A new species of Heteromyoxyuris (Nematoda: Oxyuridae), parasite of Dipodomysspp. (Rodentia: Heteromyidae) from Mexico
Una nueva especie de Heteromyoxyuris (Nematoda: Oxyuridae), parásito de Dipodomys spp. (Rodentia: Heteromyidae) de México
Jorge Falcón-Ordaz a, Jesus A. Fernandez b, Whitney Preisser c, Elizabeth A. Martínez-Salazar d, *
a Universidad Autónoma del Estado de Hidalgo, Laboratorio de Morfología Animal, Ciudad Universitaria, Carretera Pachuca Tulancingo s/n Km. 4.5, 42184 Mineral de la Reforma, Hidalgo, Mexico
b Universidad Autónoma de Chihuahua, Facultad de Zootecnia y Ecología, Departamento de Recursos Naturales, Periférico Francisco R. Almada Km. 1, 31453 Chihuahua, Chihuahua, Mexico
c Kennesaw State University, Department of Ecology, Evolution, and Organismal Biology, 370 Paulding Ave NW SC 326, MD 1202, Kennesaw, GA, 30144, USA
d Universidad Autónoma de Zacatecas, Unidad Académica de Ciencias Biológicas, Laboratorio de Colecciones Biológicas y Sistemática Molecular, Av. Preparatoria s/n, Campus Universitario II, Col. Agronómica, 98066 Zacatecas, Zacatecas, Mexico
*Corresponding author: eamskandii@gmail.com (E.A. Martínez-Salazar)
Received: 14 November 2024; accepted: 29 May 2025
http://zoobank.org/urn:lsid:zoobank.org:pub:51B9D39E-8714-4DC7-B17A-19CDCF1A1CE3
Abstract
A new species of Heteromyoxyuris, parasitic on the caecum of Dipodomys merriami and Dipodomys ordii from the Mexican Plateau, is described and illustrated. This species differs from the other 3 species by the absence of lateral alae in both sexes and resembles males of Heteromyoxyuris deserti by the presence of a pair of caudal cuticular inflations. The distribution of the 4 species of Heteromyoxyuris coincides with the geographical distribution of the heteromyid rodent subfamilies Dipodomyinae and Perognathinae across the deserts and arid lands of USA and Mexico. A key to the species of Heteromyoxyuris is provided. Heteromyoxyuris garciaprietoi n. sp. is the fourth species described for the genus and the second described for Mexico.
Keywords: Chihuahua; Guanajuato; Semi-arid regions; Heteromyoxyuris garciaprietoi n. sp.; Dipodomys spp.; Parasites
Resumen
En el presente estudio, una especie nueva del género Heteromyoxyuris, parásita del ciego intestinal de Dipodomys merriami y Dipodomys ordii del Altiplano mexicano se describe e ilustra. Esta especie se diferencia de las otras especies por la ausencia de alas laterales en ambos sexos y se asemeja a los machos de Heteromyoxyuris deserti por la presencia de 1 par de inflaciones cuticulares en la región caudal. La distribución del género Heteromyoxyuris coincide con la distribución de las subfamilias de roedores heterómidos Dipodomyinae y Perognathinae en los desiertos y tierras áridas de EUA y México. Se presenta una clave para las especies del género Heteromyoxyuris. Heteromyoxyuris garciaprietoi n. sp. es la cuarta especie del género y la segunda descrita para México.
Palabras clave: Chihuahua; Guanajuato; Regiones semiáridas; Heteromyoxyuris garciaprietoi n. sp.; Dipodomys spp.; Parásitos
Introduction
The genus Dipodomys (Heteromyide: Dipodomyinae) includes 20 species of kangaroo rats occurring throughout the arid and semi-arid regions of western North America, including both warm and cold deserts, grasslands, and chaparral, reaching semi-desert habitat at the Tehuacán–Cuicatlán Valley of southern Puebla and northern Oaxaca, Mexico (Alexander & Riddle, 2005; Fernández et al., 2012, 2014).
Ten valid species of Dipodomys have been recorded in Mexico (Fernández et al., 2014); however, their parasitic helminths have been neglected. To our knowledge, Dipodomys merriami Mearns, 1890, Dipodomys ordii Woodhouse 1853, and Dipodomys phillipsi Gray, 1841 are hosts of 2 species of cestode and 10 species of nematodes; it is noteworthy that the specific composition of these parasite communities varies among the 3 host species (further details on their parasites can be found in Preisser & Falcón-Ordaz, 2019). Notably, these 3 kangaroo rat species ecogeographically coincide within the tropical subtropical desert environment North and Central American ecosystems; D. merriami and D. phillipsii are exclusively inhabitants of the tropical and subtropical desert. In contrast, D. ordii is found in this ecoregion and extends its range to include the temperate steppe, temperate desert, and tropical and subtropical steppe (Alexander & Riddle, 2005).
To date, Heteromyoxyuris Quentin, 1973 (Nematoda: Oxyuridae) is distributed in deserts and arid lands of North America (Table 1), and comprises 3 species: Heteromyoxyuris deserti (Read and Millemann, 1953), with a primary distribution in the Chihuahuan Desert (localities detailed in Table 1); Heteromyoxyuris longejector (Hannum, 1943), found mainly in the Mexican Plateau; and Heteromyoxyuris otomii García-Prieto, Falcón-Ordaz, Lira-Guerrero, and Mendoza-Garfías, 2008, which is endemic to the Mexican Plateau. Within the family Heteromyidae, 3 rodent genera —Chaetodipus (pocket mice), Dipodomys (kangaroo rats), and Perognathus (silky pocket mice— are parasitized by these species of nematodes (García-Prieto et al., 2008; Table 1). Heteromyoxyuris longejector is the most widely distributed species in Mexico (Martínez-Salazar et al., 2016; Iturbe-Morgado et al., 2017), although both H. longejector and H. otomii have been found parasitizing 3 genera across the country (Table 1).
During a monitoring of Heteromyid rodent helminth parasites in the Mexican Plateau, many nematodes of an undescribed Heteromyoxyuris species were found in the caecum of D. merriami and D. ordii. The aim of this study is to describe a new species of the mentioned genus. In addition, a key to the species of Heteromyoxyuris is provided.
Materials and methods
In December 2015, 4 D. merriami and 1 D. ordii were trapped and collected in Sherman live-trap and subsequently euthanized following the standard procedures and techniques of the guidelines of the American Society of Mammologists (Kelt et al., 2010; Sikes & The Animal Care and Use Committee of the America Society of Mammalogists, 2016), under permit FAUT-0306 (issued to JAF), from 4.6 km S Ejido Mayran (25º37’48” N, 102º50’5.1” W; 1,091 m asl), municipality of San Pedro de las Colonias, Coahuila, Mexico. The viscera of the rodents were taken out and were fixed in 4% formalin. Intestinal samples were examined for helminths in June 2019. The skins and skulls of the specimens were deposited in the Colección de Mamíferos of the Universidad Autónoma de Chihuahua (UACH-M), with the numbers UACH-M152 for D. ordii and UACH-M153 for D. merriami.
In addition, specimens of Heteromyoxyuris sp. isolated from D. merriami and deposited at the Colección Nacional de Helmintos (CNHE), Instituto de Biología, Universidad Nacional Autónoma de México (IB-UNAM), Mexico City, Mexico, were examined. The nematodes were obtained from the caecum of the host and stored in 70% ethanol. The specimens were cleared on temporary slides by an immersion in a solution of glycerol and 70% ethanol (2:1). Transverse sections were made at different levels of the body in both sexes using a razor blade and placed in semi-permanent preparations. Photographs and measurements were taken using a Zeiss microscope, which was equipped with an Axiocam ERc5s and a calibrated ocular millimeter. For scanning electron microscopy (SEM) observations, 4 males and 4 females (2 of each sex per locality; these specimens were previously measured in the morphological study and correspond with the paratype series) were dehydrated in a series of graded ethanol solutions, followed by critical point drying with carbon dioxide. Six specimens (3 from Coahuila and 3 from Guanajuato) were coated with a gold-palladium mixture and examined in a Hitachi S-2460 N SEM (Hitachi, Tokyo, Japan) at 15 kV.
Table 1
Distribution of the genus Heteromyoxyuris in deserts and arid lands of North America (DALNA).
| Species | Host | Locality and Country | DALNA* | Reference |
| H. deserti | Dipodomys deserti | Lincoln County, Nevada, USA | MD | Quentin, 1973 |
| King & Babero, 1974 | ||||
| D. merriami | Victorville, California, USA | SD | Read & Millemann, 1953 | |
| Lincoln County, Nevada, USA | MD | King & Babero, 1974 | ||
| Quentin, 1973 | ||||
| Socorro County, New Mexico, USA | ChD | Decker et al., 2001 | ||
| Otero County, New Mexico, USA | ChD | Fedynich et al., 2001 | ||
| Dipodomys heermanni morroensis (Merriam, 1907) | Morro Bay, California, USA | ODA | Read & Millemann, 1953 | |
| D. ordii | Ward County, Texas, USA | ChD | Garner et al., 1976 | |
| Socorro County, New Mexico, USA | ChD | Decker et al., 2001 | ||
| Dipodomys panamintinus (Merriam, 1894) | Palmdale, California, USA | SD | Read & Millemann, 1953 | |
| D. spectabilis | Socorro County, New Mexico, USA | ChD | Decker et al., 2001 | |
| Perognathus flavus | Socorro County, New Mexico, USA | ChD | Decker et al., 2001 | |
| H. longejector | Chaetodipus sp. | Loreto, Zacatecas, Mexico | MP | Martínez-Salazar et al., 2016 |
| Pánuco, Zacatecas, Mexico | MP | Martínez-Salazar et al., 2016 | ||
| Villa de Cos, Zacatecas, Mexico | MP | Martínez-Salazar et al., 2016 | ||
| Ch. eremicus | Mazapil, Zacatecas, Mexico. | MP | Iturbe-Morgado et al., 2017 | |
| Ch. hispidus | Hidalgo, Durango, Mexico | MP | Iturbe-Morgado et al., 2017 | |
| San Luis de La Paz, Guanajuato, Mexico | MP | Iturbe-Morgado et al., 2017 | ||
| Villa de Reyes, San Luis Potosí, Mexico | MP | García-Prieto et al., 2008 | ||
| D. merriami | Arizona, USA | UNK | Hannum, 1941 | |
| Washington County, Utah, USA | CP | Bienek & Klikoff, 1974 | ||
| Pánuco, Zacatecas, Mexico | Martínez-Salazar et al., 2016 | |||
| Villa de Cos, Zacatecas, Mexico | MP | Martínez-Salazar et al., 2016 | ||
| Mazapil, Zacatecas, Mexico | MP | Iturbe-Morgado et al., 2017 | ||
| Perognathus sp. | Clark County, Nevada, USA | MD | Quentin, 1973 | |
| P. amplus | Mazatán, Sonora, Mexico | SD | García-Prieto et al., 2008 | |
| Perognathus californicus Merriam, 1889 | Monterrey, California, USA | ODA | Quentin, 1973 | |
| Table 1. Continued | ||||
| Species | Host | Locality and Country | DALNA* | Reference |
| Peromyscus sp. | Ojo Caliente, Zacatecas, Mexico | MP | Martínez-Salazar et al., 2016 | |
| Sylvilagus floridanus | Payne County, Oklahoma, USA | ODA | Boggs et al., 1990 | |
| H. otomii | P. flavus | San Luis de La Paz, Guanajuato, Mexico | MP | Iturbe-Morgado et al., 2017 |
| Ajacuba, Hidalgo, Mexico | MP | García-Prieto et al., 2008 | ||
| H. garciaprietoi n.sp. | D. merriami | San Luis de La Paz, Guanajuato, Mexico | MP | Iturbe-Morgado et al., 2017 |
| D. merriami | San Pedro de las Colonias, Coahuila, Mexico | MP | this study | |
| D. ordii | San Pedro las Colonias, Coahuila, Mexico | MP | this study |
*Mexican Plateau (MP); Great Basin Desert (GBD); Mohave Desert (MD); Sonora Desert (SD); Outside desert areas (ODA); Chihuahua Desert (ChD); Unknown (UNK); Colorado Plateau (CP).
The species description is based on specimens parasitizing 2 species of Dipodomys (D. ordii and D. merriami) from the states of Coahuila and Guanajuato; measurements of D. merriami specimens collected in Coahuila and Guanajuato are provided in Table 2. All measurements are given in micrometers (µm), unless otherwise indicated. The measurements are reported as a range, with the mean and standard deviation in parentheses. Also, the measurements of the holotype and allotype are included in brackets. The specimens were deposited in the CNHE, IB-UNAM.
Description
Family Oxyuridae Cobbold, 1864
Genus Heteromyoxyuris Quentin, 1973
Heteromyoxyuris garciaprietoi n. sp. Falcón-Ordaz and Martínez-Salazar
(Figs. 1-3)
http://zoobank.org/urn:lsid:zoobank.org:act:716F4F32-F0BE-473C-95AB-7297A8635510
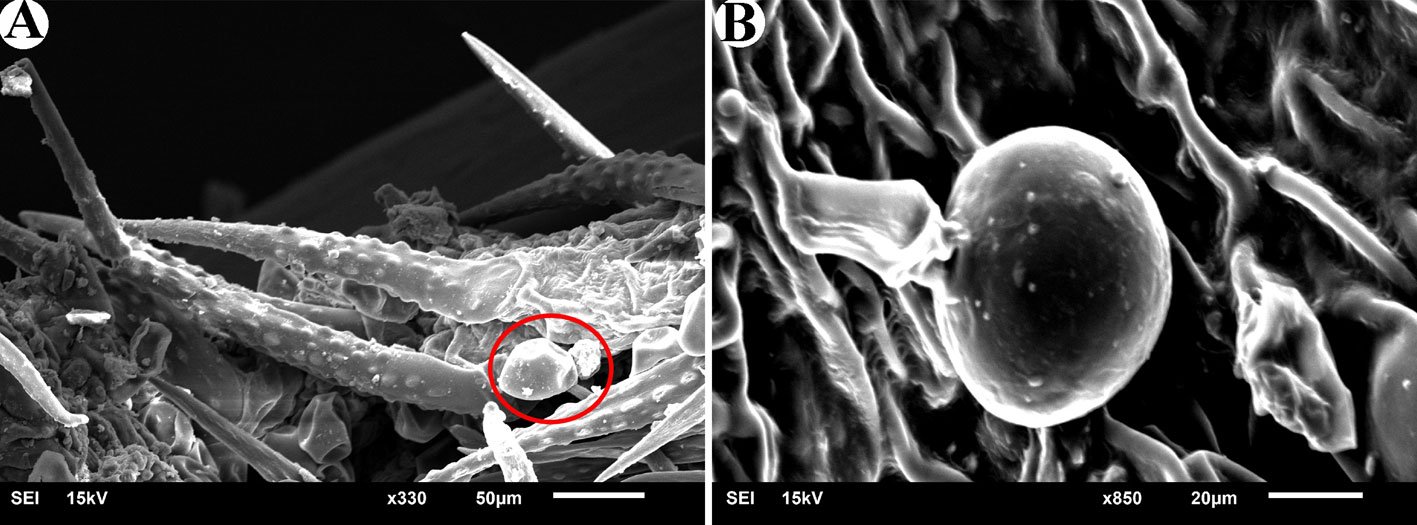
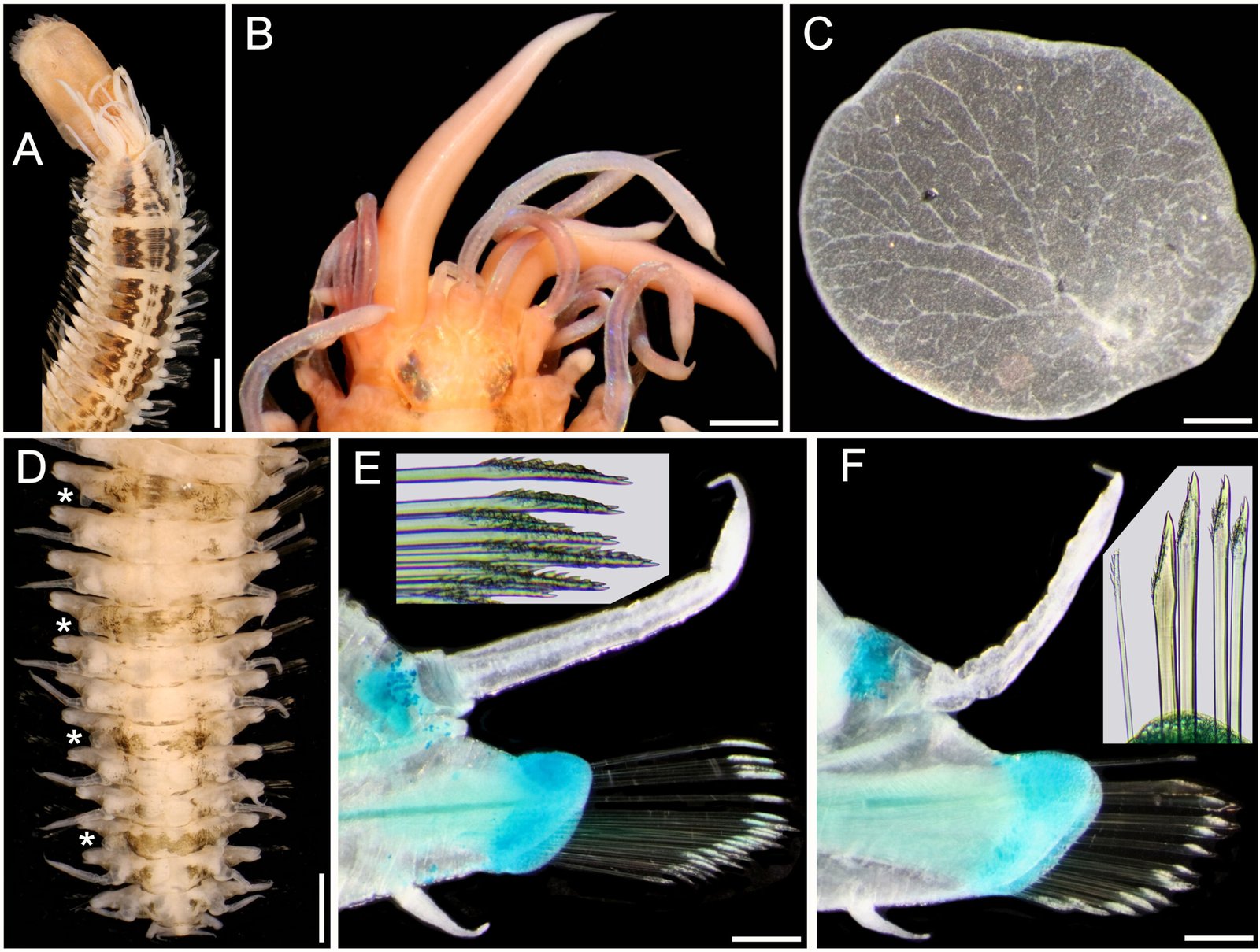
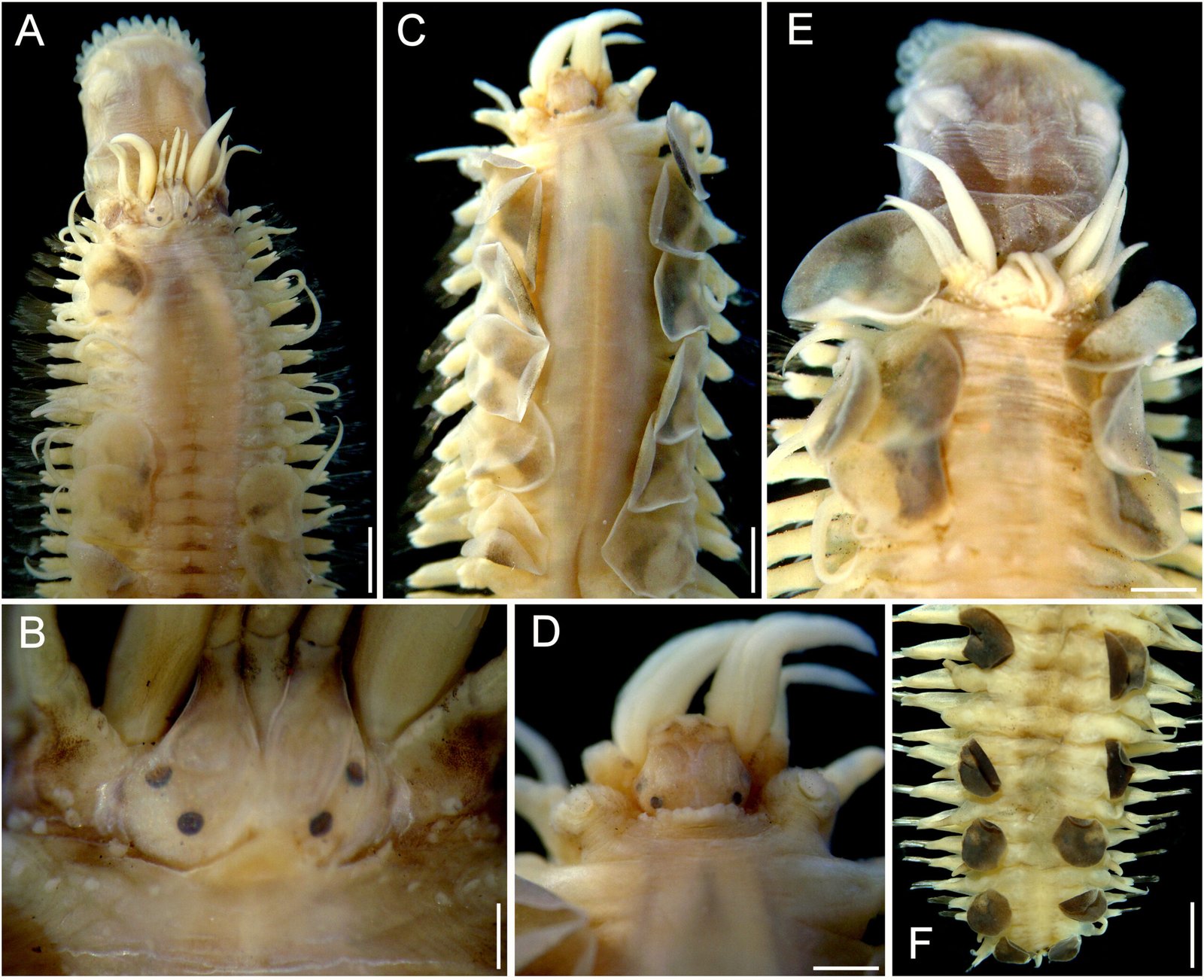
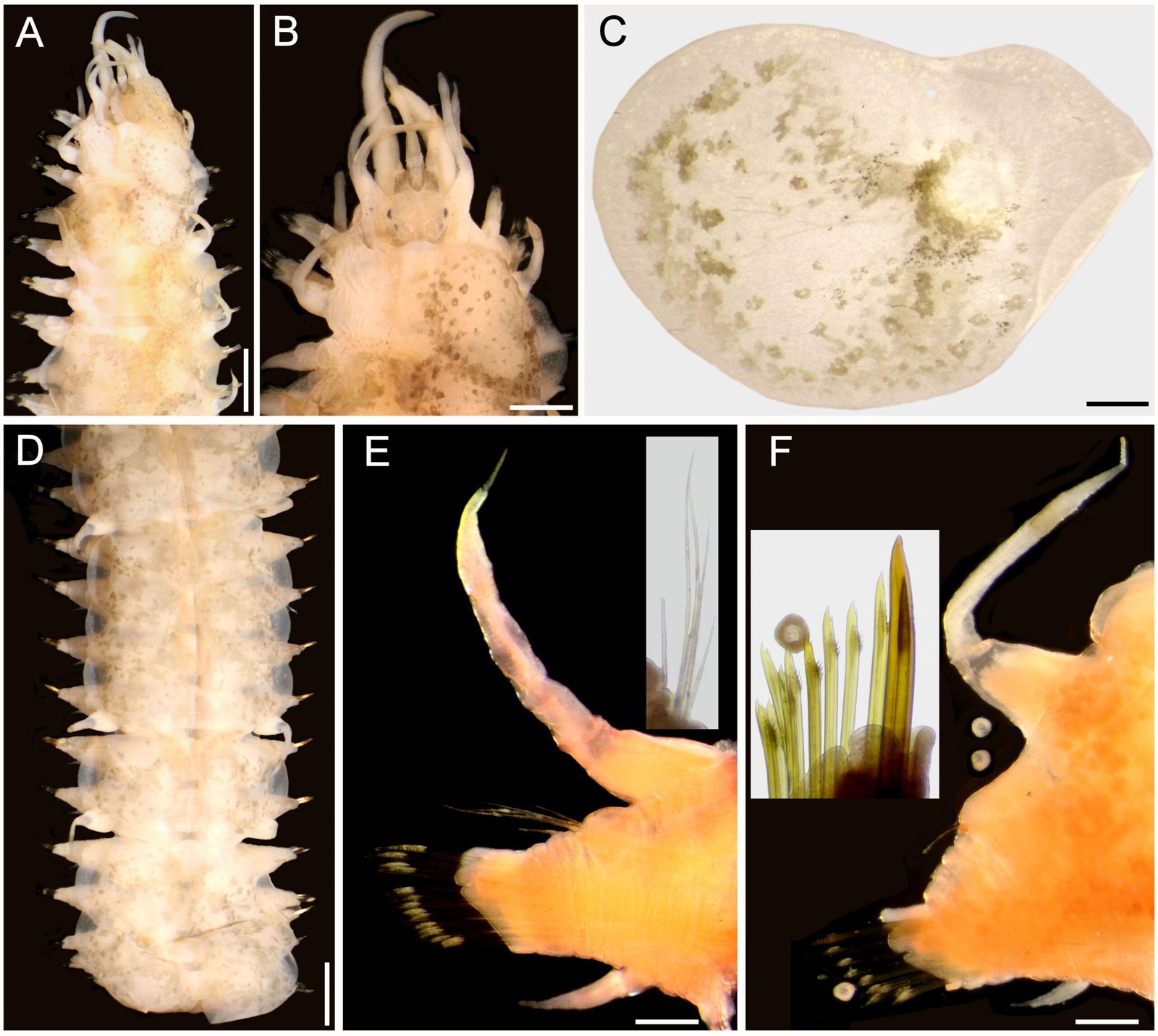
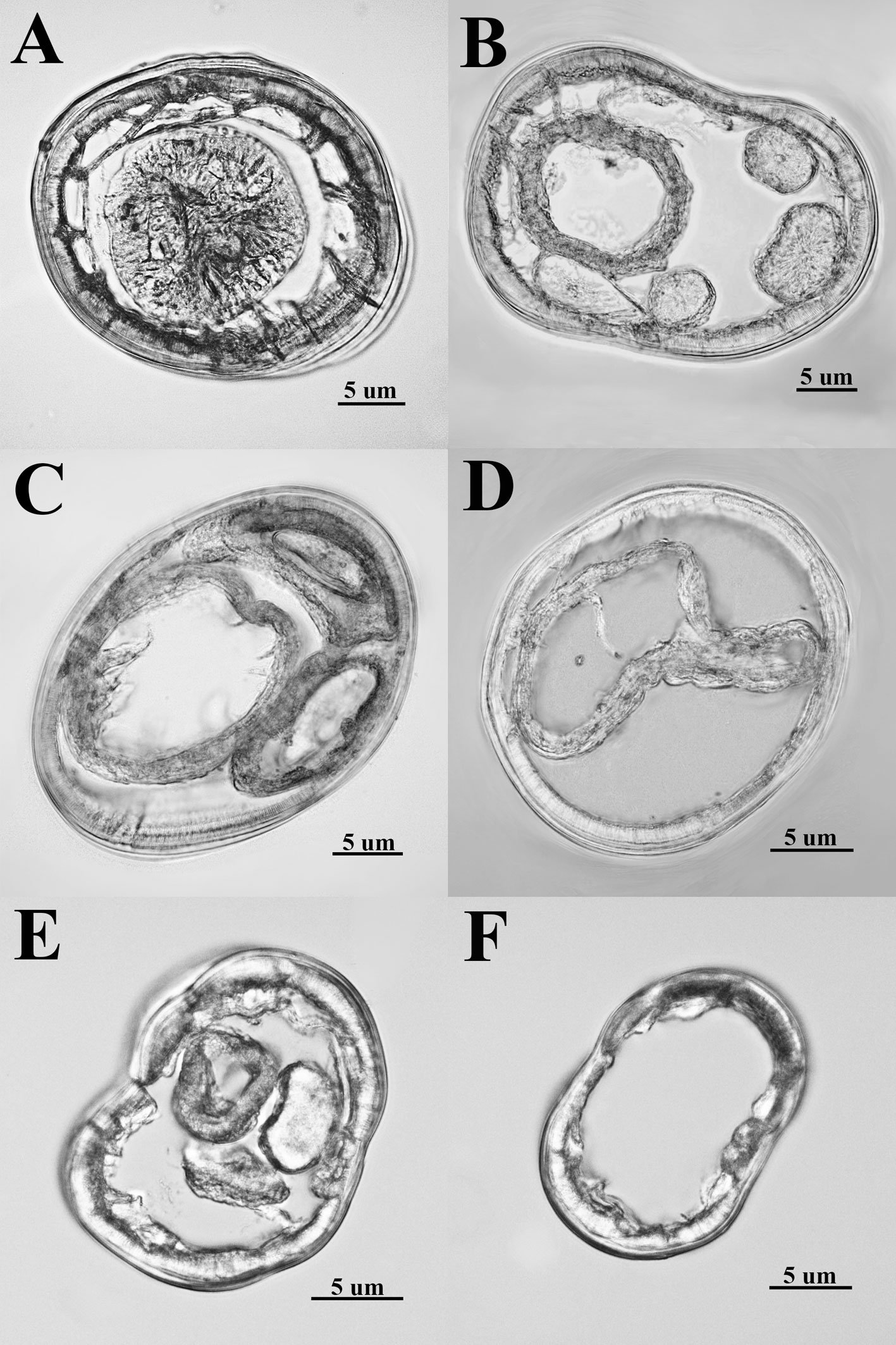
General description (based on 57 individuals, 17 males and 40 females). Nematodes small and robust. Mouth with 3 strong teeth (Fig. 1A); females with 3 interlabial processes, 2 latero-dorsal and 1 ventral (Fig. 1B). Cephalic end with 2 circles of 4 papillae each; external circle simple papillae (Fig. 1C), clearly differentiated; amphids located at external circle of papillae. Cephalic region separated from body by deep groove (Fig. 1D). Males with a pair of caudal cuticular inflations and tail acute (Fig. 1E). Females with conical tail. Excretory pore situated in first third of body. In cross sections at different levels of the body, lateral alae are not observed in either sex (Fig. 2A-F).
The measurements of the Dipodomys ordii specimens from Coahuila are given below, while those of Dipodomys merriami from Guanajuato and Coahuila are presented in Table 2.
Table 2
Measurements of paratype specimens (min.-max., mean ± standard deviation) of Heteromyoxyuris garciaprietoi n. sp., parasite of Dipodomys merriami from Guanajuato and Coahuila.
| Locality | San Luis de La Paz | San Pedro de las Colonias | ||
| State | Guanajuato | Coahuila | ||
| Sex | Male, n = 8 | Female, n = 16 | Male, n = 4 | Female, n = 8 |
| Length (mm) | 2.52-3.55 (3±0.34) | 7.30-12.45 (10.48±1.66) | 1.78-2.8 (2.5±0.5) | 9.1-11 (10.1±8) |
| Maximum diameter | 182.5-232.5 (218.12±17.81) | 265-472.5 (373.9±69.52) | 125-180 (159.3±26.3) | 360-455 (411.5±35.5) |
| Esophagus with bulb | ||||
| Length | 325-425 (381.25±32) | 550-725 (629.68±48.49) | 345-392.5 (372.5±20.1) | 570-630 606.2±21.3) |
| Esophageal bulb | ||||
| Length | 70-100 (89.37±10.66) | 100-162.5 (134.68±18.43) | 77.5-92.5 (82.5±7) | 110-132.5 (126.2±9) |
| Width | 72.5-120 (92.81±14.72) | 107.5-170 (141.25±19.4) | 72.5-90 (81.8±8.2) | 140-157.5 (150±6) |
| Nerve ring | 95-132.5 (113.43±11.72) | 130-370 (196.1±55.55) | 82.5-145 (105±28.3) | 165-195 (176.2±15.2) |
| Excretory pore | 177.5-870 (671±235.14) | 1.18-1.80 mm (1.48±0.22) | 215-350 (273.7±61.1) | 1.2-1.6 mm 1.4±0.12 |
| Spicule length | 77.5-80 (78.33±1.44) | 75-80 (77.5±3) | ||
| Gubernaculum length | 22.5-27.5 (24.5±2.09) | 25-27.5 (26.6±1.44) | ||
| Vulva | 3.0-4.8 mm (3.84±0.59) | 3.8-4.4 mm (4.1±0.22) | ||
| Ovejector | 300.0-620 (434.54±96.47) | 250-362.5 (318.4±38.2) | ||
| Anus* | 100-157.5 (129.64±20.12) | 0.85-1.47 mm (1.23±0.17) | 105-145 (126±20) | 0.96-1 mm (1±0.04) |
| Tail length | 70-127.5 (95.93±19.95) | 85-87.5 (86.2±1.4) | ||
| Caudal cuticular inflation | 70-127.5 (96±20) | 192.5-255 (221.8±28.3) | ||
| Eggs | ||||
| Length | 100-112.5 (107.63±4.23) | 97.5-112.5 (105.3±3.8) | ||
| Width | 30-40 (35.76±2.80) | 30-40 (34.2±2.4) |
*In male is cloaca and in females is anus.
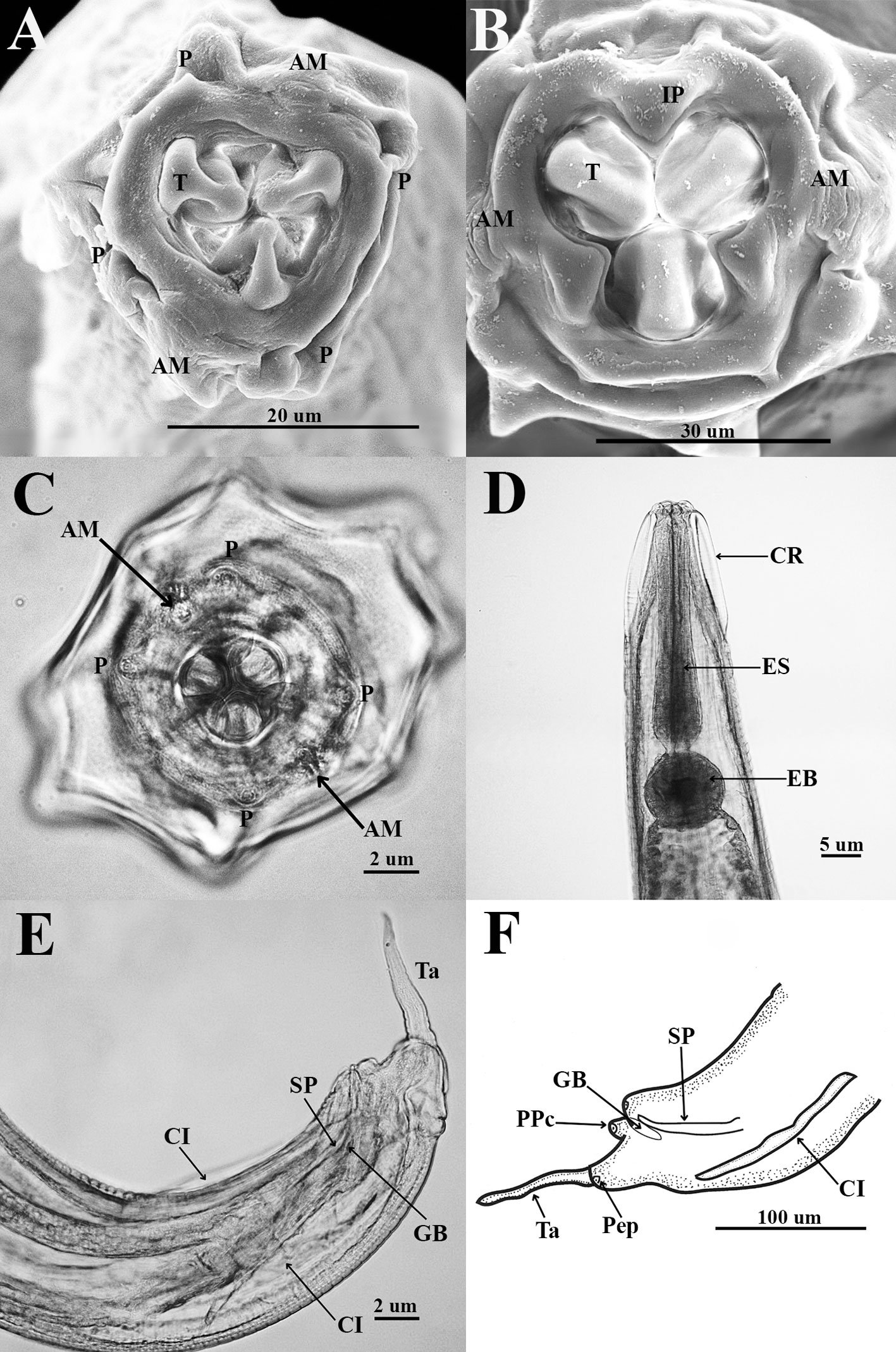
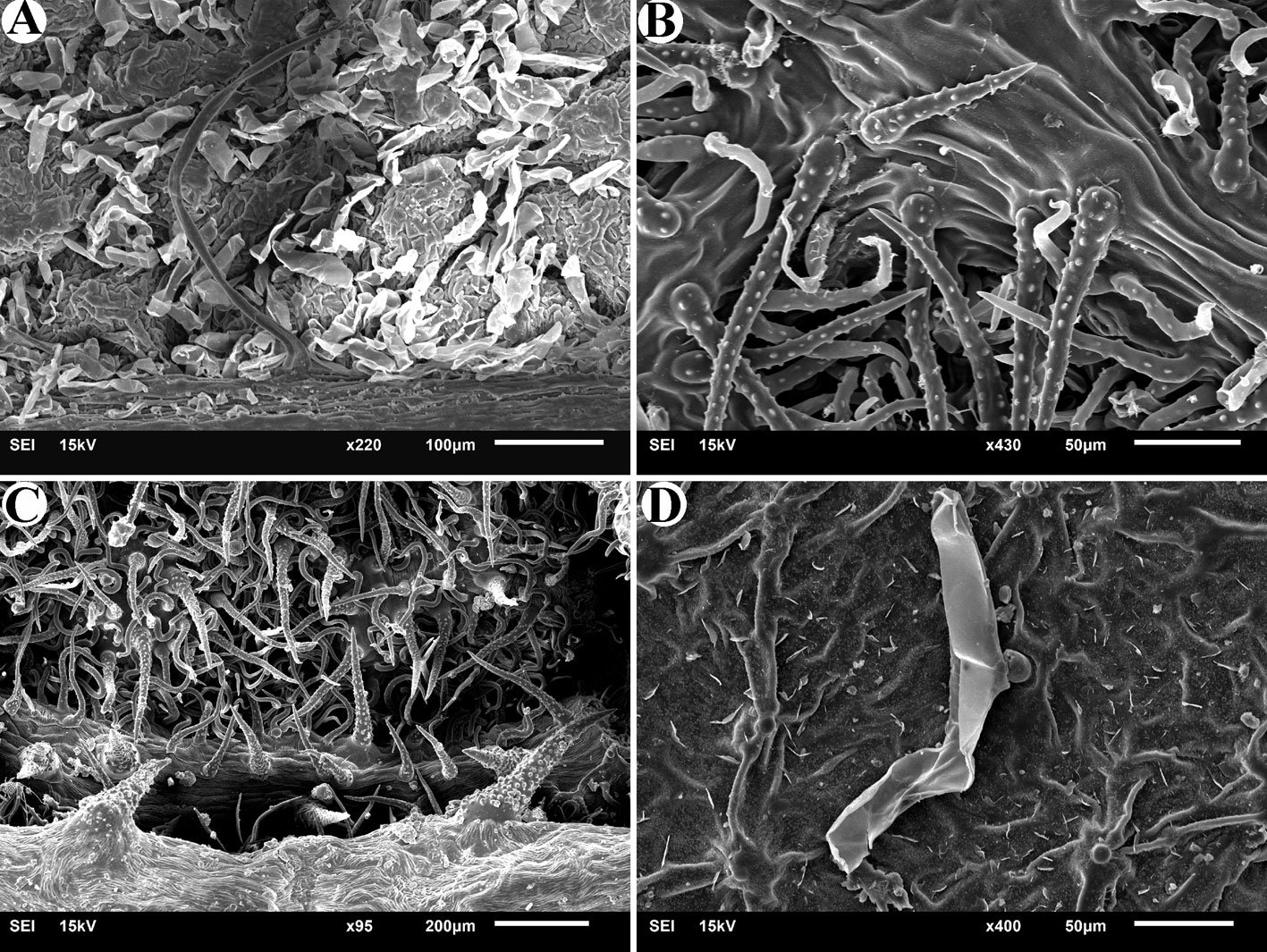
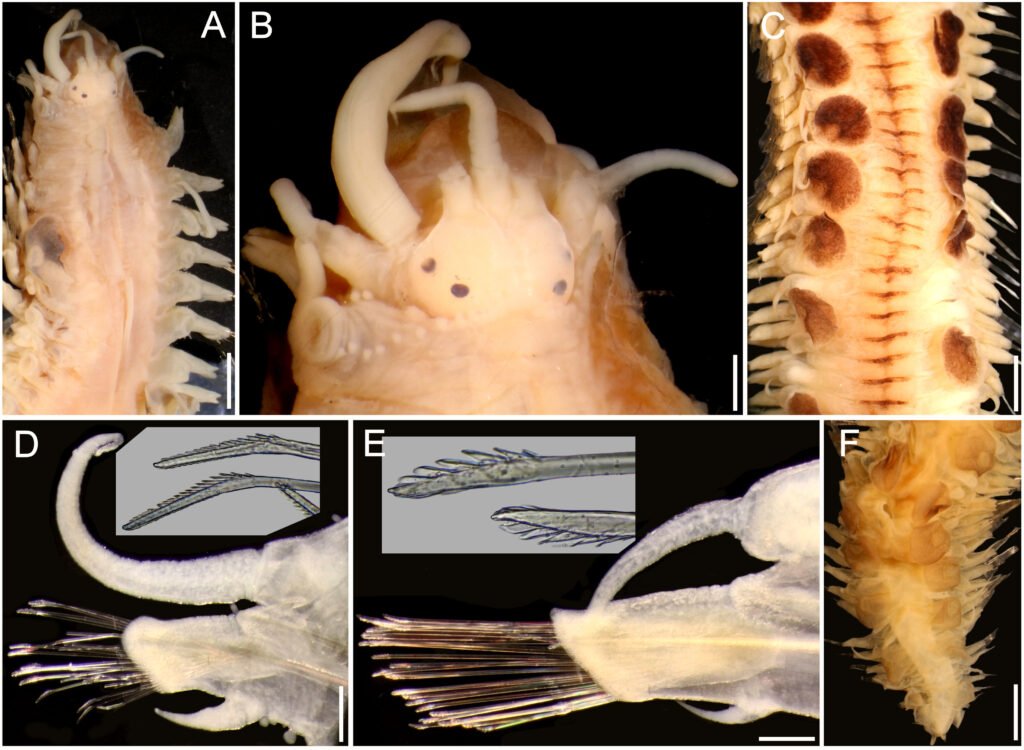
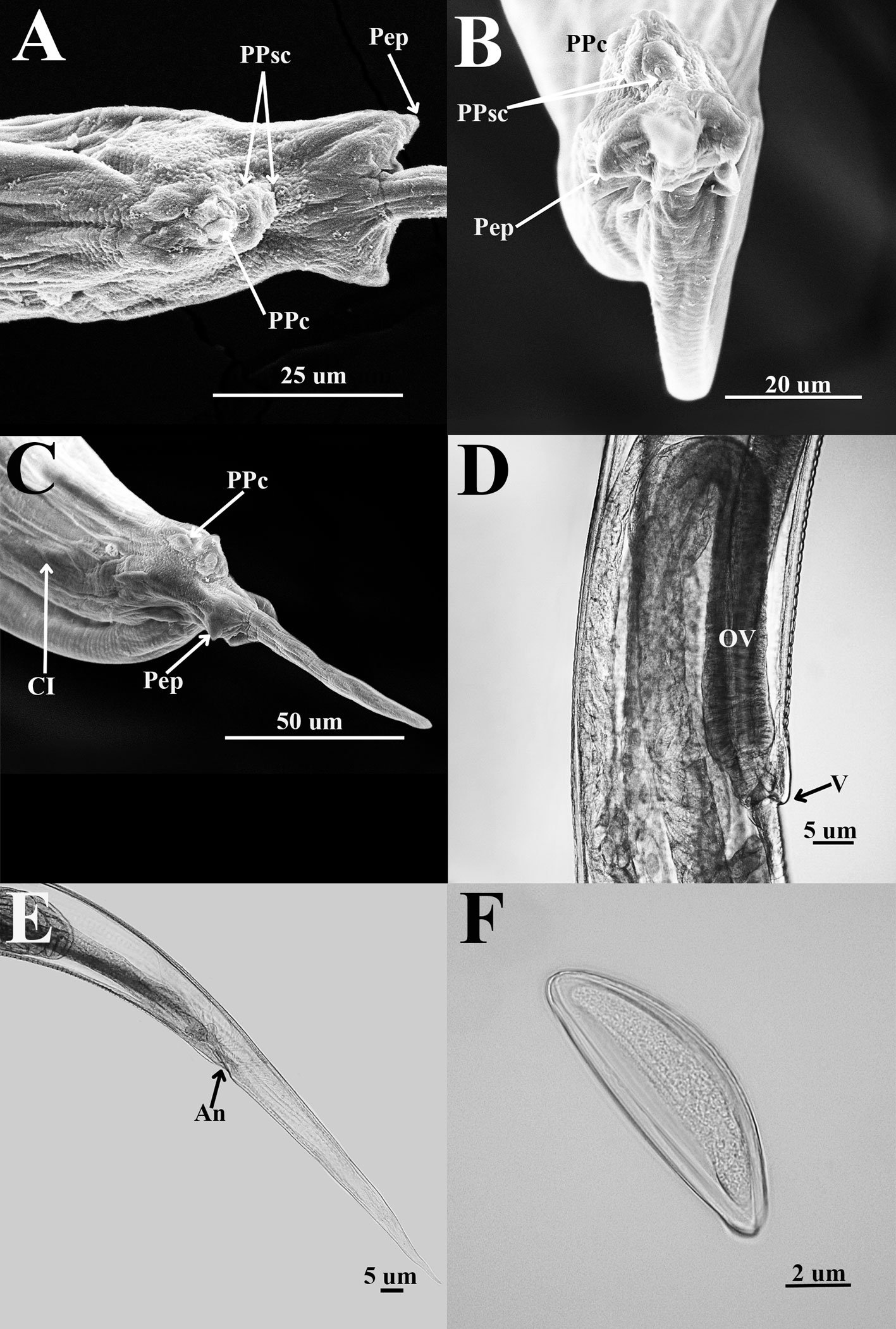
Figure 1. Heteromyoxyuris garciaprietoi n. sp., scanning electron micrographs. A, Male, apical view, amphids (AM), papillae (P), and tooth (T); B, female, apical view, interlabial processes (IP). Micrographs; C, male, apical view; D, lateral view of the cephalic region (CR), showing the esophagus (ES) and the esophageal bulb (EB); E, lateral view of the caudal region, showing cuticular inflation (CI), spicule simple (SP), gubernaculum (GB), and tail (Ta); F, line drawing of the caudal region, showing the same structures as in Figure (E), plus a pair of preanal papillae (PPc) and pedunculated papillae (Pep).
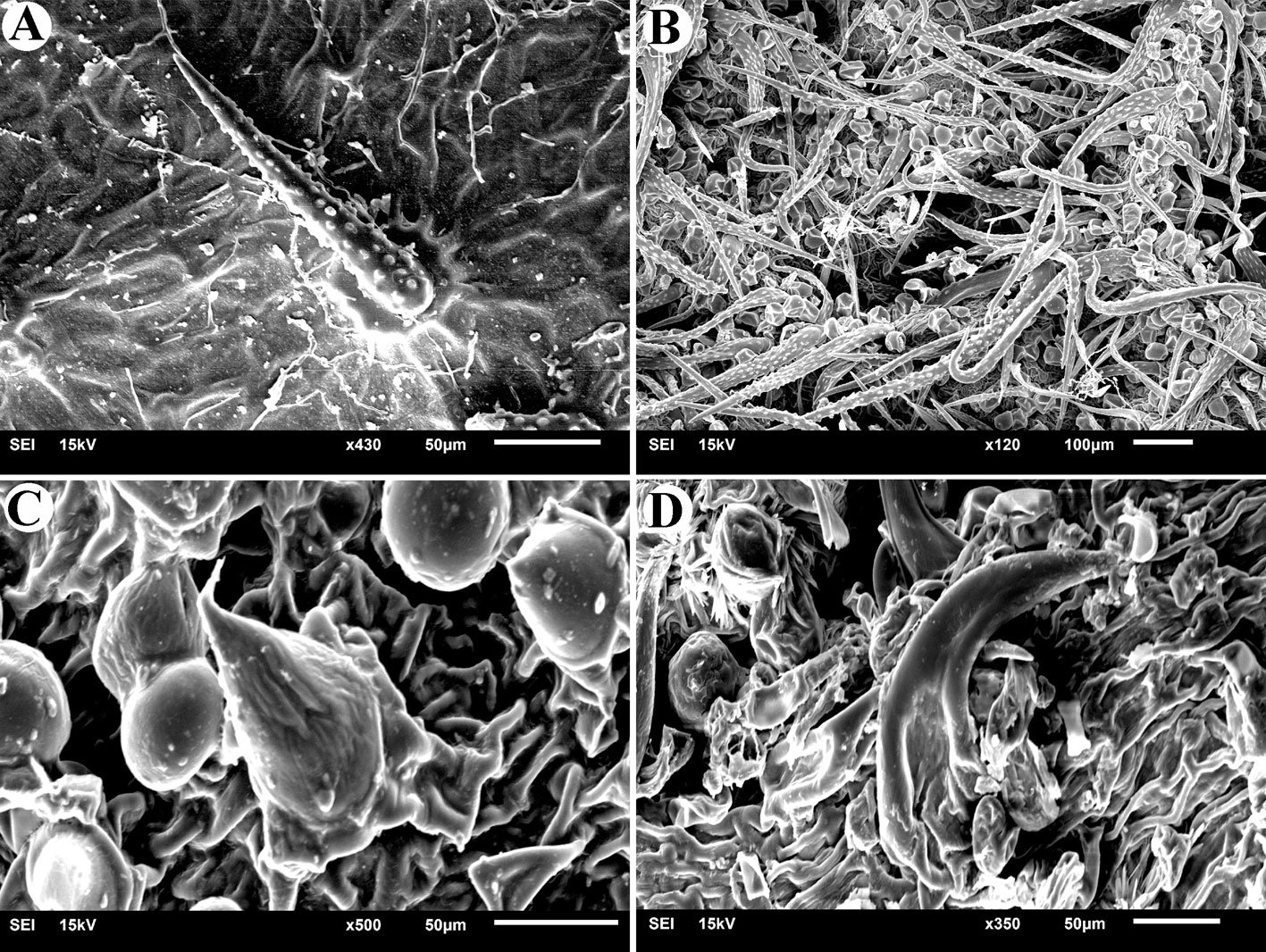
Male (based on holotype and 4 paratypes):body 2.9-3.7 (3.2±0.3) mm long and 202.5-222.5 (211±10.5) width at mid-body. Esophagus length 105-437.5 (351±139); esophageal bulb 95-110 (100.5±6.7) long and 92.5-120 (102.5±12) wide. Nerve ring and excretory pore located at 85-130 (108.5±17.2), and 490-567.5 (540.5±27), from anterior end, respectively. Spicule simple 75-82.5 (79.3±3.14) in length; gubernaculum 20-27.5 (24±3) long (Fig. 1E, F). Tail 127.5-137.5 (132±4.1) long; tail reduced to slender appendage 90-117.5 (100±16) in length. In the caudal region, the specimens exhibit a cuticular inflation 182.5-255 (207.5±17.5) long. Caudal sessile papillae arranged as follows: 1 preanal pair and 2 postanal pairs; posterior pair fused. One additional pair of pedunculated papillae located laterally at beginning of caudal filament containing phasmidia (Fig. 3A-C).
Female (based allotype and 15 paratypes):length 8.0-12.1 (11±1.4) mm, width 270-457.5 (386.2±0.7) at mid-body level. Esophagus length 590-680 (645±35.8); esophageal bulb 130-147.5 (139±7.7) long by 130-187.5 (158.1±20.6) wide. Nerve ring and excretory pore located at 125-207.5 (166.2±23), and 1.21-1.69 (1.4±0.1) mm from anterior end, respectively. Vulva opening at 3.1-5.3 (4.2±0.7) mm from anterior end. Ovejector 257.5-390 (338.7±44.7) long (Fig. 3D). Tail conical, 1.0-1.4 (1.1±0.1) mm long (Fig. 3E). Eggs without operculum, 97.5-112.5 (105.2±4.6) long by 30-40 36.3±3.2) wide (Fig. 3F).
Taxonomic summary
Type host: Dipodomys ordii (UACH-M152)
Type locality: 4.6 km S Ejido Mayran (25º37’48” N 102º50’5.1” W; 1,091 m asl), municipality of San Pedro de las Colonias, Coahuila, Mexico.
Site of infection: intestinal caecum.
Type specimens: holotype male (CNHE 12143); allotype female (CNHE 12144); paratypes: 8 males and 4 females (CNHE 12145).
Etymology: the new species was named in honor of MSc. Luis García-Prieto (IB-UNAM, Mexico) for his important and invaluable contribution to Mexican parasitology.
Other hosts and localities: Dipodomys merriami, 4.6 km S Ejido Mayran, municipality of San Pedro de las Colonias, Coahuila, Mexico (CNHE 12146); 3 km N San Luis de La Paz, Guanajuato, Mexico (CNHE: 9832).
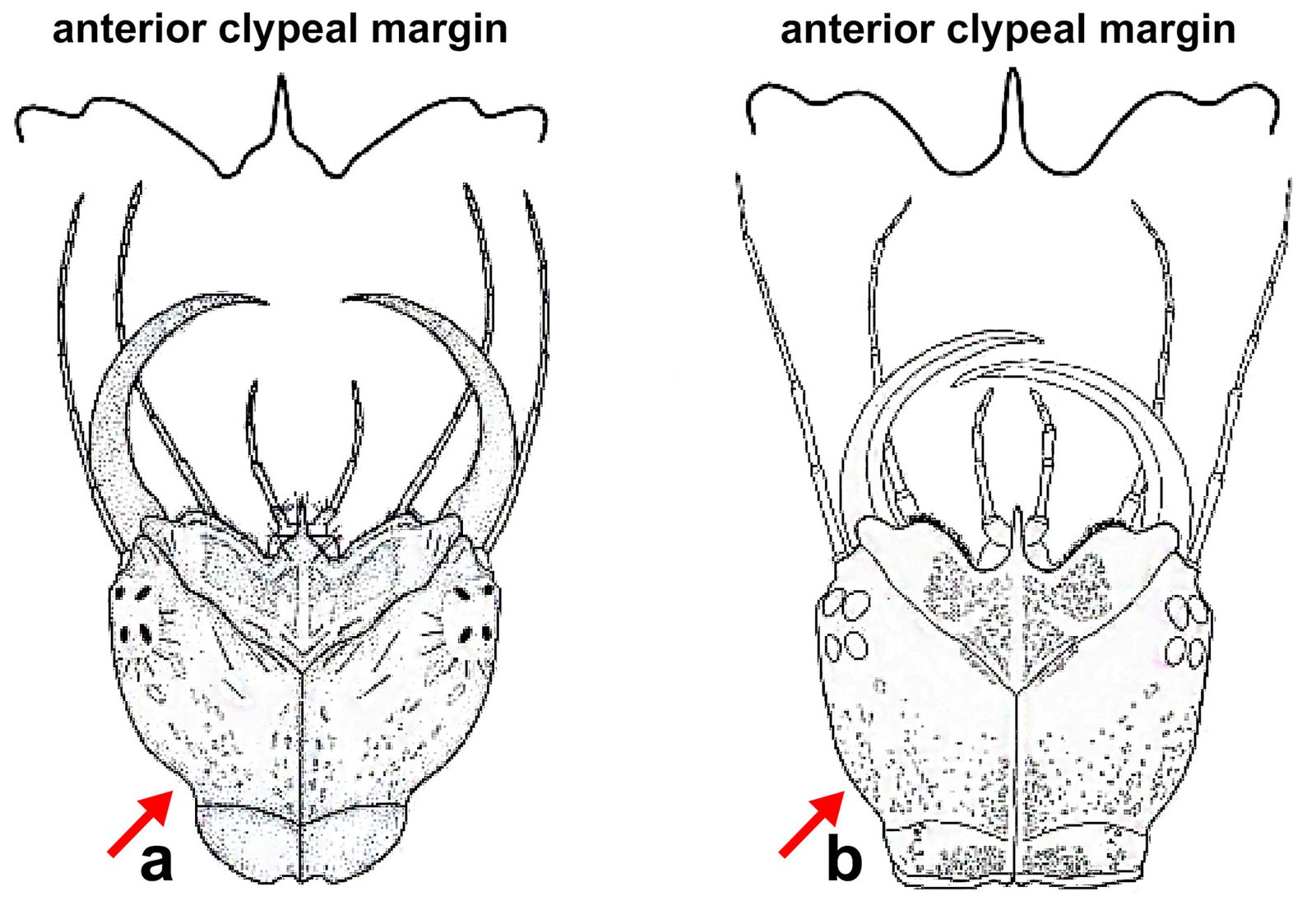
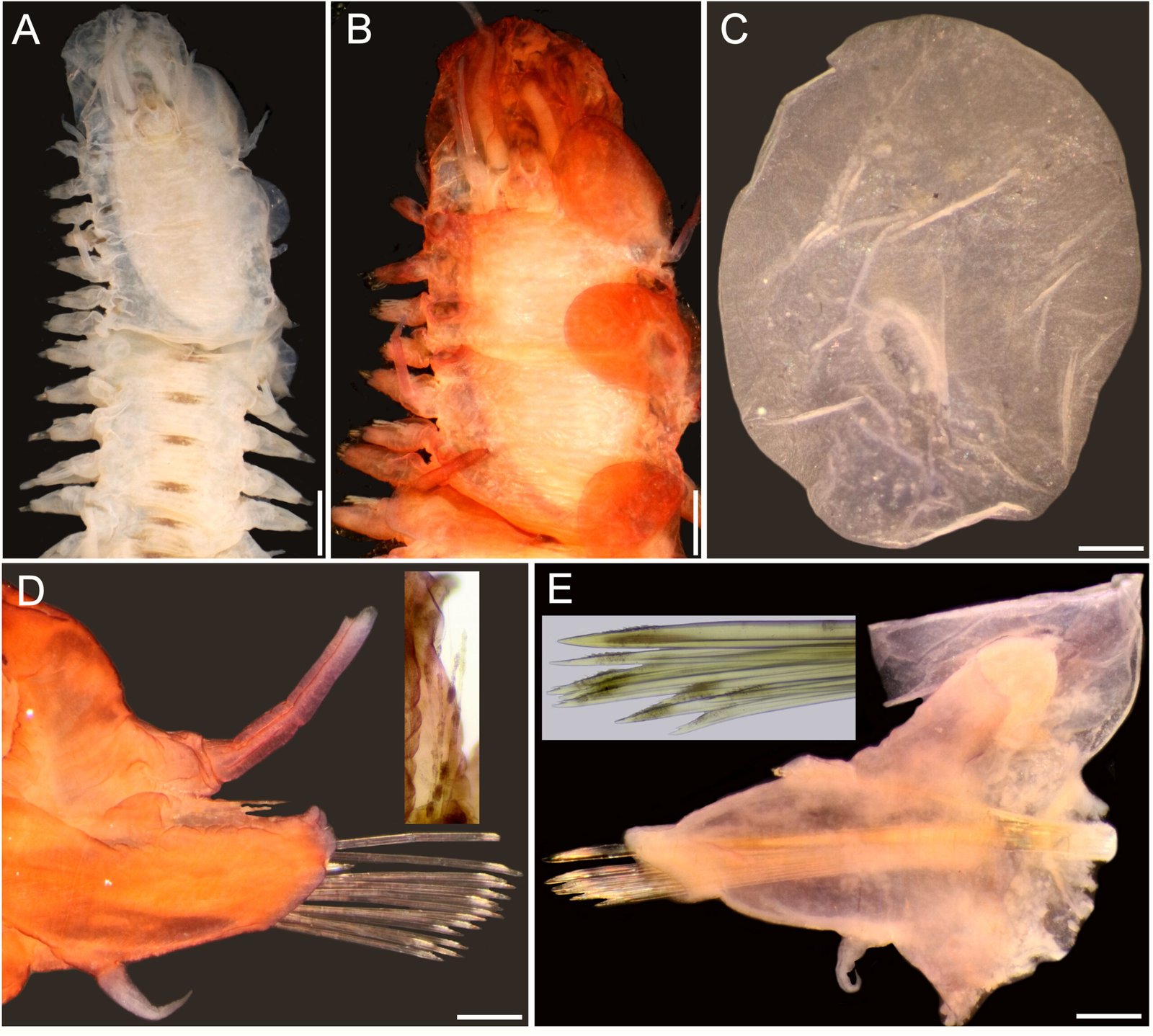
Remarks. Hereromyoxyuris was established by Quentin in 1973 and currently comprises 4 species: H. longejector, H. deserti, H. otomii (Quentin, 1973; García-Prieto et al., 2008), and H. garciaprietoi n. sp. The new species is characterized by the presence of 2 circles of 4 cephalic papillae (which are double in external circle), a mouth opening surrounded by 3 interlabia (1 ventral and 2 laterodorsals), containing 3 thick teeth, a strongly muscular vagina and nonembryonated eggs, lacking opercula. The new species is unique in that it is the only one with a smooth cuticle, not featuring lateral alae in worms of either sex. In contrast, H. deserti exhibits simple lateral alae located on both sides of the body. The males of both H. deserti and the new species feature caudal alae (caudal cuticular inflation) on the caudal region. These alae are more prominent in H. deserti (Fig. 4, in Quentin, 1973) than in the new species. In comparison to the new species, both H. longejector and H. otomii possess double alae on both sides of the body. The new species is found in 2 localities of the Mexican Plateau, where it parasitizes 2 species of the genus Dipodomys.
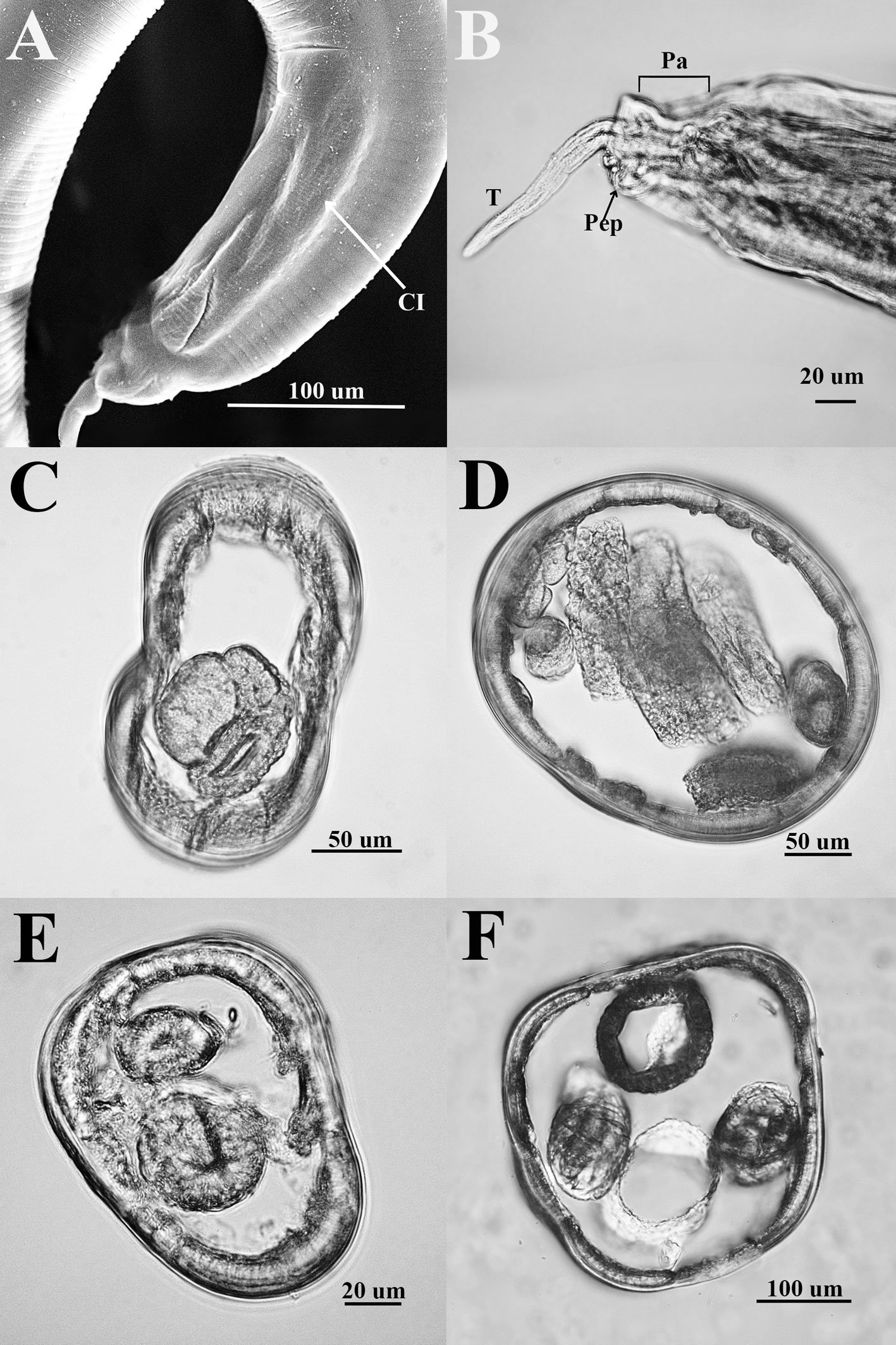
Also, the morphometry of specimens from Guanajuato and Coahuila is consistent with the same morphological characteristics (Table 2). Notably, the males present a caudal cuticular inflation (Fig. 4A); the arrangement of the papillae in the caudal region is the same as that described above (Fig. 4B). Both sexes lack lateral alae (Fig. 4 C-E).
The distribution of the genus Heteromyoxyuris coincides with the geographic distribution of the heteromyid rodent subfamilies Dipodomyinae (6 species) and Perognathinae (7 species) (Morrone, 2005; Omernik, 1987). Despite a record of H. longejector in Sylvilagus floridanus (J. A. Allen, 1890) (Lagomorpha: Leporidae] from Oklahoma, USA, this finding remains unverified due to the absence of a voucher specimen [therefore, we have excluded this data from our work; Boggs et al., 1990]).
Biogeographic studies of North American and Mexican deserts and drylands frequently reveal an east-west division of closely related species, with sister species often limited to the eastern (Chihuahuan) and western (Mojave and Sonoran) deserts (Ayoub & Riechert, 2004; Devitt, 2006; Douglas et al., 2006; Jaeger et al., 2005; Wilson & Pitts, 2010). The distribution pattern observed is presumably present in the Heteromyoxyuris host-parasite system. To confirm our hypothesis of a mirrored distribution pattern, initially we need to undertake phylogenetic analyses and exploratory studies in regions that match the host distribution.
In Mexico, Heteromyoxyuris has been found in Coahuila, Durango, Guanajuato, San Luis Potosí, and Zacatecas (Table 1). Most records of this nematode genus are associated with Heteromyidae, including hosts such as Chaetodipus eremicus (Mearns, 1898), Chaetodipus hispidus (Baird 1858), Chaetodipus sp., Dipodomys merriami, Dipodomys ordii, Perognathus amplus Osgood, 1900, and Perognathus flavus Baird, 1855. Only 1 record exists in Cricetidae, specifically in Peromyscus sp. Heteromyoxyuris deserti and H. longejector are of particular interest for biogeographic studies due to their wide distribution across the deserts and drylands of North America. The evidence suggests that both H. longejector and H. deserti are generalist parasites, infecting a wide range of hosts: 9 and 7 species, respectively (Table 1). It is noteworthy that H. deserti has been documented in Nevada, California, New Mexico, and Texas, USA, as well as in host species that are also distributed in Mexico. These include Dipodomys deserti Stephens, 1887, D. merriami, D. spectabilis Merriam, 1890, D. ordii, and P. flavus (Table 1). Therefore, it cannot be excluded that this species is also present in arid and semi-arid regions of Mexico.
Heteromyoxyuris otomii and H. garciaprietoi n. sp. are currently endemic to Mexico, but their distribution could be as wide as that of their hosts, which are not endemic to Mexico. Heteromyoxyuris otomii has only been recorded on P. flavus in Guanajuato and Hidalgo, Mexico (García-Prieto et al., 2008; Iturbe-Morgado et al., 2017). However, since its host is widespread across central Mexico and the southern USA, this species of nematode likely occurs in other regions in Mexico. Likewise, the broad distribution of D. merriami and D. ordii suggests that H. garciaprietoi n. sp. probably exists in other Mexican locations, similar to H. longejector (Table 1).
So far, there is only 1 study of the parasites of rodents in Coahuila. According to Falcón-Ordaz et al. (2024), only D. merriami has been examined for parasites in Coahuila, reporting Syphacia sp., Pterygodermatites dipodomis Tiner, 1948, and Protospirura dipodomis (Read & Millemann, 1953), and H. garciaprietoi n. sp. The diversity of parasitic helminths in rodents remains incomplete (García-Prieto et al., 2012; Falcón-Ordaz et al., 2024; Iturbe-Morgado et al., 2017; Martínez-Salazar et al., 2016), and the helminth fauna is still unknown in Campeche, Quintana Roo, Sinaloa, and Tamaulipas (González-Moreno & Falcón-Ordaz, 2025) or is under investigation, as in the Baja California Peninsula (Martínez-Salazar, Pers. Comm.).
Conservation efforts often overlook parasites, despite their ecological significance and potential as environmental indicators (Lymbery & Smit, 2023). Bridging this knowledge gap demands comprehensive updates to taxonomic inventories; this means examining not just living wildlife, but also the extensive collections of parasite and host material housed in biological repositories, since there are still species to be discovered (e.g., Museomics, see Fernández, 2019; Greiman et al., 2018). In addition, some areas are inaccessible for various reasons or due to the lack of support directed to this type of work (Martínez-Salazar et al., 2016). Undoubtedly, our understanding of host-parasite systems is incomplete, largely due to significant gaps in knowledge of species diversity (the Linnaean shortfall), their distribution (the Wallace shortfall), and their phylogenetic and evolutionary relationships (the Darwin shortfall) (Hortal et al., 2015; Rubio-Godoy & Pérez-Ponce de León, 2023). To reduce these limitations, we examined the material deposited at the CNHE (CNHE 9832), and subsequently, taxonomic analysis revealed that this material corresponds to H. garciaprietoi n. sp. The examination of museum specimens, as evidenced by the discovery of H. garciaprietoi n. sp. in CNHE material, highlights the vital role of biological collections in this effort.
In this study, it was not possible to extract DNA from the H. garciaprietoi n. sp. material due to the fixation condition of the organs examined (in formalin). Therefore, if this species is found in other regions of the country, it may offer a favorable panorama for exploring the phylogenetic relationships of the genus. Regarding this last point, it would be essential to confirm the close morphological relationship that the new species has with H. deserti. Heteromyoxyuris garciaprietoi n. sp. is the only species of the genus that does not have lateral alae in either sex. This is the fourth species described for the genus and the second described for Mexico.
Key to species of the genus Heteromyoxyuris Quentin, 1973
1a. Male and female with lateral alae on the body 2a
1b. Male and female without lateral alae on the body H. garciaprietoi n. sp.
2a. Male and female with simple lateral alae on the body H. deserti (Read and Millemann, 1953)
2b. Male and female with double lateral alae on the body 3a
3a. The lateral alae extend from esophageal bulb to anus in both sexes H. otomii García-Prieto, Falcón-Ordaz, Lira-Guerrero and Mendoza-Garfías, 2008
3b. The lateral alae in male begin at the posterior half of the body and in female start posterior to esophageal bulb ending in the anus in both sexes H. longejector (Hannum, 1943)
Acknowledgements
We are indebted to Berenit Mendoza-Garfias for her invaluable assistance in processing samples for the SEM photographs in the Laboratorio de Microscopía y Fotografía de la Biodiversidad I, at the LANABIO (IB-UNAM) and Luis Garcia-Prieto for kindly loaning specimens of the CNHE (IB-CNHE). We also want to thank Mitzi F. Aquino-Camacho for editing the figures. The treating results were obtained as part of the project “Sistemática y taxonomía de helmintos parásitos de vertebrados con enfásis en pequeños mamíferos” (number UAEH-DI-17-ICBI-BI-SF-107, J. Falcón-Ordaz, main researcher), and were partially written during the project UAZ-2023-38899 (Elizabeth A. Martínez-Salazar, main researcher). We are grateful to the Associate Editor and the anonymous reviewers for their valuable suggestions for improvements to the manuscript.
References
Alexander, L. F., & Riddle, B. R. (2005). Phylogenetics of the new world rodent family Heteromyidae. Journal of Mammalogy, 86, 366–379. https://doi.org/10.1644/BER-120.1
Ayoub, N. A., & Riechert S. E. (2004). Molecular evidence for Pleistocene glacial cycles driving diversification of a North American desert spider, Agelenopsis aperta. Molecular Ecology, 13, 3453–3465. https://doi.org/10.1111/j.1365-294X.2004.02335.x
Bienek, G. K., & L. G. Klikoff, L. G. (1974). Parasitological evidence of arthropods as food for Dipodomys merriami vulcani. American Midland Naturalist, 91, 251–253. https://doi.org/10.2307/2424421
Boggs, J. F., McMurry, S. T., Leslie, Jr. D. M., Engle, D. M., & Lochmiller, R. L. (1990). Influence of habitat modification on the intestinal helminth community ecology of cottontail rabbit populations. Journal of Wildlife Diseases, 26, 157–169. https://doi.org/10.7589/0090-3558-26.2.157
Decker, K. H., Duszynski, D. W., & Patrick, M. J. (2001). Biotic and abiotic effects on endoparasites infecting Dipodomys and Perognathus species. Journal of Parasitology, 87, 300–307. https://doi.org/10.1645/0022-3395(2001)087[0300:BAAEOE]2.0.CO;2
Devitt, T. J. (2006). Phylogeography of the Western Lyresnake (Trimorphodon biscutatus): testing arid land biogeographical hypotheses across the Nearctic-Neotropical transition. Molecular Ecology, 15, 4387–4407. https://doi.org/10.1111/j.1365-294X.2006.03015.x
Douglas, M. E., Douglas, M. R., Schuett, G. W., & Porras, L. W. (2006). Evolution of rattlesnakes (Viperidae: Crotalus) in the warm deserts of western North America shaped by Neogene vicariance and Quaternary climate change. Molecular Ecology, 15, 3353–3374. https://doi.org/10.1111/j.1365-294X.2006.03007.x
Falcón-Ordaz J., Aquino-Camacho, M. F., Ladrón de Guevara-Bárcenas, K., & Fernández, J. A. (2024). Helminths parasites of heteromyd rodents from semiarid regions of Mexico. Therya Notes, 5, 157–161. https://doi.org/10.12933/therya_notes-24-164
Fedynich, A. M., Monasmith, T., & Demarais, S. (2001). Helminth community structure and pattern in Merriam’s kangaroo rats, Dipodomys merriami Mearns, from the Chihuahuan Desert of New Mexico, U.S.A. Comparative Parasitology, 68, 116–121.
Fernández, J. A. (2019). The holistic specimen and parasites of mammals. Therya, 10, 65–67. https://doi.org/10.12933/therya-19-827
Fernández, J. A., Cervantes, F. A., & Hafner, M. S. (2012). Molecular systematics and biogeography of the Mexican endemic kangaroo rat, Dipocomys phillipsii (Rodentia: Heteromyidae). Journal of Mammalogy, 93, 560–571. https://doi.org/10.1644/11-MAMM-A-224.1
Fernández, J. A., Hafner, M. S., Hafner, D. J., & Cervantes, F. A. (2014). Conservation status of rodents of the families Geomyidae and Heteromyidae of Mexico. Revista Mexicana de Biodiversidad, 85, 576–588. https://doi.org/10.7550/rmb.36710
García-Prieto, L., Falcón-Ordaz, J., & Guzmán-Cornejo, C. (2012). Helminth parasites of wild Mexican mammals: list of species, host and geographical distribution. Zootaxa, 3290, 1–92. https://doi.org/10.11646/zootaxa.3290.1.1
García-Prieto, L., Falcón-Ordaz, J., Lira-Guerrero, G., & Mendoza-Garfias, B. (2008). A new species of Heteromyoxyuris (Nematoda: Oxyuridae), parasite of Perognathus flavus (Rodentia: Heteromyidae) from Mexico. Journal of Parasitology, 94, 860–865. https://doi.org/10.1645/GE-1452.1
Garner, H. W., Richardson, L. W., & Felts, L. A. (1976). Alimentary helminths of Dipodomys ordii: effects on the host population. The Southwestern Naturalist, 21, 327– 334. https://doi.org/10.2307/3669718
González-Moreno, C. D., & Falcón-Ordaz, J. (2025). Análisis de la riqueza de helmintos en roedores de México. Pädi Boletín Científico de Ciencias Básicas e Ingenierías del ICBI, 12, 22–27. https://doi.org/10.29057/icbi.v12i24.12640
Greiman, S. E., Cook, J. A., Tkach, V. V., Hoberg, E. P., Menning, D. M., Hope, A. G. et al. (2018). Museum metabarcoding: a novel method revealing gut helminth communities of small mammals across space and time. International Journal for Parasitology, 48, 1061–1070. https://doi.org/10.1016/j.ijpara.2018.08.001
Hannum, C. A. (1941). Nematode parasites of Arizona vertebrates (Ph.D. Thesis). University of Washington, Seattle.
Hortal, J., Bello, F., Diniz-Filho, J. A., Lewinsohn, T. M., Lobo, J. M., & Ladle, R. J. (2015). Seven shortfalls that beset large-scale knowledge of biodiversity. Annual Review of Ecology, Evolution, and Systematics, 46, 523–549. https://doi.org/10.1146/annurev-ecolsys-112414-054400
Iturbe-Morgado, J. C., Falcón-Ordaz, J., Lira-Guerrero, G., Fernández, J. A., & Acosta, R. (2017). Nematofauna of rodents of the families Heteromyidae and Cricetidae from the Mexican Plateau. Journal of Parasitology, 103, 127–131. https://doi.org/10.1645/16-105
Jaeger, J. R., Riddle, B. R., & Bradford, D. F. (2005). Cryptic Neogene vicariance and Quaternary dispersal of the red-spotted toad (Bufo punctatus): insights on the evolution of North American warm desert biotas. Molecular Ecology, 14, 3033–3048. https://doi.org/10.1111/j.1365-294X.2005.02645.x
Kelt, D. A., Hafner, M. S., & The American Society of Mammalogists’ ad hoc Committee for Guidelines on Handling Rodents in the Field. (2010). Updated guidelines for protection of mammalogists and wildlife researchers from hantavirus pulmonary syndrome (hps). Journal of Mammalogy, 91, 1524–1527. https://doi.org/10.1644/10-MAMM-A-306.1
King, S. R., & Babero, B. B. (1974). Helminths of kangaroo rats (Dipodomys spp.) in Nevada with reports of other worm parasites from these hosts. Proceedings of the Helminthological Society of Washington, 41, 241–248.
Lymbery, A. J., & Smit, N. J. (2023). Conservation of parasites: a primer. International Journal for Parasitology: Parasites and Wildlife, 21, 255–263. https://doi.org/10.1016/j.ijppaw.2023.07.001
Martínez-Salazar, E. A., Flores-Rodríguez V., Rosas-Valdez, R., & Falcón-Ordaz, J. (2016). Helminth parasites of some rodents (Cricetidae, Heteromyidae, and Sciuridae) from Zacatecas, Mexico. Revista Mexicana de Biodiversidad, 87, 1203–1211. https://doi.org/10.1016/j.rmb.2016.10.009
Morrone, J. J. (2005). Hacia una síntesis biogeográfica de México. Revista Mexicana de Biodiversidad, 76, 207–252. http://dx.doi.org/10.22201/ib.20078706e.2005.002.303
Omernik, J. M. (1987). Ecoregions of the Conterminous United States. Annals of the Association of American Geographers, 77, 118–125. https://doi.org/10.1111/j.1467-8306.1987.tb00149.x
Preisser, W. C., & Falcón-Ordaz, J. (2019). A checklist of the parasitic helminths of cricetid and heteromyid rodents in Mexico. Therya, 10, 329–341. http://dx.doi.org/10.12933/therya-19-787
Quentin, J. C. (1973). Les Oxyurinae de Rongeurs. Bulletin du Muse ́e Nationalle d’Histoire Naturelle, 12, 1045–1096.
Read, C. P., & Millemann, R. E. (1953). Helminth parasites in kangaroo rats. University of California Publications in Zoology, 59, 61‒80.
Rubio-Godoy, M., & Pérez-Ponce de León, G. (2023). Equal rights for parasites: Windsor 1995, revisited after ecological parasitology has come of age. Biological Conservation, 284, 110174. https://doi.org/10.1016/j.biocon.2023.110174
Sikes, R. S., & The Animal Care and Use Committee of the American Society of Mammalogists. (2016). Guidelines of The American Society of Mammalogists for the use of wild mammals in research and education. Journal of Mammalogy, 97, 663–688. https://doi.org/10.1093/jmammal/gyw078
Wilson, J. S., & Pitts, J. P. (2010). Phylogeographic analysis of the nocturnal velvet ant genus Dilophotopsis (Hymenoptera: Mutillidae) provides insights into diversification in the Nearctic deserts. Biological Journal of the Linnean Society, 101, 360–375. https://doi.org/10.1111/j.1095-8312.2010.01526.x