounded (Fig. 7B) (Costa Rica) ………………………………………………………………… Mesoplia tica sp. nov.
6(4). T7 with apices rounded and setae on ventral side come out a little in dorsal view (Fig. 7C) (Mexico and Central America) ………………………………………………………………… Mesoplia sapphirina Melo & Rocha-Filho, 2011
– T7 with apices flat and with ventral setae below conspicuous in dorsal view (Panama and South America) ………………………………………………………………… Mesoplia regalis (Smith, 1854)
7(1). Pubescence of the head and mesosoma mainly bright yellow; metasoma with spots of pubescence bright yellowon lateral margins of T1 and T2 (Fig. 32A, B); without spots of short white pubescence on the marginal areas of T3 to T6; middle tibial spur with long branches (Fig. 16E) (South of Mexico, Chiapas, to South America) ………………………………………………………………… Mesopliaornata (Spinola, 1841)
Pubescence of the head and mesosoma mainly whitish; without yellow pubescence on the metasoma; with 2 spots of short white pubescence in the marginal zone of T2 to T6 (Fig. 29D); branches of the middle tibial spur short (Fig. 16D) (Mexico, Guatemala and Costa Rica) ………………………………………………………………… Mesoplia carmelitae sp. nov.
Figure 32. Mesoplia (E.) ornata, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, scutellar tubercles posterior view (photos A, B by Jorge Mérida).
Females
1. Pygidial plate broad, occupying the exposed dorsal surface of T6 and covered with scales or setae (Fig. 4A-G); dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces [M. (Mesoplia)] ………………………………………………………………… 2
– Pygidial plate narrow, does not completely occupy the exposed dorsal surface of T6 and without scales or setae (Figs. 4H, 23D); dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces [M. (Eumelissa)] …………………………………………………………………9
2(1). Pronotum with a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5B) (Group azurea) ………………………………………………………………… 3
– Pronotum without a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5A) ………………………………………………………………… 5 Group bifrons (sensu Moure, 1960a, b)
3(2). With metallic scales evident on head and tegula (Mexico) (Fig. 23B) ………………………………………………………………… Mesoplia dugesi (Cockerell, 1917)
– Without metallic scales or inconspicuous on head and tegula ………………………………………………………………… 4
4(3). Setae on most of the body mainly dark; pygidial plate with broad apex (Fig. 4G) (Mexico, Oaxaca) ………………………………………………………………… Mesopliaoaxacana sp. nov.
– Setae of most of the body mainly whitish; pygidial plate with narrow apex (Fig. 4F) (Mexico to South America) ………………………………………………………………… Mesoplia rufipes (Perty, 1833)
5(2). With bright yellowpubescence on head, mesosoma and metasoma (Fig. 20) (Mexico, Veracruz) ………………………………………………………………… Mesoplia veracruzana sp. nov.
– Without bright yellow pubescence on head, mesosoma and metasoma ………………………………………………………………… 6
6(5). With metallic scales very evident on the scape and tegula; forewings evenly smoked ………………………………………………………………… 7
– Without metallic scales on the scape and tegula or not very evident; forewings with a darker spot at the apex ………………………………………………………………… 8
7(6). Head pubescence mainly black (Fig. 9C); scutellar tubercles moderately projecting upwards (Fig. 9D); truncated pygidial plate with dark setae on the surface (Fig. 4A) (Mexico) ………………………………………………………………… Mesoplia insignis (Smith, 1879)
– Head pubescence mainly whitish (Fig. 13C); scutellar tubercles strongly projecting upwards (Fig. 13A); rounded pygidial plate with scales on the surface, in some specimens they do not reach the apex due to the abrasion of these (Fig. 4B) (Costa Rica)………………………………………………………………… Mesoplia tica sp. nov.
Figure 33. Mesoplia (E.) ornata, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
8(6). Poorly developed scutellum protuberances with a shallow depression between them and low tubercles (Fig. 16A, D); the carina of the scutellum runs continuously from one side to the other (Mexico and Central America) ………………………………………………………………… Mesoplia sapphirina Melo & Rocha-Filho, 2011
– Scutellum protuberances developed with a deep depression between them and conspicuously pointed tubercles; the carina of scutellum most developed along tubercles, gradually fading to sides (Panamá and South America) ………………………………………………………………… Mesoplia regalis (Smith, 1854)
9(1). With pale yellow pubescence on the mesosoma (Fig. 34A-C); T2 to T5 without white pubescent spots; long middle tibial spur branches (South of Mexico, Chiapas, to South America) ………………………………………………………………… Mesoplia ornata (Spinola, 1841)
– Without pale yellow pubescence on the mesosoma; T2 to T5 with a couple of spots of appressed white pubescence in the marginal zone (Fig. 31A); short middle tibial spur branches (Fig. 18D) (Mexico, Guatemala and Costa Rica) ………………………………………………………………… Mesoplia carmelitae sp. nov.
Hosts. Like the rest of the members of the Ericrocidini tribe, species of the genus Mesoplia are cleptoparasites of several species of the Centridini tribe (Table 2). It should be noted that M. rufipes is the only species of the tribe that has been recorded cleptoparasitizing species of Epicharis. The cleptoparasitism relationship between M. insignis and Centris agilis was recorded for the first time in Jalisco, Mexico. The first author of this work had the opportunity to collect some specimens of this species in an aggregation of C. agilis nests in a suburban site.
Floral asossitations. The floral records reported in this section are based on information extracted directly from the collection tags of the specimens reviewed in this study (Table 3).
Figure 35. Species of Mesoplia in nature. A-B, Mesoplia (M.) insignis on Ramirezella sp. in Jalisco, México. A, male; B, female (photos by the first author). C, Mesoplia (M.) tica sp. nov. female on Stachytarpheta sp. in Alajuela, Costa Rica (photo by Daniel Garrigues). D, Mesoplia (M.) veracruzana sp. nov. female on Ipomoea sp. in Veracruz, Mexico (photo by Gerardo Quintos-Andrade). E, Mesoplia (M.) dugesi male on Tecoma stans in Querétaro, Mexico (photo by Belem Hernández). F, Mesoplia (M.) rufipes male on Duranta sp. in San José, Costa Rica (photo by “Homúnculo Daimon”).
Discussion
The complexity of the genus Mesoplia has previously been recognized for regions like the Caribbean. There is inaccurate information on its distribution due to taxonomic problems, which has caused erroneous identifications (Genaro & Breto, 2022; Genaro & Franz, 2008). For Mexico, difficulty in accessing the primary types deposited in institutions outside the country complicates the taxonomic understanding of the group. However, there are quality images available that have allowed this review, obtaining a reliable list and synonymizing the names that were required v. gr. M. insignis (Smith, 1879) and M. imperatrix (Friese, 1912), a species easily recognizable in the end.
Most of the of the species in this study present morphological characters that make them easily differentiated. However, 2 pairs of species, M. regalis and M. sapphirina; as well as M. dugesi and M. oaxacana, are quite similar to each other, particularly the females, which complicates their differentiation. Here we provide characters that allow the unequivocal identification of the mentioned species, such as scutellar tubercles and pygidial plate. Additionally, variation in characters of the male of M. rufipes, including shape of S8 and shape of the basal projection of the posterior femur, hint to the possibility that M. rufipes is a species complex, but more evidence is needed including molecular studies.
Table 3
Floral records of the species of Mesoplia Lepeletier, 1841.
Species
Plant
Family
Mesoplia (M.) dugesi (Cockerell, 1917)
Tecoma stans
Bignoniaceae
Canavalia villosa
Fabaceae
Crotalaria sp.
Libidibia sclerocarpa
Macroptilium atropurpureum
Mesoplia (M.) insignis (Smith, 1879)
Eryngium sp.
Apiaceae
Macroptilium atropurpureum
Fabaceae
Ramirezella sp.
Vitex pyramidata
Lamiaceae
Cuphea lanceolata
Lythraceae
Hibiscus tiliaceus
Malvaceae
Mesoplia (M.) regalis (Smith, 1854)
Antigonon leptopus
Polygonaceae
Mesoplia (M.) rufipes (Perty, 1833)
Ipomoea violacea
Convolvulaceae
Tabebuia rosea
Bignoniaceae
Tecoma stans
Caesalpinia sp.
Fabaceae
Haematoxylon brasiletto
Hyptis albida
Lamiaceae
Cuphea sp.
Lythraceae
Genipa americana
Rubiaceae
The association of cleptoparasitism between M. insignis and Centris agilis supports the observations of Lim et al. (2022), they observed a considerable increase in the body size of Ericocidini that could be influenced by a change of host; and supports Harrison’s rule, since both species are of similar sizes (Harrison, 1915). Therefore, in some cases where the host is unknown, it could at least be inferred. For example, M. carmelitae sp. nov. is a relatively small bee within the genus, so it is expected that its host will be a bee from the Centridini tribe of similar size (approx. 11.395 mm) and distributed in the same geographical area. There is an opportunity to study cleptoparasite-host relationships between bees of the tribe Centridini and Ericrocidini, in sites where it is known that nest aggregations periodically occur, in addition to increasing information about the immature stages of both groups.
With this research, the region studied reaches South America with 10 species of Mesoplia, despite this it is expected that there are still several species to be described for that region since Ericrocidini is richer in that region (Snelling & Brooks, 1985). Furthermore, the distribution of some of the species recognized in this work could be broader, since there is a significant information gap across some Central American countries. The area included in this study has little diversity of species of the subgenus Eumelissa, with only 2 of the 10 known species. Furthermore, the data obtained extends the distribution of Eumelissa at least to Central Mexico on the Pacific Slope (particularly in Jalisco), far north of the previous northern limit in Costa Rica (Snelling & Brooks, 1985).
Acknowledgements
This article serves as fulfillment of the first author for obtaining the degree of M.Sc. degree with emphasis on Systematics of the Posgrado en Ciencias Biológicas at the UNAM. This study was financially supported by the student grants: Beca Nacional para Estudios de Posgrado 2022 – 1 Conahcyt No. 814906 and Programa de Apoyo a los Estudios de Posgrado (PAEP) 2023. We want to thank José Luis Navarrete(CZUG), Hugo Fierros (CZUG and HFL), Jorge E. Valenzuela, Viridiana Vega, Gerardo Quintos-Andrade (IEXA), Rémy Vandame, Jorge A. Mérida Rivas, Philippe Sagot (ECOAB), Virginia Meléndez Ramírez, María José López Gómez (CER-UADY), Zachary Falin, Michael Engel, Rachel Osborn (SEMC), Maricelle Méndez, Marcela Sánchez (MNCR), Mauricio Fernández, Paul Hanson (MZUCR), Mario Gallardo, Eduardo Herrera (CINAT) for the loan of specimens and/or for the information provided about the specimens of Mesoplia deposited in their collections. We thank Stefanie Krause (ZMB) and Corey Smith (AMNH), who sent us photographs of some type specimens. We thank again Jorge Mérida (ECOAB), Marcela Sánchez (MNCR), José Luis Navarrete (CZUG), Adam Karremans (Director of Jardín Botánico Lankaster, UCR) and Nicolás Rengifo Alfonso (MZUCR) for letting and helping us to use the imaging system to take the photographs. We want to thank Daniel Garrigues, Gerardo Quintos-Andrade, Belem Hernández and “Homúnculo Daimon” for the photographs of Mesoplia in nature. Finally, we thank the reviewers that helped to improve this article.
References
Aguiar, A. M. L., & Ganglianone, M. C. (2003). Nesting biology of Centris (Centris) aenea Lepeletier (Hymenoptera, Apidae, Centridini. Revista Brasileira de Zoologia, 20, 601–606. https://doi.org/10.1590/S0101-81752003000400006
Coville, R. E., Frenkie, G. W., & Vinson, S. B. (1983). Nests of Centris segregata (Hymenoptera: Anthophoridae) with a Review of the Nesting Habits of the Genus. Journal of the Kansas Entomological Society, 56, 109–122.
Gaglianone, M. C. (2005). Nesting biology, seasonality, and flower hosts of Epicharis nigrita (Friese, 1900) (Hymenoptera: Apidae: Centridini), with a comparative analysis for the genus. Studies on Neotropical Fauna and Environment, 40, 191–200. https://doi.org/10.1080/01650520500250145
Genaro, J. A., & Franz, N. M. (2008). The bees of Greater Puerto Rico (Hymenoptera: Apidae: Anthophila). Insecta Mundi, 40, 1–24.
Genaro, J. A., & Breto, D. (2022). Descripción de dos especies nuevas de Mesoplia Lepeletier (Hymenoptera: Apoidea: Apidae) para Cuba y Las Bahamas, con notas sobre taxonomía y distribución del género en las Antillas. Insecta Mundi, 929, 1–17.
Harris, R. A. (1979). A glossary of surface sculpturing. Occasional Papers in Entomology, 28, 1–32.
Harrison, L. (1915). Mallophaga from Apteryx, and their signifcance; with a note on the genus Rallicola. Parasitology, 8, 88–100.
Hiller, B., & Wittmann, D. (1994). Seasonality, nesting biology and mating behavior of the oil-collecting bee Epicharis dejeanii (Anthophoridae, Centridini). Biociências, 2, 107–124.
Lim, K., Lee, S., Orr, M., & Lee, S. (2022). Harrison´s rule corroborated for the body size of cleptoparasitic cuckoo bees (Hymenoptera: Apidae: Nomadinae) and their hosts. Scientific Reports, 12, 10984. https://doi.org/10.1038/s41598-022-14938-9
Lobo, J. A., Fernández-Otárola, M., Chavarría, M. M., & Agraz-Hernández, C. M. (2023). Nesting biology of Centris aethyctera (Centridini, Apidae) in an estuarine environment. Apidologie, 54, 58. https://doi.org/10.1007/s13592-023-01044-6
Martins, A. C., Luz, D. R., & Melo, G. A. R. (2018). Palaeocene origin of the Neotropical lineage of cleptoparasitic bees Ericrocidini-Rhathymini (Hymenoptera, Apidae). Systematic Entomology, 43, 510–521. https://doi.org/10.1111/syen.12286
Martins, C. F., Neto V. I. S., & Cruz, R. D. M. (2019). Nesting biology and mating behavior of the solitary bee Epicharis nigrita (Apoidea: Centridini). Journal of Apicultural Research, 58, 1–11. https://doi.org/10.1080/00218839.2019.1584963
Martins, C. F., Peixoto, M. P., & Aguiar, C. M. L. (2014). Plastic nesting behavior of Centris (Centris) flavifrons (Hymenoptera: Apidae: Centridini) in an urban area. Apidologie, 45, 156–171. https://doi.org/10.1007/s13592-013-0235-4
Michener, C. D. (2007). The bees of the World(2nd Edition). Baltimore, Maryland: The Johns Hopkins University Press.
Moure, J. S. (1960a). Notas sobre os tipos de abelhas do Brasil descritas por Perty em 1833 (Hymenoptera – Apoidea). Boletim da Universidade do Paraná, Zoologia, 1, 1–23.
Moure, J. S. (1960b). Notes on the types of the Neotropical bees described by Fabricius (Hymenoptera: Apoidea). Studia Entomologica, 3, 97–160.
Moure, J. S., & Melo, G. A. R. (2023). Mesoplia Lepeletier, 1841. In J. S. Moure, D. Urban, & G. A. R. Melo (Orgs.), Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical Region. Retrieved on 30 May 2024 from: http://moure.cria.org.br/catalogue?id=113174
QGIS.org (2022). QGIS Geographic Information System. QGIS Association. http://www.qgis.org
Rocha-Filho, L. C., Morato, É. F., & Melo, G. A. R. (2009). New host records of Aglaomelissa duckei and a compilation of host associations of ericrocidini bees (hymenoptera: Apidae). Zoologia, 26, 299–304. http://dx.doi.org/10.1590/S198446702009000200012
Rocha-Filho, L. C., Silva, C. I., Gaglianone, M. C., & Augusto, S. C. (2008). Nesting behavior and natural enemies of Epicharis (Epicharis) bicolor Smith (Hymenoptera: Apidae). Tropical Zoology, 21, 227–242.
Rozen, J. G. (1969). The larvae of the Anthophoridae (Hymenoptera, Apoidea). Part 3. The Melectini, Ericrocidini, and Rhathymini. American Museum Novitates, 2382, 1–24.
Rozen, J. G. (2016). Nesting biology of the solitary bee Epicharis albofasciata (Apoidea: Apidae: Centridini). American Museum Novitates, 3869, 1–8. https://doi.org/10.1206/3869.1
Rozen, J. G., Vinson, S. B., Coville, R. E., & Frankie, G. W. (2011). Biology of the cleptoparasitic bee Mesoplia sapphirina (Ericrocidini) and its host Centris flavofasciata (Centridini) (Apidae: Apinae). American Museum Novitates, 3723, 1–36. https://doi.org/10.1206/3723.2
Silveira, F. A., Melo, G. A. R., & Almeida, E. A. B. (2002). Abelhas brasileiras: sistemática e identificação. Belo Horizonte, Minas Gerais: Ministério do Meio Ambiente & Fundação Araucária.
Snelling, R. R. (1984). Studies on the taxonomy and distribution of American Centridine bees (Hymenoptera: Anthophoridae). Contributions in Science, Natural History Museum of Los Angeles County, 347, 1–69.
Snelling, R. R., & Brooks, R. W. (1985). A review of the genera of cleptoparasitic bees of the Tribe Ericrocidini (Hymenoptera: Anthophoridae). Contributions in Science, Natural, History Museum of Los Angeles County, 369, 1–34.
Vinson, S. B., Frankie, G. W., & Coville, R. E. (1987). Nesting habits of Centris flavofasciata Friese (Hymenoptera: Apoidea: Anthophoridae) in Costa Rica. Journal of the Kansas Entomological Society, 60, 249–263.
Vivallo, F. (2010). Sistemática e filogenia da tribo de abelhas Centridini e suas relações filogenéticas com as tribos cleptoparasitas Ericrocidini e Rhathymini (Hymenoptera: Apidae)(Ph.D. Thesis). Universidade Federal do Paraná. Curitiba, Brazil.
Vivallo, F. (2014). Taxonomic revision of the cleptoparasitic bee genus Epiclopus Spinola, 1851 (Hymenoptera: Apidae: Ericrocidini). Zootaxa, 3857, 41–70. https://doi.org/10.11646/zootaxa.3857.1.2
Vivallo, F. (2021). Nesting behavior of the oil-collecting bee Epicharis (Triepicharis) analis Lepeletier (Hymenoptera: Apidae) in an urban area of Rio de Janeiro, RJ, Brazil. Journal of Apicultural Research, 60, 135–142. https://doi.org/10.1080/00218839.2020.1820150
Werneck, H. A., & Campos, L. A. O. (2020). A study of the biology of Epicharis (Epicharoides) picta using emergence-traps. Journal of Hymenoptera Research, 80, 147–167. https://doi.org/10.3897/jhr.80.56898
Phylogeny of the weevil genus Megalostylus (Coleoptera: Curculionidae: Entiminae), endemic to Mexico
Alejandro Lizama-Hernández a, Ma Ventura Rosas-Echeverría a, *, M. Guadalupe del Rio b
The first phylogenetic analysis of the weevil genus Megalostylus endemic to Mexico(Entiminae, Naupactini) is presented, based on a data matrix of 37 morphological characters of adults and 21 terminal taxa. The ingroup comprises 9 species, 4 varieties, and 5 specimens of Megalostylus whose identification at the species level is doubtful. The outgroup includes species representing closely related genera: Pantomorus albosignatus, Naupactus cervinus, and Megalostylodes hirsutus. The objectives were to test the monophyly of Megalostylus, to explore its species relationships, and to determine which synapomorphies allow its identification and differentiation from other genera. The analysis yielded a single cladogram of 61 steps, showing the following phylogenetic sequence:
Victor Cuesta-Porta a, *, George Melika b, Juli Pujade-Villar a
a Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals, Avda. Diagonal 645, 08028-Barcelona, Spain
b National Food Chain Safety Office, Plant Health Diagnostic National Reference Laboratory, Budaörsi Str. 141-145, Budapest 1118, Hungary
The North American oak gall wasp genus Femuros Kinsey is revised after its re-establishment in 2015. Morphological evidence suggests that only 3 species described by Kinsey belong to this genus: F. repandae Kinsey, F. ocri Kinsey, and F. ruidum Kinsey. The rest of the species are transferred to the genus Andricus Hartig: A. geniale (Kinsey) comb. rev., A. integrum (Kinsey) comb. rev., A. lusum (Kinsey) comb. rev., and A. perfectum (Kinsey) comb. rev. A new species with slightly swollen femora but without an apical lobe and without internal tibial carina is described: A.calvoi sp. nov. We provide descriptions, redescriptions, diagnoses, keys to species, and information on biology, phenology, and distribution.
Keywords: Hymenoptera; Gall wasp; Nearctic; Taxonomy
Se revisa el género norteamericano Femuros Kinsey que fue restituido en 2015. Solo 3 especies descritas por Kinsey pertenecen a este género: F. repandae Kinsey, F. ocri Kinsey y F. ruidum Kinsey. El resto de las especies están incluidas en el género Andricus Hartig: A. geniale (Kinsey) comb. rev., A. integrum (Kinsey) comb. rev., A. lusum (Kinsey) comb. rev. y A. perfectum (Kinsey) comb. rev. Se describe una especie nueva con fémures ligeramente hinchados, pero sin lóbulo apical y sin carina tibial interna: A. calvoi sp. nov. Se proporcionan descripciones, redescripciones, diagnosis, una clave para las especies, información sobre biología, fenología y distribución.
Kinsey (1937a, b) described several oak gall wasp genera from the Nearctic and Neotropics related to Andricus, some of which were synonymized under Andricus by Weld (1952): Femuros Kinsey, 1937, Feron Kinsey, 1937, Druon Kinsey, 1937, and Conobius Kinsey, 1938. Later, Melika and Abrahamson (2002) increased the number of synonymies: Dros Kinsey, 1937 and Erythres Kinsey, 1937. Recently morphological and phylogenetic studies have revalidated some of these genera: Erythres Kinsey, 1937 (Pujade-Villar & Melika, 2014), Femuros Kinsey, 1937(Pujade-Villar & Ferrer-Suay, 2015), Antron Kinsey, 1930 (Melika et al.,2021), Dros Kinsey, 1937 (Pujade-Villar et al., 2017), Druon Kinsey, 1937 (Cuesta-Porta et al., 2022), and Feron (Cuesta-Porta et al., 2023).
Femuros was described by Kinsey (1937a) to include 2 species (F. repandae and F. ruidum) characterized as having …“the hind femur peculiarly broadened on ventral margin near distal end, […], although it is suddenly constricted where it joins the tibia”. Later, Kinsey (1937b) described the “geniale complex” to include several species with different gall morphologies and punctured metasoma (F. lusum, F. perfectum, F. geniale, and F.integrum), and a new species belonging to “repandae complex” (F. ocri). Recently, Pujade-Villar and Ferrer-Suay (2015) mentioned another character present in the genus Femuros, an internal carina on the hind tibia, thus re-establishing Femuros as valid.
In Kinsey’s descriptions (both generic and specific), many diagnostic characters are not mentioned. The study of the type material and of the samples collected in Mexico show 2 distinct morphological models and some errors in the description of the species. The aim of this study is to redescribe the genus Femuros to adjust the diagnostic characters to the new proposed generic limits of Femuros and redescribe the species that Kinsey (1937a, b) included. We transfer and redescribe the species belonging to the “genialecomplex” to the genus Andricus, and we describe a new species of Andricus with swollen femurs and internal tibial carina: A. calvoi sp. nov. Also, the species Femuros bracteatus comb. nov. is transferred from Andricus.
Materials and methods
Fresh material was obtained from galls collected in Mexico from multiple collecting events during 2009-2019. Site locations and other details are provided in the “Material examined” section for each species. Galls were reared at room temperature in plastic containers with mesh lids to ensure ventilation. Emerging adult wasps were preserved in 70% ethanol. Type materials of previously described species was also examined, both to confirm the identity of our recently collected specimens and to help define species limits.
Type specimens are deposited at the American Museum of Natural History (AMNH) in New York City, Hungarian Natural History Museum (HNHM) in Budapest, and JP-V collection of Barcelona University (UB).
Morphological descriptions
The terminology used to describe gall wasp morphology follows other recent cynipid studies (Liljeblad et al., 2008; Melika, 2006; Melika et al., 2010). Abbreviations for forewing venation follow Ronquist and Nordlander (1989), and cuticular surface terminology follows Harris (1979). Measurements and abbreviations used here include: F1-F12 for the 1st and subsequent flagellomeres; POL (post-ocellar distance) for the distance between the inner margins of the lateral ocelli; OOL (ocellar-ocular distance) for the distance from the outer edge of a lateral ocellus to the inner margin of the compound eye; and LOL (lateral-frontal ocelli distance) for the distance between lateral and frontal ocelli. The width of the forewing radial cell is measured from the margin of the wing to the Rs vein.
The information contained in the material examined sections is a verbatim transcription of the pinned labels attached to the adult wasps including symbols and abbreviations. Alfred Kinsey included the symbol “ꚛ” to indicate the specimens naturally emerged from galls. Also, Kinsey designated most holotypes using a “Paratype” red label and crossing out the “Para-” to handwrite “Holo-” on top. We use the term “HoloParatype” to better reflect the verbatim transcription of Kinsey’s holotype labels, and to follow the terms used in previous studies (Cuesta-Porta et al., 2022, 2023, 2024).
Bright-field images of adults were produced with a digital Leica DC500 camera attached to a Leica DM2700M compound microscope using the LAS Store&Recall software, followed by processing in Adobe Photoshop 6.0. SEM images of the new species were taken with the Stereoscan Leica-360 at low voltage (700V) and without coating, in order to preserve the specimens. For all species examined in this paper, descriptions are provided according to current morphological requirements as the original descriptions are often superficial.
Nomenclature of host plants
Denk et al. (2017) reorganized the classification of Quercus, dividing the genus into the subgenera Cerris and Quercus. Subgenus Cerris includes 3 sections: Cyclobalanopsis (previously considered a subgenus), and the sections Ilex and Cerris from Menitsky (1984). Subgenus Quercus includes 5 sections: Lobatae and Protobalanus as in Nixon (1993), with the old section Quercus divided into 3 new sections: Quercus ‘sensu stricto’, Virentes, and Ponticae. The gall wasp species listed herein are associated with section Quercus subsection Leucomexicanae: Q. arizonica Sarg. (= Q. sacame Trel.), Q. chihuahuensis Trel., Q. deserticola Trel. (= Q.texcocana Trell.), Q. glaucoides Mart. & Gal., Q. grisea Liebm., Q. laeta Liebm., Q. magnoliifolia Née (= Q. haemathophebia Trel., = Q. macrophylla Née), Q.obtusata Humb. & Bonpl., Q. potosina Trel., Q. repanda Bonpl., Q. rugosa Née (= conglomerata Trel., = reticulata Bonpl., = rhodophlebia Trel.), Q. repanda Bonpl., and Q. undata Trel. There is only one record of an association to an oak in section Lobatae, Q. crassifolia Bonpl.
Results
Below we present the formal redescriptions of 4 valid species of Femuros (Cynipini), all from the Nearctic or Neotropics: F. bracteatus (Weld) comb. nov., F. ocri Kinsey, F. repandae Kinsey, and F. ruidum Kinsey. Also, 4 species currently in Femuros are placed in the genus Andricus Hartig 1840: A. geniale (Kinsey) comb. rev., A. integrum (Kinsey) comb. rev., A. lusum (Kinsey) comb. rev., and A. perfectum (Kinsey) comb. rev., along with a new Andricus species with swollen hind femurs and internal tibial carina described here: A.calvoi sp. nov.
The following identification key includes 8 Cynipini species characterized by a conspicuous transscutal articulation, toothed tarsal claws, fully-developed wings, without malar sulcus, antenna with 11-12 flagellomeres, the hind femur with a distinct lobe in posterior margin, hind tibia with a strong longitudinal carina, the second metasomal tergite with sparse setae on anterolateral area, the third and subsequent metasomal tergites micropunctured, and the ventral spine of hypopygium needle-like with its sides gradually converging into a tip.
Key to asexual Andricus and Femuros females with broadened femur and tibial carina
1. Body chestnut to black (Fig. 24a-d); lower head in frontal view sculptured, with radiating striae, carina reaching margin of the eye (Fig. 1a); occipital carina absent; all metasomal segments micropunctured posteriorly (Figs. 2c, 9b, 11b, 14b) …………………………………………………………………………………… Andricus (2)
– Body red rufous (Fig. 25a-d); lower head in frontal view almost smooth, without striae radiating from clypeus or with very inconspicuous carinae only next to clypeus (Figs. 15a, 16a, 18a, 21a); occipital carina dorsally present (Figs. 18b, 21c); second metasomal tergum smooth, without micropunctures, subsequent terga micropunctured (Figs. 15d, 16e, 20a, 23b) …………………………………………………………………………………… Femuros (6)
2. LOL subequal or shorter than diameter of lateral ocellus (Figs. 1b, 10b, 12c); hind coxae and femora swollen, sometimes hind femora with strong distal lobe (Figs. 2e-f, 11b, 14b); propodeal carinae bent outwards or converging posteriorly (Figs. 1e, 10f, 14c) …………………………………………………………………………………… 3
– LOL more than 1.2× diameter of lateral ocelli (Figs. 7c, 12c); hind coxae and femora not swollen (Fig. 9a); propodeal carinae subparallel (Figs. 6b, 8d) …………………………………………………………………………………… 5
3. Lower face rugose-reticulate, dull, with vertical carinae radiating from clypeus towards toruli (Fig. 12a, c); pronotum completely striate, with parallel curved striae (Fig. 13c); notaulus weakly impressed, incomplete, reaching slightly more than 1/2 of mesonotum length (Figs. 12f, 13a) …………………………………………………………………………………… A. perfectum, comb. rev.
– Lower face faintly reticulate, shining, carinae radiating from clypeus not extending towards toruli (Figs. 1a-b, 10a, 10b); pronotum punctured with striae only on lateral margin (Figs. 1c, 10c); notaulus deep, complete or not (Figs. 1d, 10c) …………………………………………………………………………………… 4
4. Notauli incomplete not converging posteriorly, space between notauli subequal to space between notauli and parapsidal line (Fig. 10d-e), propodeal carinae subparallel slightly bent outwards (Fig. 10a) …………………………………………………………………………………… A. lusum, comb. rev.
– Notauli complete strongly converging posteriorly, space between notauli subequal to width of notauli on posterior margin (Fig. 1d); propodeal carinae strongly converging towards posterior end (Fig. 1e) …………………………………………………………………………………… A. calvoi, sp. nov.
5. Lower face reticulate, sometimes with some sparse striae (Fig. 4a); mesopleuron smooth with sparse striae across mid-height of mesopleuron (Fig. 5a) …………………………………………………………………………………… A. geniale, comb. rev.
– Lower face rugose-reticulate, dull, with vertical carinae radiating from clypeus towards toruli (Fig. 7a); mesopleuron completely striated, except for smooth speculum (Fig. 8a) …………………………………………………………………………………… A. integrum, comb. rev.
6. POL around 1.5× OOL, OOL 1.5× diameter of lateral ocellus (Fig. 18c); transfacial distance 1.3× as long as height of eye, and height of eye 2.0× as long as malar space (Figs. 15a, 18a) …………………………………………………………………………………… 7
– POL 2.0× OOL, OOL subequal to diameter of lateral ocellus (Figs. 16b, 21c); transfacial distance subequal to height of eye; and height of eye more than 2.4× as long as malar space (Figs. 16a, 21a) …………………………………………………………………………………… 8
7. Mesonotum with infuscate stripes along anterior parallel lines and parapsidal lines (Fig. 19b); mesoscutellar foveae divided by a thin carina (Fig. 19d) ……………………………………………………………………………………F. repandae
– Mesoscutum uniformly colored (Fig. 15b); mesoscutellar foveae divided by a broad rugose elevated area …………………………………………………………………………………… F. bracteatus
8. Mesopleuron densely pubescent with visible punctures and deep piliferous points, speculum mostly pubescent except for a small central glabrous smooth area (Fig. 22a); propodeal carinae bent outwards (Fig. 22c) …………………………………………………………………………………… F. ruidum
– Mesopleuron sparsely pubescent, mostly smooth with scattered delicate shallow piliferous points; speculum smooth and glabrous at least on median 1/2 length of speculum (Fig. 16c); propodeal carinae parallel (Fig. 17c) …………………………………………………………………………………… F. ocri
Diagnosis. Andricus calvoi sp. nov. differs from all Andricus species, except for the species here transferred to Andricus, by the hind femur swollen and by the presence of a tibial carina. This species differs from Andricus geniale comb. rev., A. integrum comb. rev., A. perfectum comb. rev., and A. lusum, comb. rev. by the parallel internal margins of the eyes and the body without dense pubescence, the mesopleuron only basally pubescent; the lower face is reticulate; differs from A. geniale, comb. rev. by the weakly reticulate mesopleuron, except for the smooth speculum; and the propodeal carinae strongly converging posteriorly.
Description. Asexual female (Figs. 1a-e, 2a-g, 3a). Head, mesosoma and metasoma black (with some brown areas in a specimen dissected from a gall); antenna light brown, last flagellomere darker; tegula yellowish; legs light brown, coxa, trochanter, femur brown to black, last tarsus darker than preceding (Fig 3a). Wing veins light brown.
Head (Fig. 1a-c) transversally ovate, broadest part below toruli, 1.4× as broad as high and narrower than mesosoma in frontal view, 1.5× as broad as long in dorsal view, uniformly coriaceous, lower face with short dense white setae; frons glabrous, without setae, 1.3× as broad as high in anterior view. Gena coriaceous to microreticulate, slightly broadened behind eye, 0.89× as broad as cross diameter of eye, measuring along transfacial line. Malar space coriaceous to microreticulate, with very few striae radiating from clypeus, malar sulcus absent; eye 1.7× as high as length of malar space. Inner margins of eyes parallel. POL 1.7× as long as OOL, OOL 1.7× as long as diameter of lateral ocellus and slightly longer than LOL, all ocelli rounded, of same size. Transfacial distance nearly equal to height of eye; torulus located above mid height of head, frons shorter than height of lower face, diameter of antennal torulus 2.8× as long as distance between them, distance between torulus and eye as long as diameter of torulus or very slightly shorter; lower face and slightly elevated median area delicately reticulate, pubescent. Clypeus impressed, coriaceous, rounded ventrally, medially not incised, anterior tentorial pits distinct; epistomal sulcus and clypeo-pleurostomal line distinctly impressed. Frons uniformly coriaceous to microreticulate, without striae, with few setae; interocellar area coriaceous. Vertex and occiput coriaceous, glabrous. Antenna (Fig. 2a) shorter than length of head + mesosoma, with 11 flagellomeres; pedicel subglobose, slightly longer than broad; flagellomeres subsequently broadening until apex; F1 slightly broader distally, subequal to F2 and 2.3× as long as pedicel; subsequent flagellomeres progressively shortened; F3 = F4, F5 = F6, F7 to F10 equal in length, F11 longer than F10, sometimes F11 with a very inconspicuous suture and thus antenna with 12 flagellomeres. Placodeal sensilla on F5-F11.
Mesosoma (Fig. 1c-e) 1.2× as long as high, concave in lateral view, with dense white setae. Sides of pronotum alutaceous, with denser white setae and weak, parallel carinae in upper lateroposterior part, anterolateral rim of pronotum inconspicuous. Mesoscutum as broad as long in dorsal view; coriaceous, without rugae, pubescent, with piliferous points denser in the posterior half. Notaulus complete, deep, straight, converging posteriorly, in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; median mesoscutal line absent; parapsidal and anterior parallel lines differentiated by delicate sculpture. Circumscutellar carina complete. Mesoscutellum as long as broad, uniformly rugose; overhanging metanotum, with sparse long setae. Mesoscutellar foveae differentiated, triangular, deep, with shining bottom, median carina short. Mesopleuron completely sculptured, coriaceous to weak reticulate with delicate carinae and sparse setae anteriorly, speculum smooth and shining; mesopleural triangle smooth, glabrous, with dense, long white setae, hiding the surface sculpture; dorsal and lateral axillar areas with delicate parallel longitudinal carinae, glabrous; axillula alutaceous, pubescent; subaxillular bar smooth, glabrous, triangular, short, posteriorly as high as height of metanotal trough; metapleural sulcus reaching mesopleuron in upper 2/3 of its height; upper part of sulcus distinct; lower part of sulcus delimiting broad area with dense long white setae. Metascutellum subrectangular, rugose, strongly incised ventrally; metanotal trough alutaceous, with dense setae; ventral bar of metanotal trough coriaceous-rugose; central propodeal area nearly smooth and glabrous; lateral propodeal carinae strongly converging toward posterior end; lateral parts of propodeum uniformly alutaceous, densely pubescent. Nucha almost smooth and shining dorsally, with parallel sulci laterally. Legs (Fig. 2e-g) with broad coxa, femur strongly broadened, without apical lobe, with some short teeth on the internal margin produced by marginal sculpture; hind tibia with ventral carina reaching almost to the base of tibia; base of tarsal claws with strong tooth (Fig. 2b).
Figure 1a-e. Andricus calvoi sp. nov., asexual female. a) Head in frontal view; b) head in dorsal view; c) mesosoma in lateral view; d) mesosoma in dorsal view; e) propodeum.
Forewing (Fig. 3c) hyaline, 1.1× as long as body, pubescent, with cilia on margins; radial cell opened, around 3.0× as long as broad; veins light brown; areolet absent or indistinct.
Metasoma (Fig 2c) shorter than head+mesosoma, longer than high. metasomal tergum 2 with lateral patch of white setae, punctate in posterior 1/2-1/3, all subsequent terga uniformly and entirely punctate. Prominent part of ventral spine of hypopygium needle-like, 4.5-5.0× as long as broad, with sparse setae laterally, without apical tuft of setae (Fig. 2d). Body length: 3.0-3.4 mm (n = 3).
Figure 2a-g. Andricus calvoi sp. nov., asexual female. a) Antenna; b) tarsal claw; c) metasoma in lateral view; d) ventral spine of the hypopygium; e-g) hind leg, longitudinal carina of hind tibia marked with a white arrow.
Galls are developing in thin branches (Fig. 3b). It is a unilocular bud gall, woody, more or less cylindrical or globular, 8-18 mm in length and 7-15 mm in diameter, composed of 2 subunits; the upper and the lower third are constricted; the upper end is open with a depression, the edge is continuous. The gall is brown; the surface is covered by fine short brownish pubescence, which remains on the mature gall. The inner larval chamber is located in the constricted part of the gall. The single larval chamber is globular, with a hard woody wall, which is connected to the tissue of the gall. The emergence hole is located inside the upper depression.
Taxonomic summary
Type material. Holotype, asexual female deposited in JP-V collection (UB), labeled as “MEX, Parque Nacional Bosque del Pedregal, Ciudad de México, Mexico, M. Serrano col. 206A, 2,437 m asl, [19°17.283’ N, 99°12.256’ W]” (white label), “Ex. Q. obtusata, (24.vi.2012) 26.vi.2012” (white label); “Holotype Korlevicandricus tlalpanus desig. JP-V 2013” (red label), “Andricus calvoi Pujade-Villar, Cuesta-Porta & Melika, JP-V det., asexual gen.” (white label). Paratypes (3 females): 1 female with the same data as the holotype; 2 females collected in the same place and host than the holotype, but with different collection dates (24.vi.2012) 27.vi.2012: 1 female; (3.x.2010) extr. vi.2011:1 female.
Additional material. Two females collected in the same place and from the same host plant as the holotype (20.vi.2011). Both females were dissected from the galls on 16.vii.2013 (damaged material). Eight females deposited in the JP-V col. (UB): 1 female “Parque Nacional bosque del Pedregal, Tlalpan, Ciudad de México, Mexico, (MEX096), Ex Quercus sp., (19.iv.12) 01.v.2012, M. Serrano leg.”; 1 female “Santa Fe, Denominación de Coajomulco, Ciudad de México, Mexico, (MEX362), Ex Quercus sp., (23.iv.2017): 1 female , D. Cibrián-Tovar leg.”; 5 females “ Santa Fe, Coajomulco, Ciudad de México, Mexico, (MEX473), Ex Q. laeta, (22.iii.2019) 14.v.2019: 5 females, U. Barrera-Ruíz & D. Cibrián-Tovar leg.”; 1 female “Sanctórum de Lázaro Cárdenas, Tlaxcala, Mexico, (MEX644), Ex Quercus sp., (22.vi.2022) 30.vi.2022: 1 female, JP-V, VC-P, A. Equihua & E. Estrada-Venegas leg.”; 1 female “Peñón Blanco, Sierra Fría, San José de Gracia, Aguascalientes, Mexico (MEX772), Ex Quercus sp., (12.ii.2022) extracted: 1 female, R. Clark leg.”; 1 female “Puente Roto, Santa Catarina Ixtepeji, Oaxaca, Mexico, (MEX834), Ex Q. glabrescens, (25.i.2023) extracted: 1 female, R. Clark leg.”; 1 female “Carretera Maravatio-Morelia (Km. 177-178), Michoacán, Mexico, (MEX841), Ex Q. obtusata (SR-R det.), (18.vi.2023) 16-31.vii.2023: 1 female, A. Equihua & J. Pujade-Villar leg.”; 1 female “Peñón Blanco, Sierra Fría, San José de Gracia, Aguascalientes, Mexico, (MEX973), Ex Q. chihuahuensis, (20.ii.2023) extracted: 1 female, R. Clark leg.”.
Figure 3a-c. Andricus calvoi sp. nov. a) Asexual female, habitus; b) galls, deposited in the JP-V col. (UB); c) asexual female, fore wing.
Etymology. Species is named after Mr. Oriol Calvo Verderes, accused in 2019, one month after the protests in Barcelona against the sentence of the Catalan political prisoners.
Biology. The asexual generation is only known to induce galls on Q. obtusata Humb. & Bonpl., and Q. laeta (section Quercus, subsection Leucomexicanae, white oaks). The galls were collected between March and June, adults emerged between May and June.
Distribution. Mexico: Ciudad de México, and Estado de México.
Diagnosis. Andricus geniale differs from all the Andricus species, except for the species here transferred to Andricus, which have tibial carina and the last femur swollen. It differs from A. lusum comb. rev. by having the body scarcely pubescent with the glabrous mesopleuron on all its surface, the internal margin of eyes parallel, hind femur without distal lobe; differs from A. calvoi sp. nov., A. lusum, comb. rev., and A. perfectum comb. rev. for the LOL more than 1.2× as long as the diameter of lateral ocellus, the hind coxae and femora not swollen, and the propodeal carinae subparallel. Differs from A. integrum, comb. rev. by having the reticulated lower face, the mesopleuron is smooth with sparse striae across mid-height of mesopleuron.
Redescription. Asexual female (Figs. 4a-e, 5a-d, 6a-c). Head, mesosoma dark brown to black, antenna dark brown; legs chestnut brown, except dark brown to black coxae; metasoma chestnut brown.
Figure 4a-e. Andricus geniale comb. rev., asexual female. a) Head in frontal view; b) head in posterior view; c) head in dorsal view; d) head in lateral view; e) antenna.
Head (Fig. 4a-d) trapezoid, broadest part on the toruli level, 1.2× as broad as high, or less, and slightly narrower than mesosoma in frontal view, with sparse setae, denser on lower face and postgena; 2.4× as broad as long in dorsal view. Gena alutaceous, broadened behind eye in frontal view, nearly as broad as cross diameter of eye in lateral view. Malar space alutaceous with distinct rugae radiating from clypeus and reaching eye; eye 1.7× as high as length of malar space. Inner margins of eyes parallel or only slightly converging ventrally. POL 2.0× as long as OOL, OOL 1.5× as long as diameter of lateral ocellus and 1.2× as long as LOL, all ocelli ovate, of same size. Transfacial distance 1.2× as long as height of eye; toruli located at mid height of eyes, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye 1.3× as long as diameter of torulus; lower face with sparse setae, rugoso-reticulate, rugae orientated mainly dorsoventrally and units of impressed areas slightly elongated, smooth; without setae; slightly elevated median area sculptured like rest of lower face. Clypeus impressed, flat, trapezoid, broader than high, smooth, glabrous, with few long setae along ventral edge; ventrally rounded, not emarginate and without median incision; anterior tentorial pit rounded, distinct, epistomal sulcus distinct, clypeo-pleurostomal line well impressed. Frons uniformly alutaceous, without striae and setae, area between toruli and eye alutaceous; interocellar area rugose. Vertex rugose, occiput alutaceous, glabrous, with white long setae; postocciput and postgena glabrous, alutaceous-reticulate, with concentric lines around occipital foramen and postgenal bridge; posterior tentorial pit large, elongated, area below impressed; occipital foramen as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci, which diverge toward occipital foramen, postgenal bridge anteriorly slightly broader than occipital foramen. Antenna (Fig. 4e) as long as head+mesosoma, with 11 flagellomeres, scape 2.0× as long as pedicel, pedicel 1.4× as long as broad; flagellomeres subsequently broadening until apical end; all flagellomeres with sparse white short setae; F1 slightly longer than F2 and 1.8× as long as pedicel; F2 1.2× as long as F3; F3 slightly longer than F4, F5 = F6 = F7, F8 until F10 equal in length; F11 1.8× as long as F10; placodeal sensilla on F3-F11.
Figure 5a-d. Andricus geniale comb. rev., asexual female. a) Mesosoma in lateral view; b) mesosoma in frontal view; c) mesosoma in dorsal view; d) mesoscutellum.
Mesosoma (Figs. 5a-d, 6b) longer than high, with few white setae, setae denser along propleuron and on lateral propodeal area. Pronotum smooth, sparsely pubescent with delicate parallel striae; anterolateral impressed narrow area smooth, shining; propleuron coriaceous, smooth, shining in the dorsomedial part, with scattered white short setae. Mesoscutum uniformly alutaceous-reticulate, slightly broader than long (largest width measured across mesoscutum at level of base of tegulae), with white setae, denser along anterior parallel lines. Notaulus deep, distinct, complete, slightly obscured anteriorly; posteriorly converging; in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; anterior parallel line almost indistinct, slightly elevated, and finely alutaceous; parapsidal line impressed, extending well above tegula; median mesoscutal line absent or in a form of very short, impressed triangle; circumscutellar carina broad, reaching notaulus. Mesoscutellum elongated, longer than broad, trapezoid, broadest part in posterior 1/4 of its length; disk of mesoscutellum dull rugose laterally and posteriorly, coriaceous in anteromedian part, overhanging metanotum, with sparse setae. Mesoscutellar foveae transverse, with smooth, shining bottom, divided by narrow elevated coriaceous central area. Mesopleuron smooth, shining with delicate striae starting at ventrocentral part and going across mesopleuron to anterodorsal edge; lower half of anterior margin elevated into a strong carina, followed by impressed narrow furrow with striated bottom, speculum and posterior margin of mesopleuron smooth, glabrous, without longitudinal striae; mesopleural triangle smooth, glabrous, with dense, long white setae; dorsal and lateral axillar areas coriaceous, matt, with sparse white setae; axillula with delicate parallel longitudinal striae; subaxillular bar smooth, glabrous, with parallel sides, posteriorly as high as height of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/3 of its height, delimiting broad triangular smooth area with dense setae; upper part of sulcus distinct. Metascutellum coriaceous, higher than height of smooth, glabrous ventral impressed area; metanotal trough smooth, glabrous, with dense white setae; central propodeal area rectangular, smooth, shining, with irregular strong rugae; lateral propodeal carinae distinct, broad, blackish, subparallel; lateral propodeal area smooth, glabrous. Hind coxa and femora slightly swollen, without apical lobe; hind tibia with internal carinae. Tarsal claws toothed, with acute basal lobe.
Figure 6a-d. Andricus geniale comb. rev., asexual female (a-c). a) Fore wing; b) propodeum; c) metasoma in lateral view; d) galls, deposited in the AMNH.
Forewing (Fig. 6a) as long as body, hyaline, with very short cilia on margins, veins brown; radial cell opened, 3.1× as long as broad; Rs and R1 not reaching wing margin; areolet indistinct. Rs+M on all length indistinct, invisible.
Metasoma (Fig. 6c) as long as head+mesosoma, slightly longer than high in lateral view; 2nd metasomal tergum extending to 3/4 length of metasoma in dorsal view, with dense patch of white setae anterolaterally; all terga and hypopygium micropunctate posteriorly, prominent part of ventral spine of hypopygium 5.2× as long as broad in ventral view. Body length 2.0-2.5 mm.
Gall (Fig. 6d) unilocular, located on the buds, easily separable, elongated-cylindrical, cup-shaped (sometimes globular), almost or totally closed at the top. External surface silvery to yellow-brown, often with a rough surface; darker when more mature, completely bare, densely scaly or purple. Base of galls enlarged but joined by a small central point to the branch. Internally, the base of the gall is quite solid and woody, containing a centrally located circular larval chamber which, although heavily encrusted, has a distinct cell wall; upper half hollow, cup-shaped, bounded by thin to fairly thick walls occupying more than one-third the diameter of the entire gall; the walls of the upper half of the gall are quite thick and often wrinkled. Diameter up to 8.0 mm and length up to 11.0 mm.
Taxonomic summary
Type material examined. Holotype: female deposited in the AMNH “Durango, 2 N Dgo6500. Gall 11.8.31, female Spr.’32”, “Q. undata Kinsey Coll.” (white labels), “F. geniale HoloParatype” (red label), “AMNH_IZC 00322882” (white label with a QR code). Paratypes: 2 females deposited in the AMNH: “Durango, 2 N Dgo6500. Mex ꚛ, 11.8.31, female Spr.’32”, “Q. undata Kinsey Coll.” (white labels), “F. geniale Paratype” (red label); a female with the same data at HMNH.
Figure 7a-f. Andricus integrum comb. rev., asexual female. a) Head in frontal view; b) head in lateral view; c) head in dorsal view; d) head in posterior view; e) antenna; f) propleuron and first coxae.
Biology. Only the asexual generation is known; adults emerge in spring. Galls on Q. undata Trell. (section Quercus, subsection Leucomexicanae, white oaks) (Kinsey 1937b).
Distribution. Mexico: state of Durango (Kinsey, 1937b).
Diagnosis. Differs from all the Andricus species, except for the species here transferred to Andricus, which have tibial carina and the last femur swollen. It differs from A. lusum comb. rev. by scarcely pubescent body, with mesopleuron glabrous on its entire surface, the internal margin of eyes parallel, the hind femur without distal lobe; differs from A. calvoi sp. nov., A. lusum comb. rev., and A. perfectum comb. rev. by the LOL which is more than 1.2× as long as the diameter of lateral ocellus, the hind coxae and femora are not swollen, and the propodeal carinae subparallel. Differs from A. geniale comb. rev. by dull, rugose-reticulate lower face, with vertical carinae radiating from clypeus; the mesopleuron is completely striated, except for the smooth speculum.
Redescription. Asexual female (Figs. 7a-f, 8a-e, 9a-b). Head, mesosoma, antenna dark brown to black; legs dark brown, except dark brown to black coxae; metasoma chestnut brown.
Head (Fig. 7a-d) trapezoid, broadest part on toruli level, 1.5× as broad as high and slightly narrower than mesosoma in frontal view, with sparse setae, denser on lower face and postgena; 2.2× as broad as long in dorsal view. Gena alutaceous, units of sculpture orientated dorsoventrally, elongated, giving striate aspect in lower part; gena broadened behind eye in frontal view, slightly narrower than cross diameter of eye in lateral view. Malar space with distinct rugae radiating from clypeus and reaching eye, area between rugae matte, alutaceous; eye 1.9× as high as length of malar space. Inner margins of eyes parallel or only slightly converging ventrally. POL 2.0× as long as OOL, OOL 1.5× as long as diameter of lateral ocellus and only slightly longer than LOL. Transfacial distance 1.2× as long as height of eye; toruli located at mid height of eyes, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye 1.5× as long as diameter of torulus; lower face with sparse setae, rugoso-reticulate, rugae radiating from clypeus and reaching toruli, slightly elevated median area sculptured like rest of lower face. Clypeus impressed, flat, rectangular, broader than high, smooth, glabrous; ventrally rounded, not emarginate and without median incision; anterior tentorial pit rounded, distinct, epistomal sulcus distinct, clypeo-pleurostomal line well impressed. Frons uniformly alutaceous, without striae and setae, area between toruli and torulus and eye alutaceous; interocellar area rugose. Vertex rugose, occiput alutaceous, glabrous, with white long setae; postocciput with delicate transverse striae, postgena glabrous, alutaceous-reticulate, around occipital foramen and postgenal bridge smooth, shining; posterior tentorial pit large, elongated, area below impressed; occipital foramen 2.0× as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci, which diverge toward occipital foramen, postgenal bridge anteriorly broader than occipital foramen. Antenna (Fig. 7e) as long as head+mesosoma, with 11 flagellomeres, scape 2.0× as long as pedicel, pedicel subglobose, as long as broad; flagellomeres subsequently broadening until apical end; all flagellomeres with sparse white short setae; F1 slightly longer than F2 and 1.8× as long as pedicel; F2 1.2× as long as F3; F3 slightly longer than F4, F5 = F6 = F7, F8 until F10 equal in length; F11 1.8× as long as F10; placodeal sensilla on F3-F11.
Figure 8a-e. Andricus integrum comb. rev., asexual female. a) Mesosoma in lateral view; b) mesosoma in dorsal view; c) mesoscutellum; d) propodeum; e) fore wing.
Mesosoma (Figs. 7f, 8a-d) longer than high, with few white setae, setae denser along propleuron and on lateral propodeal area. Pronotum smooth, glabrous dorsally, with transverse striae laterally; anterolateral impressed narrow area smooth, shining; propleuron alutaceous, smooth shining, with scattered white short setae. Mesoscutum uniformly alutaceous-reticulate, slightly broader than long (largest width measured across mesoscutum at level of base of tegulae), with white setae, denser along anterior parallel lines. Notaulus deep, distinct, complete; posteriorly converging; in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; anterior parallel line, glabrous, indistinct; parapsidal line impressed, extending well above tegula; median mesoscutal line absent or in a form of a very short, impressed triangle; circumscutellar carina broad, reaching notaulus. Mesoscutellum elongated, longer than broad, trapezoid, with subparallel sides; broadest part in posterior 1/4 of its length; disk of mesoscutellum dull rugose laterally and posteriorly, reticulated in anteromedian part, overhanging metanotum, with sparse setae. Mesoscutellar foveae transverse, with smooth, shining bottom, divided by narrow elevated smooth central area. Mesopleuron shining in between striae going across mesopleuron, anteriorly mesopleuron with narrow foveolate stripe, lower half of anterior margin elevated into a strong carina, followed by an impressed narrow furrow with bottom striated, speculum smooth, glabrous, without transverse longitudinal striae; mesopleural triangle smooth, glabrous, with dense, long white setae; dorsal and lateral axillar areas alutaceous, matte, with sparse white setae; axillula with delicate parallel longitudinal striae; subaxillular bar smooth, glabrous, with parallel sides, posteriorly as high as height of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/3 of its height, delimiting broad triangular smooth area with dense setae; upper part of sulcus distinct. Metascutellum with longitudinal rugae, 2.0× as high as height of smooth, glabrous ventral impressed area; metanotal trough smooth, glabrous, with dense white setae. Central propodeal area rectangular, smooth, shining, without irregular rugae; lateral propodeal carinae slightly bent outwards (parallel in the holotype), distinct, broad, blackish; lateral propodeal area smooth, glabrous, with long dense white setae. Nucha dull rugose dorsally, with parallel sulci laterally. Hind coxa and femora swollen, femora without apical lobe (Fig. 9a). Tarsal claws toothed, with acute basal lobe.
Figure 9a-c. Andricus integrum comb. rev., asexual female (a-b). a) Hind leg; b) metasoma in lateral view; c) galls, deposited in the AMNH.
Forewing (Fig. 8e) above as long as body, hyaline, with very short cilia on margin, veins brown; radial cell opened, 2.8× as long as broad; Rs and R1 not reaching wing margin; areolet indistinct. Rs+M on all length indistinct, invisible.
Metasoma (Fig. 9b) as long as head+mesosoma, slightly longer than high in lateral view; 2nd metasomal tergum extending to 3/4 length of metasoma in dorsal view, with dense patch of white setae anterolaterally; all terga and hypopygium micropunctured, prominent part of ventral spine of hypopygium 7.2× as long as broad in ventral view. Body length 2.1-2.6 mm (according to Kinsey, 1937b).
Gall (Fig. 9c). Similar to A. geniale but a little smaller: diameter up to 7 mm, length up to 10 mm.
Figure 10a-f. Andricus lusum comb. rev., asexual female. a) Head in frontal view; b) head in dorsal view; c) mesosoma in lateral view; d) mesosoma in dorsal view; e) mesoscutellum; f) propodeum.
Taxonomic summary
Type material examined. Holotype: female deposited in the AMNH “S. Luis Potosí, S.L.P. 15W, 8000, Mex. ꚛ, gall 11.30.31, female 1932”, “Q. potosina Kinsey Coll.” (white labels), “F. integrum HoloParatype” (red label), “AMNH_IZC 00322883” (white label with a QR code). Paratypes: 2 females deposited in the AMNH “S. Luis Potosí, S.L.P. 15W, 8000, Mex. ꚛ, gall 10.30.31, female 1932”, “Q. potosina Kinsey Coll.” (white labels), “F. integrum Paratype” (red label); a female with the same data at HMNH.
Additional material. One female deposited in the AMNH “San Felipe, Gto., 20 SW, 3000’, Mex. ꚛ, 12.7.31, female, spr. 32”, “Q. chihuahuens Kinsey coll.”, “F. integrum Kinsey coll. 37”.
Biology. The asexual generation is only known to induce galls on Q. potosina Trel., Q. chihuahuensis Trel., Q. undata Trel., and Q. rugosa Né (= Q. reticulata Née) (section Quercus, subsection Leucomexicanae, white oaks), according to Kinsey (1937b). Adults probably emerge in spring.
Distribution. México: Guanajuato and San Luis Potosí (Kinsey, 1937b).
Diagnosis. Differs from all the Andricus species, except for the species here transferred to Andricus, which have tibial carina and the last femur swollen. This species belongs to the group of species that have the hind femur with a distal lobe and the LOL shorter than diameter of lateral ocelli, along with A. calvoi sp. nov. and A. perfectum comb. rev.It differs from A. perfectum comb. rev. by a faintly reticulate, shining lower face, carinae radiating from clypeus not extending towards toruli, the pronotum punctured with striae only on lateral margin, and the notaulus incomplete reaching more than 3/4 of length of mesonotum. Differs from A.calvoi sp. nov., by the space between notauli in the posterior margin on mesoscutum, which is subequal to the space between notauli and parapsidal lines, and parapsidal lines parallel or slightly bent outwards.
Redescription. Asexual female (Figs. 10a-f, 11a-b). Head, mesosoma dark brown to black, antenna dark brown; legs chestnut brown, except dark brown to black coxae; metasoma chestnut brown.
Head (Fig. 10a-c) trapezoid, broadest part below the toruli level, 1.5× as broad as high and slightly narrower than mesosoma in frontal view, with sparse setae, denser on lower face and postgena; 2.6× as broad as long in dorsal view. Gena finely reticulate, shining, broadened behind eye in frontal view, nearly as broad as cross diameter of eye in lateral view. Malar space alutaceous with distinct rugae radiating from clypeus and reaching eye; eye 2.1× as high as length of malar space. Inner margins of eyes parallel. POL 2.0× as long as OOL, OOL 1.5× as long as diameter of lateral ocellus and 1.3× as long as LOL, all ocelli ovate, of same size. Transfacial distance slightly longer than height of eye; toruli located at mid height of eyes, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye 1.2× as long as diameter of torulus; lower face with dense setae, shining, faintly reticulate-alutaceous, rugae orientated mainly dorsoventrally and units of impressed areas slightly elongated, smooth; slightly elevated median area sculptured like rest of lower face. Clypeus impressed, flat, rectangular, broader than high, smooth, glabrous, with few long setae along ventral edge; ventrally rounded, not emarginate and without median incision; anterior tentorial pit rounded, distinct, epistomal sulcus distinct, clypeo-pleurostomal line well impressed. Frons uniformly reticulated, without setae, area between toruli and torulus and eye alutaceous; interocellar area rugose. Vertex rugose, occiput alutaceous, glabrous, with white long setae; postocciput and postgena glabrous, alutaceous-reticulate, with concentric lines around occipital foramen and postgenal bridge. Antenna of the type material broken past F4, scape 1.5× as long as pedicel, pedicel 1.2× as long as broad; flagellomeres subsequently broadening until apical end; all flagellomeres with sparse white short setae; F1 slightly longer than F2 and 2.4× as long as pedicel; F2 slightly longer than F3; F3 slightly longer than F4, subsequent flagellomeres are missing.
Mesosoma (Fig. 10c-f) longer than high, with dense white setae, setae denser along propleuron and on lateral propodeal area. Pronotum with delicate parallel striae and impressed setose pits; anterolateral impressed narrow area smooth, shining; propleuron coriaceous, smooth shining in the dorsomedial part, with scattered white short setae. Mesoscutum uniformly alutaceous-reticulate, units transversally oriented on anterior half, posterior half alutaceous-reticulate with impressed setal points; slightly broader than long (largest width measured across mesoscutum at level of base of tegulae), with white setae. Notaulus deep, distinct, incomplete, absent in anterior 1/4-1/5 of mesoscutum length, anterior insertion into parascutal carina visible, posteriorly converging; in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; anterior parallel line distinct, slightly elevated, extending to 1/2 of mesoscutum length; parapsidal line impressed, extending until shortly passed the tegula; median mesoscutal line absent; circumscutellar carina broad, reaching notaulus. Mesoscutellum elongated, longer than broad, trapezoid, broadest part in posterior 1/4 of its length; disk of mesoscutellum uniformly dull rugose, overhanging metanotum, with sparse setae. Mesoscutellar foveae transversally ovate, with smooth, shining bottom, divided by broad elevated rugose central area, half as wide as mesoscutellar fovea. Mesopleuron smooth, shining with delicate striae going across mesopleuron; anteriorly with narrow foveolate stripe, speculum and posterior margin smooth, glabrous, without transverse longitudinal striae; mesopleural triangle coriaceous, glabrous, with dense, long white setae; dorsal and lateral axillar areas alutaceous with faint rugae, dull, with sparse white setae; axillula with delicate parallel longitudinal striae; subaxillular bar smooth, glabrous, triangular, posteriorly as high as height of metanotal trough. Metapleural sulcus reaching mesopleuron in upper 1/3 of its height, delimiting broad triangular smooth area with dense setae; upper part of sulcus distinct. Metascutellum rugose, as high as the smooth, glabrous ventral impressed area; metanotal trough smooth, glabrous, with dense white setae; central propodeal area rectangular, smooth, shining, with irregular strong rugae. Lateral propodeal carinae distinct, broad, slightly bent outwards; central propodeal area smooth with faint rugae; lateral propodeal area punctured-smooth, uniformly pubescent. Nucha dull rugose dorsally, with parallel sulci laterally. Hind legs (Fig. 11b) with coxa broad, femur strongly broadened, with apical lobe; hind tibia with ventral carina reaching almost to the base of the tibia; tarsal claws toothed, with acute basal lobe.
Forewing (Fig. 11a) above as long as body, hyaline, with very short cilia on margins, veins brown; radial cell opened, 3.2× as long as broad; Rs and R1 not reaching wing margin; areolet indistinct. Rs+M on all length indistinct, invisible.
Metasoma (Fig. 11b) as long as head+mesosoma, slightly longer than high in lateral view; 2nd metasomal tergum extending to 3/4 length of metasoma in dorsal view, with dense patch of white setae anterolaterally, with micropunctures; all subsequent terga and hypopygium with dense micropunctures, prominent part of ventral spine broken. Body length 3.0 mm (according to Kinsey, 1937b).
Gall (Fig. 11c). According to Kinsey (1937b): galls are cylindric, with the thin-walled top half of the gall widely open or flaring, making the gall wide open cup-shaped with a thick and solid base, the open cavity almost as wide as the whole gall. The gall silvery to yellowish-brown; young galls quite scurfy, older galls naked. The diameter of the body of the gall up to 15.0 mm, averaging nearer 11.0 mm; the flared top up to 19.0 mm in diameter; length up to 19.0 mm, averaging near 15.0 mm.
Taxonomic summary
Type material examined. Holotype asexual female deposited in the AMNH: “Morelia, 14E; Mich 7000; Mex gall 12.27.31”, “Q. conglomerata, Kinsey Coll.” (white labels), “Femuros lusum HoloParatype” (red label), “AMNH_IZC 00267205” (white label with a QR code). Paratypes: only galls with the same data as the holotype.
Biology. The asexual generation is known for inducing galls on a single species of white oaks: Q. rugosa Née (= conglomerata Trell.; section Quercus, subsection Leucomexicanae, white oaks). The single adult of this species (holotype) probably obtained after gall cut; galls were collected on December 27, 1931 (Kinsey, 1937b).
Diagnosis. Differs from all the Andricus species, except for the species here transferred to Andricus, with tibial carina and the last femur swollen. This species belongs to the group of species that have the hind femur with a distal lobe and the LOL shorter than the diameter of lateral ocellus, A. calvoi sp. nov., and A. lusum comb. rev. Differs from those 2 species by dull rugose-reticulate lower face, with vertical carinae radiating from the clypeus towards the toruli; the pronotum completely striate with parallel curved striae; and the notaulus incomplete reaching slightly more than half-length of mesonotum.
Redescription. Asexual female (Figs. 12a-f, 13a-c, 14a-c). Head, antenna dark brown; mesosoma blackish; legs chestnut brown, except dark brown to black coxae; metasoma chestnut brown.
Head (Figs. 12a-d, 13c) trapezoid, broadest part on the toruli level, 1.4× as broad as high and as broad as mesosoma in frontal view, with sparse setae, denser on postgena; 2.4× as broad as long in dorsal view. Gena alutaceous, units of sculpture orientated dorsoventrally, elongated, giving striate aspect of gena in its lower part; gena slightly broadened behind eye in frontal view, slightly broader than cross diameter of eye in lateral view. Malar space with distinct rugae radiating from clypeus and reaching eye, area between rugae smooth, shining; eye 2.2× as high as length of malar space. Inner margins of eyes parallel. POL 2.1× as long as OOL, OOL 1.7× as long as diameter of lateral ocellus and only slightly longer than LOL, lateral ocelli bigger than median ocellus. Transfacial distance subequal to height of eye; toruli located slightly above height of eyes, frons at least 2.0× shorter than height of lower face, diameter of antennal torulus 2.4× as long as distance between them, distance between torulus and eye 1.4× as long as diameter of torulus; lower face with sparse setae, delicately coriaceous, striae radiating from clypeus extending to 1/3 of lower face height; slightly elevated median area delicately coriaceous, on both sides with delicate striae radiating from clypeus and extending to 1/3 of lower face height. Clypeus convex, rectangular, broader than high, delicately coriaceous; ventrally rounded, not emarginate and without median incision; anterior tentorial pit rounded, distinct, epistomal sulcus distinct, clypeo-pleurostomal line well impressed. Frons dull rugose, without striae and setae, area between toruli and torulus and eye smooth, shining; interocellar area rugose. Vertex rugose, occiput alutaceous, glabrous, with white long setae; postocciput and postgena glabrous, alutaceous-reticulate, around occipital foramen and postgenal bridge smooth, shining; posterior tentorial pit large, elongated, area below impressed; occipital foramen as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci which diverge toward occipital foramen, postgenal bridge anteriorly broader than occipital foramen. Antenna (Fig. 12e) slightly longer than head+mesosoma, with 11 flagellomeres, scape 2.7× as long as pedicel, pedicel subglobose, as long as broad; flagellomeres; flagellomeres from F1 until F7 of same width, with sparse white short setae; F1 = F2 and 2.4× as long as pedicel; F2 1.65× as long as F3; F3 slightly shorter than F4, F5 = F6 = F7, from F8 flagellomeres subsequently broadening until apical end, F8 until F10 equal in length; F11 1.8× as long as F10; placodeal sensilla on F2-F11.
Mesosoma (Figs. 12f, 13a-c, 14c) longer than high, with few white setae, setae denser along propleuron and on lateral propodeal area. Pronotum smooth, glabrous dorsally, laterally delicately uniformly coriaceous, without transverse striae; anterolateral impressed narrow area smooth, shining; propleuron alutaceous, smooth shining, with scattered white short setae. Mesoscutum uniformly alutaceous-reticulate, with distinct piliferous points scattered all over the mesoscutum, slightly broader than long (largest width measured across mesoscutum at level of base of tegulae), with white setae, denser along anterior parallel lines. Notaulus weakly impressed, narrow, incomplete, reaching to 3/4 of mesoscutum length; posteriorly converging; in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; anterior parallel line indistinct; parapsidal line impressed, extending to the level of tegula; median mesoscutal absent; circumscutellar carina broad, reaching notaulus. Mesoscutellum elongated, longer than broad, trapezoid, with subparallel sides; broadest part in posterior 1/4 of its length; disk of mesoscutellum uniformly dull rugose, overhanging metanotum, with sparse setae. Mesoscutellar foveae transverse, with smooth, shining bottom, divided by narrow elevated coriaceous central area. Mesopleuron alutaceous, delicate short transverse striae ventrally only in the anterolateral part, anteriorly mesopleuron with narrow foveolate stripe, speculum smooth, glabrous, without transverse longitudinal striae; mesopleural triangle delicately coriaceous, with dense, long white setae; dorsal and lateral axillar areas delicately coriaceous, matte, with sparse white setae; axillula with delicate parallel longitudinal striae; subaxillular bar smooth, glabrous, triangular, in the highest posterior part as high as height of metanotal trough; metapleural sulcus indistinct, reaching mesopleuron in upper 1/3 of its height, delimiting broad triangular smooth area with dense setae; upper part of sulcus indistinct. Metascutellum with longitudinal rugae, as high as height of smooth, glabrous ventral impressed area; metanotal trough smooth, glabrous, with dense white setae; central propodeal area rectangular, smooth, shining, without irregular rugae; lateral propodeal carinae slightly bent outwards, distinct, broad, blackish; lateral propodeal area smooth, glabrous, with long dense white setae. Nucha dull rugose dorsally, with parallel sulci laterally. Hind femora (Fig. 14b) strongly broadened, hind tibia with ventral carina reaching almost to the base of the tibia. Tarsal claws toothed, with acute basal lobe.
Forewing (Fig. 14a) above as long as body, hyaline, with dense cilia on margins, veins whitish, hardly traceable; radial cell opened, 2.8-3.2× as long as broad; Rs and R1 not reaching wing margin; areolet indistinct and Rs+M, indistinct invisible.
Metasoma (Fig. 14b) as long as head+mesosoma, longer than high in lateral view; 2nd metasomal tergum extending to 3/4 length of metasoma in dorsal view, with numerous white setae anterolaterally, smooth, shining; all subsequent terga and hypopygium with micropunctures, prominent part of ventral spine of hypopygium 4.5× as long as broad in ventral view. Body length 2.4-3.5 mm (according to Kinsey, 1937b).
Gall (Fig. 14d). Similar to A. geniale and A. perfectum, but larger (diameter up to 16 mm, never more than 13 mm in length), nearly globular or slightly elongated or flattened. In section, the upper cavity is exceedingly small occupying the upper half of the gall, never wider than larval chamber (often narrower), almost completely closed by very thick and solid walls, although a central pucker at the apex of the gall still indicates the opening to the cavity; outside of the gall silvery grey to purplish brown, conspicuously covered with a purplish-white bloom.
Taxonomic summary
Type material examined. Holotype: female deposited in the AMNH “Chilpancingo, Gro 6S, 4000’, Mex; gall 1.8.32, 56 females 5.10.32”, “Q. macrophylla Kinsey Coll.” (white labels), “F. perfectum HoloParatype” (red labels), “AMNH_IZC 00322886” (white label with a QR code). Paratypes: 33 females deposited in the AMNH “Chilpancingo, Gro 6S, 4000, Mex; gall 1.8.32, 56 females 5.10.32”, “Q. macrophylla Kinsey Coll.” (white labels), “F. perfectum Paratype” (red label).
Biology. The asexual generation is only known to induce galls on Q. magnoliifolia Née (= Q. macrophylla Née; section Quercus, subsection Leucomexicanae, white oaks). Adults emerge from May to July (Kinsey, 1937b).
Distribution. México: Guerrero (Kinsey, 1937b).
Femuros Kinsey, 1937
Type species: Femuros repandae Kinsey, 1937.
Diagnosis. Only asexual females known. Femuros is characterized as having a lower head smooth or almost smooth, without radiating carina from clypeus or very short, neither malar sulcus, occipital carina visible behind the ocelli, with transscutal articulation, winged forms, hind femur broadened on ventral margin near distal end, and an internal carina on the hind tibia. Some species of Andricus, Amphibolips, and Odontocynips can have broadened hind femur but in these cases the lower face is always sculptured (rugose and/or with radiating carina from clypeus). Very few Cynipini genera with transscutellar articulation lack facial striae and a malar groove: Dros, Cynips, Eumayriella, Heteroecus, and Kokkocynips. Eumayriella, unlike Femuros, is brachypterous, lacks scutellar pits and has a dorso-ventrally flat mesosoma. Heteroecus and Kokkocynips have sculptured face and simple tarsal claws, unlike Femuros; furthermore, the metanotum is sculptured in Kokkocynips (without sculpture in Femuros). Cynips have the head sculptured and, the mesoscutum and mesopleuron smooth or practically smooth (head smooth and sculptured mesoscutum and mesopleuron in Femuros); also, agamic forms of Cynips have a wide forming ventral spine provided with a dense truncated tuft in agamic forms (long spine without the apical tuft in Femuros). Dros has the head in frontal view smooth and mesoscutum also smooth but in Femuros the fronts and mesoscutum are sculptured. Finally, Sphaeroteras and some Neuroterus species have the lower face smooth, as Femuros, but have also malar sulcus (absent in Femuros); besides, in Neuroterus the transscutal articulation is absent (present in Femuros).
Redescription. Asexual female. Head, mesosoma dark brown, chestnut brown to black, rusty brown or yellowish to yellow amber; antenna of the same color or paler than head; metasoma dark brown to reddish brown; mouthparts, legs yellowish-brown, light brown to dark brown, with coxae always darker.
Head alutaceous, with sparse setae, denser on lower face, rounded, trapezoid or transverse, 1.2-1.4× as broad as high and slightly broader than mesosoma in frontal view; 2.0-2.2× as broad as long in dorsal view. Gena alutaceous, either not or broadened behind eye in frontal view, narrower than transverse diameter of eye in lateral view. Malar space alutaceous, with striae radiating from clypeus and reaching eye, malar sulcus absent (Figs. 15a, 16a, 18a, 21a); eye 2.8-4.0× as high as length of malar space. Eyes parallel or slightly converging ventrally. POL 1.8-3.6× as long as OOL, OOL slightly shorter or 1.3-1.6× as long as diameter of lateral ocellus and nearly equal to LOL, all ocelli ovate, of same size (Figs. 16b, 18c, 21c). Transfacial distance slightly shorter or equal to height of eye; toruli located at mid-height or level with the lower half of eye; frons higher than lower face, diameter of antennal torulus 1.3-2.0× as long as distance between them, distance between torulus and eye slightly shorter or 1.2-1.5× as long as diameter of torulus; lower face smooth, glabrous, alutaceous to delicately coriaceous, with white setae, with or without short delicate striae; slightly elevated median area alutaceous to delicately coriaceous, with few setae. Clypeus rounded, quadrangular or rectangular, nearly 2.0× as broad as high, smooth, glabrous, with few setae; ventrally rounded, not emarginate and without median incision; anterior tentorial pit large, rounded, distinct, epistomal sulcus distinct, clypeo-pleurostomal line well impressed. Frons uniformly alutaceous, without striae and setae, interocellar area alutaceous. Vertex, occiput and postocciput alutaceous; postgena smooth or alutaceous, with or without setae; posterior tentorial pit large, elongated, area below impressed; occipital foramen slightly shorter or as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci which strongly diverge toward occipital foramen, postgenal bridge anteriorly slightly broader than occipital foramen. Antenna longer than head+mesosoma, with 11-12 flagellomeres (in some specimens the suture between F12 and F11 hardly visible), pedicel longer than broad; flagellomeres subsequently broadening towards apical end; F1 1.6× as long as pedicel and 1.3× as long as F2; F2 1.1× as long as F3; F3 = F4 = F5, F5 1.1× as long as F6, F6 to F10 nearly equal in length; F11 2.1× as long as F10; placodeal sensilla on F3-F11(F12).
Mesosoma slightly longer than high, with a few white setae, denser along propleuron and laterally. Pronotum smooth, glabrous, with parallel striae and sparse setae laterally; propleuron smooth, glabrous. Mesoscutum uniformly and entirely reticulate, with a few white setae along notauli (Fig. 17a), slightly longer than broad (greatest width measured across mesoscutum level with base of tegulae). Notaulus complete, in some species indistinct in anterior 1/3, impressed in posterior 2/3 of mesoscutum length, notaular furrow alutaceous or smooth; posteriorly strongly converging; anterior parallel line invisible or indistinctly impressed and extending to 1/3 of mesoscutum length; parapsidal line marked with broad impressed area; median mesoscutal line absent or short and impressed only posteriorly; parascutal carina broad, reaching pronotum. Mesoscutellum ovate, slightly longer than broad; disk of mesoscutellum, rugose, reticulated or dull rugose only laterally and posteriorly, overhanging metanotum, glabrous or with sparse long setae; circumscutellar carina complete. Mesoscutellar foveae transverse, about 2.0× as broad as high, with smooth glabrous bottom, divided by narrow rugose elevated central carina. Mesopleuron entirely covered with delicate interrupted parallel striae, without setae or only setose ventrally (Figs. 19a, 22a); mesopleural triangle smooth, glabrous, with a few strong irregular striae and long white setae; dorsal and lateral axillar areas smooth, glabrous, without setae; axillula with delicate parallel longitudinal striae; subaxillular bar smooth, glabrous, triangular, posteriorly as high as height of metanotal trough; metapleural sulcus reaching mesopleuron at mid-height or slightly above, upper part of sulcus also distinct, sulcus separating smooth, glabrous area, with some setae in lower part. Metascutellum smooth, glabrous or coriaceous, as high as height of smooth, glabrous ventral impressed area; metanotal trough smooth, glabrous, without setae; central propodeal area smooth, glabrous, with or without delicate rugae in posterolateral part; several parallel indistinct, interrupted, incomplete lateral propodeal carinae bent outwards in mid-height or posterior 1/3 (Figs. 19c-22c); in some cases lateral carinae absent; lateral propodeal area smooth, with long white setae and piliferous points. Nucha with strong longitudinal sulci dorsally and laterally. Tarsal claws with basal lobe.
Forewing longer than body, hyaline, with distinct dense cilia on margin, veins dark brown, radial cell open, 3.2-4.5× as long as broad; Rs not reaching wing margin, R1 indistinct, less pigmented, not reaching wing margin; areolet small, triangular, closed and distinct. Rs+M inconspicuous, its projection reaching basalis slightly below or in lower half of its height.
Metasoma as long as or longer than head+mesosoma, slightly longer or shorter than high in lateral view; 2nd metasomal tergum extending to half-length of metasoma in dorsal view, with few white setae anterolaterally, without micropunctures; third and fourth terga smooth, glabrous, without micropunctures; subsequent terga without or with rare delicate micropunctures (Figs. 15d, 20b). Hypopygium with or without micropunctures, prominent part of ventral spine of hypopygium 2.3-5.1× as long as broad in ventral view, with a few white setae ventrally. Body length 1.5-2.5 mm.
Femuros bracteatus (Weld, 1944), comb. nov.
Fig. 15a-d
Andricus bracteatus Weld, 1944: 8.
Diagnosis. Femuros bracteatus is most closely related to F. repandae and both differ from F. ruidum and F. ocri in having the POL around 1.5× as long as OOL, OOL 1.5× longer than diameter of lateral ocellus, the height of eye 2.0× longer than malar space, and the transfacial distance 1.3× as long as height of eye. Femuros bracteatus differs from F. repandae by the anterior parallel lines and parapsidal lines of the same color as the rest of the mesoscutum; mesoscutellar foveae are divided by a wide rugose elevated area, while in F. bracteatus the mesoscutum with the anterior parallel lines and parapsidal lines are darkened, chestnut colored; the mesoscutellar foveae are divided by a thin and sharp carina.
Redescription. Asexual female (Figs. 15a-b, 15d). Head, mesosoma, metasoma and legs reddish-brown, without darker areas in anterior parallel lines and parapsidal lines; antenna dark brown, last flagellomere darker; tegula yellowish. Wing veins brown.
Head (Fig. 15a-c) transversally oval, broadest part below toruli, broader than high and narrower than mesosoma in frontal view, with short dense white setae; 1.3× as broad as high in anterior view and head 2.0× as broad as long in dorsal view. Gena alutaceous to smooth, slightly broadened behind eye, as broad as cross diameter of eye, measuring along transfacial line. Malar space weak alutaceous, without striae radiating from clypeus, malar sulcus absent; eye 2.1× as high as length of malar space. Inner margins of eyes parallel. POL 2.6× as long as OOL, OOL as long as diameter of lateral ocellus and as long as LOL, all ocelli rounded, of same size. Transfacial distance 1.1× as long as height of eye; toruli located above mid height of head, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye almost as long as diameter of torulus. The lower face almost smooth, raised central part below the toruli alutaceous, pubescent. Clypeus impressed, coriaceous, rounded ventrally, medially not incised, anterior tentorial pits distinct; epistomal sulcus and clypeo-pleurostomal line distinctly impressed. Frons uniformly coriaceous, without striae, with few setae, piliferous points visible; interocellar area coriaceous with piliferous points; space between lateral ocellus to compound eye almost smooth and shining. Vertex and occiput coriaceous; postocciput and postgena glabrous, alutaceous-reticulate, with concentric lines around occipital foramen and postgenal bridge; posterior tentorial pit large, elongated, area below impressed; occipital foramen as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci which strongly diverge toward occipital foramen, postgenal bridge anteriorly slightly broader than occipital foramen. Antenna (Fig. 15a-c) shorter than length of head + mesosoma, with 11 flagellomeres; pedicel 1.5× as long as broad; flagellomeres not broadening until apical end; F1 1.2× as long as F2 and 3.6× as long as pedicel; subsequent flagellomeres progressively shortened; F3 = F4, F5 = F6, F7 to F10 equal in length F11 longer than F10, sometimes F11 with a very inconspicuous suture and thus antenna with 12 flagellomeres; placodeal sensilla on F5-F11.
Mesosoma (Fig. 15b-c) 1.2× as long as high, concave in lateral view, pubescent. Sides of pronotum alutaceous, with white setae and weak and parallel carinae in upper lateroposterior part, anterolateral rim of pronotum inconspicuous. Mesoscutum 1.2× as broad as long in dorsal view; punctured, without rugae, pubescent, with dense piliferous points. Notaulus complete, deep, straight, converging posteriorly, in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; median mesoscutal line present; parapsidal lines and anterior parallel lines differentiated by alutaceous sculpture. circumscutellar carina absent. Mesoscutellum as long as broad, uniformly rugose; overhanging metanotum, with sparse long setae. Mesoscutellar foveae differentiated, delimited posteriorly by a broad rugose area, triangular, deep, with smooth bottom, separated by a median carina. Mesopleuron completely pubescent, weakly reticulate with delicate carinae and sparse setae anteriorly, speculum smooth and shining; mesopleural triangle alutaceous, with dense white setae, hiding the surface sculpture; dorsal and lateral axillar areas with delicate parallel longitudinal carinae, glabrous; axillula alutaceous, pubescent; subaxillular bar smooth, glabrous, triangular, posteriorly as high slightly shorter than high of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/2 of its height; upper part of sulcus distinct; lower part of sulcus delimiting big area with dense long white setae. Metascutellum subrectangular, coriaceous with some weak longitudinal carina, curved ventrally; metanotal trough alutaceous, with dense setae; ventral bar of metanotal trough coriaceous dorsally and smooth ventrally; central propodeal area nearly smooth and glabrous; lateral propodeal carinae parallel by converging toward posterior end; lateral parts of propodeum uniformly alutaceous, densely pubescent. Nucha almost smooth and shining dorsally, with parallel sulci laterally. Legs (Fig. 15d) with hind coxa broad, hind femur strongly broadened, with apical lobe; hind tibia with ventral carina on the ventral part extending to half the length of the tibia; base of tarsal claws with strong tooth.
Forewing (Fig. 15b) hyaline, 1.1× as long as body, pubescent, with cilia on margins; radial cell opened, around 2.6× as long as broad; veins light brown; areolet present. Rs+M vein obsolete heading towards the middle of the basal vein.
Metasoma (Fig. 15d) shorter than head+mesosoma, longer than high. 2nd metasomal tergum occupying 3/4 parts on the metasoma, with lateral patch of white setae, without micropunctures or micropunctures obsolete, all subsequent terga uniformly and entirely smooth or with obsolete micropunctures. Prominent part of ventral spine of hypopygium needle-like, around 2.6× as long as broad, with sparse setae laterally, without apical tuft of setae. Body length: 2.9 mm (Weld 1944).
Gall (Fig. 15c). Globular, bud gall up to 8.4 mm in diameter, more or less with short bracts usually on apical half. The internal parenchyma is cavernous with numerous lamellae connecting the outer shell and the larval chamber.
Taxonomic summary
Type material examined. Holotype, asexual female deposited in the USMN with the following labels “Young, Ariz.” (white label), “Type No. 56377, U.S.N.M.” (red label), “Andricus bracteatus 1942 Weld” (white label), “USNMENT 00779931” (white label with a QR code). Holotype examined through the images available at the website of the Entomology Department in the USNM: http://n2t.net/ark:/65665/3e449e73d-2c5a-49ee-8f2b-94f3ac9f237c
Biology. Only the asexual generation is known. The holotype was collected from an unknown Quercus, but the paratypes are reported as inducing galls on Q. oblongifolia Torr. and Q. rugosa (= diversicolor Trel.). According to Weld (1944) similar galls are also reported in Q. arizonica Sarg., and Q. toumeyi Sarg. (section Quercus, subsection Leucomexicanae, white oaks). The galls were collected between December and February, and the adults emerge in late March (Weld, 1944).
Figure 11a-c. Andricus lusum comb. rev., asexual female (a-b). a) Fore wing; b) metasoma in lateral view; c) galls, deposited in the AMNH.
Diagnosis. Differs from F. repandae by the POL 2.0× as long as OOL, OOL similar in length to the diameter of lateral ocellus, height of eye 2.5× as long as malar space, and the transfacial distance subequal to the height of the eye. Differs from F. ruidum by the sparsely pubescent mesopleuron, which is smooth, with scattered piliferous points, the speculum smooth and glabrous at least on half of its length, and the propodeal carinae are parallel.
Figure 12a-f. Andricus perfectum comb. rev., asexual female. a) Head in frontal view; b) head in posterior view; c) head in dorsal view; d) head in lateral view; e) antenna; f) mesosoma in dorsal view.
Redescription. Asexual female (Figs. 16a-e, 17a-c). Head, mesosoma, metasoma and legs reddish-brown (with some darker areas in anterior parallel lines and parapsidal lines); antenna light brown, last flagellomere darker; tegula yellowish. Wing veins brown.
Head (Fig. 16a-c) transversally ovate, broadest part below toruli, broader than high and narrower than mesosoma in frontal view, with short dense white setae; 1.3× as broad as high in anterior view and head 2.0× as broad as long in dorsal view. Gena alutaceous to smooth, slightly broadened behind eye, as broad as cross diameter of eye, measuring along transfacial line. Malar space weak alutaceous, without striae radiating from clypeus, malar sulcus absent; eye 2.1× as high as length of malar space. Inner margins of eyes parallel. POL 2.6× as long as OOL, OOL as long as diameter of lateral ocellus and as long as LOL, all ocelli rounded, of same size. Transfacial distance 1.1× as long as height of eye; toruli located above mid height of head, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye slightly longer than diameter of torulus. Lower face almost smooth raised central part below the toruli alutaceous, pubescent. Clypeus impressed, coriaceous, rounded ventrally, medially not incised, anterior tentorial pits distinct; epistomal sulcus and clypeo-pleurostomal line distinctly impressed. Frons uniformly coriaceous, without striae, with few setae, piliferous points visible; interocellar area coriaceous with piliferous points; space between lateral ocellus to compound eye almost smooth and shining. Vertex and occiput coriaceous. Antenna (Fig. 16d) shorter than length of head + mesosoma, with 11 flagellomeres; pedicel 1.5× longer than broad; flagellomeres not broadening until apical end; F1 1.2× as long as F2 and 3.6× as long as pedicel; subsequent flagellomeres progressively shortened; F3 = F4, F5 = F6, F7 to F10 equal in length F11 longer than F10, sometimes F11 with a very inconspicuous suture and thus antenna with 12 flagellomeres. Placodeal sensilla on F5-F11.
Figure 13a-c. Andricus perfectum comb. rev., asexual female. (a) Mesosoma in dorsal view; b) mesoscutellum; c) mesosoma in lateral view.
Mesosoma (Figs. 16e, 17a-c) 1.2× as long as high, concave in lateral view, pubescent. Sides of pronotum alutaceous, with white setae and weak, parallel carinae in upper lateroposterior part, anterolateral rim of pronotum inconspicuous. Mesoscutum 1.2× as broad as long in dorsal view; weak coriaceous, without rugae, pubescent, with piliferous points more visible in the posterior half between notauli. Notaulus complete, deep, straight, converging posteriorly, in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; median mesoscutal line present; parapsidal lines and anterior parallel lines differentiated by alutaceous sculpture. circumscutellar carina absent. Mesoscutellum as long as broad, uniformly rugose; overhanging metanotum, with sparse long setae. Mesoscutellar foveae differentiated, delimited posteriorly by a weak carinae, triangular, deep, with weak alutaceous to smooth bottom, separated by a median carina. Mesopleuron sculptured, coriaceous to weak reticulate with delicate carinae and sparse setae anteriorly, speculum smooth and shining; mesopleural triangle alutaceous, with dense white setae, hiding the surface sculpture; dorsal and lateral axillar areas with delicate parallel longitudinal carinae, glabrous; axillula alutaceous, pubescent; subaxillular bar smooth, glabrous, triangular, posteriorly as high slightly shorter than high of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/2 of its height; upper part of sulcus distinct; lower part of sulcus delimiting big area with dense long white setae. Metascutellum subrectangular, coriaceous with some weak longitudinal carina, curved ventrally; metanotal trough alutaceous, with dense setae; ventral bar of metanotal trough coriaceous dorsally and smooth ventrally; central propodeal area nearly smooth and glabrous; lateral propodeal carinae parallel by converging toward posterior end; lateral parts of propodeum uniformly alutaceous, densely pubescent. Nucha almost smooth and shining dorsally, with parallel sulci laterally. Legs (Fig. 16e) with hind coxa broad, hind femur strongly broadened, with apical lobe; hind tibia with ventral carina on the ventral part extending to half the length of the tibia; base of tarsal claws with strong tooth.
Figure 14a-d. Andricus perfectum comb. rev., asexual female (a-c). a) Fore wing; b) metasoma in lateral view; c) propodeum; d) galls, deposited in the AMNH.
Forewing hyaline, 1.1× as long as body, pubescent, with cilia on margins; radial cell opened, around 2.6× as long as broad; veins light brown; areolet present. Rs+M vein obsolete heading towards the middle of the basal vein.
Metasoma (Fig. 16e) shorter than head+mesosoma, longer than high. 2nd metasomal tergum occupying 3/4 of metasoma, with lateral patch of white setae, without or with obsolete micropunctures, all subsequent terga uniformly and entirely smooth or with obsolete micropunctures. Prominent part of ventral spine of hypopygium needle-like, around 3.5-4.0× as long as broad, with sparse setae laterally, without apical tuft of setae. Body length: 3.0-4.5 mm (Kinsey, 1937b).
Gall (Fig. 17d). Similar to F. repandae but smaller, diameter up to 12 mm.
Taxonomic summary
Type material examined. Holotype: female, deposited in the AMNH “Pabellon, 20W, Ags 7000, Mex; gall 11.26.31, female Spr. ‘32”, “Q. sacame Kinsey Coll.” (white labels), “F. ocri HoloParatype” (red label), “AMNH_IZC 00267206” (white label with a QR code). Paratypes (4 females): 3 females with same data as holotype (AMNH); 1 female, “Sayula, 13S W, Jal 7700’, Mex, gall 12.21.31, female 2.10.32”, “Q. rhodophleb Kinsey Coll.”, “F. ocri Kinsey coll 37” (white labels), “F. ocri paratype, in type box” (red label), (JP-V col).
Figure 15a-d. Femuros bracteatus comb. nov., asexual female (a-b, d). a) Head in frontal view; b) mesosoma in dorsal view; d) habitus; pictures extracted from the entomology collection website of the Smithsonian Institution [https://collections.nmnh.si.edu/search/ento/], pictures taken by Matthew Buffington; c) galls extracted from Weld, 1944.
Additional materialexamined. Three females, deposited in the AMNH “Sayula, 13S W, Jal 7700’, Mex. ꚛ, gall 12.21.31, female 2.10.32”, “Q. rhodophleb Kinsey Coll.”, “F. ocri Kinsey coll 37”; 1 female “Guadalajara, Jal., 25W, 6000’, Mex. ꚛ, 12.23.31”, “Q. haematophlebKinsey coll.”, “F. ocri Kinsey coll 37” white labels. New material deposited in the JP-V col. (UB): 11 asexual females: 3 females “Santa Fe, Delegación de Cuajimalpa, Ciudad de México, México, (MEX181), Ex Q. laeta, (28.ii.2014) iii.2014: 3 females, D. Cibrián-Tovar leg.; 3 females “Fracción de Bosques de Santa Fe, Delegación de Cuajimalpa, Ciudad de México, México, (MEX336), Ex Q. laeta, (10.xii.2015) 15-24.ii.2016: 3 females, D. Cibrián-Tovar”; 1 female “Nativitas, Texcoco, Estado de México, (MEX400), Ex Quercus sp., (13.i.2015) 19.i.2015: 1 female, A. Equihua-Martinez & E. Estrada-Venegas leg.”; 1 female “Nativitas, Texcoco, Estado de México, (MEX401), Ex Quercus sp., (13.i.2015) 19.i.2015: 1 female, A. Equihua-Martinez & E. Estrada-Venegas leg.”; 3 females “Peñon Blanco, 22°10’12.6” N, 102°31’14.8” W, San José de Grácia, Aguascalientes, México, (MEX645), Ex Q. grisea, (10.i.2021) 15.ii.21-20.iii.21: 3 females, R. Clark leg.”, “Femuros ocri Kinsey, 1937, JP-V det. 2022”.
Biology. The asexual generation is only known to induce galls on Q. arizonica Sargent (= Q. sacame Trel.), Q. grisea Liebm, Q. laeta Liebm., Q. magnoliifolia Née (= Q. haemathophlebia Trel.), and Q. rugosa Née (= Q. rhodophebia Trel.) (section Quercus, subsection Leucomexicanae, white oaks). The galls were collected between November and January; adults emerge between January and March (Kinsey, 1937b).
Figure 16a-e. Femuros ocri, asexual female. a) Head in frontal view; b) head in dorsal view; c) mesosoma and head in lateral view; d) antenna; e) lateral habitus, with detail of the ventral spine of the hypopygium.
Distribution. México: Aguascalientes and Jalisco (Kinsey, 1937b).
Diagnosis. Femuros repandae is most closely related to F. bracteatus and both differ from F. ruidum and F. ocri by the POL which is around 1.5× as long as OOL, OOL 1.5× as long as diameter of lateral ocellus, the height of eye is 2.0× as long as the malar space, and the transfacial distance 1.3× as long as the height of the eye. Femuros repandae differs from F. bracteatus by the mesoscutum with the anterior parallel lines and parapsidal lines darkened, chestnut colored; mesoscutellar foveae divided by a thin and sharp carina, while in F. bracteatus the anterior parallel lines and parapsidal lines are of the same color as the rest of the mesoscutum; and the mesoscutellar foveae are divided by a wide rugose elevated area.
Redescription. Asexual female (Figs. 18a-f, 19a-d, 20a-b). Head, mesosoma, metasoma and legs reddish-brown (with some darker areas in anterior parallel lines and parapsidal lines); antenna light brown, last flagellomere darker; tegula yellowish. Wing veins brown.
Head (Fig. 18a-d) transversally ovate, broadest part below toruli, broader than high and narrower than mesosoma in frontal view, with short dense white setae; 1.3× as broad as high in anterior view and head 2.0× as broad as long in dorsal view. Gena alutaceous to smooth, slightly broadened behind eye, as broad as cross diameter of eye, measuring along transfacial line. Malar space weak alutaceous, without striae radiating from clypeus, malar sulcus absent; eye 2.1× as high as length of malar space. Inner margins of eyes parallel. POL 2.6× as long as OOL, OOL as long as diameter of lateral ocellus and as long as LOL, all ocelli rounded, of same size. Transfacial distance 1.1× as long as height of eye; toruli located above mid height of head, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye slightly longer than diameter of torulus. Lower face almost smooth raised central part below the toruli alutaceous, pubescent. Clypeus impressed, coriaceous, rounded ventrally, medially not incised, anterior tentorial pits distinct; epistomal sulcus and clypeo-pleurostomal line distinctly impressed. Frons uniformly coriaceous, without striae, with few setae, piliferous points visible; interocellar area coriaceous with piliferous points; space between lateral ocellus to compound eye almost smooth and shining. Vertex and occiput coriaceous; postocciput and postgena glabrous, alutaceous-reticulate, with concentric lines around occipital foramen and postgenal bridge; posterior tentorial pit large, elongated, area below impressed; occipital foramen as high as height of postgenal bridge; hypostomal carina emarginate, continuing into postgenal sulci which strongly diverge toward occipital foramen, postgenal bridge anteriorly slightly broader than occipital foramen. Antenna (Fig. 18e) shorter than length of head + mesosoma, with 11 flagellomeres; pedicel 1.5× longer than broad; flagellomeres not broadening until apical end; F1 1.2× as long as F2 and 3.6× as long as pedicel; subsequent flagellomeres progressively shortened; F3 = F4, F5 = F6, F7 to F10 equal in length F11 longer than F10, sometimes F11 with a very inconspicuous suture and thus antenna with 12 flagellomeres. Placodeal sensilla on F5-F11.
Figure 17a-d. Femuros ocri, sexual female (a-c). a) Mesosoma in dorsal view; b) mesoscutellum; c) propodeum; d) galls, deposited in the AMNH.
Mesosoma (Figs. 18f, 19a-d) 1.2× as long as high, concave in lateral view, pubescent. Sides of pronotum alutaceous, with white setae and weak and parallel carinae in upper lateroposterior part, anterolateral rim of pronotum inconspicuous. Mesoscutum 1.2× as broad as long in dorsal view; weak coriaceous, without rugae, pubescent, with piliferous points more visible in the posterior half between notauli. Notaulus complete, deep, straight, converging posteriorly, in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; median mesoscutal line present; parapsidal lines and anterior parallel lines differentiated by alutaceous sculpture. Circumscutellar carina absent. Mesoscutellum as long as broad, uniformly rugose; overhanging metanotum, with sparse long setae. Mesoscutellar foveae differentiated, delimited posteriorly by a weak carinae, triangular, deep, with weak alutaceous to smooth bottom, separated by a median carina. Mesopleuron completely sculptured, coriaceous to weak reticulate with delicate carinae and sparse setae anteriorly, speculum smooth and shining; mesopleural triangle alutaceous, with dense white setae, hiding the surface sculpture; dorsal and lateral axillar areas with delicate parallel longitudinal carinae, glabrous; axillula alutaceous, pubescent; subaxillular bar smooth, glabrous, triangular, posteriorly as high slightly shorter than high of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/2 of its height; upper part of sulcus distinct; lower part of sulcus delimiting big area with dense long white setae. Metascutellum subrectangular, coriaceous with some weak longitudinal carina, curved ventrally; metanotal trough alutaceous, with dense setae; ventral bar of metanotal trough coriaceous dorsally and smooth ventrally; central propodeal area nearly smooth and glabrous; lateral propodeal carinae parallel by converging toward posterior end; lateral parts of propodeum uniformly alutaceous, densely pubescent. Nucha almost smooth and shining dorsally, with parallel sulci laterally. Legs with hind coxa broad, hind femur strongly broadened, with apical lobe; hind tibia with ventral carina on the ventral part extending to half the length of the tibia; base of tarsal claws with strong tooth.
Figure 18a-f. Femuros repandae, asexual female. a) Head in frontal view; b) head in posterior view; c) head in dorsal view; d) head in lateral view; e) antenna; f) mesosoma in frontal view.
Forewing (Fig. 20a) hyaline, 1.1× as long as body, pubescent, with cilia on margins; radial cell opened, around 2.6× as long as broad; veins light brown; areolet present. Rs+M vein obsolete heading towards the middle of the basal vein.
Metasoma (Fig. 20b) shorter than head+mesosoma, longer than high. 2nd metasomal tergum occupying 3/4 parts of metasoma length, with lateral patch of white setae, without or with obsolete micropunctures; all subsequent terga uniformly and entirely smooth or with obsolete micropunctures. Prominent part of ventral spine of hypopygium needle-like, around 3.5-4.0× as long as broad, with sparse setae laterally, without apical tuft of setae. Body length: 3.2-4.0 mm (Kinsey, 1937a).
Gall (Fig. 20c). Bud gall, globular to spherical, unilocular, sessile, located on main or side twigs, solitary or occasionally in groups of 2 or 3 galls. It is normally spherical (diameter up to 17 mm), distorted by the bracts, deformed leaves, and petioles, which are fused on the surfaces. Green touched with red when young, silvery and purple-brown when mature (similar to the twig bark). In section, a hard wall is observed that surrounds the gall, a spongy-opened and lax tissue and a large spherical central larval chamber thick and hard walled, up to 7 mm in diameter even in small galls; the spongy tissue usually is darker in mature galls.
Figure 19a-d. Femuros repandae, asexual female. a) Mesosoma in lateral view; b) mesosoma in dorsal view; c) propodeum; d) mesoscutellum.
Taxonomic summary
Type material examined. Holotype asexual female deposited in the AMNH: “Mex. (City), 32E; D.F. 10,600; gall 1.3.32, 13 females, 4.24.32 ”, “Q. repanda, Kinsey Coll.”, red handwriting label “Femuros repandae HoloParatype” “AMNH_IZC 00267207” (white label with a QR code). Paratypes: 15 females with same data as holotype, except for 6 females with different emergence date “4.16.32” (13 females deposited in the AMNH, 2 paratype females deposited I the JP-V col., UB). Type material from F. ruidum belonging to F. repandae. Seven females deposited in the AMNH “Mex. ꚛ, Cuernavaca 8N, Mor. 8700; gall 1.7.32, 1 female, 4.10.32”, “Q. texcocana, Kinsey Coll.”, red handwriting label “Femuros ruidum Paratype”.
Additional material. Material deposited in JP-V col. (UB), 15 asexual females: 1 female “San Juan Coajomulco, 19°45’21.0” N, 99°59’06.4” W, Jocotitlan, Estado de México, México, (MEX254), Ex Q. martinezii, (16.x.2013) 10.vi.2014: 1f, D. García-Martiñón leg.”; 2 females “San Juan Coajomulco, 19°45’21.0” N, 99°59’06.4” W, Jocotitlan, Estado de México, México, (MEX255), Ex Q. rugosa, (22.ix.2013) 30.iii.2014: 1f, D. García-Martiñón leg.”; 4 females “San Juan Coajomulco, 19°45’21.0” N, 99°59’06.4” W, Jocotitlan, Estado de México, México, (MEX256), Ex Q. obtusata,(18.i.2014) 28.i-5.iii.2014: 1f, D. García-Martiñón leg.”; 3 females “La Cumbre, 17°10’01” N, 96°35’58” W, Santa Catarina Ixtepeji, Oaxaca, México, (MEX649), Ex Q. rugosa, (02.ii.2020) 10.vi.2020: 3 females, R. Clark leg.”; 3 females “Santa María Yavesía, 17°16’04.8” N, 96°26’06.5” W, Santa María Yavesía, Oaxaca, México, (MEX650), Ex Q. crassifolia, (20.i.2022) 20.iii-20.iv.2022: 3 females, R. Clark leg”; 2 females “La Congoja, 22°09’34.5” N 102°33’05.2” W, San José de Grácia, Aguascalientes, México, (MEX651), Ex Q. grisea, (12.xii.2021) 15.ii-03.iii.2022: 2 females, R. Clark leg.”.
Figure 20a-c. Femuros repandae, asexual female (a-b). a) Fore wing; b) metasoma in lateral view; c) galls, deposited in the AMNH.
Biology. The asexual generation is only known for inducing galls on Q. deserticola Trel. (= texcocana Trel.), Q. grisea Liebm., Q. martinezii C.H.Muller, Q. obtusata Bonpl., Q. repanda Bonpl., Q. rugosa Née (section Quercus, subsection Leucomexicanae, white oaks), and 1 oak species from section Lobatae, Q. crassifolia Bonpl.. The adult galls were collected between September and February; adults emerge between January and April (Kinsey 1937a).
Diagnosis. Femuros ruidum differs from F. repandae by having the POL 2.0× as long as OOL, the OOL is equal to length of lateral ocellus, the malar space 0.4× as long as height of the eye; the transfacial distance subequal to the height of the eye. Differs from F. ocri by the densely pubescent and distinctly punctured mesopleuron; the speculum pubescent and punctured except for a small smooth central area; propodeal carinae bent outwards.
Redescription. Asexual female (Figs. 21a-f, 22a-d, 23a-b). Head, mesosoma, metasoma and legs reddish-brown (without darker areas); antenna light brown, last flagellomere darker; tegula yellowish. Wing veins brown.
Figure 21a-f. Femuros ruidum, asexual female. a) Head in frontal view; b) head in posterior view; c) head in dorsal view; d) mesosoma in frontal view; e) head in lateral view; f) antenna.
Head (Fig. 21a-e) transversally oval, broadest part below toruli, broader than high and narrower than mesosoma in frontal view, with short dense white setae; 1.3× as broad as high in anterior view and head 2.4× as broad as long in dorsal view. Gena alutaceous, slightly broadened behind eye, as broad as cross diameter of eye, measuring along transfacial line. Malar space weak alutaceous to smooth, without striae radiating from clypeus, malar sulcus absent; eye 2.1× as high as length of malar space. Inner margins of eyes parallel. POL 1.6× as long as OOL, OOL 2.6× as long as diameter of lateral ocellus and longer than LOL, all ocelli rounded, of same size. Transfacial distance 1.2× as long as height of eye; toruli located above mid height of head, frons shorter than height of lower face, diameter of antennal torulus 2.0× as long as distance between them, distance between torulus and eye slightly longer than diameter of torulus. Lower face alutaceous to weak coriaceous, raised central part below the toruli coriaceous, pubescent. Clypeus impressed, coriaceous, rounded ventrally, medially not incised, anterior tentorial pits distinct; epistomal sulcus and clypeo-pleurostomal line distinctly impressed. Frons uniformly coriaceous, without striae, with few setae, piliferous points visible; interocellar area coriaceous with piliferous points; space between lateral ocellus to compound eye almost smooth and shining. Vertex and occiput coriaceous. Antenna (Fig. 21f) shorter than length of head + mesosoma, with 11 flagellomeres; pedicel 1.5× longer than broad; flagellomeres from F5 broadening until apical end; F1 1.4× as long as F2 and 4.1× as long as pedicel; subsequent flagellomeres progressively shortened; F11 longer than F10, sometimes F11 with a very inconspicuous suture and thus antenna with 12 flagellomeres. Placodeal sensilla on F5-F11.
Figure 22a-d. Femuros ruidum, asexual female. a) Mesosoma in lateral view; b) mesosoma in dorsal view; c) propodeum; d) mesoscutellum.
Mesosoma (Figs. 21d, 22a-d) 1.2× as long as high, pubescent, concave in lateral view. Sides of pronotum alutaceous, with white setae and inconspicuous carinae in upper lateroposterior part, anterolateral rim of pronotum inconspicuous. Mesoscutum 1.1× as broad as long in dorsal view; weak coriaceous, without rugae, pubescent, with piliferous points more visible in the posterior half between notauli. Notaulus complete, deep, straight, converging posteriorly, in most posterior part distance between notauli shorter than distance between notaulus and side of mesoscutum; median mesoscutal line absent; parapsidal lines and anterior parallel lines differentiated by alutaceous sculpture, almost smooth. Circumscutellar carina absent. Mesoscutellum slightly longer than broad, uniformly rugose; overhanging metanotum, with sparse long setae. Mesoscutellar foveae differentiated, triangular, relatively deep, not defined posteriorly with alutaceous bottom, separated by a median carina. Mesopleuron alutaceous to smooth, pubescent with piliferous points, speculum smooth and shining; mesopleural triangle alutaceous, with dense white setae, hiding the surface sculpture; dorsal and lateral axillar areas with delicate parallel longitudinal carinae, glabrous; axillula alutaceous, pubescent; subaxillular bar smooth, glabrous, triangular, posteriorly as high slightly shorter than high of metanotal trough; metapleural sulcus reaching mesopleuron in upper 1/2 of its height; upper part of sulcus distinct; lower part of sulcus delimiting big area with dense long white setae. Metascutellum subrectangular, coriaceous with some weak longitudinal carina, curved ventrally; metanotal trough alutaceous, with dense setae; ventral bar of metanotal trough coriaceous; central propodeal area nearly alutaceous with some rugae and glabrous; lateral propodeal carinae parallel by converging toward posterior end; lateral parts of propodeum uniformly alutaceous to smooth, densely pubescent. Nucha almost smooth and shining dorsally, with parallel sulci laterally. Legs (Fig. 23b) with hind coxa broad, hind femur strongly broadened, with apical lobe; hind tibia with ventral carina on the ventral part extending to 4/5 the length of the tibia; base of tarsal claws with strong tooth.
Forewing (Fig. 23a) hyaline, 1.1× as long as body, pubescent, with cilia on margins; radial cell opened, around 2.6× as long as broad; veins light brown; areolet present. Rs+M vein obsolete heading towards the middle of the basal vein.
Metasoma (Fig. 23b) shorter than head+mesosoma, slightly longer than high. Metasomal tergite 2 occupying 2/3 parts of the metasoma, with lateral patch of white setae, without micropunctures or micropunctures obsolete, all subsequent tergites uniformly and entirely smooth or with obsolete micropunctures. Prominent part of ventral spine of hypopygium needle-like, around 5.5× as long as broad, with sparse setae laterally, without apical tuft of setae. Body length: 3.0-4.3 mm (Kinsey, 1937a).
Gall (Fig. 23c). Similar to F. repandae but larger, diameter up to 23 mm.
Figure 23a-c. Femuros ruidum, asexual female (a-b). a) Fore wing; b) metasoma in lateral view; c) galls, deposited in the AMNH.
Taxonomic summary
Type material of F. ruidum. Holotype asexual female: “Mex. ꚛ, Cuernavaca 8N, Mor. 8700; gall 1.7.32, 8 females, 3.5.32”, “Q. texcocana, Kinsey Coll.”, red handwriting label “Femuros ruidum HoloParatype” (deposited in AMNH: AMNH_IZC 00267208). Paratypes: 3 females and 3 galls “Mex. ꚛ, Cuernavaca 8N, Mor. 8700; gall 1.7.32, 1 female, 4.10.32”, “Q. texcocana, Kinsey Coll.”, red handwriting label “Femuros ruidum Paratype” (1 female in AMNH; 1 female in JP-V col; 1 female and galls in HMNH); 3 females “Mex (City) 12W, D.F. 8500’, Mex. ꚛ, gall 1.13.32, 10 females, 4.10.32”, “Q. texcocana, Kinsey Coll.”, “Femuros ruidum Kinsey coll.” (1 female in JP-V col; 2 females in HMNH). Type material of F. repandae belonging to F. ruidum. Paratypes: 1 female “Mex. (City), 32E; D.F. 10600; gall 1.3.32, 9 females, 4.16.32”, “Q. repanda, Kinsey Coll.”, red handwriting label “Femuros repandae Paratype” (deposited in JP-V col).
Figure 24a-e. Lateral habitus of holotypes. a) Andricus calvoi sp. nov.; b) Andricus geniale comb. rev.; c) Andricus integrum comb. rev.; d) Andricus lusum comb. rev.; e) Andricus perfectum comb. rev.
Additional material. Seven asexual females deposited in the JP-V col. (UB) with the label “Universidad de la Sierra Juárez, 17°18’54” N, 96°28’58” W, Ixtlán de Juárez, Oaxaca, México, (MEX646-648), Ex Q. glaucoides, R. Clark leg.” and different collection and emergence dates: 1 female “(10.iii.2018) 27.iv.2018”; 3 females “(08.xii.2020) 10.i- 20.ii.2021”; and 3 females “(10.xii.2018) 10.i.2019”.
Biology. The asexual generation is only known to induce galls on Q. deserticola (= texcocana Trell.) and Q. rugosa Née (= conglomerata Trel. = rhodophlebia Trel.), according to Kinsey (1937a); and Q. glaucoides Mart. & Gal. (section Quercus, subsection Leucomexicanae, white oaks) in this study. The galls were collected between December and March, and the adults emerge between January and April.
Distribution. México: México City, Morelos, and Oaxaca (Kinsey, 1937a).
Figure 25a-d. Lateral habitus of holotypes. a) Femuros bracteatus comb. nov.; b) Femuros ocri; c) Femuros repandae; d) Femuros ruidum.
Discussion
After carefully studying the type materials of the Femuros species described by Kinsey (1937a, b), we have observed distinct morphologies. The species F. repandae, F. ruidum, and F. ocri have a smooth lower face of the head, while the rest of the species (A. geniale, A. integrum, A. lusum, and A. perfectum) have a sculptured lower face with carinae radiating from the clypeus. This latter group of species has been provisionally transferred to the Andricus genus. Andricus is a polyphyletic genus, and although several species have recently been transferred to other genera (such as Striatoandricus, Disholandricus, Druon, Dros, Feron, Protobalandricus, and Trichoteras), there remains a significant number of species groups within Andricus that require further assessment. The morphological characteristics of the species currently included in Andricus display excessive variation, making it difficult to determine reliable diagnostic characters. The species transferred into Andricus in this study exhibit the radiating carinae from the clypeus, which is one of the few characters broadly used to define Andricus.
The Cynipini genera are known for exhibiting a high level of homoplasy (Liljeblad et al., 2008), making it challenging to identify exclusive diagnostic characters for most genera. Recognition of these genera relies on a combination of characters rather than any single defining feature, further complicating the task of generic delimitation. Although, the morphological distinctiveness of Femuros compared to its closest genera is evident (see diagnosis). The etymology of Femuros refers to the broadening of the hind femur on the ventral margin near the distal end, although this feature is not exclusive to Femuros (see diagnosis). Additionally, the presence of an internal carina on the hind tibia, previously considered exclusive to Femuros, loses its exclusivity after transferring some species to the genus Andricus. This character is also found in the unrelated genus Odontocynips. Regardless of its distinctiveness, little is known about the relationships between Femuros and the rest of the Cynipini genera. It is suspected to be closely related to Andricus, probably to the species transferred in this study. If that is the case, the distinctiveness of Femuros should be assessed with the support of molecular data and would also be dependent on the taxonomy of Andricus and how this macro genus is assessed.
The galls of Femuros are now exclusively round bud galls with small bracts partially covering their surface with radiating lamellae connecting the larval chamber and the lignified outer shell. There is a certain degree of variation in terms of the gall size between the species; F. ruidum usually induces the biggest galls (up to 23 mm in diameter), but the size distribution overlaps with the galls of F. repandae and F. ocri. Hence, the galls of Femuros are almost impossible to tell apart without the confirmation of the adult wasp.
The galls of the studied Andricus speciesare mostly barrel-shaped or cup-shaped bud galls, with an apical depression, forming a central cavity that can be widely open to the exterior or almost closed, with thick lignified parenchyma. The apical walls of the outer shell of the gall can be opened forming a distinct cup or cover the apical part of the central cavity. The larval chamber is closer to the base of the gall.
The host plant associated with Femuros and the newly transferred Andricus are mainly oak species from section Quercus, subsection Leucomexicanae, except for a single record of F. repandae galling on Q. crassifolia (section Lobatae). Species galling on 2 different sections are extremely rare and in many cases such records have been corrected. This last record on Q. crassfolia may be attributed to an accidental host swap since F. repandae galls mainly in section Quercus. Further samplings should confirm this record and the occurrence of it if ever repeated.
Acknowledgments
VCP was supported by the University of Barcelona, grant number APIF 2018-2019. We thank all the staff and colleagues of the AMNH during VCP’s visit for their help and support: James Carpenter, Christine Lebeau, Melody Doering, Morgan H Chase, Marcel Hermes, Nayeli Gutiérrez, and Royce Cumming. We are also thankful to Matt Buffington (Smithosnian Institution, USA) and Mar Ferrer-Suay for their pictures of the type materials. This research was funded by “PID2021-128146NB-I00/MCIN/AEI/10.13039/501100011033/” and “FEDER una manera de hacer Europa”.
References
Cuesta-Porta, V., Cooke-McEwen, C., Melika, G., Romero-Rangel, S., Equihua-Martínez, A., Estrada-Venegas, E. G. et al. (2024). Review of the Mexican species of Dishol- caspis Dalla Torre & Kieffer with the description of eleven new species. Zootaxa, 5498, 1–100. https://doi.org/10.11646/zootaxa.5498.1.1
Cuesta-Porta, V., Melika, G., Nicholls, J. A., Stone, G. N., & Pujade-Villar, J. (2022). Re-establishment of the Nearctic oak cynipid gall wasp genus Druon Kinsey, 1937 (Hymenoptera: Cynipidae: Cynipini), with description of 5 new species. Zootaxa, 5132, 1–92. https://doi.org/10.11646/zootaxa.5132.1.1
Cuesta-Porta, V., Melika, G., Nicholls, J. A., Stone, G. N., & Pujade-Villar, J. (2023). Re-establishment of the Nearctic oak cynipid gall wasp genus Feron Kinsey, 1937 (Hymenoptera: Cynipidae: Cynipini), including the description of six new species. Zootaxa, 5366, 1–174. https://doi.org/10.11646/zootaxa.5366.1
Denk, T., Grimm, G. W., Manos, P. S., Deng, M., & Hipp, A. L. (2017). An updated infrageneric classification of the oaks: review of previous taxonomic schemes and synthesis of evolutionary patterns. In E. Gil-Pelegrín, J. Peguero-Pina, & D. Sancho-Knapik (Eds.), Oaks physiological ecology. Exploring the functional diversity of genus Quercus L. (pp. 13–38). Cham Switzerland: Tree Physiology. Springer Nature.
Harris, R. (1979). A glossary of surface sculpturing. Occasional Papers in Entomology, 28, 1–31.
Kinsey, A. C. (1937a). New Mexican gall wasps (Hymenoptera, Cynipidae). Revista de Entomologia, 71, 39–79.
Kinsey, A. C. (1937b). New Mexican gall wasps (Hymenoptera, Cynipidae) II. Revista de Entomologia, 74, 428–471.
Liljeblad, J., Ronquist, F., Nieves-Aldrey, J. L., Fontal-Cazalla, F., Ros-Farre, P., Gaitros, D. et al. (2008). A fully web-illustrated morphological phylogenetic study of relationships among oak gall wasps and their closest relatives (Hymenoptera: Cynipidae). Zootaxa, 1796, 1–73. https://doi.org/10.11646/zootaxa.1796.1.1
Melika, G. (2006). Gall wasps of Ukraine. Vols. 1 & 2. Vestnik Zoologii, 21 (Suppl.), 1–644.
Melika, G., & Abrahamson, W. (2002). Review of the world genera of oak cynipid wasps (Hymenoptera: Cynipidae, Cynipini). In G. Melika, & C. Thuróczy (Eds.), Parasitic wasps: evolution, systematics, biodiversity and biological control (pp. 150–190). Budapest: Agroinform.
Melika, G., Nicholls, J. A., Abrahamson, W. G., Buss, E. A., & Stone, G. N. (2021). New species of Nearctic oak gall wasps (Hymenoptera: Cynipidae, Cynipini). Zootaxa, 5084, 1–131. https://doi.org/10.11646/zootaxa.5084.1.1
Melika, G., Pujade-Villar, J., Abe, Y., Tang, C., Nicholls, J., Wachi, N. et al. (2010). Palaearctic oak gallwasps galling oaks (Quercus) in the section Cerris: re-appraisal of generic limits, with descriptions of new genera and species (Hymenoptera: Cynipidae: Cynipini). Zootaxa, 2470, 1–79. https://doi.org/10.11646/zootaxa.2470.1.1
Menitsky, L. L. (1984). Oaks of Asia. St. Petersburg: Leningosizdat Sciences.
Nixon, K. C. (1993). Infrageneric classification of Quercus (Fagaceae) and typification of sectional names. Annals of Forest Science, 50 (Suppl.), 25s–34s. https://doi.org/10.1051/forest:19930701
Pujade-Villar, J., & Ferrer-Suay, M. (2015). Adjudicació genérica d’espècies mexicanes d’ubicació dubtosa descrites per Kinsey i comentaris sobre la fauna mexicana (Hymenop- tera: Cynipidae: Cynipini). Butlletí de la Institució Catalana d’Història Natural, 79, 7–14.
Pujade-Villar, J., Lobato-Vila, I., & Ferrer-Suay, M. (2017). Restablecimiento del género Dros Kinsey (Hymenoptera: Cynipidae: Cynipini) como género válido para especies americanas. Entomología Mexicana, 4, 752–758.
Pujade-Villar, J., & Melika, G. (2014). Re-establishment of Erythres Kinsey, 1937 as a valid genus of gallwasps from Mexico (Hymenoptera: Cynipidae: Cynipini). Dugesiana, 21, 155–160. https://doi.org/10.32870/dugesiana.v21i2.4155
Ronquist, F., & Nordlander, G. (1989). Skeletal morphology of an archaic cynipoid, Ibalia rufipes (Hymenoptera: Ibaliidae). Entomologica Scandinavica. Supplementum, 33, 1–60.
Weld, L. H. (1952). Cynipoidea (Hym.) 1905-1950 being a Supplement lo the Dalla Torre and Kieffer monograph, the Cynipidae in Das Tierreich, Leifening 24, 1910 and bringing the systematic literature of the world up to date, including keys to families and subfamilies and list of new generic, specific and variety names. Ann. Arbor, Michigan: Privately printed.
José Ángel Zamarripa-Fernández a, b, *, Ismael Hinojosa-Díaz c, Moisés Armando Luis-Martínez d, Ricardo Ayala a
a Universidad Nacional Autónoma de México, Instituto de Biología, Estación de Biología Chamela (Sede Colima), Km 59, Carr. Fed. 200, Melaque – Pto. Vallarta, La Huerta, 48895 Chamela, Jalisco, Mexico
b Universidad Nacional Autónoma de México, Posgrado en Ciencias Biológicas, Edificio D, 1°Piso, Circuito de Posgrados, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
c Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, 3er. Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
d Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Biología Evolutiva, Museo de Zoología “Alonso L. Herrera”, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
A taxonomic revision of the cleptoparasitic bee genus Mesoplia Lepeletier, 1841 from Mexico and Central America is presented. We recognize 4 new species, resulting in a total of 10 species for the study area: Mesoplia (Mesoplia) insignis (Smith, 1879), M. (M.) sapphirina Melo & Rocha-Filho, 2011, M. (M.) regalis (Smith, 1854), M. (M.) dugesi (Cockerell, 1917), M. (M.) rufipes (Perty, 1933), M. (M.) veracruzana sp. nov., M. (M.) oaxacana sp. nov., M. (M.) tica sp. nov., M.(Eumelissa) ornata (Spinola, 1841)and M.(E.) carmelitae sp. nov. We propose the following synonymies: M. imperatrix (Friese, 1913) = M. (M.) insignis, and M. guatemalensis (Cockerell, 1912) = M. (M.) rufipes. We describe the previously unknown female of M. (M.) dugesi and provide diagnoses for both sexes based on the holotype. An identification key for the species, illustrations, distributional records (available as supplementary material), and lists of hosts and floral associations are provided.
Revisión taxonómica y distribución de las abejas cleptoparásitas del género Mesoplia (Apidae: Ericrocidini) de México y Centroamérica
Resumen
Se presenta una revisión taxonómica del género de abejas cleptoparásitas Mesoplia Lepeletier de México y Centroamérica. Se reconocen 4 especies nuevas, dando un total de 10 especies para el área en estudio: Mesoplia (Mesoplia) insignis (Smith, 1879), M. (M.) sapphirina Melo & Rocha-Filho, 2011, M. (M.) regalis (Smith, 1854), M. (M.) dugesi (Cockerell, 1917), M. (M.) rufipes (Perty, 1933), M. (M.) veracruzana sp. nov., M. (M.) oaxacana sp. nov., M. (M.) tica sp. nov., M. (Eumelissa) ornata (Spinola, 1841) y M. (E.) carmelitae sp. nov. Se encontraron las siguientes sinonimias: M. imperatrix (Friese, 1913) = M. (M.) insignis y M. guatemalensis (Cockerell, 1912) = M. (M.) rufipes. Se describe la hembra de M. (M.) dugesi, previamente desconocida, y se proporcionan las diagnosis para ambos sexos basados en el tipo. Se aportan una clave de identificación de las especies, ilustraciones, registros de distribución (disponible como material complementario), una lista de huéspedes y asociaciones florales.
The tribe Ericrocidini is composed mainly of cleptoparasitic bees that have Neotropical affinity, with the exception of the genus Ericrocis Cresson, 1887, which is Nearctic (Martins et al., 2018). The greatest diversity occurs in the Amazon region (Silveira et al., 2002; Snelling & Brooks, 1985). Their known hosts are species of the tribe Centridini (Rocha-Filho et al., 2009). The most easily recognizable morphological character of the tribe is the large, bifurcate or multidentate apex of the middle tibial spur (Michener, 2007).
The genus Mesoplia Lepeletier, 1841 is the richest in the tribe with 19 described species (Table 1), although it is estimated that it could include more species (Rozen et al., 2011). It is distributed from northern Mexico (Sinaloa and Tamaulipas) to Argentina (Snelling & Brooks, 1985). Mesoplia is recognized by the following characters: internal orbits more or less parallel; presence of 2 tubercles on the scutellum of variable development; meso and metabasitarsus without lateral patches of short, flattened setae; tiny metallic scales blue or green, mainly in the dorsal area of the metasoma, frequently also with patches of yellowish or whitish pubescence; S5 of male evidently emarginated and hidden under S4 (Genaro & Breto, 2022; Michener, 2007; Snelling & Brooks, 1985; Vivallo, 2010).
The subgeneric classification was proposed by Snelling and Brooks (1985). They proposed 2 subgenera based on morphological characters, Mesoplia and Eumelissa. Males of the former have posterior tibial spurs that do not extend beyond the basal third of the basitarsus, the posterior tibia has an internal distal patch of setae and S5 evidently emarginate and hidden under S4. Females of the same subgenus have a broad pygidial plate, which occupies the exposed dorsal surface of T6, covered with setae or scales and dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces. In contrast, males of the subgenus Eumelissa have long posterior tibial spurs reaching to or beyond middle of basitarsus. Females of the same subgenus have narrow pygidial plate, does not completely occupy the exposed dorsal surface of T6, without scales or setae and dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces (Michener, 2007). Furthermore, Moure (1960a, b) recognizes 2 groups: bifrons and azurea, both of which are part of the subgenus Mesoplia s. str.Males of the bifrons grouphave conspicuous hind tibia setal brush, 2 spurs on the posterior tibia, ventral basal surface of posterior femur without a robust posterior projection. Females have pronotum without a carina extending laterally from the pronotal collar to the pronotal lobe.On the other hand, males of the azurea grouphave inconspicuous hind tibia setal brush, 1 spur on the posterior tibia and ventral basal surface of posterior femur with a robust posterior projection. Females of this group have pronotum with a carina extending laterally from the pronotal collar to the pronotal lobe.
The known hosts include several species of Centridini (Rocha-Filho et al., 2009). For Mesoplia, as in the rest of the tribe, the biology of most species is practically unknown and their geographical distribution uncertain, since some of the information available in the literature is based on mistaken identifications (Vivallo, 2014).
In this work we update the knowledge of the species of Mesoplia forMexico and Central America, providing diagnoses, descriptions, distributional data, identification keys, and a summary of floral and host records. We describe 4 new species and establish 2 new synonyms.
Materials and methods
We reviewed 363 specimens from the following collections where studied specimens are deposited (names of curators and/or collection managers are in parentheses): EBCh, Estación de Biología Chamela, IBUNAM, Jalisco, Mexico (R. Ayala Barajas); CNIN, Colección Nacional de Insectos, Instituto de Biología, UNAM, Mexico City (I.A. Hinojosa Díaz); MZFC, Museo de Zoología “Alfonso L. Herrera”, Facultad de Ciencias, UNAM, Mexico City (M.A. Luis Martínez); CZUG, Centro de Estudios en Zoología, CUCBA, Universidad de Guadalajara, UDG, Jalisco, Mexico (José Luis Navarrete, Hugo Fierros); HFL, Colección Particular de Hugo Eduardo Fierros López, Jalisco, Mexico (Hugo Fierros); IEXA, Colección Entomológica del Instituto de Ecología, A.C., INECOL, Veracruz, Mexico (Jorge E. Valenzuela, Viridiana Vega); ECOAB, Colección de Abejas de El Colegio de la Frontera Sur, Chiapas, Mexico (Rémy Vandame, Jorge A. Mérida Rivas, Philippe Sagot); CER-UADY, Colección Regional Entomológica de la Universidad Autónoma de Yucatán, UADY, Yucatán, Mexico, (Virginia Meléndez Ramírez, María José López Gómez); SEMC, Snow Entomological Collection, Division of Entomology, University of Kansas Natural History Museum, KU, Lawrence, Kansas, USA (Zachary Falin, Michael Engel, Rachel Osborn); MNCR, Museo Nacional de Costa Rica, Heredia, Costa Rica (Maricelle Méndez, Marcela Sánchez); MZUCR, Museo de Zoología, Universidad de Costa Rica, San José, Costa Rica (Mauricio Fernández, Paul Hanson); CINAT, Centro de Investigaciones Apícolas Tropicales, Universidad Nacional de Costa Rica, UNA, Heredia, Costa Rica (Mario Gallardo, Eduardo Herrera).
Table 1
Recognized species of Mesoplia Lepeletier, 1841. * New synonyms.
Dominica, Guadeloupe, Martinique and Saint Vincent
Mesoplia chalybaea (Friese, 1912)
Both
Brazil
Mesoplia dugesi (Cockerell, 1917)
Both
Mexico
Mesoplia guatemalensis (Cockerell, 1912)*
–
–
Mesoplia oaxacana sp. nov.
Female
Mexico
Mesoplia rufipes (Perty, 1833)
Both
Mexico to South America
Eumelissa
Mesoplia alboguttata (Ducke, 1905)
Both
Bolivia and Brazil
Mesoplia carmelitae sp. nov.
Both
México and Costa Rica
Mesoplia decorata (Smith, 1854)
Both
Brazil, Ecuador, Guatemala, Panama and Venezuela
Mesoplia friesei (Ducke, 1902)
Both
Brazil, Colombia and Paraguay
Mesoplia guedesii (Ducke, 1902)
Female
Brazil
Mesoplia ornata (Spinola, 1841)
Both
Mexico to French Guiana
Type specimens of M. sapphirina were studied. For the rest of the species high-resolution images of holotypes were available, which allowed verification of the species that were delimited and that had been previously described: for M. insignis (both sexes)and M. regalis on the website of the Natural History Museum (London, England); the holotype of M. dugesi on the website of the National Museum of Natural History, Smithsonian Institution (Washington D.C., USA); for M. imperatrix images of holotype from the Museum für Naturkunde (Berlin, Germany) were provided by Stefanie Krause; and the holotype images of M. guatemalensis from the American Museum of Natural History (New York, USA) were provided by Corey Smith. Delimitation of other species followed original descriptions and specimens previously identified by taxonomists who worked with the genus and compared their specimens with types deposited in the mentioned collections. Due to the morphological characteristics of the species of this genus, it is possible to be certain of their identification.
Morphological terminology follows Michener (2007); for integument sculpture we follow Harris (1979). Metasomal terga and sterna are designated as T and S respectively, numbered from anterior to posterior. We used F for antennal flagellum, with F1 following the pedicel. Male genitalia were dissected along with S7 and S8. The subgeneric classification follows Snelling and Brooks (1985) and the classification of groups follows Moure (1960a). All measurements are given in millimeters (mm). Images for most of the structures and habitus were taken with a Leica MC 170 HD camera adapted to a Leica M205C stereoscopic microscope except some of the habitus photographs were taken with a Cannon EOS 70D DSLR camera and a Canon EF-S 60 mm macro lens. For Mesoplia tica sp. nov., images were taken using an Olympus DP74 camera attached to an Olympus SZX16 stereoscopic microscope. Images were processed with Adobe Photoshop CS5®, Photopea.com, HeliconSoft and Leica Application Suite. The scale bar indicates 0.5 mm in the genitalia images. Distribution maps were produced from specimen locality data (available as supplementary material) using QGis software and the base map of Esri Physical and were prepared by the first author. Geospatial coordinates are presented in DMS. Plant associations were based on the information present in the specimen label. For the cleptoparasite-host associations we reviewed and compiled the published information categorized following Rocha-Filho et al. (2009).
Description
We recognize 10 species of Mesoplia for Mesoamerica; 4 are new to science herein described and illustrated. In addition, 2 synonyms are recognized, Mesoplia imperatrix (Friese, 1913) = M. insignis (Smith, 1854) and M. guatemalensis (Cockerell, 1912) = M. rufipes (Perty, 1833).
Genus Mesoplia Lepeletier, 1841
Mesoplia Lepeletier, 1841: 457. Type species: Mesocheira azurea Lepeletier & Serville, 1825: Museo Regionale di Scienze Naturale, Torino, Italy (♀♂), monobasic.
Melissa Smith, 1854: 279. Type species: Mesocheira azurea Lepeletier & Serville, 1825, designated by Sandhouse, 1943: 570.
Diagnosis. Maxillary palps elongated, with 3 or 4 segments; internal orbits more or less parallel; scutellum bituberculate (in some species not very evident); medial basitarsus with a flattened distal process on the posterior margin, more evident in females (Figs. 1, 2); dorsal region of the metasoma covered with tiny scales that give it a metallic blue or green color, regularly with small patches of yellow or white pubescence; S5 of the male evidently emarginate and hidden under S4; body length from 10 to 25 mm.
Subgenus Mesoplia Lepeletier, 1841
Diagnosis. Male: distal inner surface of the posterior tibia with a setal brush, not very evident in some species (Fig. 3A-E); hind tibial spurs not reaching beyond basal third of basitarsus, some species with only 1 spur present. Female: pygidial plate broad, occupying the exposed dorsal surface of T6; pygidial plate covered with scales or setae (Fig. 4A-G); dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces.
M. bifrons group(sensu Moure, 1960a, b)
Diagnosis. Male: hind tibia with setal brush conspicuous (Fig. 3A-C) and 2 spurs; posterior femur with ventral basal margin without a robust posterior projection; posterior basitarsus with inner surface without a longitudinal carina. Female: without a carina between pronotal collar and the pronotal lobe (Fig. 5A).
Mesoplia (Mesoplia) insignis (Smith, 1879)
(Figs. 3A, 4A, 5A, 6, 7A, 8-10, 35)
Melissa insignis Smith, 1879: 108. Syntypes: British Museum Natural History, London, England (♀♂). Type locality: Mexico, Veracruz, Orizaba.
Melissa imperialis Friese, 1912: 205. Holotype: Museum für Naturkunde, Berlin, Germany (♀). Type locality: Mexico, Morelos. New junior synonym.
Melissa imperatrix Friese, 1913: 89. nom. nov. for Melissa imperialis Friese, 1912. New junior synonym.
Melissa imperatrix Friese, 1921: 266. nom. nov. for Melissa imperialis Friese, 1912. New junior synonym.
Diagnosis. Male: metallic greenish body, with pubescence on head and mesosoma mostly bright yellow; integument of antenna, pronotal lobe, tegula and legs black; scutellar tubercles conical, inconspicuous due to dense pubescence (Fig. 6D), with perceptible cleft between them; robust forked medial tibial spur, with dark integument, wide anterior projection and with between 2 and 4 internal teeth; pubescence of the terga mainly whitish with a spot of bright yellow pubescence on both lateral margins of T1, at least also present in T2 (Fig. 6A); surface of T7 without scales but with black pubescence; apex of T7 slightly forked, both apices truncated and bare at tip (Fig. 7A); S4 shallowly emarginate, with several rows of compound pubescence on the apical margin, dark in color in the center and becoming light at the laterally; digitiform lobe of the gonostylus triangular and broad, projecting upwards (Fig. 8C, D).
Figure 1. Middle leg basitarsus, male: A, Mesoplia (M.) sapphirina. B, Mesoplia (M.) dugesi. C, Mesoplia (M.) rufipes. D, Mesoplia (E.) carmelitae sp. nov. E, Mesoplia (E.) ornata.
Female: body dark metallic blue and black pubescence, but with some small patches of whitish pubescence behind the antennal area, preoccipital area, lateral margins of the scutum to the level of tegula, and lateral margins of S2-S4 (Fig. 9A-C); black integument on antenna pronotal lobe, tegula and legs; scape with dark metallic blue; integument of scutum and tegula with scales that give a dark metallic blue appearance; scutellar tubercles mamilliform; tergal pubescence mainly dark; ventral surface of mesosoma and metasoma with metallic blue scales; middle tibial spur robust with black integument, forked, anterior projection with 1 to 4 internal teeth; pygidial plate with a broad and quadrangular apex, without scales on the surface, with blackish pubescence that does not reach the apex, black integument (Fig. 4A).
Variation. Some females from Morelos with white setae on lateral edges of the labrum and posterolateral edges of the propodeum. Pubescence and the presence of metallic scales in some body areas of males seem to be relatively variable, labral setae can be yellow, whitish or dark; the clypeus may have yellow setae or metallic blue scales, as well as the dorsal surface of the scutellar tubercles.
Material examined. Mexico: Jalisco: 22.5 km al NE de Magdalena, 20°58’46.17” N, 104°6’52.33” W, 1,066.8 m, 19-VII-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1254912); Tala, Bosque Escuela, 20°35’44.9988” N, 103°38’15” W, 1,499 m, 7-IX-2012, H. E. Fierros-López, 1♀ (CZUG); ANP Bosque la Primavera, S. Nopales, 20°36’3.4” N, 103°38’20.4” W, 1,500 m, 26-IX-2013, O. Serrano-Pineda, 1♀ (CZUG); Tequila, Volcán de Tequila, 20°51’52” N, 103°50’36” W, 1,300 m, 29-VII-1995, H. E. Fierros-López, 1♀ (CZUG 580); 20°49’36” N, 103°50’45” W, 1,300 m, 29-VIII-1995, H. E. Fierros-López, 1♀ 1♂ (HFL), 20°51’36” N, 103°51’0” W, 1,370 m, 12-VI-2022, K. Sanchez, 1♂ (ECOAB 87968); Tlajomulco de Zúñiga, Fraccionamiento Nueva Galicia coto Valencia, 20°34’39” N, 103°26’40” W, 1,618 m, 24-IX-2022, J. A. Zamarripa-Fernández, C. Sánchez-Bastidas e I. I. Alfaro-Ríos, 5♀ (CZUG); Zapopan, Barranca la Experiencia, 20°57’42.804” N, 103°24’5.292” W, 1,300 m, 14-IX-1996, G. Mercado, 1♀ (CZUG 1112), 1♂ (CZUG 1113), 22-VIII-2009, G. Mercado, 1♀ (CZUG EXP0440), 1♂ (CZUG EXP0446), 5-IX-2009, G. Mercado, 1♂ (CZUG EXP0445), 1,350 m, 29-VII-2009, G. Mercado, 1♂ (HFL), 29-VII-2009, G. Mercado, 1♂ (CZUG); Parque ecológico Huilotán, 20°57’42.804” N, 103°24’5.292” W, 1,121 m, 7-IX-2023, J. A. Zamarripa-Fernández, C. Morales & J. M. Sandoval, 3♀ 4♂ (CZUG). Morelos: Jojutla de Juárez, Cerro del Higuerón, 18°34’0.3876” N, 99°9’55.2816” W, 1,060 m, 26-VII-1981, C. Deloya, 1♀ (MZFC 28612); Yautepec de Zaragoza, 18°50’55.6116” N, 99°4’42.0888” W, 1,249 m, 15-XI-1980, J. M. Labougle, 4♀ (CNIN 3042-3045) 2♂ (CNIN 3046; 3047). Oaxaca: Santo Domingo Tehuantepec, 17 km W of Tehuantepec, 16°21’59.328” N, 95°22’59.088” W, 8-IX-1965, D. Janzen, 2♂ (SEMC 1254910; 1254911).
Distribution. This species is endemic to Mexico and distributed from the center of country to the north of the Isthmus of Tehuantepec (Fig. 10) between 1,060-1,616 m of altitude, a distribution that corresponds to the Trans-Mexican Volcanic Belt and Sierra Madre del Sur.
Figure 3. Hind leg, male: A, Mesoplia (M.) insignis. B, Mesoplia (M.) tica sp. nov. C, Mesoplia (M.) sapphirina. D, Mesoplia (M.) dugesi. E, Mesoplia (M.) rufipes. F, Mesoplia (E.) carmelitae sp. nov. G. Mesoplia (E.) ornata.
Taxonomic comments. This species is morphologically similar to Mesoplia tica sp. nov. Males differ in the apex of T7 slightly forked with both apices truncated and bare at tip (Fig. 7A), and the pubescence of the mesosoma almost entirely bright yellow (Fig. 6). The female pygidial plate has a broadly quadrangular apex, with blackish pubescence on the surface (Fig. 4A), and the scutellar tubercles are less projected upward than in Mesoplia tica sp. nov.
Diagnosis. Male: metallic greenish body, with bright yellow pubescence on the clypeus, anterior surface of the mesepisternum, pronotal lobe, anterior region of the scutellar disc, posterior region of the scutellum, scutellum and lateral areas of the propodeum (Fig. 11); tegula with black setae on inner anterior and posterior edge; scutellar tubercles conical and strongly projecting upwards, with a perceptible cleft between them; robust forked medial tibial spur, with blackish integument, wide anterior projection and with between 3 and 4 internal teeth; tergal pubescence mainly whitish, with a spot of bright yellowpubescence on both lateral margins of T1 (Fig. 11B); tergal scales metallic greenish-copper; apex of T7 noticeably forked, both apices rounded (Fig. 7B); dorsal lobe of gonostyle triangular and broad projected upwards (Fig. 12C, D).
Female: body dark metallic and blue pubescence, with scales blue-greenish on head and mesosoma, and metallic blue to the metasoma; with patches of whitish pubescence on labrum, anterior lateral edges of the clypeus, behind the antennal insertion, preoccipital region, upper region of the gena, lateral surface of the mesepisternum, lateral margins of the scutum at the level of the tegula, posterior region of the scutellum, lateral apex of T5 and lateral apices of S2-S4 (Fig. 13); black integument on antenna, pronotal lobe, tegula and legs; scape with blue-green scales; tegula with metallic blue scales; scutellar tubercles subconical, strongly projecting upwards (Figs. 13A, D, 35C); middle tibial spur robust with black integument, forked, anterior projection with 2 to 3 internal teeth; pygidial plate with apex more or less rounded, with scales on the surface that do not reach the apex, and black integument (Fig. 4B).
Figure 6. Mesoplia (M.) insignis, male. A, Lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: male. Body length 17.5 mm, forewing length 13.4 mm, head length 4.2 mm, intertegular distance 4.3 mm, T2 length 2.5 mm. Head: mandible with an internal tooth. Labrum subrectangular, concave surface, with a very evident preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles conical, strongly projecting upwards, cleft between them evident. Vertical metanotum and propodeum. Middle tibial spine rounded and short. Medial tibial spur robust forked, anteriorly projecting with 3 to 4 internal teeth. Middle basitarsus with posterior margin smooth and with short and straight flattened distal process. Hind tibia with 2 short spurs at the inner apex. Metasoma: T7 with a flattened surface, noticeably forked apex, with a moderately deep central cleft, medium distance between the apices, both ends rounded. Color: in general, with black integument. Head: labrum, rostrum, scape, pedicel and f1 black, remainder of flagellum greyish. Mandible with apical third yellowish brown, the rest black. Mesosoma: pronotal lobe, tegula, wing veins, legs, middle tibial spur, black. With smoked wings. Forewings without dark spots evident. Metasoma: with black sternum. Surface sculpture: head, labrum with most of surface rugose, by basal edge polished. Mesosoma: tegula with stippling. The propodeum strigulate. Pubescence: black and bright yellow. Head: labrum central surface with white plumose setae and scattered simple black setae, lower edge with a pair of long black tufts, lateral edges with simple black setae, upper edge bare. Clypeus densely covered with bright yellow plumose setae. Supraclypeal area, forehead close to the lower edge of the ocelli and vertex with metallic bluish-green scales. Supraclypeal area with black setae. With a spot of bright yellow setae in the upper part of the paraocular area. Preoccipital region and upper gena region with bright yellow setae. Lower region of the gena with black setae. Mesosoma: anterior surface of the mesepisternum, pronotal lobe, region anterior to the central disc of the scutum, posterior region of the scutellum, metascutum and lateral areas of the propodeum covered almost exclusively by bright yellow setae. The rest covered by black setae. Central disc of the scutum and most of the surface of the tegula with metallic green scales. Both the anterior and posterior inner edges of the tegula with black setae. Naked propodeal triangle. Internal apex of posterior tibia with very evident short blackish setal brush (Fig. 3B). Metasoma: terga covered with metallic greenish-copper scales. Anterior edge of T1 with scattered simple black and white setae, with a spot of yellow pubescence on both lateral margins. Surfaces of T3 to T6 with several series of rows of simple black setae. S2 to S4 with a short white patch of pubescence on the lateral edges. Scattered black sternum setae. S4 with long, branched black setae near the posterior edge and with several rows of simple black setae on the surface. S2 to S4 with a more or less triangular white spot on the lateral margins; dorsal lobe of gonostyle triangular and broad projected upwards.
Allotype: female. Body length 16.8 mm, forewing length 12.6 mm, head length 4 mm, intertegular distance 4.5 mm, T2 length 2.3 mm. Head: mandible with an internal tooth; labrum subrectangular with flat surface, and with a very evident preapical margin; ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles with the conical shape strongly projecting upwards, an evident cleft between them. Vertical metanotum and propodeum. Middle tibial spine pointed and short. Medial tibial spur robust forked, anteriorly projecting with 2 to 3 internal teeth. Medium basitarsus with slightly wavy posterior margin and a flattened curved distal process. Metasoma: pygidial plate with apex more or less rounded. Color: integument black. Head: labrum, rostrum, scape, pedicel and F1 black, remainder of flagellum greyish. Apical third of mandible dark reddish brown with nearby yellowish brown spot, basal half black. Mesosoma: pronotal lobe, tegula, wing veins, legs, middle tibial spur, black. Apex of distal process of middle basitarsus dark reddish brown. Evenly smoked wings. Forewings without obvious dark spots. Metasoma: black sternum. Surface sculpture: head. Rugulous labrum. Mesosoma: finely punctuate tegula. Dotted propodeum. Metasoma: dotted pigidial plate. Pubescence: mainly black, but head covered mainly with white setae. Head: labrum surface with white setae branched and few scattered simple black setae, with a pair of tufts of black setae on the anterior margin and with setae on the lateral edges. Scape almost entire surface of anterior area with metallic blue-green scales. Mesosoma: covered mainly with black setae, except for patches of white setae on the collar of the pronotum, lateral region of the mesepisternum, lateral edges of the scutum, near the tegula, posterior region of the scutellum. Scales of the central disc of the scutum and the pronotal lobe metallic blue-green, of the same tone as those of the face. Scales of the tegula, mesepisternum and scutellum metallic blue. Metasoma: virtually the entire surface covered with metallic blue scales and the setae present are almost exclusively black, except for a spot of branched white setae on the lateral edges of T1, several rows of simple white pubescence on the surface of T4 and T5, a triangular spot of short branched white pubescence on the lateral ends of S3 to S5 and another small spot on the lateral edge of T5. Sterna also with metallic blue scales. Pygidial plate with metallic scales on the surface.
Variation. Some males have bright yellow setae on the external surface of anterior tibia and little pale-yellow pubescence on the lateral edge of S4.
Figure 8. Mesoplia (M.) insignis, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Taxonomic summary
Type material. Holotype: ♂ MNCR INB0004333459, CR. CART (Costa Rica, Cartago), Ochomongo, San Nicolás, Finca Kirqua, 9°54’41.29” N, 83°56’1.81” W, 14-II-2010/1-III-2010, W. Porras (MNCR). Allotype: ♀ MNCR INB0004333299, CR. HER [Costa Rica, Heredia], Sarapiquí, La Isla, 10°24’50.79” N, 84°7’42.24” W, 100-200 m, 22-II-2017/17-IV-2017, I. A. Chacón (MNCR). Paratypes: same data as the holotype ♀ MNCR INB0004333460 (MNCR), ♀ MNCR A5002767; CR. ALAJ [Costa Rica, Alajuela], Virgen del Socorro, 10°17’0.20” N, 84°10’38.22” W, 750 m, 15-IX-1982, I. A. Chacón (MNCR); ♂ MNCR A5057872 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, Estación Biológica La Selva, 10°25’50.61” N, 84°10’38.21” W, 61 m, 24-VI-1988, I. A. Chacón (MNCR), ♀ SEMC 1254906, 10°25’35.81” N, 84°1’1.16” W, 80 m, 19-V-1993, J. Ashe & A. Ashe (SEMC); ♀ MZUCR HY6823 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, La Isla, 10°14’7.70” N, 83°54’43.45” W, 221 m, VI-2011/VII-2011, I. A. Chacón (MZUCR); ♀ CINAT JZ344 CR. HER [Costa Rica, Heredia], Sarapiquí, Puerto Viejo, La Tirimbina, 10°23’52.20” N, 84°8’9.16” W, 186 m, 1997, Sergio (CINAT), ♀ MNCR JZ357, same location as the previous one, 25-VI-1997, Posada (MNCR); ♀ MZUCR HY6827 CR. PUNT [Costa Rica, Puntarenas], Reserva Forestal Golfo Dulce, 24 km W. Piedras Blancas, 8°45’17.43” N, 83°29’5.51” W, 200 m, IV-1992/V-1992, P. Hanson (CINAT).
Etymology. The specific epithet tica refers to the demonym with which the inhabitants of Costa Rica recognize themselves.
Distribution. Registered only for Costa Rica and it is possible that this species be endemic to this country (Fig. 10), present between 61 to 750 m altitude.
Taxonomic comments. This species is morphologically like M. insignis, the main differences are that the female has the scutellar tubercles strongly projecting upwards (Fig. 6A), and the pygidial plate covered by metallic scales and its apex is rounded (Fig. 4B); and the male has the apex of T7 noticeably forked, both apices rounded (Fig. 7B).
Figure 9. Mesoplia (M.) insignis, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Mesoplia (Mesoplia) regalis (Smith, 1854)
Melissa regalis Smith, 1854: 280. Syntypes: British Museum Natural History, London, England (♀♂). Type locality: Brazil, Pará, Santarém.
Diagnosis. Male: body metallic green and pubescence white and brown; reddish brown on antennal integument, pronotal lobe, tegula and legs; scutum and tegula with metallic green scales; scutellar tubercles with a flattened surface, median slit between them; bifurcated middle tibial spur, with integument brown; anterior projection with 2 to 4 internal teeth; anterior edge of T1 with scattered white setae, lateral edges with a brown and white spot; T7 with the flat tips, with setae below it that are noticeable in dorsal view.
Figure 10. Map with the known distribution of Mesoplia (Eumelissa) carmelitae sp. nov., M. (E.) ornata, M. (Mesoplia) dugesi, M. (M.) insignis, M. (M.) oaxacana sp. nov., M. (M.) regalis, M. (M.) tica nov. sp., and M. (M.) veracruzana sp. nov.
Female: body metallic green, with white pubescence; antennal integument, pronotal lobe, tegula and legs reddish brown; scutum and tegulas with scales metallic green; scutellar tubercles more developed than in M. sapphirina with a deep depression between them and tubercles conspicuously pointed the delimiting carina is more developed only along the tubercles gradually fading to the sides; bifurcated middle tibia spur, integument brown; anterior projection with 2 to 4 internal teeth; pygidial plate with rounded apex and metallic scales that do not reach the apex (Melo & Rocha-Filho, 2011).
Taxonomic summary
Material examined. 1♂: Panama: Islas de las Perlas, Isla del Rey, 8°23’34.70” N, 78°54’24.11” W, 139 m, 13-IV-1981, R. W. Brooks, 1♂ (SEMC 1254921).
Distribution. Mainly South American, with the northern limit in Panama (Moure & Melo, 2023).
Taxonomic comments. According with Melo and Rocha-Filho (2011), this is a species similar morphologically to M. sapphirina, the main difference is that in this species the tubercles of the scutellum have the distal edge projecting upwards, more evident in females; the dorsal surface of the T7 in males bulging, with flat apices and with setae inserted underneath, distinguishable even in dorsal view. Specimens from Brazil deposited in SEMC were analyzed, whose distribution data were not included because they are from a distribution area not included in this study.
Mesoplia sapphirina Melo & Rocha-Filho, 2011: 30. Holotype: Department of Zoology, Universidade Federal do Pará, Curitiba, Brazil (♀). Type locality: Panamá, ‘Old Panama’.
Figure 11. Mesoplia (M.) tica sp. nov. Holotype, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Diagnosis. Male: body metallic blue, with whitish setae (Fig. 14); F1 light reddish brown; remainder of flagellum, pronotal lobe, tegula, most of legs and sterna dark reddish brown; scutum and tegula with metallic blue scales; scutellar tubercles poorly developed, posterior carina conspicuous; middle tibial spur forked, both apical branches with robust teeth, outer branch with 2 small internal teeth (Fig. 16A); dorsal surface of T7 bulging, its projected ventral portion conspicuously excavated; with a pair of apical projections of T7 with rounded apices, emargination between them relatively deep (Fig. 7C); dorsal and digitiform lobes of the gonostylus well developed (Fig. 15C, D).
Female: body metallic blue, with whitish setae (Fig. 17); F1 light reddish brown; remainder of flagellum, pronotal lobe, tegula, most of legs and sterna dark reddish brown; scutellar tubercles weakly developed, its surface only slightly convex and its tubercles low and broadly rounded; the carina that delimits the tubercles posteriorly well developed and runs continuously from one side of the scutellum to the other; middle tibial spur forked, both apical branches with robust teeth, outer branch with 2 small internal teeth (Fig. 18A); pygidial plate with oval apex and covered with decumbent bluish metallic scaly hairs, except for the apex which is naked (Fig. 4C).
Figure 12. Mesoplia (M.) tica sp. nov. Holotype male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Variation. Some males have yellow setae on the posterior surface of the scutellar tubercles.
Taxonomic summary
Material examined. Costa Rica: Cartago: Cachí, 9°49’52.06” N, 83°48’18.12” W, 1,016 m, 28-V-1974, G. Rivera, 1♀ (MZUCR HY6608). Guanacaste: Bagaces: Barrio el Brasil, 10°32’57.36” N, 85°15’54.11” W, 114 m, 5-V-2022, K. Borbón, 1♀ (CINAT JZ340), 3♂ (CINAT JZ341 – CINAT JZ343). 10°32’58.60” N, 85°15’52.42” W, 110 m, 8-IV-2023, K. Borbón, 1♀ (MZUCR HY6131), 1♂ (MZUCR HY6131). Filadelfia, 10°26’51.27” N, 85°32’59.86” W, 22 m, 28-IX-1959, M. Leiva, 1♂ (MZUCR HY6816); La Cruz: Cuajiniquil, Bahía Tomás, Salinitas, in mangrove, 10°55’15.60” N, 85°42’57.96” W, 0 m, 28-I-2022, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2249), 7-II-2022/7-III-2022, M. M. Chavarría & P. Hanson, 2♀ (MZUCR HY5067), (MZUCR HY5068), 25-III-2021, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2345), V-2022, M. M. Chavarría & P. Hanson, 1♀ (MZUCR HY6838); Nicoya: Sámara, 9°52’51.60” N, 85°31’36.30” W, 17-I-2009, C. Reichie, 1♀ (MZUCR HY5051). San José: San Pedro de Montes de Oca, 9°56’11.41” N, 84°2’58.50” W, 1,201 m, 20-VI-1959, Y. Gómez, 1♀ (MZUCR HY5061). Mexico: Chiapas: La Concordia, 16°7’12.00” N, 92°39’0.00” W, 616 m, 1-IV-2016, P. Sagot, 1♀ (ECOAB 58710), 16°3’36.00” N, 92°33’36.00” W, 550 m, 5-III-2020, P. Sagot, 1♂ (ECOAB 134687); Mazatan, 14°55’12.00” N, 92°37’48.00” W, 6 m, 2-II-2019, P. Sagot, 1♂ (ECOAB 130647), 14°55’48.00” N, 92°37’12.00” W, 8 m, 2-II-2019, I. Pérez, 1♂ (ECOAB 130665); Tonalá, 15°30’0.00” N, 92°24’0.00” W, 3 m, 21-IV-2011, 1♀ (ECOAB 56616). Jalisco: La Huerta: Chamela, 19°29’54.77” N, 105°2’41.22” W, 10-IX-1981, S. Bullock, 1♀ (EBCH SB736), 19-X-1985, S. Bullock, 1♀ (SEMC 1451101), 27-IX-1985, R. Ayala, 1♀ (SEMC 1451080); Estación Biológica Chamela, 19°29’54.77” N, 105°2’41.22” W, 25-IX- 1985, C. D. Michener, 1♀ (EBCH JZ65); La Manzanilla, 19°17’0.03” N, 104°47’13.06” W, 1 m, 16-XI-1994, A. Moreno, 1♀ (HFL 302), 1♀ (CZUG 302), 16-XI-1994, H. E. Fierros-López, 1♀ (HFL JZ7), 1♂ (HFL JZ8), 1♀ (CZUG 309); Playa Careyitos, 19°29’36.18” N, 105°1’29.56” W, 27-X-1985, R. Ayala, 1♀ (EBCH RA219), 27-IX-1985, R. Ayala, 1♀ (EBCH RA218), 30-IX-1985, C. D. Michener, 1♀ (SEMC 1451078); Playa Careyitos 6 Km. South of Chamela, 19°29’36.18” N, 105°1’29.56” W, 27-IX-1985, A. Rodríguez P., 1♀ (EBCH JZ63), R. B. Roberts, 1♀ (EBCH JZ64); Playa Teopa, 19°23’48.69” N, 105°1’9.27” W, 4-X-1985, J. G. Rozen 1♀ (EBCH JZ62), R. Ayala, 1♀ (EBCH JZ69). Oaxaca: San Pedro Huamelula, 15°55’48.00” N, 95°46’48.00” W, 22 m, 21-XII-2019, P. Sagot, 1♂ (ECOAB 133378). Sinaloa: San Ignacio, Autopista 4 Río-Piaxtla, 23°55’59.28” N, 104°25’59.88” W, 1,184 m, 18-III-1990, W. Bell, D. Conlon & R. L. Mickley, 1♀ (SEMC 1451099). Tabasco: Tenosique, 17°18’0.00” N, 91°10’12.00” W, 126 m, 23-V-2018, A. Tapia, 1♀ (ECOAB 120153). Tamaulipas: Altamira, Playa Altamira, 22°29’59.53” N, 97°51’41.04” W, 3-VI-1985, M. S. Wasbauer & J. E. Slansky, 1♀ (SEMC 1442315); Llera de Canales, 23°19’24.79” N, 99°1’19.66” W, 267 m, 19-VII-1954, Univ. Of Kans. Mex. Exped., 1♂ (SEMC 1451076); Soto la Marina, La Pesca, 23°46’31.53” N, 97°44’21.63” W, 4 m, 1-VII-1981, B. Miller, B. Porter & L. Stage, 6♀ (SEMC 1451070 – SEMC1451075); Tampico, 22°13’57.47” N, 97°53’30.53” W, 2 m, 10-VI-1951, H.E. Evans, 6♀ (SEMC 1451089 – SEMC 1451094), 1♂ (SEMC 1451088). Veracruz: without coordinates, 25-VII- 1995, 1♀ (CERUADY 239ver). 25-VII-1995, 1♂ (CERUADY 230ver); Actopan: Cofre de Perote, 19°35’45.96” N, 96°22’47.28” W, 10 m, VI-2019, P. Luna & W. Dáttilo, 1♀ (IEXA JZ38); La Mancha, 19°21’12.60” N, 96°13’27.48” W, 3 m, VI-2022, B. Ratoni & W. Dáttilo, 1♂ (EBCH 622 1), 19°21’16.92” N, 96°13’29.64” W, 15 m, VI-2022, B. Ratoni & W. Dáttilo, 1♀ (EBCH 623 7), 1♂ (EBCH 623 6), V-2023, C. Pinilla, 2♂ (EBCH JZ355, EBCH JZ356). Boca del Río: 8 km SE of Boca del Río, 19°3’13.58” N, 96°2’42.53” W, 9 m, 22-VII-1990, W. Bell, D. Conlon & R. L. Mickley, 1♂ (SEMC 1451100); San Andrés Tuxtla: Estación biológica Los Tuxtlas, 18°31’0.00” N, 95°7’59.59” W, 136 m, 16-IX-2018, I. Huerta, 1♀ (CZUG JZ 13); Tecolutla: 20°28’47.00” N, 97°0’36.00” W, H. E. Evans, 1♀ (SEMC 1451095), 2.41402 km N of Tecolutla, 20°29’33.25” N, 97°1’34.21” W, 4.572 m, 12-VI-1985, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451077). Yucatán: Tizimín: El Cuyo, 21°31’5.99” N, 87°40’59.00” W, 25-VII-1995, H. Moo, 2♀ (CERUADY 009406(02450)), (009411(02477)), 1♂ (009414(02451)), R. Tut, 7♀ (CERUADY 009574(02453), (009546(02454)), (009543(02455)), (009539(02456)), (009541(02474)), (009545(02475)), (009536(02476)), 1♂ (009544(02461)), 14-VI-1995, R. Tut, 8♀ (CERUADY 007998(02452)), (007987(02462)), (007992(02457)), (007986(02465)), (007980(02466)), (007982(02467)), (008004(02469)), (008000(02473)), 5♂ (007996(02458)), (007979(02460)), (007994(02463)), (007983(02472)), (008006(02479)), 24-V-1995, R. Tut, 1♀ (006649(02468)), 1♂ (006651(02478)), H. Moo, 1♂ (006844(02464)), 4-IV-1995, R. Tut, 1♀ (005166(02470)), 1♂ (005175(02459)). Nicaragua: León: Campus Agropecuário UNAN-León, 12°25’22.14” N, 86°57’8.68” W, 16-V-2022, R. Varela, 1♀ (LEECAV JZ292). Panama: Chimán: Brujas: Punta Bruja, Zona del Canal, 8°35’6.51” N, 78°31’56.69” W, 4 m, 25-I-1929, C. H. Curran, 1♂ (SEMC 1451102). Panamá: Panamá Viejo, 9°0’22.09” N, 79°29’6.29” W, 5 m, 16-IV-1945, C. D. Michener, 1♂ (SEMC 1451079). Panamá Oeste: Fort Kobbe, 8°55’0.30” N, 79°34’59.59” W, 8 m, 11-I-1960, W. J. Hanson, 2♀ (SEMC 14510972), (SEMC 1451098), 2♂ (SEMC 1451096), (SEMC 1427034).
Figure 13. Mesoplia (M.) tica sp. nov. Allotype, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Distribution. The northernmost distribution has been recorded in Sonora, along the Pacific Ocean slope and in Tamaulipas on the Gulf of Mexico slope, to the south the limit of its distribution it is in the center of Panama (Fig. 19).
Taxonomic comments. It is a species morphologically similar to M. regalis, the main difference is that the apices of the scutellar tubercles are rather flattened (Fig. 16A, D), not projected upwards (this character can be confused if the scales in this area of the body have been lost), while males do not present setae that protrude in dorsal view below T7 and the apices of this are rounded (Fig. 7C).
Diagnosis. Female: body metallic blue, with pubescence mostly blackish, but bright yellow setae on head and mesosoma, especially on the pronotum, mesepisternum, posterior region of the scutellum, metanotum and lateral areas of the propodeum (Fig. 20); integument black in antenna, pronotal lobe, tegula and legs; posterior apex of scutellar tubercles projecting upwards (Fig. 20C); middle tibial spur forked, with black integument, the anterior projection with 3 inner teeth; tergal pubescence mainly dark with a bright yellow spot on both lateral margins of T1 and T2 (Fig. 20A); pygidial plate with rounded apex, but with a notch in the medial part of the apex, with scales that does not reach the apex, and with black integument (Fig. 4D).
Figure 14. Mesoplia (M.) sapphirina, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: female. Body length 17.9 mm, forewing length 12.6 mm, head length 5 mm, intertegular distance 4.2 mm, T2 length 2.5 mm. Head: mandible with an internal tooth. Labrum subtriangular, surface slightly concave, with a raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: scutum convex in lateral view. Scutellar tubercles not very evident due to the large amount of pubescence, a noticeable cleft between them. Metanotum and propodeum vertical. Spine of the middle tibia long pointed. Medial tibial spur robust and forked, anterior projection with 3 internal teeth. Medial basitarsus with smooth posterior margin and a flattened distal process that is slightly curved externally. Metasoma: broad pygidial plate at the base, apex subtruncate with a slight central notch. Color: integument blackish. Head: black on face, labrum and antenna; the flagellum with integument opaque. Apical third of mandible yellowish brown, except for black teeth. Mesosoma: almost all black. Apex of distal process of middle basitarsus reddish brown. Wings overall evenly smoked (Fig. 20A, B); forewings with the anterior edge of the marginal cell darker. Veins dark. Metasoma: between the terga and sterna reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Reticulated propodeum. Metasoma: dotted pigidial plate. Pubescence: mostly bright yellow and black. Head: labrum discal area with white plumose setae and scattered simple black setae, lower edge with a pair of long dark tufts, lateral edges with simple black setae, upper edge bare. Clypeus with lower margin with white plumose setae, sides with pale yellow plumose setae, central surface with light metallic blue scales. Suplaclypeal area with few long and scattered black simple setae and with scales equal to those of the clypeus. Frons with black setae and light metallic green scales near the lower edge of the ocelli. Paraocular area with pale yellow plumose setae. Vertex and preoccipital region with bright yellowcompound setae. Gena with setae bright yellow, except for the lower region that has black. Mesosoma: pronotal lobe with the anterior and external lateral margin with bright yellow setae, posterior margin with interspersed white and black setae, inner margin with mainly black setae, surface with dark metallic blue scales (Fig. 20). Mesepisternum upper region with bright yellow setae, lower region black. Scutum covered mainly by black setae, except for the lateral edges near the posterior two-thirds of the tegula that have interspersed bright yellow setae, central disc with light metallic green scales. Tegula black on the anterior and inner edge and with dark metallic blue scales on the surface. Upper of scutellum with few black setae and light metallic blue scales and bright yellow on posterior region, that projects a little between the tubercles. On the metanotum bright yellow. Propodeum upper region naked, posterolateral region bright yellow. Legs are mainly dark and with dark metallic blue scales on the external surfaces, with a tuft of white setae on the posterior margin of the femurs of the forelegs and on the external apices of the middle femurs. Metasoma: terga covered with metallic bluish-green scales. Anterior edge of T1 with scattered white setae, lateral edges with a bright yellow spot; T2 with a smaller pale-yellow spot in the lateral marginal area. T4 and T5 with few white simple setae in lateral view. T6 with thick black setae on the sides of the pygidial plate. Pygidial plate with light metallic green scales that do not reach the apex. Sterna with scattered black setae. S2 to S4 with a sub triangular white spot on the lateral margins.
Figure 15. Mesoplia (M.) sapphirina, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Male: unknown.
Taxonomic summary
Type material. Holotype: ♀ IEXA JZ39 MEX. VER [Mexico, Veracruz], Coatepec, 19°26’42.36” N, 96°57’52.13” W, 1,200 m, 6-VI-2020, G. Quintos-Andrade (IEXA). Paratypes: 3♀: UNAM-LT JZ352 MEX. VER [Mexico, Veracruz], San Andrés Tuxtla, Estación Biológica Los Tuxtlas, 18°35’5.50” N, 95°4’26.58” W, 160 m, 1985-VII-6, A. Ibarra (UNAM-LT). Same location as the previous one, 2♀ UNAM-LT JZ353, 1985-VI-2, P. Sinica. UNAM-LT JZ354, 1986-VII-20, P. Sinica.
Etymology. The name veracruzana refers to the known distribution of this species, the state of Veracruz, Mexico.
Distribution. Only known for the localities of Coatepec and San Andrés Tuxtla (Fig. 10), although there are photographic records in iNaturalist near Xalapa and Orizaba, localities with cloud forest and tropical forest.
Taxonomic comments. The female of this species has a pubescent color similar to that in the male of M. insignis, so they could be confused. In this species, the bright yellow pubescence on the metasoma is only located in the pronotum, mesepisternum, posterior region of the scutellum, metanotum and lateral areas of the propodeum (Fig. 20), unlike M. insignis whose metasoma is almost completely covered by yellow setae (Fig. 6).
M. azurea group(sensu Moure, 1960a, 1960b)
Diagnosis. Male: hind tibia with an inconspicuous setal brush and with 1 spur; ventral basal surface of posterior femur with a robust posterior projection (Fig. 3D, E); inner surface of basitarsus with a longitudinal carina. Female: with carina extending laterally from the pronotal collar, to the pronotal lobe (Fig. 5B).
Mesoplia (Mesoplia) dugesi (Cockerell, 1917)
(Figs. 1B, 3D, 4E, 5B, 7D, 10, 16B, 21-23, 35E)
Mesonychium dugesi Cockerell, 1917: 477. Holotype: National Museum of Natural History, Smithsonian Institution, Washington DC, United States of America (♂). Type locality: Mexico, Guanajuato.
Diagnosis. Male: body dark metallic blue, pubescence mainly black (Fig. 21), except for some white spots on the central surface of the labrum, anterior edge of the superior gena and clypeus; integument blackish on antenna, pronotal lobe, tegula and legs, except for the F1 which is usually dark reddish brown; tegula with metallic blue scales; wing with black veins; scutellar tubercles practically flattened and without a cleft between them; middle tibial spur forked with dark reddish-brown integument, anterior projection with 1 to 3 internal teeth (Fig. 16B); femur posterior with a robust posterior projection on the ventral basal surface (Fig. 3D); apex of T7 bifurcated, both with an acute angle and close to each other (Fig. 7D); dorsal lobe of gonostylus reduced (Fig. 22C, D).
Female: body dark metallic blue, with dark pubescence at first glance (Fig. 23), in a deeper examination some areas with whitish pubescence can be seen on labrum, superior area of the gena, internal margin of the pronotal lobe and a small spot on the posterior surface of the scutellum and between the tubercles; integument of antenna, pronotal lobe, tegula and legs blackish (dark reddish); labrum, vertex, preoccipital area and tegula with metallic blue scales; with a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5B); wings with black veins (Fig. 23A, B); scutellar tubercles conical, slightly projecting with a shallow cleft between them (Fig. 23D); middle tibial spur forked with dark reddish-brown integument, anterior projection with 3 internal teeth; T1 with black setae; terga with metallic blue scales; pygidial plate with a narrow apex and completely covered by metallic bluish-green scales (Fig. 4E).
Description. Female: body length, 16.4 mm, forewing length 12.7 mm, head length 4.1 mm, intertegular distance 3.9 mm, T2 length 1.7 mm. Head: mandible with an internal tooth. Labrum subrectangular, surface slightly concave, with a raised preapical margin. Ocelli aligned with the upper margin of the eyes, protruding slightly above the head in lateral view. Mesosoma: with carina that runs laterally from the collar of the pronotum to the pronotal lobe. Convex scutum in lateral view. Scutellar tubercles rounded, without posterior carina, with a median slit between them. Metanotum and propodeum vertical. Medial tibial spine short rounded. Medial tibial spur bifurcated, anterior projection with 3 internal teeth. Medial basitarsus with pentadentate external margin and with a flattened distal process that is slightly curved externally. Metasoma: broad pygidial plate at the base, narrow rounded apex. Color: integument blackish to dark reddish brown. Head: labrum with reddish brown lateral and anterior edges, blackish surface. Apical region of mandible blackish (yellowish brown) reddish brown in basal region. Scape and pedicel blackish to reddish brown. F1 reddish brown, remainder of flagellum brown. Mesosoma: pronotal lobe and tegula dark reddish brown. Almost transparent wings. Forewings with the anterior edge and the distal apex of the marginal cell darker, also presenting a darker spot that goes from the external edge of the 2r-m to the wing apex (Fig. 23B). Dark veins. Inner surfaces of legs dark reddish brown, outer surfaces blackish. Apex of the distal process of the medial basitarsus ferruginous and medial tibial spur reddish brown. Metasoma: blackish sterna, except for S6 which is reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Strigulate propodeum. Metasoma: dotted pigidial plate. Pubescence: mostly blackish. Head: labrum central surface with white feathery setae and scattered simple black setae, lower edge with a pair of long dark tufts, lateral edges with simple black setae, upper edge bare. Clypeus and supraclypeal area with metallic blue scales and long black simple setae. Scape with short black setae. Frons with black setae and metallic blue scales on the lower edge of the ocelli. Paraocular area mainly with simple black setae, with a tuft of setae near the antennal alveolus. Vertex and preoccipital region with simple black setae and metallic blue scales. Gena with black setae, except for the middle region near the eyes which has white setae. Mesosoma: with black setae except for the inner margin of the pronotal lobe and a small spot on the posterior surface of the scutellum and between the tubercles. Pronotal lobe with metallic blue scales. Anterior region of the scutum with dense setae coverage. Central disc with scattered setae and light metallic blue scales. Tegula with black setae on the anterior edge and metallic blue scales on the inner edge. Scutellum upper region with few black setae and metallic blue scales, posterior region also with metallic blue scales. Naked propodeal triangle. Legs are mainly black, with only a patch of white setae on the external apex of the anterior tibia and on the external apex of the middle femur. External surface of the tibiae of all legs with metallic blue scales and on the posterior basitarsus. Metasoma: with black setae except for the lateral margins of S2 to S4 which have a triangular spot of short white setae. Sterna with scattered black setae.
Variation. Some males have a greater proportion of white setae in some body regions, on all margins of the pronotal lobe, in the anterior region of the scutum, anterior and posterior apex of the tegula, between the tubercles and in the posterior region of the scutellum, metanotum and on the supralateral margins of the propodeum.
Taxonomic summary
Material exanimated. Mexico: Jalisco: Mazamitla, Puerta del Zapatero, 19°50’29.00” N, 103°5’52.00” W, 1,615 m, 27-VIII-2005, H. E. Fierros-López, 1♂ (HFL); Teocuitatlán: Laguna de Sayula, 19°56’55.00” N, 103°27’56.00” W, 1,350 m, 28-VIII-2011, A. Aguilar, 1♂ (HFL 1111); Zapopan: Barranca la Experiencia, 20°44’6.13” N, 103°19’38.38” W, 1,200 m, 23-VIII-1997, G. Mercado, 1♀ (HFL 1107), 23-VIII-1996, G. Mercado, 1♂ (HFL), 14-IX-1996, G. Mercado, 1♂ (CZUG 1108), 1,350 m, 27-IX-2009, G. Mercado, 1♀ (CZUG). Michoacán: Nahuatzen: Zacapu, 19°45’22.46” N, 101°46’52.61” W, 2,152 m, 10-IX-2014, G. Mérida, 1♂ (ECOAB 25661).
Distribution. Only known from Jalisco, Guanajuato and Michoacán in México, although there is at least one record in iNaturalist for Querétaro (Fig. 35E). Its distribution seems to be restricted to the center of the country (Fig. 10).
Taxonomic comments. Is the darkest species distributed in the region, both the general pubescence of the body and the integument of the legs are mainly blackish.
Mesoplia guatemalensis Cockerell, 1912: 567. Holotype: National Museum of Natural History, Smithsonian Institution, Washington D.C., United States of America (♀). Type locality: Guatemala, Gualan. New junior synonym.
Diagnosis. Male: body metallic blue-greenish coloration and with whitish pubescence (Fig. 24); integument of the scape, pedicel and f1 ferruginous (in some specimens inner margin of the scape darker); integument ferruginous on apical flagellomere in posterior view, pronotal lobe and tegula; integument of legs dark brown, on external surface and ferruginous on internal; tegula without scales (Fig. 24B); wing veins ferruginous; with rounded scutellar tubercles, with posterior carina and no cleft between them; femur posterior with a robust posterior projection on the ventral basal surface (Fig. 3E, 35F); terga with blue-green scales; apex of T7 bifurcated, both with an acute angle and close to each other (Fig. 7E); dorsal lobe of gonostylus reduced (Fig. 25C, D).
Female: body metallic blue-greenish coloration and with whitish pubescence (Fig. 26); integument ferruginous on scape, pedicel and f1, pronotal lobe, tegula and legs (some populations with dark brown integument); with a carina extending laterally from the pronotal collar to the pronotal lobe; tegula without metallic scales (Fig. 26B); wing veins ferruginous; conical tubercles slightly projected with a shallow cleft between them; terga with dorsal pubescence whitish; middle tibial spur bifurcated with ferruginous integument, anterior projection with 2 or 3 internal teeth (Fig. 18B); pubescence of the anterior edge of T1 whitish; terga with green scales; pygidial plate with a narrow apex and covered by metallic bluish-green scales (Fig. 4F).
Notes made by Moure (1960a) from the holotype: “The pubescence of the clypeus and paraocular area, especially in the upper part, tends to very pale yellow. With few dark hairs in the supraclipeal area, next to the ocelli and on the frons. The pubescence of the thorax is pale on the pronotum, with dark setae on the lobes. Dark setae in the anterior third of the mesoscutum, but divided in the middle by a line of white setae and bordered posteriorly by white setae. Mesoscutum disc with fine bronze-green scales and few setae; The punctuation is very evident there in two sizes, the small denser dots and the thick ones very dispersed. On the mesepisternum there is white pubescence above and below an irregular disc area of dark setae and ventrally the setae are also dark. The terga are covered with bronze-green scales and in certain light give the impression of being dark at the base of each tergum, each side of the terga with a small longitudinal line of white setae on the border with the ventral surface which is covered with setae quite short white ones. The long setae of T1 toT5 are white, in the last two black. The integument of mandibles, labrum and legs is rusty; the tegulae and pronotal lobes of a lighter rust color.
Figure 20. Mesoplia (M.) veracruzana sp. nov. Holotype, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
The spur of the middle tibia widened towards the apex and bifurcated, with the inner ramus having a single tooth and the outer ramus having a toothed inner edge. The middle basitarsus with the posterior angle between the external and internal surfaces only edged, not laminated. Hind femurs thickened near base projecting backwards at a large finger-like angle. The tibia moderately thickened at the apex, internally more or less flattened, distally on the internal surface a short imperfect tuft and with a relatively short spur. The posterior basitarsus with the inner side flattened and smooth, largely glabrous and separated from the posterior surface by a thin carina. T7 with the apex narrowly emarginated in a paraboloid shape, the tips relatively close together and strongly rounded. The S4 widely emarginated, procurved, with a wide short band ending at the level of the edge. S5 with deeper emargination, very pubescent, hairs curved towards the center. Approximate length 13.2 mm, forewings including tegula 13 mm; head width 4.28 mm, abdomen 5.45 mm.”
Variation.The shape of the S8 seems to be higly variable; and the shape of the basal projection of the posterior femur on males, in some individuals it is wider, while in others it is narrower.
Material examined. Costa Rica: no location data, 1♂ (MZUCR HY6836). Guanacaste: Bagaces, 8 km al NE de Bagaces, Pan Am Hwy, 10°32’17.67” N, 85°19’53.58” W, 116 m, 23-II-2000, P. Ronchi & G. W. Frankie, 1♂ (MZUCR HY5770), 24-II-2000, P. Ronchi & G. W. Frankie, 2♂ (MZUCR HY3446), (MZUCR HY3448), 12-II-2000, P. Ronchi & G. W. Frankie, 2♂ (MZUCR HY3450), (MZUCR HY3447), 9-III-2000, P. Ronchi & G. W. Frankie, 1♂ (MZUCR HY6835); Urban Bagaces, 10°31’39.84” N, 85°15’25.26” W, 83 m, 12-III-2004, L. Sandoval & G. W. Frankie, 2♂ (MZUCR HY6825), (MZUCR HY6824); Hacienda Monteverde 9 km NW Bagaces, 10°33’17.91” N, 85°16’12.50” W, 91 m, 24-II-1996, G. W. Frankie, 1♀ (MZUCR HY5697), 18-III- 2022, J. Lobo, 1♀ (MZUCR HY6828); Carrillo: Filadelfia, 11.2654 SW of Filadelfia, 10°22’21.18” N, 85°37’29.14” W, 85 m, 3-III-1954, A. Wille & H. Daly, 1♀ (SEMC 1451159); Hojancha: Lajas, 9°59’34.55” N, 85°27’5.06” W, 200 m, 18-V-2014, S. Galbraith, 2♀ (INB0004444317 – INB0004444319); La Cruz, Cuajiniquil, Bahía Tomás, Salinitas, in mangrove, 10°55’17.40” N, 85°42’57.96” W, 0 m, 25-III-2021, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY2792), 7-II-2022/7-III-2022, M. F. Otárola & J. Lobo, 1♀ (MZUCR HY5064), 25-III-2021, M. F. Otárola & J. Lobo, 2♀ (MZUCR HY2348), (MZUCR HY2349), 18-III-2022/-IV-2022, M. M. Chavarría & P. Hanson, 1♀ (MZUCR HY5065), 9-IV-2022/7-V-2022, M. M. Chavarría & P. Hanson, 3♀ (MZUCR HY5063), (MZUCR HY5066), (MZUCR HY5066); Liberia, 10°38’55.46” N, 85°26’39.36” W, 149 m, 21-II-2013, 1♀ (MZUCR HY3449); Barrio La Arena, 10°36’21.96” N, 85°25’30.23” W, 142 m, 15-V-1980, E. Ponce, 1♂ (MZUCR HY6817); Pan Am Hwy, 10°38’32.38” N, 85°27’13.36” W, 132 m, 22-II-2004, G. W. Frankie & S. B. Vinson, 1♂ (MZUCR HY5696); Santa Cruz, Playa Junquillal, 10°10’7.04” N, 85°48’47.87” W, 3 m, 3-III-1976, R. M. Bohart, 1♀ (SEMC 1442317). Heredia: CINAT Lagunilla, 9°58’23.58” N, 84°7’45.55” W, 1,038 m, 30-VII-1997, A. Ortiz, 1♂ (CINAT JZ337), 27-III-2017, R. Díaz, 1♀ (CINAT JZ339); Sarapiquí: Puerto Viejo, Estación Biológica La Selva (OET), 10°25’49.70” N, 84°0’24.96” W, 50 m, 17-VII-1995, P. Hanson, 1♀ (MZUCR HY6826), 2-IV-1993, P. Hanson & C. Godoy, 1♀ (MZUCR HY6818). Puntarenas: Golfo Dulce, 8°42’13.42” N, 83°29’8.86” W, 10 m, XII-1989/III-1990, P. Hanson, 1♀ (MZUCR HY6821); Gromaco, 34 km S of Potrero Grande en río, 8°50’49.49” N, 83°2’41.43” W, 836 m, 21-VII-1963, C. D. Michener & W. Kerfoot, 1♀ (SEMC 1451161); Rincon, 5 km S of Rincon, 8°40’57.72” N, 83°28’24.94” W, 21 m, 15-III-1973, E. Borrows, 1♂ (SEMC 1451162). San José, 9°40’0.01” N, 84°0’0.00” W, C. D. Michener, 1♀ (SEMC 1451160); Pérez Zeledón, Esperanza, 9°16’30.64” N, 83°42’9.86” W, 807 m, II-1997, N. Siles, 1♂ (CINAT JZ336), 13-IX-1999, A. Ortiz, 1♂ (CINAT JZ338); San Antonio de Escazú, 9°53’23.88” N, 84°8’17.66” W, 1,454 m, W. Edward, 1♂ (MZUCR HY6834). Guatemala: without locality, 28-III-1923, 1♀ (SEMC 1451087). Huehuetenango: La Libertad, 15°34’12.00” N, 91°51’0.00” W, 915 m, 27-IV-2010, M. Hurtado, 1♂ (ECOAB 25637), 28-IV-2010, C. Balboa, 2♂ (ECOAB 25639), (ECOAB 38876), L. Aguilar, 1♂ (ECOAB 25642), 26-II-2010, C. Balboa 2♂ (ECOAB 25643), (ECOAB 25646); San Antonio Huista, 15°36’36.00” N, 91°44’24.00” W, 1,192 m, 23-II-2010, D. Sánchez, 1♂ (ECOAB 25645). Honduras: Ocotepeque: Lucerna, Autopista 4, 9 km N of Lucerna, 14°37’36.00” N, 89°3’42.00” W, 792.48 m, 26-V-2003, D. Brzoska, 1♀ (SMO 418386). Mexico: No location data, J. Mérida, 1♀ (ECOAB 25656). Campeche: Champotón, 18°33’0.00” N, 89°56’24.00” W, 155 m, 8-VI-2018, J. Mérida, 3♀ (ECOAB 121736 – ECOAB 121738), 18°27’0.00” N, 89°53’24.00” W, 168 m, 6-VI-2018, 7♀ (ECOAB 121803 – ECOAB 121806), (ECOAB 121813 – ECOAB 121816). Chiapas: Acacoyagua, 15°24’0.00” N, 92°39’0.00” W, 862 m, 15-V-2007, R. Ayala, 1♂ (ECOAB 56633); Arriaga: 16°19’12.00” N, 93°58’48.00” W, 367.5 m, 10-III-2009, M. Guzmán, 1♂ (ECOAB 25641), 19-V-2009, C. Balboa, 1♀ (ECOAB 25657); Ej. López Mateos, 16°20’46.60” N, 93°58’26.80” W, 255 m, 13-III-2009, C. Balboa, M. Guzmán y M. Cigarroa, 2♂ (EBCH ECO-TAB57347), (EBCH ECO-TAB57276); Cacahoatán, 5 km. sur de Cacahoatán, 14°57’0.00” N, 92°9’0.00” W, 340 m, 18-IV-1993, A. Rodríguez P., 2♀ (EBCH JZ70), (SMO 317151), 2♂ (EBCH JZ77), (SMO 317154); La Concordia, 15°45’0.00” N, 92°58’12.00” W, 921 m, 23-IV-2021, P. Sagot, 1♂ (ECOAB 125000), J. Mérida, 2♂ (ECOAB 125229), (ECOAB 125230), 5-III-2020, P. Sagot, 2♂ (ECOAB 134690), (ECOAB 134691), E. Gómez, 2♂ (ECOAB 134756), (ECOAB 134757), 16°2’24.00” N, 92°33’36.00” W, 580 m, 5-III-2020, P. Sagot, 1♂ (ECOAB 134785); Mapastepec, 15°31’12.00” N, 92°48’0.00” W, 316 m, 22-II-2006, J. Esponda, 1♂ (ECOAB 25644), 1-IV-2005, M. Rincón, 1♀ (ECOAB 25660); Motozintla de Mendoza, 15°22’48.00” N, 92°19’12.00” W, 1,230 m, 7-II-2011, P. Sagot, 1♂ (ECOAB 25638), 15°19’12.00” N, 92°20’24.00” W, 1,000 m, 3-II-2019, P. Sagot, 1♂ (ECOAB 131078); Ocosingo, 16°54’36.00” N, 93°40’48.00” W, 950 m, 15-IV-2010, J. Mérida, 1♂ (ECOAB 56637); San Juan Cancuc, 16°54’0.00” N, 92°21’36.00” W, 1,170 m, 6-IV-2010, J. Mérida, 1♂ (ECOAB 25654); Siltepec, 15°34’48.00” N, 92°39’0.00” W, 1,709 m, 9-III-2022, L. Arroyo, 1♀ (ECOAB 71706), 1♂ (ECOAB 71705); Tapachula, 14°53’24.00” N, 92°17’24.00” W, 127 m, 9-VI-1989, A. Aquino, 1♂ (ECOAB 25648), 14°54’0.00” N, 92°18’36.00” W, 102 m, 14-XII-1988, A. Aquino, 2♂ (ECOAB 25649), (ECOAB 25651), 18-I-1989, A. Aquino, 1♂ (ECOAB 25650), 1♀ (ECOAB 25658), M. Cigarroa, 2♂ (ECOAB 25652), (ECOAB 25653), 1♀ (ECOAB 25659); Tonalá, 16°8’24.00” N, 93°40’48.00” W, 407.5 m, 21-V-2009, C. Balboa, 1♂ (ECOAB 56635), 12-VII-2009, C. Balboa, 1♀ (ECOAB 56636), 9-III-2009, M. Guzmán, 1♀ (ECOAB 25655), 16°30’0.00” N, 93°40’48.00” W, 407.5 m, 12-II-2009, C. Balboa, 1♀ (ECOAB 61187), 15°56’24.00” N, 93°49’12.00” W, 6 m, 31-I-2019, I. Pérez, 1♂ (ECOAB 130806), 25-I-1989, M. Cigarroa, 1♂ (ECOAB:66072); Tuxtla Chico, 14°58’12.00” N, 92°10’48.00” W, 395 m, 18-II-1992, M. Medina, 1♀ (ECOAB 56638), 1♂ (ECOAB:25647); Venustiano Carranza, 16°18’60.00” N, 92°33’59.76” W, 570 m, 21-IV-1993, J. L. Neff, 1♀ (EBCH JZ71). Guerrero: Iguala, 18°20’37.00” N, 99°32’21.01” W, 731.52 m, 4-II-1954, R. R. Dreisbach, 1♂ (SEMC 1451084); Tlapa de Comonfort, 17°33’0.00” N, 98°34’48.00” W, 1,063 m, 17-IV-1989, A. Aquino, 1♂ (ECOAB 56634). Jalisco: Cabo Corrientes, 19°39’0.00” N, 105°10’48.00” W, 49 m, 8-IX-2018, J. Mérida 1♀ (ECOAB 121569); El Tuito, 20°21’20.50” N, 105°18’59.96” W, 18-VI-1989, A. Borquez, 1♂ (EBCH JZ79); Jardín Botánico, 20°27’57.64” N, 105°17’30.89” W, 360 m, 26-VII-2014, C. E. Alatorre-Bracamontes y M. Vásquez-Bolaños, 1♂ (CZUG JZ12); La Huerta: Chamela, 19°29’54.77” N, 105°2’41.22” W, 20-VII-1985, R. Ayala, 2♂ (EBCH RA178bis JZ73), 1♂ (EBCH RA178bis JZ76), 19-XI-1989, G. Rodríguez, 2♂ (EBCH JZ74), (EBCH JZ75); Tequila: Volcán de Tequila, 20°51’50.99” N, 103°50’35.00” W, 1,300 m, 15-III-1996, H. E. Fierros-López, 1♂ (HFL JZ11), 15-III-1995, H. E. Fierros-López, 1♂ (CZUG 344); Zapopan: Barranca la Experiencia, 20°44’6.13” N, 103°19’38.38” W, 30-I-1997, G. Mercado, 1♂ (CZUG 1110), 31-VII-1996, G. Mercado, 1♂ (CZUG 1109). Morelos: Temixo: Cuentepec, 18°51’38.16” N, 99°19’30.72” W, 1,485 m, 31-X-2019, D. A. Hernández-Márquez, 1♀ (MZFC 33199), 18°51’27.72” N, 99°19’40.80” W, 1,455 m, 22-XII-2018, D. A. Hernández-Márquez, 1♀ (MZFC 32554). Querétaro: Jalpan de Serra, 21°13’0.01” N, 99°28’21.00” W, 770 m, 24-V-1989, D. Yanega, 2♀ (SEMC 1451144), (SEMC 1451145). Nayarit: San Blas, 20 km al sur de San Blas, 21°23’25.00” N, 105°6’14.00” W, 10 m, 7-IV-1994, F. Noguera, 1♂ (SMO 328474). Oaxaca: San Pedro Totolapa, 8.04672 km al noroeste de San Pedro Totolapa, 16°43’23.12” N, 96°19’14.27” W, 1,158.24 m, 6-VII-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451086); Putla Villa de Guerrero, 17°0’36.00” N, 97°57’0.00” W, 849 m, 24-V-2020, Wilebaldo, V., 1♀ (ECOAB 58886). Puebla: Cuetzalan del Progreso, 20°3’36.00” N, 97°28’12.00” W, 413 m, 26-III-2020, L. Arroyo, 1♂ (ECOAB 137985). Sonora: Álamos, La Aduana, 27°2’27.16” N, 109°0’57.91” W, 567 m, 15-III-1962, L. A. Stange, 1♀ (SEMC 1442316). Tabasco: Huimanguillo, 17°49’48.00” N, 93°55’12.00” W, 21 m, 3-VI-2015, P. Sagot, 1♂ (ECOAB 77134). Veracruz: without locality, 1♀ (CERUADY 037ver); Acayucan, 48.2803 km al sur de Acayucan, 17°38’6.41” N, 94°54’14.40” W, 56 m, 21-IV-1962, L. A. Stage, 1♀ (SEMC 1442318); Coatepec, 19°26’42.36” N, 96°57’52.13” W, 1,200 m, 1-VI- 2020, G. Quintos-Andrade, 3♀ (IEXA JZ40 – IEXA JZ42), 3-VI- 2020, G. Quintos-Andrade, 1♀ (IEXA JZ43). 4-VI- 2020, G. Quintos-Andrade, 1♀ (IEXA JZ44), 7-VI-2020, G. Quintos-Andrade, 1♀ (IEXA JZ45), 6-VI- 2020, G. Quintos-Andrade, 3♀ (IEXA JZ46 – IEXA JZ48); Medellín: Paso del Toro, 4.82803 km SW of Paso del Toro, 19°0’46.43” N, 96°8’39.30” W, 15.24 m, 23-VI-1961, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451081); Teocelo, Llano Gande, 19°22’13.01” N, 96°52’50.99” W, 833 m, 17-VI-1990, G. Rodríguez, 1♀ (SEMC1451143); Xalapa, 35.4056 km SW of Xalapa, 19°24’20.92” N, 96°38’46.46” W, 335.28 m, 29-VI-1953, Univ. of Kans. Mex. Exped., 1♂ (SEMC 1451176); Veracruz, 18°54’29.30” N, 96°8’16.01” W, 27-III-1951, W. P. Stephen, 1♂ (SEMC 1451083). Yucatán: Izamal, Plaza Izamal, 20°56’1.00” N, 89°1’5.00” W, 15 m, 24-V-1995, W. May, 1♀ (CERUADY 22841(02486)), 1♂ CERUADY 22838(02482), A. Canto, 2♂ (CERUADY 22488(02482)), (CERUADY:22481(02484)); Kinchil, Parque Central Kinchil, 20°55’1.23” N, 89°56’50.18” W, 8 m, 24-V-1995, F. León, 1♂ (CERUADY 019671(02480)), 8-V-1995, R. Rodríguez, 1♂ (CERUADY 021772(02481)); Xmatkuil, CCBA, 20°52’2.86” N, 89°37’28.10” W, 11 m, 18-V-2022, A. Sansores, 1♂ (CERUADY JZ206), 16-V-2022, 1♀ (CERUADY JZ211), 1♂ (CERUADY JZ207). Nicaragua: Rivas: Cárdenas, 11°14’24.00” N, 85°42’36.00” W, 90 m, 26-V-2015, E. Dorgay, 1♀ (ECOAB 66669); San Juan del Sur, 11°6’36.00” N, 85°45’36.00” W, 1 m, 6-IV-2017, M. Calero, 1♀ (ECOAB 80382), 0 m, 6-IV-2017, M. Calero, 1♂ (ECOAB 80393). Panama: Colón, 3.21869 km NW of Gamboa, 9°7’42.46” N, 79°42’55.28” W, 65 m, 21-I-1958, W. J. Hanson, 1♀ (SEMC 1451152); 8 km NW of Gamboa, pipeline road, 9°8’38.63” N, 79°43’38.22” W, 61 m, 12-I-1981, C. D. Michener, 1♀ (SEMC 1451153). Panamá: Zona del Canal, Juan Mina, 9°13’39.67” N, 79°57’2.59” W, 106 m, 22-VI-1945, C. D. Michener, 1♂ (SEMC 1451082); Curundú, 8°59’26.73” N, 79°32’27.29” W, 34 m, 31-III-1981, R. W. Brooks, 1♂ (SMO 725157), 19-V-1981, R. W. Brooks, 4♂ (SMO 725158), (SEMC 1254922 – SEMC1254924). Panamá Oeste: Laguna, Chame, 8°39’53.99” N, 80°6’10.90” W, 792.48 m, 29-IV-1945, C. D. Michener, 1♂ (SEMC1451085); Zona del Canal, Río Corona, 8°28’55.22” N, 80°1’42.15” W, 609.6 m, 3-V-1952, C. W. Rettenmeyer, 1♀ (SEMC 1451146); Isla Barro Colorado, 9°11’0.00” N, 79°51’0.00” W, 21-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1451147), 16-IV-1956, C. W. Rettenmeyer, 1♀ (SEMC 1451148), 15-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1254915), 19-II-1956, C. W. Rettenmeyer, 1♀ (SEMC 1254916), 7-III-1967, M. Naumann, 1♀ (SEMC 1451149); Fort Kobbe, 8°55’0.30” N, 79°34’59.59” W, 8 m, 11-I-1960, W. J. Hanson, 1♀ (SEMC 1451150), 1♂ (SEMC 1451151).
Figure 22. Mesoplia (M.) dugesi, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Distribution. Registered from south Sonora in northern Mexico, to Brazil, according to opinion of Melo and Genaro (Genaro & Franz, 2008), its distribution is restricted to continental America, so its presence on the islands of the continent would be ruled out (Fig. 27).
Taxonomic comments. Within the genus, it is the species with the widest distribution (Fig. 27), largest number of registered hosts (Table 2). This is possibly a species complex; we did not find morphological characters that allow us to delimit it in a group of species.
Diagnosis. Female: body metallic blue but black in face and mesosoma and metasoma (Fig. 28); scape with dark integument on the base and reddish brown on apex; pedicel with dark integument; integument of f1 ferruginous, remainder of flagellum dark brown; labrum, without metallic blue scales on vertex, occiput and tegula (or not very evident on tegula (Fig. 28B)); tegula integument dark brown; legs with integument on outer surface of blackish and inner surface dark reddish brown; with a carina extending laterally from the pronotal collar to the pronotal lobe; conical scutellar tubercles moderately projected, with a median cleft between them (Fig. 28D); tergal pubescence blackish; middle tibial spur forked with dark reddish-brown integument, anterior projection with 4 internal teeth; pygidial plate with a broad apex and almost completely covered by metallic bluish-green scales (Fig. 4G).
Figure 23. Mesoplia (M.) dugesi, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Description. Holotype: female. Body length 13.5 mm, forewing length 12.5 mm, head length 3.5 mm, intertegular distance 4.4 mm, T2 length 2.8 mm. Head: mandible with an internal tooth. Labrum subrectangular, surface slightly concave, with a raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding above the head in lateral view. Mesosoma: with a carina extending laterally from the pronotal collar to the pronotal lobe. Scutum convex in lateral view. Scutellar tubercles conical, without posterior carina, with an evident cleft between them. Vertical metanotum and propodeum. Short pointed middle tibial spine. Medial tibial spur bifurcated, anterior projection with 4 internal teeth. Medium basitarsus with tridentate external margin and with a thin flattened distal process slightly curved externally. Metasoma: broad pygidial plate at the base, and with a broad rounded apex (Fig. 4G). Color: integument blackish to dark reddish brown. Head: scape mostly blackish, with dark reddish brown upper outer edge. Pedicel also bicolor. F1 ferruginous, remainder of flagellum brown. Basal and apical region of the mandible dark, with a yellowish brown area in the base of the teeth and reddish brown near the base. Mesosoma: pronotal lobe and tegula blackish. Wings almost transparent. Forewings with the anterior edge and the distal apex of the marginal cell darker, also presenting a darker spot that goes from the external edge of the 2r-m to the wing apex (Fig. 28B). Very dark veins. Inner surfaces of legs dark reddish brown, outer surfaces blackish. Apex of distal process of middle basitarsus blackish. Middle tibial spur reddish-brown. Metasoma: sterna blackish, except for S6 which is reddish brown. Surface sculpture: head. Labrum with most surface rugose, basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: dotted pigidial plate. Pubescence: approximately the same proportion of blackish and whitish. Head: labrum discal area with white plumose setae and scattered simple black setae; lower edge with a pair of long dark tufts; lateral edges with simple black setae and upper edge bare. Clypeus and supraclypeal area, with lateral with white plumose setae, discal surface with little pubescence, some elongated black simple setae evident in lateral view. Frons with brown setae. Paraocular area with white plumose setae. Vertex and preoccipital region with white setae. Gena with white setae, except for the darks on lower area. Mesosoma: pronotal lobe, with the inner and posterior margins with white setae, rest with blackish setae, with light metallic green scales, only slightly evident in frontal view. Mesepisternum with blackish setae, with a thick longitudinal white line. Scutum covered mainly by black setae, except for the lateral edges near the posterior two-thirds of the tegula that have white setae, a central disc with light metallic green scales. Tegula with black setae on the anterior margin and white on the posterior, and with few light metallic green scales on the inner margin, only slightly evident in the opposite lateral view. Scutellum, upper area with few black setae and light metallic blue scales, posterior part with white setae project a little between the tubercles, also with light metallic blue scales in posterior view. Metanotum blackish. Propodeum upper region naked, posterolateral region blackish. Mainly black legs. Outer surface of anterior tibia with a few light metallic green scales, some white plumose setae and simple blackish elongated setae and also present on the basitarsus. With light metallic blue scales evident on the outer surface of the tibia and basitarsus of the middle and hind legs. Metasoma: terga covered with metallic blue-green scales and setae almost black and simple, except on the side of T5, S2 to S4. T6, with thick black setae on the sides; pygidial plate with light metallic green scales that do not reach the apex (Fig. 4G); sterna with scattered black setae.
Figure 24. Mesoplia (M.) rufipes, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Male: unknown.
Taxonomic summary
Type material. Holotype: ♀ ECOAB 74052 MEX. OAX [Mexico, Oaxaca], Santiago Nacaltepec, 17°28’12.00” N, 96°49’12.00” W, 1,377 m, 4-XII-2014, L. M. Reyes (ECOAB).
Etymology. The name oaxacana makes reference to the known distribution of this species, the state of Oaxaca.
Distribution. It is only known from the type locality (Fig. 10).
Taxonomic comments. This species is similar to M. dugesi since the majority of the pubescence is dark. But this species has few metallic scales on the face (Fig. 28C) and tegula (Fig. 28B), and the apex of the pygidial plate is broad (Fig. 4G).
Subgenus Eumelissa Snelling & Brooks, 1985
Eumelissa Snelling & Brooks, 1985: 21. Type species: Melissa decorata Smith, 1854: British Museum Natural History, London, England (♀), by original designation.
Diagnosis. Male: posterior tibia, distal inner surface without a conspicuous setal brush (Fig. 3F, G); hind tibial spurs reaching to or beyond middle of basitarsus, both present. Female: pygidial plate narrow, does not completely occupy the exposed dorsal surface of T6; pygidial plate glabrous (Figs. 4H, 23D); dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces.
Diagnosis. Male: body metallic blue, with mostly whitish pubescence; whitish-yellowish pubescence on the face, except for the frons, which has brown pubescence; scape and pedicel with integument dark; basal and apical flagellomere, pronotal lobe, tegula and legs with integument ferruginous; anterior region of the scutellar disc pubescence with a color pattern that goes from blackish in the anterior region, brownish-yellowish to whitish (Fig. 29B); scutellar tubercles flattened; forked spur of middle tibia ferruginous, becoming dark at apex, both branches short, external apex with 3 or 4 internal teeth (Fig. 16D). Terga pubescence whitish, with a pair of white spots with short pubescence in the marginal area of T2 to T6 (Fig. 29D); forked apex of T7, both tips truncated and bare, median cleft shallow (Fig. 7F); S4 bilobed, with a band of very short, dark hairs on the distal margin; dorsal lobe of the gonostylus elongated and projected towards the penis valvae (Fig. 30C, D).
Figure 25. Mesoplia (M.) rufipes, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
Female: body metallic blue, with mostly whitish pubescence; F1 shorter than F2; scape and pedicel integument reddish brown; F1, pronotal lobe, tegula and legs integument ferruginous; pubescence of the anterior region of the scutum short and branched, with a characteristic pattern that transversely intersperses spots of whitish and blackish setae (Fig 20B); scutellar tubercles flattened with a superficial cleft between them and with short, bushy black pubescence on the apical edge; terga pubescence whitish; with a pair of white spots with short pubescence in the marginal area of T2 to T4 (Fig. 31A); mesosoma and metasoma, ventral surface with metallic blue scales; middle tibial spur forked with ferruginous integument, becoming dark at apex, both branches short, external apex with a series of 3 or 4 short internal teeth (Fig. 18D); pygidial plate narrow at the base, does not cover the entire posterior margin of T5, glabrous, without scales or hairs (Fig. 4H).
Description. Holotype: male. Body length, 11.3 mm (10.8-11.9; n = 4), forewing length 9.4 mm (9.1-10; n = 4), head length 3.4 mm (3.4-3.5; n = 4), intertegular distance 3.3 mm (2.8-3.5; n = 4), T2 length 1.729 mm (1.6-1.8; n = 4). Head: mandible with an internal tooth. Labrum subrectangular, flat surface, with a slightly elevated preapical margin. Ocelli aligned to the upper margin of the eyes, protruding only a little above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles with a flattened surface, a superficial cleft between them. Vertical metanotum and propodeum. Medial tibial sine short pointed (Fig. 1D). Medial tibial spur bifurcated near the apex, noticeably elongated, internal tip with a single thin elongated tooth, external tip with a series of 3 or 4 internal teeth (Fig. 16D). Medial basitarsus with posterior margin smooth and with short and straight flattened distal process (Fig. 1D). Posterior femur thin. Posterior tibia with 2 spurs at the apex, the inner one longer than the outer one (Fig. 3F). Metasoma: T7 with a flattened surface, bifurcated, with a shallow central cleft, short distance between the apices, both ends truncated (Fig. 7F). Color: blackish to ferruginous integument. Head: ferruginous labrum. Blackish face. Ferruginous scape (some specimens with a reddish spot at the apex). Dark brown pedicel. Ferruginous F1 and F11, remainder of flagellum brown. Apical third of the mandible with apical third blackish (some specimens with yellowish-brown tints), rest dark reddish brown. Mesosoma: ferruginous pronotal lobe and tegula. Blackish mesepisternum. Apex of distal process of middle basitarsus reddish brown. Spur of the middle tibia mainly ferruginous, with blackish apices. Wings almost transparent. Forewings with the anterior edge of the marginal cell darker, also presenting a subtly darkened spot that goes from the outer apex of the marginal cell to the wing apex. Brown veins. Ferruginous legs. Blackish propodeum. Metasoma: ferruginous sterna. Surface sculpture: head. Labrum with most of the surface rugose, the basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: polished T7 apical surface. Pubescence: mainly whitish. Head: labrum discal area with pale yellowish plumose setae; lower edge with a pair of long brown tufts; lateral edges with simple brown setae and upper edge bare. Clypeus and suplaclypeal area, with pale yellowish plumose setae. Frons with brown setae. Paraocular area with pale yellowish plumose and simple setae, some specimens with bright yellow setae in the upper region. Vertex and preoccipital region with white compound setae. Gena with white compound setae, except for the brown on lower area. Mesosoma: pronotal lobe, with yellowish anterior margins, white lateral and posterior margins, practically naked posterior surface. Mesepisternum anterior margin with bright yellow and brown setae, some specimens with the lower portion with a black spot, rest of the surface white except for a spot of black setae below the tegula. Scutum anterior area with a characteristic pattern that goes from blackish, passing through yellowish-brown, until reaching whitish in the region of the discal surface, the latter with metallic blue scales and scattered setae (Fig. 29B). Axila lateral edges with black setae. Tegula with white setae on the inner anterior edge and at the posterior apex, and black on the inner and outer anterior edge, rest of the surface bare. Scutellum upper region with few scattered short black setae and metallic blue scales, scutellar tubercles posterior apices with long black setae and with a line of short, branched white setae between them, posterior area with long white, branched setae (Fig. 29B). Metanotum with white setae. Propodeum upper region naked, posterolateral region whitish-light brown. Legs mainly dark. Anterior tibia outer surface scaleless, with a tuft of short plumose white setae on the outer basal surface, with long simple dark setae running from the outer apex of the tibia and extending along the basitarsus. External surface of middle and posterior tibia and basitarsus with metallic blue scales evident on the external surface. Coxa and trochanter with simple long white setae scattered. Metasoma: terga and sterna with metallic blue scales. Sternum disc with scattered long thin white setae. Anterior edge of T1 with thin white moderately bushy setae, lateral edges with a white patch of short plumose setae. Marginal zone of T2 to T4 with a pair of white spots with short pubescence, from T4 onwards they tend to merge with a lateral spot (Fig. 29D). T7 with simple black setae on the sides. T1 to T6 with a lateral stripe of short, branched setae. Sterna setae scattered black. S4 with short, branched brown setae near the apex and with several rows of simple black setae. S2 to S4 with a more or less triangular white spot on the lateral margins.
Figure 26. Mesoplia (M.) rufipes, female: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, posterior view of the scutellar tubercles.
Allotype: female. Body length 12.3 mm (11-14.3; n = 3), forewing length 12.3 mm (9.3-17.5; n = 3), head length 3.5 mm (n = 3), intertegular distance 3.5 mm (3.4-3.6; n = 3), T2 length 1.8 mm (1.6-1.9; n=3). Head: mandible with an internal tooth. Labrum subrectangular, surface flattened, with a subtly raised preapical margin. Ocelli aligned to the upper margin of the eyes, protruding only a little above the head in lateral view. Mesosoma: convex scutum in lateral view. Scutellar tubercles flattened, with a superficial cleft between them (Fig. 31A, D). Vertical metanotum and propodeum. Medial tibial spine short pointed. Medial tibial spur notably elongated bifurcated near apex, inner tip with a single slender tooth elongated and curved posteriorly, outer apex has a series of 3 or 4 short inner teeth curved anteriorly (Fig. 18D). Basitarsus medium with smooth posterior margin and with flattened distal process slightly curved externally (Fig. 2D). Metasoma: narrow pygidial plate at the base, forming an angle of approximately 30°, apex rounded (Fig. 4H). Color: blackish to ferruginous integument. Head: scape with dark brown inner edge, rest ferruginous. Pedicel also bicolor, dark brown and ferruginous. Flagellum goes from ferrugous in the basal region to dark brown in the distal region. Apical third of mandible blackish, base dark reddish brown, with a yellowish-brown region at base of teeth extending approximately to end of first third. Mesosoma: pronotal lobe, tegula, upper region of the mesepisternum ferruginous. Wings almost transparents; forewings with the anterior edge of the marginal cell darker, also presenting a subtly darkened spot that goes from the outer apex of the marginal cell to the wing apex. Brown veins. Ferruginous legs. Blackish propodeum. Metasoma: dark pygidial plate. Ferruginous sterna. Surface sculpture: head. Labrum with most of the surface rugose, basal edge polished. Mesosoma: punctured tegula. Dotted propodeum. Metasoma: polished pygidial plate. Pubescence: mostly whitish with some blackish areas. Head: labrum with a thick transverse central band of branched white setae and with few scattered brown simple setae; lower edge with a pair of long brown tufts and some short simple brown setae; lateral edges with simple short brown setae and upper edge bare. Clypeus and supraclypeal area with short white plumose setae, discal area with little pubescence, some simple elongated white setae evident in lateral view. Scape with a tuft of short white plumose setae at the outer edge. Frons with brown setae. Paraocular area with short white plumose setae and some long simple. Vertex and preoccipital region with white compound setae. Gena with white compound setae, except for the lower region that has brown. Mesosoma: pronotal lobe with lateral and posterior margins with short white compound setae, with scattered simple blackish setae (Fig. 31C); anterior surface with metallic blue scales, evident in frontal view; discal surface practically naked. Surface surrounding the pronotal lobe with a thick strip of light brown branched short pubescence. Mesepisternum upper surface mainly with white branched short setae, lower surface with metallic blue scales. Scutum anterior area with short and branched setae, with a characteristic pattern, central area with a thin whitish stripe, followed on both sides by a more or less rectangular blackish spot, these followed by a more or less rectangular whitish spot, after them, a quadrangular blackish spot bordered by whitish setae (the whitish areas have interspersed dark setae); discal area with scattered short simple white setae and metallic blue scales, lateral edges with branched short white setae. Tegula with white setae on the inner anterior edge and at the posterior apex, and black on the inner and outer anterior edge, rest of the surface bare. Scutellum upper region with few scattered short black setae and metallic blue scales; bushy black setae at the posterior apices of the tubercles and with a line of short, branched white setae between them, posterior region with long white, branched setae (Fig. 31B, D). Metanotum with white setae. Propodeum upper region naked, posterolateral region whitish-light brown. Legs mainly dark. Anterior tibia outer surface scaleless, with a tuft of short plumose white setae on the outer basal surface, with long simple dark setae running from the outer apex of the tibia and extending along the basitarsus. External surface of middle and posterior tibia and basitarsus with metallic blue scales evident on the external surface. Coxa and trochanter with simple long white setae scattered. Metasoma: terga and sterna with metallic blue scales. Anterior edge of T1 with white simple long setae, lateral apices with a whitish light brown spot. Marginal zone of T2 to T4 with a pair of white spots with short pubescence, in T4 they tend to merge with a lateral spot. Lateral margins of T2 toT5 and S2 to S4with a strip of short white setae (Fig. 31A). T6 with thick black setae on the sides of the naked pygidial plate. Sterna with scattered black setae.
Variation. Some males from Costa Rica also present yellow setae on the white spots of T4 to T6, the white pubescence spots appear from T1 and they all tend to be fused together.
Figure 27. Map with the known distribution of Mesoplia (M.) rufipes.
Taxonomic summary
Type material. Holotype: ♂ ECOAB 566228, MEX. CHIS [Mexico, Chiapas], Arriaga, 16°13’48.00” N, 93°58’48.00” W, 367.5 m, 10-III-2009, R. Vargas (ECOAB). Allotype: ♀ ECOAB 134689, MEX. CHIS [Mexico, Chiapas] La Concordia, 16°3’36.00” N, 92°34’48.00” W, 550 m, 5-III-2020 P. Sagot (ECOAB). Paratypes: ♀ MZUCR HY6837, CR. GUA [Costa Rica, Guanacaste], La Cruz, Cuajiniquil Bahía Tomás, Salinitas, in mangrove, 10°55’17.40” N, 85°42’57.96” W, 7-III-2022/18-III-2022, M. M. Chavarría & P. Hanson (MZUCR); ♂ MZUCR HY10213, CR. GUA [Costa Rica, Guanacaste], La Cruz, Cuajiniquil, Parque Nacional Santa Rosa, 10°54’19.51” N, 85°46’11.78” W, 80 m, 20-II-2024, M. F. Otárola, M. Solano & M. M. Chavarría (MZUCR), ♂ MZUCR HY10221, 10°54’49.68” N, 85°48’15.41” W, 33 m, 21-II-2024, M. F. Otárola, M. Solano & M. M. Chavarría (MZUCR), ♂ MZUCR HY10257, 10°50’20.40” N, 85°37’4.80” W, 295 m, 7-II-2019, J. Lobo (MZUCR), same data as the previous one, ♀ MZUCR HY10257; ♂ ECOAB 566230, GU. HUE [Guatemala, Huehuetenango], San Antonio Huista, 15°40’12.00” N, 91°57’36.00” W, 1,007 m, 20-II-2010, J. Mérida (ECOAB). Same locality as the holotype, 2♀ ECOAB 566231, 566232, 13-III-2009, C. Balboa (ECOAB); same data as the allotype, ♀ ECOAB 134688 (ECOAB); ♂ ECOAB 566229, MEX. CHIS [Mexico, Chiapas], Tonalá, 16°8’24.00” N, 93°40’48.00” W, 407.5 m, 19-I-2009, C. Balboa (ECOAB); ♂ ECOAB 67237, MEX. CHIS [Mexico, Chiapas], Chiapilla, 16°33’0.00” N, 92°0’0.00” W, 561 m, 1-I-2017, P. Sagot; ♂ ECOAB 67320, MEX. CHIS [Mexico, Chiapas], Venustiano Carranza, 16°21’0.00” N, 92°37’12.00” W, 597 m, 1-II-2017, P. Sagot (ECOAB); ♂ ECOAB 67390 MEX. CHIS [Mexico, Chiapas], Solcotenango, 16°9’0.00” N, 92°22’48.00” W, 723 m, 14-I-2017, P. Sagot. (ECOAB); ♂ ECOAB 125155, MEX. CHIS [Mexico, Chiapas], Nicolás Ruiz, 16°28’12.00” N, 92°36’0.00” W, 890 m, 8-III-2020, P. Sagot (ECOAB); ♀ ECO-TAE57096, MEX. CHIS [Mexico, Chiapas], Arriaga, ej. López Mateos, 16°20’46.60” N, 93°58’26.80” W, 255 m, 10-III-2009, C. Balboa, M. Guzmán & M. Cigarroa (EBCH). Same data as the previous one, 2♀ ECO-TAE57326, ECO-TAE57327, 3♂ ECO-TAE67405, ECO-TAE57406, ECO-TAE57407. ♀ HFL JZ5, MEX. JAL [Mexico, Jalisco], La Huerta, Chamela, Estación biológica, vereda Chachalaca, 19°29’37.20” N, 105°2’44.94” W, 300 m, 9-IV-2014/11-IV-2014, E. Ramírez & A. Estrada (CZUG); ♀ CNIN-AP 009852, MEX, OAX [Mexico, Oaxaca], Santa Maria Huatulco, Parque Nacional Huatulco, 15°45’59.40” N, 96°12’11.00” W, 14-IV-2022, A. Saldivar & P. Benítez (CNIN).
Other material examined. ♂ MZUCR HY5700, no collection data (MZUCR).
Etymology. This species is dedicated to Luz del Carmen Fernández Rodríguez, mother of the first author. The noun is feminine and declined in the genitive case.
Distribution. It is only known from Jalisco, Oaxaca and Chiapas in Mexico, Huehuetenango in Guatemala and Guanacaste in Costa Rica, it is highly probable that it is also found in Belize, El Salvador, Honduras and Nicaragua (Fig. 10).
Taxonomic comments. This species is easily recognizable by the middle tibial spur with 2 short arms in respect to the rest of the spur (Figs. 29D, 33D) and with spots of whitish setae on the sublateral area of T2 to T6 in males (Fig. 29D) and T2 to T4 in females (Fig. 31A).
Table 2
Known host associations for species of Mesoplia Lepeletier, 1841.
Cleptoparasite
Hosts
Relationship record
Type of evidence
Reference
Mesoplia (M.) bifrons (Fabricius, 1804)
Centris (Centris) caxiensis Ducke, 1907
Brazil
Confirmed
Rocha-Filho et al. (2009)
Mesoplia (M.) cubensis Genaro & Breto, 2022
Centris (Centris) poecila Lepeletier, 1841
Cuba
Probable
Genaro and Breto (2022)
Mesoplia (E.) decorata (Smith, 1854)
C. (Centris) flavofasciata Friese, 1900
Costa Rica, Guanacaste
Presumed
Vinson et al. (1987)
Mesoplia (M.) dugesi (Cockerell, 1917)
C. (Exallocentris) aterrima Smith, 1854
Mexico
Suspected
Snelling (1984)
Mesoplia (M.) insignis (Smith, 1879)
C. (Acritocentris) agilis Smith, 1874
Mexico, Jalisco
Probable
This work
Mesoplia (M.) regalis (Smith, 1854)
C. (Centris) flavofasciata Friese, 1900
Costa Rica, Guanacaste
Confirmed
Vinson et al. (1987)
C. (Centris) flavifrons (Fabricius 1775)
Brazil, Paraiba
Confirmed
Martins et al.(2014)
Mesoplia (M.) rufipes (Perty, 1833)
C. (Centris) aenea Lepeletier, 1841
Brazil, Cerrado
Probable
Aguiar and Gaglianone (2003); Rocha-Filho et al. (2009)
Mesocheira ornata Spinola, 1841: 145. Holotype: Museo Regionale di Scienze Naturale, Torino, Italy (♀). Type locality: French Guiana, Cayenne.
Diagnosis. Male: body metallic green, bright yellow and black pubescence (Fig. 32); integument of the antenna, pronotal lobe, tegula and legs black, except for some internal regions of the hind leg which are dark reddish; pubescence of the head and mesosoma mostly bright yellow; conical scutellar tubercles, with a noticeable cleft between them; short and wide forked medial tibial spur, with dark reddish integument that becomes blackish at the apex, wide anterior projection, with 2 to 3 internal teeth (Fig. 3G); pubescence of the terga mainly whitish with a bright yellow maculation on both lateral margins of T1 and T2 (Fig. 32A, B); apex of T7 forked, both pointed and bare at tip, median cleft shallow but fairly wide, separating apices considerably (Fig. 7G); S4 bilobed, with several rows of compound pubescence on the apical margin, dark in color in the center and becomes light in the middle of the apices and continues that color towards the lateral edges of the lobes; digitiform and dorsal lobe of the gonostylus of similar size (Fig. 33C, D).
Female: body metallic blue, black and pale yellow pubescence (Fig. 34); integument of antenna, pronotal lobe, tegula and legs black; F1 a little longer than F2; yellowish pubescence in preoccipital region, gena, collar of the pronotum, a broad band in the center of the mesepisternum, the posterior region of the scutellum and metascutum; conical scutellar tubercles, projecting upwards with an evident cleft between them; robust medial tibial spur, bifurcated practically in half, internal tip curved posteriorly, has an internal tooth that in some cases is located on the bifurcation, external apex wider, has 2 internal teeth; pygidial plate, narrow at the base, rounded apex, forming an angle of approximately 35°, glabrous, without scales or hairs (Fig. 34D).
Figure 29. Mesoplia (E.) carmelitae sp. nov. Holotype, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, detail of the metasoma.
Taxonomic summary
Material examined. Costa Rica: Guanacaste: Estación Biológica Maritza, 10°57’25.06” N, 85°29’42.01” W, 573 m, 1988/1999, 1♀ (MZUCR HY6820), 1990, 1♀ (MZUCR HY6822). Mexico: Chiapas: La Concordia, 15°58’48.00” N, 92°48’36.00” W, 581 m, 2-III-2016, P. Sagot, 1♂ (ECOAB 76697).
Distribution. Previously it had only been recorded in South America. If the identity of this species is confirmed, its distribution would expand to southern Mexico in Chiapas (Fig. 10).
Taxonomic comments. The male is similar to M. insignis due to the yellow coloration of the mesosoma. It is differentiated among other characters by the lack of pubescence brush on the inner margin of the posterior tibia (Fig. 3G) and by the shape of the T7 whose internal notch resembles a semicircle (Fig. 7G).
Key for species of Mesoplia Lepeletier, 1841 of Mexico and Central America.
Figure 30. Mesoplia (E.) carmelitae sp. nov. Holotype, male: A, S7. B, S8.C and D, genital capsule, dorsal and lateral view, respectively.
Males
1. Distal inner surface of the posterior tibia, with a setal brush (Fig. 3A-E); hind tibial spurs not reaching beyond basal third of basitarsus, some species with only 1 spur present [M. (Mesoplia)] ………………………………………………………………… 2
– Distal inner surface of the posterior tibia, without a setal brush (Fig. 3F, G); hind tibial spurs reaching to or beyond middle of basitarsus (Fig. 3F), both present M. (Eumelissa) …………………………………………………………………7
2(1). Inconspicuous hind tibia setal brush (Fig. 3D, E); 1 spur on the posterior tibia; ventral basal surface of posterior femur with a robust posterior projection; inner surface of posterior basitarsus with a longitudinal carina (Group azurea) ………………………………………………………………… 3
– Conspicuous hind tibia setal brush (Fig. 3 A-C); 2 spurs on the posterior tibia; ventral basal surface of posterior femur without a robust posterior projection; inner surface of posterior basitarsus without a longitudinal carina (Group bifrons) ………………………………………………………………… 4
3(2). Setae of most of the body mainly dark; metallic scales on the tegula present (Fig. 21B); integument of legs blackish (Mexico) Mesoplia dugesi (Cockerell, 1917)
– Setae of most of the body mainly whitish; metallic scales on the tegula ausent (Fig. 24B); integument of legs redish brown (Mexico to South America) Mesoplia rufipes (Perty, 1833)
– Pubescence of the mesosoma mainly blackish and whitish, if it presents yellowish setae they are restricted to the posterior region of the scutellum ………………………………………………………………… 6
Figure 31. Mesoplia (E.) carmelitae sp. nov. Allotype, female: A, lateral habitus; B, frontal habitus; C, dorsal habitus; D, posterior view of the scutellar tubercles.
5(4). Pubescence of the mesosoma almost exclusively bright yellow; central disc of scutum and dorsal surface of scutellar tubercles with yellow setae; metasoma with a spot of bright yellow pubescence on lateral margins of T1 and T2 (repeated in some specimens until T4) (Fig. 6A); T7 concavity shallow; T7 apices broadly rounded (Fig. 7A) (México) ………………………………………………………………… Mesoplia insignis (Smith, 1879)
– Pubescence of the mesosoma bright yellow interrupted by a broad transverse band in the anterior region; central disc of scutum and dorsal surface of scutellar tubercles with scattered black setae; metasoma with a bright yellow pubescent spot on both lateral margins of T1 (some specimens also with a very small spot on T2) (Fig. 11A); T7 concavity deep; T7 apices narrowly rounded (Fig. 7B) (Costa Rica) ………………………………………………………………… Mesoplia tica sp. nov.
6(4). T7 with apices rounded and setae on ventral side come out a little in dorsal view (Fig. 7C) (Mexico and Central America) ………………………………………………………………… Mesoplia sapphirina Melo & Rocha-Filho, 2011
– T7 with apices flat and with ventral setae below conspicuous in dorsal view (Panama and South America) ………………………………………………………………… Mesoplia regalis (Smith, 1854)
7(1). Pubescence of the head and mesosoma mainly bright yellow; metasoma with spots of pubescence bright yellowon lateral margins of T1 and T2 (Fig. 32A, B); without spots of short white pubescence on the marginal areas of T3 to T6; middle tibial spur with long branches (Fig. 16E) (South of Mexico, Chiapas, to South America) ………………………………………………………………… Mesopliaornata (Spinola, 1841)
Pubescence of the head and mesosoma mainly whitish; without yellow pubescence on the metasoma; with 2 spots of short white pubescence in the marginal zone of T2 to T6 (Fig. 29D); branches of the middle tibial spur short (Fig. 16D) (Mexico, Guatemala and Costa Rica) ………………………………………………………………… Mesoplia carmelitae sp. nov.
Figure 32. Mesoplia (E.) ornata, male: A, lateral habitus; B, dorsal habitus; C, frontal habitus; D, scutellar tubercles posterior view (photos A, B by Jorge Mérida).
Females
1. Pygidial plate broad, occupying the exposed dorsal surface of T6 and covered with scales or setae (Fig. 4A-G); dorsal surface of posterior coxa angulate or carinate at the junction with the lateral surfaces [M. (Mesoplia)] ………………………………………………………………… 2
– Pygidial plate narrow, does not completely occupy the exposed dorsal surface of T6 and without scales or setae (Figs. 4H, 23D); dorsal surface of posterior coxa abruptly rounded at the junction with lateral surfaces [M. (Eumelissa)] …………………………………………………………………9
2(1). Pronotum with a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5B) (Group azurea) ………………………………………………………………… 3
– Pronotum without a carina extending laterally from the pronotal collar to the pronotal lobe (Fig. 5A) ………………………………………………………………… 5 Group bifrons (sensu Moure, 1960a, b)
3(2). With metallic scales evident on head and tegula (Mexico) (Fig. 23B) ………………………………………………………………… Mesoplia dugesi (Cockerell, 1917)
– Without metallic scales or inconspicuous on head and tegula ………………………………………………………………… 4
4(3). Setae on most of the body mainly dark; pygidial plate with broad apex (Fig. 4G) (Mexico, Oaxaca) ………………………………………………………………… Mesopliaoaxacana sp. nov.
– Setae of most of the body mainly whitish; pygidial plate with narrow apex (Fig. 4F) (Mexico to South America) ………………………………………………………………… Mesoplia rufipes (Perty, 1833)
5(2). With bright yellowpubescence on head, mesosoma and metasoma (Fig. 20) (Mexico, Veracruz) ………………………………………………………………… Mesoplia veracruzana sp. nov.
– Without bright yellow pubescence on head, mesosoma and metasoma ………………………………………………………………… 6
6(5). With metallic scales very evident on the scape and tegula; forewings evenly smoked ………………………………………………………………… 7
– Without metallic scales on the scape and tegula or not very evident; forewings with a darker spot at the apex ………………………………………………………………… 8
7(6). Head pubescence mainly black (Fig. 9C); scutellar tubercles moderately projecting upwards (Fig. 9D); truncated pygidial plate with dark setae on the surface (Fig. 4A) (Mexico) ………………………………………………………………… Mesoplia insignis (Smith, 1879)
– Head pubescence mainly whitish (Fig. 13C); scutellar tubercles strongly projecting upwards (Fig. 13A); rounded pygidial plate with scales on the surface, in some specimens they do not reach the apex due to the abrasion of these (Fig. 4B) (Costa Rica)………………………………………………………………… Mesoplia tica sp. nov.
Figure 33. Mesoplia (E.) ornata, male: A, S7. B, S8. C and D, genital capsule, dorsal and lateral view, respectively.
8(6). Poorly developed scutellum protuberances with a shallow depression between them and low tubercles (Fig. 16A, D); the carina of the scutellum runs continuously from one side to the other (Mexico and Central America) ………………………………………………………………… Mesoplia sapphirina Melo & Rocha-Filho, 2011
– Scutellum protuberances developed with a deep depression between them and conspicuously pointed tubercles; the carina of scutellum most developed along tubercles, gradually fading to sides (Panamá and South America) ………………………………………………………………… Mesoplia regalis (Smith, 1854)
9(1). With pale yellow pubescence on the mesosoma (Fig. 34A-C); T2 to T5 without white pubescent spots; long middle tibial spur branches (South of Mexico, Chiapas, to South America) ………………………………………………………………… Mesoplia ornata (Spinola, 1841)
– Without pale yellow pubescence on the mesosoma; T2 to T5 with a couple of spots of appressed white pubescence in the marginal zone (Fig. 31A); short middle tibial spur branches (Fig. 18D) (Mexico, Guatemala and Costa Rica) ………………………………………………………………… Mesoplia carmelitae sp. nov.
Hosts. Like the rest of the members of the Ericrocidini tribe, species of the genus Mesoplia are cleptoparasites of several species of the Centridini tribe (Table 2). It should be noted that M. rufipes is the only species of the tribe that has been recorded cleptoparasitizing species of Epicharis. The cleptoparasitism relationship between M. insignis and Centris agilis was recorded for the first time in Jalisco, Mexico. The first author of this work had the opportunity to collect some specimens of this species in an aggregation of C. agilis nests in a suburban site.
Floral asossitations. The floral records reported in this section are based on information extracted directly from the collection tags of the specimens reviewed in this study (Table 3).
Figure 35. Species of Mesoplia in nature. A-B, Mesoplia (M.) insignis on Ramirezella sp. in Jalisco, México. A, male; B, female (photos by the first author). C, Mesoplia (M.) tica sp. nov. female on Stachytarpheta sp. in Alajuela, Costa Rica (photo by Daniel Garrigues). D, Mesoplia (M.) veracruzana sp. nov. female on Ipomoea sp. in Veracruz, Mexico (photo by Gerardo Quintos-Andrade). E, Mesoplia (M.) dugesi male on Tecoma stans in Querétaro, Mexico (photo by Belem Hernández). F, Mesoplia (M.) rufipes male on Duranta sp. in San José, Costa Rica (photo by “Homúnculo Daimon”).
Discussion
The complexity of the genus Mesoplia has previously been recognized for regions like the Caribbean. There is inaccurate information on its distribution due to taxonomic problems, which has caused erroneous identifications (Genaro & Breto, 2022; Genaro & Franz, 2008). For Mexico, difficulty in accessing the primary types deposited in institutions outside the country complicates the taxonomic understanding of the group. However, there are quality images available that have allowed this review, obtaining a reliable list and synonymizing the names that were required v. gr. M. insignis (Smith, 1879) and M. imperatrix (Friese, 1912), a species easily recognizable in the end.
Most of the of the species in this study present morphological characters that make them easily differentiated. However, 2 pairs of species, M. regalis and M. sapphirina; as well as M. dugesi and M. oaxacana, are quite similar to each other, particularly the females, which complicates their differentiation. Here we provide characters that allow the unequivocal identification of the mentioned species, such as scutellar tubercles and pygidial plate. Additionally, variation in characters of the male of M. rufipes, including shape of S8 and shape of the basal projection of the posterior femur, hint to the possibility that M. rufipes is a species complex, but more evidence is needed including molecular studies.
Table 3
Floral records of the species of Mesoplia Lepeletier, 1841.
Species
Plant
Family
Mesoplia (M.) dugesi (Cockerell, 1917)
Tecoma stans
Bignoniaceae
Canavalia villosa
Fabaceae
Crotalaria sp.
Libidibia sclerocarpa
Macroptilium atropurpureum
Mesoplia (M.) insignis (Smith, 1879)
Eryngium sp.
Apiaceae
Macroptilium atropurpureum
Fabaceae
Ramirezella sp.
Vitex pyramidata
Lamiaceae
Cuphea lanceolata
Lythraceae
Hibiscus tiliaceus
Malvaceae
Mesoplia (M.) regalis (Smith, 1854)
Antigonon leptopus
Polygonaceae
Mesoplia (M.) rufipes (Perty, 1833)
Ipomoea violacea
Convolvulaceae
Tabebuia rosea
Bignoniaceae
Tecoma stans
Caesalpinia sp.
Fabaceae
Haematoxylon brasiletto
Hyptis albida
Lamiaceae
Cuphea sp.
Lythraceae
Genipa americana
Rubiaceae
The association of cleptoparasitism between M. insignis and Centris agilis supports the observations of Lim et al. (2022), they observed a considerable increase in the body size of Ericocidini that could be influenced by a change of host; and supports Harrison’s rule, since both species are of similar sizes (Harrison, 1915). Therefore, in some cases where the host is unknown, it could at least be inferred. For example, M. carmelitae sp. nov. is a relatively small bee within the genus, so it is expected that its host will be a bee from the Centridini tribe of similar size (approx. 11.395 mm) and distributed in the same geographical area. There is an opportunity to study cleptoparasite-host relationships between bees of the tribe Centridini and Ericrocidini, in sites where it is known that nest aggregations periodically occur, in addition to increasing information about the immature stages of both groups.
With this research, the region studied reaches South America with 10 species of Mesoplia, despite this it is expected that there are still several species to be described for that region since Ericrocidini is richer in that region (Snelling & Brooks, 1985). Furthermore, the distribution of some of the species recognized in this work could be broader, since there is a significant information gap across some Central American countries. The area included in this study has little diversity of species of the subgenus Eumelissa, with only 2 of the 10 known species. Furthermore, the data obtained extends the distribution of Eumelissa at least to Central Mexico on the Pacific Slope (particularly in Jalisco), far north of the previous northern limit in Costa Rica (Snelling & Brooks, 1985).
Acknowledgements
This article serves as fulfillment of the first author for obtaining the degree of M.Sc. degree with emphasis on Systematics of the Posgrado en Ciencias Biológicas at the UNAM. This study was financially supported by the student grants: Beca Nacional para Estudios de Posgrado 2022 – 1 Conahcyt No. 814906 and Programa de Apoyo a los Estudios de Posgrado (PAEP) 2023. We want to thank José Luis Navarrete(CZUG), Hugo Fierros (CZUG and HFL), Jorge E. Valenzuela, Viridiana Vega, Gerardo Quintos-Andrade (IEXA), Rémy Vandame, Jorge A. Mérida Rivas, Philippe Sagot (ECOAB), Virginia Meléndez Ramírez, María José López Gómez (CER-UADY), Zachary Falin, Michael Engel, Rachel Osborn (SEMC), Maricelle Méndez, Marcela Sánchez (MNCR), Mauricio Fernández, Paul Hanson (MZUCR), Mario Gallardo, Eduardo Herrera (CINAT) for the loan of specimens and/or for the information provided about the specimens of Mesoplia deposited in their collections. We thank Stefanie Krause (ZMB) and Corey Smith (AMNH), who sent us photographs of some type specimens. We thank again Jorge Mérida (ECOAB), Marcela Sánchez (MNCR), José Luis Navarrete (CZUG), Adam Karremans (Director of Jardín Botánico Lankaster, UCR) and Nicolás Rengifo Alfonso (MZUCR) for letting and helping us to use the imaging system to take the photographs. We want to thank Daniel Garrigues, Gerardo Quintos-Andrade, Belem Hernández and “Homúnculo Daimon” for the photographs of Mesoplia in nature. Finally, we thank the reviewers that helped to improve this article.
References
Aguiar, A. M. L., & Ganglianone, M. C. (2003). Nesting biology of Centris (Centris) aenea Lepeletier (Hymenoptera, Apidae, Centridini. Revista Brasileira de Zoologia, 20, 601–606. https://doi.org/10.1590/S0101-81752003000400006
Coville, R. E., Frenkie, G. W., & Vinson, S. B. (1983). Nests of Centris segregata (Hymenoptera: Anthophoridae) with a Review of the Nesting Habits of the Genus. Journal of the Kansas Entomological Society, 56, 109–122.
Gaglianone, M. C. (2005). Nesting biology, seasonality, and flower hosts of Epicharis nigrita (Friese, 1900) (Hymenoptera: Apidae: Centridini), with a comparative analysis for the genus. Studies on Neotropical Fauna and Environment, 40, 191–200. https://doi.org/10.1080/01650520500250145
Genaro, J. A., & Franz, N. M. (2008). The bees of Greater Puerto Rico (Hymenoptera: Apidae: Anthophila). Insecta Mundi, 40, 1–24.
Genaro, J. A., & Breto, D. (2022). Descripción de dos especies nuevas de Mesoplia Lepeletier (Hymenoptera: Apoidea: Apidae) para Cuba y Las Bahamas, con notas sobre taxonomía y distribución del género en las Antillas. Insecta Mundi, 929, 1–17.
Harris, R. A. (1979). A glossary of surface sculpturing. Occasional Papers in Entomology, 28, 1–32.
Harrison, L. (1915). Mallophaga from Apteryx, and their signifcance; with a note on the genus Rallicola. Parasitology, 8, 88–100.
Hiller, B., & Wittmann, D. (1994). Seasonality, nesting biology and mating behavior of the oil-collecting bee Epicharis dejeanii (Anthophoridae, Centridini). Biociências, 2, 107–124.
Lim, K., Lee, S., Orr, M., & Lee, S. (2022). Harrison´s rule corroborated for the body size of cleptoparasitic cuckoo bees (Hymenoptera: Apidae: Nomadinae) and their hosts. Scientific Reports, 12, 10984. https://doi.org/10.1038/s41598-022-14938-9
Lobo, J. A., Fernández-Otárola, M., Chavarría, M. M., & Agraz-Hernández, C. M. (2023). Nesting biology of Centris aethyctera (Centridini, Apidae) in an estuarine environment. Apidologie, 54, 58. https://doi.org/10.1007/s13592-023-01044-6
Martins, A. C., Luz, D. R., & Melo, G. A. R. (2018). Palaeocene origin of the Neotropical lineage of cleptoparasitic bees Ericrocidini-Rhathymini (Hymenoptera, Apidae). Systematic Entomology, 43, 510–521. https://doi.org/10.1111/syen.12286
Martins, C. F., Neto V. I. S., & Cruz, R. D. M. (2019). Nesting biology and mating behavior of the solitary bee Epicharis nigrita (Apoidea: Centridini). Journal of Apicultural Research, 58, 1–11. https://doi.org/10.1080/00218839.2019.1584963
Martins, C. F., Peixoto, M. P., & Aguiar, C. M. L. (2014). Plastic nesting behavior of Centris (Centris) flavifrons (Hymenoptera: Apidae: Centridini) in an urban area. Apidologie, 45, 156–171. https://doi.org/10.1007/s13592-013-0235-4
Michener, C. D. (2007). The bees of the World(2nd Edition). Baltimore, Maryland: The Johns Hopkins University Press.
Moure, J. S. (1960a). Notas sobre os tipos de abelhas do Brasil descritas por Perty em 1833 (Hymenoptera – Apoidea). Boletim da Universidade do Paraná, Zoologia, 1, 1–23.
Moure, J. S. (1960b). Notes on the types of the Neotropical bees described by Fabricius (Hymenoptera: Apoidea). Studia Entomologica, 3, 97–160.
Moure, J. S., & Melo, G. A. R. (2023). Mesoplia Lepeletier, 1841. In J. S. Moure, D. Urban, & G. A. R. Melo (Orgs.), Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical Region. Retrieved on 30 May 2024 from: http://moure.cria.org.br/catalogue?id=113174
QGIS.org (2022). QGIS Geographic Information System. QGIS Association. http://www.qgis.org
Rocha-Filho, L. C., Morato, É. F., & Melo, G. A. R. (2009). New host records of Aglaomelissa duckei and a compilation of host associations of ericrocidini bees (hymenoptera: Apidae). Zoologia, 26, 299–304. http://dx.doi.org/10.1590/S198446702009000200012
Rocha-Filho, L. C., Silva, C. I., Gaglianone, M. C., & Augusto, S. C. (2008). Nesting behavior and natural enemies of Epicharis (Epicharis) bicolor Smith (Hymenoptera: Apidae). Tropical Zoology, 21, 227–242.
Rozen, J. G. (1969). The larvae of the Anthophoridae (Hymenoptera, Apoidea). Part 3. The Melectini, Ericrocidini, and Rhathymini. American Museum Novitates, 2382, 1–24.
Rozen, J. G. (2016). Nesting biology of the solitary bee Epicharis albofasciata (Apoidea: Apidae: Centridini). American Museum Novitates, 3869, 1–8. https://doi.org/10.1206/3869.1
Rozen, J. G., Vinson, S. B., Coville, R. E., & Frankie, G. W. (2011). Biology of the cleptoparasitic bee Mesoplia sapphirina (Ericrocidini) and its host Centris flavofasciata (Centridini) (Apidae: Apinae). American Museum Novitates, 3723, 1–36. https://doi.org/10.1206/3723.2
Silveira, F. A., Melo, G. A. R., & Almeida, E. A. B. (2002). Abelhas brasileiras: sistemática e identificação. Belo Horizonte, Minas Gerais: Ministério do Meio Ambiente & Fundação Araucária.
Snelling, R. R. (1984). Studies on the taxonomy and distribution of American Centridine bees (Hymenoptera: Anthophoridae). Contributions in Science, Natural History Museum of Los Angeles County, 347, 1–69.
Snelling, R. R., & Brooks, R. W. (1985). A review of the genera of cleptoparasitic bees of the Tribe Ericrocidini (Hymenoptera: Anthophoridae). Contributions in Science, Natural, History Museum of Los Angeles County, 369, 1–34.
Vinson, S. B., Frankie, G. W., & Coville, R. E. (1987). Nesting habits of Centris flavofasciata Friese (Hymenoptera: Apoidea: Anthophoridae) in Costa Rica. Journal of the Kansas Entomological Society, 60, 249–263.
Vivallo, F. (2010). Sistemática e filogenia da tribo de abelhas Centridini e suas relações filogenéticas com as tribos cleptoparasitas Ericrocidini e Rhathymini (Hymenoptera: Apidae)(Ph.D. Thesis). Universidade Federal do Paraná. Curitiba, Brazil.
Vivallo, F. (2014). Taxonomic revision of the cleptoparasitic bee genus Epiclopus Spinola, 1851 (Hymenoptera: Apidae: Ericrocidini). Zootaxa, 3857, 41–70. https://doi.org/10.11646/zootaxa.3857.1.2
Vivallo, F. (2021). Nesting behavior of the oil-collecting bee Epicharis (Triepicharis) analis Lepeletier (Hymenoptera: Apidae) in an urban area of Rio de Janeiro, RJ, Brazil. Journal of Apicultural Research, 60, 135–142. https://doi.org/10.1080/00218839.2020.1820150
Werneck, H. A., & Campos, L. A. O. (2020). A study of the biology of Epicharis (Epicharoides) picta using emergence-traps. Journal of Hymenoptera Research, 80, 147–167. https://doi.org/10.3897/jhr.80.56898