In her 1945 study on the Tertiary fossil mollusks of northeastern Mexico, Gardner listed only 2 taxa of land snails: Holospira eva Gardner, 1945 and “Helix” sp. The latter is recognized here as a new species of Lysinoe Adams & Adams, 1855 (Helicoidea, Xanthonychidae). The new species can be diagnosed from its congeners by its flattened shell profile, a lower body whorl with a faint angulation on its median portion, a ridge-like angulation around the umbilicus, and a likely wide umbilicus. The species is known to occur in 2 nearby Upper Middle Oligocene outcrops in the eastern part of the state of Nuevo León.
Keywords: Mollusca; Land snail; Fossil; Nuevo León; Stylommatophora; Xanthonychidae
Una especie nueva de Lysinoe (Gastropoda: Helicoidea) del Oligoceno de México
Resumen
En su estudio de 1945 sobre los moluscos fósiles del Terciario del noreste de México, Gardner enumeró solo 2 taxones de caracoles terrestres: Holospira eva Gardner, 1945 y “Helix” sp. Esta última se reconoce aquí como una especie nueva de Lysinoe Adams et Adams, 1855 (Helicoidea, Xanthonychidae). La nueva especie se puede diagnosticar a partir de sus congéneres por el perfil aplanado de su concha, la vuelta del cuerpo es más baja y con una leve angulación en su porción media, una angulación en forma de cresta alrededor del ombligo y un ombligo probablemente ancho. La especie se encuentra en 2 afloramientos cercanos del Oligoceno Medio Superior en la parte oriental del estado de Nuevo León.
Palabras clave: Mollusca; Caracol terrestre; Fósil; Nuevo León; Stylommatophora; Xanthonychidae
Introduction
In 1945, Julia Gardner published an important paper describing the Tertiary fossil molluscan fauna of northeastern Mexico (Gardner, 1945), with information and specimens collected by the United States Geological Survey (USGS). Virtually all fossils were marine or freshwater, with 2 exceptions: the land snail taxa Holospira eva Gardner, 1945 (Urocoptidae) and “Helix” sp.
Gardner (1945) assigned the fossils the broad identity of “Helix” sp. to signify they were helicoid snails (superfamily Helicoidea). Still, she alluded to the possibility that those fossils were closely related to the Mexican and Central American genus Lysinoe Adams & Adams, 1855 (Helicoidea, Helminthoglyptidae). Later, Roth (1984) described a new species, Lysinoe breedlovei Roth, 1984, from the Vieja Group (Upper Eocene; Texas, USA) and included Gardner’s “Helix” sp. in that species. It is uncertain why Roth (1984) did that, as the morphological differences between his new species and Gardner’s “Helix” sp. are quite striking. Furthermore, the discrepancy in age between the 2, alongside the large geographical distance, would also have been good indicators that they were 2 separate species.
Herein, Gardner’s “Helix” sp. is recognized as a distinct species belonging to the genus Lysinoe and described as new.
Materials and methods
The single specimen representative of the new species is housed in the Paleobiology Collection of the National Museum of Natural History (NMNH, Smithsonian Institution, Washington D.C., USA) under the registration number USNM MO 497132. A historical background and further details regarding the USGS collection efforts in Mexico can be found in Gardner (1945).
Description
Family Xanthonychidae Strebel & Pfeffer, 1880
Genus Lysinoe Adams & Adams, 1855 (type species Helix ghiesbreghti Nyst, 1841)
Shell large (height 23.0 mm, width 41.3 mm), helicoid, of ca. 4 whorls, with a low spire. Whorls growing regularly in size, but final ½ whorl suddenly expands towards aperture. Body whorl slightly flattened, with faint angulation on its median portion and light ridge-like angulation around the umbilicus. Aperture seemingly rounded, prosocline. Umbilicus seemingly deep and relatively wide. The fossil is an internal mold, so not all shell structures can be observed; features such as the umbilicus’ width might be misleading.
Even though Gardner (1945: 18, 267) alludes to further material, no specimens beyond the holotype (Gardner’s figured specimens) were available for study at the NMNH. According to that author’s remarks, a few additional morphological features could be gleaned from the specimens then available (Gardner, 1945: 267): the number of whorls could grow a little beyond 4; shell moderately heavy (observed in fragments of shell adhered to some of the molds); the body whorl angulation in juvenile shells was more pronounced than in the adults.
Taxonomic summary
Type locality: Mexico, Nuevo León, General Bravo, USGS station 13517: “[a]t and just west of Rancho Paulino Ríos” (Gardner, 1945: 23, quadrant N-15 on map).
Type material: USNM MO 497132 (holotype).
Etymology: the specific epithet refers to the type locality.
Type stratum: base of Upper Middle Oligocene sandstone, described as a “hard conglomeratic, shaly, locally silicious sandstone” (Gardner, 1945: 18).
Age: Middle Oligocene.
Distribution: also known from nearby USGS station 14023: Nuevo León, General Bravo, Zacate, “[o]n a high hill 3,750 meters S. 68° W. from Zacate well No. 1” (Gardner, 1945: 32, quadrant N-13 on map).
Diagnosis: Shell large, helicoid with a flattened profile. Body whorl low, with faint median angulation. Ridge-like angulation around umbilicus. Umbilicus likely wide.
Previous mentions in the literature: “Helix” sp.: Gardner, 1945: 267, pl. 18, figs. 1-3. Helix s. l. near Lysinoe sp.: Gardner, 1945: 18. Lysinoe breedlovei [in part]: Roth, 1984: 203.
Remarks
Roth (1984, 1988) included Lysinoe and related forms in Helminthoglyptidae (Sei et al., 2017), while current classification (MolluscaBase, 2023; Thompson, 2011) places it in Xanthonychidae instead. That interpretation of Xanthonychidae was shown to be polyphyletic, with branches scattered throughout Helicoidea (Calcutt et al., 2020; Cuezzo, 1998; Zhang et al., 2024). The classification of Lysinoe in Xanthonychidae is maintained here, but it is noted that this could potentially change with new phylogenetic studies.
Considering the overall shell shape, the present fossil is consistent with helminthoglyptid snails, in particular with Lysinoe spp., which include animals with large helicoid shells with similar features, including the wide umbilicus and the periumbilical angulation (Schileyko, 2004; Zilch, 1960). Species in related genera, e.g., Leptarionta Fischer & Crosse, 1872 and Helminthoglypta Ancey, 1887, have smaller shells (Schileyko, 2004; Zilch, 1960). The shells of Helminthoglypta spp. are also much more globular. Leptarionta spp. often display a median angulation on the body whorl, but the shells typically have higher spires and a more conical profile.
Figure 1. Lysinoe bravoensis sp. nov., holotype USNM MO 497132. A, Apertural view; B, lateral view; C, apical view; D, umbilical view. Scale bar = 2 cm.
The holotype and only specimen available is an internal mold of the shell, so interpretations of the species systematics must be done with caution. While the classification in Lysinoe seems the best solution at the moment, it must remain tentative until better-preserved fossils are found. There are minor morphological differences observed in the fossil (more flattened shell, light median angulation of body whorl; Fig. 1) that are not seen in recent representatives of Lysinoe: L. eximia (Pfeiffer, 1844), L. ghiesbreghti (Nyst, 1841), L. sebastiana (Dall, 1897), and L. starretti Thompson, 1963. Those differences, alongside the old age of the fossil, could indicate that this species belongs to a still unrecognized and extinct genus. Nevertheless, only the discovery of additional material (both, of the new species and L. breedlovei), in a better state of preservation, can shed more light on this matter.
Therefore, the same shell features mentioned above (i.e., more flattened shell, light median angulation of body whorl) can be used to diagnose Lysinoe bravoensis sp. nov. from its extant congeners. Importantly, the more flattened shell, lower spire, the median angulation on the body whorl, the ridge-like angulation surrounding the umbilicus, and the larger umbilicus (Fig. 1) diagnose Lysinoe bravoensis sp. nov. from Roth’s Lysinoe breedlovei.
Discussion
Living Lysinoe spp. are distributed from central and southern Mexico to El Salvador and Honduras (Schileyko, 2004; Thompson, 2011; Zilch, 1960). As such, an occurrence in the Oligocene of northern Mexico is outside the genus’s current distribution but hardly surprising, particularly considering that Lysinoe breedlovei is from the Late Eocene of Texas, close to the border with Mexico (Roth, 1984).
Still, Lysinoe bravoensis sp. nov. represents the first fossil record of the genus in Mexico. The oldest record of the genus (and of other closely-related xanthonychid genera) is Lysinoe breedlovei. The superfamily Helicoidea, to which they belong, originated in North America, with its oldest records (family Labyrinthidae) dating from the Paleocene and Eocene (Calcutt et al., 2020; Roth, 1988).
Gardner (1945: 18) noted that in USGS station 14023, Lysinoe bravoensis sp. nov. was part of an assemblage with marine mollusks (both gastropods and bivalves) and the freshwater snail Hemisinusmiralejas Gardner, 1945. Thus, Gardner (1945) suggested an estuarine paleoenvironment for the area, such as the mouth of a stream where sea shells could have accumulated alongside freshwater and terrestrial snail shells. Further, fossil Hemisinus spp. are also common in other deltaic deposits of northeastern Mexico (e.g., Vega & Perrilliat, 1992), supporting Gadner’s (1945) interpretation. At the type locality USGS station 13517, no accompanying fauna was found (Gardner, 1945); however, considering the sedimentological similarity between the 2 stations (Gardner, 1945), a similar paleoenvironment could be expected.
An actualistic genus-level approach can be used to hypothesize further paleoenvironmental aspects (Rasser et al., 2019), though this must be done with caution considering the relatively old age of the fossil and its uncertain generic affinities. Based on the current distribution and habitats of living Lysinoe spp., a humid forest environment would have been expected for the region in the past (cf. Roth, 1984), contrary to today’s xeric shrublands of the Tamaulipan mezquital.
Acknowledgements
I am very grateful to Mark Florence and Jessica Nakano (NMNH) for the loan of the material; to Erik Kjellman and Andreas Altenburger (UiT) for the help with the photographs of the specimen; and to the two anonymous reviewers and the editor, Xochitl Vital, for their helpful comments.
References
Calcutt, J., Cuezzo, M. G., Jackson, M., & Salvador, R. B. (2020). Phylogenetic relationships and classification of Solaropsidae (Gastropoda: Stylommatophora). Archiv für Molluskenkunde, 149, 181–193. https://doi.org/10.1127/arch.moll/149/181-193
Cuezzo, M. G. (1998). Cladistic analysis of the Xanthonychidae (=Helminthoglyptidae) (Gastropoda: Stylommatophora: Helicoidea). Malacologia, 39, 93–111.
Gardner, J. (1945). Mollusca of the Tertiary formations of northeastern Mexico. Geological Society of America Memoir, 11, 1–332. https://doi.org/10.1130/MEM11-p1
Rasser, M. W., Höltke, O., & Salvador, R. B. (2019). Gastropod paleohabitats of Miocene Lake Randeck Maar and its hinterland defined by an actualistic genus-level approach. Lethaia, 53, 229–241. https://doi.org/10.1111/let.12353
Roth, B. (1984). Lysinoe (Gastropoda: Pulmonata) and other land snails from Eocene-Oligocene of Trans-Pecos Texas, and their paleoclimatic significance. The Veliger, 27, 200–218.
Roth, B. (1988). Camaenid land snails (Gastropoda: Pulmonata) from the Eocene of southern California and their bearing on the history of the American Camaenidae. Transactions of the San Diego Society of Natural History, 21, 203–220.
Schileyko, A. A. (2004). Treatise on recent terrestrial pulmonate molluscs. Part 12. Bradybaenidae, Monadeniidae, Xan- thonychidae, Epiphragmophoridae, Helminthoglyptidae, Elonidae, Humboldtianidae, Sphincterochilidae, Cochli- cellidae. Ruthenica, 2 (Suppl.), 1627–1763.
Sei, M., Robinson, D. G., Geneva, A. J., & Rosenberg, G. (2017). Doubled helix: Sagdoidea is the overlooked sister group of Helicoidea (Mollusca: Gastropoda: Pulmonata). Biological Journal of the Linnean Society, 122, 697–728. https://doi.org/10.1093/biolinnean/blx082
Thompson, F. G. (2011). An annotated checklist and bibliography of the land and freshwater snails of México and Central America. Florida Museum of Natural History Bulletin, 50, 1–299. https://doi.org/10.58782/flmnh.bnej7351
Vega, F. J., & Perrilliat, M. C. (1992). Freshwater gastropods from Early Eocene Difunta Group, Northeastern Mexico. Journal of Paleontology, 66, 603–609. https://doi.org/10. 1017/S002233600002446X
Zhang, G., Naggs, F., Andrus, P. S., & Wade, C.M. (2024). Phylogenetic insights into the terrestrial snails Helicoidei (Gastropoda: Stylommatophora) with special emphasis on the Camaenidae. Zoological Journal of the Linnean Society, 200, 1–13. https://doi.org/10.1093/zoolinnean/zlae027
Zilch, Α. (1960). Euthyneura. In O. H. Schindewolf (Ed.), Handbuch der Paläozoologie (pp. 1–835). Berlin: Gebrüder Borntraeger.
Jesús García-Jiménez a, Olivia Ayala-Vásquez a, b, Javier Isaac de la Fuente a, b, Roberto Garibay-Orijel c, Fortunato Garza-Ocañas d, Edgardo Ulises Esquivel-Naranjo e, Felipe Manuel Ferrusca-Rico e, Fidel Landeros e, *
a Tecnológico Nacional de México, Instituto Tecnológico de Ciudad Victoria, Boulevard Emilio Portes Gil #1301, 87010 Ciudad Victoria, Tamaulipas, Mexico
b Colegio de Postgraduados, Microbiología, Edafología, Km 36.5 Carretera México-Texcoco, Montecillo, 56230 Texcoco, Estado de México, Mexico
c Universidad Nacional Autónoma de México, Instituto de Biología, Circuito exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
d Universidad Autónoma de Nuevo León, Campus Linares, Facultad de Ciencias Forestales, Carretera Nacional Km 145, 67700 Linares, Nuevo León, Mexico
e Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales, Carretera a Chichimequillas s/n, 76140 Santiago de Querétaro, Querétaro, Mexico
Cyanoboletus abieticola is described as a new species to science. This species is characterized by the viscid pileus, cylindric to bacilliform basidiospores ([14.1] 16.2 ± 1.1 [17.5] × [5] 5.5 ± 0.3 [5.9] µm, Q [2.8] 3.0 ± 0.1 [3.2]), and its gregarious habit in mixed coniferous forests dominated by Abies religiosa or A. guatemalensis. Photographs, drawings, and the phylogenetic analysis of 3 genetic data sets (ITS, nucLSU, and RPB2) of the new species are presented.
Cyanoboletus abieticola (Boletaceae, Basidiomycota), una especie nueva de México
Resumen
Cyanoboletus abieticola es descrita como una especie nueva para la ciencia. Esta especie se caracteriza por el píleo víscido, basidiosporas cilíndricas a baciliformes ([14.1] 16.2 ± 1.1 [17.5] × [5] 5.5 ± 0.3 [5.9] µm, Q [2.8] 3.0 ± 0.1 [3.2]) y su hábito gregario en bosques mixtos de coníferas dominados por Abies religiosa o A. guatemalensis. Se presentan fotografías, dibujos y el análisis filogenético de 3 conjuntos de datos genéticos (ITS, nucLSU y RPB2) de la nueva especie.
The family Boletaceae comprises pileate-stipitate or sequestrate species, with poroid or lamellate hymenophore. Boletaceae species are an important component of ecosystems because most of them form ectomycorrhizal associations with a great diversity of angiosperms and gymnosperms (Binder & Hibbet, 2006; Ortíz-Santana et al., 2007). They are common in different vegetation types, such as deciduous, conifer, and mixed forest; mediterranean shrublands, tropical and montane cloud forests (Bessette et al., 2010; Gelardi, 2020; Leonardi et al., 2020; Wu et al., 2016). In addition to their ecological importance, many species are valued edibles in several regions of the world (Garibay-Orijel et al., 2009; Gelardi, 2020).
Wu et al. (2014) recognized 7 major clades at the subfamily level and 59 genus-level clades, placing Boletus pulverulentus within the “Pulveroboletus group”, which also includes the genera Butyriboletus D. Arora & J.L. Frank, Cacaoporus Raspé & Vadthanarat, Caloboletus Vizzini, Crocinoboletus N.K. Zeng, Zhu L. Yang & G. Wu, Gymnogaster J.W. Cribb, Lanmaoa G. Wu & Zhu L. Yang, Pulveroboletus Murrill, Rubroboletus Kuan, Rugiboletus G. Wu & Zhu L. Yang, Suillellus Murrill, and Sutorius Halling, Nuhn & N.A. Fechner. Members of the “Pulveroboletus group” share certain common characteristics such as poroid hymenium, usually yellowish, reddish or brown hymenophore, Boletus-type hymenophoral trama (Snell & Dick, 1958), and smooth spores.
Gelardi et al. (2014) proposed the new genus Cyanoboletus, within the “Pulveroboletus group”, for those species characterized mainly by their strong blue reaction of basidiomata and context to cutting. They included 3 species in this genus: C. pulverulentus (Opat.) Gelardi, Vizzini & Simonini, C. sinopulverulentus (Gelardi & Vizzini) Gelardi, Vizzini & Simonini, and C. rainisiae (Bessette & O.K. Mill.) Gelardi, Vizzini & Simonini. The latter one was transferred to the genus Xerocomellus, but without type studies, and is currently known as X. rainisiae (Bessette & O.K. Mill.) N. Siegel, C.F. Schwarz & J.L. Frank (see discussion in Frank et al. [2020]).
Later, 4 more species were transferred to this genus: C. cyaneitinctus (Murrill) A. Farid, A.R. Franck & J.A. Bolin (Farid et al., 2021), C. instabilis (W. F. Chiu) G. Wu & Zhu L. Yang (Wu et al., 2016), C. poikilochromus (Pöder, Cetto & Zuccherelli) M. Carbone, D. Puddu & P. Alvarado (Carbone et al., 2023), and C. flavosanguineus (Lavorato & Simonini) Pierotti (Pierotti, 2015). However, the latter species was placed in the genus Neoboletus in 2021: N. flavosanguineus (Lavorato & Simonini) Biketova, Wasser, Simonini & Gelardi (Biketova et al., 2021).
Additionally, 5 Cyanoboletus species have been recently described: C. brunneoruber G. Wu & Zhu L. Yang from China (Wu et al., 2016), C. hymenoglutinosus D. Chakr., K. Das, A. Baghela, S.K. Singh & Dentinger nom. inval. from India (Li et al., 2016), C. bessettei A.R. Bessette, L.V. Kudzma, & A. Farid from the USA (Farid et al., 2021), C. macroporus Sarwar, Naseer & Khalid from Pakistan (Sarwar et al., 2021), and C. mediterraneensis Biketova, A. Rinaldi & Simonini from Israel and Italy (Biketova et al., 2016, 2022). Therefore, 10 species are currently known for the genus. These species are distributed mainly in Asia, Europe and North America and associated with Abies, Arbutus, Buxus, Carya, Castanopsis, Cistus, Crataegus, Cryptomeria, Erica, Juniperus, Lithocarpus, Ostrya, Pinus, Pistacea,and Quercus trees, as well as with Halimium shrub (Biketova et al., 2022; Farid et al., 2021; García et al., 1998; Li et al., 2016; Wu et al., 2016).
This paper describes Cyanoboletus abieticola as a new species from Mexico associated with mixed conifer forests dominated by Abies religiosa (Kunth) Schltdl. & Cham. and A. guatemalensis Rehder.
Materials and methods
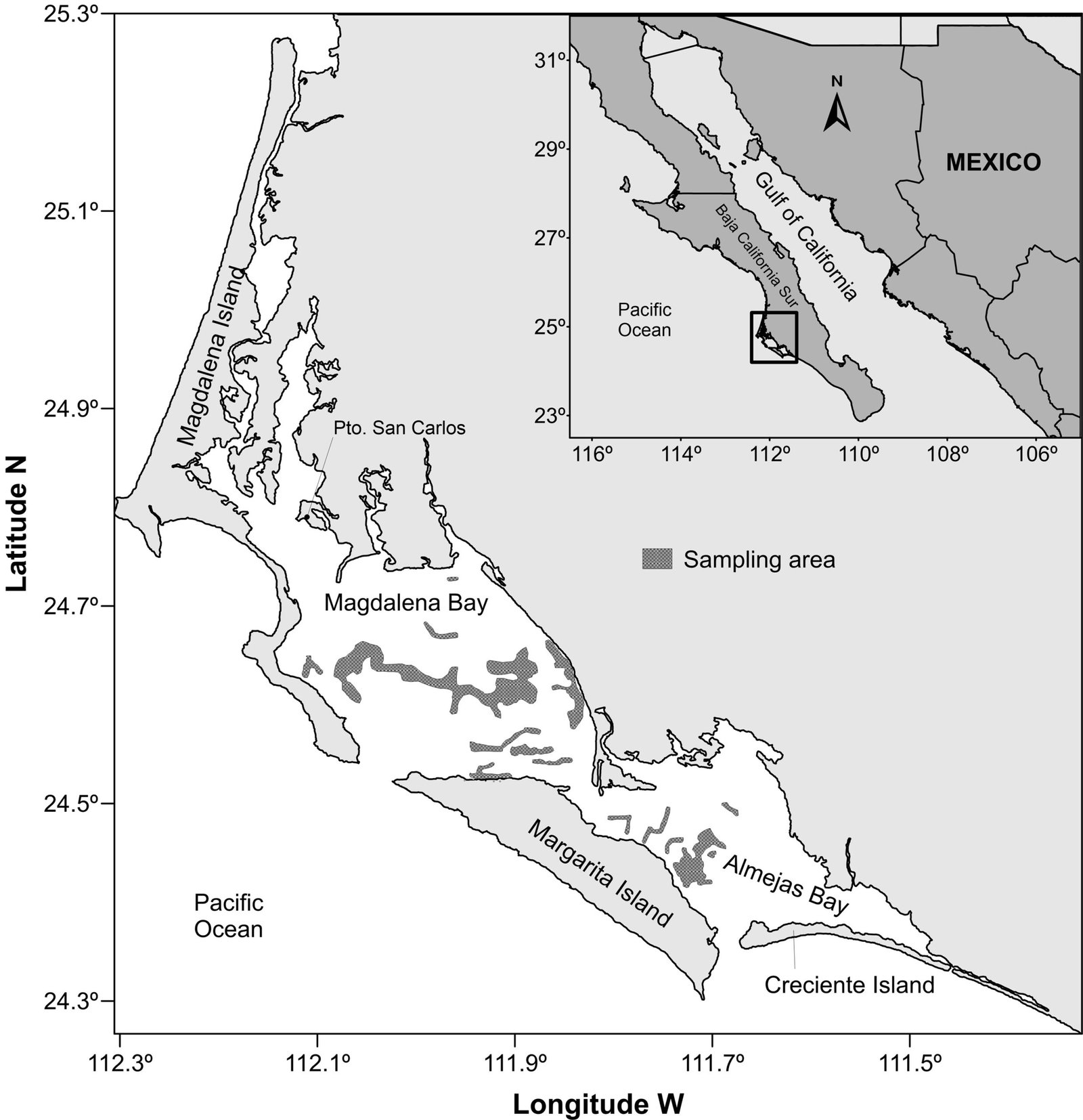
Collections were carried out in central and southern Mexico (Fig. 1), in mixed coniferous forests, during the rainy season from June to October (2000-2017). Morphological characteristics were described according to Largent (1977) and Lodge et al. (2004). Chemical reactions with KOH 5% and Melzer reagent were tested on pileus, hymenophore, and stipe surface. Photographs of basidiomata were taken on site, as well as data regarding plant species. The colors for the taxonomic description were based on Kornerup and Wanscher (1978). A light microscope (Carl Zeiss GmbH 37081, Germany) was used to observe the microscopic characteristics. Only structures from mature basidiomes were measured. Twenty basidiospores, basidia, pleurocystidia, cheilocystidia and caulocystidia were measured by specimen. If the specimen consisted of several basidiomes, only the structures of 2 of them were measured. The mean of each variable of every collection was computed. Dimensions are given according to the following format: (lower mean) mean of means ± standard deviation (upper mean), Q = (lower mean) mean of means ± standard deviation (upper mean).
Vouchers were deposited in the “Herbario Nacional de México” (MEXU), in the José Castillo Tovar herbarium (ITCV), and in the mycological herbarium of the Universidad Autónoma de Querétaro (MUAQ). Additional materials were obtained in loan from the following herbaria: Escuela Nacional de Ciencias Biológicas-Instituto Politécnico Nacional herbaria (ENCB-IPN), Instituto de Biología Universidad de Guadalajara (IBUG), Instituto Nacional de Ecología (INECOL) and Universidad Autónoma de Nuevo León (UNL). All acronyms of herbaria and mycological collections follow Index Herbariorum (Thiers, 2024).
Samples of dehydrated basidiomata were used for DNA extraction. The DNA was extracted using the DNeasy Power-Soil kit (QIAGEN). Cell lysis was performed by grinding in a mortar with liquid nitrogen. Three nuclear loci (ITS, LSU and RPB2) were amplified with Platinum Taq DNA Polymerase (Invitrogen-Thermo Fisher Scientific) or Taq & Load PCR Mastermix (MP Biomedicals) in a thermal cycler (BIO-RAD). The PCR parameters were as follows: 95 ºC initial denaturation 4 min; 35 cycles of denaturation at 94 ºC for 1 min, annealing at 54 ºC for 1 min, extension at 72 ºC for 1 min, and a final extension step at 72 ºC for 10 min. The primers ITS1/ITS4 (White et al., 1990) were used for the ITS region; LR0R/LR5 (Vilgalys & Hester, 1990) for nucLSU; and RPB2-B-F2/RPB2-B-R (Wu et al., 2014) for the RPB2 gene. The PCR products were examined by 1% agarose gel electrophoresis, the gel was stained with GelRed (Biotium) and observed under an UVP Multidoc-It transilluminator (Analytikjena). Successful PCR products were cleaned with diluted 1:1 ddH2O: ExoSAP-IT (Thermo Fisher Scientific) and incubated at 37 ºC for 45 min and 80 ºC for 15 min. Sanger sequencing of clean PCR products was performed in the “Laboratorio de secuenciación genómica de la biodiversidad y la salud” at Instituto de Biología, Universidad Nacional Autónoma de México. Samples were sequenced in both directions with PCR primers using BigDye Terminator v3.1 (Thermo Fisher Scientific). Sequences were edited in Chromas Pro Vers. 1.41 (Technelysium Pty, Ltd, Tewantin, Qld, Australia).
For phylogenetic analyses we used Cyanoboletus sequences from the ITS, nucLSU and RPB2 regions listed in the Table 1. A total of 32 concatenated sequences were used (Supplementary material 1). Two Lanmaoa species were chosen as an outgroup. The sequences of each locus were aligned separately using MacClade 4.0 (Maddison & Maddison, 2000) and concatenated in Mesquite 3.40 (Maddison & Maddison, 2018). The alignments were manually edited to eliminate ambiguous regions.
Table 1
List of DNA sequences used for Cyanoboletus phylogenetic analyses.
Species
Code
Country
GenBank
Reference
ITS
nucLSU
RPB2
Lanmaoa sublurida
USF 288426
USA
MW675740
MW662575
MW737499
Farid et al., 2021
L. sublurida
USF 300104
USA
MW675736
MW662572
MW737498
Farid et al., 2021
Cyanoboletus abieticola
MUAQ13 Paratype
Mexico
MW750332
MW750369
–
This study
C. abieticola
ITCV-1010 Paratype
Mexico
–
MW750367
–
This study
C. abieticola
MEXU-30111 Paratype
Mexico
MW209739
MW750366
PP108649
This study
C. abieticola
MEXU-30106 Paratype
Mexico
MW209740
MW750365
–
This study
C. abieticola
MEXU-30109 Holotype
Mexico
MW209738
MW750368
PP108650
This study
Table 1. Continued
Species
Code
Country
GenBank
Reference
ITS
nucLSU
RPB2
C. abieticola
MEXU-26275 Paratype
Mexico
KC152077
–
–
This study
C. abieticola
MEXU-26276 Paratype
Mexico
KC152076
–
–
This study
C. abieticola
MEXU-26278 Paratype
Mexico
KC152075
–
–
This study
C. bessettei
USF 301500 (A) Holotype
USA
MW675737
MW662571
MW737457
Farid et al., 2021
C. bessettei
USF 301500 (B) Holotype
USA
MW675738
–
MW737458
Farid et al., 2021
C. brunneoruber
HKAS63504
China
–
KF112368
KF112702
Wu et al., 2014
C. brunneoruber
HKAS80579 1
China
–
KT990568
KT990401
Wu et al., 2016
C. brunneoruber
HKAS80579 2
China
–
KT990569
KT990402
Wu et al., 2016
C. cyaneitinctus
USF 288424
USA
MW675739
MW662574
MW737461
Farid et al., 2021
C. cyaneitinctus
USF 301499 Epitype
USA
MW675744
MW662579
MW737503
Farid et al., 2021
C. cyaneitinctus
JAB184
USA
MW675731
MW662584
MW737467
Farid et al., 2021
C. cyaneitinctus
JAB324
USA
MW675732
MW662586
MW737469
Farid et al., 2021
C. cyaneitinctus
JAB325
USA
MW675733
–
MW737470
Farid et al., 2021
C. hymenoglutinosus
AB2016
India
KT907355
KT860060
–
Li et al., 2016
C. instabilis
FHMU1839
China
MG030473
MG030466
–
Chai et al., 2018
C. instabilis
HKAS59554
China
–
KF112412
KF112698
Wu et al., 2014
C. macroporus
DC21-02
India
OQ860238
OQ860239
ON364552
Das et al., 2023
C. macroporus
DC21-04
India
OQ860240
OQ860241
OQ876894
Das et al., 2023
C. mediterraneensis
K-M000265123 Holotype
Israel
–
OM801212
–
Biketova et al., 2023
C. mediterraneensis
K-M000265124 Paratype
Israel
OM801199
–
–
Biketova et al., 2023
C. mediterraneensis
TUR-A 209199
Italy
MZ265183
MZ265198
MZ277228
Carbone et al., unpublished
C. paurianus
KD22-008
India
–
OQ859920
OQ914389
Das et al., 2023
C. paurianus
KD22-009 Holotype
India
–
OQ859919
OQ914388
Das et al., 2023
C. poikilochromus
GS10070
Italy
KT157051
KT157060
KT157068
Gelardi et al., 2015
C. poikilochromus
GS11008
Italy
KT157050
KT157059
KT157067
Gelardi et al., 2015
C. poikilochromus
TO HG 100091987 Epitype
Italy
KT157047
KT157056
–
Gelardi et al., 2015
C. pulverulentus
MG126a
Italy
KT157053
KT157062
–
Gelardi et al., 2015
C. pulverulentus
MG 456a
Portugal
KT157054
KT157063
–
Gelardi et al., 2015
C. pulverulentus
MG 628a
Italy
KT157055
KT157064
KY157069
Gelardi et al., 2015
C. sinopulverulentus
HKAS59609
China
–
KF112366
KF112700
Wu et al., 2014
C. sinopulverulentus
HMAS266894
China
KC579402
–
–
Gelardi et al., 2015
Cyanoboletus. sp.
HKAS76850
China
–
KF112343
KF112697
Wu et al., 2014
Phylogenetic analyses were partitioned for both Maximum Likelihood (ML) and Bayesian Inference (BI) analyses. Best evolutionary models were selected with jModelTest 2.1.10 (Darriba et al., 2012), using the Akaike information criteria. Table 2 summarizes the evolutionary model of each region. The ML analysis was performed in RAxML 7.2.6 (Stamatakis, 2006), with 1,000 bootstrap replicates. The BI analysis was performed in MrBayes 3.2.5 (Ronquist & Huelsenbeck, 2003) with 5,000,000 generations, 4 Monte Carlo chains and sampling every 100 trees, chain convergence was determined using Tracer 1.7.2 (Rambaut et al., 2018), we discarded the first 25% of trees as burning. ML bootstrap support (BS) over 50% and Bayesian posterior probability (PP) values exceeding 0.90 are reported in the resulting tree.
Table 2
Nucleotide substitutions models by each partition obtained from jModelTest with the corrected Akaike information criterion.
Gene/Partition
Length
Evolutifon model
Nst-rates
nucLSU
747 pb
TIM1+G
6 – Gamma
ITS
603 pb
TPM3ef+G
6 – Gamma
RPB2
688 pb
TIM3ef+G
6 – Gamma
Results
The phylogenetic analyses (Fig. 1) based on the ITS, nucLSU and RPB2 concatenated dataset using ML analyses and BI showed that all Cyanoboletus species cluster in a monophyletic clade with strong support (PP 1 / BS 100). Trees obtained from separate ITS and RPB2 analyses are presented in Supplementary material 2. Sequences of all C.abieticola sp. nov. samples form a strongly supported clade (PP 1/BS 99) within the genus Cyanoboletus. Consequently, based on morphological, phylogenetic and ecological data, we describe it as a new species, which is also the first species of the genus Cyanoboletus reported from Mexico.
To determine whether the ITS or RPB2 region is more informative as a barcode, in Table 3 we show the percentages of nucleotide similarity between species. The RPB2 region commonly has more variation and generates fewer ambiguous regions than the ITS. Even while the ITS region has been considered as the fungal genetic barcode, the RPB2 region has higher resolution for species delineation in Cyanoboletus.
Description
Cyanoboletus abieticola J. García, Ayala-Vásquez & Landeros,sp. nov.
Diagnosis. Pileus 13-50 mm in diameter, viscid, widely convex, convex to plane convex, brown, yellow, pale brown, reddish-brown, cinnamon. Stipe 40-75 × 7-8 mm, viscid, cylindrical, yellow when young, middle section and apex yellow when mature, basal area red brown to red-vinaceous, and basidiospores (14.1) 16.2 ± 1.1 (17.5) × (5) 5.5 ± 0.3 (5.9) µm. It grows on the ground of neotropical Abies forests.
Description. Macroscopic characters (Fig. 2). Pileus 13-50 mm diameter, widely convex, convex to plane convex, brown yellow (4B6), pale brown (6B5), reddish-brown, cinnamon, furfuraceous, very viscid when young, incurved margin, sterile. Hymenophore attached, pores 0.3-0.7 mm in diameter, pale yellow (2A8-2A4), yellow (3A8) to yellow-olive (30B8-30B7) it stains dark blue (22F8) when touched, with some brown tones, young specimens exude a somewhat acidic astringent-flavored yellow liquid from hymenophore; tubes 2-6 mm diameter, concolor to pores, immediately changing to dark blue (25F8) when cut. Context white, 5 mm thick, dark blue (22F8) when cut; stipe context pale yellow (3A6) base red-vinaceous (10F8), turning blue (25F8) when cut. Stipe 40-75 × 7-8 mm, cylindrical, yellow when young, middle section and apex yellow when mature, basal area red brown (9C8-9C5) to red-vinaceous (10F8), surface pruinose to furfuraceous, immediately turning dark blue (22F8) when touched. Mycelium white.
Chemical reactions: pileus surface and context turning dark brown (6F8) with KOH 5%, hymenophore turning brown (5F3) with KOH 5%.
Microscopic characters (Fig. 3). Basidiospores 14.1-17.5 × 5-5.9 µm, mean values 16.2 ± 1.1 × 5.5 ± 0.3 µm, Q 2.8-3.2, means values 3.0 ± 0.1, cylindric to bacilliform, yellow in KOH, inamyloid with Melzer’s reagent, with visible suprahilar depression (Fig. 3A). Basidia 27.3-37.6 × 9.5-10.7 µm, mean values 33.3 ± 4.0 × 10.3 ± 0.5 µm, clavate, hyaline in KOH, tetrasporic (Fig. 3B). Hymenophoral trama divergent (Boletus-type), with a medium and lateral stratum of cylindrical hyphae, hyaline to yellowish brown in KOH, inamyloid with Melzer’s reagent, with gelatinized wall. Pleurocystidia 52.2-71.2 × 10.6-13.9 µm, mean values 63.2 ± 7.5 × 12.4 ± 1.4 µm, arise from subhymenium, mucronate, clavate, fusoid-ventricose, hyaline to brown in KOH (Fig. 3C), with reddish brown incrustations on Melzer. Cheilocystidia 39.6-58.4 × 8.9-11 µm, mean values 47.7 ± 7.5 × 9.9 ± 0.8 µm, fusoid-ventricose, mucronate, clavate, reddish to brown on KOH, with reddish brown incrustations (dextrinoid) in Melzer’s reagent, thick-walled (Fig. 3D). Pileipellis formed by an ixotrichoderm 250-300 µm thick, with terminal cells 34.8-45.4 × 4-5.6 µm, mean values 38.4 ± 4.8 × 4.7 ± 0.6 µm, cylindrical, yellow-reddish brown in KOH, reddish brown (dextrinoid) with Melzer’s reagent, some with thick wall, sometimes gelatinized (Fig. 3E). Stipitipellis 100-120 µm thick, ixocutis, hyphae subparallel to loosely intermingled, formed of caulocystidia 34-47 × 9.6-11.4 µm, mean values 40.6 ± 5.2 × 10.3 ± 0.7 µm, in clusters, fusoid-mucronate, clavate, some ventricose, arise from the middle or surface, hyaline to brown in KOH, with reddish brown incrustations (dextrinoid) in Melzer’s reagent.
Figure 1. Bayesian tree using the concatenated alignment (LSU, ITS and RPB2). The values in the phylogram branches represent the maximum likelihood bootstrap (MLB)/Bayesian posterior probabilities (BPP). Only MLB 50 ≥ and 90 BPP ≥ are displayed. Cyanoboletus abieticola is in bold.
Taxonomic summary
Holotype: Mexico, Oaxaca, Santa Catarina Ixtepeji, La Cumbre, Abiesguatemalensis, 17º11’ N, 96º38’ W, 2,902 m asl, 4 November 2017, Ayala-Vásquez (MEXU-30109).
Isotype: ITCV-1141
Mycobank: MB 838523.
Etymology: the name of the species is due to its association with Abies forests.
Habitat: scattered or solitary in mixed neotropical conifer forests dominated by Abies religiosa and A. guatemalensis.
Table 3
Percent nucleotide similarity between Cyanoboletus species based on their sequences, ITS/RPB2 DNA sequences. Above ITS and below RPB2 percentage ranges (WD: without data).
C. abieticola
C. bessettei
C. brunneoruber
C. cyaneitinctus
C. hymenoglutinosusus
C. instabilis
C. macroporus
C. mediterraneensis
C. paurianus
C. poikilochromus
C. pulverulentus
C. sinopulverulentus
C. abieticola
0.0-0.5 0.0
C. bessettei
5.6-6.2 8.7
0.0 0.0
C. brunneoruber
WD 5.9
WD 8.7
WD 0.0
C. cyaneitinctus
6.7-7.2 7.0
8.1 10.2
WD 5.9
0.0 0.0
C. hymenoglutinosus
3.6-3.9 WD
4.6 WD
WD WD
5.7 WD
WD WD
C. instabilis
5.7-7.0 7.5
5.7 8.3
WD 7.8
6.3 8.7
6.3 WD
WD WD
C. macroporus
4.4-4.6 6.0
5.4 8.8
WD 5.3
4.9 3.6
1.8 WD
7.0 7.9
0.0 0.0
C. mediterraneensis
2.9-3.9 2.7
2.9-3.4 7.0
WD 5.4
2.0-5.9 6.0
2.3 WD
5.1 5.8
3.1-3.2 5.4
0.0-0.3 WD
C. paurianus
WD 6.2
WD 9.2
WD 4.6
WD 3.6
WD WD
WD 8.0
WD 2.7
WD 4.9
WD 0.0
C. poikilochromus
4.2-5.9 6.9-7.1
3.9 8.5-8.7
WD 7.1-7.4
5.9-7.6 7.7-7.8
2.0-3.4 WD
7.0-7.1 7.8-8.0
3.3-4.6 7.5
1.7-3.0 6.3-6.7
WD 7.1-7.2
0.0 0.0-0.5
C. pulverulentus
3.6-3.9 6.5
4.9-5.2 9.1
WD 5.2
4.6 2.2
2.1-2.3 WD
6.3 8.4
1.8-2.1 2.8
2.3-2.8 5.1
WD 2.8
3.6-4.9 7.8
0.0-0.3 WD
C. sinopulverulentus
3.8-4.1 6.1
5.2 9.3
WD 4.6
4.0 3.9
2.8 WD
6.3 7.4
1.0 2.7
2.3-3.1 4.7
WD 2.1
3.1-4.7 7.0-7.1
1.3-1.6 3.3
WD WD
Additional material studied. Mexico, Hidalgo: El Chico National Park, Abies religiosa, 13 August 1988, J. García (ITCV-5874), 13 August 1988, J. García (ITCV-5878), 23 August 2018, J. García (ITCV-21996); Jalisco: Road to Tamazula, 15 km to Mazamitla, between Guayabos and Cabañas, A. religiosa, 24 August 1974, G. Guzmán (ENCB-11869); Road from San Sebastián del Oeste to Santa Ana, A. religiosa, 29 August 1994, L. Guzmán-Dávalos (IBUG-5349); Nevado de Colima, A. religiosa, 13 October 1984, A. Tamayo and R. González (IBUG); 11 August 1990, J. García (ITCV-6633); Estado de México: Road to Chalma, Lagunas de Zempoala National Park, A. religiosa, 1 August 1982, R. E. Chio (ENCB-368, ITCV-368), 23 September 1985, E. Perez-Silva (MEXU-19733), 17 July 1985, García (ITCV-4784); Nevado de Toluca National Park, near Ranchería La Puerta, A. religiosa, 21 August 1983, L. Colón 208-a (ENCB); Naucalpan-Toluca highway, road to Villa Alpina, La Glorieta, A. religiosa, without date, A. González-Velázquez (ENCB-965); Mpio. Amanalco, Corral de Piedras, A. religiosa, without date, A. González-Velázquez (ENCB-1454); Zone E of San Rafael Atlixco, Escualango, A. religiosa, 7 October 1983, A. Hernández (ENCB-145); La Marquesa, A. religiosa, 7 July 1963, Gispert-Imaz (MEXU-9873), 12 August 1962, G. Guzman (ENCB-3198); Ciudad de México: Former Mexico-Cuernavaca highway 3 km S. of Parres, A. religiosa, 17 July 1982, S. Chacón (ENCB-183); Michoacán: Morelia-Cd Road. Hidalgo, Sierra de Mil Cumbres, km 45, A. religiosa, 21 July 1983, J. García (UNL-3665, ITCV-3665), 15 July 1982, J. García (UNL-1998, ITCV-1998); Morelos: Road to Chalma, 5 km Huitzilac, A. religiosa, 17 July 1982, S. Chacón (ENCB-234); Oaxaca: Santa Catarina Ixtepeji, La Cumbre, 17°11’ N, 96°38’ W, A. guatemalensis, 18 July 2017, Ayala-Vásquez (ITCV-846), 20 August 2017, Ayala-Vásquez (ITCV-1002), 21 August 2017, Ayala-Vásquez and Victores-Aguirre, (ITCV-1010); 8 October 2017, Ayala-Vásquez, (MEXU-30106, ITCV-1129), Ayala-Vásquez (MEXU-30111, ITCV-1128); 4 November 2017, Ayala-Vásquez (ITCV-1136); Ayala-Vásquez (MEXU-30109, ITCV-1141), Ayala-Vásquez, (ITCV-1142); Querétaro: Mpio. Cerro El Zamorano, A. religiosa, 16 September 1995, J. García (ITCV-9560), 23 September 2017, Ferrusca 390 (MUAQ13), 19 october 2019, Ferrusca 462 (MUAQ14), Ferrusca 467 (MUAQ15), 16 september 2022, Landeros 3770 (MUAQ16); Tlaxcala: road from Tlaxco to Chignahuapan, Cerro de Teapa, El Conejo, A. religiosa, 26 June 1979, J. García (UNL-496, ITCV-496); Huamantla, road to La Malinche hilltop, A. religiosa, 29 August 2010, T. Bruns (MEXU-26275) (MEXU-26276) (MEXU-26278); Veracruz: Xico, Los Gallos, 1.5 km to N. from Ingenio El Rosario, Zona del Cofre de Perote, A. religiosa, 27 October 1983, Villarreal 1024 (INECOL-1068), 31 July 1990, J. García (ITCV-6459).
Figure 2. Cyanoboletus abieticola morphology. A) Basidiomata (holotype), B) context, C) basidiomata at different stages. Bar = 10 mm.
Figure 3. Microscopic characteristics of Cyanoboletus abieticola. A) Basidiospores, B) basidia, C) cheilocystidia, D) pleurocystidia, E) stipitipellis, F) pileipellis. Bar = 10 µm.
Remarks
Cyanoboletus abieticola is a new species with a strong phylogenetic support (PP1 / BS 99) and distinctive taxonomic characters. This species is characterized by a small pileus 13-50 mm diameter, very viscid, pale brown, yellowish-brown, brown, red to cinnamon; pileus sterile margin somewhat involute when young; hymenophore distills a liquid with sweaty acidic flavor when young, pores and tubes yellow; stipe yellow cylindrical in the middle and apex, basal area red brown to red-vinaceous, when touched immediately turns to dark blue. It is distributed in mixed coniferous forests in central and southern Mexico at altitudes ranging from 2,700 to 3,000 m asl (Fig. 4).
Figure 4. Distribution of Cyanoboletus abieticola in Mexico.
Previously, only 2 species of Cyanoboletus (C. bessettei and C. cyaneitinctus) have been recorded from North America (Farid et al., 2021). Cyanoboletus bessettei and C. cyaneitinctus have similar small basidiomata as C. abieticola, nevertheless, both species are associated with oak forests in the United States, in contrast, C. abieticola is distributed from central to southern Mexico in mixed conifer forests with a strong association with Abies. Morphological and microscopical comparisons of the American species of Cyanoboletus are shown in the Table 4. They are similar by morphology, however, C. bessettei has blue-green then reddish-brown staining in its stipe when handled, while C. abieticola and C. cyaneitinctus are bluing, also their basidiospores are bigger than those in C. bessettei (9-11 ´ 3.5-5 mm). Cyanoboletus cyaneitinctus and C. abieticola are very similar by morphology and microscopical characters, however they are not phylogenetically close, and the first one has shorter basidiospores in length (11.5-15 mm) and associates with Carya and Quercus.
Table 4
North American Cyanoboletus species morphology comparison.
Characteristic
C. bessettei
C. cyaneitinctus
C. abieticola
Pileus color
Buffy brown when young, darker brownish at maturity
Most Cyanoboletus species are morphologically similar, so to separate them, a combination of microscopic, geographic and molecular characteristics is necessary. With the description of this new species, the genus Cyanoboletus reaches its southernmost distribution in America, developing in forests of central and southern Mexico forests. Its occurrence in Abies forests is also highlighted, since in North America the genus has been recorded in mixed Pinus–Quercus forests (Farid et al., 2021).
Acknowledgements
We thank Instituto Tecnológico de Ciudad Victoria, Universidad de Quintana Roo, Universidad Nacional Autónoma de México and Universidad Autónoma de Nuevo León for supporting this research. The DNA sequences produced at IB-UNAM were financed by project Conacyt 239266 assigned to RGO. AVO thanks Conahcyt for the postdoctoral funding 3129307. The last author wants to thank the Universidad Autónoma de Querétaro for the project FNB-2022-06.
References
Bessette, A., Roody, W. C., & Bessette, A. R. (2010). Boletes of Eastern North America: a color guide to the fleshy pored mushrooms. China: Syracuse University Press.
Biketova, A. Y., Kosakyan, A., Wasser S. P., & Nevo, E. (2016). New, noteworthy, and rare species of the genus Boletus in Israel. Plant Biosystems, 150, 876–886. https://doi.org/10.1080/11263504.2014.990537
Biketova, A. Y., Wasser, S. P., Simonini, G., & Gelardi, M. (2021). Nomenclatural novelties: Neoboletus flavosanguineus (Lavorato & Simonini) Biketova, Wasser, Simonini & Gelardi, comb.nov. Index Fungorum, 505, 1.
Biketova A. Y., Rinaldi A. C., & Simonini, G. (2022). Nomenclatural novelties: Cyanoboletus mediterraneensis Biketova, A. Rinaldi & Simonini, sp. nov. Index Fungorum, 516, 1.
Carbone, M., Puddu, D., & Alvarado, P. (2023). Nomenclatural novelties: Cyanoboletus poikilochromus (Pöder, Cetto & Zuccherelli) M. Carbone, D. Puddu & P. Alvarado, comb. nov. Index Fungorum, 534, 1.
Darriba, D., Taboada, G. L., Doallo, R., & Posada, R. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nature Methods, 9, 772.
Farid, A., Bessette, A. E., Bessette, A. R., Bolin, J. A., Kudzma, L. V. Franck, A. R. et al. (2021). Investigations in the boletes (Boletaceae) of southeastern USA: four novel species and three novel combinations. Mycosphere, 12, 1038–1076. https://doi.org/10.5943/mycosphere/12/1/12
Frank, J. L., Siegel, N., Schwarz, C. F., Araki, B., & Vellinga, E. C. (2020). Xerocomellus (Boletaceae) in western North America. Fungal Systematics and Evolution, 5, 265–288. http://dx.doi.org/10.3114/fuse.2020.06.13
García, J., Pedraza, D., Silva, C. I., Andrade, R. L., & Castillo, J. (1998). Hongos del estado de Querétaro. Querétaro: Universidad Autónoma de Querétaro.
Garibay-Orijel, R., Martínez-Ramos, M., & Cifuentes, J. (2009). Disponibilidad de esporomas de hongos comestibles en los bosques de pino encino de Ixtlán de Juárez, Oaxaca. Revista Mexicana de Biodiversidad, 80, 521–534. https://doi.org/10.22201/ib.20078706e.2009.002.615
Gelardi, M. (2020). Diversity, biogeographic distribution, ecology, and ectomycorrhizal relationships of the edible porcini mushrooms (Boletus s. str., Boletaceae) worldwide: state of the art and an annotated checklist. In J. Pérez-Moreno, A. Guerin-Laguette, R. Flores Arzú, & F.Q. Yu (Eds.), Mushrooms, humans and nature in a changing World (pp. 223–271). Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-030-37378-8_8
Kornerup, A., & Wanscher, J. H. (1978). Methuen handbook of colour. London: Methuen Publishing.
Largent, D., Johnson, D., & Watling, R. (1977). How identify mushrooms to genus III: microscopic features. Eureka, California: Mad River Press.
Leonardi, M., Marinho-Furtado, A. N., Comandini, O., Geml, J., & Rinaldi, A. C. (2020). Halimium as an ectomycorrhizal symbiont: new records and an appreciation of known fungal diversity. Mycological Progress, 19, 1495–1509. https://doi.org/10.1007/s11557-020-01641-0
Li, G. J., Hyde, K. D., Zhao, R. L., Hongsanan, S., Abdel-Aziz, F. A., Abdel-Wahab, M. A. et al. (2016). Fungal diversity notes 253-366: taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity, 78, 1–237. http://doi.org/10.1007/s13225-016-0366-9
Lodge, D. J., Ammirati, J. F., Dell, T. O., & Mueller, G. M. (2004). Terrestrial and lignicolous macrofungi: collecting and describing macrofungi. In G. Mueller, G. F. Bills, & M. S. Foster (Eds.), Biodiversity of Fungi. Inventary and monitoring methods (pp. 128–158). New York: Elsevier Academic Press.
Maddison, D. R., & Maddison, W. P. (2000). MacClade 4: analysis of phylogeny and character evolution. Sunderland, Massachussetts: Sinauer Associates.
Maddison, W. P., & Maddison, D. R. (2018). Mesquite: a modular system for evolutionary analysis. Version 3.40. http://www.mesquiteproject.org
Pierotti, A. (2015). Nomenclatural novelties: Cyanoboletus flavosanguineus (Lavorato & Simonini), in Pierotti, comb. nov. Index Fungorum, 263, 1.
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., & Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biology, 67, 901-904. http://doi.org/10.1093/sysbio/syy032
Ronquist, F., & Huelsenbeck, J. P. (2003). MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19, 1572–1574. https://doi.org/10.1093/bioinformatics/btg180
Sawar, S., Naaser, N., & Khalid, A. N. (2021). Cyanoboletus macroporus (Boletaceae), a new bolete species from Pakistani forests. Karstenia, 59, 78–87.
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22, 2688–2690. https://doi.org/10.1093/bioinformatics/btl446
Vizzini, A. (2014). Nomenclatural novelties: Cyanoboletus Gelardi, Vizzini & Simonini, gen. nov. Index Fungorum, 176, 1.
Wu, G., Feng, B., Xu, J. P., Zhu, X. T., Li, Y. C., Zeng, N. K., Hosen, M. I., & Yang, Z. L. (2014). Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Diversity, 69, 93–115. https://doi.org/10.1007/s13225-014-0283-8
Wu, G., Li, Y. C., Zhu, X. T., Zhao, K., Han, L. H, Cui, Y. Y., Li, F., Xu, J. P., & Yang, Z. L. (2016). One hundred noteworthy boletes from China. Fungal Diversity, 81, 25–188. https://doi.org/10.1007/s13225-016-0375-8
Sergio I. Salazar-Vallejo a, * Víctor M. Conde-Vela b, Daniel A. López-Sánchez a
a El Colegio de la Frontera Sur, Departamento de Sistemática y Ecología Acuática, Ave. Centenario Km 5.5, Chetumal, Quintana Roo, México
b Department of Invertebrate Zoology, Smithsonian Institution, National Museum of Natural History, 10th St. & Constitution Ave. NW, Washington, DC, 20560, USA
Received: 17 January 2024; accepted: 29 April 2024
Abstract
Aaron Treadwell described Lagisca crassa from Punta Arenas, Chile 100 years ago, based on an incomplete specimen. The species was later regarded as belonging in Eunoe Malmgren, 1865, but the species has not been found after its original description. The rediscovery of specimens collected in Punta Arenas has allowed us to evaluate its morphology to clarify some incompletely known features. Further, a comparison of the holotype of L. crassa with specimens of Hermadion magalhaensi Kinberg, 1856, led us to conclude these 2 species-group names are synonyms. We provide additional information and illustrations, and make some remarks on Eunoe and Hermadion Kinberg, 1856.
Redescubrimiento de Eunoe crassa y aclaración de algunos atributos morfológicos (Annelida: Aphroditiformia: Polynoidae)
Resumen
Aarón Treadwell describió Lagisca crassa de Punta Arenas, Chile hace 100 años con un ejemplar incompleto. Luego la especie fue considerada como parte de Eunoe Malmgren, 1865, pero no fue recolectada de nuevo después de su descripción original. El redescubrimiento de ejemplares recolectados en Punta Arenas nos permitió evaluar su morfología y aclarar algunos atributos poco conocidos. Además, la comparación del material tipo de L. crassa con ejemplares de Hermadion magalhaensi Kinberg, 1856, nos hizo concluir que estas 2 especies nominales son sinónimas. En esta contribución proporcionamos información e ilustraciones adicionales y hacemos algunos comentarios sobre los géneros Eunoe y Hermadion Kinberg, 1856.
The family Polynoidae Kinberg, 1856 is one of the largest groups of marine annelids, and although there is some discrepancy about the generic definition for several taxa, and many synonyms were introduced without revisions, or after using a rather wide definition for genera, Polynoidae includes 12-13% of all polychaete genera, and about 8% or all polychaete species. Thus, Polynoidae would include almost 180 genera and about 900 species in some sources (Pamungkas et al., 2019), or 167 genera and 870 species (Read & Fauchald, 1924), but the most relevant feature is that the number of new taxa proposed per year is still growing (Pamungkas et al., 2019). One problem in identifying marine scaleworms is that they often break in parts, detach their elytra, or both, and this is widespread in specimens from the intertidal to abyssal depths. For this reason, many species have been described based on incomplete specimens. Further, as indicated by Barnich and Fiege (2009), as a result of a low number of taxonomic publications or revisions, for many polynoid genera “neither the respective generic nor specific identification characters have been critically evaluated.” This is further complicated because about 30% of all polynoid species are only known after the original description (Hourdez, 2024 pers. comm.).
The polynoin genera Lagisca Malmgren, 1865 and Eunoe Malmgren, 1865 are very similar by having 15 pairs of elytra but final segments without elytra, notochaetae as thick as, or thicker than neurochaetae, never with pilose or capillary tips, neurochaetae without semilunar pockets, with tips uni- or bidentate, ventral cirri digitate, venter smooth (Fauchald, 1977). The main difference separating them is the type of neurochaetae; in Lagisca there are at least some bidentate ones, whereas in Eunoe all are unidentate. Pettibone (1963) regarded both, Eunoe and Lagisca as subgenera of Harmothoe Kinberg, 1856. Malmgren (1865: key) separated Lagisca from Harmothoe because of the extent of dorsal cover by elytra. In Lagisca, the last segments are exposed, whereas in Harmothoe they are always covered. These 2 genera are regarded as synonyms because besides both having bidentate neurochaetae, larger specimens of Harmothoe usually have the last segments exposed (Barnich et al., 2006). However, as indicated by Fauvel (1916), the diagnostic relevance of this feature implies there are 12-19 chaetigers uncovered, as in Hermadion Kinberg, 1856, against a few (up to 5) in Harmothoe and other genera.
On the other hand, Eunoe also resembles Hermadion Kinberg, 1856, another subgenus in Harmothoe after Pettibone (1963), which is regarded as distinct by Wehe (2006). They are similar to each other by having the same number of elytra, final segments without elytra, and notochaetae as thick as, or thicker than neurochaetae. The main differences between them are that Eunoe species have less segments (40 vs. 50 or more), and neurochaetae are all unidentate in Eunoe, whereas they were regarded as uni- or bidentate in Hermadion. However, Bock et al. (2010) revised Hermadion, redefined the genus, and concluded it is monotypic, with H. magalhaensi Kinberg, 1856, as its type species. They did not provide an emended diagnosis but after their key, the diagnostic features would include body short, with up to 50 segments, prostomium without cephalic peaks, anterior eyes towards anterior margin (after figure), notochaetae with blunt tips, and neurochaetae denticulate, without semilunar pockets.
Hartman (1938) studied the holotype of Lagisca crassa and although she hesitated about its placement in Eunoe Malmgren, 1865, she completed the original description, corrected some details and included illustrations for the prostomium, 1 parapodium, and tips of 1 notochaeta and 2 neurochaetae. The prostomium has the anterior eyes ventral, under anterior prostomial margin, and chaetae were depicted with better definition of their fine details; she also indicated that palps, antennae and dorsal cirri were smooth, and that aciculae are exposed. Later, Hartman (1956, 1959) listed Treadwell’s species in Eunoe and hence confirmed the new combination.
Rozbaczylo (1985) noted that 3 species of Eunoe had been recorded for Chile: E. crassa (Treadwell, 1924), E. opalina M’Intosh, 1885, and E. rhizoicola Hartmann-Schröder, 1962. However, regarding E. crassa, after the original description, the species has been listed for Chile by Wesenberg-Lund (1962) but no additional specimens have been found.
On the other hand, Hermadion magalhaensi has more records for Chile, but only the original description (and the one for its junior synonym H. longicirratus Kinberg, 1856, plus 2 figures by Fauvel [1916]) has been illustrated, with all other records only listing the species in several Chilean localities from the intertidal to 200 m water depth (Rozbaczylo, 1985).
In this contribution, we document the discovery of some specimens of Eunoe crassa (Treadwell, 1924), collected in the type locality, and deposited in the University of Miami Voss Museum of Marine Invertebrates collection. Because the specimens are well-preserved, some remarks are introduced in the diagnosis of the involved genera, and the diagnostic features are clarified, explained, and accompanied by some illustrations. We also conclude, after the study of the holotype of E. crassa, that it is a junior synonym of H. magalhaensi.
Materials and methods
During part of the cruise 23 of the USNS Eltanin, some specimens were collected in Punta Arenas, Chile. They were deposited in the University of Miami Voss Museum of Marine Invertebrates (UMML). Additional specimens for comparison of H. magalhaensi (USNM 57798) and the holotype of L. crassa Treadwell, 1924 (USNM 19101) were examined at the National Museum of Natural History, Smithsonian Institution.
Specimens were observed with stereomicroscopes. Some detached elytra, parapodia and chaetae were observed in compound microscopes. Digital photos were stacked with HeliconFocus8, and plates were prepared with PaintShopPro and Photoshop CS.
Diagnosis (slightly modified after Barnich and Fiege [2010]). Body depressed, short, with up to 50 segments; dorsum more or less covered by elytra or short posterior region uncovered. Fifteen pairs of elytra on segments 2, 4, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 26, 29, and 32. Prostomium with or without distinct cephalic peaks and 3 antennae; lateral antennae inserted ventrally to median antenna. Anterior pair of eyes dorsolateral at widest part of prostomium, posterior pair dorsal near hind margin. Parapodia with elongate acicular lobes with both aciculae with tips exposed; neuropodia with a supra-acicular process. Notochaetae stout with distinct rows of spines, tips blunt. Neurochaetae more numerous and thinner, with distinct rows of spines distally and exclusively unidentate tips.
Remarks
Eunoe Malmgren, 1865 includes 46 species-group names distributed along all oceans from the intertidal to abyssal depths (Read & Fauchald, 2024). Malmgren (1865) included a key to genera and the diagnostic features for Eunoe were 15 pairs of elytra, covering dorsum, less than 45 segments; lateral antennae subventral; notochaetae with transverse rows of spines; neurochaetae unidentate with tips falcate, thinner than notochaetae. The same features were completed for the diagnosis (Malmgren, 1865).
There are no keys for identifying all Eunoe species. Species have been sorted out after the position of the anterior eyes (under anterior margin vs. median prostomial area), palp surface (papillose vs. spinulose), elytral features (pigmentation, fimbriae, macrotubercles), tips of notochaetae (tapered, mucronate, elongate), tips of neurochaetae (acute, swollen), and size or extent of subdistal denticulate region (short or long). Jimi et al. (2021) described a dimorphic species, and noted that after some molecular indicators, 2 groups of species can be recognized in Eunoe.
Malmgren (1865) included 2 species in Eunoe: E. oerstedi (Fig. 3A-D, in his plate 8), a replacement name for Lepidonote (sic) scabra Örsted, 1843, and the new combination of E. nodosa (Sars, 1861) for Polynoe nodosa Sars, 1861 (Fig. 4A-D in his plate 8). The main difference between these 2 species is the type of macrotubercles because in E. nodosa they have granulose tips, whereas they are spiny in E. oerstedi. On the other hand, the replacement name, E. oerstedi was unnecessary because there was no homonymy (ICZN, 1999, Art. 52.2) or matching combinations, but it has become accepted in recent publications and redescriptions (Barnich & Fiege, 2010; Pettibone, 1954, 1963).
The generic diagnosis included above indicates that prostomium has or lacks cephalic peaks. The type species, E. nodosa (Sars, 1861) has cephalic peaks “rather inconspicuous” (Barnich & Fiege, 2010).
On the other hand, the above diagnosis, slightly modified after Barnich and Fiege (2010), leaves out the species with eyes present towards the anterior prostomial region, as in E. crassa (Treadwell, 1924), not along the widest prostomial area. Other species having anterior eyes displaced anteriorly include E. alvinella Pettibone, 1989; E. barbata Moore, 1910; E. clarki Pettibone, 1951; E. hubrechti (McIntosh, 1900); E. papillosa Amaral & Nonato, 1982; E. rhizoicola Hartmann-Schröder, 1962; E. senta (Moore, 1902); E. spinosa Imajima, 1997 and E.subtruncata Annenkova, 1937. Further, the only species having anterior eyes displaced anteriorly and directed ventrally are E. barbata, E. clarki, E. rhizoicola, E. senta, and E. spinicirris. Another alternative, which should be based upon the study of type materials, would be to transfer these species to Hermadion, but this is beyond our current objectives.
The study of the type material of Lagisca crassa Treadwell, 1924 allowed us to conclude it belongs in Hermadion, and that it is a junior synonym of H. magalhaensi Kinberg, 1856, as redescribed elsewhere (Bock et al., 2010).
Type species. Hermadion magalhaensi Kinberg, 1856 by subsequent designation (Hartman, 1959: 79).
Diagnosis. Body depressed, short, with up to 50 segments, posterior region without elytra. Fifteen pairs of elytra on segments 2, 4, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 26, 29, and 32. Prostomium without cephalic peaks, and 3 antennae, lateral antennae inserted ventrally to median one. Anterior pair of eyes ventrolateral, posterior pair towards hind margin. Parapodia with elongate acicular lobes with acicular tips exposed; neuropodia without supracicular process. Notochaetae stout with distinct rows of spines, tips blunt. Neurochaetae thinner, with distinct rows of spines distally and only unidentate tips.
Remarks
Hermadion Kinberg, 1856 is currently regarded as a monotypic genus (Bock et al., 2010). If the species listed above become regarded as members of Hermadion, then the potential diagnostic features would be centered in papillation of dorsal cirri and elytral ornamentation (fimbriae, and macro- and microtubercles); however, as indicated above, revising these species is beyond our current objectives.
Diagnosis. Hermadion with elytra without fimbriae, surface covered by short microtubercles, round and elongate spine-like; dorsal cirri with small papillae, tips mucronate, smooth.
Additional material. Punta Arenas, Chile. Four specimens (UMML), USNS Eltanin, Cruise 23, Sta. P4-3 (53°11’ S, 70°50’ W), shore collection, by hand, 0.8-1.6 km south of commercial pier, 30 Mar. 1966, fixed in isopropyl alcohol, McSween, coll. 11 specimens (USNM 57798), Cobble Beach, Magellanes, 6 May 1965, J. Mohr, coll.
Holotype of Lagisca crassa. The holotype (USNM 19101) is posteriorly incomplete, 7.3 mm long, 2.4 mm wide, 32 segments (Fig. 1A, C). Most elytra, cephalic appendages and dorsal cirri detached, some parapodia previously dissected, pharynx everted.
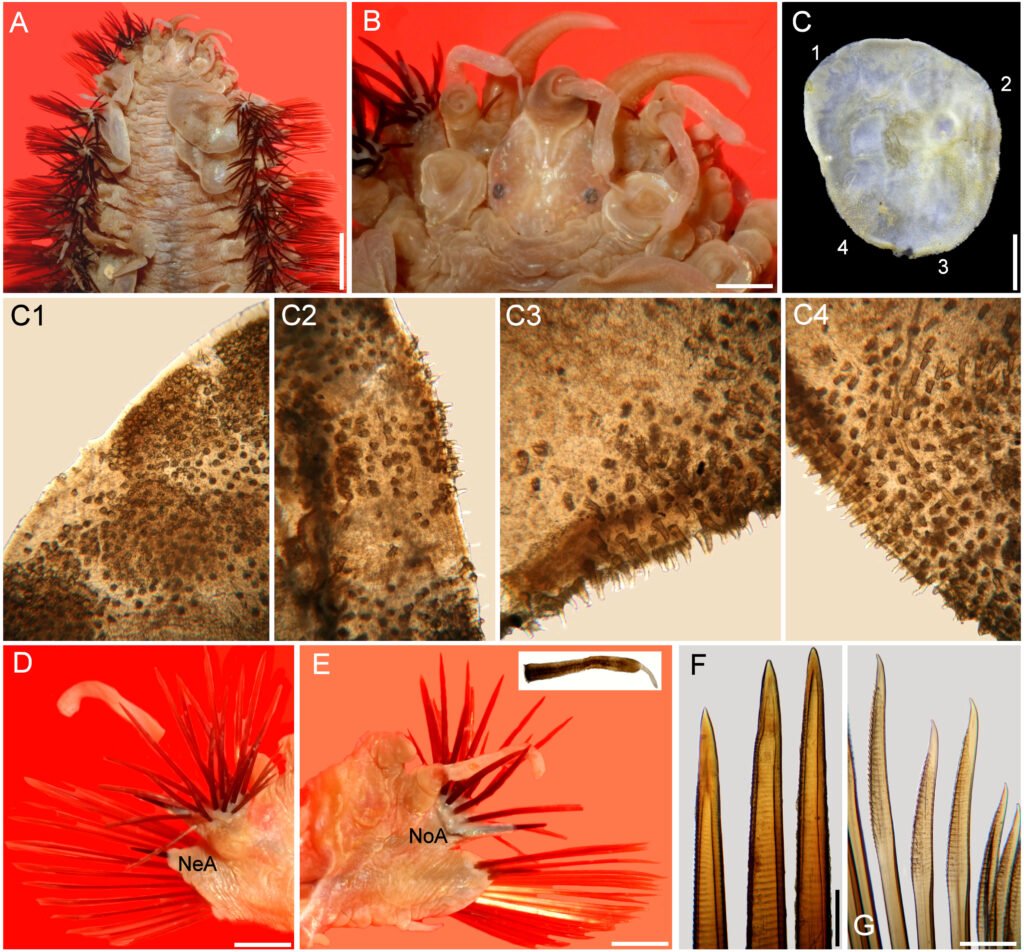
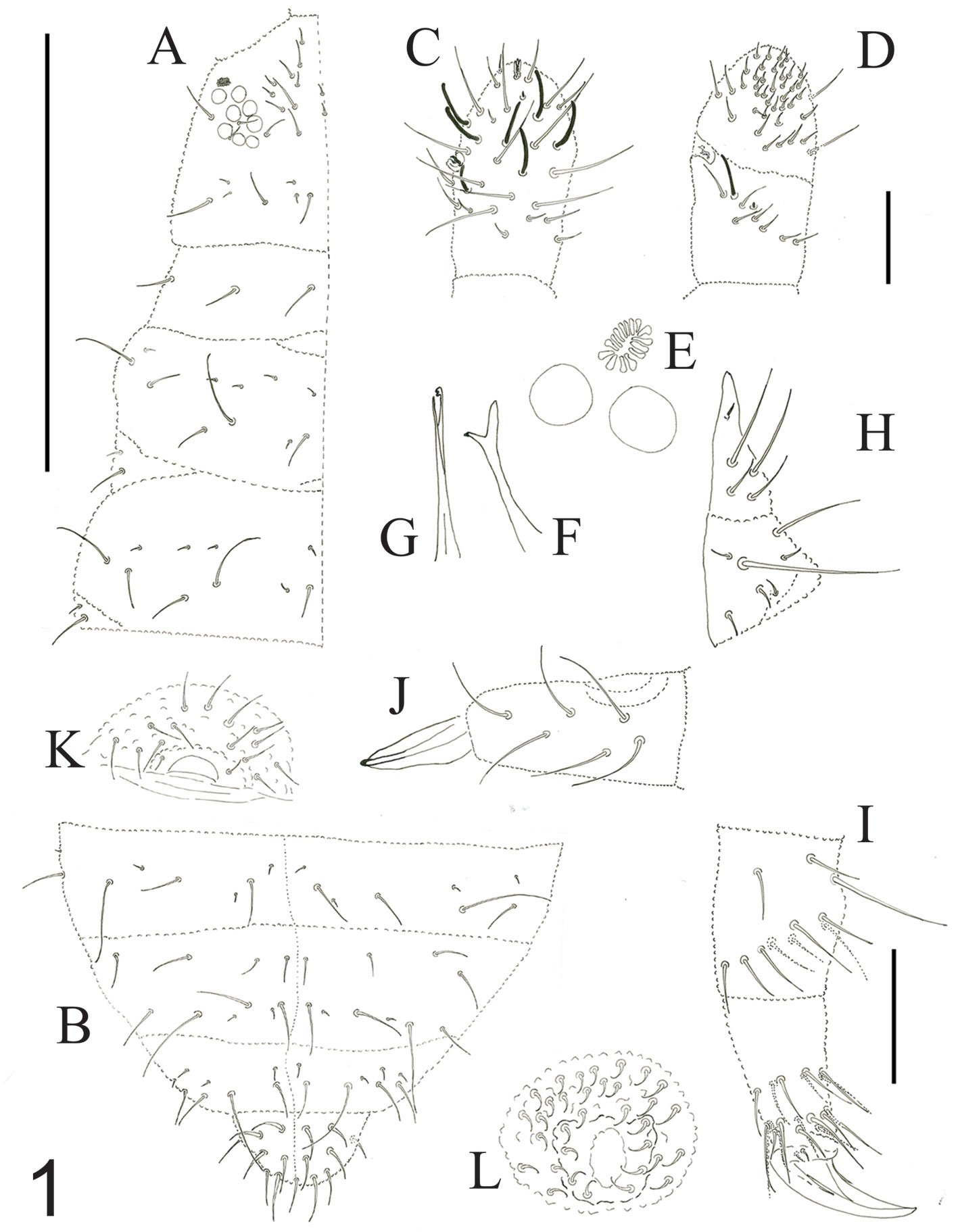
Figure 1. Hermadion magalhaensi Kinberg, 1856. Holotype of Lagisca crassa Treadwell, 1924 (USNM 19101). A, Whole specimen, dorsal view; B, C, elytra from anterior segments, seen from above; D, right cirrigerous chaetiger from middle segment, posterior view; E, right elytrigerous segment from middle segment, anterior view; F, anterior end, dorsal view; G, same, ventral view; H, I, notochaetae from middle segment; J, K, tips of same; L, M, supra-acicular neurochaetae from middle segment; N, O, sub-acicular neurochaetae from middle segment; P, tip of supra-acicular neurochaeta; Q, tip of sub-acicular neurochaeta. Scale bars: A, 5 mm; B-E, 1 mm; F, G, 2 mm; H, I, L-O, 0.1 mm; J, K, P, Q, 50 μm.
Prostomium longer than wide (Fig. 4F); eyes almost faded out, anterior eyes anteroventral (Fig. 1F). Ceratophore of median antenna with a V-shaped depression; ceratostyle missing. Lateral antenna ventral, ceratophores about half as wide as median one; ceratostyles 1.5x longer than ceratophores (Fig. 1F). Palps lost.
Tentacular cirri with chaetae; cirrostyles distally incomplete, slightly longer than cirrophores (Fig. 1F). Facial tubercle not visible dorsally.
Pharynx fully exposed (Fig. 1F, G); no pigments observed, slightly expanded distally; 9 pairs of marginal papillae. Jaws dark brown (Fig. 1G), blunt tips, without accessory denticles.
Elytra pale, non-fimbriate (Fig. 1B, C), with eccentric insertions. Surface almost fully covered with microtubercles; microtubercles rounded, rod-like, or distally truncate.
Parapodia biramous from segment 2. Few dorsal cirri remain attached, all without tips (Fig. 1D). Both notacicular and neuracicular lobes projected, tips of aciculae exposed (Fig. 1D, E). Ventral cirri tapered, reaching base of neuracicular lobe (Fig. 1D, E). Nephridial lobes blunt, present from segment 9 throughout body.
Notochaetae light brown, of different sizes (Fig. 1H, I), each blunt, with series of denticles, margin finely spinulose, tips delicately bent, entire (Fig. 1J, K). Neurochaetae light brown to transparent, subdistally expanded (Fig. 1L-O), with rows of denticles leaving tip smooth (Fig. 1P, Q); tip falcate, unidentate (Fig. 1P, Q).
Posterior end lost (Fig. 1A).
Additional material. The UMML and some USNM specimens were complete, body wall brittle, most elytra detached, some cephalic appendages and dorsal cirri lost, some with some portions removed likely after predatory attacks, especially along anterior region including right parapodia (Fig. 2), or right lateral antenna (Fig. 3B). Some specimens bent laterally, others bent ventrally, 2 with pharynx exposed. Body 35-47 mm long, 9-17 mm wide, 43-47 segments. Elytra overlapping laterally leaving middorsal area exposed in some specimens (Fig. 2A), fully covering it in others (Fig. 4A, B).
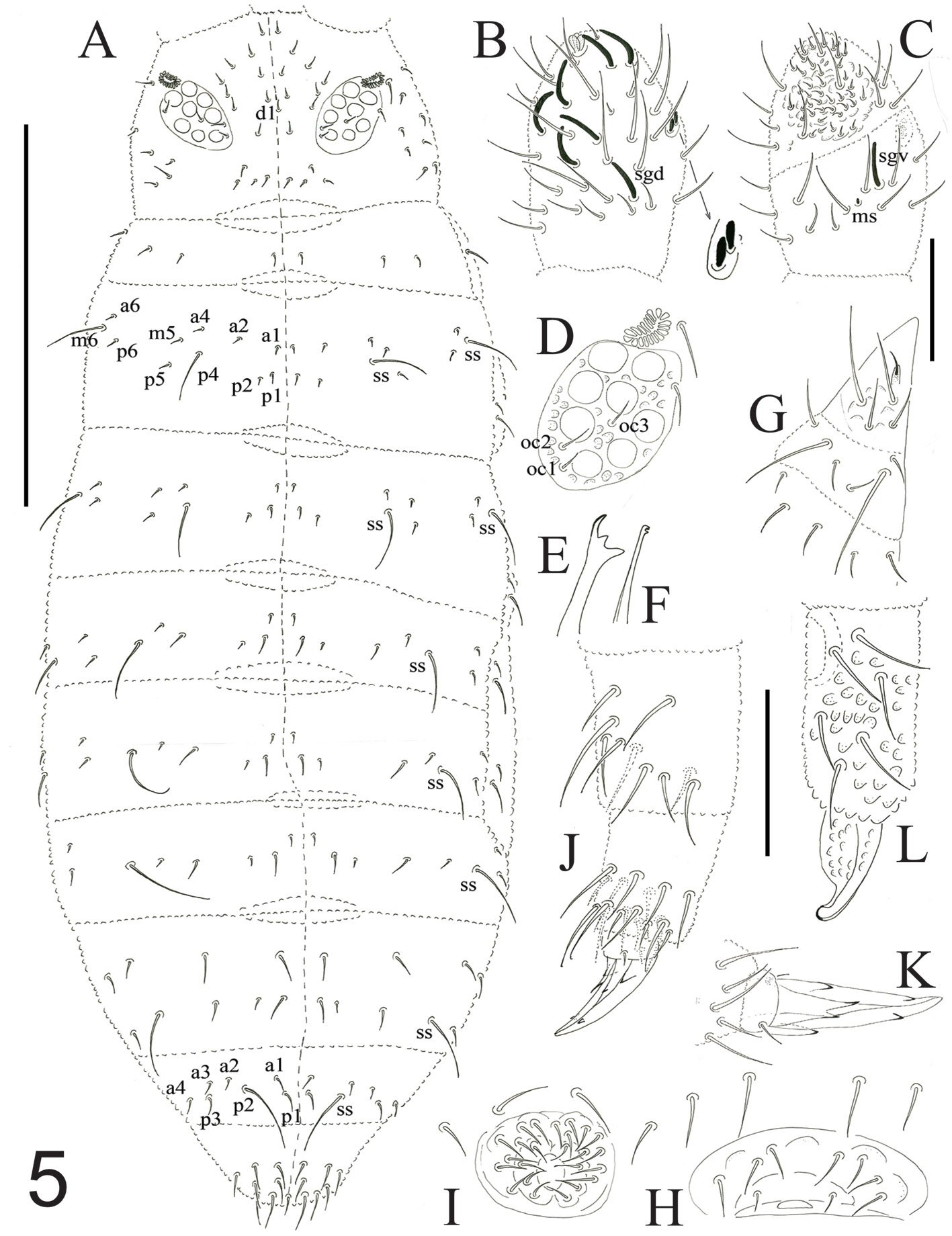
Figure 2. Hermadion magalhaensi Kinberg, 1856, topotype specimen (UMML). A, Anterior región, dorsal view; B, anterior end, dorsal view; C, right elytron 6, seen from above (1-4: sections enlarged in C1-C4); D, chaetiger 18, right parapodium, anterior view (NeA, neuracicular lobe); E, same, posterior view (inset: dorsal cirrus; NoA, notacicular lobe); F, tips of notochaetae; G, tips of neurochaetae. Scale bars: A, 2.1 mm; B, 0.6 mm; C, 1.1 mm; D, E, 1 mm; F, G, 180 µm.
Prostomium longer than wide. Eyes black, anterior eyes ventrolateral, not visible dorsally (Fig. 2B), better perceived in frontal view (Fig. 3A, B). Antennae and cirri cylindrical, tips mucronate, blunt. Median antenna with ceratophore forming a V-shaped depression, about 4 times wider than ceratostyle, ceratostyle about as long as prostomium. Lateral antennae ventral, ceratophores about half as wide as median one; ceratostyles lost. Palps thick, short, about as long as median antennae, finely papillate, but papillae not arranged in rows.
Tentacular cirri with cirrophores fused, with chaetae exposed; cirrostyles cylindrical mucronate. Facial tubercle pale, not visible dorsally, better defined after pharynx is exposed.
Elytra barely pigmented, non-fimbriate (Figs 2C, 4D), with variable amount of sediment particles. Surface covered by abundant microtubercles (Fig. 4D), small globular along anterior regions (Fig. 3C1, 2), progressively longer along posterior region, projected beyond elytral margin (Figs. 3C3, 4; 4E). Other specimens with a diffuse spot surrounding central area. Insertion area eccentric, displaced anteriorly and laterally, to the right in right elytra, to the left in left ones.
Figure 3. Hermadion magalhaensi Kinberg, 1856, topotype specimen (UMML). A, Anterior región and pharynx, dorsal view; B, anterior end, frontal view; C, pharynx opening, frontal view (Pa, papilla). Scale bars: A, 1 mm; B, 0.6 mm; C, 0.9 mm.
Parapodia biramous from segment 2. Dorsal cirri finely papillate, papillae not arranged in rows, tip smooth. Notopodia with dorsal cirri cylindrical, tip mucronate (Fig. 2D, E). Notacicular lobe projected, aciculae exposed (Fig. 4F, G). Neuropodia with neuropodial lobe projected, rarely with a long prechaetal subacicular lobe (Fig. 4G). Neuracicular lobe projected, aciculae exposed (Fig. 4F, G). Ventral cirri tapered, short, reaching base of neuracicular lobe. Nephridial lobes blunt, short, present from chaetiger 8-9, continued along body.
Notochaetae dark brown, abundant, roughly verticillate, each blunt, with series of denticles, margin finely spinulose, tips delicately bent, entire (Figs. 2F; 4H, I). Neurochaetae brownish, subdistally expanded, with rows of denticles leaving tip smooth; tip falcate, unidentate (Figs. 2G; 4J-M).
The pharynx is fully exposed; it is 9 mm long in a 46 mm long specimen (Fig. 3A). The outer surface looks maculate but the spots correspond with adsorbed crystals on the surface. The pharynx tube is slightly expanded distally, its margins are eroded and only the lateral papillae are left after erosion of most marginal integument (Fig. 3C); it was described with 9 pairs of marginal papillae. Jaws dark brown, tips blunt, without accessory denticles (Fig. 3C).
Posterior end tapered (Fig. 4C); pygidium with anus terminal; anal cirri short, resembling dorsal cirri.
Remarks
Hermadion magalhaensi Kinberg, 1856 and H. longicirratus Kinberg, 1856 were both described from the same locality and depth in Saint York Bay, Magellan Strait. The main differences between these species were that H. magalhaensi has smooth elytra, and smooth notochaetae, whereas H. longicirratus has elytra minutely tuberculate and spiny notochaetae; the former species was based on a 52 mm long specimen, whereas the latter on a 14 mm long specimen. The size difference might explain some features present in the smaller specimen and lost after abrasion in the larger specimen. However, the microtubercles in elytra might accumulate sediment and look smooth if they are not carefully cleaned. Fauvel (1916: 425) studied several specimens of different size and concluded H. magalhaensi and H. longicirratus were the same species.
Figure 4. Hermadion magalhaensi Kinberg, 1856, topotype specimen (USNM 57798). A, Whole specimen, dorsal view; B, anterior end, dorsal view; C, posterior end, dorsal view; D, right elytron from middle segment, seen from above; E, microtubercles from elytral margin; F, right cirrigerous chaetiger from middle segment, anterior view; G, right elytrigerous chaetiger from middle segment, anterior view; H, I, tips of notochaetae from middle segment; J, K, tips of supra-acicular neurochaetae from middle segment; L, M, tips of sub-acicular neurochaetae from middle segment. Scale bars: A, 3 mm; B, C, 1 mm; D, F, G, 0.5 mm; E, 0.1 mm; H-M, 50 μm.
Treadwell (1924) described Lagisca crassa from Punta Arenas, Chile based on an incomplete specimen, without most of its elytra, and included figures for the anterior end, 1 cirrigerous parapodium, and tips of 1 notochaetae (tapered), and 1 neurochaetae (unidentate), but no elytra were illustrated. Ceratostyles of antennae and dorsal cirrostyles were shown with a subdistal brown ring but were not subdistally expanded. The median antenna is longer than laterals, its base marks a deep V-shaped depression over prostomium, and the parapodium shows acicular lobes projected, but tips of aciculae were not emergent. The pharynx was indicated as having 9 pairs of marginal papillae, but no details of the jaws were provided.
On the other hand, what has been regarded as E. crassa and E. rhizoicola Hartmann-Schröder, 1962 are the 2 only species described and recorded from shallow water depths in Chile. The latter species was also described from Punta Arenas, with a 21 mm long specimen. These 2 species are very similar by having anterior eyes displaced anteriorly, similar types of noto- and neurochaetae, and dorsal cirri with black bands. They differ because in E. crassa the dorsal cirri have a single subdistal black band, against 2 in E. rhizoicola, and its tip is short, whereas it is longer in E. rhizoicola. The main difference is in the presence of fimbriae; there are no fimbriae in E. crassa, whereas E. rhizoicola has some short filaments along posterior margins. It is likely that E. rhizoicola is another junior synonym of H. magalhaensi because it resembles H. longicirratus in having longer dorsal cirri, but this might be a size dependent feature, becoming relatively shorter in larger specimens. Further, H. magalhaensi has been found living in Macrocystis rhizoids (Pratt, 1901), which was the habitat also for E. rhizoicola.
We think that the main reason for the confusion regarding the affinities between what was described as L. crassa and H. magalhaensi is because there were only 1 set of illustrations of the species (Kinberg, 1858), and since during many years, the proposals for new records or new species did not include the study of type or topotype specimens (Fauchald, 1989).
Distribution. Originally described from Puntarenas, Chile, in shallow depths (0-200 m), it ranges along subantarctic localities including the Falkland and Kerguelen Islands.
Acknowledgments
Geoff Read, Igor Jirkov and Oscar Díaz-Díaz kindly provided useful publications. William Moser found some additional field data for the Eltanin station where the specimens were found. Stéhane Hourdez and an anonymous referee carefully read this contribution and suggested several important modifications. The technical editorial issues were masterfully made by María Antonieta Arizmendi.
References
Amaral, A. C., & Nonato, E. F. (1982). Anelideos poliquetos da costa brasileira, 3. Aphroditidae e Polynoidae. Brazilia, Conselho Nacional de Desenvolvimento Científico e Tecnológico.
Annenkova, N. P. (1937). Fauna Polychaeta severnoii chasti Yaponskogo Morya. Issledovaniya Morei SSSR, 23, 139–216.
Augener, H. (1932). Antarktische und Antiboreale Polychaeten nebst einer Hirudinee. Scientific Results of the Norwegian Antarctic Expeditions 1927-1928 et sqq., instituted and financed by Consul Lars Christensen, 9, 1–85.
Averintsev, V. G. (1972). Donnye mnogotchetinkovye chervi Errantia Antarktiki i Subantarktiki no materialam sovetskoi Antarktitcheskoi Exspeditsii. Issledovaniya Fauny Morei, 11(19). Resul’taty biologicheskikh issledovanii Sovetskikh Antarktitseskikh Exspeditsii, 5, 85–293.
Baird, W. (1865). Contributions towards a monograph of the species of Annelides belonging to the Aphroditacea, containg a list of the known species, and a description of some new species contained in the National Collection of the British Museum. Journal of the Linnean Society, Zoology, 8, 172–202. https://doi.org/10.1111/j.1096-3642.1865.tb02438.x
Barnich, R., & Fiege, D. (2003). The Aphroditoidea (Annelida: Polychaeta) of the Mediterranean Sea. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft Frankfurt am Main, 559, 1–167.
Barnich, R., & Fiege, D. (2009). Revision of the genus Harmothoe Kinberg, 1856 (Polychaeta: Polynoidae) in the Northeast Atlantic. Zootaxa, 2104, 1–76. https://doi.org/10.11646/zoo taxa.2104.1.1
Barnich, R., & Fiege, D. (2010). On the distinction of Harmothoe globifera (G.O. Sars, 1873) and some other easily confused polynoids in the NE Atlantic, with the description of a new species of Acanthicolepis Norman in McIntosh, 1900 (Polychaeta, Polynoidae). Zootaxa, 2525, 1–18. https://doi.org/10.11646/zootaxa.2525.1.1
Barnich, R., Fiege, D., Micaletto, G., & Gambi, M. C. (2006). Redescription of Harmothoe spinosa Kinberg, 1856 (Polychaeta: Polynoidae) and related species from Subantarctic and Antarctic Waters, with the erection of a new genus. Journal of Natural History, 40, 33–75. https://doi.org/10.1080/00222930500445044
Bock, G., Fiege, D., & Barnich, R. (2010). Revision of Hermadion Kinberg, 1856, with a redescription of Hermadion magalhaensi Kinberg, 1856, Adyte hialina (G.O. Sars, 1873) n. comb., and Neopolynoe acanellae (Verrill, 1881) n. comb. (Polychaeta: Polynoidae). Zootaxa, 2554, 45–61. https://doi.org/10.11646/zootaxa.2554.1.4
Dales, R. P. (1962). The polychaete stomodeum and the inter-relationships of the families of Polychaeta. Proceedings of the Zoological Society of London, 139, 389–428. https://doi.org/10.1111/j.1469-7998.1962.tb01837.x
Ehlers, E. (1897). Hamburger Magalhaensischen Sammelreise 1892/93 3(Bryozoen und Würmer). Polychaeten. Hamburg: Friederichsen & Co.
Fauchald, K. (1977). The polychaete worms: definitions and keys to the orders, families and genera. Natural History Museum of Los Angeles County, Science Series, 28, 1–188.
Fauchald, K. (1989). The second annual Riser lecture: eclecticism and the study of polychaetes. Proceedings of the Biological Society of Washington, 102, 742–752.
Fauvel, P. (1916). Annélides polychètes des Iles Falkland recueillies par M. Rupert Vallentin Esq (1902-1910). Archives de Zoologie Expérimentale et Générale, 55, 417–482. https://doi.org/10.5962/bhl.part.11511
Fauvel, P. (1923). Polychètes errantes. Faune de France, 5,1–488.
Gravier, C. (1911). Deuxième Expédition Antarctique Française (1908-1910), commandé par le Dr. Jean Charcot. Annélides polychètes. Paris: Masson et Cie.
Hartman, O. (1938). The types of the polychaete worms of the families Polynoidae and Polyodontidae in the United States National Museum and the description of a new genus. Proceedings of the United States National Museum, 86, 3046, 107–134. https://doi.org/10.5479/si.00963801.86-3046.107
Hartman, O. (1956). Polychaetous annelids erected by Treadwell, 1891 to 1948, together with a brief chronology. Bulletin of the American Museum of Natural History, 109, 239–310.
Hartman, O. (1959). Catalogue of the polychaetous annelids of the World. Allan Hancock Foundation Publications, Occasional Paper, 23, 1–628.
Hartman, O. (1964). Polychaeta Errantia of Antarctica. Antarctic Research Series, 3, 1–131. https://doi.org/10.1029/ar003
Hartmann-Schröder, G. (1962). Zur Kenntnis des Eulitorals der chilenischen Pazifikküste und der argentinischen Küste Südpatagoniens unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Die Polychaeten des Eulitorals. Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, 60, 57–270.
Imajima, M. (1997). Polychaetous annelids from Sagami Bay and Sagami Sea collected by the Emperor Showa of Japan and deposited at the Showa Memorial Institute, National Science Museum, Tokyo. Families Polynoidae and Acoetidae. National Science Museum Monographs, 13,1–131.
ICZN (International Commission of Zoological Nomenclature). (1999). International Code of Zoological Nomenclature. London, Natural History Museum. Available at: www.code.iczn.org
Jimi, N., Hookabe, N., Moritaki, T., Kimura, T., & Imura, S. (2021). First evidence of male dwarfism in scale worms: A new species of Polynoidae (Annelida) from hermit crab and molluscan shells. Journal of Zoological Systematics and Evolutionary Research, 59,801–818. https://doi.org/10.1111/jzs.12463
Jirkov, I. A. (2001). Polikhety Severnogo Ledovitogo Okeana. Tromsø, Akvaplan-Niva.
Kinberg, J. G. H. (1856). Nya slägten och arter af Annelider. Öfversigt af Kongliga Vetenskaps-Akademiens Förhhandlingar Stockholm, 12, 381–388.
Kinberg, J. G. H. (1858). Kongliga Svenska Fregatten Eugenies resa Omkring Jorden under befäl af C.A. Virgin, Ånen 1851-1853, Vetenskapliga Iakttagelser, Zoologi, 3. Annulater. Stockholm.
Levinsen, G. M. R. (1883). Systematisk-geografisk Oversigt over de nordiske Annulata, Gephyrea, Chaetognathi og Balanoglossi. Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i Kjøbenhavn, 1882, 160–251. https://doi.org/10.5962/bhl.title.16117
Malmgren, A. J. (1865). Nordiska Hafs-Annulater. Öfversigt af Kongl. Vetenskaps-Akademiens Förhandlingar, 22,51–110,
M’Intosh, W. C. (1885). Report on the Annelida Polychaeta collected by H.M.S. Challenger during the years 1873-1876. Reports on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–76, Zoology, 12,i-xxxvi.
McIntosh, W. C. (1900). A monograph of British Annelids, 1(2) Polychaeta Amphinomidae to Sigalionidae. London: Ray Society of London.
Moore, J. P. (1902). Descriptions of some new Polynoidae, with a list of other Polychaeta from North Greenland waters. Proceedings of the Academy of Natural Sciences of Philadelphia, 54, 258–278.
Moore, J. P. (1910). The polychaetous annelids dredged by the U.S.S. “Albatross” off the coast of Southern California in 1904: 2. Polynoidae, Aphroditidae and Segalionidae (sic). Proceedings of the Academy of Natural Sciences of Philadelphia, 62, 328–402.
Örsted, A. S. (1843). Grönlands Annulata dorsibranchiata. Det Kongelige Danske videnskabernes selskabs. Naturviden- skabelige og Mathematiske Afhandlinger, 10, 153–216.
Pamungkas, J., Glasby, C. J., Read, G. B., Wilson, S. P., & Costello, M. J. (2019). Progress and perspectives in the discovery of polychaete worms (Annelida) of the world. Helgolander Marine Research, 73, 4. https://doi.org/10.1186/s10152-019-0524-z
Pettibone, M. H. (1951). A new species of polychaete worm of the family Polynoidae from Point Barrow, Alaska. Journal of the Washington Academy of Sciences, 41,44–45.
Pettibone, M. H. (1954). Marine polychaete worms from Point Barrow, Alaska, with additional records from the North Atlantic and North Pacific. Proceedings of the United States National Museum, 103, 203–356. https://doi.org/10.5479/si.00963801.103-3324.203
Pettibone, M. H. (1963). Marine polychaete worms of the New England región, 1. Families Aphroditidae through Trochochaetidae. Bulletin, United States National Museum, 227, 1–356. https://doi.org/10.5479/si.03629236.227.1
Pettibone, M. H. (1989). Two new species of Harmothoinae (Polychaeta: Polynoidae) from the East Pacific Rise, collected by Alvin dives 2000 and 2003. Proceedings of the Biological Society of Washington, 102,305–310.
Pratt, E. M. (1901). A collection of Polychaeta from the Falkland Islands. Manchester Memoirs, Memoirs and Proceeding of the Manchester Literary & Philosophical Society, 45, 1–18.
Rozbaczylo, N. (1985). Los anélidos poliquetos de Chile: Índice sinonímico y distribución geográfica de especies. Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Monografías Biológicas, 3, 1–284.
Sars, M. (1861). Om de ved Norges Kyster forekommende Arter af Annelideslaegten Polynoë. Forhandlinger i Videnskabs-Selskabet i Christiania, 1860, 54–62.
Uschakov, P. V. (1965). Polychaeta of the Far Eastern Seas of the U.S.S.R. Zoological Institute of the Russian Academy of Sciences, Keys to the Fauna of the U.S.S.R., 56, 1–419.
Wehe, T. (2006). Revision of the scale worms (Polychaeta: Aphroditoidea) occurring in the seas surrounding the Arabian Peninsula, 1. Polynoidae. Fauna of Arabia, 22, 23–197.
Wesenberg-Lund, E. (1962). Reports of the Lund University Chile Expedition 1948-49, 43. Polychaeta Errantia. Lunds Universitets Årsskrift, neue folge, Series 2, 57, 1–139.
Fernanda Chávez-Samayoa a, Jaime Antonio Escoto-Moreno a, Edmundo González-Santillán b, *
a Universidad Autónoma de Aguascalientes, Colección Zoológica, Av. Universidad 940, Ciudad Universitaria, 20100 Aguascalientes, Aguascalientes, Mexico
b Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Arácnidos, Tercer Circuito Universitario s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
Received: 18 October 2023; accepted: 19 February 2024
Abstract
The scorpion fauna of the state of Aguascalientes was revised based on specimens deposited in scientific collections and fieldwork. We evaluated the completeness of the scorpion species inventory of the Colección Zoológica of the Universidad Autónoma de Aguascalientes (CZUAA) based on nonparametric estimators. The results indicate that the number of species increased from 11 to 13, recording for the first time Chihuahuanus glabrimanus (Sissom & Hendrixson, 2005), a psammophilic species, and Centruroides balsasensis Ponce-Saavedra and Francke, 2004, a medically relevant species for public health. Finally, an identification key to the scorpion species of Aguascalientes is provided and a recount of ecological and biological traits of the species herein is studied.
Keywords: Chihuahuanus glabrimanus; Centruroides balsasensis; Species list; Parthenogenesis; Scientific collections
Análisis de riqueza y completitud de la escorpiofauna de Aguascalientes, México, con una clave de identificación de especies
Resumen
Se revisó la escorpiofauna del estado de Aguascalientes con base en ejemplares depositados en colecciones científicas y trabajo de campo. Se evaluó la completitud del inventario de especies de escorpiones de la Colección Zoológica de la Universidad Autónoma de Aguascalientes (CZUAA) mediante métodos no paramétricos. Los resultados indican un aumento del inventario de 11 a 13 especies; se registra por primera vez a la especie psamofílica Chihuahuanus glabrimanus (Sissom y Hendrixson, 2005) y la especie de importancia para la salud pública Centruroides balsasensis Ponce-Saavedra y Francke, 2004. Finalmente, se brinda una clave de identificación de especies de escorpiones de Aguascalientes, así como un recuento de caracteres ecológicos y biológicos de las especies estudiadas.
With an area of 5,589 km2 (INEGI, 2019), the state of Aguascalientes lies in the central region of Mexico. It is the fourth smallest state in Mexico with 11 municipalities (Sandoval-Ortega et al., 2017; Sigala-Rodríguez & Greene, 2009). Despite this overshadowing territory compared to other states in Mexico, such as Chihuahua, Aguascalientes comprises 3 biogeographic provinces: Sierra Madre Occidental (SMO), Chihuahuan Desert (CD), and Pacific Lowlands (PL) (Morrone et al., 2017) (Fig. 1). Three general types of climatic groups are associated with predominant plant communities in the state: arid zone covered by xerophytic vegetation, temperate zone with oak forest or mixed forests, and the tropical zone covered by secondary subtropical vegetation (Siqueiros-Delgado et al., 2017).
Although formal scorpion fauna studies have been done in the state, the first mention of scorpions distributed in Aguascalientes was presented by Hoffmann (1931, 1932, 1936), who listed 7 species in total: Centruroides infamatus C. L. Koch, 1844; Chihuahuanus bilineatus Pocock, 1898; Diplocentrus zacatecanus Hoffmann, 1931; Mesomexovis spadix Hoffmann, 1931; Paruroctonus gracilior Hoffmann, 1931; Thorellius intrepidus Thorell, 1876 and Vaejovis nigrescens Pocock, 1898. Later, several authors contributed to the knowledge of the species list (Contreras-Félix et al., 2015; Chávez-Samayoa et al., 2022; Escoto-Rocha & Delgado-Zaldívar, 2008), which is currently composed of 11 species. At present, there are no published identification keys for the scorpions of Aguascalientes, therefore researchers must rely on other works (González-Santillán & Prendini, 2013, 2018; Ponce-Saavedra & Francke, 2013a; Ponce-Saavedra et al., 2016), where part of the scorpion fauna is described and keyed.
Figure 1. Biogeographic provinces of the state of Aguascalientes (Morrone, 2017). Chihuahuan Desert (CD) is highlighted in blue, Pacific Lowlands (PL) in yellow, and Sierra Madre Occidental (SMO) in green.
Figure 2. Chelicerae of C. infamatus (A), M. spadix (B) and P. gracilior (C). Abbreviations: b, basal; d, distal; ed, external distal; id, internal distal; m, medial, sd, subdistal, vm, ventral median.
Established in 1978, the Colección Zoológica of the Universidad Autónoma de Aguascalientes (CZUAA), preserves specimens collected during the formation of biology students (De la Riva-Hernández, 2014). The first scorpion collected and deposited in the CZUAA dates to 1979. Subsequently, numerous biology students, professors, and the public have contributed with samples recording the state’s species richness. Escoto-Rocha and Delgado-Zaldívar (2008) presented a species list derived from the study of the specimens deposited in the CZUAA, where they listed 9 species. In this contribution we update the knowledge of the scorpion species distributed in Aguascalientes and provide an identification key at the species and genus levels for the first time. Furthermore, an evaluation of the alpha diversity using non-parametric estimators is computed. Although, examined material deposited in the CZUAA is the primary source, we studied scorpions deposited in the Colección Nacional de Arácnidos (CNAN) to corroborate taxonomical identities.
Materials and methods
To determine the species taxonomic identity, the following identification keys and species descriptions were used: Buthidae: Ponce-Saavedra and Francke (2013a), Ponce-Saavedra et al. (2016); Diplocentridae: Santibáñez-López and Francke (2013); Vaejovidae: Contreras-Félix and Francke (2019), González-Santillán and Prendini (2013, 2018), Sissom and González-Santillán (2004), Sissom and Hendrixson (2005). Digital microphotographs were taken with a Leica EZ4HD stereoscope and habitus with a Nikon Z7. All photographic edition was made with Adobe Photoshop CS5.1.
Nomenclature and homology follow González-Santillán and Prendini (2013), except for cheliceral dentition (Sissom, 1990). We proposed a name to identify a distinctive but unnamed denticle of the family Buthidae as the ventral medial denticle of the cheliceral fixed finger (Fig. 2A). This denticle lies between the distal and subdistal denticles but on the ventral surface. This denticle is a diagnostic character —and probably a synapomorphy— for the family Buthidae (Sissom, 1990) and thus is only present in the 2 species of the genus Centruroides.
Several authors have used museum and collection label data to estimate species richness through nonparametric estimators, overcoming the fact that this kind of data suffers from a standardized sampling effort, but at the same time, recognizing the value of this information as an estimate of inventory completeness (Basualdo, 2011; Beck & Kitching, 2007; Fattorini, 2013; Guralnick & Van Cleve, 2005; Meier & Dikow, 2004; Petersen & Meier, 2003; Petersen et al., 2003; Soberón-Mainero et al., 2000).
Collection records deposited in the CZUAA were georeferenced remotely using Google Earth Pro 7.3.4.8248 and, in some cases, directly registered during fieldwork using a Garmin eTrexH GPS. We generated 2 incidence databases, one considering spatial and another temporal species accumulation. For the spatial perspective, we created a layer with a grid of 0.025° × 0.025° —approximately 10 × 10 km— cells, recording the appropriated incidence (0 = absence, 1 = presence) per species (Appendix 1). For the temporal species accumulation, we divided the species incidence into classes of 5 years from 1979 to 2022 (Appendix 2). Diversity analyses were conducted using the function specpool of the R package vegan (Oksanen et al., 2019). Centruroides balsasensis was excluded from this analysis because we consider it an introduced species. Distributional maps with an elevation model (Fick & Hijmans, 2017) were generated with QGIS 3.16.
Acronyms used in the records are as follow: AMCC, Ambrose Monell Cryocollection at AMNH; AMNH, American Museum of Natural History; BM, British Museum; CAS ARA, California Academy of Sciences, Arachnid collection; CIBUAEM, Colección de Entomología del Laboratorio de Parasitología Vegetal del Centro de Investigaciones Biológicas de la Universidad Autónoma del Estado de Morelos; CNAN-SC, Colección Nacional de Arácnidos, Scorpion Collection; CZUAA SCO, Colección Zoológica de la Universidad Autónoma de Aguascalientes, Scorpion Collection; ENCB IPN, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional; FSCA, Florida State Collection of Arthropods; GL, Graeme Lowe’s personal collection; INDRE, Instituto Nacional de Diagnóstico y Referencia Epidemiológica; JCC, J. C. Cockendolpher’s personal collection; MES, Michael E. Soleglad’s personal collection; MNHN, Museum National d´ Historie Naturelle; MWSU, Midwestern State University; SAM, Indiana University School of Medicine; SAS, S. A. Stockwell’s personal collection; TMM, Texas Memorial Museum; TOR, Museo ed Instituto di Zoologia Sistematica della Universita di Torino, Italy; UAQ, Universidad Autónoma de Querétaro; UCLB, University of California Long Beach; VF, Victor Fet’s personal collection; WDS, W. D. Sissom’s personal collection; WTAMU, West Texas A & M University. Notice that some records from the literature are devoid of numerical code because they were not found in the CNAN despite the authors listing them in their publications.
Results
We examined 934 specimens and identified 13 species corresponding to the families Buthidae, Diplocentridae, and Vaejovidae (Table 1). The results of analyses of non-parametric estimators using spatial and temporal approaches reflected similar completeness of Aguascalientes biodiversity (Table 2). Under the 10 × 10 km grid approach, all estimators produced coverage of over 80%, the lowest for first order Jackknife at 85.7% and the highest for Jackknife 2 at 100%. On the other hand, the estimation constructed through the 5-year classes lays a higher percentage of coverage, the lowest for Bootstrap at 92.3% and the highest Jackknife 2 at 100%. The overall most conservative estimation was 85.7%, and the least conservative one was 100%.
Nonparametric estimators of species richness. Abbreviations as follows: Boot = bootstrap estimator; Boot.se = standard error for bootstrap estimator; Chao = Chao estimator; Chao.se = standard error for chao estimator; DB = database, (1) 10 × 10 km grids as spatial units, (2) 5 years classes; Jack1 = first order Jackknife; Jack1.se = standard error for first order Jackknife; Jack2 = second order Jackknife; N = number of units; Sp = number of observed species.
DB
Sp.
Chao
Chao.se
Jack1
Jack1.se
Jack2
Boot
Boot.se
N
1
12
12.24
0.71
12.98
0.98
12.04
12.72
0.77
61
2
12
12
0
12
0
10.63
12.24
0.72
9
Figure 3. Habitus of female of C. balsasensis, scale = 1 cm.
Holotype specimen: was not localized in the CNAN. The only available information from the original description is as follow: “El holotipo es un macho adulto recolectado en Churumuco, municipio del mismo nombre en Michoacán, México el 08 de junio de 2000 por Erwin P. Miranda López y será depositado en la Colección Nacional de Arácnidos del Instituto de Biología de la Universidad Nacional Autónoma de México (CNA-IBUNAM)”.
Records from literature: Córdova-Athanasiadis (2005): México, Morelos, Municipality of Amacuzac: Huajuntlán, 18°36’36.00” N, 99°25’48.00” W, 1,510 m, 4-09-2004, M. Córdova, A. Gotilla, 5 ♀, CNAN-SC106. San Gabriel de las Palmas, 18°36’36.00” N, 99°21’00.00” W, 1,420 m, 4-09-2004, M. Córdova, A. Gotilla, 8 ♀, CNAN-SC131. Municipality of Axochiapan: Quebrantadero, 18°31’48.00” N, 98°47’24.00” W, 1,280 m, 2-09-2003, M. Córdova, D. Morán, 8 ♀, 6 ♂, 2 juv., CNAN-SC105. Municipality of Coatlán del Río: El Oyanco, 18°43’48.00” N, 99°25’48.00” W, 1,022 m, 6-08-2004, M. Córdova, O. Sotelo, 17 ♀, 6 ♂, CNAN-SC132. Municipality of Emiliano Zapata: Temimilcingo, 18°44’24.00” N, 99°09’00.00” W, 1,060 m, 28-04-2004, M. Córdova, A. Jaimes, 6 ♀, 4 ♂, CNAN-SC74. Municipality of Jonacatepec: Chalcatzingo, 18°40’48.00” N, 98°46’12.00” W, 1,371 m, 5-04-2004, M. Córdova, D. Morán, 11 ♀, 6 ♂, CNAN-SC978. Las Trancas, 13-11-1938, 2 ♀, CNAN-SC185. Municipality of Miacatlán: Palpan, 18°51’00.00” N, 99°25’12.00” W, 1,587 m, 7-08-2004, M. Córdova, O. Vázquez, 3 ♀, CNAN-SC93. Municipality of Temixco: Pueblo Viejo, 18°52’12.00” N, 99°15’00.00” W, 1,378 m, 11-06-2004, M. Córdova, A. Jaimes, O. Sotelo, 6 ♀, 6 ♂, 1 juv., CIBUAEM. Municipality of Tepalcingo: Cerro del Venado, 18°36’36.00” N, 98°51’36.00” W, 1,380 m, 1-09-2003, M. Córdova, D. Morán, 17 ♀, 9 ♂, CNAN-SC80. El Limón, 18°32’24.00” N, 98°55’48.00” W, 1,191 m, 12-09-2004, M. Córdova, G. Obregón, 4 ♀, 7 ♂, CIBUAEM. Municipality of Tlaltizapán: Barranca Honda, 18°49’12.00” N, 99°06’00.00” W, 1,200 m, 20-06-2004, M. Córdova, M. Barrios, 4 ♀, 2 ♂, CNAN-98; M. Córdova, M. Barrios, 1 ♀, 18 juv., CNAN. Municipality of Tlalquiltenango: Chimalacatlán, 18°27’36.00” N, 99°06’00.00” W, 1,066 m, 11-07-2004, M. Córdova, O. Sotelo, 4 ♀, 5 ♂, CNAN-SC976. Huautla, 18°23’24.00” N, 99°01’48.00” W, 976 m, 12-07-2004, M. Córdova, A. Jaimes, 17 ♀, 8 ♂, 1 juv., CNAN-SC81; 1 ♀, 24 juv., CNAN; 18°26’24.00” N, 99°01’12.00” W, 11-06-1994, A. Burgos, 3 ♀, CIBUAEM. Quilamula, 18°31’12.00” N, 99°01’12.00” W, 1,100 m, 7-06-2004, M. Córdova, E. Maison, 2 ♀, 3 juv., CIBUAEM; 15-08-2003, M. Córdova, A. Jaimes, H. Lagunas, 11 ♀, 9 ♂, 2 juv., CNAN-SC165. Xicatlacotla, 18°33’36.00” N, 99°13’48.00” W, 812 m, 26-06-2004, M. Córdova, 1 ♀, 28 juv., CNAN-SC917; 3♀, CIBUAEM. Xalostoc, 18°43’12.00” N, 98°54’00.00” W, 1,602 m, 21-11-2003, M. Córdova, 6 ♀, 1 ♂, CNAN-SC159. Municipality of Yautepec: San Isidro, 18°49’12.00” N, 99°06’00.00” W, 1,378 m, 20-06-2004, M. Córdova, M. Barrios, 5 ♀, 1 ♂, CNAN-SC96; 1 ♀, 19 juv., CNAN. Municipality of Zacatepec: Galeana, 15-08-1991, G. Peña, 4 ♀, 3 ♂, 2 juv., CIBUAEM. Zacatepec, 30-06-1981, F. Guerrero, 3 ♀, CNAN-SC118. Municipality of Zacualpan de Amilpas: El Curiel, 18°46’48.00” N, 98°45’36.00” W, 1,668 m, 31-07-2004, M. Córdova, O. Sotelo, A. Sotelo, 18 ♀, 4 ♂, 1 juv., CNAN-SC79. Ponce-Saavedra & Francke (2004): México, Guerrero, Municipality of Tzirándaro: 18°28’25” N, 100°57’17” W. Michoacán, Municipality of Apatzingán: Altamira, El Tesorero, 18°57’42” N, 102°22’58” W. Las Anonas, 18°51’36” N, 102°35’00” W. Valle Verde, 19°02’54” N, 102°23’18” W. Municipality of Arteaga: El Descansadero, 19°34’17” N, 101°49’17” W. El Puerto, road to Tumbiscatío, 18°09’18” N, 102°23’18” W. Las Cañas, 18°33’20” N, 101°58’19” W. Municipality of Buenavista Tomatlán: Buenavista, 19°120’24” N, 102°35’12” W. El Puerto, 19°14’48” N, 102°35’37” W. Municipality of Carácuaro: 19°00’54” N, 102°07’05” W. Km 4 from San Antonio de las Huertas towards Carácuaro, 19°06’13” N, 101°12’06” W. Municipality of Churumuco: Cerro de Las Letras, 18°39’37” N, 101°39’17” W. Churumuco, 18°39’48” N, 101°38’64” W. El Ahuijote, 18°42’30” N, 101°46’00” W. Potrero de Corpus, El Chocolate, 18°52’48” N, 101°37’47” W. Municipality of Gabriel Zamora: Cerro de la Cruz, 19°09’55” N, 102°04’03” W. El Jagüey, 19°10’49” N, 102°01’24” W. El Ranchito, 19°11’00” N, 102°03’11” W. Km 128 Morelia-L.C road, 19°04’51” N, 102°00’59” W. Municipality of Huacana: Presa de Zicuirán, 18°55’18” N, 101°55’49” W. Municipality of Huetamo: road to Baztán, near El Rodeo, 18°46’13” N, 100°59’54” W. Cerro de Turitzio, near Arúa, 18°31’49” N, 100°55’45” W. Municipality of Mújica: Km 135 road La Huacana-Cuatro Caminos, 18°31’49” N, 100°55’45” W. Nueva Italia, 19°01’24” N, 102°05’30” W. Municipality of Nuevo Urecho: breach towards Nueva Jerusalem, 19°05’06” N, 101°30’15” W. Km 3.8 road Puruarán-Turicato, 19°’05’48” N, 101°30’24” W. Municipality of Parácuaro: La Tuna, 19°05’48” N, 102°13’55” W. Municipality of San Lucas: 18°35’12” N, 100°47’11” W. Municipality of Tacámbaro: Arroyo Frío, 19°10’53” N, 101°28’30” W. Municipality of Taretán: 19°20’00” N, 101°55’05” W. Municipality of Tepalcatepec: El Taixtán, 19°00’49” N, 102°59’12” W. Estado de México, Municipality of Tejupilco: 1 km SE of Mango Matus, 18°40’00” N, 100°26’39” W. Quijano-Ravell & Ponce-Saavedra (2016): México, Michoacán, Municipality of Churumuco: 18°39’39.00” N, 101°38’46.00” W, 4 ♀, 3 ♂. Municipality of Huetamo: Arúa, 18°32’25.00” N, 100°55’35.00” W, 3 ♀, 4 ♂. Reyes-Moya et al. (2021): Mexico, Chihuahua, Municipality of Ciudad Juárez: 31°38’31” N,106°26’34.4” W, 03/04-2019.
Examined material: México, Aguascalientes, Municipality of Aguascalientes: Universidad Autónoma de Aguascalientes, 18-05-2012, N. Ubario-Guevara, 1 ♀, CZUAA SCO-107; 15-05-2016, A. A. Martínez-Durón, 1 ♀, CZUAA SCO-411. Jardín Botánico Rey Netzahualcóyotl, 25-04-2014, J. E. Gutiérrez-Montoya, 1 ♀, CZUAA SCO-106; 04-2015, J. E. Díaz-Plascencia, 3 ♀, CZUAA SCO-108. Fraccionamiento Casa Blanca, 13-04-2016, M. A. Delgado, 1 ♀, CZUAA SCO-434. Fraccionamiento El Dorado, 14-10-2019, 1 ♀, CZUAA SCO-551. Tlaxcala #107, colonia San Marcos, 21-06-2020, V. Esparza, 4 ♀, CZUAA SCO-535. Pozo Hondo 141, colonia Pozo Bravo, 29-06-2020, N. Palacios, 1 ♀, CZUAA SCO-546. Tlaloc 519, colonia Prados del Sur, 8-03-2022, I. Martínez, 1 ♀, CZUAA SCO-672. Municipality of Calvillo: Colomos, plantación de guayaba, 21°52’44.11” N, 102°39’16.70” W, 1,798 m, 27-01-2022, I. J. Rodríguez-Elizalde, J. Brito-Ruiz de Velasco, 1 ♀, CZUAA SCO-686; 29-08-2021, I. J. Rodríguez-Elizalde, J. Brito-Ruiz de Velasco, L. I. Rodríguez-Rodríguez, M. E. Herrera-Rodríguez, 1 ♀, CZUAA SCO-692. El Terrero, 17-04-2011, J. Ruvalcaba-Calderón, 1 ♀, CZUAA SCO-104. Los Adobes, 21°48’39.22” N, 102°41’21.34” W, 1,875 m, 17-09-2019, F. Chávez-Samayoa, L. R. Haro-Tapia, 4 ♀, CZUAA SCO-509; 21°48’39.22” N, 102°41’57.34” W, 1,875 m, 11-10-2019, F. Chávez-Samayoa, 2 ♀, CZUAA SCO-698. Calvillo, 10-11-2021, R. García, 2 ♀, CZUAA SCO-670. Municipality of Jesús María: Alegría #122, colonia Gómez Portugal, 23-03-2022, 1 ♀, CZUAA SCO-695. Cerro del Muerto, 8-05-2005, A. Ramírez, 1 ♀, CZUAA SCO-111. Michoacán, Municipality of Uruapan: F. A. Guerrero-Roque, 1 ♀, CZUAA SCO-54. Zacatecas, Municipality of Jalpa: 21-04-2016, V. E., 1 ♀, CZUAA SCO-407. Municipality of Juchipila: 25-04-2010, C. Y. Gómez-Aranzazú, 2 ♀, CZUAA SCO-102; 13-05-2000, D. Magaña-Ortíz, 1 ♀, CZUAA SCO-110.
Remarks
Other than the Churumuco site (18°39’48” N, 101°38’64” W), in the Michoacan Balsas Depression, C. balsasensis has been recorded in the states of Guerrero, Mexico, Morelos, and Puebla (Córdova-Athanasiadis, 2005; Ponce-Saavedra & Francke, 2004; Ponce-Saavedra et al., 2022; Santibáñez-López et al., 2015). However, the records from Morelos are questionable (González-Santillán & Possani, 2018).
Reyes-Moya et al. (2021) recently recorded C. balsasensis in Ciudad Juárez, Chihuahua, indicating that the species collected may represent an “accidental introduction” into the city likely due to (Ciudad Juárez is) “the northern largest and busiest border crossing of Mexico and the USA, experiencing massive local and non-local human transit”. We were able to examine some of the specimens cited in that contribution and concluded that they are not conspecific with C. balsasensis. We base our conclusion on the comparison of the following set of the characters readily observed in their published figures: 1) there is an areola created by pigmentation circling each median ocellus on the carapace in Reyes-Moya et al. (2021, their Fig. 1A), whereas in the original description of Ponce-Saavedra & Francke, 2004 (their Fig. 9), in Quijano-Ravell & Ponce-Saavedra 2016 (their Fig. 6[2]), and in our figure 4A, the areola is not complete with the retrolateral area lacking pigmentation; 2) the body length of the females in Reyes-Moya et al. (2021) is 37 and 40 mm, whereas in the original description, the body length is 60 and 73 mm, a difference of more than 20 mm; 3) the pectens’ basal piece of the females is wider than longer (Reyes-Moya et al., 2021), but those of the original description are trapezoid, almost as long as wide, with a deep medial depression. The description of the specimens provided by Reyes-Moya et al. (2021), however, is more like Centruroides sculpturatus, particularly due to the pigmentation pattern of the carapace and tergites exhibited by phase 2 or 3 identified by Stahnke (1971). The likelihood of introduction from the Balsas Basin to Ciudad Juárez is considerably lower compared to the introduction of this species from Sonora or even Arizona because of the proximity of these areas. Our conclusion is that the specimens reported and illustrated by Reyes-Moya et al. (2021) are conspecific to C. sculpturatus instead of C. balsasensis.
Figure 4. Carapace of C. balsasensis (A) and C. infamatus (B).
We recorded C. balsasensis for the first time in Aguascalientes with 20 samples (32 individuals) from the Municipalities of Aguascalientes, Calvillo, and Jesús María. The first sample dates to 2005 (CZUAA SCO 111), and the most recent one to May 2023 (CZUAA SCO-732). Specimens deposited in the CZUAA were associated mainly with urban areas or meadows. Remarkably, we only observed adult or juvenile females; males are absent in the collection. It is likely that this species was transported accidentally and has invaded Aguascalientes and other northern states (we also examined 3 samples from Zacatecas: CZUAA SCO-102, CZUAA SCO-110, CZUAA SCO-407). However, further investigation to elucidate the establishment of this species is needed. Our data also suggest that the establishment of the population in Aguascalientes may have occurred by parthenogenesis due to the presence of only females in colonized urban and suburban areas. This hypothesis finds support by the occurrence of the same phenomenon in the South American species Tityus serrulatus, Lutz and Mello, 1922, which exhibits indigenous sexual populations and asexual populations in colonized habitats (Braga-Pereira & Santos, 2021). Intriguingly, this is also a medically relevant species in Brazil, suggesting parallel ecological mechanisms to establish in novel locations in different parts of the world.
Centruroides infamatus (C. L. Koch, 1844) (Fig. 5)
Taxonomic summary
Holotype specimen: Zoologisches Museum, Humbolt Universität, Berlin, Germany. Sex and type locality unknown (Fet & Lowe, 2000).
Records from literature: Esposito et al. (2018): México, Michoacán, Municipality of Tangamandapio: Los Tabanos, 19°58’29.64” N, 102°50’32.14” W, 223 m, 31-05-2006, O. F. Francke, H. Montaño, A. Valdez, A. Ballesteros, 2 ♀, AMCC (LP6420). Municipality of Tingambato: Tingambato 4 km W, under stones, 23-03-2000, E. González, 1 ♂, AMCC (LP1822). Ponce-Saavedra & Francke (2004): México, Guanajuato, Municipality of Romita: San Antonio del Pochote, 20°55’47.00” N, 101°33’48.00” W. Michoacán, Municipality of Coalcomán: 18°48’47.00” N, 103°09’57.00” W. Municipality of Morelia: 19°42’00.00” N, 101°11’18.00” W. Tiripetío, 19°32’00.00” N, 101°21’00.00” W. Municipality of Salvador Escalante: Salvador Escalante, 19°24’00.00” N, 101°38’18.00” W. Municipality of Uruapan: La Cofradía, 19°26’00.00” N, 102°01’00.00” W. Zumpimito, 19°22’18.00” N, 102°03’19.00” W. Ponce-Saavedra et al. (2009): México, Michoacán, Municipality of Uruapan: Zumipito, 19°21’25.00” N, 102°04’13,00” W, GenBank AF439753. Quijano-Ravell & Ponce-Saavedra (2016): México, Guanajuato, Municipality of León: 21°01’02.00” N, 101°37’44.00” W, 3 ♂ 2 ♀. Michoacán, Muncipality of Uruapan: Zumipito, 21°07’30.00” N, 101°01’49.00” W, 4 ♂ 4 ♀.
Figure 5. Habitus of male (top) and female (bottom) of C. infamatus, scale = 1 cm.
Examined material: Mexico, Aguascalientes, Municipality of Aguascalientes: Central de abastos, 02-1996, J. G. Estrada-A., 1 ♂, CZUAA SCO-54. Cerro de los Gallos, 21-10-1989, L. Silva-López, 1 ♀, CZUAA SCO-117; 26-04-1984, H. Gallegos-Rangel, 1 ♀, CZUAA SCO-118. Municipality of Calvillo: Cascada Los Huenchos, 21°52’52.90” N, 102°47’06.25” W, 1756 m, 22-05-2021, J. L. Reyes-Hernández, 1 ♂, CZUAA SCO-614. Cercano a balneario “La Cueva”, 21°48’47.70” N, 102°49’34.82” W, 28-09-2019, F. Chávez-Samayoa, L. R. Haro-Tapia, 1 ♀, CZUAA SCO-483; 15-10-2019, F. Chávez-Samayoa, 19 ♂, 10 ♀, CZUAA SCO-495; 19-10-2019, F. Chávez-Samayoa, 23 ♂, 12 ♀, CZUAA SCO-503. Cercano a restaurante “La Fragua”, 21°59’52.32” N, 102°40’35.36” W, 1,825 m, -05-2021, T. Huerta, 3 ♂, 2 ♀, CZUAA SCO-644. El Cuervero, 19-06-2004, A. D. González, 1 ♀, CZUAA SCO-162. El Zapote, 8-12-2021, R. García, 1 ♀, CZUAA SCO-666. Calvillo, 29-04-2017, E. M. Flores-Villalpando, 1 ♀, CZUAA SCO-431; 4-05-2017, A. Hernández, 1 ♀, CZUAA SCO-435. Crucero de las Pilas, 11-11-2020, R. García, 1 ♂, CZUAA SCO-700. La Calixtina, 16-05-2017, I. Rodríguez-Ortiz, 1 ♀, CZUAA SCO-424. La Labor, 10-04-2011, M. P. Ramírez-Guillen, 1 ♂, CZUAA SCO-139. La Mezquitera, 21°51’37.47” N, 102°46’04.20” W, 30-08-2019, F. Chávez-Samayoa, J. A. Escoto-Moreno, 1 ♂, 1 ♀, CZUAA SCO-484; 5-10-2019, F. Chávez-Samayoa, 23 ♂, 8 ♀, CZUAA SCO-508; 7-09-2019, F. Chávez-Samayoa, R. Haro-Tapia, 4 ♂, 2 ♀, CZUAA SCO-514; 10-10-2019, F. Chávez-Samayoa, 2 ♂, CZUAA SCO-558. Los Adobes, 21°48’39.22” N, 102°41’21.34” W, 1-09-2015, J. E. Díaz-Plascencia, 1 ♀, CZUAA SCO-330; 11-10-2019, F. Chávez-Samayoa, 15 ♂, 4 ♀, CZUAA SCO-480; 17-09-2019, F. Chávez-Samayoa, R. Haro, 26 ♂, 13 ♀, CZUAA SCO-497; 6-09-2019, J. A. Escoto-Moreno, F. Chávez-Samayoa, 1 ♂, CZUAA SCO-555. Los Alisos, 9-10-1989, J. Moreno-Lara, 1 ♂, CZUAA SCO-121; 2-06-1982, Z. M. Vela-Ortiz, 1 ♂, CZUAA SCO-133; 8-09-1984, Morales, Franco, Montoya, 1 ♂, CZUAA SCO-134; 19-06-2004, G. Guerrero, 1 ♀, CZUAA SCO-154. Los Lobos, 21-10-1990, J. A. Sánchez-García, 1 ♀, CZUAA SCO-129. Presa Malpaso, 19-06-2004, G. Villalobos, 1 ♂, CZUAA SCO-123; 3-05-1999, R. P. Arteaga-Hernández, 1 ♂, CZUAA SCO-125; 13-06-2004, E. Ortiz-Martínez, 1 ♂, CZUAA SCO-126; 20-03-1999, M. Morga, 1 ♂, CZUAA SCO-131; 3-05-1999, M. Martínez-Garnica, 1 ♀, CZUAA SCO-135; 26-11-1986, E. Alvizo-Flores, 1 ♀, CZUAA SCO-138. Palo Alto, 18-11-2021, R. García, 1 ♂, 3 ♀, CZUAA SCO-671; 20-11-2021, R. García, 4 ♂, CZUAA SCO-667. Presa La Codorniz, 22°00’39.07” N, 102°40’15.19” W, 1,894 m, 1-11-2020, E. González-Martínez, J. L. Reyes-Hernández, I. E. Gallegos-Vieyra, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 5 ♂, 5 ♀, CZUAA SCO-589; 21°59’50.78” N, 102°40’42.78” W, 2,068 m, 5-06-2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, C. Huerta, 5 ♂, 10 ♀, CZUAA SCO-612. Presa La Ordeña, 25-04-2005, C. A. Díaz-Delgado, 1 ♀, CZUAA SCO-145. Presa de los Serna, 20-10-2003, S. García, 1 ♀ CZUAA SCO-124; 14-06-2004, S. García, 1 ♀, CZUAA SCO-137. San Tadeo, -04-1996, J. Martínez-de Lara, 1 ♀, CZUAA SCO-127; -05-2015, C. A. Franco-Servín de la Mora, 1 ♂, 2 ♀, CZUAA SCO-149. Sierra del Laurel, 25-10-1982, E. Padilla-R., 1 ♂, CZUAA SCO-128. Terrero de la Labor, 22°01’32.02” N, 102°39’58.00” W, 22-10-2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, 25 ♂, 20 ♀, CZUAA SCO-655. Municipality of Jesús María: Cerro El picacho, 23-05-2014, E. A. Hernández-Medrano, 1 ♂, CZUAA SCO-109; 29-04-2016, D. Martínez, 1 ♂, CZUAA SCO-418. Municipality of Rincón de Romos: 17-04-2004, N. Arroyo-Chávez, 1 ♂, CZUAA SCO-112; 17-04-2004, A. F. Ramos-Martínez, 1 ♂, CZUAA SCO-113. Guanajuato, Municipality of Comonfort: 04-2007, D. García, 1 ♂, CZUAA SCO-402. Municipality of León: Fraccionamiento La Luz, 27-04-2013, V. H. González-Sánchez, 1 ♀, CZUAA SCO-75; 27-04-2003, K. A. de Luna-Gómez, 1 ♀, CZUAA SCO-85. León, 30-05-2009, C. García-Balderas, 1 ♀, CZUAA SCO-77; 13-06-2004, C. Moreno, 1 ♂, CZUAA SCO-84; 21-06-2004, M. A. Gómez, 1 ♂, CZUAA SCO-87; 4-06-2000, M. Chávez-Andrade, 1 ♂, CZUAA SCO-88; 7-06-2004, S. Flores, 1 ♂, CZUAA SCO-90; 15-06-2004, M. A. B. López, 1 ♂, CZUAA SCO-94; 16-06-2006, Miguel, 1 ♀, CZUAA SCO-96; R. A. Carbajal-Márquez, 1 ♀, CZUAA SCO-99. Municipality of Moroleón: Presa Quiahuyo, 25-03-2016, K. Rosales, 1 ♀, CZUAA SCO-445. Municipality of Pénjamo: 21-09-1983, O. A. Rodríguez, M. G. Flores, 1 ♀, CZUAA SCO-79. Municipality of Salamanca: L. Flores, 1 ♂, CZUAA SCO-70; J. A. Berlín-Diosdado, 1 ♀, CZUAA SCO-71. Municipality of Yuriria: 13-06-1991, V. Villalobos-Sánchez, 1 ♀, CZUAA SCO-74. Michoacán, Municipality of Uruapan: M. A. Domínguez-de la Riva, 1 ♀, CZUAA SCO-51. Municipality of Tancítaro: El Cuate, -04-1996, R. E. Martínez, 1 ♀, CZUAA SCO-58. Zacatecas, Municipality of Guadalupe Victoria: 9-11-1986, M. Torres-Romero, 1 ♂, CZUAA SCO-60. Municipality of Juchipila: 16-04-2016, K. I. Molina, 1 ♂, CZUAA SCO-413; 25-03-2016, E. A. Bernal-Montoya, 1 ♀, CZUAA SCO-425; 15-05-2016, A. Barba, 1 ♂, CZUAA SCO-448. Municipality of Nochistlán: 2-03-2010, E. F. Pulido-Hornedo, 1 ♀, CZUAA SCO-53. Municipality of Tabasco: 10-06-2010, A. Lara, 1 ♂, CZUAA SCO-57. Municipality of Valparaiso: 4-04-2017, S. D. López-González, 1 ♂ CZUAA SCO-416.
Remarks
Known records for C. infamatus include the southern half of Aguascalientes, northwestern Guanajuato, Jalisco, Michoacán, and the southern third of Zacatecas (González-Santillán & Possani, 2018). State records without precise locality information were published for Colima, Durango, Nayarit, Oaxaca, Puebla, Querétaro, and Sinaloa (Ponce-Saavedra & Francke, 2013a; Ponce-Saavedra et al., 2022; Santibáñez-López et al., 2015).
Centruroides infamatus is a component of Aguascalientes species richness, and before our work, it was the only species of medical importance reported for the state. This species inhabits the area corresponding to the Trans Mexican Volcanic Belt biogeographic province. However, its distribution in Aguascalientes is restricted to the southwestern municipalities, most abundantly in Calvillo, which corresponds to the lower lands of the Pacific biogeographic province. Centruroides infamatus was recorded in high densities in altitudes between 1,700 and 2,100 m on the slopes of a canyon formed by the Sierra del Laurel and Sierra Fría. We hypothesized that as part of the diversification of the infamatus species group of the genus Centruroides (sensu Ponce-Saavedra & Francke, 2019), the populations established in this area of Aguascalientes and most likely in Zacatecas, may represent a separate species from the Trans Mexican Volcanic Belt biogeographic province populations due to their geographical disjunction created by the Sierra del Laurel to the east and the arid zone corresponding to the Chihuahuan desert, north to the Aguascalientes City. In another scenario, we surmise that this species may be a component of the Pacific Lowlands province that has penetrated the mainland and most likely represents a distinctively independent taxon from the populations inhabiting the Trans Mexican Volcanic Belt. Further molecular and ecological studies may clarify these hypotheses.
Figure 6. Habitus of male (top) and female (bottom) of D. zacatecanus, scale = 1 cm.
Diplocentridae Karsch, 1880
Diplocentrus zacatecanus Hoffmann, 1931 (Fig. 6)
Taxonomic summary
Lectotype: deposited in the CNAN. México: Aguascalientes: Tepezalá (22°13.3629’ N, 102°10.0149’ W, 2,100 m.), no date or collector 1 ♂ (CNAN-T0761). Paralectotypes: 2 ♂♂ and 1 ♀ (CNAN-T0762) same locality and collection event (Santibáñez-López & Francke, 2013).
Records from literature: Hoffmann (1931): México, Aguascalientes, Municipality of Tepezalá. Zacatecas, Municipality of Pinos. Ponce-Saavedra et al. (2009): México, Aguascalientes, Municipality of Tepezalá: 22°13’20.94” N, 102°10’17.53” W, 2100 m. Durango, Municipality of San Juan de Guadalupe: Loma Alta, 24°37’58.05” N, 102°45’2.80” W, 1,560 m. Vicente Guerrero, 23°43’53.99” N, 103°59’14.55” W, 1,922 m. Estado de México, Municipality of Aculco: La Loma, 20°06’07.03” N, 99°51’25.71” W, 2,450 m. Municipality of Soyaniquilpan: San José Deguedo, 20°05’09.04” N, 99°33’48.78” W, 2,514 m. Guanajuato, Municipality of Coroneo, 20°11’54.81” N, 100°21056.60” W, 2,280 m. Hidalgo, Municipality of Tula de Allende: Michimaloya, 20°05’34.88” N, 99°23’57.93” W, 2,078 m. Michoacán, Municipality of Contepec: 19°57’10.44” N, 100°09’48.90” W, 2,480 m. Querétaro, Municipality of Cadereyta: Cadereyta, 20°41’38.29” N, 99°48’36.21” W, 2037 m. El Organal, 20°37’09.49” N, 99°38’27.98” W, 1,733 m. Vizarrón de Montes, 20°50’03.63” N, 99°43’09.27” W. Municipality of Huimilpan: 20°22’22.23” N, 100°16’27.48” W, 2,288 m. San Luis Potosí, Municipality of Villa de Ramos: 22°49’53.21” N, 101°54’36.01” W, 2,200 m. Zacatecas, Municipality of Cañitas de Felipe Pescador: 23°36’20.10” N, 102°43’47.50” W, 2,027 m. Municipality of Sain Alto: Emiliano Zapata, 23°34’16.66” N, 103°18’02.75” W, 2,067 m. Municipality of Zacatecas: 22°46’19.52” N, 102°34’32.01” W, 2,440 m. Santibáñez-López and Francke (2013): México, Aguascalientes, Municipality of Tepezalá: 22°13’21.77” N, 102°10’00.89” W, 2,100 m, CNAN-T0761, lectotype, 1 ♂; CNAN-T0762, paralectotypes, 2 ♂, 1 ♀; 1 km N, 22°14’20.88” N, 102°10’28.02” W, 2,048 m, 4 ♂, CNAN-S03075, 4-07-2005, O. F. Francke, J. Ponce-Saavedra, M. Córdova, A. Jaimes, G. Francke, V. Capovilla, 2 ♂, AMNH. Zacatecas, Road Sombrerete-Durango: km 179, 23°40’47.88” N, 103°41’42.72” W, 2,448 m, 9-08-2005, O. F. Francke, W. D. Sissom, C. Lee, K. McWest, L. Jarvis, C. Durán, H. Montaño, A. Ballesteros, 3 ♀, CNAN-SC1731; 3 ♀, AMNH.
Examined material: Mexico, Aguascalientes, Municipality of Asientos: Asientos, 10-02-2013, V. Martínez-Salazar, 1 ♀, CZUAA SCO-06; 17-03-2013, C. Franco-Servín de la Mora, 1 ♀, CZUAA SCO-10; 22-02-2013, R. A. Rosales, 1 ♀, CZUAA SCO-12. Municipality of El Llano: Juan el Grande, 21°56’21.00” N, 101°55’06.00” W, 2,495 m, 18-10-2019, N. A. Pulido-Hornedo, X. A. Ruiz, 3 ♀, CZUAA SCO-473; 18-10-2019, F. Chávez-Samayoa, 4 ♂, 6 ♀, CZUAA SCO-475; 21°55’54.55” N, 101°56’09.28” W, 2,117 m, 22-09-2019, F. Chávez-Samayoa, 2 ♂, 4 ♀, CZUAA SCO-490; 21°56’14.57” N, 101°55’24.35” W, 2,285 m, 1-06-2020, P. C. Hernández-Romero, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-541; 21°56’21.08” N, 101°55’17.87” W, 2,395 m, -10-2019, E. González-Martínez, 1 ♂, 2 ♀, CZUAA SCO-543; 22-01-2021, F. Chávez-Samayoa, A. Palacios, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-633; 21°56’14.57” N, 101°55’24.35” W, 2,284 m, -04-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-645; -03-2021, F. Chávez-Samayoa, D. M. Reyes-Páramo, 1 ♀, CZUAA SCO-652. Municipality of Tepezalá: Cerro del Capulín, 22°14’45.06” N, 102°09’14.90” W, 2,189 m, 6-10-2019, F. Chávez-Samayoa, I. Villalobos-Juárez, E. González-Martínez, 2 ♂, 2 ♀, CZUAA SCO-488; 13-10-2019, F. Chávez-Samayoa, 5 ♀, CZUAA SCO-505; 22°14’01.00” N, 102°09’27.58” W, 2,130 m, 21-08- 2020, F. Chávez-Samayoa, 2 ♀, CZUAA SCO-525; 30-11-2020, I. Villalobos-Juárez, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-595; 25-04-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, I. Villalobos-Juárez, 1 ♂, CZUAA SCO-662. Tepezalá, -04-2006, O. Ortega, 1 ♂, CZUAA SCO-01; -03-2006, M. de la Riva, 1 ♂, CZUAA SCO-02; 27-04-2006, L. Reséndiz, 1 ♂, CZUAA SCO-04; -04-2006, R. A. Carbajal, 1 ♀, CZUAA SCO-07; -06-2006, L. Flores, 1 ♀, CZUAA SCO-09; -06-2006, J. Salazar-López, 1 ♀, CZUAA SCO-11; 15-06-2006, E. Medina-Alcantar, 1 ♂, CZUAA SCO-15; 15-06-2006, V. Revilla-Castellanos, 1 ♀, CZUAA SCO-16; 28-02-2006, M. Domínguez-De la Riva, 1 ♀, CZUAA SCO-17; -03-2006, D. Rendón, 1 ♀, CZUAA SCO-18; -05-2006, F. A. Guerras-Roque, 1 ♀, CZUAA SCO-20. San Luis Potosí, Muncipality of San Luis Potosí: Cerro de San Pedro, 7-04-2007, L. Argüello, 1 ♀, CZUAA SCO-449.
Remarks
This species is recorded from Aguascalientes, Durango, Estado de México, Guanajuato, Hidalgo, Michoacán, Querétaro, San Luis Potosí, and Zacatecas (Ponce-Saavedra et al., 2009; Santibáñez-López & Francke, 2013).
Formerly a subspecies of Diplocentrus keyserlingii Karsch, 1880, it was described by Hoffmann from specimens collected in the Municipality of Tepezalá, Aguascalientes. We confirmed its presence in the type locality Tepezalá and additionally in the Municipalities of El Llano and Asientos. Diplocentrus zacatecanus inhabits the portion of the state that belongs to the Chihuahuan Desert biogeographic province (Morrone et al., 2017), which is nearly half of Aguascalientes’s surface.
The external morphology and the hemispermatophore of D. zacatecanus are either conserved or too variable to delimit species (Santibáñez-López & Francke, 2013). One possible explanation is the propensity of being substrate specialists and their strong adaptation to construct galleries where they spend most of their life exhibiting scarcely superficial activity. Diplocentrus zacatecanus is a substrate specialist requiring a particular type of soil, like other pelophilous species (Polis, 1990; Prendini, 2001). Soil-type discontinuity hinders its dispersal, promoting isolated populations and may promote allopatric speciation. Due to its wide distribution range in Mexico, D. zacatecanus may comprise a species complex that needs molecular studies to clarify and guide the correct species delimitation within such a broad distribution area.
Vaejovidae Thorell, 1876
Syntropinae Kraepelin, 1905
Chihuahuanus bilineatus (Pocock, 1989) (Fig. 7)
Taxonomic summary
Holotype: British Museum of Natural History, London, UK: San Diego Texas, USA. 1 ♀. Williams (1970) and Sissom (2000) indicated that this is a spurious locality and is not found in Texas.
Figure 7. Habitus of male (top) and female (bottom) of C. bilineatus, scale = 1 cm.
Records from literature: Hoffmann (1931): México, Aguascalientes, Municipality of Tepezalá: 20 exemplars (♂, ♀). González-Santillán and Prendini (2013): México, Tamaulipas, Municipality of Jaumave: El Salto, between Palmillas and Jaumave, 23°21’26.39” N, 99°30’56.45” W, 1,115 m, 23-08- 2006, O. F. Francke, W.D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, 1 ♂, 1 ♀, CAS(ARA-1883). González-Santillán and Prendini (2015): México, Guanajuato, Municipality of San Luis de la Paz: San Luis de La Paz, 7 km E, 21°18’42.84” N, 100°25’36.06” W, 2,117 m, 24-08-2006, O. F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, A. Ballesteros, 1 ♀, AMCC (LP 6594). San Luis Potosí, Municipality of Ciudad del Maíz: Puerto Santa Catarina, 22°18’13.38” N, 99°37’16.08” W, 1,214 m, 22-08-2006, O. F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, A. Ballesteros, 1 ♀, AMCC (LP 6507). Tamaulipas, Municipality of Jaumave: El Salto, between Palmillas and Jaumave, 23°21’26.39” N, 99°30’56.45” W, 1,115 m, 23-08-2006, O.F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, 1 ♂, 1 ♀, AMNH (ARA 1129). Zacatecas, Municipality of Fresnillo: Colonia Guanajuato, 23°04’44.94” N, 103°05’08.34” W, 2,250 m, 6-07-2005, O. F. Francke, J. Ponce, M. Córdova, A. Jaimes, G. Francke, V. Capovilla, 1 ♀, AMCC (LP 5306). Yahia & Sissom (1996): México, Aguascalientes, Municipality of Asientos: 3.2 km W Asientos, 2,225 m, 9-06-1956, B. Banta, 2 ♂, 1 ♀, 19 juv., AMNH. Muncipality of Tepezalá: Tepezalá, C. C. Hoffmann, 4 ♂, 4 ♀, AMNH. Coahuila, Municipality of Saltillo: 8.6 km W Buñuelos in Valle de Guerra, 15-07-1977, E. A. Liner, Chaney, 2 ♀, FSCA. Nuevo León, Municipality of La Asención: 7.24 km N La Ascension, 19-07-1975, E. A. Liner, 2 ♀, 9 juv., FSCA. 4.34 km N, 3.86 km SE La Ascension on La Caballada Road, 19-07-1975, E. A. Liner et al., 2 ♂, 3 ♀, FSCA. 12.39 km N La Ascension, 19-07-1975, E. A. Liner et al, 2 ♂, 1 ♀, FSCA. 11.1 km W El Carmen, 15-06-1976, E. A. Liner et al., 1 ♀, FSCA. 3 km S San Roberto, under cactus, 13-08-1972, N.V. Horner, 1 ♀, WDS. San Luis Potosí: 35.40 km S Huizache, 20-09-1979, J.C., J. E. Cokendolpher, 1 ♂, WDS. km 20 on highway 70, 1 juv, -03-1972, 2 ♀, 1 ♀, AMNH. Hwy 70, 112.65 km W Valles, 19-02-1970, J. A. L. Cooke, R. W. Mitchell, 1 ♂, 2 ♀, 1 juv., AMNH. Municipality of Ciudad de Maíz: near Ciudad del Maíz, 19-08-1947, C. & M. Goodnight, 1 ♀, AMNH; km 50 on Hwy 57, 18-03-1972, J. M. Rowland, TMM 1 ♂. Tamaulipas: Km 14 on Hwy 101, 22-02-1973, W. Graham, T. R. Mollhagen, C. McConnell, 3 ♂, 8 ♀, 2 juv., AMNH. Km 53 on Hwy 101, 23-02-1973, T. R. Mollhagen, 1 ♂ subadult 1 ♀, AMNH. Km 92 on Hwy 101, 22-02-1973, T. R. Mollhagen, 1 ♂, 8 ♀, AMNH. Km 15 on Hwy 19, 18-03-1972, J. A. L. Cooke, 1 ♀, AMNH. Municipality of Ciudad Victoria: Ciudad Victoria, -06-1977, F. D. White, 1 ♀, WDS. 1 km NW La Presita, 20-09-1979, J. C. & J. E. Cokendolpher, 2 ♀, WDS. Palmillas, T. Raines, 2 ♀, AMNH. Municipality of Jaumave: 6.43 km N Jaumave, 20-09-1979, J.C. & J. E. Cokendolpher, 1 ♀, WDS.
Examined material: Mexico, Aguascalientes, Municipality of Asientos: Cerro Altamira, 2015, J. E. Díaz-Plascencia 1 ♀, CZUAA SCO-470. Asientos, 21-06-2015, J. E. Díaz-Plascencia, J. L. Aguilar-Aguilar, 1 ♀, CZUAA SCO-325. Municipality of El Llano: Juan el Grande, 21°56’21.00” N, 101°55’05.99” W, 2,493 m, 18-10-2019, N. A. Pulido-Hornedo, X. A. Ruiz, 5 ♂, 20 ♀, CZUAA SCO-474; 21°56’21.00” N, 101°55’06.00” W, 2,148 m, 8-08-2020, F. Chávez-Samayoa, 1 ♂, CZUAA SCO-524; 21°56’20.00” N, 101° 55’08.90” W, 2,492 m, 20-05-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-540; 21°56’01.53” N, 101°55’42.29” W, 2,155 m, 25-09-2020, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 4 ♀, CZUAA SCO-577; 21°56’08.88” N, 101°55’30.72” W, 2,228 m, 22-01-2021, D. F. Simijaca-Salcedo, A. Palacios, F. Chávez-Samayoa, 2 ♀, CZUAA SCO-628; 21°56’21.88” N, 101°55’18.44” W, 2,384 m, -03-2021, F. Chávez-Samayoa, D. M. Reyes-Páramo, 2 ♂, 1 ♀, CZUAA SCO-648. Municipality of Tepezalá: Cerro del Capulín, 22°14’45.06” N, 102° 09’14.90” W, 2,189 m, 13-10-2019, F. Chávez-Samayoa, I. Villalobos-Juárez, 15 ♂, 16 ♀, CZUAA SCO-494; 22°14’19.46” N, 102° 09’58.03” W, 2,082 m, 31-05-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, I. Villalobos-Juárez, 1 ♂, CZUAA SCO-520; 22°14’01.00” N, 102°09’27.58” W, 2,129 m, 21-08-2020, F. Chávez-Samayoa, 4 ♀, CZUAA SCO-527; 22°13’52.88” N, 102°09’36.76” W, 2,125 m, 23-10-2020, F. Chávez-Samayoa, I. Villalobos-Juárez, I. E. Gallegos-Vieyra, D. F. Simijaca-Salcedo, 1 ♀, CZUAA SCO-575. Cerro de San Juan, ladera N, 8-10-1988, J. A. Rodríguez-Ávalos, 3 ♀, CZUAA SCO-305. Tepezalá, 22°13’54.26” N, 102°10’03.06” W, 2,116 m, 7-08-2015, M. Rodríguez, CNAN; Tepezalá, 15-06-2006, V. Revillo-Castellanos, 1 ♀, CZUAA SCO-301; 28-03-2006, Z. Y. González-Saucedo, 1 ♀, CZUAA SCO-302. Nuevo León, Municipality of Galeana: 2 km hacia mina “La Huiche”, 24°41’33.72” N, 100°03’19.91” W, 1,945 m, 28-09-2013, O. F. Francke, A. Valdez, D. Barrales, J. Cruz, A. Guzmán, CNAN-SC003911. Zacatecas, Municipality of Luis Moya: Griegos community, 15-11-1996, J. R. Treviño, 1 ♀, CZUAA SCO-303.
Remarks
Chihuahuanus bilineatus is distributed in Aguascalientes, Coahuila, Guanajuato, Nuevo León, San Luis Potosí, and Tamaulipas (Sissom, 2000; Yahia & Sissom, 1996).This species was reported for the first time in Aguascalientes by Hoffmann (1931) for the Municipality of Tepezalá. This species has an affinity for arid environments and could be classified as a lapidicolous species (González-Santillán & Prendini, 2013). It also inhabits mainly the Chihuahuan Desert province, and collection records in the state include the municipalities of Aguascalientes, Asientos, El Llano, Jesús Maria, and Tepezalá.
Chihuahuanus coahuilae (Williams, 1968) (Fig. 8)
Taxonomic summary
Holotype: California Academy of Sciences: Mexico: Coahuila: 0.5 km SW Cuatro Ciénegas de Carranza, 1 ♂ and 19 ♂♂, 25 ♀♀ paratypes (type No. 10170).
Figure 8. Habitus of male (top) and female (bottom) of C. coahuilae, scale = 1 cm.
Records from literature: González-Santillán and Prendini (2013): México, Coahuila, Municipality of Cuatro Ciénegas: Ojo de Agua, ejido El Oso, 27°00’03.35” N, 102°00’13.57” W, 1,039 m, 19-07-2006, O. F. Francke, W. D. Sissom, K. McWest, B. Hendrixson, S. Grant, A. Jaimes, M. Córdova, 2 ♂, 2 ♀, CNAN-T84). González-Santillán and Prendini (2015): Mexico, Coahuila, Municipality of Cuatro Ciénegas: Ojo de Agua, Ejido el Oso, 27°00’03.35” N, 102°00’13.57” W, 1,039 m, 19-07-2006, O. F. Francke, W. D. Sissom, K. McWest, B. Hendrixson, S. Grant, A. Jaimes, M. Córdova, 2 ♂, 2 ♀, AMNH (ARA 1120); 19-07-2006, O. F. Francke, W. D. Sissom, K. McWest, B. Hendrixson, S. Grant, A. Jaimes, M. Córdova, E. González, A. Ballesteros, 1 ♂, AMCC(LP6600). Durango, Municipality of Gómez Palacio: Dinamita, 25°44’05.34” N, 103°40’24.00” W, 1,328 m, 29-07-2005, O. F. Francke, H. Montaño, J. Ballesteros, C. Durán, 1 ♂, AMCC (LP 5353). USA, New Mexico, Luna County: Little Florida Mountains, 1.6 km SE on Bonita Road from turnoff of Gap Road, 32°09’29.4” N, 107°35’18.9” W, 1,761 m, L. Prendini, J. Huff, 1 juv., AMCC(LP7095). McWest et al. (2017): USA; Texas, Armstrong County: SE Claude, Palo Duro Canyon, Mulberry Creek arm, S side of Mulberry Creek, 12.8 km S junction US 287/FM 28889, 34°57’01.01” N, 101°13’32.99” W, 11-06-2004, 3 ♂, 1 juv., WTAMU. Tx 207, 23.9 km S US 87, 34°53’42.40” N, 101°22’06.71” W, 1-09-2000, 1 ♀, WTAMU. Briscoe County: Caprock Canyons State Park, 34°26’40.78” N, 101°04’34.54” W, 1 ♂ 3 ♀, WTAMU. Lake Mackenzie, 34°32’40.49” N, 101°26’25.55” W, 14-10-1996, 1 ♀, WTAMU; 16-09-1998, 4 ♀ 4 juv., WTAMU. Tx 256, 22.2 km SW of South Brice, 34°31’28.20” N, 101°00’08.64” W, 739 m, 24-07-2009, 4 ♂, WTAMU. 32.1 km SW of South Brice, 34°28’20.57” N, 101°05’11.04 W, 818 m, 24-06-2009, 1 ♀, WTAMU. Cottle County: Matador Wildlife Management Area, 34°07’09.01” N, 100°27’10.01” W, 1 ♂, WTAMU; Matador WMA, Headquarters Pasture, 34°07’00.16” N, 100°21’32.98” W, ?-05~09-2009, 4 ♂ 3 ♀, 1 juv., WTAMU. Crosby County: White River Lake, 33°27’33.48” N, 101°05’28.61” W (approximate), 23-09-1973, 1 ♀, AMNH; White River Lake, 12-04-1985, Taylor, S. W., SAS. Dickens-Crosby County line: FM 2794, 33°28’09.34” N, 101°02’19.79” W, 26-09-2009, 3 ♂ 3 ♀, 1 juv. ♀, WTAMU. Garza County: Post, 33°11’27.20” N, 101°22’41.56” W (approximate), 11-04-1983, 2 ♀ (one with juveniles), AMNH. 1.6 km S Justiceburg, 33°01’47.53” N, 101°11’41.82” W (approximate), 10-06-1984, 1 ♂, SAS. Justiceburg, 33°02’34.30” N, 101°12’10.44” W (approximate), 16-09-1975, 1 ♂, AMNH. FM 2458, 1.1 km E jct US 84, 33°02’58.67” N, 101°11’43.08” W, 26-09-2009, 6 ♂ 3 ♀, WTAMU; FM 2458, 1.4~1.7 km E jct US 84, 26-09-2009, 3 ♂ 5 ♀, WTAMU; FM2458, 2.2 km E jct US 84, 33°02’46.00” N, 101°10’58.01” W, 26-09-2009, 3 ♀, WTAMU; jct FM 2458 and FM 3519, 33°02’46.46” N, 101°09’37.51” W, 26-09-2009, 3 ♂ 4 ♀, WTAMU. Hall County: N Estelline, sand dunes along S side of Red River at US 287, 34°33’38.30” N, 100°26’33.83” W, 545 m, 19-09-1996, 2 ♂ 1 ♀, WTAMU. Turkey, 34°23’33.18” N, 100°53’51.54” W (approximate), 18-06-1970, 1 ♀, AMNH. TX 70/256, 34°40’17.15” N, 100°55’25.68” W, 650 m, 24-07-2009, 8 ♂ 2 ♀, WTAMU; TX 70/256 6.4 km SW South Brice, 34°38’44.88” N, 100°55’52.90” W, 658 m, 24-07-2009, 2 ♂, WTAMU. King County: Outskirts, E side of Guthrie, 33°37’01.31” N, 100°18’58.32” W, -10-2006, 6 ♂ 22 ♀ 1 juv., WTAMU. Lubbock County: Yellowhouse Canyon, 33°37’33.67” N, 101°53’05.64” W (approximate), 26-04-1973, 1 ♂, AMNH. Motley County: Starkey Ranch, 3.2 km W and 1.6 km S Flomot, 34°12’40.93” N, 101°01’27.62” W (approximate), 19-09-1996, 1 ♂ 3 ♀, WTAMU. 6.4 km NW Matador, 34°03’35.71” N, 100°52’00.88” W (approximate), 11-06-1979, 1 ♀, AMNH. Oldham County: Dirt road off US 385, on grade edge, SE of Canadian River, S Boys Ranch, 35°31’16.64” N, 102°15’26.03” W, 07-04-1996, 1 juv., WTAMU. FM 1061, 7.5 km E jct US 385, 35°28’39.00” N, 102°11’03.98” W, 14-08-1999, 1 ♂, WTAMU. Potter County: Crossbar ranch, E-facing slope above W Amarillo Creek, 35°25’14.99” N, 101°54’38.99” W, 1 subadult ♂, WTAMU. Randall County: Amarillo (south side), 35°09’30.46” N, 101°53’26.81” W (approximate), 07-07-1998, 1 ♀ 1 juv. ♀, WTAMU. Canyon, 34°58’49.04” N, 101°55’07.50” W (approximate), 04-02-1968, 1 juv., WTAMU. RR 1541, 35°01’57.58” N, 101°50’51.65 W, 1,053 m, 24-06-2009, 6 ♂ 1 ♀, WTAMU. Ceta Canyon, 27-09-1998, 1 ♀, WTAMU. FM 1541, N side of Palo Duro Creek bridge, 35°01’58.55” N, 101°50’51.29” W, 1,055 m, 14-10-2005, 1 ♂ 1 juv. ♂, WTAMU. About 11.2 km NE Canyon, FM 1541, S side of canyon at Palo Duro Creek, 35°01’53.29” N, 101°50’53.63” W, 01-08-1997, 1 ♂, WTAMU. Lake Tanglewood, 35°03’24.19” N, 101°46’11.57” W, 26-09-2006, 2 ♂ 7 ♀ 2 juv., WTAMU, 35°02’37.43” N, 101°45’31.57” W, 08-07-2002, 2 ♂, WTAMU; 35°03’10.01” N, 101°45’07.99” W, 11-07-2002, 1 ♂, WTAMU; 35°02’26.40” N, 101°45’56.04” W, 11-07-2002, 2 ♂, WTAMU; 35°03’11.23” N, 101°45’16.49” W, 08-07-2002, 2 ♂, WTAMU; 35°03’08.89” N, 101°45’35.10” W, 08-07-2002, 1 ♂, WTAMU. Palo Duro Canyon State Park, 34°56’01.39” N, 101°38’34.58” W, 05-06-1996, 1 ♂, WTAMU; 34°55’56.21” N, 101°38’05.89” W, 05-06-1996, 2 ♂ 3 ♀ 1 subadult ♀, WTAMU. Tangle Aire Estates, 35°04’05.12” N, 101°47’49.49” W, 22-10-1998, 1 ♀, WTAMU. Timbercreek Canyon, 19.3 km NE Canyon, 14-10-1998, 2 juv, WTAMU, 35°02’54.49” N, 101°49’41.41” W, 24-09-2006, 3 ♂ 22 ♀ 2 juv., WTAMU. Scurry County: 11.7 km W Ira, 32°35’54.67” N, 101°07’08.65” W (approximate), 21-04-1979, 1 ♀ 2 juv., WDS. Yoakum County: Plains, Co. Rd 230, about 0.8 km S jct US 82/US 380, 33°10’49.62” N, 102°50’09.28” W, 17-07-2009, 1 ♂ 1 ♀, WTAMU; Co. Rd 230, about 2.5 km S Plains, jct US 82/US 380, 33°10’04.08” N, 102°50’21.16” W, 17-07-2009, 1 ♀, WTAMU. Santibáñez-López et al. (2019): México, Zacatecas, Municipality of Concepción del Oro: Anahuac, 24°31’51.60” N, 101°19’09.12” W, -04-2015, H. Carmona. Stockwell (1986): México, Coahuila, Municipality of Cuatro Ciénegas: 0.5 km SW of Cuatro Ciénegas de Carranza, 28-07-1967, S. C. Williams, W. S. Brown, 1 ♂ paratype 1 ♀ paratype, AMNH. 1.6 km S of Cuatro Ciénegas de Carranza, 12-08-1968, M. A. Cazier, J. Bigelow, 1 ♂ 1 ♀, AMNH. Cuatro Ciénegas Basin, 22-04-1973, J. Landy, 1 ♀, AMNH. USA, Arizona, Cochise County: Peloncillo mountains., 4-08-1976, M. A. Cazier, J. Hubbard, 1 ♂, AMNH. New Mexico, Bernardillo County: 5 km W of Albuquerque, 9-08-1981, R. A. Bradley, 1 ♀, AMNH; 2-09-1981, R. A. Bradley, 2 ♀, AMNH. Dona Ana County: 32.1 km N of Las Cruces, 31-05-1970, R. Smith, 6 ♂, 7 ♀, AMNH; Las Cruces, 28-05-1970, R. Smith, 3 ♂, AMNH; 16.09 km. S of Las Cruces, 15-05-1970, R. Smith, 3 ♂, AMNH. Eddy County: Guadalupe Mts., 1 ♀, AMNH. Carlsbad Caverns National Park, 6-10-1976, V. Roth, 1 ♂, AMNH. 24.14 km E of Loving, Summer 1978, C. Rudolf, 3 ♂ 6 ♀, AMNH. Hidalgo County: 1.6 km N of Rodeo, 20-07-1976, J. Hubbard, M. A. Cazier, 21 juv., AMNH; 25-07-1976, J. Hubbard, M. A. Cazier, 2 ♂, 2 ♀, AMNH. Lincoln County: Valley of Fires State Park, 19-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 1 ♂, AMNH. Luna County: 17.70 km E of Deming, J. Bigelow, 1 ♀, AMNH. Valencia County: 32.83 km NW of Los Lunas, 17-06-1970, M. A. Cazier, et al., 1 ♀, AMNH. Texas, Brewster County: Alpine, 2-10-1955, H. L Stahnke, 2 ♂, CAS; 7-10-1964, J. Scudday, 1 ♀, CAS. 4 km S of Alpine, 19-08-1968, S. C. Williams, J. Bigelow, M. M. Bentzien, 1 ♀, 7 ♂, CAS. 35.4 km S of Alpine (Babcock Ranch), 25-04-1964, S. Sikes, 1 ♀, CAS; 26-04-1964, S. Sikes, 1 ♂, CAS. Marathon, 20-07-1956, H. L. Stahnke, 1 ♂, CAS. 41.8 km S of Alpine, 28-07-1978, O. F. Francke, J. V. Moody, 1 ♀, AMNH. 12.87 km NE of Marathon, 27-07-1978, O. F. Francke, J. V. Moody, 1 ♂, AMNH; Big Bend National Park, 4-05-1966, E. Lehnert, 1 ♀, CAS. Big Bend National Park, Grapevine Hills, 18-08-1968, S. C. Williams, 2 ♀, CAS. Big Bend National Park, N base of Grapevine Mt., 7-09-1969, M. A. Cazier, et al., 35 ♂ 25 ♀, AMNH. Big Bend National Park, Burrow Spring Road, 22-03-1986, S. A. Stockwell, J. M. Steele, 1 ♀, CAS. Big Bend National Park, K-Bar Ranch, 25-07-1972, R. C. Stephens, 1 ♀, UCLB. Big Bend National Park, Panther Jct., 23-07-1956, H. L. Stahnke, 1 ♂, CAS. Big Bend National Park, Panther Jct., 17-08-1968, S. C. Williams, M. M. Bentzien, 29 ♀ 39 ♂, CAS; Big Bend National Park, 19.31 km SE of Panther Jct., 17-08-1968, S. C. Williams, M. M. Bentzien, 1 ♀, CAS. Big Bend National Park, 21.24 km SE of Panther Jct., 17-08-1968, S. C. Williams, M. M. Bentzien, 2 ♀, CAS. Big Bend National Park, Basin Jct., 17-08-1968, M. A. Cazier, J. Bigelow, 2 ♀ 5 ♂, CAS. Big Bend National Park, Rio Grande Village, 21-06-1970, M. A. Cazier, L. Welch, O. F. Francke, 27 ♂, 24, ♀, AMNH; 16-06-1974, M. A. Cazier, O. F. Francke, 19 ♂, 9 ♀, AMNH; -07-1978, 1 ♀, AMNH; 29-07-1978, O. F. Francke, J. V. Moody, 1 ♂, 1 ♀, AMNH. Big Bend National Park, 27-01-1973, C. McConnell, 1 ♂, AMNH. Crocket County: 16.09 km N of Iraan, 15-09-1985 S. A. Stockwell, 7 ♀, 1 ♂, SAS; 17.7 km N of Iraan, 29-09-1985, S. A. Stockwell, 1 ♀, SAS. 16.1 km N of Iraan, 8-03-1986, S. A. Stockwell, 3 ♀, SAS. 24.1 km S of Rankin, 21-03-1978, O. F. Francke, J. V. Moody, Hall, 1 ♂, 1 ♀, AMNH. Crosby County: White River Lake, 23-09-1973, J. M. Rowland, 1 ♀, AMNH; 12-04-1985, S. W. Taber, 1 ♀, 1 ♂, SAS. Bar X Ranch, 17-09-1977, O. F. Francke, J. R. Reddell, F. W. Wagner, 1 ♀, AMNH. Culberson County: Guadalupe Mts. National Park, McKittrick Canyon, 14-05-1978, O. F. Francke, J. V. Moody, 2 ♂, AMNH. Guadalupe Mts., Pine Springs Canyon, 18-06-1974, M. A. Cazier, O. F. Francke, 4 ♂, 1 ♀, AMNH. Guadalupe Mts., 5.6 km S of Pine Springs, 3-09-1972, O. F. Francke, J. Davidson, 2 ♂, AMNH. Guadalupe Mts., 12.07 km S of Pine Springs, 3-09-1972, O. F. Francke, J. Davidson, 1 ♂, AMNH. Guadalupe Mts. foothills below El Capitan, 30-08-1969, W. S. Parker, 1 ♀, CAS. Kent, 23-07-1956, H. L. Stahnke, 1 ♀, CAS; 1.6 km N of Kent, 23-06-1970, M. A. Cazier, L. Welch, O. F. Francke, 4 ♂, 6 ♀, AMNH. 6.4 km NE of Kent, 14-03-1981, N. V. Horner, 1 ♀, 1 ♂, MWSU. 125.5 km NE of Van Horn, 6-07-1979, O. F. Francke, J. V. Moody, Merikel, 1 ♀, AMNH. 31.38 km NE of Van Horn, 2-06-1978, O. F. Francke, J. V. Moody, Hall, 1 ♂, 1 ♀, AMNH. Van Horn, 28-06-1964, M. H. Muma, 1 ♂, CAS. El Paso County: Anthony, 19-06-1970, M. A. Cazier, L. Welch, O. F. Francke, 1 ♂, 1 ♀, AMNH; Franklin Mts., 19-05-1978, O. F. Francke, J. V. Moody, 1 ♂, AMNH; Franklin Mts. McKelligan Canyon Park, 13-04-1969, W. S. Parker, 1 ♂, CAS, 22-06-1969, W. S. Parker, 4 ♂, CAS, 12-07-1969, W. S. Parker, 1 ♀ 1 ♂, CAS. Garza County: Post, 11-04-1983, F. Rose, 1 ♂ 1 ♀ with young, AMNH; Justiceburg, 16-09-1975, Webb, 1 ♂, AMNH; 1.6 km mi. S of Justiceburg, 10-06-1984, S. A. Stockwell, 1 ♂, SAS. Hall County: Turkey, 18-06-1970, D. Kiser, 1 ♀, AMNH. Hudspeth County: 57.9 km E of El Paso, 15-03-1986, S. A. Stockwell, J. M. Steele, 1 ♀, SAS. 9.6 km E of Salt Flat, 8-09-1972, O. F. Francke, J. Davidson, 1 ♀, AMNH. 9.6 km E of Hueco, 5-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 5 ♂, 4 ♀, AMNH. 22.5 km N of Sierra Blanca, 21-05-1978, O. F. Francke, J. V. Moody, 1 ♀, AMNH. 4.5 km S of Sierra Blanca, 21-05-1978, O. F. Francke, J. V. Moody, 1 ♂, 3 ♀, AMNH. Jeff Davis County: 16.1 km S of Kent, 18-04-1980, W. W. Dalquest, 1 ♀, MWSU. 20 km S of Toyahvale, 13-03-1977, J. C. Cokendolpher, 1 ♂, MWSU. 40.1 km S of Van Horn, 3-06-1978, O. F. Francke, J. V. Moody, Hall, 3 ♀, AMNH. 50.4 km S of Van Horn, 2-06-1978, O. F. Francke, J. V: Moody, Hall, 1 ♂, AMNH. 6.4 km W of Fort Davis, 6-06-1978, O. F. Francke, Hall, J. V. Moody, 2 ♀, AMNH. Fort Davis, 7-06-1978, O. F. Francke, J. V. Moody, Hall, 2 ♀, AMNH. Fort Davis National Park, 11-09-1963, “Staff”, 1 ♀, 1 ♂, CAS. Davis Mts. State Park, 20-06-1970, M. A. Cazier, L. Welch, O. F. Francke, 3 ♀, AMNH; 20-08-1974, J. V. Moody, 1 ♂, 1 ♀, AMNH; Davis Mts. State Park Limpia Canyon, 5-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 3 ♀, AMNH. Loving County: 12.9 km N of Mentone, 1-06-1978, O. F. Francke, J. V. Moody, Hall, 1 ♀, AMNH. Lubbock County: Yellowhouse Canyon, 26-04-1973, T. R. Mollhagen, 1 ♂, AMNH. Motley County: 6.4 km NW of Matador, 11-06-1979, O. F. Francke, J. V. Moody, W. D. Sissom, 1 ♀, AMNH. Pecos County: 16.1 km N of Ft. Stockton, Will Banks Ranch, 27-12-1966, R. Winokur, 2 ♀ paratypes, CAS. 6.4 km E of Sheffield, along Pecos River, 7-06-1974, M. A. Cazier, L. Draper, O. F. Francke, 7 ♂, 5 ♀, AMNH. Presidio County: 76.6 km N of Candelaria, 3-06-1978, O. F. Francke, J. V. Moody, Hall, 1 ♀, AMNH. Candelaria, 14-06-1977, R. Wilson, 3 ♀, 1 ♂, CAS; R. Wilson, 2 ♀, 5 ♂, one ♀ with 40 first instar young, CAS; 29-07-1977, R. Wilson, 4 ♀ 1 ♂, CAS; 14-08-1977, R. Wilson, 3 ♀, 2 ♂, CAS; 23-08-1977, R. Wilson, 3 ♀, CAS; 26-09-1977, R. Wilson, 2 ♂, CAS. 61.1 km NE of Presidio, 5-06-1978, O. F. Francke, J. V. Moody, Hall, 1 ♂, 2 ♀, AMNH. 49.8 km NE of Presidio, 5-06-1978, O. F. Francke, J. V. Moody, Hall, 2 ♀, AMNH. 5.3 km N of Presidio, 2-09-1972, O. F. Francke, J. Davidson, 2 ♂, AMNH. 53.1 km S of Marfa, 5-06-1978, O. F. Francke, J. V. Moody, Hall, 2 ♂, AMNH. 0.8 km N of Shafter, 2-09-1972, O. F. Francke, J. Davidson, 3 ♂ 4 ♀, AMNH. Reeves County: Pecos, 13-07-1973, J. J. Landye, 1 ♀, AMNH. Balmorhea State Park, 26-08-1971, K. M., M. A. Cazier, 4 ♂ 1 ♀, AMNH. Terrel County: 30.6 km S of Sheffield, 8-06-1974, O. F. Francke, 1 ♂, AMNH. 8 km W of Sanderson, 8-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 3 ♀, AMNH. 40.2 km N of Dryden, 28-07-1970, R. C. Stephens, 1 ♂, UCLB. 12.8 km N of Dryden, 21-10-1978, J. V. Moody, Meeks, 1 ♀, AMNH. Val Verde County: 1.6 km SE of Langtry, 7-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 6 ♂, 7 ♀, AMNH. Wrinkler County: 9.6 km N of Winkler, 1-06-1978, O. F. Francke, J. V. Moody, Hall, 2 ♀, AMNH. 20.9 km S of Kermit, 7-06-1979, O. F. Francke, 1 ♂, AMNH. Williams (1968): México, Coahuila, Cuatro Ciénegas Municipality: Laguna Tio Candido, 14 km S of Cuatro Ciénegas, 15-04-1965, W. L. Minckley, 2 ♀, CAS. Laguna Churince, 16 km S of Cuatro Ciénegas, 24-12-1965, W. L. Minckley, C. O. Minckley, 3 ♀, CAS. Town of Cuatro Ciénegas, 05-04-1967, L. E. Cullum, 3 ♀, CAS. 4 km E of Cuatro Ciénegas, 27-07-1967, S. C. Williams, W. S. Brown, 10 ♂ 17 ♀, CAS. Poza la Becerra, 14 km SW of Cuatro Ciénegas, 28-07-1967, S. C. Williams, W. S. Brown, 2 ♂ 1 ♀, CAS. Gypsum sand dunes, 13 km SW of Cuatro Ciénegas, 27-07-1967, S. C. Williams, W. S. Brown, 2 ♂ 2 ♀, CAS. Travertine Ridge, 12 km SW of Cuatro Ciénegas, 15-08-1967, W. L. Minckley, 1 ♀, CAS. Above Rio Cañon, 4.3 km NW of Cuatro Ciénegas, 15-08-1967, W. L. Minckley, 1 ♂, 2 ♀. USA, Texas, Pecos County: Will Banks Ranch, 16 km N of Fort Stockton, 27-12-1966, R. Winkour, 1 ♂, 1 ♀, CAS.
Examined material: Mexico, Aguascalientes, Municipality of Aguascaientes: El Chicalote, 22°00’43.26” N, 102°15’11.84” W, 1,895 m, -10-2019, D. A. Perales-Olivares, 1 ♂, 1 ♀, CZUAA SCO-504. Universidad Autónoma de Aguascalientes, edificio 132, 28-06-2022, M. Martínez, 1 ♂, CZUAA SCO-673. Cerro de Los Gallos, -04-2015, C. A. Franco-Servín de la Mora, 1 ♂, CZUAA SCO-321. Area NO de la ciudad, 15-06-2006, M. A. López, 1 ♀, CZUAA SCO-322. Municipality of Asientos: Cerro Altamira., 2015, J. E, Díaz-Plascencia, 1 ♂, CZUAA SCO-404. Ojo de Agua de los Sauces, 25-04-2016, G. S. García-Macias, 1 ♂, CZUAA SCO-433. Municipality of Cosío: Estación de Adames, 16-08-2016, I. Chávez, 1♀, CZUAA SCO-412. Municipality of El Llano: Juan el Grande, 21°56’21.00” N, 101°55’06.00” W, 18-10-2019, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-553. Municipality of Jesús María: Los Arquitos, 21°55’21.68” N, 102°23’17.92” W, 1,931 m, 5-09- 2019, F. Chávez-Samayoa, I. Villalobos-Juárez, 1 ♀, CZUAA SCO-486; 21°55’11.50” N, 102°23’26.02” W, 2,014 m, 23-01-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo; 1 ♀, CZUAA SCO-617, 26-03-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♀, CZUAA SCO-653. Municipality of Tepezalá: Cerro del Capulín, 22°14’01.00” N, 102°09’27.58” W, 2,129 m, 21-08- 2020, F. Chávez-Samayoa, 1 ♂, 1 ♀ juv., CZUAA SCO-526, 23-10-2020, F. Chávez-Samayoa, I. Villalobos-Juárez, I. E. Gallegos-Vieyra, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-573; 22°13’57.46” N, 102°09’40.41” W, 2,119 m, 26-09-2020, F. Chávez-Samayoa, I. Villalobos-Juárez, 1 ♂, CZUAA SCO-582; 22°14’30.30” N, 102°09’13.93” W, 2,184 m, 31-05-2020, F. Chávez-Samayoa, I. Villalobos-Juárez, P. C. Hernández-Romero, D. F. Simijaca-Salcedo, 1 ♀, CZUAA SCO-584; 22°13’54.26” N, 102°10’03.06” W, 2,116 m, 7-08-2015, M. Rodríguez, CNAN. Coahuila, Municipality of Cuatro Ciénegas: Ejido el Oso, 27°03’35.64” N, 102°13’34.90” W, 1,039 m, O.F. Francke et al., 5 ♂, CNANSC2437.
Remarks
Chihuahuanus coahuilae (Williams, 1968)is distributed inAguascalientes, Chihuahua, Coahuila, and Durango (Sissom, 2000) and in the USA in Texas and New Mexico (Sissom & Hendrixson, 2005; Stockwell, 1986).
Although Escoto-Rocha & Delgado-Zaldívar (2008) reported Chihuahuanus coahuilae for the first time in Aguascalientes, subsequent national and regional scale studies (Francke, 2019; Ponce-Saavedra & Francke, 2013b; Santibáñez-López et al., 2015) fail to enlist this species as a part of Aguascalientes scorpion richness. Herein, we confirm the presence of C. coahuilae in the municipalities of Aguascalientes, Asientos, El Llano, Jesús María, and Tepezalá. Chihuahuanus coahuilae is a lapidicolous species that prefers arid environments within the Chihuahuan Desert province. We observed during fieldwork C. coahuilae in sympatry with D. zacatecanus, C. bilineatus, C. glabrimanus Sissom & Hendrixson, 2005, M. spadix, P. gracilior, and Vaejovis aguazarca Díaz-Plascencia & González-Santillán, 2022. Interestingly, no buthid scorpion was sympatric with C. coahuilae in Aguascalientes, suggesting a distinctive transition between scorpion faunas. The current distribution of the scorpion fauna established in Aguascalientes can be correlated to the geological history that created the interdigitation between the Sierra Madre Occidental, Pacific lowlands, and the Chihuahuan Desert biogeographic provinces.
Figure 9. Habitus of male (top) and female (bottom) of C. glabrimanus, scale = 1 cm.
Holotype: Florida State Collection of Arthropods, Gainesville, Florida, USA. México, Nuevo Léon: 7.8 km N La Ascension, 24°19’0.00” N, 99°54’00.00” W, 18-07-1972, E.A. Liner, R. M. Johnson, A. H. Chaney, 1 adult ♂. Paratype Mexico, Coahuila: 10.8 km S, 0.16 km E Arteaga at Los Pinos, 25°17’00.00” N, 100°50’00.00” W, 17-09-1975, leg. E.A. Liner et al.
Records from literature: González-Santillán and Prendini (2013): Mexico, Guanajuato, Municipality of San Diego de la Union: San Diego de la Unión, 8 km S, 21°23’08.67” N, 100°53’27.14” W, 2,078 m, 28-07-2006, E. González, M. Córdova, A. Jaimes, A. Ballesteros, O. F. Francke, 1 ♂, AMNH. San Luis Potosí, Municipality of Villa de Arista: Villa de Arista, 10 km E, 22°30’45.65” N, 100°45’29.99” W, 30-08-2006, O. F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, A. Ballesteros, 4 ♂ 1 ♀, CAS (ARA1909). González-Santillán and Prendini (2015): México, Guanajuato, Municipality of San Diego de la Unión: San Diego de la Unión, 8 km S, 21°23’08.64” N, 100°53’27.12” W, 2078 m, 28-07-2006, O. F. Francke, M. Córdova, A. Jaimes, E. González, A. Ballesteros, 1 juv. AMCC (LP 6611). Nuevo León, Municipality of Galeana: Providencia, 3 km E, 25°08’39.06” N, 100°39’02.40” W, 1,980 m, 17-07-2006, O. F. Francke, E. González, M. Córdova, J. Ballesteros, A. Jaimes, 1 ♂, AMCC (LP6558). San Luis Potosí, Muncipality of Ciudad del Maíz: Puerto Santa Catarina, 22°18’13.38” N, 99°37’16.08” W, 1,214 m, 22-08-2006, O. F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, A. Ballesteros, 1 juv, AMCC (LP6610).
Examined material: Mexico, Aguascalientes, Municipality of Aguascalientes: Jardín Botánico Rey Netzahualcoyotl, 10-08-2004, V., 1 ♂, CZUAA SCO-172; 8-06-, G. García, 1 ♂, CZUAA SCO-178; 8-01-2004, J. Escoto, 1 ♀, CZUAA SCO-180; 27-06-2015, J. E. Díaz-Plascencia, 1 ♂, 1 ♀, CZUAA SCO-183. Universidad Autónoma de Aguascalientes, edificio 1 A, 13-10-2015, M. Croce-Hernández, 1 ♀, CZUAA SCO-373; edificio 132, 28-08-2019, I. Villalobos-Juárez, 1 ♂, CZUAA SCO-469; 23-06-2015, G. De la Riva-Hernández, 1 ♀, CZUAA SCO-181; 21-09-2017, M. Martínez, 1 ♂, 1 ♀, CZUAA SCO-438. Avenida Circunvalación lado E, 24-11-1985, O. Rosales-Carrillo, 1 ♀, CZUAA SCO-170. Fraccionamiento Asturias, 18-12-2002, J. A. Escoto-Moreno, 1 ♂, CZUAA SCO-175. Carretera Aguascalientes-San Luis Potosí km 2.5, 15-08-2004, J. A. Escoto-Moreno, 1 ♂, 1 ♀, CZUAA SCO-171. Los Pericos, 29-05-2000, V. Herrera, 1 ♀, CZUAA SCO-165; 28-05-2000, R. M. Morales-Contreras, 1 ♀, CZUAA SCO-169. Rancho El Cocuyo, 23-10-1993, L. Delgado-Saldívar, 1 ♀, CZUAA SCO-173. Centro de Salud Urbano, colonia Guadalupe Peralta, 15-06- 2023, I. Martínez, 2 ♂, CZUAA SCO-734. Municipality of Asientos: Francisco Villa, hogar particular, 22°05’19.00” N, 101°58’05.00” W, 15-10-2002, F. Aguinaga, CNAN-SC4043. Municipality of Calvillo: Cercano a balneario “La Cueva”, 21°49’57.00” N, 102°48’58.82” W, 15-10-2019, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-560. Presa Malpaso, 10-09-1985, C. Ortiz-de L., 1 ♀, CZUAA SCO-164; 1 ♀, 26-09-1993, J. M. González-Martínez CZUAA SCO-174. Municipality of Cosío: Soledad de Abajo, unidad médica, 25-10-2002, F. Aguinaga, CNAN-SC4044. Casa del centro, 1-01-2002, F. Aguinaga, CNAN-SC4045. Municipality of El Llano: Juan el Grande, 21°55’58.40” N, 101°55’42.49” W, 2,198 m, 8-08-2020, F. Chávez-Samayoa, 1 ♂, CZUAA SCO-523. Municipality of Jesús María: Los Arquitos, 21°55’21.68” N, 102°23’17.92” W, 4-09-2019, I. Villalobos-Juárez, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-559. Buenavista, 11-06-2000, R. M. Muñoz, 1 ♂, CZUAA SCO-179. Municipality of Pabellón de Arteaga: Río San Pedro, 23-06-2015, J. L. Aguilar-Aguilar, 1 ♂, CZUAA SCO-182. Municipality of Tepezalá: Cerro del Capulín, 22°14’30.30” N, 102°09’30.13” W, 31-05-2020, I. Villalobos-Juárez, P. C. Hernández-Romero, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 1 ♂, CZUAA SCO-515; 22°14’16.19” N, 102°09’15.88” W, 6-06-2020, F. Chávez-Samayoa, 1 ♂, CZUAA SCO-542; 22°14’07.25” N, 102°09’25.16” W, 26-09-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, I. Villalobos-Juárez, 1 ♀, CZUAA SCO-563. Tepezalá, 21°13’54.41” N, 102°10’03.06” W, 2,116 m, 7-08-2015, M. Rodríguez. Guanajuato, Municipality of San Luis de la Paz: 7 km E of San Luis de la Paz, 21°18’42.84” N, 100°25’36.06” W, 2,117 m, 24-08-2006, O. F. Francke, H. Montaño, C. Santibáñez, A. Ballesteros, 2 ♂, CNAN-SC02455. Nuevo León, Municipality of Galeana: Carretera San Roberto-Galeana, km 78, 24°40’57.6” N, 100°06’24.0” W, 1,944 m, O. F. Francke, W. D. Sissom, K. McWest, B. Hendrixson, S. Grant, E. González S., M. Córdova, A. Jaimes, A. Ballesteros, 1 ♀, CNAN-SC2136. Providencia, 3 km NE, 25°08’39.06” N, 100°39’02.40” W, 1,980 m, 17-07-2006, O. F. Francke, E. González, M. Córdova, J. Ballesteros, A. Jaimes, 1 ♂, AMCC (LP 6558). 3km NW of La Providencia, 25°08’39.12” N, 100°39’02.88” W, 1,980 m, 17-07-2006, O. F. Francke, E. González, A. Jaimes, M. Córdova, A. Ballesteros, 5 ♂, CNAN-SC3108. Querétaro: Estación de Microondas Santa Lucia hacia Peña Miller, matorral espinoso, 21°03’28.30” N, 99°47’01.90” W, 13-11-2009, O. F. Francke, J. Cruz-López, C. Santibáñez, A. Valdez, 1 ♀ juv., CNAN-SC2133. San Luis Potosí, Municipality of Ciudad del Maíz: Puerto Santa Catarina, 22°18’13.38” N, 99°37’16.08” W, 1,214 m, 22-08- 2006, O. F. Francke, W. D. Sissom, G. Casper, T. Anton, V. Torti, H. Montaño, C. Santibáñez, A. Ballesteros, 1 juv., AMCC (LP 6610). Municipality of Villa de Arista: 10 km E of Villa de Arista, 22°38’45.60” N, 100°45’29.94” W, 1,624 m, 20-08-2006, O. F. Francke, D. W. Sissom, T. Anthon, V. Totti, G. Casper, H. Montaño, C. Santibáñez, A. Ballesteros, 2 ♂, CNAN-SC03107. Zacatecas, Municipality of Fresnillo: 23-05-2009, S. R. López-Navarro, 1 ♀, CZUAA SCO-168; 21-06-2009, R. López-Navarro, 1 ♂, CZUAA SCO-176. Municipality of Jalpa: 15-04-2005, J. Silva, 1 ♀, CZUAA SCO-167. Municipality of Saín Alto: Emiliano Zapata, 9-09-1971, S. Guijosa, 1 ♀, CNAN SC02456.
Remarks
Chihuahuanus glabrimanus is distributed in Aguas-calientes, Coahuila, Guanajuato, Nuevo León, Querétaro, and San Luis Potosí (González-Santillán & Prendini, 2013, 2015).This speciesexhibits somewhat variable external morphology but is not correlated with its distribution, complicating its separation into morphospecies. Unlike diplocentrids, C. glabrimanus and C. coahuilae present some degree of substrate-specialism, having more and less elongated and alienated setae on the telotarsi, which is characteristic of the psammophilic ecomorphotype, an adaptation to digging in sandy soils (Polis, 1990; Prendini, 2001). Although we collected specimens in sandy areas, we failed to find a significant relationship between their presence and sandy habitats.
Figure 10. Habitus of male (top) and female (bottom) of M. spadix, scale = 1 cm.
Mesomexovis spadix (Hoffmann, 1931) (Fig. 10)
Taxonomic summary
Holotype: deposited in the American Museum on Natural History, New York, USA. México, Zacatecas, Municipality of Jalpa, 1 ♂, paratypes same locality deposited in UNAM 053, 1 ♂, 1 ♀ (Sissom, 2000).
Records from literature: González-Santillán & Prendini (2013): México, Guanajuato, Municipality of León: León, -04-2004, P. Berea, 1 ♂, 1 ♀, CNAN. Hoffmann (1931): México, Zacatecas, Jalpa. Santibáñez-López et al. (2015): México, Aguascalientes, Jalisco, Zacatecas. Sissom (2000): México, Aguascalientes, Jalisco, Zacatecas.
Examined material: Mexico, Aguascalientes, Municipality of Aguascalientes: Cerro de Los Gallos, 16-09-2014, J. E. Díaz-Plascencia, 2 ♀, CZUAA SCO-197; 27-11-1997, V. Villalobos-Sánchez, 1 ♀, CZUAA SCO-289; 26-11-1984, H. Gallegos-Rangel, 2 ♀, CZUAA SCO-295; 21-10-1989, M. E. Serafín-Luevano, 2 ♂, CZUAA SCO-297; -04-2015, C. A. Franco-Servín de la Mora, 2 ♀, CZUAA SCO-316. Cerro del Muerto, 23-05-2014, E. A. Hernández-Medrano, 1 juv. ♂, CZUAA SCO-248; 14-06-1979, J. Á. Uyoa, 1 ♀, CZUAA SCO-259; 10-04-2010, L. Consuelo-Márquez, 1 ♀, CZUAA SCO-268; 19-10-1988, J. A. Rodríguez-Avalos, 3 ♂, 4 ♀, CZUAA SCO-275; 21-10-1982, M. E. Serafín-Luevano, 2 ♂, CZUAA SCO-297. El Sauz, km 35, 19-04-1996, F. Arenas-Luevano, 1 ♂, CZUAA SCO-227. Municipality of Calvillo: Peña Blanca, 21°54’09.76” N, 102°45’15.48” W, 31-01-2022, I. J. Rodríguez-Elizalde, J. Brito-Ruiz de Velasco, 1 ♂, CZUAA SCO-724. Sierra Escondida, 21°43’32.12” N, 102°43’14.52” W, 10-02-2022, I. J. Rodríguez-Elizalde, J. Brito-Ruiz de Velasco, 1 ♂ juv., CZUAA SCO-725. 7 km NE of Temazcal, 22°02’36.46” N, 102°44’07.22” W, 2,246 m, 6-09-2021, I. J. Rodríguez-Elizalde, J. Brito-Ruiz de Velasco, 1 ♀, CZUAA SCO-682. Cercano a balneario “La Cueva”, 21°49’57.00” N, 102°48’58.82” W, 1,781 m, 28-10-2019, F. Chávez-Samayoa, S. S. Gutiérrez-Martínez, 1 ♀, CZUAA SCO-482; 15-10-2019, F. Chávez-Samayoa, 6 ♂, 22 ♀, CZUAA SCO-485. Los Adobes, 21°48’39.22” N, 102°41’21.34” W, 1,874 m, 11-10-2019, F. Chávez-Samayoa, L. R. Haro-Tapia, 2 ♀, CZUAA SCO-487. La Mezquitera, 21°51’39.85” N, 102°46’03.47” W, 5-10-2019, F. Chávez-Samayoa, M. E. Samayoa-Sepúlveda, L. F. Chávez-Rodríguez, 2 ♀, CZUAA SCO-511; 7-09-2019, F. Chávez-Samayoa, 7♂, 5♀, CZUAA SCO-513; 30-08-2019, J. A. Escoto-Moreno, F. Chávez-Samayoa, 4 ♀, CZUAA SCO-592. Presa La Codorniz, 21°59’50.78” N, 102°40’42.78” W, 2,068 m, 5-07-2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, C. Huerta, 10 ♂, 12 ♀, CZUAA SCO-613; 22°00’39.07” N, 102°40’19.51” W, 1-11-2020, E. González-Martínez, J. L. Reyes-Hernández, I. E. Gallegos-Vieyra, D. F. Simijaca-Salcedo, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-590. Presa Los Alamitos, 21°43’49.44” N, 102°42’50.04” W, 14-06-2021, M. E. Samayoa-Sepúlveda, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-635; 17-05-2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, 1 ♀, CZUAA SCO-649; 13-02-2021, D. G. Castillo-Arceo, N. D. Morales, 1 ♀, CZUAA SCO-663. Los Alisos, 4-03-2005, M. Orozco, 1 ♀, CZUAA SCO-311. El Ocote, 13-06-2009, M. Gómez, 1 ♀, CZUAA SCO-208; 11-06-2004, G. Guerrero, 1 ♀, CZUAA SCO-215; 12-06-2004, Benjamín, 1 ♀, CZUAA SCO-217; 13-06-2004, A. Del Río, 1 ♀, CZUAA SCO-255; 13-06-2004, S. Saucedo, 1 ♀, CZUAA SCO-267; -06-2002, E. N. Manzo-Mata, 2 ♀, CZUAA SCO-284; 13-06-2004, R. S. Pérez, 1 ♂, 1 ♀, CZUAA SCO-293. El Pilar, 2-05-2004, P. O. Hernández, 1 ♀, CZUAA SCO-214. La labor, caseta de vigilancia, 17-06-1992, J. A. Castorena Rangel, 1 ♀, CZUAA SCO-260. Las lomas, 3.5 km hacia Presa de los Serna, 14-10-1992, H. J. Cruz-González., 1 ♂, 1 ♀, CZUAA SCO-256. Las moras, 15-03-2007, S. Álvarez-Betancourt, 1 ♀, CZUAA SCO-442. Los Alisos, 12-09-1988, J. A. Rodríguez-Avalos, 2 ♀, CZUAA SCO-219; 22-01-2014, A. Farfán-Castañeda, 1 ♂, CZUAA SCO-224; 19-05-1982, V. Quintero, A. Salado, 2 ♂, CZUAA SCO-231; 9-10-1989, M. Y. Quezada, J. I. Soriano, 4 ♂, 4 ♀, CZUAA SCO-294; 3-10-2014, J. E. Díaz-Plascencia, 2 ♀, CZUAA SCO-417. Presa Malpaso, 13-06-2004, Hernández-Camacho, 1 ♀, CZUAA SCO-210; 1-06-2005, C. C. Herrada, 2 ♂, 3 ♀, CZUAA SCO-225; 13-06-2004, E. J. Ortiz-Martínez, 1 ♀, CZUAA SCO-244; 13-06-2004, L. Muro, 2 ♀, CZUAA SCO-250. Mesa Montoro, 4-05-1996, F. Arenas-Luevano, 1 ♀, CZUAA SCO-291. Río Gil, 11-06-2004, A.E.G.E., 2 ♀, CZUAA SCO-296. Sierra del Laurel, 18-04-2010, A. G. Pérez D., 1 ♀, CZUAA SCO-237; 29-08-2015, J. E. Díaz-Plascencia, 4 ♂, 3 ♀, CZUAA SCO-319; 11-02-2017, V. M. Arano-Sierra, 1 ♂, CZUAA SCO-415. Municipality of El Llano: Juan el Grande, 21°56’21.00” N, 101°55’06.00” W, 18-10-2019, F. Chávez-Samayoa, E. González-Martínez, 1 ♂, 1 ♀, CZUAA SCO-476; 21°56’22.45” N, 101°55’22.22” W, 23-08-2020, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-534; 21°56’21.08” N, 101°55’17.87” W, -10-2019, E. González-Martínez, 6 ♂, 5 ♀, CZUAA SCO-544; 21°56’20.00” N, 101°55’09.84” W, 2,488 m, 27-02-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-642. Municipality of Jesús María: Los Muñozes, 21°57’00.14” N, 102°34’00.66” W, 2,179 m, 12-10-2019, F. Chávez-Samayoa, M. J. Morán-Gutiérrez, V. M. Arano-Sierra, 3 ♂, 9 ♀, CZUAA SCO-478; 16-10-2019, F. Chávez-Samayoa, 2 ♂, 7 ♀, CZUAA SCO-479. Los Arquitos, 21°55’21.68” N, 102°23’17.92” W, 1,931 m, 4-09-2019, F. Chávez-Samayoa, 6 ♂, 22 ♀, CZUAA SCO-485; 4-09-2019, I. Villalobos-Juárez, 4 ♀, CZUAA SCO-512; 21°55’12.51” N, 102°23’47.53” W, 2,034 m, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 3 ♀, CZUAA SCO-639; 21°55’18.66” N, 102°23’34.79” W, 1,964 m, 26-03-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 2 ♀, CZUAA SCO-640. Pileta, 21°57’20.92” N, 102°30’02.92” W, 2,167 m, 6-09-2019, J. A. Escoto-Moreno, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-593. Campamento Yuca, 22°02’20.37” N, 102°24’43.12” W, 2,262 m, 7-10-2019, M. J. Perales-Olivares, 1 ♀, CZUAA SCO-465. Cerro El Picacho, 18-10-1986, P. Serna-Pérez, 1 ♀, CZUAA SCO-263; 13-11-1985, J. G. Vázquez A., 1 ♀, CZUAA SCO-267; -08-2020, C. Ramírez, 1 ♀, CZUAA SCO-587. La Posta, 5-06-2014, E. Águila-Lira, 1 ♂, CZUAA SCO-236. Rancho Seco, 21-04-2019, V. M. Arano-Sierra, 1 ♂, 1 ♀, CZUAA SCO-472. Municipality of Pabellón de Arteaga: 25-06-1999, C. A. Ramírez, 1 ♀, CZUAA SCO-228. Municipality of Rincón de Romos: Sierpe, Y. Calvillo Lezama, 1 ♂, 1 ♀, CZUAA SCO-298. Rincón de Romos, 17-04-2004, N. Arroyo-Chávez, 1 ♂, CZUAA SCO-243; 20-06-1999, P. Hernández, 1 ♀, CZUAA SCO-258; 20-11-1999, L. F. de Alba, 1 ♀, CZUAA SCO-271; 23-03-1999, B. Saucedo, 1 ♀, CZUAA SCO-274. Municipality of San Francisco de los Romo: 6-06-2016, A. Bernal, 1 ♀, CZUAA SCO-446. Municipality of San José de Gracia: Estación Biológica Agua Zarca, 22°05’32.3” N, 102°33’20.3” W, 2,149 m, 20-03-2019, E. C. Ramírez, 1 ♂, 4 ♀, CZUAA SCO-547; 12-03-2003, Y. Gámez-Roldán, 1 ♂, CZUAA SCO-198; -02-2003, A. Morales, 1 ♀, CZUAA SCO-201; 18-03-2003, C. Quiñones-Valles, 1 ♀, CZUAA SCO-203; 29-03-2003, C. A. Morales-Asencio, 1 ♀, CZUAA SCO-204; 30-03-2014, J. Montero, 1 ♀, CZUAA SCO-211; 5.-04-2014, J. E. Díaz-Plascencia, 1 ♂, 2 ♀, CZUAA SCO-213; 31-05-2014, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-238; 31-05-2014, R. González-Muñoz, 1 ♀, CZUAA SCO-241; -02-2003, A.P.R.A., 1 ♂, CZUAA SCO-288; 30-05-2014, A. Mejía-Arellano, 1 ♂, CZUAA SCO-314; 28-04-2004, L. Rodríguez, 1 ♀, CZUAA SCO-315; 13-05-2016, O. Herrera, 1 ♀, CZUAA SCO-406; 20-05-2016, L. R. Haro-Tapia, 1 ♂, CZUAA SCO-409; 18-05-2018, F. S. Álvarez-Solís, 1 ♀, CZUAA SCO-414; 27-05-2016, I. Rodríguez-Ortiz, 1 ♀, CZUAA SCO-428. Guanajuato, Municipality of León: Colonia La Luz, V. H. González, 1 ♂, CZUAA SCO-230; León, 25-06-2007, M. Ávila, 1 ♀, CZUAA SCO-410. Municipality of Guanajuato: 26-05-2007, D. García, 1 ♀, CZUAA SCO-426. Municipality of Salamanca: J. A. Berlín-Diosdado, 1 ♀, CZUAA SCO-226. Jalisco, Municipality of Lagos de Moreno: Cerro La Bola, 18-02-2012, R. E. Rea-Reyes, 1 ♀, CZUAA SCO-222. Lagos de Moreno, 24-04-2016, C. E. Valdivia-Muñoz, 1 ♀, CZUAA SCO-451. Municipality of Guadalajara: 6-06-2017, O. Gutiérrez-Muñoz, 1 ♂, 1 ♀, CZUAA SCO-423. Zacatecas, Municipality of Guadalupe: Cerro de la Virgen, 1-06-2003, C. A. Barrón, 1 ♀, CZUAA SCO-262. Municipality of Jerez de García: Sierra de Los Cardos, 14-10-2015, R. A. Rosales-García, 1 ♂, 1 ♀, CZUAA SCO-313. Cerro de la Campana, 17-03-2014, R. A. Rosales-García, 1 ♂, 1 ♀, CZUAA SCO-209. Jerez, 2-11-1986, R. Macías-Ramírez, 1 ♀, CZUAA SCO-234. Municipality of San Pedro Piedra Gorda: Chiquihuitillo, 22-09-1990, M. G. Flores Olivares, 1 ♂, CZUAA SCO-245.
Remarks
Mesomexovis spadix occurs in Aguascalientes, Guanajuato, Jalisco, and Zacatecas (González-Santillán & Prendini, 2013; Hoffmann, 1931; Santibáñez-López et al., 2015; Sissom, 2000).
Hoffmann (1931) synonymized Vaejovis punctatus spadix and Vaejovis punctatus variegatus Pocock, 1898 with Vaejovis punctatus punctatus, although he recognized some differences, he did not consider them enough to separate species. Later, González-Santillán and Prendini (2013) erected Mesomexovis and elevated them to species rank as M. spadix and Mesomexovis variegatus. Some authors have enlisted Mesomexovis punctatus as part of Aguascalientes species richness (Ponce-Saavedra & Francke, 2013b; Contreras-Félix et al., 2015), probably confounding the former nominotypical subspecies.
Mesomexovis spadix is a very abundant lapidicolous species covering most of the territory of Aguascalientes and tolerates disturbed environments. We collected specimens in 7 of the 11 municipalities of the state: Aguascalientes, Calvillo, El Llano, Jesús María, Pabellón de Arteaga, Rincón de Romos, and San José de Gracia. Although M. spadix occupies the 3 biogeographical provinces in Aguascalientes, the Chihuahuan desert appears to limit its distribution to the northern territories, which is congruent with the prediction of González-Santillán and Prendini (2013), who mentioned that Chihuahuanus ecologically substitutes Mesomexovis in more arid areas.
Figure 11. Habitus of male (top) and female (bottom) of T. intrepidus, scale = 1 cm.
Thorellius intrepidus (Thorell, 1876) (Fig. 11)
Taxonomic summary
Holotype: deposited in the Naturhistoriska Riksmuseets Stockholm, Sweden. The only information associated to the holotype is: México 1 ♀ (Sissom, 2000).
Records from literature: González-Santillán and Prendini (2013): México, Colima, Municipality of Armería: Mine la Salada, NW Ixtlahuacan, 19°01’40.85” N, 103°47’02.21” W, 275 m, mine staff, 1 ♂, 1 ♀, AMNH. Municipality of Tecomán: Tecomán, 18°54’30.00” N, 103°52’28.00” W, 1 ♂, 1 ♀, AMNH. González-Santillán & Prendini (2018): México, Aguascalientes, Municipality of Calvillo: Chiquihuitero, 21°52’27” N, 102°42’09” W, 1,680 m, 29-06-2007, A. Valle Colis , 1 ♂, INDRE; El Rodeo, 21°52’09” N, 102°44’50” W, 1,620 m, 25-06-2007, A. Valle Colis, 1 ♀, INDRE; La Labor, 21°57’43” N, 102°41’46” W, 1,740 m, 15-18-06-2007, I. Escalante-Piña, 1 ♂, 1 ♀, INDRE; Rancho Media Luna, 21°47’56” N, 102°48’05” W, 1,560 m, 9-11-2008, I. Franchini, 1 ♀, INDRE; Mesa Grande, 21°48’18” N, 102°43’18” W, 1,760 m, 24-06-2003, G. Vargas, 1 ♀, INDRE, 24-06-2007, M. Sema, 1 ♂, INDRE. Colima, Municipality of Armeria: Armeria, 18°56’03” N, 103°57’52” W, 40 m, 2-01-2003, H. Rodríguez, 1 ♂, 1 ♀, CNAN SC2628. Mina La Salada, NW Ixtlahuacán, at mine headquarters, 19°01’40.80” N, 103°47’02.16” W, 275 m, 2005, mine staff, 11 ♀, AMNH. Municipality of Camotlán: Punta de Agua, 19°09’00.00” N, 104°16’00.00” W, 2-09-2003, E. González, 1 ♀, CNAN SC2242. Municipality of Colima: Colima, 19°14’36.00” N, 103°43’28.98” W, 30-12-1927, J. C. Chamberlin, 1 ♂, AMNH. Colima, 16 km S, 19°14’37” N, 103°43’51” W, 30-07-1954, W. J. Gertsch, 1 ♂, AMNH. Los Ortices, 19°06’46.8” N, 103°44’22.6” W, 343 m, 9-04-2004, P. Berea, 1 ♂, AMCC (LP 6379). Municipality of Comala: Comala, 19°19’00.00” N, 103°45’00.00” W, 9-04-2004, P. Berea, 1 juv. ♀, AMCC (LP 6377). Municipality of Coquimatlán: near Coquimatlán, 19°18’00.00” N, 104°06’00.00” W, 2008, L. L. Valdez, 1 ♂, CNAN SC2921. Municipality of Manzanillo: Los Parajes, 19°14’07” N, 104°24’52” W, 230 m, 3-02-2008, 1 ♂, INDRE. Manzanillo, 19°03’08” N, 104°18’57” W, 5 m, 1 ♀, INDRE, 17-01-1947, 2 ♂, 4 ♀, 1 subad. ♂CNAN SC2634. Manzanillo, 10 km of El Colomo, 19°03’08” N, 104°18’57” W, 16-04-2008, P. Berea, 1 ♀, CNAN SC2619. Colonia Valle de Las Garzas, 19°03’08” N, 104°18’57” W, 29-09-2008, 5 m, 1 ♀, INDRE. Manzanillo, Sector 7, 19°03’08” N, 104°18’57” W, 5 m, 6-02-2004, 1 ♂, INDRE. Nuevo Petatero Ranch, 19°03’38.94” N, 104°16’36.12” W, 4-02-2008, 1 ♀, INDRE. Potrero Grande, 19°10’19” N, 104°32’27” W, 6 m, 27-11-2008, 1 ♀, INDRE. Veladero de Otates, 19°15’38” N, 104°20’14” W, 280 m, 4-02-2008, 1 ♀, INDRE. Municipality of Minatitlán: Junction to Saus, on way to Minatitlán, 19°25’22.74” N, 103°58’59.94” W, 1,006 m, 26-05-2006, O. F. Francke, A. Valdez, H. Montaño, A. Ballesteros, 1 ♂, AMCC (LP 6469). Municipality of Tecomán: Tecomán, 18°54’30” N, 103°52’28” W, 8-05-1961, E. López, 1 subad. ♀, INDRE, -03-1992, Orozco, R. G., 1 ♂, INDRE, 30 m, 9-03-1992, R. Lezama, 3 ♂, 6 ♀, CNAN SC2629, around village, 29-11-1995, E. López, 1 ♀ 1 subad. ♂, CNAN SC2611, 12-01-1996, M. González, 1 ♀, CNAN SC2605, 18-02-1996, H. Arredondo, 1 ♀, CNAN SC2641, 19-06-1996, H. Arredondo, 1 ♀, INDRE. Municipality of Villa de Álvarez: Villa de Álvarez, 19°16’00.00” N, 103°44’15.00” W, 530 m, 11-02-1995, E. Polanco, 1 ♀, CNAN SC2618. Guanajuato, Municipality of León: Hacienda de Arriba, NE of León, in house, 2008, L. Olguín, 2 ♀, CNAN SC2922; León, 21°07’00.00” N, 101°41’00.00” W, 2-06-2006, 1 ♂, CNAN SC2612. Jalisco, Municipality of Jocotepec: San Juan Cosola, 1.6 km E, 20°17’12.00” N, 103°20’13.02” W, 10-07-1959, C. M. Bogert, 1 ♀, AMNH. Muncipality of La Huerta: Ejido E. Zapata, 19°22’58” N, 104°57’55” W, 10 m, 9-07-1994, I. Ramírez, 1 ♀, CNAN SC2606. Estación Biológica Chamela, 19°29’52.50” N, 105°02’36.48” W, 5-04-1981, M. V. Julia, 1 ♀, CNAN-SC2244, 20 m, 19-05-1985, A. Pescador, 1 subad. ♀, CNAN-SC2607; 26-04-1993, E. Ramírez, 1 ♂, CNAN SC2614; 25-03-1998, S. H. Bullock, 1 ♀, CNAN-SC2920; 14-09-1999, A. M. Corona, R. Ayala, 1 ♀, CNAN-SC2621; 19-09-1999, A. M. Corona, 1 ♀, CNAN-SC2622; 1-09-2011, O. F. Francke, A. Valdez, H. Montaño, A. Ballesteros, C. Santibáñez, 3 ♂, 1 ♀, CNAN-SC2245; 19°29’52.50” N, 105°02’36.48” W, 97 m, 30-08-2007, O. Francke et al., 1 juv. ♀, AMCC (LP 7670); Estación Biológica Chamela, El Tejon Trail, 19°29’50.16” N, 105°02’29.46” W, 18-08-2005, J. L. Castelo, UV detection at night, 1 juv., AMCC (LP 5309). Michoacán, Municipality of Aquila: Aquila, 18°35’57” N, 103°30’15” W, 180 m, 21-02-1992, R. Mares, 1 ♀, CNAN-SC2636. Arroyo de la Cruz, 5 km NE of Maquili, 18°34’33.5” N, 103°35’47.8” W, 24-01-2004, 1 ♂, 2 ♀ 1 subad. ♂ 1 subad. ♀, CNAN-SC2992. El Faro de Bucerías, tropical dry forest, 18°21’00.00” N, 103°29’00.00” W, 13/14-01-2002, E. González, 1 ♀, AMCC (LP 2022); El Faro de Bucerías, near La Llorona 18°20’40.74” N, 103°30’32.04” W, 17-01-2006, G. Suárez, CNAN-SC2624. Pomaro, 18°20’28” N, 103°18’29” W, 380 m, 1 ♂, 18-02-1992, R. Álvarez, 1 ♀, CNAN-SC2608. Municipality of Coahuayana: Coahuayana, 18°45’00.00” N, 103°40’00.00” W, 24-06-1991, M. J. Nereida, 1 ♀, CNAN-SC2615. Municipality of Jiquilpan: Jiquilpan, 19°59’37” N, 102°43’02” W, 1,560 m, G. Ignacio, 1 ♂, 1 ♀, CNAN-SC2616. University of Michoacán, 19°41’27.96” N, 101°12’09.78” W, 1 ♂, CNAN-SC2613. Municipality of Uruapan: Uruapan, 19°25’10” N, 102°03’30” W, 1,620 m, J. Julia, 1 ♂, CNAN-SC2610. Nayarit, Municipality of Tepic: La Loma motel, 21°30’30.00” N, 104°53’34.98” W, C.M. Bogert, 1 ♀, AMNH. Santibáñez-López et al. (2019): México, Colima, Municipality of Coquimatlán: El Palapo, 19°11’54.60” N, 102°54’49.68” W, 3-04-2015, J. Valencia, J. López, E. González. Soleglad & Fet (2008): México, Nayarit, Municipality of Tepic: 1 ♀, MES, 1 ♀, GL.
Examined material: Mexico, Aguascalientes, Municipality of Calvillo: Calvillo, 9-06-2021, I. J. Rodriguez-Elizalde, 1 ♂, CZUAA SCO-609. Terrero de la Labor, 22°01’32.02” N, 102°39’58.00” W, 1,927 m, 22-10-2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, 1 ♀, CZUAA SCO-656. Presa de los Serna, 3-10-2003, S. García, 1 ♀, CZUAA SCO-188; 3-10-2003, S. García, 1 ♂, CZUAA SCO189. Calvillo, 8-11-2021, R. García, 1 ♂, CZUAA SCO-669. Jalisco, Municipality of Encarnación de Diaz: 5-06-2009, E. Duar, 1 ♀, CZUAA SCO-378. Municipality of San Juan de los Lagos: 14-04-2011, F. A. Rubalcaba-Castillo, 1 ♀, CZUAA SCO-194.
Remarks
Thorellius intrepidus inhabits the states of Aguascalientes, Colima, Guanajuato, Jalisco, Michoacán, and Nayarit, inhabiting tropical dry forests, deciduous and broadleaf forests between 5 and 1,760 m altitude (González-Santillán & Prendini, 2018).
In Aguascalientes, we recorded T. intrepidus at 1,927 m of altitude (CZUAA SCO-656), which may represent most likely its altitudinal distribution limit at circa 2,000 m. Thorellius intrepidus is restricted to the tropical deciduous forest of the Calvillo Municipality. Like C. infamatus, T. intrepidus inhabits the Pacific Lowlands province, which may be a second component that invaded or diversified within that area. However, all the species of Thorellius are distributed in tropical to subtropical habitats (González-Santillán & Prendini, 2018) connected to the Pacific Lowlands, which suggest that the linage may have originated in the Pacific Lowlands province.
Smeringurinae Soleglad & Fet, 2008
Figure 12. Habitus of male (top) and female (bottom) of P. gracilior, scale = 1 cm.
Paruroctonus gracilior Hoffmann, 1931 (Fig. 12)
Taxonomic summary
Lectotype: deposited in the American Museum of Natural History, New York, USA. Tepezalá, Aguascalientes, México, 1 ♂ and 2 paralectotypes ♂♂.
Records from literature: Brown et al. (2002): USA, Texas, Terrel County: Chandler Independence Creek Preserve, 30°26’30.00” N, 101°43’26.00” W. Bryson Jr. (2014): USA, Arizona, Cochise County. Fet et al. (2006): México, Coahuila, Municipality of Cuatro Ciénegas: 2 ♂, MES. USA, New Mexico, Hidalgo County: Lordsburg, 1 ♂, GL. Gertsch & Soleglad (1966): México, Aguascalientes, Municipality of Tepezalá: C. C. Hoffmann, 3 ♂ cotypes, AMNH. Coahuila, Municipality of Saltillo: 32.1 km E of Saltillo, 16-07-1965, W. J. Gertsch, V. Roth, 1 ♂. USA, Arizona, Cochise County: Portal, 1-07- to 4-09-1965, R. M. Hastings, W. J. Gertsch, V. Roth, 27 ♂, 4 ♀. New Mexico, Hidalgo County: Rodeo, 29-08-1964, R. Hastings, 1 ♂. Eddy County: Hope, 23-09-1950, W. J. Gertsch, 1 ♀. Hoffmann (1931): México, Aguascalientes, Municipality of Tepezalá: 4 ♀. Sissom & Francke (1981): USA, New Mexico, Chaves County: 30.8 km W Caprock, 21-03-1980, J. E. Cokendolpher, 1 ♀, JCC. Eddy County: Carlsbad Caverns National Monument, 8-09-1969, M. A. Cazier, J. Bigelow, 4 ♂, 5 ♀, AMNH. 24.1 km E Loving (creosote scrub and dunes), -07-1978, C. Rudoph, 9 ♂, AMNH. Hidalgo County: 20.9 km N Rodeo, 25-06-1973, O. F. Francke, 2 imm., AMNH. Hope, 23-09-1950, W. J. Gertsch, 1 ♀. Rodeo, 29-08-1964, R. Hastings, ♂. Luna County: Rock Hound State Park, 14.4 km E Deming, J. Bigelow, 1 ♂, 1 ♀, AMNH. Texas, Brewster County: Castolon, Big Bend National Park, 8-08-1979, O. F. Francke, J. V. Moody, F. Merickel, 1 ♂, AMNH; Grapevine Ranch, N base of Grapevine Mountain, Big Bend National Park, 7-09-1969, M. A. Cazier, J. Bigelow, 15 ♂, 3 ♀, 2 imm, AMNH. Culberson County: 1.6 km N Kent, 23-06-1970, M. A. Cazier, L. Welch, O. F. Francke, 4 ♀, 3 imm., AMNH. Jeff Davis County: Panthon Spring, 25-06-1968, J. C. Lewis, 1 ♀, AMNH. 5.3 km N Presidio, 2-09-1972, J. Davidson, O. F. Francke, 8 ♂, 11 ♀, AMNH. 4.6 km E Presidio, 2-09-1972, J. Davidson, O. F. Francke, 6 ♂, 3 ♀, 1 imm., AMNH. 57.9 km S Marfa, 2-05-1980, L. Robbins, 2 juv., AMNH. Val Verde County: 0.8 km S Langry, 14-06-1974, L. Draper, A. Cazier, O. F. Francke, 1 ♀, AMNH. Winkler County: 4.8 km W Wink, 7.-04-1979, W. D. Sissom, 2 ♂, WDS. Soleglad and Fet (2008): México, Coahuila, Municipality of Cuatro Ciénegas: 1 ♂ 1 ♀, MES. USA, New Mexico: 1 ♂, MES. Texas, Brewster County: Lajitas, 1 ♂, GL. Stockwell (1986): USA, Texas, Brewster County: 4 km S of Alpine, 19-08-1968, S. C. Williams, J. Bigelow, M. M. Bentzien, 1 ♂, CAS. Big Bend National Park Grapevine Ranch, N base of Grapevine Mt., 7-09-1969, J. Bigelow, M. A. Cazier, 19 ♂, 5 ♀, AMNH; Big Bend National Park, 19.3 km SE Panther Jct., 17-08-1968, S. C. Williams, M. A. Cazier, 3 ♂, 3 ♀, CAS. 21.2 km SE of Panther Jct., 17, 17-08-1968, S. C. Williams, M. M. Bentzien, 14 ♂, 10 ♀, CAS. Big Bend National Park, Castalon, 8-08-1979, O. F. Francke, J. V. Moody, 1 ♂, AMNH. Crockett County: 10 mi. N of Iraan, 8-03-1986, S. A. Stockwell, J. M. Steele, 2 ♂, 4 ♀, SAS. Culberson County: 19.3 km E of Van Horn, 3-07-1965, M. H. Muma, 1 ♀, CAS. 1.6 km N of Kent, 25-06-1970, M. A. Cazier, Welch, O. F. Francke, 3 ♂, 4 ♀, AMNH. Hudspeth County: 12.8 km W of Van Horn, 11-06-1982, W. D. Sissom, 1 ♀, AMNH. Jeff Davis County: Phantom Springs, 25-06-1968, J. C. Lewis, 1 ♀, AMNH. Pecos County: 16.1 km N, 3 mi. W of Fort Stockton, 6-08-1967, W. L. Minckley, 1 ♂, CAS. Presidio County: Candelaria, 26-09-1977. R. Wilson, et al., 2 ♂, 1 ♀, CAS. 57.9 km S of Marfa, 2-05-1980, L. Robbins, 2 ♀, MWSU. 11.26 km N of Presidio, 27-11-1975, J. C. Cokendolpher, 1 ♀, MWSU. 5.3 km N of Presidio, 2.ix.1972, J. Davidson, O. F. Francke, 10 ♂, 11 ♀, AMNH. Val Verde County: 0.8 km S of Langtry, 14-06-1974, L. Draper, M. A. Cazier, O. F. Francke, 1 ♀, AMNH.
Examined material: Mexico, Aguascalientes, Municipality of Tepezalá: Cerro del Capulín, 22°14’30.30” N, 102°09’30.13”, 2159 m, 31-05-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, P. C. Hernández-Romero, 1 ♂, CZUAA SCO-517; 22°14’16.19” N, 102° 09’15.88” W, 2,138 m, 29-06-2020, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-521; 22°14’06.83” N, 102°09’15.77” W, 2,130 m, 21-08-2020, F. Chávez-Samayoa, 2 ♂, 3 ♀, CZUAA SCO-539; 22°14’07.25” N, 102° 09’25.16” W, 2,117 m, 26-09-2020, F. Chávez-Samayoa, 1 ♀, CZUAA SCO-562; 22°13’52.88” N, 102°09’36.77” W, 2,127 m, 23-10-2020, I. Villalobos-Juárez, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♂, CZUAA SCO-576. San Luis Potosí, Municipality of Charcas: 23°16’00.36” N, 100°54’06.06” W, 2,029 m, -08-2010, O. F. Francke, D. W. Sissom, T. Anthon, G. Casper, V. Totti, H. Montaño, C. Santibáñez-López, A. Ballesteros, CNANSC2008. San Luis Potosí Municipality: 6-11-2009, D. Muro, 1 ♂, CZUAA SCO-394.
Remarks
Paruroctonus gracilior is distributed in Aguascalientes, Chihuahua, Coahuila, Durango, San Luis Potosí and the southern states of Arizona, New Mexico, and Texas in the USA (Francke, 2019).
Paruroctonus gracilior type locality is Tepezalá, the only municipality with specimens deposited in the CZUAA. Clearly, a psammophilous species component of the Chihuahuan desert adapted to sandy soils. We infer that populations could be established only in places with adequate substratum. In consequence, we suspect that, like D. zacatecanus, P. gracilior is a case of a complex of substrate-specialist species and is an excellent candidate to evaluate under a multispecies coalescence model (Degnan & Rosenberg, 2009) to test whether the species represents 1 or multiple lineages in such a wide distribution area, encompassing almost all Chihuahuan Desert.
Vaejovinae Thorell, 1876
The genus Vaejovis C. L. Koch in Aguascalientes is represented in the state by the mexicanus (Contreras-Félix & Francke, 2019) and nitidulus (Sissom & González-Santillán, 2004) species group.
Vaejovis nigrescens Pocock, 1898 (Fig. 13)
Taxonomic summary
Holotype: deposited in the British Museum of Natural History, England, UK. The only information associated to the specimen is a label indicating Mexico, 1 ♀ (Sissom, 2000).
Vaejovis nigrescens is distributed in Aguascalientes, Estado de México, Guanajuato, Jalisco, Michoacán, Querétaro, and Zacatecas (Sissom & González-Santillán, 2004).
Records from literature: Fet et al. (2006): México, Hidalgo, Muncipality of Pachuca: 2 ♀, MES. Sissom and Francke (1985): México, Aguascalientes, unspecified municipality: Río de Pirules, 1 ♀, AMNH. Ciudad de México: Mexico City, 4 ♂, 2 ♀, TOR Sc 516, ex. 617. Durango, Muncipality of Gómez Palacio: Dinamita, TOR Sc 517, ex. 656. Guanajuato, Muncipality of Guanajuato: Duges, 4 ♀, MNHN RS-0668; 2 ♀, MNHN RS-0678; 1 ♀, MNHN RS-0684; 1 ♂, 5 ♀, 7 first instars, MNHN RS-0671; in houses, Duges, 3 ♂ 4 ♀, TOR Sc 515; ex. 616, 2 ♀, BM; 1 ♀, BM; -06-1963, S. A. Minton, 1 ♀, SAM. Sissom and González-Santillán (2004): Mexico, Guanajuato, Municipality of Apaseo de Alto: 3 km S Huapango, 12-09-98, L. Ramírez, 1 ♂, UAQ LE00043. Municipality of Apaseo El Grande: El Tunal, 28-05-2000, A. R. Arias, 1 ♀, INDRE. Municipality of Atarjea: El Pilar (El Pilón) on wall of house, 12-05-2001, M. Suárez, 1 ♀, INDRE. Municipality of Celaya: 23-06-1979, M. Robledo, 2 ♂, 3 ♀, ENCB-IPN No. 49. Municipality of León: 27-05-1965, 1 ♂, 3 ♀, INDRE. Municipality of Irapuato: Rancho Cuchichuato, 25-04-2001, E. Martínez Zavala, 1 ♂, INDRE. Municipality of Samalanca: 20-05-2001, V. Jaime, 1 ♂, 1 ♀, ENCB-IPN. Municipality of Salvatierra: Emenguaro, -02-1969, 1 ♀, INDRE Sc 88199. Salvatierra, 31-10-1999, J. Callzontzin, 1 ♀ juvenile, UAQ. San José del Carmen, -01-1969, 2 ♂, 1 ♀, INDRE Sc 88195. San Pedro de los Naranjos, -02-1969, 3 ♂, 12 ♀, 2 juv. ♀, INDRE. Santiago Maravatio, -02-1969, 1 ♀, 2 juv. ♀, INDRE Sc 88198. Municipality of Victoria: Agua Fría, in house, 10-05-2001, M. Rivera-Díaz, 1 ♂, 1 ♀, INDRE. Jalisco, Municipality of La Huerta: Chamela, 04-04-1977, 1 ♀, CNAN-SC2497. Municipality of San Juan de los Lagos: 31-03-1979, H. Plascencia, 1 ♂, ENCB-IPN no. 189. Estado de México, Municipality of Ixtapan del Oro: 8-06-1941, CNAN-SC2499. Michoacán, Municipality of Apatzingan: 30-06-1989, L. Rios, 2 ♂, CNAN. Municipality of Hidalgo: Ciudad Hidalgo, 12-06-1987, I. Estrella, 1 ♂, CNAN-SC2505. Municipality of Huandacareo: 18-05-1989, D. A. Fernández, 1 ♂, CNAN; 15-05-1989, 2 ♀, CNAN. Municipality of Juárez: Parícuaro, 31-06-1988, B. Castro-Zarco, 1 ♀, CNAN -SC2506. Municipality of Junganpeo: Agua Blanca, in house, 1 ♂, CNAN-SC2508. Municipality of La Piedad: 25-06-1987, M. Méndez, 1 ♀, CNAN-SC2504. Municipality of Morelia: 15-06-1987, Á. Tinoco, 1 ♂ juv., CNAN-SC2502; 7-09-1986, J. Fabela, 1 ♂, CNAN; 30-06-1987, Ma. De J. Nereyda, 1 ♂, 1 ♀, CNAN-SC2496; 28-05-1987, R. Maya, 2 ♂, CNAN-SC2501; 7-06-1987, I. Tinoco, 1 ♂, CNAN-SC2494; 12-05-1986, 2 ♀, CNAN-SC2498; 11-07-1987, O. Barriga, 1 ♀, CNAN-SC2491; 8-08-1984, J. López, 1 ♂, CNAN-SC2516. El Reolito, 2 juv., J. Ponce, CNAN-SC2510. Municipality of Taretan: 12-07-1996, 1 ♂, CNAN-SC2492. Municipality of Uruapan: 3-05-1988, M. Moreno G, 1 ♂, CNAN- SC2511. Municipality of Zamora: 2-06-1991, A. R. Santos, 1 ♀, CNAN-SC2509. Municipality of Zitacuaro: 6-06-1985, 1 ♀, CNAN-SC2500. Querétaro, Municipality of Querétaro: 7-09-1998, R. Barron, Cid, Morales, 1 ♀, UAQ-LE0008. Zacatecas: Aguas Frías, -06-1963, 1 ♂, INDRE Sc 88203.
Examined material: Mexico, Aguascalientes, Municipality of Aguascalientes: Puertecito de la Virgen, 21°57’41.90” N, 102°16’04.37” W, 20-10-2020, C. Ramos, D. Rivero, 1 ♂, CZUAA SCO-569; -05-2015, L. Ruíz, 1 ♂, CZUAA SCO-354. Cañada Honda, 12-05-2010, N. Parga-Flores, 1 ♀, CZUAA SCO-353. Jardín Botánico Rey Netzahualcóyotl, 25-04-2014, R. Rivera-Martínez, 1 ♂, CZUAA SCO-346. Aguascalientes, 29-05-2012, S. Valencia-Chico, 1 ♀, CZUAA SCO-348; 12-05-2014, L. P. Cervantes-Zavalsa, 1 ♂, CZUAA SCO-351. Municipality of Asientos: Presa Charcos Verdes, 6-04-1994, A. Muñoz, 1 ♀, CZUAA SCO-361. Municipality of Calvillo: Presa Malpaso, 10-06-2004, F. Ramos-Martínez, 1 ♀, CZUAA SCO-345; 13-06-2023, E. Calvillo, 1 ♂, 2 ♀, CZUAA SCO-735. Municipality of Jesús María: Maravillas, 7-05-2004, L. F. López, 1 ♀, CZUAA SCO-350; Jesús María área urbana, A. Carmona, 1 ♂, CZUAA SCO-344. Municipality of Rincón de Romos: 17-04-2004, C. Arias-Camino, 1 ♀, CZUAA SCO-352. Guanajuato, Municipality of Guanajuato: 25-06-1999, L. L. O., 1 ♀, CZUAA SCO-336; 15-04-2014, C. D. Andrade-Moreno, 1 ♀, CZUAA SCO-341; 25-06-2007, M. C. Flores-Banderas, 1 ♂, CZUAA SCO-427. Municipality of Irapuato: 25-04-1999, M. Martínez-Garnica, 1 ♂, CZUAA SCO-333; 05-09-1987, Eliazarrás-Rivera, González-López, 1 ♂, CZUAA SCO-338. Municipality of Jaral del Progreso: 18-04-2009, C. García-Balderas, 1 ♀, CZUAA SCO-339; 18-04-2009, D. Valdés-Jiménez, 1 ♀, CZUAA SCO-342. Municipality of León: 21°00’32.73” N, 101°14’57.65” W, 3-06-2015, Ernestina, 1 ♀, CZUAA SCO-359. León, -06-2007, M. Ontiveros, 1 ♀, CZUAA SCO-439; 25-05-2007, M. C. Flores-Balderas, 1 ♂, CZUAA SCO-459. Municipality of Silao: -06-1986, M. Hernández A, 1 ♀, CZUAA: SCO-337. Jalisco, Municipality of Belén del Refugio: -08-1986, S. Ruvalcaba, 1 ♀, CZUAA SCO-355. Municipality of Encarnación de Díaz: 5-03-2013, C. Franco-Servín de la Mora, 1 ♀, CZUAA SCO-356; 25-05-2000, R. Guerrero-Alba, 1 ♀, CZUAA SCO-357; 15-04-2013, J. E. Díaz-Plascencia, 1 ♀, CZUAA SCO-363; 22-05-2005, K. G. Torres-Hernández, 1 ♀, CZUAA SCO-364; 7-06-2000, R. Guerrero-A, 1 ♂, CZUAA SCO-365. Zacatecas, Municipality of Valparaíso: 5-03-1999, J. G. Macías-Barragán, 1 ♀, CZUAA SCO-367.
Remarks
The municipalities recorded here include Aguas-calientes, Asientos, Calvillo, Jesús María, Rincón de Romos, and San Francisco de los Romo. Vaejovis nigrescens is a lithophilic species distributed in the transitional area between the Trans Mexican Volcanic Belt, the Chihuahuan desert, and the Sierras Madre Occidental provinces. Like M. spadix, V. nigrescens exhibits a wide distribution in Aguascalientes. Unlike the species of Mesomexovis, several species of the nitidulus species group of Vaejovis are distributed in the Chihuahuan desert, suggesting a different origin and species diversification compared to the genus Mesomexovis, maladapted to xeric environments (González-Santillán & Prendini, 2013).
Figure 14. Habitus of male (top) and female (bottom) of V. aguazarca, scale = 1cm.
Holotype: deposited in the Colección Nacional de Arácnidos, UNAM, México City. México, Aguascalientes, Municipality of San José de Gracia: Sierra Fría, Estación Biológica Agua Zarca, 22°05’26.00” N, 102°33’22.67” W, 2,173 m, 31-08-2015, J. E. Díaz-Placencia, J. L. Aguilar, ♂, CNAN-T01492. Paratypes 22°05’31.96” N, 102°33’27.86” W, 2,190 m, 7-09-2020. F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♂, 4 ♀, CNAN-T01493; 22°05’26.00” N, 102°33’22.67” W, 2,190 m, 9-11-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, P. García-Macías, 1 ♂, 1 ♀, CNAN-T01494; 22°05’34.58” N, 102°33’40.03” W, 2,181m, 19-02-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1♂, CNAN-T01495.
Examined material: Mexico, Aguascalientes, Municipality of Jesús María: Presa Abelardo Rodríguez, 21°55’7.64” N, 102°24’28.62” W, 1,927 m, 8-06-2003. J. E. Valencia, 1 ♀, CZUUA SCO-382. Los Arquitos, 21°55’17.94” N, 102°23’49.39” W, 1935 m, 08-2020, F. Chávez-Samayoa, S. S. Gutiérrez-Ramírez, D. F. Simijaca-Salcedo, 1 ♀, CZUAA SCO-583; 21°55’17.94” N, 102°23’49.39” W, 1935 m, 28-11-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 1 ♀, CZUAA SCO-606. Municipality of Rincón de Romos: Pabellón de Hidalgo, 22°11’18.48” N, 102°21’22.56” W, 2,046 m, 15-05-2013, H. Puga-Arévalo, 1 ♀, CZUUA SCO-386. Municipality of San Francisco de los Romo: El Chicalote, 22°00’43.26” N, 102°15’11.84” W, 1,896 m, -10-2019., M. J. Perales-Olivares, 1 ♀, CZUAA SCO-500. Muncipality of San José de Gracia: Estación Biológica Agua Zarca, 22°05’31.96” N, 102°33’27.86” W, 2,190 m, 7-09-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, 3 ♂, 6 ♀, CZUAA SCO-552; 22°05’33.18” N, 102°33’26.21” W, 2,190 m, 9-09-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, P. García-Macías, 5 ♂, CZUAA SCO-571; 22°05’34.58” N, 102°33’40.03” W, 2,181 m, 19-02-2021, F. Chávez-Samayoa, D. Fernando Simijaca-Salcedo, 1 ♂, CZUAA SCO-608; 22°05’31.92” N, 102°33’27.72” W, 1-04-2021, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, A. Jaime, 23 ♀, CZUAA SCO-702; 22°05’33.73” N, 102°33’53.21” W, 2,201 m, J. E. Díaz-Plascencia, C. Franco-Servín de la Mora, 1 ♂, 2 ♀, CNAN-S4005.
Remarks
Vaejovis aguazarca presents the most accentuated sexual dimorphism with body size at 23 ♂ and 32 ♀ mm among the mexicanus group species distributed in Aguascalientes. Vaejovis aquascalentensis Chávez-Samayoa & González-Santillán (2022), in contrast, is a medium size scorpion at 28 ♂ and 30 ♀ mm, and V. tenamaztlei Contreras-Félix et al. (2015), the smallest of them, is at 20 ♂ and 25 ♀ mm. The original description of Vaejovis aguazarca includes a detailed description of the natural history of the 3 species of this species group (Chávez-Samayoa et al., 2022). The authors indicated that the species uses a broad habitat, including oak bark nooks, under rocks, the base of yucca trees, and leaf litter, among others, as a refuge. The species also presents a peak of superficial activity during the rainy season, and it appears to be a well-separated altitudinal niche use when contrasted with V. aquascalentensis and V. tenamaztlei. Vaejovis aguazarca uses altitudinal levels between 1,927 and 2,201 m, whereas V. tenamaztlei uses between 2,390 and 2,864 m, a sharp difference of 100 m. However, V. aquascalentensis overlaps both altitudinal ranges from 1,601 to 2,367 m. Another difference is that V. tenamaztlei and V. aguazarca extend their distribution within the Sierra Madre Occidental and the Chihuahuan desert provinces (Chávez-Samayoa et al., 2022, their Fig. 22), whereas V. aquascalentensis seems to occupy the Pacific lowlands within Aguascalientes.
Figure 15. Habitus of male (top) and female (bottom) of V. aquascalentensis, scale = 1 cm.
Taxonomic summary
Holotype: deposited in the Colección Nacional de Arácnidos, UNAM, México City. Mexico, Aguascalientes, Municipality of Calvillo: Presa Los Alamitos, 21°43’49.85” N, 102°42’50.51” W, 2,367 m, 24-05-2021, F. Chávez-Samayoa, M. E. Samayoa-Sepúlveda ♂, CNAN-T 01496. Paratypes: 21°43’49.85” N, 102°42’50.51” W, 2,367 m, 17-05-2021, F. Chávez-Samayoa, D. Ortíz-Álvarez, 2 ♂, 1 ♀, CNAN-T01497. Presa el Adobe, inside a narrow canyon, 21°48’20.12” N, 102°40’55.99” W, 2,026 m, 3-08-2017, J. E. Díaz-Plascencia, E. González-Santillán, 3 ♂, CNAN-T01498.
Remarks
Vaejovis aquascalentensis was first recorded in the state as an undescribed species of Pseudouroctonus by Escoto-Rocha & Delgado-Saldívar (2008). However, Chávez-Samayoa et al. (2022) examined the specimens and concluded that the species belongs to the mexicanus species group. Likely, this species is also distributed in Jalisco and Zacatecas, although more fieldwork is required to confirm this supposition.
Figure 16. Habitus of male (top) and female (bottom) of V. tenamaztlei, scale = 1 cm.
Holotype: deposited in the Colección Nacional de Arácnidos, UNAM, México City, México. México, Aguascalientes, Municipality of Calvillo: 3 km to the east of Alamitos dam, Sierra del Laurel, 21º44’07.12” N, 102º41’51.11” W, 2,440 m, 24-07-2012, O. Francke, G. Contreras, D. Barrales, A. Valdez, 1 ♂, CNAN-T0871, paratypes same locality, 1 ♂, 3 ♀, CNAN T-0872 and Los Alisos, Sierra del Laurel, 21º43’32.20” N, 102º42’01.40” W, 2,415 m, 10-01-2011. J. C. Arenas. 1 ♂ deposited in the American Museum of Natural History.
Examined material: Mexico, Aguascalientes, Muni-cipality of Calvillo: Presa Los Alamitos, 21°43’49.44” N, 102°42’50.04” W, 2,368 m, 14-06-2021 F. Chávez- Samayoa, M. E. Samayoa-Sepúlveda, 2 ♂, 5 ♀, CZUAA SCO-616; 17.v.2021, F. Chávez-Samayoa, D. Ortiz-Álvarez, 5 ♂, 5 ♀, CZUAA SCO-632. Municipality of El Llano: Juan el Grande, 21°55’54.01” N, 101°55’06.18” W, 2,495 m, 18-10-2019, F. Chávez-Samayoa, E. González-Martínez, 1 ♀, CZUAA SCO-586; 21°56’22.88” N, 101°55’18.19” W, 2,390 m, 27-09-2020 F. Chávez-Samayoa, D. F. Simijaca Salcedo, I. E. Gallegos Vieyra, 2 ♂, 1 ♀, CZUAA SCO-566; 21°56’21.77” N, 101°55’18.30” W, 2,386 m, 25-09-2020, F. Chávez-Samayoa, D. F. Simijaca-Salcedo, P. C. Hernández-Romero, 7 ♂, 1 ♀, CZUAA SCO-567. Municipality of San José de Gracia: Monte Grande, 22°15’44.50” N, 102°37’43.03” W, 2,864 m, 19-09-2019, F. Chávez-Samayoa, J. J. Ayala-Rodríguez, R. Rosales, C. Franco-Servín de la Mora, E. Quiroz, 2 ♂, 3 ♀, CZUAA SCO-506; 22°15’44.50” N, 102°37’43.03” W, 2,864 m, 8-10-2019, F. Chávez-Samayoa, J. J. Ayala-Rodríguez, E. Quiroz, E. González-Martínez, 3 ♂, 2 ♀, CZUAA SCO-481.
Remarks
Vaejovis tenamaztlei was the first member of the mexicanus species group recorded in Aguascalientes and most likely is also distributed in Jalisco and Zacatecas. Like other species of the group, this species can be found in oak forests of the municipalities of Calvillo, El Llano and San José de Gracia (Chávez-Samayoa et al., 2022).
Key to identification of the scorpion genera and species of Aguascalientes Mexico.
– Cheliceral movable finger ventral medial denticle absent (Fig. 2B, C); pedipalp chela fixed and movable fingers denticles aligned as a straight line of retrolateral and prolateral denticles (Fig. 17B); pedipalp patella trichobothrial set v1-v3 present (Fig. 18B); pentagonal sternum (Diplocentridae/ Vaejovidae) 3
2. Carapace with each median ocelli surrounded by an incomplete areola of pigmentation; (Fig. 4A); pectinal basal piece with a central depression (Fig. 19A), pectinal tooth count 24-28 in males and 22-25 in females Centruroides balsasensis
– Carapace with each median ocelli surrounded by a complete areola of pigmentation; (Fig. 4B); pectinal basal piece without central depression (Fig. 19B); pectinal tooth count 23-24 in males and 20-21 in females Centruroides infamatus
6. Telson, dorsal surface with small fusiform, whitish glandular area anterior to base of aculeus (Fig. 27A) (minute to obsolete in Chihuahuanus bilineatus) [Chihuahuanus] 7
– Telson, dorsal surface without small fusiform, whitish glandular area anterior to base of aculeus (Fig. 27B) (Mesomexovis/Thorellius) 9
-Pedipalp patella dorsal retrolateral carinae granular or weakly granular (Fig. 28B); metasomal segments III-IV ventral carinae granular (Fig. 29C); telson caudal gland fusiform and evident 8
8. Carapace, pedipalp, terguites, and legs with evident dorsal patterns of infuscation (Fig. 30A); sternite VII with setae non arranged in a linear path (Fig. 31A) Chihuahuanus coahuilae
-Carapace and terguites with faint dorsal patterns of infuscation (Fig. 30B), pedipalps and legs immaculate; sternite VII with setae arranged in a linear path (Fig. 31B) Chihuahuanus glabrimanus
9. Chelicerae dorsal manus with 2 or more macrosetae (Fig. 2B); pedipalp chela carinae obsolete, fingers without median lobe, movable finger with a single denticle on seventh position; pectinal tooth count 20-21 in males, 17-18 in females; metasomal segments I-IV ventral carinae obsolete but with marked infuscation (Fig. 32) Mesomexovis spadix
– Chelicerae dorsal manus with 1 macrosetae (Fig. 2C); pedipalp chela carina granular, fingers with median lobe present, movable finger with a pair of denticles on seventh position; pectinal tooth count 23-25 in males, 20-22 in females; metasomal segments I-IV ventral carina costate without infuscation (Fig. 33) Thorellius intrepidus
10. Carapace anterior margin with 4 pairs of macrosetae; general coloration dark brown; pectinal tooth count 19-21 in males and 17-21 in females; metasomal segments I-IV ventral submedian carinae obsolete (Fig. 25B) Vaejovis nigrescens
– Carapace anterior margin with 3 pairs of macrosetae; general coloration light brown to reddish; pectinal tooth count 14-16 in males and 12-14 in females; metasomal segments I-IV ventral submedian carinae granular (Fig. 29C) 11
11. Pedipalp chela dorsal carinae immaculate; telotarsi ventral spinules grouped as a cluster (Fig. 34A); adult male telson dorsal surface with conspicuous hyaline glandular area (Fig. 27C) Vaejovis tenamaztlei
– Pedipalp chela dorsal carinae infuscated; telotarsi ventral spinules disposed as a straight row (Fig. 34B); adult male telson dorsal surface without conspicuous hyaline glandular area (Fig. 27B) 12
12. Chelicerae dorsal manus with 1 macrosetae (Fig. 2A, C); pedipalp femur retrolateral dorsosubmedian carina composed of 6 or 7 granules; chela movable finger with 7 prolateral and 6 retrolateral denticles Vaejovis aguazarca
– Chelicerae dorsal manus with 2 macrosetae (Fig. 2B); pedipalp femur retrolateral dorsosubmedian carinae costate in males or weakly granular in females; chela movable finger with 7 prolateral and retrolateral denticles Vaejovis aquascalentensis
Figure 17. Movable finger of C. infamatus (A) showing imbricated tooth rows, and M. spadix (B) showing aligned retrolateral and prolateral denticles.
Discussion
Despite being one of the smallest states of Mexico, the topography, climatic, and environmental conditions create a combination of factors that enhance biological diversity in Aguascalientes. Currently, Aguascalientes scorpion fauna comprises 13 species within the families Buthidae, Diplocentridae, and Vaejovidae (Table 1). Nonparametric estimations led us to conclude that the CZUAA currently holds a complete sample of the state’s diversity. Although these estimations must be taken, cautionary, as a lower bound of the actual species richness (Chao, 1984), this is especially true in taxa like scorpions with low vagility (Bryson Jr. et al., 2013), some of which are specialized in distinct microhabitats, making them less prone to be collected. In these terms, if we consider the upper limit that results from the sum of the standard error to the expected species number of each estimator reported in Table 2, the completeness of the CZUAA’s species inventory might be between 85.7-100% under the 10 × 10 km grids approach and between 92.3-100% under the 5-year classes approach. This last result can be explained by the fact that, under this approach, P. gracilior and V. tenamaztlei are doubletons, and thus, the calculation of Jackknife 2 resulted above 100%. Another factor to consider is that we are taking political boundaries as species limits, which make little sense in biological systems but result in a pragmatic approach. Our goal is to produce information that can be used for decision-makers.
Figure 18. Ventral view of pedipalp patella of C. infamatus (A), and Chihuahuanus glabrimanus (B).
Figure 19. Coxosternal area and pectinal basal piece of C. balsasensis (A), and C. infamatus (B).
Figure 20. Dorsal view of pedipalp patella of D. zacatecanus (A), V. aquascalentensis (B), and T. intrepidus (C).
Figure 21. Ventral view of leg III of D. zacatecanus (A), P. gracilior (B), and V. nigrescens (C).
Figure 22. Lateral view of telson of D. zacatecanus (A), and M. spadix (B).
Figure 23. Chela fingers of T. intrepidus (A), V. nigrescens (B), and P. gracilior (C). Trichobothria signaled as follows: 1 basal, 2 = internal terminal.
Figure 24. Carapace of D. zacatecanus (A), P. gracilior (B), and Vaejovis tenamaztlei (C).
Figure 25. Lateral (A), and ventral (B) view of the metasoma of V. nigrescens.
Figure 26. Lateral view of the metasoma of P. gracilior.
Figure 27. Dorsal view of telson of C. glabrimanus (A), P. gracilior (B), and V. tenamaztlei (C).
Figure 28. Dorsal view of pedipalp patella of C. bilineatus (A), and C. glabrimanus (B).
Figure 29. Ventral view of metasoma of C. bilineatus (A, B), and C. glabrimanus (C).
The key provided by Hoffmann (1931) in his original monography distinguishes M. spadix from M. punctatus Karsch based on the body base color and a higher pectinal tooth count, which led him to consider it as a subspecies. However, more recent contributions found further characteristics that differentiate these species. For instance, Fet et al. (2006) presented SEM micrographs of M. punctatus with 2-8 denticles conforming laterobasal aculear serrations (LAS) on the aculeus; however, the specimens of M. spadix examined under UV light during this study do not possess LAS. Finally, M. variegatus is devoid of ventral median carinae in metasomal segment V (Santibáñez-López & Sissom, 2010), which is present in M. spadix (Fig. 32).
Figure 30. Carapace of C. coahuilae (A), and C. glabrimanus (B).
Figure 31. Sternite VII of C. coahuilae (A), and C. glabrimanus (B).
Figure 32. Ventral view of the metasoma of M. spadix.
Figure 33. Ventral view of the metasoma of T. intrepidus.
This contribution is one of the few that present a taxonomic identification key to the species of a political state in Mexico. Currently, only the Mexican states of Baja California, Baja California Sur, Estado de México, and Morelos have a taxonomic key that allows determination to species level (Córdova-Athanasiadis, 2005; González-Santillán, 2004; Williams, 1980).
Natural history collections (NHC) are an invaluable source of information for evolutionary, ecological, taxonomical, and biogeographical research, as well as future applications and yet unexplored fields (Miller et al., 2020). Researchers focused on diverse taxonomical groups such as mammals (McLean et al., 2016), corals (Hoeksema et al., 2011), insects (Kharouba et al., 2019), parasites (Harmon et al., 2019), among others have also stressed the importance of NHC for the evaluation of impacts on biodiversity due to global change and human impact (Shaffer et al., 1998). At the same time, several authors have pointed out a decline in collecting activities and the deposition of specimens due to factors ranging from decline of taxonomical expertise to funding (Rohwer et al., 2022; Salvador & Cunha, 2020; Thompson et al., 2021), increasing regulations and ethical concerns. This contribution epitomizes the importance of local and regional NHCs because without the empirical evidence provided by the CZUAA and the CNAN, this contribution would not be possible, and valuable resources and information, such as a taxonomic key and the recognition of introduced species of public health importance, such as C. balsasensis, would have required comparatively more effort and time to discover.
Figure 34. Ventral view of the telotarsi of V. tenamaztlei (A), and V. aquascalentensis (B).
Acknowledgements
This contribution received funding from the doctoral scholarship 717372 granted to the first author by the Consejo Nacional de Humanidades, Ciencia y Tecnología (Conahcyt) and the project 321671 of Conahcyt and PIB22-2 of the Departamento de Biología de la Universidad Autónoma de Aguascalientes. We thank Fernando Álvarez Padilla of the Laboratorio de Aracnología of the Facultad de Ciencias, UNAM, and José Jesús Sigala Rodríguez of the CZUAA for their help and advice during the photography process of this contribution. We thank Daniel Alexander Carrillo Martínez, Mónica Croce and Patricia García Macías for their assistance during laboratory work. We thank all the collectors, especially Emmanuel González-Martínez, Luis Rodrigo Haro-Tapia, Pablo César Hernández-Romero, Diego Fernando Simijaca-Salcedo, and Iván Villalobos-Juárez, for their constant help. Collection permit FAUT-0213 from Semarnat to JAEM.
Appendix 1. Database of species incidence on a 10 × 10 km grid covering the state of Aguascalientes number of grids (n) = 61. Abbreviations as follows: C. inf = Centruroides infamatus, C. bil = Chihuahuanus bilineatus, C. coa = Chihuahuanus coahuilae, C. gla = Chihuahuanus glabrimanus, D. zac = Diplocentrus zacatecanus, M. spa = Mesomexovis spadix, P. gra = Paruroctonus gracilior, T. int = Thorellius intrepidus, V. agu = Vaejovis aguazarca, V. aqu = Vaejovis aquascalentensis, V. nig = Vaejovis nigrescens, V. ten = Vaejovis tenamaztlei.
C. inf
D. zac
C. bil
C. coa
C. gla
M. spa
P. gra
T. int
V. agu
V. aqu
V. nig
V. ten
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
0
0
1
1
0
0
0
1
1
0
1
0
1
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
1
0
0
0
0
0
0
0
1
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
1
0
1
1
1
1
0
0
1
0
1
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
1
0
0
1
0
0
0
0
0
0
1
1
0
0
0
1
0
1
0
1
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
Appendix 1. Continued
C. inf
D. zac
C. bil
C. coa
C. gla
M. spa
P. gra
T. int
V. agu
V. aqu
V. nig
V. ten
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
1
0
0
0
0
0
1
0
0
0
0
0
1
0
0
0
0
0
0
0
1
0
0
1
1
1
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
1
1
1
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
1
0
0
0
0
0
0
0
0
0
1
1
0
1
1
0
0
0
0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
Appendix 2. Database of species incidence in Aguascalientes in 5 years classes, number of classes (n) = 9. Abbreviations as follows: C. inf = Centruroides infamatus, C. bil = Chihuahuanus bilineatus, C. coa = Chihuahuanus coahuilae, C. gla = Chihuahuanus glabrimanus, D. zac = Diplocentrus zacatecanus, M. spa = Mesomexovis spadix, P. gra = Paruroctonus gracilior, T. int = Thorellius intrepidus, V. agu = Vaejovis aguazarca, V. aqu = Vaejovis aquascalentensis, V. nig = Vaejovis nigrescens, V. ten = Vaejovis tenamaztlei.
C. inf
D. zac
C. bil
C. coa
C. gla
M. spa
P. gra
T. int
V. agu
V. aqu
V. nig
V. ten
1979-83
1
1
0
0
1
1
0
0
0
1
0
0
1984-88
1
0
1
0
1
1
0
0
0
0
1
0
1989-93
1
1
0
1
1
1
0
0
0
0
0
0
1994-98
1
0
0
1
1
1
0
0
0
0
1
0
1999-03
1
0
1
0
1
1
0
1
1
0
0
0
2004-08
1
1
1
1
1
1
0
0
1
0
1
0
2009-13
1
1
0
0
1
1
0
0
1
1
1
0
2014-18
1
1
1
1
1
1
1
1
1
1
1
1
2019-22
1
1
1
1
1
1
1
1
1
1
1
1
References
Basualdo, C. V. (2011). Choosing the best non-parametric richness estimator for benthic macroinvertebrates databases. Revista de la Sociedad Entomológica Argentina, 70 , 27–38.
Beck, J., & Kitching, I. J. (2007). Estimating regional species richness of tropical insects from museum data: a comparison of a geography-based and sample-based methods. Journal of Applied Ecology, 44, 672–681. https://doi.org/10.1111/j. 1365-2664.2007.01291.x
Braga-Pereira, G. F., & Santos, A. J. (2021). Asexual reproduction in a sexual population of the Brazilian yellow scorpion (Tityus serrulatus, Buthidae) as evidence of facultative parthenogenesis. Journal of Arachnology, 49, 185–190. https://doi.org/10.1636/JoA-S-20-001
Brown, C. A., Davis, J. M., O’Conell, D. J., & Formanowiz Jr., D. A. (2002). Surface density and nocturnal activity in a west Texas assemblage of scorpions. The Southwestern Naturalist, 47, 409–419. https://doi.org/10.2307/3672498
Bryson, R. W. Jr. (2014) Bacterial endosymbiont infections in ‘living fossils’: a case study of North American vaejovid scorpions. Molecular Ecology Resources, 14, 789–793. https://doi.org/10.1111/1755-0998.12220
Bryson, R. W. Jr., Riddle, B. R., Graham, M. R., Smith, B. T., & Prendini, L. (2013). As Old as the Hills: Montane scorpions in southwestern North America reveal ancient associations between biotic diversification and landscape history. Plos One, 8, e52822. https://doi.org/10.1371/journal.pone.0052822
Chao, A. (1984). Nonparametric estimation of the number of classes in a population. Scandinavian Journal of Statistics, 11, 265–270.
Chávez-Samayoa, F., Díaz-Plascencia, J. E., & González-Santillán, E. (2022). Two new species of Vaejovis (Scorpiones: Vaejovidae) belonging to the mexicanus group from Aguascalientes, Mexico, with comments on the homology and function of the hemispermatophore. Zoologischer Anzeiger, 298, 148–169. https://doi.org/10.1016/j.jcz.2022.04.005
Contreras-Félix, G. A., Francke, O. F., & Bryson Jr., R. W. (2015). A new species of the “mexicanus” group of the genus Vaejovis C. L. Koch, 1836 from the Mexican state of Aguascalientes (Scorpiones: Vaejovidae). Zootaxa, 3936, 131–140. https://doi.org/10.11646/zootaxa.3936.1.8
Contreras-Félix, G. A., & Francke, O. F. (2019). Taxonomic revision of the “mexicanus” group of the genus Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae). Zootaxa, 4596, 1–100. https://doi.org/10.11646/zootaxa.4596.1.1
Córdova-Athanasiadis, M. (2005). Escorpiofauna (Arachnida: Scorpiones) del estado de Morelos, México(Bachelor Thesis). Facultad de Ciencias Biológicas, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos.
Degnan, J. H., & Rosenberg, N. A. (2009). Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends in Ecology & Evolution, 24, 332–340. https://doi.org/10.1016/j.tree.2009.01.009
De la Riva-Hernández, G. (2014). Colección Zoológica de la Universidad Autónoma de Aguascalientes. In J. J. Sigala-Rodríguez, M. D. Barba-Ávila, E. Meza-Rangel, G. Castorena-Esparza, & G. González-Adame (Eds.), La biología en la UAA: 40 años de desarrollo (pp. 77–82). Aguascalientes: Universidad Autónoma de Aguascalientes.
Escoto-Rocha, J., & Delgado-Saldívar, L. (2008). Insectos y arácnidos. InH. Ávila-Villegas, E. D. Melgarejo, & A. Cruz-Angón (Eds.), La biodiversidad en Aguascalientes: estudio de caso (pp. 126–128). Aguascalientes:Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Instituto del Medio Ambiente del Estado de Aguascalientes/ Universidad Autónoma de Aguascalientes.
Esposito, L. A., Yamaguti, H. Y., Pinto-Da-Rocha, R., & Prendini, L. (2018). Plucking with the plectrum: phylogeny of the New World buthid scorpion subfamily Centruroidinae Krauss, 1955 (Scorpiones: Buthidae) reveals evolution of three pecten-sternite stridulation organs. Arthropod Systematics & Phylogeny, 76, 87–122. https://doi.org/10.3897/asp.76.e31942
Fattorini, S. (2013). Faunistic knowledge and insect species loss in an urban area: the tenebrionid beetles of Rome. Journal of Insect Conservation, 17, 637–643. https://doi.org/10.1007/s10841-013-9569-5
Fet, V., & Lowe, G. (2000). Family Buthidae C. L. Koch, 1837. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the world (1758-1998) (pp. 54–286). New York: New York Entomological Society.
Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37, 4302–4315. https://doi.org/10.1002/joc.5086
Francke, O. F. (2019). Scorpions (Arachnida: Scorpiones) from the Cuatro Ciénegas Basin. In F. Alvarez, & M. Ojeda (Eds.), Animal diversity and biogeography of the Cuatro Ciénegas Basin: an endangered hyperdiverse oasis (pp. 53–59). Switzerland: Springer International Publishing. https://doi.org/10.1007/978-3-030-11262-2_4
Gertsch, W. J., & Soleglad, M. (1966). The Scorpions of the Vejovis boreus Group (Subgenus Paruroctonus) in North America (Scorpionida, Vejovidae). American Museum Noti- vitates, 2278, 1–54.
González-Santillán, E. (2004). Escorpiofauna del Estado de México(Master Thesis). Instituto de Biología, Universidad Nacional Autónoma de México. México D.F.
González-Santillán, E., & Possani, L. D. (2018). The North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Tropica, 187, 264–274. https://doi.org/10.1016/j.actatropica.2018.08.002
González-Santillán, E., & Prendini, L. (2013). Redefinition and Generic revision of the North American Vaejovid scorpion subfamily Syntropinae Kraeplin, 1905, with descriptions of six new genera. Bulletin of the American Museum of Natural History, 382, 1–71. https://doi.org/10.1206/830.1
González-Santillán, E., & Prendini, L. (2015). Phylogeny of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, based on morphology, mitochondrial and nuclear DNA. Cladistics, 31, 341–405. https://doi.org/ 10.1111/cla.12091
González-Santillán, E., & Prendini, L. (2018). Systematic revision of the North American syntropine vaejovid scorpion genera Balsateres, Kuarapu, and Thorellius, with descriptions of three new species. Bulletin of the American Museum of Natural History, 420, 1–81. https://doi.org/10.1206/0003-0090-420.1.1
Guralnick, R. P., & Van Cleve, J. (2005). Strengths and weaknesses of museum and national survey data sets for predicting regional species richness: Comparative and combined approaches. Diversity and Distributions, 11, 349–359. https://doi.org/10.1111/j.1366-9516.2005.00164.x
Harmon, A., Littlewood, D. T., & Wood, C. L. (2019). Parasites lost: using natural history collections to track disease change across deep time. Frontiers in Ecology and the Environment, 17, 157–166. https://doi.org/10.1002/fee.2017
Hoeksema, B. W., Van der Land, J., Van der Meij, S. E. T., Van Ofwegen, L. P., Reijnen, B. T., Van Soest, R. W. M. et al. (2011). Unforeseen importance of historical collections as baselines to determine biotic change of coral reefs: the Saba Bank case. Marine Ecology, 32, 135–141. https://doi.org/10.1111/j.1439-0485.2011.00434.x
Hoffmann, C. C. (1931). Monografías para la entomología médica de México. Monografía Núm. 2, Los escorpiones de México, Primera Parte: Diplocentridae, Chactidae, Vejovida. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, 2, 291–408.
Hoffmann, C. C. (1932). Monografías para la entomología médica de México Monografía NUM. 2 los escorpiones de México, Segunda Parte: Buthidae. Anales del Instututo de Biología, Universidad Nacional Autónoma de México, 3, 103–361.
Hoffmann, C. C. (1936). La distribución geográfica de los “alacranes peligrosos” en la República Mexicana. Boletín del Instituto de Higiene, 2, 321–330.
Kharouba, H. M., Lwethwaite, J. M. M., Guralnick, R., Kerr, J. T., & Vellend, M. (2019). Using insect natural history collections to study global change impacts: challenges and opportunities. Philosophical Transactions B., 374, 20170405. http://dx.doi.org/10.1098/rstb.2017.0405
McLean, B. S., Bell, K. C., Dunnnum, J. L., Abrahamson, B., Colella, J. P., Deardorff, E. R. et al. (2016). Natural history collections-based research: progress, promise, and best practices. Journal of Mammalogy, 97, 287–297. https://doi.org/10.1093/jmammal/gyv178
McWest, K. J., Valois, Z. J., & Sissom, W. D. (2017). Scorpions (Arachnida) of the high plains and adjacent canyonlands of northwestern Texas. Texas Journal of Science, 67, 3–38.
Meier, R., & Dikow, T. (2004). Significance of specimen databases from taxonomic revisions for estimating and mapping the global species diversity of invertebrates and repatriating reliable specimen data. Conservation Biology, 18, 478–488.
Miller, S. E., Barrow, L. N., Ehlman, S. M., Goodheart, J. A., Greiman, S. E., Lutz, H. L. et al. (2020). Building Natural History Collections for the Twenty-First Century and beyond. Bioscience, 70, 674–687. https://doi.org/10.1093/biosci/biaa069
Morrone, J. J., Escalante, T., & Rodríguez-Tapia, G. (2017). Mexican biogeographic provinces: Map and shapefiles. Zootaxa, 4277, 277–279. https://doi.org/10.11646/zootaxa.4277.2.8
Oksanen, J., Simpson, G. L., Guillaume-Blancjet, F., Kindt, R., Legendre, P., Minchin, P. R. et al. (2019). ‘Vegan’ Community Ecology Package. R package version 2.6-4. Accessed November 9th, 2022: https://github.com/vegan devs/vegan
Petersen, F. T., & Meier, R. (2003). Testing species-richness estimation methods on single-sample collection data using the Danish Diptera. Biodiversity and Conservation, 12, 667–686. https://doi.org/10.1023/A:1022495610021
Petersen, F. T., Meier, R., & Larsen, M. N. (2003). Testing species richness estimation methods using museum label data on the Danish Asilidae. Biodiversity and Conservation, 12, 687–701. https://doi.org/10.1023/A:1022464710930
Polis, G. A. (1990). The biology of scorpions. Standford, California: Standford University Press.
Ponce-Saavedra, J., & Francke, O. F. (2004). Una nueva especie de alacrán del género Centruroides Marx (1890) (Scorpiones, Buthidae) de la depresión del Balsas, México. Acta Zoológica Mexicana (nueva serie), 20, 221–232. https://doi.org/10.21829/azm.2004.2022340
Ponce-Saavedra, J., & Francke, O. F. (2013a). Clave para la identificación de especies de alacranes del género Centruroides Marx, 1890 (Scorpiones: Buthidae) en el Centro Occidente de México. Biológicas, 15, 52–62.
Ponce-Saavedra, J., & Francke, O. F. (2013b). Actualización taxonómica sobre alacranes del Centro Occidente de México. Dugesiana, 20, 73–79.
Ponce-Saavedra, J., & Francke, O. F. (2019). Una nueva especie del género Centruroides (Scorpiones: Buthidae) del noroeste de México. Revista Mexicana de Biodiversidad, 90, e902660. https://doi.org/10.22201/ib.20078706e.2019.90.2660
Ponce-Saaveedra, J., Francke, O. F., Cano-Camacho, H., & Hernández-Calderón, E. (2009). Evidencias morfológicas y moleculares que validan como especie a Centruroides tecomanus (Scorpiones, Buthidae). Revista Mexicana de Biodiversidad, 80, 71–84. http://dx.doi.org/10.22201/ib.20078706e.2009.001.585
Ponce-Saavedra, J., Francke, O. F., & Quijano-Ravell, A. F. (2009). Nuevos registros y distribución actualizada de Diplocentrus zacatecanus Hoffmann, 1931 (Scorpiones: Diplocentridae). Entomología Mexicana, 8, 57–60.
Ponce-Saavedra, J., Francke, O. F., Quijano-Ravell, A. F., & Cortés-Santillán, R. (2016). Alacranes (Arachnida: Scorpiones) de importancia para la salud pública en México. Folia Entomológica Mexicana (nueva serie), 2, 45–70.
Ponce-Saavedra, J., Linares-Guillén, J. W., & Quijano-Ravell, A. F. (2022). Una nueva especie de alacrán del género Centruroides Marx (Scorpiones: Buthidae) de la costa Noroeste de México. Acta Zoológica Mexicana (nueva serie), 38, 1–24. https://doi.org/10.21829/azm.2022.3812517
Prendini, L. (2001). Substratum specialization and speciation in southern African scorpions: the effect hypothesis revisited. In V. Fet, & P. A. Selden (Eds.), Scorpions 2001: In Memoriam Gary A. Polis (pp. 113–138). Plymouth, Devon: British Arachnological Society.
Quijano-Ravell, A. F., de Armas, L. F., Francke, O. F., & Ponce-Saavedra, J. (2019). A new species of the genus Centruroides Marx (Scorpiones, Buthidae) from western Michoacán state, México using molecular and morphological evidence. Zookeys, 859, 31–48. https://doi.org/10.3897/zookeys.859.33069
Quijano-Ravell, A. F., & Ponce-Saavedra, J. (2016). A new species of scorpion of the genus Centruroides (Scorpiones: Buthidae) from the state of Michoacán, México. Revista Mexicana de Biodiversidad, 87, 49–61. https://doi.org/10. 1016/j.rmb.2016.01.022
Reyes-Moya, A. G., Mejía-Zúñiga, D., Moncada-Hernández, L. J., Flores-Mendoza, F. J., Saldaña-Alcocer, J. A., González-Acosta, C. et al. (2021). First Report of Centruroides balsasensis in Northern Mexico. Southwestern Entomologist, 46, 275–278. https://doi.org/10.3958/059.046.0129
Rohwer, V. G., Rohwer, Y., & Dillman, C. B. (2022). Declining growth of natural history collections fails future generations. Plos Biology, 20, e3001613. https://doi.org/10.1371/journal.pbio.3001613
Salvador, R. B., & Cunha, C. M. (2020). Natural history collections and the future legacy of ecological research. Oecologia, 192, 641–646. https://doi.org/10.1007/s00442-020-04620-0
Sandoval-Ortega, M. H., Siqueiros-Delgado, M. E., Sosa-Ramírez, J., & Cerros-Tlatilpa, R. (2017). Amaranthaceae (Caryophyllales) richness and distribution in the state of Aguascalientes, Mexico. Botanical Sciences, 95, 203–220. https://doi.org/10.17129/botsci.909
Santibáñez-López, C. E., González-Santillán, E., Monod, L., & Sharma, P. P. (2019). Phylogenomics facilitates stable scorpion systematics: reassessing the relationships of Vaejovidae and a new higher-level classification of Scorpiones (Arachnida). Molecular Phylogenetics and Evolution, 135, 22–30. https://doi.org/10.1016/j.ympev.2019.02.021
Santibáñez-, López, C. E., & Francke, O. F. (2013). Redescription of Diplocentrus zacatecanus (Scorpiones: Diplocentridae) and limitations of the hemispermatophore as a diagnostic trait for genus Diplocentrus. The Journal of Arachnology, 41, 1–10. https://doi.org/10.1636/Ha12-65.1
Santibáñez-López, C. E., Francke, O. F., Ureta, C., & Possani, L. D. (2015). Scorpions from Mexico: from species diversity to venom complexity. Toxins, 8, 2. https://doi.org/10.3390/toxins8010002
Santibáñez-López, C. E., & Sissom, W. D. (2010). A new species of the Vaejovis eusthenura group in Oaxaca, Mexico (Scorpiones: Vaejovidae). Zootaxa, 2493, 49–58. https://doi.org/10.11646/zootaxa.2493.1.4
Shaffer, H. B., Fisher, R. N., & Davidson, C. (1998). The role of natural history collections in documenting species declines. Trends in Ecology & Evolution, 13, 27–30. https://doi.org/10.1016/S0169-5347(97)01177-4
Sigala-Rodríguez, J. J., & Greene, H. W. (2009). Landscape change and conservation priorities: Mexican herpetofaunal perspectives at local and regional scales. Revista Mexicana de Biodiversidad,80, 231–240. https://doi.org/10.22201/ib.20078706e.2009.001.599
Siqueiros-Delgado, M. E., Rodríguez-Avalos, J. A., Martínez-Ramírez, J., Sierra-Muñoz, J. C., & García-Regalado, G. (2017). Vegetación del estado de Aguascalientes. Aguascalientes: Universidad Autónoma de Aguascalientes.
Sissom, W. D. (1990). Systematics, biogeography and paleontology. In G. A. Polis (Ed.), The biology of scorpions (pp. 31–80). Stanford, CA: Standford University Press.
Sissom, W. D. (2000). Family Vaejovidae. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758-1998) (pp. 503–553). New York: The New York Entomological Society.
Sissom, W. D., & Francke, O. F. (1981). Scorpions of the genus Paruroctonus from New Mexico and Texas (Scorpiones, Vaejovidae). Journal of Arachnology, 9, 93–108.
Sissom, W. D., & Francke, O. F. (1985). Redescription of some poorly known species of the nitidulus group of the genus Vaejovis (Scorpiones, Vaejovidae). Journal of Arachnology, 13, 243–266.
Sissom, W. D., & González-Santillán, E. (2004). A new species and new records for the Vaejovis nitidulus group, with a key to the mexican species (Scorpiones, Vaejovidae). Texas Memorial Museum Speleological Monographs, 6, 1–8.
Sissom, W. D., & Hendrixson, B. E. (2005). A new species of Vaejovis (Scorpiones: Vaejovidae) from Coahuila and Nuevo León, and a key to the vaejovid species from northwestern and north-central Mexico. Zootaxa, 1088, 33–43. https://doi.org/10.11646/zootaxa.1088.1.5
Soberón-Mainero, J., Llorente Bousquets, J., & Oñate, L. (2000). The use of specimen-label databases for conservation purposes: An example using Mexican Papilionid and Pierid butterflies. Biodiversity and Conservation, 9, 1441–1466. https://doi.org/10.1023/A:1008987010383
Stahnke, H. L. (1971). Some observations of the genus Centruroides Marx (Buthidae, Scorpionida) and C. sculpturatus Ewing. Entomological News, 82, 281–307.
Stockwell, S. A. (1986). The scorpions of Texas (Arachnida, Scorpiones) (Master Thesis). Texas Tech University. Retrieved in: September 5th, 2023 from http://hdl.handle.net/2346/22278
Thompson, C. W., Phelps, K. L., Allard, M. W., Cook, J. A., Dunnum, J. L., Ferguson, A. W. et al. (2021). Preserve a voucher specimen! The critical need for integrating Natural History Collections in infectious disease studies. mBio, 12, e02698. https://doi.org/10.1128/mBio.02698-20
Williams, S. (1968). Scorpions from Northern Mexico: five new species of Vejovis from Coahuila, Mexico. Occasional Papers of the California Academy of Sciences, 68, 1–24.
Williams, S. (1970). A redescription of the scorpion Vejovis bilineatus Pocock (Scorpionida: Vejovidae). The Pan Pacific Entomologist, 46, 238–241.
Williams, S. (1980). Scorpions of Baja California, Mexico and adjacent islands. Occasional Papers of the California Academy of Sciences, 135, 1–127.
Yahia, N., & Sissom, W. D. (1996). Studies on the systematics and distribution of the scorpion Vaejovis bilineatus Pocock (Vaejovidae). The Journal of Arachnology, 24, 81–88.
Angélica Rodríguez-Cifuentes a, Jovana M. Jasso-Martínez a, Valeria B. Salinas-Ramos a, Juan José Martínez b, Alejandro Zaldívar-Riverón a, *
a Universidad Nacional Autónoma de México, Instituto de Biología, Colección Nacional de Insectos, Tercer Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
b Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de La Pampa, Uruguay 151, L6300LB, Santa Rosa, La Pampa, Argentina
Received: 17 November 2023; accepted: 15 March 2024
Abstract
Galls are an abnormal growth of plant tissue in response to the presence generally of an inducing insect, which ensures food and protection during specific periods of its life. Besides gall formers, a vast community of arthropods are associated with galls, including inquilines and parasitoids. Few studies have assessed the gall diversity and its associated insect community in Neotropical vascular plants. Here, we characterised the leaf gall diversity of Coccoloba barbadensis Jacq. (Polygonaceae) in a Mexican tropical dry forest, as well as their associated entomofauna based on morphology and DNA barcoding. Five different gall morphotypes were observed during both dry (April-June) and rainy (November) seasons. A total of 34 and 38 species of Diptera, Hymenoptera, Coleoptera, and Lepidoptera were delimited with the 2% divergence criterion and the GMYC model, respectively. Based on our rearing observations and literature, Cecidomyiidae (Diptera) species might induce all leaf gall morphotypes, whereas hymenopterans are represented by parasitoid and probably inquiline species of the families Braconidae, Eulophidae, Eupelmidae, Platygastridae and Torymidae. Our results highlight the importance of performing integrative species delineation studies of arthropods present in galls to have an accurate knowledge of their diversity and trophic interactions.
Keywords: Trophic interactions; Host; Parasitoid; DNA barcode; Gall former
Diversidad de agallas de hojas en la planta neotropical Coccoloba barbadensis (Polygonaceae) y riqueza de especies de sus insectos asociados
Resumen
Las agallas son un crecimiento anormal de tejido de plantas por la presencia de un insecto inductor que le asegura alimento y protección durante periodos específicos. Además de los formadores de agallas, una vasta comunidad de artrópodos está también asociada, incluidos inquilinos y parasitoides. Pocos estudios han evaluado la diversidad de agallas y su comunidad de insectos en plantas vasculares neotropicales. Aquí se caracteriza la diversidad de agallas foliares de Coccoloba barbadensis Jacq. (Polygonaceae) en un bosque seco tropical mexicano, así como su entomofauna asociada basada en morfología y el código de barras del DNA. Se observaron 5 morfotipos de agallas durante las temporadas seca (abril-junio) y lluviosa (noviembre). Se delimitó un total de 34 y 38 especies de Diptera, Hymenoptera, Coleoptera y Lepidoptera con el criterio de divergencia de 2% y el modelo GMYC, respectivamente. Según las observaciones y datos de literatura, especies de Cecidomyiidae (Diptera) inducen todos los morfotipos de agallas, y los himenópteros están representados por especies parasitoides y probablemente inquilinas de las familias Braconidae, Eulophidae, Eupelmidae, Platygastridae y Torymidae. Los resultados resaltan la importancia de estudios integradores para la delimitación de especies de artrópodos de agallas para tener conocimiento preciso de su diversidad e interacciones tróficas.
Palabras clave: Interacciones tróficas; Hospedero; Parasitoide; Código de barras del DNA; Formador de agallas
Introduction
Ecological interactions among species form the basis of ecosystem functioning and underlie evolutionary and ecological principles of conservation biology (Clare et al., 2013). Three types of biological networks have been defined based on interactions and the types of organisms involved (Ings et al., 2009): traditional or antagonistic food webs (e.g., predators and prey/food webs), mutualistic networks (e.g., seed dispersal and pollination), and host-parasitoid networks. The study of these biological networks provides a whole ecosystem outline to examine the effects of biodiversity loss on communities and ecosystem functions (Ings et al., 2009).
Understanding the links of a network relies on the idea that descriptors of interaction structure are unbiased and accurate (Wirta et al., 2014). In practice, however, networks are difficult to create, especially using traditional methods. Taxonomic resolution and the methodology employed to delimit species are crucial to reconstruct interaction structure (Paine, 1980; Kaartinen & Roslin, 2011). If the links are poorly resolved and multiple taxa are inadvertently grouped within the nodes of a web, there is a risk of misunderstanding its composition and thus its functioning system (Kaartinen & Roslin, 2011; Wirta et al., 2014).
An astonishing number of arthropod taxa depend on plants as food resources or closely interact with them. Among these are species that form enclosed structures known as galls (Mani, 1964; Raman, 2011). These structures are defined as abnormal growth of tissues of host plants in response to the activity or presence of an inducing organism (Nieves-Aldrey, 1998; Price, 2005; Redfern et al., 2002). Galls can be found in several plant structures such as flowers, roots, fruits, leaves, thorns, or stems. Arthropods induce galls to ensure food resources and to protect themselves against predators or unfavourable environmental conditions during certain periods of their life cycle (Nieves-Aldrey, 1998; Raman & Withers, 2003).
Gall induction in insects mainly occurs in species of Hymenoptera, but also in species of Diptera, Hemiptera and Thysanoptera, and less frequently in Coleoptera and Lepidoptera (Raman, 2011). Besides gall formers, there is an intricate community of different insect species that also are associated with galls, including inquilines (i.e., species that develop within galls made by other insects and feed on plant tissue), parasitoids of gall formers and inquilines and hyperparasitoids (i.e., parasitoids of other parasitoid species) (Forbes et al., 2015). Despite the great ecological importance of galls in most terrestrial ecosystems due to the extraordinary arthropod diversity that they comprise, to date, most of this species diversity and the interactions that are involved are largely unknown, especially in tropical and subtropical regions.
In recent years, molecular techniques have provided detailed analyses of interaction reconstruction, allowing precise identification of members of natural communities and the structure of networks (Clare et al., 2013; Kaartinen et al., 2010; Wirta et al., 2014). The DNA barcoding locus, a fragment of the cytochrome oxidase I (COI) mitochondrial DNA gene, is the most employed genetic marker for species discrimination of closely related animal species (Hebert et al., 2003; Ratnasingham & Hebert, 2013). This marker is a valuable tool for the rapid identification of megadiverse, poorly known taxa (Hebert et al., 2003). Moreover, it allows the association of morphologically distinct semaphoronts (e.g., insect larvae and adults; Yeo et al., 2018) and sexes of the same species (e.g., Sheffield et al., 2009).
Coccoloba barbadensis is a widely distributed Neotropical vascular plant species that in Mexico occurs in tropical regions from central to southeast Mexico (Howard, 1959). In this study, the diversity of galls on the leaves of the vascular plant C. barbadensis Jacq. (Polygonaceae) in a Mexican tropical dry forest was characterised and their associated entomofauna assessed using both morphological and DNA barcoding data. We highlight the necessity to perform integrative species delimitation studies of arthropods present in galls, particularly in the tropics, to have a more accurate knowledge of their species richness.
Materials and methods
The study was conducted in the Biological Station of Chamela (EBCH), Jalisco, Mexico (19’29” N, 105’01” W; Noguera et al., 2002), owned by the Instituto de Biología, Universidad Nacional Autónoma de México. The Chamela region is mainly composed of tropical dry forest and is characterised by having 85% of the ~ 750 mm of yearly rain from July to November and a mean annual temperature of 24.9 °C (14.8-32 °C) (Méndez-Alonzo et al., 2013). Tropical dry forests frequently show extreme changes in the physiognomy and available resources during the rainy and dry seasons, therefore altering the composition and diversity of their fauna (Razo-González et al., 2014).
We carried out 2 collecting trips at the EBCH, one during the dry (from March to June 2013) and the other during the rainy season (November 2013). Fifteen trees belonging to C. barbadensis were located and marked, all of which were situated near seasonal streams. We collected 1-5 leaves with galls from different parts of the selected trees. By in situ photographs we documented the presence/absence of galls for each tree, as well as general leaf features such as colour and size. The main gall’s features, including shape, size, colour, and pubescence, and the number of all collected galls present on the leaves were classified by morphotypes and recorded.
The levels of infestation, presence, and type of galls were weekly recorded, and the total number of galls per leaf was recorded. Galls were subsequently dissected or maintained in the laboratory to rear their insects. All reared insects were preserved in 96% ethanol and stored at -20 °C.
All collected insects were sorted out into adults, larvae, or pupae, and were counted and discriminated into morphospecies with a Zeiss™ Stemi DV4 (Göttingen, Germany) stereomicroscope. Larvae and adults of Hymenoptera were identified to order and genus level, respectively, using the specialised literature. Larvae and adults of Diptera and larvae of Coleoptera and Lepidoptera could only be identified to family and order level, respectively.
Gall abundance
The relationship between both the total number of collected galls and the average number of each gall morphotype concerning the time of collection during the dry season was evaluated using a simple linear regression with the statistical program R (R Core Team, 2013). Data of rainy season were not statistically analysed, since there were not enough samples to perform statistical tests. The insect family frequency throughout the sampled period was also analysed and then a canonical correspondence analysis to characterise the association between insect species and gall morphotypes with the program Statistica version 10 (StatSoft, Inc., 2011).
Molecular data
It is widely recognised that species misidentifications have negative consequences in ecological studies (Bortolus, 2008; Vink et al., 2012). Species delimitation and identification can be considerably improved using morphology-based taxonomy coupled with DNA sequence data (Dexter et al., 2010). Some representative specimens of most identified morphospecies were molecularly characterised generating for them DNA sequences of a fragment belonging to the barcoding locus COI mitochondrial gene (Hebert et al., 2003). This gene marker has been proved to be a generally reliable tool for the rapid delimitation of animal species, including insects (Hebert et al., 2003, 2004).
The genomic DNA from 1-6 specimens belonging to each discriminated insect morphotype was extracted. DNA extractions were conducted with the kit Tissue and tissue plus SV mini (Gene All®, Seoul, Korea), by placing each individual in 20 μl of proteinase K and 200 μl of TL Buffer, at 56 °C for 8 h. The larvae bodies were completely degraded after digestion, whereas the pupae exoskeletons and the adult individuals were subsequently washed with distilled water, placed back in 96% ethanol, and stored at -20 °C until they were mounted and labelled.
The COI fragment was amplified using the LCO1460/HCO2198 primers (Folmer et al., 1994). The following PCR conditions were used: initial denaturation at 95 ºC for 3 min, followed by 30 cycles of denaturation at 94 °C for 40 s, 40 s annealing at 45 °C, 40 s extension at 72 °C, and a final extension at 72 °C for 7 min. PCRs were prepared in a final volume of 15 μl of reaction mixture, which included 1.5 μl of 10X buffer, 0.75 μl of MgCl 2 (50 mM), 0.3 μl of dNTPs (10 mM), 0.24 μl of each primer (10 μM), 0.12 μl of Taq Platinum polymerase (Invitrogen®, Carlsbad, CA, USA), 8.85 μl of water and 3 μl of DNA template.
Unpurified PCR products were sent for DNA sequencing to the High-Throughput Genomics Unit of the University of Washington, Seattle, USA (http://www.htseq.org/). Sequences were edited and aligned manually based on their translated amino acids and compared individually with the sequences available in GenBank (http://www.ncbi.nlm.nih.gov/genbank) using the BLAST online program (Altschul et al., 1990; http://blast.ncbi.nlm.nih.gov/Blast.cgi).
DNA sequence-based species delimitation of dipterans and hymenopterans was carried out separately with the barcoding locus using 2 approaches, the General Mixed Yule Coalescence (GMYC) model (Pons et al., 2006) and the 2% genetic divergence criterion (Hebert et al., 2003).
The GMYC model requires an ultrametric tree, which was obtained with the program BEAST version 1.7.4 (Drummond et al., 2012), running the analysis for 10 million generations, sampling trees every 1,000 generations, using an uncorrelated lognormal clock and a coalescent tree prior. Only 1 partition was considered, which used the GTR+Γ+I evolutionary model. The duplicated haplotypes from the matrix were removed using the program Collapse 1.2 (Posada, 2004). The first 2,500 trees were eliminated as “burn-in” and the remaining trees were used to reconstruct a maximum clade credibility tree with the program TreeAnnotator version 1.7.4 (part of the BEAST 1.7.4 package). The GMYC model implemented in the SPLITS package (http://r-forge.r-project.org/projects/splits/) was performed with the R program version 2.10.1 (R core Team, 2021).
For the 2% divergence criterion, the allocation of molecular taxonomic units (Hebert et al., 2003) was made depending on the percentage of similarity in the genetic distances of the analysed sequences. If the percentage was less than 2, those MOTUs were considered to belong to the same barcoding species. Uncorrected COI divergences were obtained with the program PAUP* version 4.0 (Swofford, 2003). A Neighbor-Joining (NJ) distance tree for Diptera and Hymenoptera was reconstructed separately with the above program to visualise the genetic distances obtained. The trees obtained from 2 species delineation approaches were visualised with the program Figtree version 1.4.4 (Bouckaert et al., 2014).
Results
Description and abundance of galls
A total of 11,044 and 1,127 galls were dissected from 480 and 50 leaves obtained from the 15 trees that were sampled during the dry and rainy seasons, respectively. Five gall morphotypes from the leaves of C. barbadensis were identified (Fig. 1A-F): 1) capsule-shaped (Fig. 1B), green on both sides, glabrous, with a central inner elongated canal; 2) conical (Fig. 1C), pale green on both sides, ending on a sharp tip on the beam, glabrous, with an internal round chamber; 3) flattened (Fig. 1D), greenish-yellow on the beam and brown on the underside, glabrous, with a horizontal centrally elongated chamber; 4) spherical (Fig. 1E), glabrous, flat, indistinct and brown on the beam, brown and spherical on the underside, distributed irregularly along the leaf; and 5) rounded (Fig. 1F), brown on both sides, with abundant whitish pubescence. All gall morphotypes were recorded in both seasons except gall morphotype 5, which was not recorded during the rainy season.
Figure 1. Leaf gall morphotypes found on Coccoloba barbadensis Jacq. (Polygonaceae). A) Leaves of C. barbadensis; B) capsule-shaped gall; C) conical gall; D) flattened gall; E) spherical gall; F) rounded gall.
A significant relationship (R2 = 0.731, F = 21.796, p < 0.001) between the average number of total galls collected during the sampled weeks was observed, with the presence of galls in the leaves gradually increasing throughout the weeks during the dry season. The number of galls was relatively constant during the first 4 weeks (March 25-29 to April 22-23), but from the fifth week (April 29-30) it gradually increased (Fig. 2A).
Figure 2. A) Simple linear regression showing the average number of collected leaf galls on C. barbadensis during the time of collect in the dry season; B) graphic showing the average number of each leaf gall morphotype during the time of collect in the dry season.
Most galls collected during the dry season corresponded to gall morphotypes 1 and 4 (35% and 32%, respectively) and were present in 87% and 80% of the examined trees, respectively. Cecidomyiid exuviates adhered to the latter 2 gall morphotypes during the first 2 sampling weeks (25-26th of March – 1-2nd of April) and the last week of February, respectively. On the other hand, 13%, 18.5%, and 1.5% of the remaining galls belonged to gall morphotypes 2, 3, and 5 and occurred in 73%, 80%, and 13% of the trees, respectively. During the rainy season, 17.5%, 26.4%, 30.7%, and 25.4% of the collected galls corresponded to morphotype galls 1 to 4, respectively.
The average number of each gall morphotype varied over time during the dry season. There were significant differences between each type of gall (F3,128 = 8.616, p < 0.001) (Fig. 2B). The frequency of gall morphotype 1 gradually increased during the first weeks, though by the last week of April they were no longer registered. In contrast, gall morphotype 2 showed a lower abundance but its number was relatively constant throughout the sampling period, whereas gall morphotypes 3 and 4 had a low frequency but their number increased towards the first week of May. Gall morphotype 5 was only observed during the last 2 sampling weeks.
Integrative taxonomy
A total of 2,008 larvae and 356 adult insects were obtained from the dissected galls during both seasons. The morphospecies including larvae, pupae, and adults were first discriminated, for which were subsequently generated 230 COI sequences (151 sequences of Hymenoptera, 77 of Diptera, 1 of Coleoptera, and 1 of Lepidoptera (GenBank accession numbers in Appendix) resulting in 125 haplotypes. The number of species per family delimited by the 2 DNA sequence-based species delimitation approaches is provided in Table 1. The 2% COI divergence criterion and the GMYC model discriminated between 34 and 38 species, respectively. The NJ distance tree derived from the examined COI sequences of Hymenoptera and Diptera is shown in figures 3 and 4, respectively. There were 2 inconsistencies between both approaches. The 2% COI divergence criterion delimited 1 species of Chrysonotomyia (Entedoninae: Eulophidae) and 2 of Teniupetiolus (Eurytominae: Eurytomidae), whereas the GMYC model divided them into 3 and 2 species, respectively.
DNA sequence data supported most of the delimited species using larvae and adults, except for the only species of Torymus Dalman (Hymenoptera: Torymidae), the 3 species of Cecidomyiidae (Diptera), and the single species of Lepidoptera and Coleoptera, for which we only generated sequences of larvae.
Insect-gall association
Gall morphotype 3 had the highest insect species richness (20 species), which comprised all sampled families except the hymenopteran species of Braconidae (Allorhogas coccolobae Martínez & Zaldívar-Riverón) and Torymidae (Torymus sp.). This gall morphotype also had the highest number of eulophid species (8), from which 5 belong to the subfamily Tetrastichinae and was only present in this gall morphotype. Gall morphotype 1 had 18 associated insect species, with Platygastridae and Eurytomidae (Hymenoptera) being the families with more species (5 and 4 species, respectively; Table 2). Gall morphotypes 2 and 4, on the other hand, registered 9 and 12 species, respectively, whereas gall morphotype 5 only had 4 species, 1 belonging to Eulohpidae (Hymenoptera), 1 to Platygastridae (Hymenoptera) and 2 to Cecidomyiidae (Diptera) (Table 2).
Larvae of Cecidomyiidae were highly abundant throughout the dry season. The abundance of larvae and adults of the families Eurytomidae and Braconidae, on the other hand, considerably increased towards the sixth week of the dry season. In contrast, the presence of immature stages of Eulophidae was more frequent at the beginning of the dry season, though adults were also observed throughout this sampling period. Platygastridae was the hymenopteran family that was most frequently found in both larval and adult stages. Only immature individuals of Cecidomyiidae (Diptera) and Eulophidae (Hymenoptera) were collected during the rainy season.
A statistically significant difference (c2 = 912,989, df = 102, p <0.001) was observed between the 4 morphotypes of galls recorded during the dry season and their associated insect species. Fourteen out of the 37 insect species delimited with 2% barcoding were associated with a single gall morphotype. These included a species of Tenuipetiolus (Eurytomidae) in gall morphotype 1, 5 of Tetrastichinae (Eulophidae) in gall morphotypes 3, 1 species of Chrysonotomyia (Eulophidae) in gall morphotypes 1, 2 and 3, respectively, the single species of Eupelmidae and Torymidae in gall morphotypes 1 and 4, respectively, 2 and 1 species of Platygastridae in gall morphotypes 1 and 3, respectively, and 1 species of Cecidomyiidae in gall morphotype 3. Five delimited species of Cecidomyiidae were recorded in gall morphotypes 3 and 4, whereas 4, 3, and 2 were present in gall morphotypes 2, 1, and 5, respectively.
Discussion
A considerable leaf gall diversity in C. barbadensis is recorded here. Based on the gathered information, the inducers of these 5 types of leaf galls were species of Cecidomyiidae (Diptera) (see below). Other cecidomyiid galls that have been reported on species of Coccoloba include those found on stems of C. mosenii Lindl., on leaves of Coccoloba cf. warmingii Meisn., C. diversifolia Jacq., C. swartzii Kuntze, and C. uvifera Meins., and on inflorescences of C. alnifolia Casar (Mead, 1970; Maia et al., 2008; Ramos-Rodrigues et al., 2014). The number of different types of galls found in C. barbadensis (5 morphotypes) is higher than those reported for the above species (≤ 2 morphotypes). However, based on their general appearance, the gall morphotypes 1 and 2 may be variants made by the same inducer species. Further rearing observations and molecular characterisation of gall inducers will help to confirm the actual gall diversity that is present on the leaves of this plant species.
Figure 3. Neighbour Joining phenogram reconstructed for specimens of Cecidomyiidae that were reared from the 4 leaf gall morphotypes found in C. barbadensis. Bars refer to the 7 species of Cecidomyiidae that were delimited with the 2% barcoding approach.
There is considerable morphological diversity in the leaf gall morphotypes found on C. barbadensis (Fig. 1A-F). All these gall morphotypes were located on the leaf blade, with 4 of them being located on the beam and 1 on the underside. The considerable morphological diversity of these leaf galls can be attributed to several factors, among which include the high synthetic activity, rapid growth, structural and functional features, and characteristic morphogenetic patterns of leaf development (Mani, 1964). Further studies on these leaf galls are therefore necessary for a better understanding of the structural and histological differences among the leaf galls found on C. barbadensis.
Insect species richness
The high morphological diversity found on leaf galls favours the existence of a complex insect community (Mani, 1964); however, few studies have assessed in detail the insect community associated with leaf galls of Neotropical plant species (e.g., Maia, 2012). In this research, the extensive insect rearing, and examination of both molecular and morphological information helped to thoroughly characterise the insect species diversity found in the 5 types of galls present on leaves of C. barbadensis. Despite that, was not possible to confirm at this stage the biology of the reared taxa, and then their probable role is based on the relevant literature and the field observations.
Cecidomyiids, commonly known as gall midges, represent by far the group of insects with the most gall-inducer species (Gagné & Jaschhof, 2021). Species of Cecidomyiidae are mainly gall-formers, though members of the tribe Cecidomyiini are known to have a wide range of biologies, including only simple and complex gall formers, free-living, mycophagous, inquiline phytophagous, predator species of mites, aphids, and coccids, as well as internal parasitoids of aphids and psyllids (Kim et al., 2014; Uechi et al., 2011). The field observations and the insect species diversity found in the 5 leaf gall morphotypes of C. barbadensis strongly suggest that they are induced by cecidomyiid species. Among this evidence was that most of the dissected galls had a single cecidomyiid larva. Moreover, several galls belonging to morphotype 2 had a cecidomyiid pupal exuviae hanging outside of a small opening, which is a common feature of many galls with former cecidomyiids. Since the 5 leaf gall morphotypes were formed by cecidomyiid species, the remaining species of this family delimited in the study probably are inquilines.
Figure 4. Neighbour Joining phenogram reconstructed for specimens of Hymenoptera that were reared from the 4 leaf gall morphotypes found in C. barbadensis. Bars refer to the 25 species of Hymenoptera that were delimited with the 2% barcoding approach.
The cecidomyiid larvae and their emerging adults could not be identified at the genus level; however, a BLAST similarity search of the barcoding locus for the delimited cecidomyiid species suggests that they are closely related to species of the Cecidomyiini genera Contarinia Geer and Macrodiplosis Kieffer. The gall-inducing species of the genus Contarinia are cosmopolitan and can be either monophagous or polyphagous with a wide range of hosts (Uechi et al., 2011). Most species of this genus live gregariously in the floral parts of the plant or in the galls that they induce on the leaves (Gagné & Jaschoff, 2021). Species of Macrodiplosis, on the other hand, are mainly gall inducers on leaves of plant species of the genus Quercus (Kim et al., 2014).
Table 1
Number of insect species discriminated by DNA sequence-based species delineation approaches conducted in this study (2% COI divergence criterion; GMYC method).
Order/Family
Subfamily
Genus
2% DC
GMYC
Diptera
Cecidomyiidae
–
–
7
7
Lepidoptera
–
–
1
–
Coleoptera
–
–
1
–
Hymenoptera
Braconidae
Doryctinae
Allorhogas coccolobae
1
1
Eulophidae
Entedoninae
Chrysonotomyia spp.
6
8
Tetrastichinae
Quadrastichus spp.
5
5
Eupelmidae
Brasema sp.
1
1
Eurytomidae
Eurytominae
Tenuipetiolus spp.
5
7
Platygastridae
Synopeas spp.
3
3
Inostemma spp.
1
1
Undetermined
2
2
Torymidae
Torymus sp.
1
1
Total
34
38
Table 2
Insect species richness by order and family associated with 5 gall morphotypes found on leaves of Coocoloba barbadensis Jacq.
Gall morphotypes (GM)
Insect Order/Family
GM 1
GM 2
GM 3
GM 4
GM 5
TOTAL
Coleoptera
–
–
1
–
–
1
Lepidoptera
–
–
1
–
–
1
Diptera
Cecidomyiidae
3
4
5
5
2
19
Hymenoptera
15
5
13
7
2
42
Eurytomidae
4
2
3
3
–
12
Eulophidae
3
2
8
1
1
15
Eupelmidae
1
–
–
–
–
1
Braconidae
1
–
–
1
–
2
Platygastridae
5
1
2
2
1
11
Torymidae
1
–
–
–
–
1
Total
18
9
20
12
4
63
Currently, 4 gall midge species are known to be associated with species of Coccoloba. The genus Ctenodactylomyia Felt (supertribe Cecidomyiidi, unplaced tribe) has 2 leaf gall inducer species on C. diversifolia Jacq., C. swartzii, and C. uvifera L. from the Caribbean (Gagné & Jaschoff, 2021). Moreover, Marilasioptera tripartite Möhn and Meunieriella magdalenae Wünsch (supertribe Lasiopteridi, tribe Alycaulini) were described as inquilines on galls induced by other insects on species of Coccoloba from El Salvador and Colombia, respectively (Gagné & Jaschoff, 2021). The present study increases the number of gall midge species associated with Coccoloba to 11.
Our study also found a considerable number of hymenopteran species reared from the 5 examined leaf gall morphotypes, most of which probably are parasitoids of the cecidomyiid species. These parasitoid species belong to the wasp families Eulophidae, Eupelmidae, Platygastridae, and Torymidae, whereas the only reared braconid species probably are phytophagous inquiline. Some members of the Platygastridae are known to be koinobiont endoparasitoids of gallery cecidomyiid eggs, and they are known to be closely associated with the parts of the plant where the host gall is found (Masner & Huggert, 1989; Masner, 1993). Eulophidae are also mainly parasitoids of holometabolous insect larvae (though in some cases also of eggs, prepupae, and pupae) of Lepidoptera, Hymenoptera, Diptera, and Coleoptera (Graham, 1991); however, some species have been reported to be phytophagous or predators (Gibson, 1993). Finally, members of the eupelmid subfamily Eupelminae, from which the genus Brasema belongs, mainly are parasitoids of larval stages of various insect hosts (Gibson, 1993).
The wasp family Eurytomidae is represented by entomophagous species that parasitise larval or pupal stages of Coleoptera, Diptera, and Hymenoptera as solitary endoparasitic idiobionts, though some species can be phytophagous feeding on seeds, or inquilines feeding on both their host and gall tissue (Lotfalizadeh et al., 2007; Gates & Delvare, 2008). Species of Tenuipetiolus have been reported to be parasitoids of gall inducer insects, including cecidomyiids and cynipids (Gates & Hanson, 2006; Zhang et al., 2014). Similarly, several species of Torymidae are known to be ectoparasitoids of gall-forming insects of the latter 2 families (Gibson, 1993).
The braconid species A. coccolobae (Doryctinae) was described a decade ago from the Chamela region in Jalisco, Mexico, based on specimens reared from leaf galls of C. barbadensis (Martínez & Zaldívar-Riverón, 2013), that correspond to the morphotype 2. Here, this braconid species was reared not only from the above leaf gall morphotype but also from morphotype 4. Several species of Allorhogas have been confirmed to feed on various plant families either by being gall formers or inquilines of galls made by other insect taxa (Centrella & Shaw, 2010; Chavarría et al., 2009; de Mâcedo et al., 1998; Zaldívar-Riverón et al., 2014). Since none of the Allorhogas species with recorded biology are known to be parasitoids, and due to the low abundance of this species compared with the cecidomyiid species that were reared from the same gall morphotypes, it is presumed that it could be a phytophagous inquiline.
The considerably high, mostly undescribed species diversity of cecidomyiid dipterans and parasitoid hymenopterans that we reared from the 5 leaf gall morphotypes of C. barbadensis remark the necessity to carry out more studies that focus on the species diversity of galls, particularly in tropical regions. Moreover, this study highlights the importance of performing integrative species delineation studies of insects present in galls to have a more accurate knowledge of their actual diversity and trophic interactions.
Acknowledgments
We thank L. Márquez, N. López, and A. Jiménez for their assistance in the laboratory (Lanabio), and Cristina Mayorga for her assistance at the CNIN-IBUNAM. Angélica Rodríguez was supported by a MSc scholarship given by the Consejo Nacional de Humanidades, Ciencias y Tecnologías (Conahcyt, Mexico), and she thanks the Posgrado en Ciencias Biológicas, UNAM, for support during her MSc studies. This study was finished thanks to the grants given by UNAM-DGAPA (PAPIIT Convocatoria 2022, Project IN201622), and Conahcyt, Mexico (Convocatoria Ciencia de Frontera, project number 58548) to AZR.
Appendix. Continued.
Order (Insecta)
Family, Genus
Species
DNA voucher No.
Genbank accession No.
Diptera
Cecidomyiidae
sp.
CeciAN255_L2
PP659768
Diptera
Cecidomyiidae
sp.
CeciAN23_L2
PP659794
Diptera
Cecidomyiidae
sp.
CeciAN193_L2
PP659795
Diptera
Cecidomyiidae
sp.
CeciAN291_A3
PP659802
Diptera
Cecidomyiidae
sp.
CeciAN292_A1
PP659803
Diptera
Cecidomyiidae
sp.
CeciAN257_L2
PP659804
Diptera
Cecidomyiidae
sp.
CeciAN259_L3
PP659805
Diptera
Cecidomyiidae
sp.
CeciAN262_L4
PP659806
Diptera
Cecidomyiidae
sp.
CeciAN207_L4
PP659797
Diptera
Cecidomyiidae
sp.
CeciAN290_A1
PP659801
Diptera
Cecidomyiidae
sp.
CeciAN160_L3
PP659807
Diptera
Cecidomyiidae
sp.
CeciAN02_L4
PP659731
Diptera
Cecidomyiidae
sp.
CeciAN05_L4
PP659732
Diptera
Cecidomyiidae
sp.
CeciAN159_L1
PP659733
Diptera
Cecidomyiidae
sp.
CeciAN165_L2
PP659734
Diptera
Cecidomyiidae
sp.
CeciAN166_L2
PP659735
Diptera
Cecidomyiidae
sp.
CeciAN167_L3
PP659736
Diptera
Cecidomyiidae
sp.
CeciAN168_L4
PP659737
Diptera
Cecidomyiidae
sp.
CeciAN169_L4
PP659738
Diptera
Cecidomyiidae
sp.
CeciAN171_L1
PP659739
Diptera
Cecidomyiidae
sp.
CeciAN173_L1
PP659740
Diptera
Cecidomyiidae
sp.
CeciAN176_L1
PP659741
Diptera
Cecidomyiidae
sp.
CeciAN180_L2
PP659742
Diptera
Cecidomyiidae
sp.
CeciAN183_L1
PP659743
Diptera
Cecidomyiidae
sp.
CeciAN186_L5
PP659744
Diptera
Cecidomyiidae
sp.
CeciAN188_L1
PP659745
Diptera
Cecidomyiidae
sp.
CeciAN191_L1
PP659747
Diptera
Cecidomyiidae
sp.
CeciAN231_L1
PP659750
Diptera
Cecidomyiidae
sp.
CeciAN233_L1
PP659751
Diptera
Cecidomyiidae
sp.
CeciAN234_L2
PP659752
Diptera
Cecidomyiidae
sp.
CeciAN237_L1
PP659753
Diptera
Cecidomyiidae
sp.
CeciAN245_L1
PP659755
Diptera
Cecidomyiidae
sp.
CeciAN267_L1
PP659756
Diptera
Cecidomyiidae
sp.
CeciAN06_A3
PP659791
Diptera
Cecidomyiidae
sp.
CeciAN209_L5
PP659749
Diptera
Cecidomyiidae
sp.
CeciAN43_A2
PP659792
Diptera
Cecidomyiidae
sp.
CeciAN189_L3
PP659746
Diptera
Cecidomyiidae
sp.
CeciAN194_L3
PP659748
Diptera
Cecidomyiidae
sp.
CeciAN239_L4
PP659754
Hymenoptera
Platygaster
sp.
TeleAN08_L3
PP481223
Hymenoptera
Platygaster
sp.
TeleAN19_P3
PP481224
Hymenoptera
Platygaster
sp.
TeleAN46_A1
PP481225
Hymenoptera
Platygaster
sp.
TeleAN45_A3
PP481226
Hymenoptera
Platygaster
sp.
TeleAN50_A3
PP481227
Hymenoptera
Platygaster
sp.
TeleAN30_A3
PP481228
Hymenoptera
Platygaster
sp.
TeleAN132_A3
PP481229
Hymenoptera
Platygaster
sp.
TeleAN133_A1
PP481230
Hymenoptera
Platygaster
sp.
TeleAN134_A3
PP481231
Hymenoptera
Platygaster
sp.
TeleAN135_A1
PP481232
Hymenoptera
Platygaster
sp.
TeleAN137_A1
PP481233
Hymenoptera
Platygaster
sp.
TeleAN138_A3
PP481234
Hymenoptera
Platygaster
sp.
TeleAN141_A3
PP481235
Hymenoptera
Platygaster
sp.
TeleAN151_A3
PP481236
Hymenoptera
Platygaster
sp.
TeleAN152_A3
PP481237
Hymenoptera
Platygaster
sp.
TeleAN153_A3
PP481238
Hymenoptera
Platygaster
sp.
TeleAN154_A3
PP481239
Hymenoptera
Platygaster
sp.
TeleAN156_A3
PP481240
Hymenoptera
Platygaster
sp.
TeleAN215_L3
PP481241
Hymenoptera
Platygaster
sp.
TeleAN216_L3
PP481242
Hymenoptera
Platygaster
sp.
TeleAN217_L3
PP481243
Hymenoptera
Platygaster
sp.
TeleAN218_L3
PP481244
Hymenoptera
Platygaster
sp.
TeleAN224_L1
PP481245
Hymenoptera
Platygaster
sp.
TeleAN225_L1
PP481246
Hymenoptera
Platygaster
sp.
TeleAN275_L3
PP481247
Hymenoptera
Platygaster
sp.
TeleAN276_L3
PP481248
Hymenoptera
Platygaster
sp.
TeleAN281_A3
PP481249
Hymenoptera
Platygaster
sp.
TeleAN158_A1
PP481250
Hymenoptera
Platygaster
sp.
TeleAN110_A1
PP481251
Hymenoptera
Platygaster
sp.
TeleAN111_A3
PP481252
Hymenoptera
Synopeas
sp.
PlatyAN22_A1
PP481253
Hymenoptera
Synopeas
sp.
PlatyAN32_A1
PP481254
Hymenoptera
Synopeas
sp.
PlatyAN52_A1
PP481255
Hymenoptera
Synopeas
sp.
PlatyAN14_A1
PP481256
Hymenoptera
Synopeas
sp.
PlatyAN157_A1
PP481257
Hymenoptera
Synopeas
sp.
PlatyAN53_A1
PP481258
Hymenoptera
Synopeas
sp.
PlatyAN99_A1
PP481259
Hymenoptera
Synopeas
sp.
PlatyAN101_A1
PP481260
Hymenoptera
Synopeas
sp.
PlatyAN102_A1
PP481261
Hymenoptera
Synopeas
sp.
PlatyAN103_A1
PP481262
Hymenoptera
Synopeas
sp.
PlatyAN105_A1
PP481263
Hymenoptera
Synopeas
sp.
PlatyAN190_L2
PP481264
Hymenoptera
Synopeas
sp.
PlatyAN219_L1
PP481265
Hymenoptera
Inostemma
sp.
PlatyAN221_L4
PP481266
Hymenoptera
Synopeas
sp.
PlatyAN223_L2
PP481267
Hymenoptera
Synopeas
sp.
PlatyAN220_L5
PP481268
Hymenoptera
Inostemma
sp.
PlatyAN112_A4
PP481269
Hymenoptera
Synopeas
sp.
PlatyAN116_A1
PP481270
Hymenoptera
Synopeas
sp.
PlatyAN117_A1
PP481271
Hymenoptera
Synopeas
sp.
PlatyAN118_A1
PP481272
Hymenoptera
Inostemma
sp.
PlatyAN126_A1
PP481273
Hymenoptera
Synopeas
sp.
PlatyAN214_L1
PP481274
Hymenoptera
Platygaster
sp.
PlatyAN222_L3
PP481275
Hymenoptera
Synopeas
sp.
PlatyAN272_A1
PP481276
Hymenoptera
Synopeas
sp.
PlatyAN273_L
PP481277
Hymenoptera
Synopeas
sp.
PlatyAN274_L
PP481278
Hymenoptera
Synopeas
sp.
PlatyAN277_L5
PP481279
Hymenoptera
Synopeas
sp.
PlatyAN278_L2
PP481280
Hymenoptera
Synopeas
sp.
PlatyAN279_L1
PP481281
Hymenoptera
Tenuipetiolus
sp.
EurytAN96_A1
PP481282
Hymenoptera
Tenuipetiolus
sp.
EurytAN265_L3
PP481283
Hymenoptera
Chrysonotomyia
sp.
EntedAN01_P1
PP481284
Hymenoptera
Chrysonotomyia
sp.
EntedAN07_A1
PP481285
Hymenoptera
Chrysonotomyia
sp.
EntedAN29_A1
PP481286
Hymenoptera
Chrysonotomyia
sp.
EntedAN11_A1
PP481287
Hymenoptera
Chrysonotomyia
sp.
EntedAN31_A1
PP481288
Hymenoptera
Chrysonotomyia
sp.
EntedAN51_A1
PP481289
Hymenoptera
Chrysonotomyia
sp.
EntedAN59_P1
PP481290
Hymenoptera
Chrysonotomyia
sp.
EntedAN61_P1
PP481291
Hymenoptera
Chrysonotomyia
sp.
EntedAN67_P1
PP481292
Hymenoptera
Chrysonotomyia
sp.
EntedAN69_A1
PP481293
Hymenoptera
Chrysonotomyia
sp.
EntedAN70_A1
PP481294
Hymenoptera
Chrysonotomyia
sp.
EntedAN71_A1
PP481295
Hymenoptera
Chrysonotomyia
sp.
EntedAN72_A1
PP481296
Hymenoptera
Chrysonotomyia
sp.
EntedAN75_A1
PP481297
Hymenoptera
Chrysonotomyia
sp.
EntedAN94_A1
PP481298
Hymenoptera
Chrysonotomyia
sp.
EntedAN95_A1
PP481299
Hymenoptera
Chrysonotomyia
sp.
EntedAN25_A2
PP481300
Hymenoptera
Chrysonotomyia
sp.
EntedAN64_P4
PP481301
Hymenoptera
Chrysonotomyia
sp.
EntedAN73_A3
PP481302
Hymenoptera
Chrysonotomyia
sp.
EntedAN142_A1
PP481303
Hymenoptera
Chrysonotomyia
sp.
EntedAN143_A1
PP481304
Hymenoptera
Chrysonotomyia
sp.
EntedAN144_A1
PP481305
Hymenoptera
Chrysonotomyia
sp.
EntedAN145_A1
PP481306
Hymenoptera
Chrysonotomyia
sp.
EntedAN146_A2
PP481307
Hymenoptera
Chrysonotomyia
sp.
EntedAN198_L3
PP481308
Hymenoptera
Chrysonotomyia
sp.
EntedAN236_L1
PP481309
Hymenoptera
Chrysonotomyia
sp.
EntedAN243_L1
PP481310
Hymenoptera
Chrysonotomyia
sp.
EntedAN244_L1
PP481311
Hymenoptera
Chrysonotomyia
sp.
EntedAN288_A2
PP481312
Hymenoptera
Chrysonotomyia
sp.
EntedAN289_A2
PP481313
Hymenoptera
Chrysonotomyia
sp.
EntedAN58_P1
PP481314
Hymenoptera
Chrysonotomyia
sp.
EntedAN55_A1
PP481315
Hymenoptera
Chrysonotomyia
sp.
EulopAN125_A5
PP481316
Hymenoptera
Tetrastichinae
sp.
TetrasAN84_A3
PP481317
Hymenoptera
Tetrastichinae
sp.
TetrasAN62_P3
PP481318
Hymenoptera
Tetrastichinae
sp.
TetrasAN63_P3
PP481319
Hymenoptera
Tetrastichinae
sp.
TetrasAN66_P3
PP481320
Hymenoptera
Tetrastichinae
sp.
TetrasAN83_A3
PP481321
Hymenoptera
Tetrastichinae
sp.
TetrasAN60_P3
PP481322
Hymenoptera
Tetrastichinae
sp.
TetrasAN269_L3
PP481323
Hymenoptera
Tetrastichinae
sp.
TetrasAN270_L3
PP481324
Hymenoptera
Tenuipetiolus
sp.
EurytAN77_A1
PP481325
Hymenoptera
Tenuipetiolus
sp.
EurytAN81_A3
PP481326
Hymenoptera
Tenuipetiolus
sp.
EurytAN93_A3
PP481327
Hymenoptera
Allorhogas
coccolobae
AllorAN18_L4
PP481328
Hymenoptera
Allorhogas
coccolobae
AllorAN28_A4
PP481329
Hymenoptera
Allorhogas
coccolobae
AllorAN35_A4
PP481330
Hymenoptera
Allorhogas
coccolobae
AllorAN48_A4
PP481331
Hymenoptera
Allorhogas
coccolobae
AllorAN56_A4
PP481332
Hymenoptera
Allorhogas
coccolobae
AllorAN107_A4
PP481333
Hymenoptera
Allorhogas
coccolobae
AllorAN108_A4
PP481334
Hymenoptera
Allorhogas
coccolobae
AllorAN109_A4
PP481335
Hymenoptera
Allorhogas
coccolobae
AllorAN121_A4
PP481336
Hymenoptera
Allorhogas
coccolobae
AllorAN122_A4
PP481337
Hymenoptera
Allorhogas
coccolobae
AllorAN195_L1
PP481338
Hymenoptera
Allorhogas
coccolobae
AllorAN248_L4
PP481339
Hymenoptera
Allorhogas
coccolobae
AllorAN249_L4
PP481340
Hymenoptera
Tenuipetiolus
sp.
EurytAN212_L3
PP481341
Hymenoptera
Tenuipetiolus
sp.
EurytAN228_L3
PP481342
Hymenoptera
Tenuipetiolus
sp.
EurytAN246_L3
PP481343
Hymenoptera
Tenuipetiolus
sp.
EurytAN285_A3
PP481344
Hymenoptera
Tenuipetiolus
sp.
EurytAN286_A3
PP481345
Hymenoptera
Tenuipetiolus
sp.
EurytAN37_A1
PP481346
Hymenoptera
Tenuipetiolus
sp.
EurytAN147_A3
PP481347
Hymenoptera
Tenuipetiolus
sp.
EurytAN148_A4
PP481348
Hymenoptera
Tenuipetiolus
sp.
EurytAN149_A3
PP481349
Hymenoptera
Tenuipetiolus
sp.
EurytAN130_A1
PP481350
Hymenoptera
Tenuipetiolus
sp.
EurytAN87_A2
PP481351
Hymenoptera
Tenuipetiolus
sp.
EurytAN91_A4
PP481352
Hymenoptera
Tenuipetiolus
sp.
EurytAN80_A3
PP481353
Hymenoptera
Tenuipetiolus
sp.
EurytAN82_A4
PP481354
Hymenoptera
Tenuipetiolus
sp.
EurytAN90_A4
PP481355
Hymenoptera
Tenuipetiolus
sp.
EurytAN85_A1
PP481356
Hymenoptera
Torymus
sp.
EurytAN57_A4
PP481357
Hymenoptera
Tenuipetiolus
sp.
EurytAN164_L1
PP481358
Hymenoptera
Tenuipetiolus
sp.
EurytAN227_L1
PP481359
Hymenoptera
Tenuipetiolus
sp.
EurytAN230_L1
PP481360
Hymenoptera
Tenuipetiolus
sp.
EurytAN283_A2
PP481361
Hymenoptera
Tenuipetiolus
sp.
EurytAN33_A4
PP481362
Hymenoptera
Tenuipetiolus
sp.
EurytAN34_A1
PP481363
Hymenoptera
Tenuipetiolus
sp.
EurytAN15_L1
PP481364
Hymenoptera
Tenuipetiolus
sp.
EurytAN114_A3
PP481365
Hymenoptera
Tenuipetiolus
sp.
EurytAN79_A1
PP481366
Hymenoptera
Tenuipetiolus
sp.
EurytAN78_A1
PP481367
Hymenoptera
Tenuipetiolus
sp.
EurytAN24_L1
PP481368
Hymenoptera
Chrysonotomyia
sp.
EurytAN88_A3
PP481369
Hymenoptera
Brasema
sp.
ToryAN170_L1
PP481370
Hymenoptera
Brasema
sp.
ToryAN196_L1
PP481371
Hymenoptera
Torymus
sp.
EurytAN229_L4
PP481372
Hymenoptera
Torymus
sp.
EurytAN266_L4
PP481373
Coleoptera
AN184
PP897664
Lepidoptera
AN250_L3
PP897665
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Bouckaert, R., Heled, J., Kühnert, D., Vaughan, T., Wu, C. H., Xie, D., Suchard, M. A., Rambaut, A., & Drummond, A. J. (2014). BEAST 2: A software platform for Bayesian evolutionary analysis. Plos Computational Biology, 10, e1003537. https://doi.org/10.1371/journal.pcbi.1003537
Centrella, M. L., & Shaw, S. R. (2010). A new species of phytophagous braconid Allorhogas minimus (Hymenoptera: Braconidae: Doryctinae) reared from fruit galls on Miconia longifolia (Melastomataceae) in Costa Rica. International Journal of Tropical Insect Science, 30, 101–107. https://doi.org/10.1017/S1742758410000147
Chavarría, L., Hanson, P., Marsh, P., & Shaw, S. (2009). A phytophagous braconid, Allorhogas conostegia sp. nov.(Hymenoptera: Braconidae), in the fruits of Conostegiaxalapensis (Bonpl.) D. Don (Melastomataceae). Journal of Natural History, 43, 2677–2689. https://doi.org/10.1080/ 00222930903243996
Clare, E. L., Schiestl, F. P., Leitch, A. R., & Chittka, L. (2013). The promise of genomics in the study of plant-pollinator interactions. Genome Biology, 14, 1–11. https://doi.org/10. 1186/gb-2013-14-6-207
de Macêdo, M. V., & Monteiro, R. F. (1989). Seed predation by a braconid wasp, Allorhogas sp. (Hymenoptera). Journal of the New York Entomological Society, 97, 358–362.
Dexter, K. G., Pennington, T. D., & Cunningham, C. W. (2010). Using DNA to assess errors in tropical tree identifications: How often are ecologists wrong and when does it matter? Ecological Monographs, 80, 267–286. https://doi.org/10.1890/09-0267.1
Drummond, A. J., Suchard, M. A., Xie, D., & Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution, 29, 1969–1973. https://doi.org/10.1093/molbev/mss075
Folmer O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
Forbes, A. A., Hall, M. C., Lund, J., Hood, G. R., Izen, R., Egan, S. P. et al. (2016). Parasitoids, hyperparasitoids, and inquilines associated with the sexual and asexual generations of the gall former, Belonocnema treatae (Hymenoptera: Cynipidae). Annals of the Entomological Society of America, 109, 49–63. https://doi.org/10.1093/aesa/sav112
Gates, M. W., & Hanson, P. E. (2006). Familia Eurytomidae, In P. E. Hanson, & I. D. Gauld (Eds.), Hymenoptera de la region tropical (pp. 380–387). Memoirs of the American Entomological Institute, 77, 1–994.
Gates, M., & Delvare, G. (2008). A new species of Eurytoma (Hymenoptera: Eurytomidae) attacking Quadrastichus spp. (Hymenoptera: Eulophidae) galling Erythrina spp. (Fabaceae), with a summary of African Eurytoma biology and species checklist. Zootaxa, 1751, 1–24. https://doi.org/10.11646/zootaxa.1751.1.1
Gibson, L. (1993). Superfamilies Mymarommatoidea and Chalcidoidea. In H. Goulet, & J. T. Huber (Eds.), Hymenoptera of the world: an identification guide to families (pp.570–655). Research Branch, Agriculture, Canada, Publication 1894.
Graham, M. W. R. V. (1991). A reclassification of the European Tetrastichinae (Hymenoptera: Eulophidae). Revision of the remaining genera. Memoirs of the American Entomological Institute, 49, 1–322.
Hebert, P. D., Cywinska, A., Ball, S. L., & DeWaard, J R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 2070, 313–321. https://doi.org/10.1098/rspb.2002.2218
Hebert, P. D., Penton, E. H., Burns, J. M., Janzen, H. D., & Hallwachs, W. (2004). Ten species in one: DNA barcoding reveals cryptic species in the Neotropical skipper butterfly Astrapes fulgerator. Proceeding of the National Academy of Sciences, 101, 14812–14817. https://doi.org/10.1073/pnas. 0406166101
Howard, R. A. (1959). Studies in the genus Coccoloba. VII A synopsis and key to species in Mexico and Central America. Journal of the Arnold Arboretum, 40, 176–203.
Ings, T. C., Montoya, J. M., Bascompte, J., Blüthgen, N., Brown, L., Dormann, C. F. et al. (2009). Ecological networks-beyond food webs. Journal of Animal Ecology, 78, 253–269. https://doi.org/10.1111/j.1365-2656.2008.01460.x
Kaartinen, R., Stone, G. N., Hearn, J., Lohse, K., & Roslin, T. (2010). Revealing secret liaisons: DNA barcoding changes our understanding of food webs. Ecological Entomology, 35, 623–638. https://doi.org/10.1111/j.1365-2311.2010.01224.x
Kaartinen, R., & Roslin, T. (2011). Shrinking by numbers: landscape context affects the species composition but not the quantitative structure of local food webs. Journal of Animal Ecology, 80, 622–631. https://doi.org/10.1111/ j.1365-2656.2011.01811.x
Kim, W., Yukawa, J., Harris, K. M., Minami, T., Matsuo, K., & Skrzypczyńska, M. (2014). Description, host range, and distribution of a new Macrodiplosis species (Diptera: Cecidomyiidae) that induces leaf-margin fold galls on deciduous Quercus (Fagaceae) with comparative notes on Palaearctic congeners. Zootaxa, 3821, 222–238. https://doi.org/10.11646/zootaxa.3821.2.3
Lotfalizadeh, H., Delvare, G., & Rasplus, J. Y. (2007). Phylogenetic analysis of Eurytominae (Chalcidoidea: Eurytomidae) based on morphological characters. Zoological Journal of the Linnean Society, 151, 441–510. https://doi.org/10.1111/j.1096-3642.2007.00308.x
Maia, V. C., Magenta, M. A. G., & Martins, S. E. (2008). Ocorrência e caracterização de galhas de insetos em áreas de restinga de Bertioga (São Paulo, Brasil). Biota Neotropica, 8, 167–197. https://doi.org/10.1590/S1676-06032 008000100020
Maia, V. C. (2012). Richness of hymenopterous galls from South America. Papéis Avulsos de Zoologia, 52, 423–429. https://doi.org/10.1590/S0031-10492012021500001
Mani, M. S. (1964). Ecology of plant galls. The Hague: W. Junk.
Martínez, J. J., & Zaldívar-Riverón, A. (2013). Seven new species of Allorhogas (Hymenoptera: Braconidae: Doryctinae) from Mexico. Revista Mexicana de Biodiversidad, 84, 117–139. https://doi.org/10.7550/rmb.31955
Masner, L., & Huggert, L. (1989). World review and keys to genera of the subfamily Inostemmatinae with the reassignment of the taxa to the Platygastrinae and Sceliotrachelinae (Hymenoptera: Platygastridae). The Memoirs of the Entomological Society of Canada, 121, 3–216.
Masner, L. (1993). Superfamily Platygastroidea. In H. Goulet, & J. T. Huber (Eds.), Hymenoptera of the world: an identification guide to families (pp.558–565). Research Branch, Agriculture, Canada, Publication 1894.
Mead, F. W. (1970). Ctenodactylomyia watsoni Felt, a gall midge of seagrape, Coccoloba uvifera L. Florida (Diptera: Cecidomyiidae). Entomology Circular, 97. Florida Department of Agriculture & Consumer Services Division of Plant Industry. Florida, USA.
Méndez-Alonzo, R., Pineda-García, F., Paz, H., Rosell, J. A., & Olson, M. E. (2013). Leaf phenology is associated with soil water availability and xylem traits in a tropical dry forest. Trees, 27, 745–754. https://doi.org/10.1007/s00468-012-0829-x
Nieves-Aldrey, J. L. (1998). Insectos que inducen la formación de agallas en las plantas: Una fascinante interacción ecológica y evolutiva. Boletín de la Sociedad Entomológica Aragonesa, 23, 3–12.
Noguera, F. A., Vega-Rivera, J. H., García-Aldrete, A. N., Quesada-Avendaño, M. (Eds.). (2002). Historia natural de Chamela. Mexico City: Universidad Nacional Autónoma de México.
Paine, R. T. (1980). Food webs: linkage, interaction strength, and community infrastructure. Journal of Animal Ecology, 49, 667–685. https://doi.org/10.2307/4220
Pons, J., Barraclough, T. G., Gomez-Zurita, J., Cardoso, A., Duran, D. P., Hazell, S., Kamoun, S., Sumlin, W. D., & Vogler, A. P. (2006). Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Systematic Biology, 55, 595-609. https://doi.org/10.1080/10635150600852011
Posada, D. (2004). Collapse: describing haplotypes from sequence alignments. Vigo, Spain: University of Vigo. https://dposada.webs.uvigo.es/
Prosser, S., Martínez-Arce, A., & Elías-Gutiérrez, M. (2013). A new set of primers for COI amplification from freshwater microcrustaceans. Molecular Ecology Resources, 13, 1151–1155. https://doi.org/10.1111/1755-0998.12132
Raman, A., & Withers, T. M. (2003). Oviposition by introduced Ophelimus eucalypti (Hymenoptera: Eulophidae) and morphogenesis of female-induced galls on Eucalyptus saligna (Myrtaceae) in New Zealand. Bulletin of Entomological Research, 93, 55–63. https://doi.org/10.1079/BER2002217
Raman, A. (2011). Morphogenesis of insect-induced plant galls facts and questions. Flora-Morphology, Distribution, Functional Ecology of Plants, 206, 517–533. https://doi.org/10.1016/j.flora.2010.08.004
Ratnasingham, S., & Hebert, P. D. N. (2013). A DNA-based registry for all animal species: the barcode index number (BIN) system. Plos One, 8, e66213. https://doi.org/10.1371/journal.pone.0066213
Razo-González, M., Castano-Meneses, G., Callejas-Chavero, A., Pérez-Velázquez, D., & Palacios-Vargas, J. G. (2014). Temporal variations of soil arthropods community structure in El Pedregal de San Ángel ecological reserve, Mexico City, Mexico. Applied Soil Ecology, 83, 88–94. https://doi.org/10.1016/j.apsoil.2014.02.007
Redfern, M., Shirley, P., & Bloxham, M. (2002). British plant galls: identification of galls on plants and fungi. Shrewsbury, UK: FSC publications.
Rodrigues, A. R., Maia, V. C., & Couri, M. S. (2014). Insect galls of restinga areas of Ilha da Marambaia, Rio de Janeiro, Brazil. Revista Brasileira de Entomologia, 58, 173–197. https://doi.org/10.1590/S0085-56262014000200010
Sheffield, C. S., Hebert, P. D. N., Kevan, P. G., & Packer, L. (2009). DNA barcoding a regional bee (Hymenoptera: Apoidea) fauna and its potential for ecological studies. Molecular Ecology Resources, 9, 196–207. https://doi.org/10.1111/j.1755-0998.2009.02645.x
Statsoft, Inc. (2011). STATISTICA (Data Analysis Software System, version 10. http://www.statsoft.com
Swofford, D. L. (2003). PAUP*. Phylogenetic Analysis using parsimony (*and other methods). Version 4.0. Sinauer Associates, Sunderland, Massachusetts.
Uechi, N., Yukawa, J., Tokuda, M., Ganaha-Kikumura, T., & Taniguchi, M. (2011). New information on host plants and distribution ranges of an invasive gall midge, Contarinia maculipennis (Diptera: Cecidomyiidae), and its congeners in Japan. Applied Entomology and Zoology, 46, 383–389. https://doi.org/10.1007/s13355-011-0050-1
Vink, C. J., Paquin, P., & Cruickshank, R. H. (2012). Taxonomy and irreproducible biological science. Bioscience, 62, 451–452. https://doi.org/10.1525/bio.2012.62.5.3
Wirta, H. K., Hebert, P. D., Kaartinen, R., Prosser, S. W., Várkonyi, G., & Roslin, T. (2014). Complementary molecular information changes our perception of food web structure. Proceedings of the National Academy of Sciences, 111, 1885–1890. https://doi.org/10.1073/pnas.1316990111
Yeo, D., Puniamoorthy, J., Ngiam, R. W. J., & Meier, R. (2018). Towards holomorphology in entomology: rapid and cost-effective adult–larva matching using NGS barcodes. Systematic Entomology, 43, 678–691. https://doi.org/10.1111/syen.12296
Zaldívar-Riverón, A., Martínez, J. J., Belokobylskij, S. A., Pedraza-Lara, C., shaw, S. R., Hanson, P. E., & Varela-Hernández, F. (2014). Systematics and evolution of gall formation in the plant-associated genera of the wasp subfamily Doryctinae (Hymenoptera: Braconidae). Systematic Entomology, 39, 633–659. https://doi.org/10.1111/syen.12078
Zhang, M. Y., Gates, M. W., & Shorthouse, J. D. (2014). Testing species limits of Eurytomidae (Hymenoptera) associated with galls induced by Diplolepis (Hymenoptera: Cynipidae) in Canada using an integrative approach. The Canadian Entomologist, 146, 321–334. https://doi.org/10.4039/tce. 2013.70
Carlos Hiram Rábago-Quiroz a, Adrián Felipe González-Acosta b, *, Gorgonio Ruiz-Campos c, Jonathan Franco-López d, Juan Antonio García-Borbón a, Joaquín D. Magaña e
a Instituto Mexicano de Investigación en Pesca y Acuacultura Sustentables, Centro Regional de Investigación Acuícola y Pesquera, La Paz, Km. 1 Carretera Pichilingue s/n, Col. Esterito, 23020 La Paz, Baja California Sur, Mexico
b Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, Av. Instituto Politécnico Nacional s/n, Col. Playa Palo de Santa Rita, 23096 La Paz, Baja California Sur, Mexico
c Universidad Autónoma de Baja California, Facultad de Ciencias, Colección Ictiológica, Carretera Transpeninsular Ensenada-Tijuana Núm. 3917, Colonia Playitas, 22860 Ensenada, Baja California, Mexico
d Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Iztacala, Laboratorio de Ecología, Av. De los Barrios Núm. 1, Colonia Los Reyes Ixtacala, 54090 Tlalnepantla, Estado de México, Mexico
e University of Belize, Belmopan Campus, P.O. Box 340, Cayo, Belize
Received: 20 June 2023; accepted: 21 February 2024
Abstract
We present an updated systematic checklist of fishes from the Bahía Magdalena-Almejas lagoon system (Mexico), including notes on taxonomy, zoogeography, and conservation status, supported by field samplings and the critical review of published and online data. The ichthyofauna is composed by 2 classes, 30 orders, 104 families, 240 genera and 416 species, mainly represented by the class Actinopterygii. Zoogeographically, the fishes showed highest affinity to the San Diego (87.2%), Cortez (81.4%) and Mexican (70%) provinces, reflecting the ancient relationship between the Californian and Tropical Eastern Pacific regions and the existence of a transitional zone in the Baja California Peninsula for fish assemblages from boreal-temperate and warm-tropical derivation, standing out the presence of the endemic Paraclinus magdalenae. The 91% of the ichthyofauna is included by the IUCN Red List as Lower Concern, Data Deficient and Not Evaluated, while Holacanthus clarionensis and Hippocampus ingens, are under Special Protection by the Mexican regulation.Ecologically, 233 species are marine-euryhaline and 177 marine-stenohaline, which preferentially inhabit soft (57%) and rocky (27.5%) bottoms; most species are demersal (60%), benthic (25%) and pelagic (14.4%). The updated checklist could improve the design and implementation of effective fishing regulation strategies and conservation programs for fishes inhabiting this coastal ecosystem.
Keywords: Fish diversity; Conservation status; Taxonomy; Mexican Pacific; Checklist
Composición y zoogeografía de peces marino-estuarinos de un sistema lagunar en una zona de transición templado-tropical del Pacífico oriental
Resumen
Se presenta la lista sistemática actualizada de los peces del sistema lagunar Bahía-Magdalena-Almejas, con notas taxonómicas, zoogeografía y estado de conservación; basada en recolectas biológicas y la revisión crítica de datos publicados. La lista la integran 2 clases, 30 órdenes, 104 familias, 240 géneros y 416 especies, mayormente representados por Actinopterygii. La mayor afinidad zoogeográfica con las provincias de San Diego (87.2%), de Cortés (81.4%) y Mexicana (70%), resulta de la relación ancestral entre las regiones de California y Pacífico oriental tropical y debido a la existencia de una zona de transición en la península de Baja California, para conjuntos ícticos de derivación boreal-templada y subtropical-tropical, donde destaca el endémico Paraclinus magdalenae. De acuerdo con la Lista Roja de la UICN, 91% de la ictiofauna es de preocupación menor, datos deficientes y no evaluado; mientras que Holacanthus clarionensis e Hippocampus ingens, están bajo protección especial por la legislación mexicana.Ecológicamente, 233 especies son marino-eurihalinas y 177 marino-estenohalinas, habitan preferentemente sobre fondos suaves (57%) y rocosos (27.5%); 60% son demersales, 25% bentónicas y 14% y pelágicas. La lista sistemática actualizada permitirá formular una estrategia efectiva de regulación pesquera y programas de conservación en este ecosistema.
Palabras clave: Diversidad íctica; Estado de conservación; Taxonomía; Pacífico mexicano; Listado sistemático
Introduction
The western coast of the Baja California peninsula (BCP) is recognized as a biogeographic region with high fish diversity (Castro-Aguirre & Espinosa-Pérez, 2006; Hastings, 2000; Ruiz-Campos et al., 2010), and one of the most productive fishing areas worldwide (Finkbeiner & Basurto, 2015; Ojeda-Ruiz et al., 2018). Within this region, the Bahía Magdalena-Almejas (BMA) lagoon system represents the largest and the main fishing zone in the northwestern Mexican Pacific region, including the Baja California Sur state (Avendaño-Ibarra et al., 2004; Cota-Nieto et al., 2015; Ojeda-Ruiz et al., 2018).
The privileged geomorphology and geographic location of the BMA lagoon system, in the transition zone between the Californian and Tropical Eastern Pacific regions (Briggs, 1974; Castro-Aguirre & Espinosa-Pérez, 2006; Castro-Aguirre et al., 1992, 1993; González-Acosta, Ruiz-Campos et al., 2015; Hastings, 2000; Parrish et al.,1981; Ruiz-Campos et al., 2010), promote the existence of a large extension (17 million m2; Acosta-Velázquez & Ruiz-Luna, 2007) of mangrove biotopes that house a great diversity of fishes from temperate and tropical derivation (Etnoyer et al., 2004; Funes-Rodríguez et al., 2007; Hernández-Velasco et al., 2016; Malakoff, 2004; Whitmore et al., 2005). Thereby, plays an important role as the primary habitat or nursery grounds for feeding, spawn, and recruitment for young and adult of commercial fish species as well as for those subject to ecological conservation (Cota-Nieto et al., 2015; Hastings & Fischer, 2001); which mostly are representative of pelagic and demersal fish species that use these habitats alternating seasonality their abundances, generating critical links in the food chains between the lagoon system and the adjacent sea (González-Acosta, Ruiz-Campos et al., 2015).
The BMA lagoon system is the second most studied and richest fish area in BCP (Hinojosa-Medina et al., 2007), after Bahía de La Paz (González-Acosta, Balart et al., 2018; González-Acosta, Ruiz-Campos et al., 2015); on this basis, the fish fauna previously reported for BMA varies between 92 to 302 species (Castro-Aguirre et al., 1993; Galván-Magaña et al., 2000). However, given that the specific richness of this ecosystem has substantially increased since its fish fauna has been more deeply studied and new taxa are reported or formally described, this study aims to document and update the systematic checklist of the marine and estuarine fish species occurring in the BMA lagoon system, including notes on taxonomy, zoogeography, and conservation status.
Material and methods
The BMA is an extensive estuarine lagoon system (114,600 ha) bordered by islands located parallel to the coast (Magdalena, Margarita and Creciente), coastal dunes, and extensive sandbars with sparse vegetation. This coastal ecosystem is located in the southwestern portion of the BCP (24°21’ – 24°46’ N, 110°30’- 112°15’ W; Fig. 1), which comprises 3 subsystems: the Northwestern zone (137 km2), characterized by the presence of negative estuaries (locally named “esteros”), marshes and channels with a mean depth of 3.6 m; the Central zone that comprises Bahía Magdalena (883 km2) with a mean depth of 12 – 15 m, which connects with the adjacent sea via a 5.6 km wide inlet (≥ 40 m deep); and the Southern portion comprised by Bahía Almejas (370 km2) that is characterized by shallow plains that are exposed at low tides, as well as a deep (~ 30 m) and wide (2-3 km) channel that communicates with BM, and empties into the sea through a shallow inlet (Álvarez-Borrego et al., 1975; Castro-Aguirre & Torres-Orozco, 1993; Lewis & Ebeling, 1971; Ojeda-Ruiz et al., 2018).
The hydrology of the BMA lagoon system is influenced during the Spring-Summer seasons by the cool California Current and from Autumn-Winter by the warm subtropical California Countercurrent with high evaporation rates in the shallow areas and high-water transport in deep zones of channels and lagoons, and upwellings in front of the mouth (Álvarez-Borrego et al., 1975; Avendaño-Ibarra et al., 2004; Funes-Rodríguez et al., 2007; Gómez-Valdez & Vélez-Muñoz, 1982; Lluch-Belda et al., 2000). The surface temperature ranges from 18 to 29 °C and the dissolved oxygen from 0.04 to 2.56 mg/l, tides are semi-diurnal. The salinity varies from 34 to 40 ups, due to the absence of rain and river runoffs; therefore, this lagoon system behaves throughout the year as a negative estuary (Álvarez-Borrego et al., 1975; Castro-Aguirre & Torres-Orozco, 1993), as is common in this arid region (Brusca et al., 2005; González-Acosta, Rabadán-Sotelo et al., 2015; González-Acosta, Ruiz-Campos et al., 2015).
The systematic checklist of fishes inhabiting the BMA lagoon system was based on fish specimens collected during the study period from March 2014 to June 2018 in several sites along this ecosystem, using a shrimp trawl net (16 -17 m long headline and 3.5 cm mesh size), deployed at 2 knots for 1 hour. All caught specimens were frozen and transported to the fish laboratory at Centro Regional de Investigación Acuícola y Pesquera-La Paz of the Instituto Mexicano de Investigación en Pesca y Acuacultura Sustentables (IMIPAS-CRIAP), to be fixed with 10% formalin and later preserved in 70% ethanol. The taxonomic identification was based on specialized taxonomic keys and fish catalogs (Allen & Robertson, 1994; Castro-Aguirre et al., 1999; Fischer et al., 1995; Love & Passarelli, 2020; Love et al., 2021), and online databases (e.g., Robertson & Allen, 2015). Some specimens of each species were housed as vouchers in the Fish Collection of the IMIPAS.
The systematic checklist also includes documented fish records (including larvae) compiled throughout the critical taxonomic review of previous and current checklist species published in specialized literature (Avendaño-Ibarra et al., 2004; Castro-Aguirre & Torres-Orozco, 1993; Castro-Aguirre et al., 1992; De la Cruz-Agüero et al., 1994; Funes-Rodríguez et al., 2007; Galván-Magaña et al., 2000; González-Acosta, Ruiz-Campos et al., 2015), and online databases of reference fish collections. The taxonomic arrangement (nomenclature and classification) of the systematic checklist follows to Page et al. (2013) and Fricke et al. (2023), respectively. The geographic distribution records for each species were confirmed in the online databases of Peces Costeros del Pacífico Oriental Tropical (Robertson & Allen, 2015), the Global Biodiversity Information Facility (GBIF, 2023), the Eschmeyer’s Catalog of Fishes (Fricke et al., 2023) and FISHBASE data (Froese & Pauly, 2023); thereby, the zoogeographic affinity of the fish fauna was determined following Briggs´s regionalization (Briggs, 1974), as well as by their distributional patterns based on Castro-Aguirre et al. (2005, 2006). Conservation status and ecology of the fish species here included were documented based on the Red List of Threatened Species of the International Union for Conservation of Nature (IUCN, 2022) and the Mexican Flora and Fauna Protection Regulation NOM-059-SEMARNAT-2010 (DOF, 2010, 2019), with complementary information obtained from published studies involving local fish fauna (Castro-Aguirre & Torres-Orozco, 1993; Castro-Aguirre et al., 1992, 1993; Fischer et al. 1995; Froese & Pauly, 2023; Funes-Rodríguez et al., 2007; González-Acosta, Ruiz-Campos et al., 2015; Love & Passarelli, 2020; Love et al., 2021; Robertson & Allen, 2015).
Results
The BMA fish fauna is composed of 2 classes, 30 orders, 104 families, 240 genera and 416 species (Table 1, Fig. 2). The class Chondrichthyes groups 45 species, 25 genera, 18 families, 8 orders, and the subclass Elasmobranchii, represented by 2 divisions: Selachii (sharks) composed by 5 orders, 7 families, 12 genera and 23 species, most of them corresponding to the order Carcharhiniformes (3 families, 8 genera and 18 species) and the family Carcharhinidae (Carcharhinus Blainville, 1816 and Sphyraena Artedi, 1793 with 4 species each); and Batomorphi (rays and skates) represented by 3 orders, 11 families,13 genera and 22 species, predominating the order Myliobatiformes (15 species, 8 genera and 7 families) and the family Urotrygonidae (2 genera and 7 species).
The class Actinopterygii encompassess 89.2% of the total fish fauna of the BMA lagoon system, represented by 22 orders, 86 families, 215 genera and 371 species, with the representativeness of the order Perciformes (45 families, 134 genera and 224 species); while the families: Sciaenidae (27 species), Haemulidae (22 species), Carangidae (18 species), Gobiidae (15 species), Serranidae (12 species), and Epinephelidae (11 species), comprise 46.9% of the order, 28.3% of the class and 25.2% of the total richness in this lagoon system. Fish larvae are represented by 76 species (20.5% of the total actinopterygian fishes) grouped in 58 genera, 36 families and 11 orders, most of the species correspond to the order Perciformes, of which Gobiidae and Labrisomidae are the most diverse families with 5 species each.
Based on the assessment of the zoogeographic affinity, the fish fauna of BMA lagoon system shows affinity to the Auletian (18 species, 4.3%), Oregonian (53 species, 13%), San Diego (362 species, 87.2%), Cortez (338 species, 81.4%), Mexican (290 species, 70%), Panamian (262 species, 63.1%), Galapagos (81 species, 20%), and Chilean-Peruvian (8 species, 1.9%) provinces. According to their reported distribution range, only the Magdalena blenny Paraclinus magdalenae Rosenblatt & Parr, 1969 (0.2%) is endemic, 38 species are circumtropical (9.2%), 4 amphipacific (1%), and 5 amphiamerican (1.2%).
Figure 1. Location of the Bahía Magdalena-Almejas lagoon system in the northeastern Pacific of Mexico. Sampling trawl fishing sites are indicated in grey.
Based on the Red List criteria of the IUCN (IUCN, 2022), 3 (0.7%) species are “Critically Endangered” (2 chondrichthyans: the Scalloped hammerhead, Sphyraena lewini (Grifit and Smith, 1834]) and the Great hammerhead, Sphyrna mokarran (Rüpell, 1832), and the Giant seabass Stereolepis gigas Ayres, 1859); 19 (4.6%) are “Vulnerable” (6 selachians, 6 batoids and 7 actinopterygians); 6 (1.5%) are “Endangered” (4 selachians and 2 actinopterygians); 10 (2.4%) are “Near Threatened” (3 selachians and 7 batoids); 17 (4.1%) are “Not Evaluated” (all actinopterygians); 21 (5.1%) are “Data Deficient”(1 selachian, 2 batoids and 18 actinopterygians); and 340 (81%) are of “Least Concern” (6 selachians, 6 batoids and 328 actinopterygians) (Table 1). According to the latest update of the NOM-059-SEMARNAT-2010 (DOF, 2019), 2 (0.5%) species are under “Special Protection (Pr)”: the Clarion angelfish Holacanthus clarionensis Gilbert, 1891 and the Pacific seashore Hippocampus ingens Girard, 1855.
Table 1 Systematic list of estuarine-marine fishes of the Bahía Magdalena-Almejas lagoon system (México).
Taxa
ZA
IUCN
NOM
D
S
H
Class Chondrichthyes
Subclass Elasmobranchii
Order Heterodontiformes
Family Heterodontidae
Heterodontus francisci (Girard, 1855)
SD, C, M, P
DD
ES
S
B
Heterodontus mexicanus Taylor & Castro-Aguirre, 1972
SD, C, M, P
LC
ES
S
B
Order Lamniformes
Family Lamnidae
Isurus oxyrinchus Rafinesque, 1810
CT
EN
MA
MP
Order Carcharhiniformes
Family Triakidae
Mustelus albipinnis Castro-Aguirre, Antuna-Mendiola, González-Acosta & De la Cruz-Agüero, 2005
Ophioblennius steindachneri Jordan & Evermann, 1898 *
SD, C, M, P, G
LC
MAR
R
D
Plagiotremus azaleus (Jordan & Bollman, 1890)
SD, C, M, P, G
LC
MAR
R
D
Family Clinidae
Gibbonsia elegans (Cooper, 1864)
O, SD, C
LC
MA
S
B
Family Labrisomidae
Labrisomus multiporosus Hubb, 1953 *
SD, C, M, P, G
LC
MA
R
B
Labrisomus xanti Gill, 1860*
SD, C, M, G
LC
MA
R
B
Malacoctenus hubbsi Springer, 1959 *
SD, C
LC
MA
R
B
Malacoctenus tetranemus (Cope, 1877)
SD, C, M, P, G
LC
MA
R
B
Paraclinus asper (Jenkins & Evermann, 1889) *
SD, C
LC
MA
R
B
Paraclinus integripinnis (Smith, 1880) *
SD, C, M
LC
MA
R
B
Paraclinus magdalenae Rosenblatt & Parr, 1969
SD, E
EN
MA
R
B
Paraclinus sini Hubbs, 1952
SD, C
LC
MA
R
B
Paraclinus stephensi Rosenblatt & Parr, 1969
SD, M
LC
MA
R
B
Family Chaenopsidae
Chaenopsis alepidota (Gilbert, 1890) *
SD, C, M, P
LC
MA
S
B
Cirriemblemaria lucasana (Stephens, 1963)
SD, C, M
LC
MA
R
B
Coralliozetus micropes (Beebe & Tee-Van, 1938)
SD, C
LC
MA
R
B
Coralliozetus rosenblatti Stephens, 1963
SD, C
LC
MA
R
B
Protemblemaris bicirrus (Hildebrand, 1946)
SD, C, M, P
LC
MA
R
B
Stathmonotus sinuscalifornici (Chabanaud, 1842) *
SD, C
LC
MA
R
B
Family Gobioesocidae
Gobiesox papillifer Gilbert, 1890
SD, C, M, P
LC
MA
R
B
Rimicola eigenmanni (Gilbert, 1890)
SD
NE
MA
R
B
Tomicodon zebra (Jordan & Gilbert, 1882)
SD, C, M, P
LC
MA
R
B
Family Callionymidae
Synchiropus atrilabiatus (Garmann, 1899)
SD, C, M, P, G
LC
MA
S
D
Family Eleotridae
Gobiomorus maculatus (Günther, 1859)
SD, C, M, P, G
LC
ES
S
D
Family Gobiidae
Bathygobius ramosus Ginsburg, 1947
SD, C, M, P
LC
ES
S
B
Ctenogobius sagittula (Günther, 1861)
SD, C, M, P
LC
ES
S
B
Coryphopterus urospilus Ginsburg, 1938
SD, C, M, P, G
LC
MA
R
B
Gillichthys mirabilis (Cooper, 1864) *
O, SD, C
LC
ES
S
B
Gobulus crescentalis (Gilbert, 1892)
SD, C, M, P
LC
MA
R
B
Gymneleotris seminuda (Günther, 1864)
SD, C, M, P
LC
MA
R
B
Ilypnus gilberti (Eigenmann & Eigenmann, 1889) *
O, SD
LC
ES
S
B
Lythrypnus dalli (Gilbert, 1890) *
SD, C, M, P, G
LC
MA
R
B
Lythrypnus pulchellus Ginsburg, 1938
SD, C, M, P
LC
MA
R
B
Lythrypnus zebra (Gilbert, 1890) *
O, SD, C
LC
MA
R
B
Microgobius erectus Ginsburg, 1938
SD, C, M, P
LC
ES
S
B
Microgobius miraflorensis Gilbert & Starks, 1904
SD, C, M, P
LC
ES
S
B
Microgobius tabogensis Meek & Hildebrand, 1928
SD, C, M, P
LC
ES
S
B
Quietula y-cauda (Jenkins & Evermann, 1889) *
SD, C
LC
ES
S
B
Typlogobius californiensis Steindachner, 1879
SD
NE
MA
R
B
Family Microdesmidae
Microdesmus dorsipunctatus Dawson, 1968
SD, C, M, P
DD
ES
S
B
Family Ephippidae
Chaetodipterus zonatus (Girard, 1858) *
SD, C, M, P
LC
ES
S
D
Family Zanclidae
Zanclus cornutus (Linnaeus, 1758)
CT
NE
MA
R
D
Famila Acanthuridae
Prionotus laticlavius (Valenncienes, 1846)
SD, C, M, P
LC
MA
R
D
Family Sphyraenidae
Sphyraena argentea Girard, 1854
O, SD, C
LC
MA
S
D
Sphyraena ensis Jordan & Gilbert, 1882 *
SD, C, M, P. PC
LC
MA
S
D
Sphyraena lucasana Gill, 1863
SD, C, M, P
DD
MA
S
D
Family Scombridae
Auxis thazard (Lacepède, 1800)
CT
LC
MA
D
Euthynnus affinis (Cantor, 1849)
CT
LC
MA
D
Sarda chiliensis (Cuvier, 1832)
O, SD, C, M, P, PC
LC
MA
D
Scomber japonicus Houttuyn, 1782
CT
LC
MA
D
Scomberomorus sierra Jordan & Starks, 1895 *
SD, C, M, P
LC
MA
D
Thunnus obesus (Lowe, 1839)
CT
VU
MA
D
Famillia Stromateidae
Peprilus simillimus (Ayres, 1860)
O, SD, C
LC
MA
D
Order Pleuronectiformes
Famila Paralichthydae
Acylopsetta dendritica Gilbert, 1890
SD, C, M, P
LC
ES
S
B
Citharichthys fragilis Gilbert, 1890
SD, C
LC
ES
S
B
Citharichthys gilberti Jenkins & Evermann, 1889
SD, C, M, P
LC
ES
S
B
Citharichthys sordidus (Girard, 185)
O, SD
LC
ES
S
B
Citharichthys stigmaeus Jordan & Gilbert, 1882
O, SD
LC
ES
S
B
Citharichthys xanthostigma Gilbert, 1890
SD
LC
ES
S
B
Cyclopsetta panamensis (Steindachner, 1876)
SD, C, M, P
LC
ES
S
B
Etropus crossotus Jordan & Gilbert, 1882 *
SD, C, M, P
LC
ES
S
B
Etropus peruvianus Hildebrand, 1946
SD, C, M, P
LC
ES
S
B
Hippoglossina bollmani Gilbert, 1890
SD, C, M, P, G
LC
ES
S
B
Hippoglossina tetrophthalma (Gilbert, 1890)
SD, C, M, P
LC
MA
S
B
Paralichthys aestuarius Gilbert & Scofield, 1898
SD, C, M
DD
ES
S
B
Paralichthys californicus (Ayres, 1859) *
SD, C
LC
ES
S
B
Paralichthys woolmani Jordan & Williams, 1897
SD, C, M, P, G
DD
ES
S
B
Syacium latifrons (Jordan & Gilbert, 1882)
SD, C, M, P, G
LC
ES
S
B
Syacium ovale (Günther, 1864)
SD, C, M, P
LC
ES
S
B
Xystreurys liolepis Jordan & Gilbert, 1880 *
SD, C
LC
ES
S
D
Family Pleuronectidae
Pleuronichthys coenosus Girard, 1854
O, SD
LC
MAS
R
B
Pleuronichthys guttulatus Girard, 1854
O, SD, C
LC
ES
S
B
Pleuronichthys ocellatus Starks & Thomson, 1910
SD, C
LC
ES
S
B
Pleuronichthys ritteri Starks & Morris, 1907
O, SD
LC
ES
S
B
Pleuronichthys verticalis Jordan & Gilbert, 1880
O, SD, C
LC
ES
S
B
Family Achiridae
Achirus mazatlanus *
SD, C, M, P
LC
ES
S
B
Family Bothidade
Bothus constellatus (Jordan, 1889)
AA
LC
ES
S
B
Bothus leopardinus (Günther, 1862)
SD, C, M, P
LC
ES
S
B
Monolene dubiosa Garman, 1899
SD, M, P
LC
ES
S
B
Family Cynoglossidae
Symphurus atramentatus Jorsan & Bollman, 1890
SD, C, M, P
LC
ES
S
B
Symphurus atricaudus (Jordan & Gilbert, 1880)
SD
LC
ES
S
B
Symphurus fasciolaris Gilbert, 1892
SD, C, M, P
LC
ES
S
B
Symphurus williamsi Jordan & Culver, 1895
SD, C, M, P
LC
ES
S
B
Order Tetraodonfiformes
Family Balistidae
Balistes polylepis Steindachner, 1876 *
SD, C, M, P
LC
MA
R
D
Pseudobalistes naufragium (Jordan & Starks, 1895)
SD, C, M, P, G
LC
MA
R
D
Suflamen verres (Gilbert & Starks, 1904)
SD, C, M, P, G
LC
MA
R
D
Family Monacanthidae
Aluterus monoceros (Linnaeus, 1758)
CT
NE
MA
S, R
MP
Family Tetraodontidae
Sphoeroides annulatus (Jenyns, 1842)
SD, C, M, P
LC
ES
S
D
Sphoeroides angusticeps (Jenyns, 1842)
SD, C, M, P, G
LC
ES
S
D
Sphoeroides lispus Walker, 1996
SD, C
LC
ES
S
D
Sphoeroides lobatus (Steindachner, 1870) *
SD, C, M, P, G
LC
ES
S
D
Sphoeroides sechurae Hildebrand, 1946
SD, C, M, P
LC
ES
S
D
Family Diodontidae
Chilomycterus reticulatus (Linnaeus, 1758)
CT
LC
ES
S
D
Diodon holocanthus Linnaeus, 1758
CT
LC
ES
S
D
Diodon hystrix Linnaeus, 1758
CT
LC
ES
S
D
Ecologically, 56.1% (233 species) are marine-euryhaline, and 42.6% (177 species) of marine-stenohaline derivation. Most of the fish species inhabit soft (235 species = 57%) or rocky (114 species = 27.5%) bottoms. Based on its distribution in the water column,14.4% (60 species) are pelagic, 25% (102 species) benthic and 60% (248 species) demersal; likewise, 262 species (63.1%) were classified as neritic, 55 (13.3%) as epipelagic, 26 (13.2%) as mesopelagic, 109 (26.3%) as benthopelagic, and 7 (1.7%) as bathypelagic.
Figure 2. Taxa (families, genera, and species) reported here for the BMA lagoon system.
Discussion
The BMA lagoon system is in a temperate-tropical transition zone where abrupt faunal changes occur (Castro-Aguirre et al., 1992, 1993; Hubbs, 1960; Ruiz-Campos et al., 2010), promoting the existence of a great variety of fish species ranging between 62 (Rodríguez-Romero et al., 2012) to 302 (Galván-Magaña et al., 2000), contrasting with the 416 fish species reported in this study (Table 1, Fig. 2). The great fish diversity found in this marine-estuarine ecosystem results from the coexistence of 3 faunal assemblages of temperate and tropical-subtropical (eurythermal species) derivation, which settled in this area by ancient invasive and colonization migrations (north-south and vice versa) related to geological and oceanographic episodes occurred between Miocene and Pliocene, during the evolution of the BCP and the Gulf of California (Bennett et al., 2015; Castro-Aguirre & Torres-Orozco, 1993; Ferrari et al., 2013).
Differences between the fish species richness previously reported and the one reported here (Fig. 3), result from the increase of ichthyological studies carried out in the area as well as the addition of updated literature records that include range expansion of some fish species like the Clarion angelfish H. clarionensis (Pomacanthidae) and 2 members of the family (Pomacentridae): the Acapulco damselfish Stegastes acapulcoensis (Fowler, 1944) and the Whitetail damselfish Stegastes leucorus (Gilbert, 1892). These tropical fish species reach their northernmost distribution ranges in temperate environments of the northeastern Pacific, due to the warm water intrusion into the temperate region provoked by positive temperature anomalies occurred during 2014 (Hernández-Velasco et al., 2016). Likewise, it has been established as a potential cause the ability of some species to capitalize physiological (e.g., oxygen requirements) and ecological adaptations (e.g., food availability and suitable substrata), regardless of the latitudinal limit of its distribution (Hastings et al., 2020). Other fish species recently included in the checklist of the BMA lagoon system (Love et al. 2021) are the Gulf anchovy Anchoa helleri (Hubbs, 1921) which was reported as endemic of the Gulf of California by González-Acosta, Balart et al. (2018) and the Clarion damselfish, Stegastes redemptus (Heller & Snodgrass, 1903) caught at Hughes Point East of Bahía Santa María (GBIF, 2023).
The checklist reported here includes 76 fish species in the larval stage which also are represented as juveniles and/or adults and contrast with the larval richness (105 to 120 species) previously reported by Avendaño-Ibarra et al. (2004) and Funes-Rodríguez et al. (2007). However, only 12.5% (n = 9) of the fish species listed here have been reported in larval stage, the cardinal fishes (Apogonidae): Apogon atricaudus Jordan & McGregor, 1898 and Apogon retrosella (Gill, 1862); the garibaldi (Pomacanthidae): Hypsypops rubicundus (Girard, 1864); the Mexican hogfish (Labridae): Bodianus diplotaenia (Gill, 1862); the delicate tiplefin (Tripterigiidae): Enneanectes carminalis (Jordan & Gilbert, 1882); the labrisomid blennies: Labrisomus xanti Gill, 1860 and Paraclinus integripinnis (Smith, 1880); and the tube blennies (Chaenopsidae): Chaenopsis alepidota (Gilbert, 1890) and Stathmonotus sinuscalifornici (Chabanaud, 1942). Other 4 taxa not listed here have been recorded at generic level: Melamphaes Günther, 1864 (Malamphaidae), Myripristis Cuvier, 1829 (Holocentridae), Serranus Cuvier, 1816 (Serranidae), and Chromis Cuvier, 1814 (Pomacentridae), as part of the fish fauna of BMA lagoon system (e.g., Avendaño-Ibarra et al. 2004). Thus, once these taxa can be identified to the species level, the fish diversity of this ecosystem could continue to increase.
As previously reported for different lagoon systems along the Mexican Pacific basin (Castro-Aguirre et al., 2006; Gómez-González et al., 2012; González-Acosta, Balart et al., 2018; González-Acosta, Monsalvo-Flores et al., 2021; González-Acosta, Rodiles-Hernández et al., 2018), the high diversity of actinopterygian fishes (371 species) over chondrichthyans (45 species) is remarkable in the BMA lagoon system (Fig. 2). The limited presence of cartilaginous fish in this coastal biotope could be mainly due to the reduced size of the tidal-channel inlets, as well as the poverty of suitable habitats for large-sized sharks and rays (González-Acosta, Balart et al., 2018; González-Acosta, Rabadán-Sotelo et al., 2015; González-Acosta, Rodiles-Hernández et al., 2018; González-Acosta, Ruiz-Campos et al., 2015), in addition to its limited tolerance to wide variations in salinity as is characteristic in coastal ecosystems of arid zones (González-Acosta, Ruiz-Campos et al., 2021). However, it has been reported that along the BCP there are coastal areas that favor their presence as well as the reproduction, breeding and feeding activities of some elasmobranchs. Therefore, the low number of chondrichthyans reported for BMA lagoon system also could be consequence of the discarding of species lacking commercial value during artisanal fishing activities or due to the need to make a greater effort focused on the capture and registration of these fishes that inhabit this ecosystem, as it has been reported in other coastal lagoons of the BCP (González-Acosta, Balart et al., 2018).
Figure 3. Number of families, genera, and species from the BMA lagoon system, according to different checklist and current data. In square brackets is indicated the corresponding reference: [CA] Castro-Aguirre and Torres-Orozco (1993), [DC] De la Cruz-Agüero et al. (1994), [GM] Galván-Magaña et al. (2000), [AI] Avendaño-Ibarra et al. (2004), [FR] Funes-Rodríguez et al. (2007), [RR] Rodríguez-Romero et al. (2012), and [GA] González-Acosta, Ruiz-Campos et al. (2015).
The highest species richness in the family Carcharhinidae and the genus Carcharhinus in costal ecosystems has been related to their wide distribution in oceans worldwide (Dulvy et al., 2008; Lea & Rosenblatt, 2000). While the higher richness of rays and skate species in the family Urotrygonidae and the genus Urotrygon Gill, 1863, could be associated with their preference for shallow areas with sandy bottoms and abundant vegetation (González-Acosta, Balart et al., 2018).
Otherwise, the high diversity of ray-finned fish species (Actinopterygii) in the BMA lagoon system, is due to the great availability of suitable soft-bottom and submerged vegetation habitats (e.g., mangrove biotopes), as well as favorable hydrological conditions for a variety of actinopterygian fish species, as is common pattern along the subtropical and tropical coastal ecosystems from the northeastern and the Central Mexican Pacific regions (Gómez-González et al., 2012; González-Acosta, Balart et al., 2018; González-Acosta, Rodiles-Hernández et al., 2018; González-Acosta, Ruiz-Campos et al., 2015; Tapia-García et al., 1998). Particularly, for those fish of the order Perciformes and their representative families and species (e.g., Sciaenidae, Haemulidae, Carangidae and Gobiidae) which regularly enter to the coastal ecosystems for different activities (e.g., spawning, nursery and feeding) due to their different life cycles (González-Acosta, Balart et al., 2018; González-Acosta, Ruiz-Campos et al., 2015, 2021).
Moreover, the high proportions (> 80%) in the zoogeographic affinity of the fish fauna from BMA lagoon system, between the California (San Diego and Cortez provinces) and Tropical Eastern Pacific (Mexican and Panamian provinces) regions, corroborate the recognition of this area as a transition zone where coexist fish assemblages from temperate and tropical-subtropical derivation (Briggs, 1974; Castro-Aguirre et al., 1993; González-Acosta, Balart et al., 2018; González-Acosta, Ruiz-Campos et al., 2015; Hastings et al., 2010; Hubbs, 1960; Ruiz-Campos et al., 2010), as well as the ancient relationship between 2 of the main biogeographical regions of the Eastern Pacific (Brusca et al., 2005; Hastings, 2000; Hastings et al., 2010). Particularly, for those fish species whose distribution ranges include boreal-temperate and subtropical-tropical latitudes.
Based on the patterns of distribution of the BMA fish fauna, the status of the endemic Magdalena blenny P. magdalenae was determined. This labrisomid fish species, only known from Magdalena and Santa María bays, Margarita Island and South of Punta Redonda, BCS (GBIF, 2023; Love et al., 2021; Rosenblatt & Parr, 1969), inhabits rocky bottoms at 8 to 21m depths (Robertson & Allen, 2015); however, there is no more information on this species.
Comparatively, the BMA lagoon system hosts a smaller number of marine fish species with circumtropical distribution than other coastal environments of the BCP (González-Acosta, Balart et al., 2018), due to the greater influence of the boreal-temperate region and the California Current over the tropical biogeographic region. However, the presence of fish assemblages with circumtropical affinity such as sharks: mackerel sharks (1 species), requiem sharks (5 species) and hammerheads (3 species), and actinopterygian fishes (e.g., milkfishes, lanternfishes, jacks, dolphinfishes, parrotfishes, Morish idols, mackerels and porcupinefishes) could be since these groups can reach high latitudes during warm ocean conditions in the North Pacific provoked by El Niño and post-El Niño events, as well as their condition as free-swimming fishes (e.g., sharks and jacks), and ontogenetic adaptation (e.g., parrotfishes and Morish idols) during these warm-water events (Lea & Rosenblatt, 2000). The recent range extension records of the Clarion angelfish (Pomacanthidae) and Acapulco damselfish and the whitetail damselfish (Pomacentridae) occurred during the 2014 warm-water period in the BMA lagoon system (Hernández-Velasco et al., 2016) confirm the tendency of these tropical fish families to extend their distribution ranges beyond their northern limits in the Tropical Eastern Pacific as had been documented previously by Lea and Rosenblatt (2000) for other representative species of these fish families.
The presence (although in a lower percentage) of species with amphipacific (e.g., the Pacific spiny dogfish and the reef cornetfish) and amphiamerican (e.g., the lemon shark, the bonnethead shark, the hospe mullet and the Latin grunt) distribution, corroborates the ancient connection between the Tropical Eastern Pacific with the Indo-Pacific and the Western Atlantic biogeographic regions; may be as result of different geologic and oceanographic events which occurred during the evolution of these oceanic basins (González-Acosta, Monsalvo-Flores et al., 2021).
The conservation status (sensu IUCN, 2022) of the fish fauna of the BMA lagoon system corresponds mainly to “Least Concern” species (82%), followed by “Data Deficient” (5%), “Not Evaluated” (4.1%7) and “Near Threatened” (2.4%), which include mainly bony fishes (except for the last category that includes shark and rays). The high number of fish species here reported as “Least Concern” and “Data Deficient” is similar to those reported in different coastal systems around the BCP (González-Acosta, Balart et al., 2018; González-Acosta, Ruiz-Campos et al., 2015, 2021) and the Central Mexican Pacific (González-Acosta, Rodiles-Hernández et al., 2018); these categories do not mean low risk but indicate the lack of complementary information on the basic biology for many fish species. Contrasting with 2 hammerhead shark species (S. lewini and S. mokarran) included as “Critically Endangered” which are widely distributed in warm waters of the World Ocean (GIBF, 2023) and commonly threatened due to targeted or incidental overfishing (Dulvy et al., 2008; González-Acosta, Monsalvo-Flores et al., 2021); and the actinopterygian S. gigas distributed in the Northeastern Pacific but without any ecological or commercial interest in the area (Cornish, 2004; González-Acosta, Balart et al., 2018). Nineteen species (including sharks, rays, and bony fish species) are listed as “Vulnerable” (sensu IUCN,2022), most corresponds to elasmobranchs which were updated in their risk category status in the version 2021-1 of the IUCN Red List (González-Acosta, Monsalvo-Flores et al., 2021); whereas some bony fishes included in this risk category such as H. ingens (Syngnathidae), H. clarionensis (Pomacanthidae) and S. leucorus (Pomacentridae), are commercially exploited by ornamental fishery in Mexico, as well as potentially affected by perturbation of their preferential habitat. A similar treatment was applied to shark species listed as “Endangered”, while some bony fishes such as Mycteroperca jordani (Jenkins & Evermann, 1889) which is caught by artisanal fishery and P. magdalenae recognized as endemic of this region, are included in this risk category under such criteria.
Moreover, the Clarion angelfish (H. clarionensis) and the Pacific seahorse (H. ingens) are included as species with a degree of vulnerability by the NOM-059-SEMARNAT-2010 (DOF, 2010, 2019) under the category of “Special Protection (Pr)”, coinciding with the IUCN Red Lists (IUCN, 2022) where both fish species are listed as “Vulnerable”; however, beyond its commercial importance and limited distribution there is no complementary information that allows corroborating its population status as well as its inclusion in this risk category.
The predominance of marine (stenohaline) and estuarine (euryhaline) fish assemblages in the BMA lagoon system, able to tolerate a wide range of salinities is mainly due to the absence of rain and river runoffs that together with high evaporation rates (that increase the salinity in the water column) make of this ecosystem a negative estuary throughout the year (Álvarez-Borrego et al., 1975; Castro-Aguirre & Torres-Orozco, 1993), as is common in this arid region (Brusca et al., 2005; González-Acosta, Monsalvo-Flores et al., 2021; González-Acosta, Ruiz-Campos et al., 2015); therefore, limiting the presence of fishes from primary and secondary freshwater derivation.
The analysis of distribution by habitat (substrata), allowed to determine that the soft (57%) and rocky (27%) bottoms are the preferential habitats of most fishes of BMA lagoon system, which make use of intertidal areas (including rocky reefs) and the mangrove biotopes associated to this ecosystem as nursery, protection and feeding grounds during their life cycle (larvae, juvenile and adult), alternating seasonally their abundances and generating critical links in the food chains between the lagoon system and the adjacent sea (González-Acosta, Balart et al., 2018; González-Acosta, Ruiz-Campos et al., 2015). Thereby, this habitat contributes to the recruitment of commercial fish species, as well as for those subject to ecological conservation in this region (Avendaño-Ibarra et al., 2004; Cota-Nieto et al., 2015; Hastings & Fischer, 2001).
Ecologically, most of the fish species in the BMA lagoon system exhibit benthic-demersal (84.3%) as well as neritic (63%) habits as correspond to a shallow coastal ecosystem or due to the selectivity of the sampling method commonly used to catch the fish fauna. Also notable is the importance of the BMA lagoon system as nursery and recruitment ground for mesopelagic, benthopelagic and bathypelagic fish species (some of them in larval stages), supporting the linkage between the coastal zone with the adjacent sea. Therefore, their presence indicates the migration process that several species carry out from the open sea to the interior (and vice versa) of the BMA lagoon system influenced by tidal currents.
Because the BMA lagoon system is considered as a priority natural area subject to conservation, the updated checklist here reported, including notes on taxonomy, zoogeography, and conservation status, could contribute to improving our knowledge on the fish resources inhabiting this coastal ecosystem and for the design and implementation of effective fishing regulation strategies, and conservation programs for fish species inhabiting this coastal ecosystem.
Acknowledgements
This study was partially supported by the Instituto Politécnico Nacional (Projects SIP-IPN 20220356, 20231049); IMIPAS-CRIAP, La Paz (BCS) (Pacific Shrimp Program 20142020); and Fishery Improvement Project (FIP 7142). AFGA thanks to EDI and COFAA-IPN Programs. AFGA, CHRQ, and GRC thank the support of SNI Conahcyt Program. Karen Link´s Editorial Services edited the English manuscript. We dedicate this contribution to the memory of JFL, who passed away during the editorial process.
References
Acosta-Velázquez, J., & Ruiz-Luna, A. (2007). Variación en la cobertura, distribución y estructura de los manglares del complejo lagunar Bahía Magdalena-Bahía Almejas (1990-2005). In R. Funes-Rodríguez, J. Gómez-Gutiérrez, & R. Palomares-García (Eds.), Estudios ecológicos en Bahía Magdalena (pp. 127–141). México D.F.: CICIMAR-IPN.
Álvarez-Borrego, S., Galindo-Beet, A., & Chee-Barragan, C. (1975). Características hidroquímicas de Bahía Magdalena, B.C.S. Ciencias Marinas, 2, 94–109. https://doi.org/10.7773/cm.v.212.285
Allen, G. R., & Robertson, D. R. (1994). Fishes of the tropical eastern Pacific. Bathurst, Hawaii: Crawford House Press.
Avendaño-Ibarra, R., Funes-Rodríguez, R., Hinojosa-Medina, A., González-Armas, R., & Aceves-Medina, G. (2004). Seasonal abundance of fish larvae in a subtropical lagoon in the west coast of the Baja California Peninsula. Estuarine Coastal and Shelf Science, 61, 125–135. https://doi.org/10.1016/j.ecss.2004.03.017
Bennett, S. E. K., Oskin, M. E., Vorsey, R. J., Irondo, A., & Kunk, M. J. (2015). Stratigraphy and structural development of the southwest Isla Tiburon marine basin: implications for latest Miocene tectonic opening and flooding of the northern Gulf of California. Geosphere, 11, 977–1007. https://doi.org/10.1130/GES01153.1
Briggs, J. C. (1974). Marine zoogeography. New York: McGraw-Hill.
Brusca, R. C., Findley, L. T., Hastings, P. A., Hendricks, M. E., Torre-Cosio, J., & van der Heiden, A. M. (2005). A. Macrofaunal diversity in the Gulf of California. In J. L. Cartron, G. Ceballos, & R. S. Felger (Eds.), Biodiversity, ecosystems, and conservation in northern Mexico. Madison, New York: Oxford University Press Inc.
Castro-Aguirre, J. L., & Torres-Orozco, R. (1993). Consideraciones acerca del origen de la ictiofauna de Bahía Magdalena-Almejas, un sistema lagunar de la costa occidental de Baja California Sur, México. Anales de la Escuela Nacional de Ciencias Biológicas, 38, 67–73.
Castro-Aguirre, J. L., & Espinosa-Pérez, H. (2006). Los peces de la familia Atherinopsidae (Teleostei: Atheriniformes) de las lagunas neutras e hipersalinas de México. Hidrobiológica, 16, 89–101.
Castro-Aguirre, J. L., Ramírez-Ortiz, J. C., & Martínez-Muñoz, M. A. (1992). Nuevos datos sobre la distribución de lenguados (Pisces: Pleuronectiformes) en la costa del oeste de Baja California, México, con aspectos biológicos y zoogeográficos. Anales de la Escuela Nacional de Ciencias Biológicas, 37, 97–119.
Castro-Aguirre, J. L., Schmitter-Soto, J. J., Balart, E. F., & Torres-Orozco, R. (1993). Sobre la distribución geográfica de algunos peces bentónicos de la costa oeste de Baja California Sur, México, con consideraciones ecológicas y evolutivas. Anales de la Escuela Nacional de Ciencias Biológicas, 38, 75–102.
Castro-Aguirre, J. L., Espinosa-Pérez, H., & Schmitter-Soto, J. J. (1999). Ictiofauna estuarino-lagunar y vicaria de México. México D.F.: Limusa-Noriega.
Castro-Aguirre, J. L., González-Acosta, A. F., & De la Cruz-Agüero, J. (2005). Lista anotada de las especies ícticas anfipacíficas, de afinidad boreal, endémicas y anfipeninsulares del Golfo de California, México. Revista Universidad y Ciencia, 21, 85–106. https://doi.org/10.19136/era.a21n42.335
Castro-Aguirre, J. L., González-Acosta, A. F., De la Cruz-Agüero, J., & Moncayo-Estrada, R. (2006). Ictiofauna marina-costera del Pacífico central mexicano: análisis preliminar de su riqueza y relaciones biogeográficas. In M. C. Jiménez-Quiroz, & E. Espino-Barr (Eds.), Los recursos pesqueros y acuícolas de Jalisco, Colima y Michoacán. Colima: Instituto Nacional de la Pesca, SAGARPA.
Cornish, A. (2004). Stereolepis gigas. The IUCN Red List of Threatened Species 2004: e. T20795A9230697. Retrieved on March 7th, 2017, from: http://www.iucnredlist.org/details/20795/0
Cota-Nieto, J. J., Jiménez-Esquivel, V., & Mascareñas-Osorio, I. (2015). La pesca en Bahía Magdalena Almejas: motor económico para B.C.S. DataMares, Interactive Resource. https://dx.doi.org/10.13022/MM3GW2F
De la Cruz-Agüero, J., Galván-Magaña, F., Abitia-Cárdenas, L. A., Rodríguez-Romero, J., & Gutiérrez-Sánchez, F. J. (1994). Lista sistemática de los peces marinos de Bahía Magdalena, Baja California Sur (México). Ciencias Marinas, 20, 17–31. http://dx.doi.org/10.7773/cm.v20i1.956
Diario Oficial de la Federación (DOF). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo, Secretaría del Medio Ambiente y Recursos Naturales, México, Retrieved on January 30th, 2023, from: https://www.profepa.gob.mx/innovaportal/file/435/1/NOM_059_SEMARNAT_2010.pdf
Diario Oficial de la Federación (DOF). (2919). Modificación del Anexo Normativo III, Lista de especies en riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo, publicada el 30 de diciembre de 2010. Secretaria del Medio Ambiente y Recursos Naturales, México, 2019. Retrieved on January 30th, 2023, from: https://www.dof.gob.mx/nota_detalle.php?codigo=5578808&fecha=14/11/2019#gsc.tab=0
Dulvy, N. K., Baum, J. K., Clarke, S., Compagno, L. J. V., Cortés, E., Domingo, A. et al. (2008). You can swim but you can´t hide: the global status and conservation of oceanic pelagic sharks and rays. Aquatic Conservation: Marine and Freshwater Ecosystems, 18, 459–482. https://doi.org/10.1002/aqc.975
Etnoyer, P., Canny, D., Mate, B., & Morgan, L. (2004). Persistent pelagic habitats in the Baja California to Bering Sea (B2B) ecoregion. Oceanography, 17, 90–101. https://doi.org/10.5670/oceanog.2004.71
Ferrari, L., López-Martínez, M., Orozco-Esquivel, T., Bryan, S. E., Duque-Trujillo, J., Lonsdale, P. et al. (2013). Late Oligocene to Middle Miocene rifting and synextensional magmatism in the southwestern Sierra Madre Occidental, Mexico: the beginning of the Gulf of California rift. Geosphere, 9,1–40. https://doi.org/10.1130/GES00925.1
Finkbeiner, E. M., & Basurto, X. (2015). Re-defining co-management to facilitate small-scale fisheries reform: an illustration from northwest Mexico. Marine Pollution, 51, 433–441. https://doi.org/10.1016/j.marpol.2014.10.010
Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K. E., & Niem, V. H. (1995). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Roma, Italy: FAO.
Froese, R., & Pauly, D. (Eds.). (2023). FishBase. World Wide Web electronic publication. Retrieved on January 30th, 2023, from: https://www.fishbase.org
Funes-Rodríguez, R., Leal-Espinoza, J. D., Hinojosa-Medina, A., Hernández-Rivas, M. E., & Flores-Coto, C. (2007). Composición, distribución y abundancia de larvas de peces en Bahía Magdalena. In R. Funes-Rodríguez, J. Gómez-Gutiérrez, & R. Palomares-García (Eds.), Estudios ecológicos en bahía Magdalena (pp. 205–219). México D.F.: CICIMAR-IPN.
Galván-Magaña, F., Gutiérrez-Sánchez, F., Abitia-Cárdenas, L. A., & Rodríguez-Romero, J. (2000). The distribution and affinities of the shore fishes of the Baja California Sur lagoons. In M. Manuwar, S. G. Lawrence, I. F. Manuwar, & D. E. Malle (Eds.), Aquatic ecosystems of Mexico: status and scope, ecovision World Monograph Series (pp. 383–398). Leiden, The Netherlands: Backhuys Publishers.
GBIF (Global Biodiversity Information Facility). (2023). Retrieved on January 30th, 2023 (version 02.01/2023), from: http://data.gbif.org
Gómez-González, A. E., Velázquez-Velázquez, E., Rodiles-Hernández, R., González-Díaz, A. A., González-Acosta, A. F., & Castro-Aguirre, J. L. (2012). Lista sistemática de la ictiofauna en la Reserva de la Biosfera La Encrucijada, Chiapas, México. Revista Mexicana de Biodiversidad, 83, 674–686. https://doi.org/10.7550/rmb.24468
Gómez-Valdez, J., & Vélez-Muñoz, H. S. (1982). Variaciones estacionales de la temperatura y salinidad en una región costera de la Corriente de California. Ciencias Marinas, 8, 167–178. https://doi.org/10.7773/cm.v8i2.392
González-Acosta, A. F., Balart, E. F., Ruiz-Campos, G., Espinosa-Pérez, H., Cruz-Escalona, V. H., & Hernández-López, A. (2018). Diversidad y conservación de los peces de la Bahía de La Paz. Baja California Sur, México. Revista Mexicana de Biodiversidad, 89, 705–740. https://doi.org/10.22201/ib.20078706e.2018.3.2145
González-Acosta, A. F., Monsalvo-Flores, E. A., Tovar-Ávila, J., Jiménez-Castañeda, M. F., Alejo-Plata, M. C., & De La Cruz-Agüero, G. (2021). Diversity and conservation of Chondrichthyes in the Gulf of California. Marine Biodiversity, 51, 1–16. https://doi.org/10.1007/s12526-021-01186-9
González-Acosta, A. F., Rabadán-Sotelo, J. A., Ruiz-Campos, G., Del Moral-Flores, L. F., & Borges-Souza, J. M. (2015). A systematic list of fishes from an insular mangrove ecosystem in the Gulf of California. In R. Riosmena-Rodriguez, A. F. González-Acosta, & R. Muñiz-Salazar (Eds.), The arid mangroves from Baja California Peninsula (pp. 81–92). Hauppauge, New York: Nova Science Publishers.
González-Acosta, A. F., Rodiles-Hernández, R., & González-Díaz, A. A. (2018). Checklist of the marine and estuarine fishes of Chiapas, Mexico. Marine Biodiversity, 48, 1439–1454. https://doi.org/10.1007/s12526-016-0630-y
González-Acosta, A. F., Ruiz-Campos, G., & Balart, E. F. (2015). Composition and zoogeography of fishes in mangrove ecosystems of Baja California Sur, México. In R. Riosmena-Rodríguez, A. F. González-Acosta, & R. Muñiz-Salazar (Eds.), The arid mangroves from Baja California Peninsula (pp. 63–80). Hauppauge, New York: Nova Science Publishers.
González-Acosta, A. F., Ruiz-Campos, G., Cruz-Escalona, V. H., & Urcádiz-Cázares, F. J. (2021). Lista comentada de la ictiofauna del estuario del río Mulegé, golfo de California, México. Revista Mexicana de Biodiversidad, 92, e923616. https://doi.org/10.22201/ib.20078706e.2021.92.3616
Hastings, P. A. (2000). Biogeography of the tropical eastern Pacific: distribution and phylogeny of chaenopsid fishes. Zoological Journal of the Linnean Society, 128, 319–335. https://doi.org/10.1111/j.1096-3642.2000.tb00166.x
Hastings, R. H., & Fischer, D. W. (2001). Management priorities for Magdalena Bay, Baja California, México. Journal of Coastal Conservation, 7, 193–202. https://www.jstor.org/stable/25098366
Hastings, P. A., Findley, L. T. & van der Heiden, A. M. (2010). Fishes of the Gulf of California. In R. Brusca (Ed.), The Gulf of California. Biodiversity and conservation (pp. 96–118). Tucson: University of Arizona Press.
Hastings, R. A., Rutterford, L. A., Freer, J. J., Collins, R. A., Simpson, S. D., & Genner, M. J. (2020). Climate change drives poleward increases and equatorial declines in marine species. Current Biology, 30, 1572–1577. https://doi.org/10.1016/j.cub.2020.02.043
Hernández-Velasco, A., Fernández-Rivera Melo, F. J., Melo-Merino, S. M., & Villaseñor-Derbez, J. C. (2016). Occurrence of Holacanthus clarionensis (Pomacanthidae), Stegastesleucorus, and Stegastes acapulcoensis (Pomacentridae) at Magdalena Bay, B.C.S., Mexico. Marine Biodiversity Records, 9, 49. https://doi.org/10.1186/s41200-016-0062-1
Hinojosa-Medina, A., Funes-Rodríguez, R., Aceves-Medina, G., & Gómez-Gutiérrez, J. (2007). Evolución de la investigación científica en el complejo lagunar Bahía Magdalena-Almejas. In R. Funes-Rodríguez, J. Gómez-Gutiérrez, & R. Palomares-García (Eds.), Estudios ecológicos en Bahía Magdalena (pp. 289–311). México D.F.: CICIMAR-IPN.
Hubbs, C. L. (1960). The marine vertebrates of the outer coast. Symposium: The Biogeography of Baja Californian and adjacent seas. Systematic Zoology, 9, 134–147. https://doi.org/10.2307/2411962
IUCN (International Union for Conservation of Nature). (2022). The IUCN Red List of Threatened Species, version 2022-2, 2022. Retrieved on January 30th, 2023, from: http://www.iucnredlist.org
Lea, R. N., & Rosenblatt, R. H. (2000). Observation on fishes associated with the 1997-98 El Niño off California. California Cooperative Oceanic Fisheries Investigations Reports, 41, 117–129.
Lewis, L. R., & Ebeling, P. E. (1971). Baja Sea guide. San Francisco, CA: Miller Freeman Publications.
Love, M. S., & Passarelli, J. K. (2020). Miller and Lea’s guide to the coastal marine fishes of California. Davis, CA: University of California Agriculture and Natural Resources.
Love, M. S., Bizarro, J. J., Cornthwaite, A. M., Frable, B. W., & Maslenikov, K. P. (2021). Checklist of marine and estuarine fishes from the Alaska-Yukon Border, Beaufort Sea, to Cabo San Lucas, Mexico. Zootaxa, 5053, 1–285. https://doi.org/10.11646/ZOOTAXA.5053.1.1
Lluch-Belda, D., Hernández-Rivas, M. E., & Guerrero-Caballero, R. (2000). Variabilidad de la temperatura superficial del mar en Bahía Magdalena, BCS. Oceánides, 15, 1–23.
Ojeda-Ruiz, M. A., Marín-Monroy, E. A., Hinojosa-Arango, G., Flores-Irigoyen, A., Cota-Nieto, J. J., Cavieses-Núñez, R. A. et al. (2018). Development of fisheries in Bahía Magdalena-Almejas: the need to explore new policies and management paradigms. Ocean and Coastal Management, 161, 1–10. https://doi.org/10.1016/j.ocecoaman.2018.04.014
Page, L. M., Espinosa-Pérez, H., Findley, L. T., Gilbert, C. R., Lea, R. N., Mandrak, N. E. et al. (2013). Common and scientific names of fishes from the United States, Canada, and Mexico. Bethesda, MD: American Fisheries Society.
Parrish, R., Nelson, H., & Bakun, A. (1981). A. Transport mechanisms and reproductive success of fishes in the California Current. Biological Oceanography, 1, 175–203. https://doi.org/10.1080/01965581.1981.10749438
Robertson, D. R., & Allen, G. R. (2015). Peces costeros del Pacífico Oriental tropical: sistema de información en línea. Ver. 2. Balboa, República de Panamá; Instituto Smithsonian de Investigaciones Tropicales. Retrieved on December 6th, 2022, from: https://www.biogeodb.stri.si.edu/sftep/es/pages.
Rodríguez-Romero, J., López-González, L., Galván-Magaña, F., Gutiérrez-Sánchez, F. J., López-Martínez, J., Inohuye-Riveral, R. et al. (2012). El papel ecológico de los peces en una zona de manglar de la costa occidental de Baja California Sur, México. In J. López-Martínez, & E. Morales-Bojórquez (Eds.), Efectos de la pesca de arrastre en el Golfo de California, México (pp. 93–113). Hermosillo: Centro de Investigaciones Biológicas del Noroeste, S.C./ Fundación Produce Sonora.
Rosenblatt, R. H., & Parr, T. D. (1969). The pacific species of the clinid fish genus Paraclinus. Copeia, 1, 1–20. https://doi.org/10.2307/1441691
Ruiz-Campos, G., Ramírez-Valdez, A., González-Acosta, A. F., Castro-Aguirre, J. L., González-Guzmán, S., & de la Cruz-Agüero, J. (2010). Composition, density and biogeographic affinities of the rocky intertidal fishes on the western coast of the Baja California Peninsula, Mexico. California Cooperative Oceanic Fisheries Investigations Reports, 51,210–220.
Tapia-García, M., Suárez-Núñez, C., Cerdenares-Ladrón de Guevara, G., Macuitl, M. C., & García-Abad, M. C. (1998). Composición y distribución de la ictiofauna en la laguna del Mar Muerto, Pacífico Mexicano. Revista de Biología Tropical, 46, 277–284.
Whitmore, R. C., Brusca, R. C., León-de la Luz, J. L., González-Zamorano, P., Mendoza-Salgado, R., Amador-Silva, E. S. et al. (2005). The ecological importance of mangroves in Baja California Sur: conservation implications for an endangered ecosystem. In J. L. E. Cartron, G. Ceballos, & R. S. Felger (Eds.), Biodiversity, ecosystems, and conservation in northern Mexico (pp. 298–333). Oxford, London; Oxford University Press. https://doi.org/10.22201/ib.20078706e.2018.3.2145
Martha Gual-Díaz a, María de Lourdes Rico-Arce a, b
a Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Av. Liga Periférico-Insurgentes Sur 4903, Col. Parques del Pedregal, Tlalpan, 14010 Ciudad de México, México
b Royal Botanic Gardens, Kew, Herbarium, Aceletated Taxonomy, HRA, Richmond, Surrey, TW9 3AE, Londres, Reino Unido
La subfamilia Grewioideae (Malvaceae) en México está representada por 6 géneros: Apeiba, Corchorus, Heliocarpus, Luehea, Trichospermum y Triumfetta; este último es de los más diversos en la subfamilia en los trópicos de América (ca. 50 especies). En México Triumfetta comprende 35 especies, de las cuales 18 se han registrado para Guerrero. Se estudió el material colectado de la subfamilia Grewioideae, en especial, el colectado en vida por Hubert Kruse; en ellos se encontraron ejemplares con características distintas a las especies ya conocidas para el género Triumfetta en México. Se describe e ilustra Triumfettanellydiegoae y se compara con el taxón más cercano a ella para Guerrero.
Palabas clave: Androginóforo; Chilpancingo de los Bravo; Endémica; Flora de Guerrero; Ginodioica; Tiliaceae
A new species of Triumfetta (Malvaceae: Grewioideae) from central Guerrero, México
Abstract
The Grewioideae subfamily (Malvaceae) in Mexico is represented by 6 genera: Apeiba, Corchorus, Heliocarpus, Luehea, Trichospermum, and Triumfetta; the latter, is one of the most diverse in this subfamily in the American tropics (ca. 50 species). In Mexico Triumfetta comprises 35 species, of which 18 have been registered for Guerrero. Within the collected material of the subfamily Grewioideae, especially, that collected by the late Hubert Kruse, specimens with characteristics other than the species already known for the genus Triumfetta in Mexico were found. Triumfetta nellydiegoae is described, illustrated and compared with closest taxon in Guerrero.
Keywords: Androgynophorus; Chilpancingo de los Bravo; Endemic; Flora of Guerrero; Gynodioecious; Tiliaceae
Introducción
Triumfetta es un género pantropical con cerca de 150 especies distribuidas en África, Australia y América tropical (Cronquist, 1981; Fryxell, 1998, 2001; Lay, 1950). En el Neotrópico se registran aproximadamente 50 nombres aceptados (Meijer, 2001), de los cuales 76% ha sido reportado en México (Fryxell, 1998; Gual-Díaz, 2011; Gual-Díaz y Diego, 2018; Jiménez-Ramírez et al., 2016).
Se han registrado 18 especies para Guerrero, 4 endémicas de la entidad, o con algún estado aledaño: Triumfetta acahuizotlanensis Gonz.-Martínez, J. Jiménez Ram. et Rios-Carr. (endémica del estado); T. heliocarpoides Bullock y T. guerrerensis Gual, Peralta et N. Diego (compartidas con Oaxaca) y T. mexiae C.V. Morton et Lay (con Michoacán) (Gual-Díaz y Diego, 2018).
La especie aquí propuesta fue colectada por Hubert Kruse por primera vez en los años 60, en la zona centro de Guerrero, en el municipio de Chilpancingo de los Bravo. Un aspecto característico de Kruse fue su perseverancia por conseguir material bien preparado, con etiquetas completas, muy ricas en datos relativos al ambiente y a los tipos de suelos de donde provenían, lo que ayuda en parte al entendimiento de la circunscripción de especies. Sus colecciones han sido material de respaldo para el registro de especies que han traído a la luz varias especies nuevas para la ciencia (Rzedowski, 1973); un ejemplo es la especie que se describe en este trabajo.
Materiales y métodos
Se estudió el material colectado en Guerrero de las subfamilias Brownlowioideae, Grewioideae y Tilioideae (Malvaceae) (APG IV, 2016; Tiliaceae, Cronquist, 1981) depositado en los herbarios CIIDIR, ENCB, FCME, IEB, MEXU y UAGC, así como de recursos disponibles en internet como el herbario MO (TROPICOS, 2023), Global Plants (JSTOR, 2023), NY Virtual Herbarium y toda la información publicada (revisiones del grupo, estudios florísticos y de vegetación, principalmente). Se hiceron disecciones cuidadosas, no destructivas de los materiales, cuando éste lo permitió. Estas se observaron al microscopio óptico como parte del método comparativo anatómico. Se elaboró una clave dicotómica para las especies taxonómicamente más relacionadas con la que se propone como especie nueva para la ciencia con base en una matriz de caracteres mofológicos de las especies distribuidas en México.
Para estimar el estado de conservación de la especie se utilizó la herramienta GeoCat (Bachman et al., 2011), propuesta por los Reales Jardines Botánicos de Kew con la que se calculó la extensión de presencia de la especie EOO (por sus siglas en inglés: extention of occupance) y el área que ocupa AOO (por sus siglas en inglés: area of occupance). Con base en estos valores, el tipo de hábitat y las características de la especie, se propone su estado de conservación de acuerdo con los criterios de la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (IUCN) y de la NOM-059- SEMARNAT-2010 (Semarnat, 2018).
En el material estudiado se encontraron ejemplares colectados por H. Kruse (depositados en el herbario FCME, MEXU y MO), con características distintas a las especies ya conocidas de Triumfetta en México, incluido Guerrero; entre las que sobresalen particularmente las hojas, más anchas que largas, con 3-5 lóbulos, de forma triangular, margen aserrado y de textura papirácea (recuerdan a las presentes en los géneros Liquidambar L. y Heliocarpus L.), así como el fruto globoso, donde el tamaño de sus pocas espinas y cuerpo son casi proporcionales (tamaño), conjunto de características no registradas en las especies de Guerrero y México (Gual-Díaz y Diego, 2018; Jiménez et al., 2016).
Los materiales de Kruse se encontraron previamente identificados como Heliocarpus nodiflorus (Donn. Sm.) Donn. Sm., Triumfetta galeottiana Turcz. y T. semitriloba Jacq.; esta última se distribuye desde el SE de los EUA hasta Chile, el Caribe, Bahamas (Gual y Diego, 2018). Asimismo, en todo su rango geográfico presenta una gran variación morfológica, concomitantemente se le asignan 5 nombres infraespecíficos, éstos incluyen 2 no formalmente descritos; de tal manera que en las colecciones bajo el nombre de T. semitriloba, existe un sin número de ejemplares que representan este “concepto de especie” que sin duda requiere de un estudio biosistemático incluyendo, por ejemplo, número cromosómico o de su ADN. La especie que aquí se presenta se justifica por la siguiente descripción, se ilustra y se compara con las especies afines de Triumfetta.
Descripción
Triumfetta nellydiegoae Gual et L. Rico, sp. nov. Fig. 1.
Triumfettae galeottianae Turcz., arcte affinis sed habitu arboreo vel fruticoso, foliis textura papyracea, fructu globoso biloculare 3 mm diametro, bipartitus sit eorum ignominia, medius inter papillari, spinae 3.5 mm longis tecto, differens.
Arbustos o pequeños árboles, ginodioicos, 3-6.5 m de alto; tallos jóvenes blanquecinos con abundantes lenticelas, glabros; hojas con láminas 3-5 lobadas, oblongas, 6-13 cm de largo, 3.5-7.5 cm de ancho, 5-nervadas, haz y envés glabro, de consistencia papirácea, ápices caudados o largamente acuminados, margen aserrado, base truncada, los 2 dientes basales de la lámina provistas de glándulas evidentemente redondeadas; pecíolos de 1.5-7.0 cm de largo, densamente estrellado-pubescentes; inflorescencia en dicasios compuestos, axilares o terminales, de ca. 15 cm de largo, ca. 30 cm de ancho, acompañada de brácteas foliares, con características similares a las hojas, oblongo lanceoladas, 2.5-3 8 cm de largo, de ca. 2 cm de ancho; pedúnculos y pedicelos estrellado-pubescentes; dicasios compuestos, axilares, 2 o 3 en cada axila, 2-3 cm de largo; pedicelos de 2-2.5 mm de largo cuando en flor, 2.5-3 mm de largo en el fruto, con pubescencia estrellada (densa o esparcida, glabrescente); flor hermafrodita no vista, flor femenina de ca. 1 cm de diámetro; cáliz con 4 sépalos obovados, verdes, 3-4 mm de largo, interna y externamente glabros, ápice acuminado, con apéndice de ca. 1 mm de largo; corola con 4(-5) pétalos espatulados, blancos o de color amarillo-claro, 2-2.5 mm de largo, de ca. 0.5 mm de ancho, ápice redondeado o agudo, en algunos es invaginado asimétricamente; androginóforo de 0.5-1.0 mm de largo, con 4 glándulas oblongas, blanquecinas, anillo de menos de 0.5 mm de largo, ciliado; estaminodios 30-40, filamentos filiformes, de 2-3 mm de largo; ovario 2-locular, globoso, 0.5-1.0 mm de largo, muricado-hialino, estilo rollizo, 1-1.5 mm de largo, estigma bipartido (cada lóbulo 3-lobado); cápsula esférica (globosa), equinada, cuerpo de ca. 3 mm de diámetro, con escasa pubescencia simple, casi glabra con 30-45 espinas, ca. 3.5 mm de largo, espínulas uncinadas (hialinas). Semillas piriformes, café claro, 1.5-2 mm de largo, 1-1.5 mm de ancho, 1 por lóculo (con un lóculo abortivo regularmente).
Resumen taxonómico
Paratipos: México, Guerrero, municipio de Chilpancingo de los Bravo, Acauizotla, A. Reyna 140 (FCME). Rincón de la Vía, K. Kruse 855 (FCME, MEXU). Rincón Viejo, 17°17’35” N, 99°29’10” O (17.2930555, -99.4861111), H. Kruse 777b (FCME). Soyatepec-Ocotito, C. Verduzco 458 (FCME). Soyatepec-Toro, L. C. Rodríguez 300 (FCME. Municipio Mochitlán, Agua de Obispo, puente El Mosco, 17°18’50” N, 99°28’10” O, H. Kruse 1455 (FCME, MEXU) y 17°19’10” N, 99°28’06” O (17.3194445, -99.4683334), H. Kruse1456 (FCME, MO).
Distribución y hábitat:Triumfetta nellydiegoae está restringida a la zona centro de Guerrero, solo se le conoce de los municipios de Chilpancingo de los Bravo y Mochitlán. Se le localiza en el bosque tropical subcaducifolio y de galería; asociada principalmente a Inga Mill. y Miconia Ruiz et Pav. o bien a Bursera copallifera (DC.) Bullock, en sitios con exposición este; en suelos arcillosos-arenosos, con pH = 6 y en altitudes que van de 700 a 1,450 m.
Fenología: se ha encontrado en floración en octubre y en fructificación en noviembre.
Etimología: el epíteto específico se dedica con mucho respeto y admiración a la Dra. Nelly Diego Pérez, por su sobresaliente labor docente, y de investigación llevada a cabo por consolidar la Flora del estado de Guerrero a través de más de 28 años.
Nombre común: no se han registrado usos o nombres comunes en la zona hasta el momento.
Estado de conservación: considerando que el área de distribución de Triumfetta nellydiegoae es menor de 10,000 km2, se considera como una especie microendémica. Su escasez, rareza en la naturaleza (muy poco colectada, en 2 municipios) y la tendencia al cambio de uso de suelo donde habita, la clasificarían como amenazada (A), de acuerdo con la categoría de riesgo establecidas en la NOM-059-SEMARNAT-2010 (Semarnat, 2010). Considerando los criterios de la IUCN, primero se obtuvo un EOO de 4,886 km2 y un AOO de 12,000 km2, considerando ésto junto con las amenazas a las que se enfrenta la especie en su habitat como son: disminución inferida de su extensión de presencia (i), porque la especie no se ha vuelto a encontrar recientemente, la calidad de su hábitat que está reduciéndose por el cambio de uso de suelo, los asentamientos humanos que continúan alterando la vegetación original y registrada solo en 3 localidades, se propone como especie vulnerable (VU), usando el criterio B1 ab (i,ii,iii) + B2 ab (i,ii,iii,iv).
Comentarios taxonómicos
De las 18 especies del género Triumfetta registradas en Guerrero, 50% se distribuyen en bosques templados: de galería, de Quercus, mesófilo de montaña, de Pinus, y de Pinus–Quercus (Gual-Díaz y Diego, 2018). Cerca de 28% se encuentra exclusivamente en bosques tropicales secos (tropical caducifolio y tropical subcaducifolio); el resto de las especies tiene una distribución de amplio espectro en los diversos bosques templados, tropicales, así como en vegetación derivada de estos bosques.
Triumfetta nellydiegoae se ha registrado, hasta el momento, solo en el bosque tropical subcaducifolio, de igual forma que T. acahuizotlanensis, aunque existe otra agrupación de especies que habitan este tipo de vegetación y el tropical caducifolio: T. acracantha, T. bogotensis, T. lappula y T. mexiae (Gual-Díaz y Diego, 2018).
Referente a 2 identificaciones de esta especie en colectas anteriores, una como Triumfetta semitriloba, de la cual se diferencia básicamente de T. nellydiegoae por ser hermafrodita y por tener un número mayor de espinas en el fruto (50-75). La segunda, T. galeottiana se diferencia por el tamaño más grande del fruto y aspecto piriforme, se resalta que esta última es la especie más cercana a la aquí descrita (tabla 1).
De acuerdo con la organización del género por Lay (1950), la especie nueva pertenece al complejo Galeottiana, serie Uncinatae que se diagnostica por presentar frutos con espinas cinadas en su ápice (espínulas),las especies de esta serie con distribución en México son: Triumfetta acahuizotlanensis González-Martínez, J. Jiménez Ram. et Ríos-Carrasco; T. galeottiana Turcz.; T. guerrerensis Gual, S. Peralta et N. Diego; T. heliocarpoides Bullock y T. paniculata Hook. et Arn. De las antes mencionadas, la más cercana es T. galeottiana, que sin duda difiere en un número importante de caracteres taxonómicos que se presentan en la siguiente clave sinóptica dicotómica y en la tabla 1.
Figura 1. Triumfetta nellydiegoae Gual et L. Rico. a, Rama con infrutescencia; b, pubescencia del pecíolo; c, rama con inflorescencia femenina, mostrando las brácteas foliares; d, flor femenina completa mostrando en la parte central los estaminodios; e, flor femenina desprovista de pétalos y sépalos mostrando estaminodios, estilo, estigma y fruto inmaduro; f, pubescencia del pedicelo; g, estigma bifurcado, 3-lobado; h, espina/espínula del fruto; i, estaminodio; j, pétalos de diferente tamaño, con diferentes formas de ápice; k, envés de un sépalo; l, fruto; m, corte transversal del fruto, mostrando 2 lóculos. Ilustración: Albino Luna, basada en: H. Kruse 1456 (b-k) y 855 (a, l-m) (FCME).
Tabla 1
Comparación Triumfetta nellydiegoae con su especie más cercana.
Característica
T. nellydiegoae Gual et L. Rico
T. galeottiana Turcz
Forma biológica
Arbusto o árbol pequeño
Hierba o arbusto
Sexo
Ginodioico
Ginodioico/hermafrodita
Lámina: largo (cm) ancho (cm) forma
6-13 3.5-7.5 3-lobadas, oblongas
3.5-18 2.5-11 3-lobadas, ovadas
Consistencia
Papirácea
Membranácea
Frutos cuerpo: largo/ancho mm
3 × 3
5 × 4
# lóculos
2
2
Espinas (ápice)
uncinadas
uncinadas
Núm. espinas (intervalo)
30-45
30-40
Distribución
Endémica de Guerrero
Endémica de Guerrero, Jalisco, México, Michoacán, Oaxaca, Sinaloa y Veracruz
Vegetación
Bosque tropical subcaducifolio y de galería
Bosque de Quercus-Pinus; Pinus-Quercus y de galería; así como en ecotonía de tropical caducifolio; Quercus y vegetación secundaria
Elevación
700-1,450 m snm
470-1,850 m snm
Clave sinóptica complejo Galeottiana, serie Uncinatae
1a. Hierbas o arbustos; hojas de consistencia membranáceas; fruto globoso (esférico) T. galeottiana Turcz.
1b. Arbustos o pequeños árboles; hojas de consistencia áspera, coriácea, escabrosa o suave; fruto globoso o piriforme 2
2a. Plantas hermafroditas
3a. Corteza ocasionalmente exfoliante; hojas suaves o escabrosas; flor pentámera; fruto globoso (esférico), de 7.5-8.5 mm diámetro, 5 locular, con ca. 280 espinas T. guerrerensis Gual, Peralta et Diego
3b. Corteza no exfoliante; hojas coriáceas; flor tetrámera; fruto piriforme, de 2-3 mm de diámetro, (3-) 4 locular, 22˗26 (-30) espinas T. acahuizotlanensis Gonz.-Martínez, J. Jiménez Ram. et Rios-Carr.
2b. Plantas andromonoicas o ginomonoicas 4
4a. Plantas andromonoicas; hojas ásperas; fruto de 6 mm diámetro, 2 locular, con 75 espinas T. paniculata Hook. et Arn.
4b. Plantas ginomonoicas; hojas membranáceas o papiráceas; fruto de 2-3 mm de diámetro, 2-3 locular, con 20-45 espinas 5
5a. Hojas membranáceas; fruto 2 mm diámetro y largo, 3 locular, con 20-25 espinas T. heliocarpoides Bullock
5b. Hoja papirácea; fruto 3 mm diámetro y ca. 3.5 mm de largo, 2 locular, con 30-45 espinas T. nellydiegoae Gual et L. Rico, sp. nov.
Agradecimientos
Nuestro profundo agradecimiento a Jerzy Rzedowski Rotter (fallecido) por la realización de la diagnosis en latín, así como las convenientes observaciones a la clave dicotómica que ayudaron a optimizar su manejo. A Albino Luna Sánchez por la elaboración de la detallada y cuidadosa ilustración que representa a la especie. A dos revisores y editor asociado por sus valiosas retroalimentaciones que mejoraron la claridad del texto enviado.
Referencias
APG IV. (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, 1–20. https://doi.org/10.1111/boj.12385
Bachman, S., Moat, J., Hill, A. W., de Torre, J. y Scott, B. (2011). Supporting Red List threat assessments with GeoCAT: geospatial conservation assessment tool. Zookeys, 150, 117–26. https://doi.org/10.3897/zookeys.150.2109
Cronquist, A. (1981). Tiliaceae. En A. Cronquist (Ed.), An integrated system of classification of flowering plants (pp. 350–352). New York: Columbia University Press.
Fryxell, P. A. (1998). A synopsis of the neotropical species of Triumfetta (Tiliaceae). En P. Mathew y M. Sivadasan (Eds.), Diversity and taxonomy of tropical flowering plants (pp. 167–192). Calicut, India: Mentor Books.
Fryxell, P. A. (2001). Tiliaceae. Ochnaceae to Loasaceae. Vol. 3. En W. R. Anderson (Ed.), Flora Novo–Galiciana. A descriptive account of the vascular plant of western Mexico (pp. 68–109). Ann Arbor: The University of Michigan Herbarium.
Gual-Díaz, M. (2011). Tiliaceae Juss. En A. García-Mendoza y J. A. Meave (Eds.), Diversidad florística de Oaxaca: de musgos a angiospermas (pp. 297–298). México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México/ Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Gual-Díaz, M. y Diego, P. N. (2018). Diversidad de las subfamilias Grewioideae y Tilioideae (Malvaceae) en Guerrero, México. Acta Botanica Mexicana, 122, 33–96. http://dx.doi.org/10.21829/abm122.2018.1161
Jiménez-Ramírez, J., González-Martínez, C. A. y Rios, C. S. (2016). Triumfetta acahuizotlanensis (Malvaceae), a new tree species endemic of Guerrero, Mexico. Phytotaxa, 265, 273–278. https://doi.org/10.11646/phytotaxa.265.3.8
Lay, K. K. (1950). The American species of Triumfetta L. Annals of the Missouri Botanical Garden, 37, 315–395. http://dx.doi.org/10.2307/2394513
Meijer, W. (2001). Tiliaceae. En D. W. Stevens, C. Ulloa Ulloa , A. Pool y O. M. Montiel (Eds.), Flora de Nicaragua III. Monographs in Systematic Botany from the Missouri Botanical Garden, 85, 2452–2468.
Rzedowski, J. (1973). Plantae Guerrenenses kruseanae. Ciencia XXVIII, 2, 49–56.
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (2018). Norma Oficial Mexicana NOM-059-SEMARNAT-2018, Protección ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para la inclusión, exclusión o cambio-Lista de especies en riesgo. Diario Oficial de la Federación. Ciudad de México, México.
Universidad San Francisco Xavier de Chuquisaca, Facultad Ciencias Químico Farmacéuticas y Bioquímicas, Experimental de Biología “Luis Adam Brianςon”, Calle Dalence Núm. 51, Ciudad de Sucre, Chuquisaca, Bolivia
El objetivo principal de este trabajo fue generar una lista actualizada de las especies de abejas sin aguijón (Apidae: Apinae: Meliponini) en Bolivia, abordando aspectos geográficos, taxonómicos, históricos y ecorregionales. Se realizaron ajustes en la sistemática de abejas sin aguijón y correcciones geográficas mediante un minucioso análisis de la información recopilada. Los resultados obtenidos muestran la presencia de 121 especies en Bolivia, distribuidas en 19 géneros válidos, estos datos se basan en 616 registros recopilados desde 1932 hasta 2022, se destaca que la riqueza de especies de las abejas meliponinas es notablemente alta por debajo de 1,000 m (> 90% especies). Las ecorregiones más diversas resultaron ser los Yungas y el sudoeste de la Amazonia albergando 90% de las especies. Con este trabajo, se logró generar datos actualizados de la riqueza de especies de meliponinos con información de distribución geográfica para Bolivia. La relevancia de este estudio es impulsar la consolidación de conocimientos de diversidad de abejas, con el involucramiento de instituciones científicas y la comunidad investigadores. Además, que sirva como guía para las meliponiculturas locales, para identificar y manejar especies en función de la ecorregión en la que se ubiquen.
Palabras clave: Estado de conocimiento; Meliponinos; Registros geográficos; Registros taxonómicos
Stingless bees (Apidae: Apinae: Meliponini) from Bolivia: an approach to the richness and geographical distribution
Abstract
The aim of this study was to compile an updated species inventory of stingless bees (Apidae: Apinae: Meliponini) in Bolivia, considering geographic, taxonomic, historical and ecoregional aspects. Adjustments in the systematics of stingless bees and geographic data corrections were made through exhaustive analysis of the available information. The results reveal the presence of 121 species in Bolivia, distributed in 19 valid genera, these findings are based on 616 records spanning from 1932 to 2022. Notably, the species richness of meliponinos bee’s is particularly high below 1,000 meters (> 90% species). The Yungas and southwestern Amazonia were identified as the most diverse ecoregion, with 90% of the species. This study´s significance lies in advancing the understanding of the bee diversity, and fostering collaboration among institutions and researcher community. Moreover, the gathered information aims to support meliponiculture initiatives by helping stakeholders identify and manage species within their respective ecoregion.
Keywords: Knowledge status; Meliponine bees; Geographic records; Taxonomic records
Introducción
La familia Apidae (Hymenoptera) abarca una amplia diversidad de linajes de abejas antófilas, entre los que se encuentran las abejas sin aguijón, también conocidas como meliponinos (término acuñado por Moure (1963)). Desde el punto de vista taxonómico pertenecen a la tribu Meliponini, cuyo nombre deriva del griego meli (μέλι) = miel, ponos (πόνος) = trabajo y el sufijo ini = que indica o agrupa como tribu a los animales. Estas abejas juegan un papel ecológico muy importante en la polinización en ambientes naturales y agroecosistemas, lo que las hace cruciales para mantener la salud de los ecosistemas y la seguridad alimentaria (González et al., 2018; Layek et al., 2022; Slaa et al., 2006). A nivel global constituyen más de 500 especies con distribución pantropical o subtropical (Hrncir et al., 2016; Michener, 2013) y en la región neotropical se tienen registradas más de 400 especies (Camargo et al., 2013; Freitas et al., 2009; Michener, 2013), llegando a representar la mayor riqueza (~ 80%) de todas las especies de abejas sin aguijón.
Los meliponinos se encuentran distribuidos en áreas tropicales y subtropicales en América, es decir, abarcando desde el norte de México hasta el norte de Argentina (Camargo et al., 2013). La mayor riqueza de especies en América del Sur se encuentra en Brasil con un impresionante registro de 259 especies (Nogueira, 2023; Pedro, 2014). Le sigue Perú, con 175 especies documentadas (Rasmussen y Castillo, 2003; Rasmussen y Delgado, 2019) y Ecuador con 132 especies (Vit et al., 2018). Colombia con una notable diversidad de 129 especies (Nates-Parra, 2001; Nates-Parra y Rosso-Londoño, 2013), mientras que Venezuela reporta 83 especies (Pedro y Camargo, 2013). La Guyana Francesa ha registrados 80 especies (Pauly et al., 2013) y Argentina está experimentado avances significativos en sus inventarios pasando de 33 a 37 especies catalogadas (Álvarez, 2015; Lucia y Álvarez, 2023). Este panorama subraya la importancia de la investigación continua para comprender y preservar la rica diversidad de las abejas sin aguijón en la región neotropical.
Los primeros registros de especies de abejas sin agujón para Bolivia, datan de los trabajos pioneros de Cockerell (1919), Schwarz (1932, 1948), Moure (1950), Wille (1960) y Kempf-Mercado (1962, 1968). Sin embargo, uno de los hitos más significativos en la recopilación del conocimiento de abejas sin aguijón en la región neotropical, es la publicación del catálogo de abejas de Moure et al. (2007). Este catálogo es un referente y revela que para Bolivia, la riqueza de especies de abejas sin aguijón está compuesta por 21 géneros y 88 especies (Camargo y Pedro 2007; Camargo et al., 2013). Asimismo, actualmente se disponen de otras plataformas de libre acceso, que ofrecen registros de abejas para Bolivia. Entre éstas, se destacan World Bee Diversity (Ascher y Pickering, 2020), la Asociación Brasileira de Estudios de las Abejas (A.B.E.L.H.A., 2016) y el Global Biodiversity Information Facility (GBIF, 2023). Estas plataformas son una fuente valiosa para complementar y actualizar la información sobre la diversidad de abejas sin aguijón en el contexto boliviano.
El conocimiento de la riqueza de especies también se benefició de iniciativas en el ámbito de la meliponicultura, con diversos proyectos que se gestaron a partir de la década de los 90 (Aguilera, 2019). Estos proyectos incluyen iniciativas como las llevadas a cabo en el departamento del Beni, específicamente dentro de comunidades indígenas Sirionó de Ibiato (Montaño, 1996). Asimismo, en Santa Cruz se implementaron proyectos a partir del 2000 en comunidades indígenas guaraní y quechuas, como se documenta en los trabajos de Martínez y Cuéllar (2004), Ferrufino y Aguilera (2006) y Aguilera (2019). Otras experiencias valiosas se dieron en las comunidades indígenas de Tacana en La Paz (Alipaz et al., 2006), durante el 2010 en Tarija, en regiones del Chaco (Clemente y Lahore, 2010), en Cochabamba las comunidades indígenas yuracares (GADC/ALD, 2014) y en Chuquisaca, a partir de 2015, con la formación de asociaciones de meliponicultoras(es) (Campos y Peducassé, 2017; Delgado y Martínez, 2021; PASOS, 2015). Actualmente, se observan esfuerzos recientes en el área protegida municipal Ibare-Mamoré de Trinidad, Beni (Paredes et al., 2022). Estos proyectos y experiencias en distintas regiones de Bolivia han sido fundamentales para avanzar en nuestro conocimiento de la diversidad y distribución de las abejas sin aguijón en el país.
El presente estudio se centra en la recopilación y sistematización exhaustiva de todas las evidencias procedentes de diferentes fuentes (publicadas y bases de datos online), para las especies de abejas sin aguijón en Bolivia. Con el objetivo principal de generar una lista actualizada de las especies, abordando aspectos geográficos, taxonómicos, históricos y ecorregionales. La relevancia de este estudio no solo radica en la presentación de datos actualizados, sino también en la utilidad práctica de la información, como impulsar la consolidación de conocimientos de diversidad de abejas, con el involucramiento de instituciones científicas, el estado y la comunidad de investigadores. Además, que sirva como guía para las meliponiculturas locales, permitiendo identificar y manejar especies en función de la ecorregión en la que se ubiquen y fomentar prácticas sostenibles en el manejo de abejas sin aguijón.
Materiales y métodos
El proceso de recopilación y sistematización de toda la información disponible consistió en generar una lista preliminar como base, información obtenida desde el catálogo de abejas en la región neotropical (Moure et al., 2007) y de la versión online del catálogo de abejas Moure (Melo et al., 2022). Esta información se comparó con bases de datos disponibles en la web, como el sistema de información científica sobre las abejas neotropicales del Brasil conocido como info A.B.E.L.H.A (A.B.E.L.H.A., 2016), Global Biodiversity Information Facility (GBIF, 2022) y la lista de diversidad de abejas del mundo (World Bee Diversity; Ascher y Pickering, 2020). Por otra parte, se ajustó la clasificación sistemática con base en la propuesta reciente realizada por Engel et al. (2023).
Posteriormente, se recopilaron los datos de distribución y taxonomía de diferentes publicaciones especializadas (artículos y libros) desde 1832 hasta 2022, éstas fueron sistematizadas en el programa Mendeley (ver. 1.19.8), de ellas se extrajo información como localidad, fecha de colecta y en algunos casos datos geográficos (fig. 1). Se revisó el origen geográfico de los registros (datos geográficos y la descripción de la localidad), de los cuales se determinó la calidad de la información. A aquellos registros que no tenían datos geográficos, se les aplicó el método de georreferenciación (fig. 1), con la descripción de localidad y mediante el uso de capas vectoriales de Bolivia, como son: centros poblados del censo poblacional de 2001 y 2012 (INE, 2016), los límites municipales (MA, 2015), en algunos casos, la red drenaje (ríos) de Bolivia y mapas topográficos del Instituto Geográfico Militar a escala de 1:50000 (IGM, 2021). Este proceso se realizó mediante el programa QGIS (ver. 3.18) (QGIS, 2020), aplicando un complemento adicional como el plugin Lat Lon Tools (Hamilton, 2021), para obtener datos geográficos. Para verificar especialmente las localidades georreferenciadas y altura, se utilizó el programa Google Earth Pro (ver. 7.3.6.9345) (Google, 2022).
Figura 1. Proceso de trabajo en la generación de la lista de especies de abejas sin aguijón para Bolivia.
Para los datos disponibles de GBIF, se procedió a la descarga en formato Darwin Core y en formato vectorial usando el plugin GBIF Occurrences (GBIF, 2022) para el programa QGIS. Posteriormente, se revisaron la calidad de los datos geográficos y taxonómicos, de la misma manera según las ambigüedades o inconsistencia encontradas, se procedió a corregir como anteriormente se mencionó en el procedimiento de georreferenciación. Además, se incorporaron datos adicionales como las ecorregiones de Bolivia (Ibisch et al., 2003), a través de sobreposición de la capa vectorial de los registros geográficos con las ecorregiones.
Por otra parte, se adicionó información obtenida en recolectas de campo, a través de diferentes proyectos ejecutados en los departamentos de Cochabamba y Chuquisaca (fig. 1), como el material que está resguardado en la colección biológica científica del Instituto Experimental de Biología “Luis Adam Briançon” – USFX (CB-IEBUSFX). Estos proyectos fueron realizados en el ANMI El Palmar sobre la identificación, caracterización de abejas nativas con potencial melífero para 6 comunidades del área protegida (PPD/PNUD, 2013), y otros proyectos patrocinados en 2019 y 2021 por la Dirección de Investigación, Ciencia y Tecnología (DICyT-USFX) sobre abejas sin aguijón, en el pueblo indígena Yuracaré (Cochabamba) y en asociaciones meliponicultoras Chaco Chuquisaqueño (Chuquisaca).
La base de datos fue estructurada en 7 componentes: taxonomía (género y especie), localidad geográfica (departamento, municipio y localidad), fecha de recolecta, nombre del colector, datos geográficos (latitud, longitud y altitud), indicación de si se realizó el proceso de georreferenciación (si/no) y fuente o procedencia de la información (base de datos online, publicación o fuente propia). Esta información fue organizada para ser consultada y visualizada de manera interactiva (gráficos y mapas) en la aplicación de shiny (ver 1.8.0) desarrollado a través de script simple en el programa R (Chang et al., 2023), disponible en el siguiente enlace: https://scotigera.shinyapps.io/BolAbejas/.
A partir de los datos obtenidos, se realizó el análisis exploratorio elaborando tablas de contingencia y gráficos descriptivos con el programa R (ver 4.2.3) para mostrar la eventualidad de los registros de abejas sin aguijón y el número de registros en función de la altura sobre el nivel del mar. Se realizaron cuadros que muestran: 1) la lista de especies de la tribu Meliponini para Bolivia, consensuada con la información consultada de bases de datos e información publicada; 2) la lista de especies registradas para Bolivia, con información de presencia a nivel departamental y ecorregiones, con el intervalo de tiempo y los registros fuente.
Con los datos geográficos se elaboraron mapas para mostrar los diferentes tipos de registros obtenidos, tanto aquellos que usaron datos originales, como aquellos que fueron corregidos (georreferenciados) y de fuente propia usando el programa QGIS. Además, se realizaron análisis de densidad de muestreo (point to polygon) para mostrar la riqueza de especies a nivel municipal y/o departamental, y de la misma manera, para las ecorregiones de Bolivia, usando el programa de DIVA-GIS (ver. 7.5) (Hijmans et al., 2012).
Resultados
Se revisaron 49 publicaciones, tanto artículos científicos como libros, y se obtuvieron 347 registros geográficos. Además, se incluyen 220 registros provenientes de bases de datos digitales como GBIF, WBD y A.B.E.L.H.A. Asimismo, se cuenta con 49 registros que proceden de CB-IEBUSFX, logrando así un consolidado de 616 registros.
Con respecto de la revisión de las publicaciones, las contribuciones de Schwarz (1832, 1938, 1948) aportaron 68 registros, mientras que Cockerell (1919), contribuyó con 3 registros, Wille (1960, 1962) sumó 2 registros adicionales. Posteriormente, Camargo (1988) aportó 20 registros de especies y Camargo y Pedro, entre 2002 y 2009 generaron 35 registros (Camargo y Pedro, 2002, 2003, 2005, 2007, 2009). Además, las colaboraciones de Camargo con Moure (1994, 1997) obtuvieron 19 registros. Por otra parte, Moure (1950, 1960), añadió 19 registros y en colaboración con Kempff (1968), 30 registros. Estos son considerados como pioneros fundamentales en el establecimiento del conocimiento de las abejas sin aguijón en Bolivia. Posteriormente, las contribuciones de Smith-Pardo y Engel (2001), De Albuquerque y Camargo (2007), Nogueira (2016), Melo (2016), Rasmussen y González (2017), Ribeiro (2021) y Engel (2021, 2022), han realizado importantes aportes en sistemática y han contribuido con más de 30 registros. Los trabajos realizados en Bolivia por Tejada (2006), Clemente y Lahore (2010), Ferrufino y Vit (2013), Townsend et al. (2021) y Morón et al. (2023), también presentan contribuciones muy relevantes para nuestra comprensión actual.
Figura 2. A), Eventualidad de los registros de abejas sin aguijón desde 1984 a 2021; B, número de registros de las abejas sin aguijón en función a la altura sobre el nivel del mar.
Estos registros muestran recolectas que se remontan a 1834 por el naturalista d’Orbigny, posteriormente se observan 2 periodos de particular interés, uno entre los años 1900 y 1970, y otro entre 1980 y 2022 (fig. 2A). Durante el primer periodo, varios naturalistas y recolectores contribuyeron significativamente al conocimiento de la tribu Meliponini en Bolivia. Entre ellos, se destacan Wolfgang Priewasser (1900), Mann (1921 a 1938), Peña (1942 a 1995), Kerr (1949), Kempff. (1962-1968), Bouseman y Lussenhop (1964-1965), Ballard (1967-1968), Walz (1940 – 1955), y Zischka (1950). Para el segundo periodo, sobresalen Brooks (1988), Montaño (1992), Irwin y Parker (1999) y Francois (1999).
En cuanto a la procedencia geográfica de los 616 registros, 273 fueron obtenidos como datos originales, mientras que 272 registros se georreferenciaron con base en las descripciones de los sitios de recolecta en los manuscritos, y adicionalmente, 71 registros tuvieron correcciones en su ubicación geográfica (fig. 3A). De los registros provenientes del GBIF (n = 236), se corrigieron geográficamente 72 debido a discrepancias con la ubicación de las localidades descritas (fig. 3B).
Los datos altitudinales derivados de los registros geográficos muestran que estas abejas se encuentran desde 94 hasta 2,329 m snm (fig. 2B). En este contexto, 66.7% de las especies (n = 80) se localizan en un rango altitudinal desde 94 a 500 m, abarcando el piso ecológico basal que incluye las tierras bajas y parte del subandino. El 27.5% de las especies (n = 33) se encuentra en el piso ecológico basimontano o andino montano situado entre 500 y 1,000 m. Solo 15 especies de abejas sin aguijón han sido registradas por encima de 1,000 m, encontrándose en fondos de valles desde 1,100 hasta 2,400 m, incluyendo especies como Scaptotrigona turusiri en los Yungas, abejas del género Plebeia sp. y Tetragonisca angustula en los valles secos interandinos. Y un caso sorprendente fue el registro de una abeja del género Trigona sp., que se reportó por encima de 4,000 m (altiplano).
La sistematización de los datos posibilita obtener una aproximación de la riqueza de abejas sin aguijón en Bolivia, lo cual se traduce en la identificación de 19 géneros y 121 especies (tablas 1, 2). Es importante señalar que este número podría aumentar hasta 132 especies, siempre y cuando se realice una confirmación taxonómica y biogeográfica de 11, debido a que existen registros próximos a las fronteras de Brasil y Argentina.
En cuanto a la distribución de la riqueza de especies a nivel departamental en Bolivia, se observa que estas abejas están presentes en 7 de los 9 departamentos del país. Destacan los departamentos de La Paz (76 especies), Beni (69 especies), Cochabamba (66 especies) y Santa Cruz (64 especies) como aquellos con el mayor número de especies, seguidos de Chuquisaca (20 especies), Tarija (10 especies) y Pando (9 especies) (tabla 2, fig. 3C). Al considerar los límites de las ecorregiones de Bolivia, se ha determinado que la riqueza de especies está presente en 8 de las 13 ecorregiones, la ecorregión sudoeste de la Amazonía destaca con la mayor riqueza contabilizando 95 especies (tabla 2), seguida de los Yungas (58 especies) y el Cerrado (47 especies). Posteriormente, se encuentran las Sabanas Inundables (27 especies) y el Bosque Seco Chiquitano (25 especies), mientras que las ecorregiones con una menor riqueza incluyen el Gran Chaco (19 especies), el Bosque Tucumano – Boliviano (16 especies), los Bosques Secos Interandinos (16 especies) y el Chaco Serrano (6 especies). Estos hallazgos proporcionan una visión detallada de la diversidad y distribución de las abejas sin aguijón en diferentes regiones de Bolivia.
Discusión
Con base en la revisión bibliográfica que se realizó al trabajo de Camargo y Pedro (2007), para Bolivia se había catalogado un total de 24 géneros. Sin embargo, los cambios introducidos por la reciente publicación de Engel et al. (2023), que presenta una revisión exhaustiva de la clasificación a nivel de género y subgénero de la tribu Meliponini, ha tenido un impacto en la adaptación de la lista, reduciendo el número a 19 géneros. Un ejemplo concreto de estos ajustes se observa en el género Plectoplebeia propuesto por Melo (2016) y con otras especies descritas por Engel (2022), ahora se integran al género Plebeia, debido a la sinonimia y a la presencia de características comunes que comparten, como indican Engel et al. (2023). Asimismo, el género Schwarzula ha sido incluido como subgénero dentro de Scaura, a pesar de que Camargo y Pedro (2002), con base en atributos morfológicos ya había notado fuerte parentesco al identificar sinapomorfias legítimas. No obstante, el análisis filogenético y morfológico realizado por Nogueira (2016) lo distingue como otro género válido (Nogueira, 2023), por lo que se mantiene la discusión sobre su identidad.
Figura 3. Mapa de ubicación de registros geográficos y riqueza de especies de abejas sin aguijón en Bolivia. A), Mapa con datos corregidos, georreferenciados y originales; B) mapa con registros corregidos de GBIF; C), riqueza de especies de abejas sin aguijón obtenido según los municipios en cada departamento; D), a nivel de ecorregiones de Bolivia. Beni (BE), Chuquisaca (CH), Cochabamba (CB), La Paz (LP), Pando (PA), Santa Cruz (SC), Tarija (TA), Oruro (OR) y Potosí (PO).
Otro cambio significativo afecta a los géneros Dolichotrigona, Leurotrigona y Celetrigona, que han sido reclasificados como subgéneros de Trigonisca. Sin embargo, en el caso de Dolichotrigona, ya no se le reconoce como un género valido según Engel et al. (2023). Esto ha llevado a retomar su designación genérica como Trigona propuesto por Wille (1965) y Michener (2013), respaldada por similitudes morfológicas discutidas con Moure, quien dividió estos grupos (Wille, 1979). Plebia intermedia, nombre propuesto para Bolivia por Wille (1960) y citado en el catálogo de abejas de la región neotropical como Plectoplebeianigrifacies (Camargo y Pedro, 2007), ha experimentado un cambio a Plebeia (Plebeia) nigrifacies. Este cambio se debe a que Melo (2016) la considera una sinonimia y Engel et al. (2023) no la reconoce como especie válida. Estos ajustes en la clasificación taxonómica reflejan la dinámica evolutiva del conocimiento y la importancia de las actualizaciones regulares en la nomenclatura de las abejas sin aguijón en Bolivia.
Tabla 1
Lista de especies de abejas sin aguijón (Meliponini) consensuada a partir de los diferentes registros que se han revisado como el Catálogo de abejas de Moure – A.B.E.L.H.A. (A.B.E.L.H.A., 2016; Camargo, Pedro, et al., 2013; Camargo y Pedro, 2007), World Bee Diversity (Ascher y Pickering, 2020), GBIF, 2022, la lista de especies de abejas nativas de Lisperger (2015) (Townsend, 2016) y las publicaciones revisadas. Registros de especies que están en Bolivia por su proximidad (P). Los valores en negrita corresponden a la sumatoria total del número de especies según el género y las fuentes consultadas.
Taxones
CAM – (2007-2013)
A.B.E.L.H.A. (2022)
WBA (2020)
GBIF (2022)
Revisión Pub.
Subtotal
Total
Subtribu Meliponina Lepeletier
Infratribu Meliponitae Lepeletier
Grupos de Género Paratrigona
Género Paratrigona Schwarz, 1938
7
7
7
4
7
8
8
Paratrigona (Paratrigona) glabella Camargo et Moure, 1994
Trigonisca (Trigonisca) rondoni Camargo et Pedro, 2005P
1
1
Trigonisca (Trigonisca) sachamiski Alvarez et Lucia, 2018P
1
Trigonisca (Trigonisca) vitrifrons Albuquerque et Camargo, 2007
1
1
1
Géneros
21
23
23
20
24
19
19
Especies
88
86
86
88
113
118
133
Tabla 2
Lista de especies registradas par Bolivia, con información de presencia a nivel de los límites departamentales y ecorregiones, y el intervalo de tiempo de los registros. Departamentos de Bolivia: Beni (BE), Chuquisaca (CH), Cochabamba (CB), La Paz (LP), Pando (PA), Santa Cruz (SC) y Tarija (TA). Ecorregiones: Bosque Seco Chiquitano (BSChi), Bosque Tucumano – Boliviano (BTBo), Bosques Secos Interandinos (BSIn), Cerrado (Cer), Chaco Serrano (ChaSe), Gran Chaco (GCha), Sabanas Inundables (SaIn), Sudoeste de la Amazonía (SAmz), Yungas (Yun).
Lista de taxones
Dep
Eco
Años
Referencias
Género Paratrigona Schwarz, 1938
Paratrigona glabella Camargo y Moure, 1994
CH, CB, LP SC, TA
BSChi, BTBo, BSIn, ChaSe GCha, SAmz, Yun
1952-2020
Camargo y Moure, 1994; GBIF.org, 2022; Oliveira et al., 2020
Paratrigona guigliae Moure, 1960
LP
Yun
~1950
Camargo y Moure, 1994
Paratrigona lineata (Lepeletier, 1836)
CH, CB, SC
BSChi, Cer, BTBo, BSIn, Cer, Yun
1951-2015
Camargo y Moure, 1994; GBIF.org, 2022; Oliveira et al., 2020
Paratrigona nuda (Schwarz, 1943)
LP
SAmz, Yun
1950-1956
Camargo y Moure, 1994; GBIF.org, 2022; Oliveira et al., 2020
Paratrigona onorei Camargo y Moure, 1994
CB
Yun
1950
Camargo y Moure, 1994
Paratrigona pacifica (Schwarz, 1943)
CB, LP
SAmz, Yun
1949-2001
Camargo y Moure, 1994; GBIF.org, 2022
Paratrigona prosopiformis (Gribodo, 1893)
CB, LP, SC
SAmz, Yun
1948-2001
Camargo y Moure, 1994; GBIF.org, 2022
Paratrigona subnuda Moure, 1947
CB
SAmz
WBD, 2020
Género Nogueirapis Moure, 1953
Nogueirapis butteli (Friese, 1900)
CB
SAmz
1900
GBIF.org, 2022; Wille, 1962
Género Partamona Schwarz, 1939
Paratrigona ailyae Camargo, 1980
BE, CB
SAmz, SaIn, Yun
1948-2018
GBIF.org, 2022; Stearman et al., 2008; Townsend et al., 2021
Partamona combinata Pedro y Camargo, 2003
BE, CB, LP, SC
SAmz, SaIn, Yun
1922-1990
GBIF.org, 2022; Pedro y Camargo, 2003
Partamona epiphytophila Pedro y Camargo, 2003
BE, CB, LP, PA, SC
BSIn, Cer, SAmz, Yun
1921-2000
GBIF.org, 2022; Pedro y Camargo, 2003
Partamona mulata Moure, in Camargo, 1980
BE, SC
GCha, SILM
1952-1992
GBIF.org, 2022; Pedro y Camargo, 2003
Partamona nhambiquara Pedro y Camargo, 2003
BE
SAmz
1922
GBIF.org, 2022
Partamona subtilis Pedro y Camargo, 2003
BE
SAmz
1956
GBIF.org, 2022; Pedro y Camargo, 2003
Partamona testacea (Klug, 1807)
BE, LP
SAmz
1921-1956
GBIF.org, 2022; Pedro y Camargo, 2003
Partamona vicina Camargo, 1980
LP, SC
Cer, SAmz
1959-2018
GBIF.org, 2022; Pedro y Camargo, 2003; Townsend et al., 2021
Partamona yungarum Pedro y Camargo, 2003
LP, CB
BSIn
1948-1955
GBIF.org, 2022; Pedro y Camargo, 2003
Tabla 2. Continúa
Lista de taxones
Dep
Eco
Años
Referencias
Género Oxytrigona Cockerell, 1917
Oxytrigona flaveola (Friese, 1900)
BE, LP, SC
Cer, GCha, SAmz, Yun
1921-2020
GBIF.org, 2022; Schwarz, 1948
Oxytrigona mulfordi (Schwarz, 1948)
BE, LP, SC
SAmz
1921-1956
GBIF.org, 2022; Schwarz, 1948
Oxytrigona obscura (Friese, 1900)
BE, CB, LP PA, SC
GCha, SAmz, Yun
1921-2020
GBIF.org, 2022; Schwarz, 1948
Oxytrigona tataira (Smith, 1863)
BE, LP SC, TA
GCha, SAmz, Yun
1921-2018
Camargo, 1988; Clemente y Lahore, 2010; GBIF.org, 2022; Schwarz, 1948; Townsend et al., 2021
Género Scaptotrigona Moure, 1942
Scaptotrigona bipunctata (Lepeletier, 1836)
CH, LP
SaIn, SAmz, Yun
1964 – 2020
Camargo, 1988; Tejada, 2006; este trabajo
Scaptotrigona fimbriata Engel, 2022
BE, SC
BSChi, SAmz
1988 – 2002
Engel, 2022
Scaptotrigona tricolorata Camargo, 1988
CB
SAmz
1949
Engel, 2022; GBIF.org, 2022
Scaptotrigonapolysticta Moure, 1950
BE, CH, CB, LP, SC
BSChi, Cer, SaIn, SAmz, Yun
1949 – 2020
FCBC, 2021; Ferrufino, 2013; GBIF.org, 2022; Montaño, 1996; Moure, 1950; Tejada, 2006; Townsend et al., 2021; este trabajo
Scaptotrigonadepilis (Moure, 1942)
BE, CB, LP, SC, TA
BSChi, Cer, GCha, SaIn, SAmz, Yun
1949 – 2020
Clemente y Lahore, 2010; Ferrufino, 2013; GBIF.org, 2022; Moure, 1950; Tejada, 2006; Townsend et al., 2021; este trabajo
Scaptotrigonanuda Engel, 2022
BE
SAmz
1988
Engel, 2022
Scaptotrigona grueteri Engel, 2022
LP
Yun
1955-1992
Engel, 2022
Scaptotrigonanigrohirta Nogueira y Santos-Silva, 2022
CB
SAmz
2007
Stearman et al., 2008
Scaptotrigona postica (Latreille, 1807)
BE, SC
GCha, SaIn
1992-2002
Martínez y Cuéllar, 2004; Montaño, 1996
Scaptotrigona semiflava Engel, 2022
SC
BSChi
1951
Engel, 2022
Scaptotrigona turusiri (Janvier, 1955)
CB
Engel, 2022
Scaptotrigona xanthotricha Moure, 1950
CB, LP, SC
BSChi, SAmz, Yun
2020
Camargo, 1988; Ferrufino, 2013; este trabajo
Scaptotrigona yungasensis Engel, 2022
BE, CB, LP
SaIn, SAmz, Yun
1955-1966
Engel, 2022
Género Geotrigona Moure, 1943
Geotrigona fulvohirta (Friese, 1900)
BE, LP
Cer, SAmz
1922 -2005
Camargo y Moure, 1996; GBIF.org, 2022; Tejada, 2006
Geotrigona argentina Camargo y Moure, 1996
CH, SC, TA
Cer, BTBo, GCha
1949-2015
Camargo y Moure, 1996; Clemente y Lahore, 2010
Geotrigona fulvatra Camargo y Moure, 1996
BE, LP
Cer, SAmz
1922 -1956
Camargo y Moure, 1996; GBIF.org, 2022
Geotrigona mattogrossensis (Ducke, 1925)
SC
Cer
1951
Camargo y Moure, 1996; GBIF.org, 2022
Geotrigona tellurica Camargo y Moure, 1996
CB, LP
Cer
1921-1956
Camargo y Moure, 1996; GBIF.org, 2022
Geotrigona mombuca (Smith, 1863)
CH, SC
Cer
1949 -2015
Moure, 1950
Género Ptilotrigona Moure, 1951
Ptilotrigona lurida (Smith, 1854)
BE, LP
SaIn, Yun
1964 – 2020
GBIF.org, 2022; Montaño, 1996
Género Tetragona Lepeletier y Serville, 1828
Tetragona clavipes (Fabricius, 1804)
CB, CH, LP SC
BSChi, BTBo, Cer, SAmz
2020-2018
FCBC, 2021; Nogueira et al., 2022; Stearman et al., 2008; Townsend et al., 2021; este trabajo
Tetragona gottei (Friese, 1900)
BE, CB, LP, SC
BSChi, SaIn, SAmz, Yun
2020-2018
Camargo, 1988; FCBC, 2021; Stearman et al., 2008; Townsend et al., 2021
Tetragona truncata Moure, 1971
LP, PA
SAmz, Yun
?
A.B.E.L.H.A., 2016; Camargo, 1988
Género Trigona Jurine, 1807
Trigona chanchamayoensis Schwarz, 1948
BE, CB, LP, SC
BSChi, Cer, SaIn, SAmz, Yun
1956-2021
Camargo, 1988; FCBC, 2021; GBIF.org, 2022; Montaño, 1996; Moure, 1950; Schwarz, 1948; Townsend et al., 2021; este trabajo
Trigona pallens (Fabricius, 1798)
BE, CB, SC
Cer, SaIn, SAmz
1956-1999
GBIF.org, 2022
Trigona williana Friese, 1900
BE, CB
SAmz
1900-1964
GBIF.org, 2022; Schwarz, 1948
Trigona cilipes (Fabricius, 1804)
BE, CB, LP, SC
BSChi, Cer, SaIn, SAmz, Yun
1951-1962
Almeida, 1992; Camargo, 1988; Kempf-Mercado, 1968; Stearman et al., 2008
Trigona lacteipennis Friese, 1900
BE, LP
SAmz
1922-1976
A.B.E.L.H.A., 2016; GBIF.org, 2022
Trigonadimidiata Smith, 1854
BE, CB, LP
SAmz, Yun
1900-2020
GBIF.org, 2022; Schwarz, 1948; este trabajo
Trigonafulviventris Guérin, 1844
BE, CB
SAmz, Yun
1956-1999
GBIF.org, 2022
Trigona guianae Cockerell, 1910
BE, CB, LP, SC
BSChi, SaIn, SAmz
1921-1999
GBIF.org, 2022; Schwarz, 1948
Trigona albipennis Almeida, 1995
CB, PA
SAmz
1949-1999
A.B.E.L.H.A., 2016; GBIF.org, 2022; Ribeiro, 2021
Trigona fuscipennis Friese, 1900
BE, CB, SC
BSChi, SaIn, SAmz
1992-2020
FCBC, 2021; Montaño, 1996; Stearman et al., 2008; Townsend et al., 2021
Clemente y Lahore, 2010; GBIF.org, 2022; Moure, 1950; Schwarz, 1932
Melipona boliviana Schwarz, 1932
BE, CB, LP
Yun, SAmz
1921 – 1992
GBIF.org, 2022; Schwarz, 1932
Melipona brachychaeta Moure, 1950
BE, CB, CH LP, SC, TA
Yun, SAmz
1921 – 2020
Camargo, 1988; Clemente y Lahore, 2010; Ferrufino, 2013; GBIF.org, 2022
Melipona crinita Moure y Kerr, 1950
BE, CB, PA, SC
Yun, SAmz
1949 – 2020
Camargo, Pedro, et al., 2013; Ferrufino, 2013; GBIF.org, 2022
Melipona eburnea Friese, 1900
CB, LP, PA
Yun, SAmz
1900 -2021
Cockerell, 1919; GBIF.org, 2022; Gonzalez et al., 2021
Melipona flavolineata Friese, 1900
CB, LP
Cer, SAmz
2005 -2007
Stearman et al., 2008; Tejada, 2006
Melipona fuliginosa Lepeletier, 1836
BE, CB, LP, SC
BSIn, GCha, SaIn, Yun, SAmz
1952 – 2020
Camargo y Pedro, 2008; Cockerell, 1919; GBIF.org, 2022; Townsend et al., 2021
Melipona illota Cockerell, 1919
LP
Yun
1950?
GBIF.org, 2022
Melipona nebulosa Camargo, 1988
LP
Cer, Yun
2005
Camargo, 1988; GBIF.org, 2022; Tejada, 2006
Melipona rufescens Friese, 1900
BE, CB, LP
Yun, SAmz, SaIn
1900-1984
GBIF.org, 2022; Ramos et al., 2015
Melipona rufiventris Lepeletier, 1836
BE, CB, LP, SC, TA
Yun, SAmz, SaIn
1949-2020
Clemente y Lahore, 2010; Copa – Alvaro, 2016; FCBC, 2021; GBIF.org, 2022; Montaño, 1996; Stearman et al., 2008; Townsend et al., 2021
Melipona seminigra abunensis Cockerell, 1912
BE
SAmz
1922-1956
GBIF.org, 2022; Schwarz, 1932
Melipona titania (Gribodo, 1893)
LP
Yun
1900
A.B.E.L.H.A., 2016; GBIF.org, 2022
Género Trigonisca Moure, 1950
Trigonisca longicornis (Friese, 1903)
BE
SAmz
1921-1922
Pedro y Camargo, 2009
Trigonisca muelleri (Friese, 1900)
BE, LP
SAmz, Yun
1996-2001
GBIF.org, 2022; Kempf-Mercado, 1968
Trigonisca browni Camargo y Pedro, 2005
BE, LP
SAmz, Yun,
1964-2004
Camargo y Pedro, 2005
Trigonisca duckei (Friese, 1900)
BE
SAmz
1922
Schwarz, 1948
Trigonisca intermedia Moure, 1990
CH, SC
BSIn, Cer
2018 -2021
Townsend et al., 2021; este trabajo
Trigonisca longitarsis (Ducke, 1916)
LP
Yun
1964-2004
Camargo, 1988
Trigonisca pediculana (Fabricius, 1804)
LP, BE
Yun, SAmz
1922
Albuquerque y Camargo, 2007; Camargo, 1988
Trigonisca vitrifrons Albuquerque y Camargo, 2007
CB, SC
Cer, SAmz
2018 -2021
Townsend et al., 2021; este trabajo
Riqueza y distribución
El primer intento de catalogación de las abejas pertenecientes a la tribu Meliponini en Bolivia fue realizado por Moure (1950), identificando 16 especies procedentes del Chaco y de comunidades próximas a la frontera con Brasil. Posiblemente, ése fue el inicio de contacto con Kempf-Mercado (1968), quien informa sobre 30 especies para el departamento de Santa Cruz, con la colaboración directa del Prof. J.S. Moure. Posteriormente, en 2007, el catálogo de abejas en la región neotropical proporcionó una lista para Bolivia que consiste en 88 especies (Camargo y Pedro, 2007). Un esfuerzo adicional para sistematizar la diversidad de estas abejas fue realizado por Townsend (2016), quien recopilando información de la consultoría de Lisperguer (2015), presenta una lista de 56 especies y 14 géneros. Los resultados obtenidos revelan que la diversidad de abejas sin aguijón en Bolivia podría ser considerablemente mucho mayor, alcanzando hasta 133 especies. Este incremento se atribuye a los avances actuales en sistemática y a los recientes proyectos de manejo de abejas sin aguijón en el país.
Esta gran diversidad de abejas meliponinas ha sido registrada en 7 de los 9 departamentos de Bolivia, lo que representa un alcance de 77% del territorio. Sin embargo, un hallazgo reciente de un posible nido de Melipona baeri en comunidades de valles secos interandinos del departamento de Potosí (Marcelino Pinto, com. pers.), indica que existen áreas con vacíos de información que requieren ser evaluados en cuanto a su diversidad, especialmente en bosques secos tropicales (Bosques Interandinos). Esta situación también sucede en las ecorregiones que presentan, aproximadamente, 5% de las especies como el Gran Chaco y el Bosque Tucumano-Boliviano. En la ecorregión Bosque Tucumano-Boliviano se están logrando avances importantes a través de proyectos de fortalecimiento en meliponicultura (Delgado y Martínez, 2021) y estudios ecológicos recientes (Adler et al., 2023; Urquizo et al., 2022), los cuales están aportando nuevos datos taxonómicos y geográficos. Por otro lado, las ecorregiones Sudoeste de la Amazonía y los Yungas son las que poseen mayor riqueza de especies (90%, 108 especies) y se ubican en pisos ecológicos de pie de monte y montano (fig. 3C), coincidiendo con la localización del “hot-spot” de los Andes Tropicales de Myers et al. (2000), lo que resulta muy importante para la conservación de estas ecorregiones.
Los registros altitudinales de abejas sin aguijón muestran que la mayor diversidad está por debajo de 1,000 m (94% de las especies), mientras que son escasas las especies que existen por encima de 2,000 m. Sin embargo, se ha reportado un caso poco común de Geotrigona tellurica registrada por encima de 4,000 m en Viacha (La Paz) (Camargo y Moure, 1996; Camargo, et al., 2013), el cual debe ser validado. A pesar de ello, en Bolivia y otros países de Sudamérica existen pocas evaluaciones específicas de como varía la diversidad de abejas sin aguijón en un gradiente altitudinal. Sin embrago, es posible que esta variación responda al relieve altitudinal, microclima y la estructura de la vegetación, aspectos evaluados por Cómbita et al. (2022) en mariposas. En Costa Rica se ha llevado a cabo un estudio sobre cómo varía la diversidad de especies en un gradiente altitudinal, demostrando que la elevación puede ser un factor importante que influye en la composición, especialmente en abejas de la tribu Meliponini (Conrad et al., 2021)
Registros y sesgos taxonómicos
Es importante destacar que los datos recopilados de catálogos como el de Abejas Moure, A.B.E.L.H.A., WBD y GBIF, subestiman la riqueza de especies. Esta subestimación posiblemente se deba a la necesidad de una actualización tanto en la base de datos de registros geográficos como de su sistemática. Ademas, se evidenciaron sesgos geográficos, especialmente en GBIF donde 29.6% de los registros tuvieron que corregirse y georreferenciarse. Este aspecto ha sido observado por Rocha-Ortega et al. (2021) y García-Rosello et al. (2023), quienes coinciden en que GBIF puede presentar sesgos de tipo taxonómico, geográfico y temporal. Por tanto, se recomienda evaluar la calidad de la información antes de utilizar estos registros para mejorar la idoneidad de los mismos. Un caso especial de sesgos geográfico se observa en los registros del franciscano P. Wolfgang Priewasser, asignados geográficamente a Tarata como el lugar de recolecta en diferentes trabajos (Cockerell, 1919; GBIF, 2022; Schwarz, 1932, 1948; Wille, 1962). Sin embargo, se pudo evidenciar que en 1900 visitó la localidad de Chimoré por las misiones del Colegio Apostólico de San José de Tarata (Barrado-Manzano, 1946; Hollaus, 2010), lo que sugiere que los datos geográficos registrados en Tarata posiblemente no correspondan.
Por otro lado, hubo registros de especies que estaban citados erróneamente en Bolivia, como son: 1) Camargoia nordestina reportado por Tejada (2006) en los pueblos indígenas Tacana, ha sido descartada como presente en Bolivia, según registros publicados por Camargo (1996) y A.B.E.L.H.A. (2016), ya que su distribución está restringida al este y noreste del Brasil; 2) Melipona (Melipona) favosa (Fabricius, 1798) citado por Lisperguer (2015), que no se encuentra presente en Bolivia, ya que su distribución más frecuente es hacia al norte de Sudamérica (Colombia, Venezuela, Guyana y Surinam), según los mapas de distribución que se muestran en Pedro y Camargo (2013) y A.B.E.L.H.A. (2016).
Otras especies, que generan dudas sobre su presencia en Bolivia son Cephalotrigona zexmeniae (Cockerell, 1912), probablemente un registro erróneo de GBIF y A.B.E.L.H.A., dado que su distribución se encuentra principalmente en Centroamérica (México, Panamá, entre otros) (Ayala, 1999). Aunque este registro podría corresponder a Trigona fulviventris debido a su parecido (Engel et al., 2023). También se plantea la duda sobre el registro de Partamona cupira (Smith, 1863) reportado por GBIF, ya que esta especie se distribuye al sureste de Brasil en la catinga brasileña (Camargo y Pedro, 2003). Lo mismo ocurre con Nannotrigonatestaceicornis registrado en Argentina, Brasil y Paraguay (Rasmussen y González, 2017), se tiene reportes en La Paz (San Buenaventura) y Santa cruz (Robore), en las plataformas de GBIF y A.B.E.L.H.A., lo cual debe ser validado. Igualmente, se debe evaluar la presencia de Melipona fasciculata, cuya distribución es más hacia al noreste de Brasil, pero se ha reportado en los territorios indígenas Tacana (La Paz) por Tejada (2006) y Yuqui (Cochabamba), según Stearman et al. (2008).
Otros registros con dudas biogeográficas y taxonómicas provienen de Santa Cruz, como Plebeiamosquito (Smith, 1863), identificado por Moure (1950) en Porongo y Melipona rufiventris xantina reportado por Kempf-Mercado (1968), designado como “incertae sedis”(Melo et al., 2022). Igualmente ocurre para Lestrimelitta limao (Smith, 1863), con una distribución más restringida al centro del Brasil (Cerrado y Catinga) (Marchi y Melo, 2004) y que ha sido registrado por Montaño (1996) en pueblos indígenas Sirionó (Beni), lo que posiblemente corresponda a L. rufa o L. rufipes, según registros obtenidos del mismo lugar por Townsend et al. (2021).
La inclusión de 11 especies de abejas sin aguijón para Bolivia puede deberse a su proximidad geográfica, lo cual incrementa la diversidad de especies. Por ejemplo, Scaptotrigonajujuyensis, Plebeiamansita y Trigonisca sachamiski se encuentran próximos a la frontera con Argentina (Álvarez y Lucia, 2018; Álvarez et al., 2021; Lucia y Alvarez, 2023), por lo que pueden estar presentes en el departamento de Tarija. Otras 9 especies están registradas próximas al departamento de Pando y el noreste del Beni, como: Partamona batesi, Trigona juvenili, Melipona amazonica, M. titania, M. flavolineata, Trigonisca gracilis, T. hirsuticorni, T. mendersoni y T. rondoni, Lestrimelittaglabrata (A.B.E.L.H.A., 2016; Camargo y Pedro, 2005, 2009; Pedro y Camargo, 2003, 2009; Marchi y Melo, 2006; Ribeiro, 2021). Para el departamento de Santa Cruz, cerca del puerto Quijarro, se ha registrado la especie Nannotrigonachapada en la ciudad fronteriza de Corumba (Brasil) (A.B.E.L.H.A., 2016).
Tras una exhaustiva revisión de artículos científicos, libros y bases de datos digitales como el Catálogo de Abejas Moure, GBIF, WBD y A.B.E.L.H.A., junto con los registros obtenidos de CB-IEBUSFX en los diversos proyectos desarrollados en Cochabamba y Chuquisaca, se han recopilado un total de 616 registros. A partir de esta compilación de información, se tienen identificadas 118 especies de abejas sin aguijón en Bolivia, distribuidas en 19 géneros. Sin embargo, es importante destacar que esta cifra podría ser aún mayor con la posible adición de 15 especies descritas cerca de las fronteras con Brasil y Argentina.
La temporalidad de los registros abarca desde 1832 hasta 2022 y es digno de mención el trabajo de destacados colectores como W. M. Mann y Peña Luis, cuyos datos de campo han sido fundamentales. Asimismo, las contribuciones significativas en términos de sistemática, biología y biogeografía realizadas por autores como Schwarz, Moure, Kempff, Camargo, Pedro y Engel, han enriquecido considerablemente nuestro entendimiento sobre las abejas sin aguijón en Bolivia.
En cuanto a la distribución altitudinal, se ha observado que la mayor riqueza de especies se encuentra por debajo de 1,000 m, aunque también se han registrado algunas especies por encima de 2,000 m. Las ecorregiones de tierras bajas más diversas son los Yungas y el sudoeste de la Amazonia, que concentran 90% de las especies y abarcan los departamentos de La Paz, Beni, Cochabamba y Santa Cruz, sin embargo, del departamento de Pando se tiene muy poca información. De manera similar, se ha observado una carencia de inventarios publicados en los departamentos de Chuquisaca y Tarija, los cuales corresponden a ecorregiones de los bosques secos tropicales (Bosques Secos Interandinos, Gran Chaco y Bosque Tucumano Boliviano), donde se sugiere una subestimación de la riqueza de especies en estas áreas.
Agradecimientos
A Alexandria Saravia por la revisión y las valiosas sugerencias para mejorar el manuscrito, así como a Daniela Morón por sus valiosos aportes y comentarios sobre las especies de abejas sin aguijón del Bosque Seco Chiquitano. Este trabajo se ha beneficiado de las becas de investigación otorgadas por la Dirección de Investigación, Ciencia y Tecnología – USFX durante las gestiones 2019, 2021 y 2022, cuyo apoyo se agradece sinceramente. Además, se reconoce el respaldo brindado por la Facultad de Ciencias Químico Farmacéuticas y Bioquímicas de la Universidad San Francisco Xavier de Chuquisaca.
Referencias
A.B.E.L.H.A. (Asociación Brasileña de Estudios de las Abejas) (2016). Sistema de informação científica sobre abelhas Neotropicais, Abelhas No Brasil. Recuperado el 14 enero, 2023 de: http://abelha.cria.org.br/
Adler, M., Escóbar-Márquez, L., Solis-Soto, M. T. y Pinto, C. F. (2023). Stingless bees: uses and management by meliponiculturist women in the Chaco region of Bolivia. Journal of Ethnobiology and Ethnomedicine, BioMed Central Ltd, 19, 1–15. https://doi.org/10.1186/s13002-022-00574-0
Aguilera, F. J. (2019). Meliponicultura en Bolivia, Sociedad Boliviana Entomología, Santa Cruz de la Sierra, Bolivia. Recuperado el 13 noviembre, 2022 de: https://youtu.be/WPFi_Alf57s
Álvarez, L. J. (2015). Diversidad de las abejas nativas de la tribu Meliponini (Hymenoptera, Apidae) (Tesis doctoral). Universidad Nacional de La Plata, Buenos Aires, Argentina. https://doi.org/10.35537/10915/52025
Álvarez, L. J., Flores, F. F. y Rasmussen, C. (2021). A new species of Plebeia Schwarz (Hymenoptera: Apidae) from the Argentine yungas. Revista de La Sociedad Entomologica Argentina, 80, 35–41. http://dx.doi.org/10.25085/rsea.800206
Álvarez, L. J. y Lucia, M. (2018). Una especie nueva de Trigonisca y nuevos registros de abejas sin aguijón para la Argentina (Hymenoptera: Apidae). Caldasia, 40, 232–245. http://dx.doi.org/10.15446/caldasia.v40n2.70870
Ayala, R. (1999). Revisión de las abejas sin aguijón de México (Hymenoptera: Apidae: Meliponini). Folia Entomológica Mexicana, 106, 1–123.
Barrado-Manzano, A. (1946). Las misiones Franciscanas en Bolivia, Real Monasterio de Guadalupe. Sevilla, España: Imprenta San Antonio. Recuperado el 22 enero, 2022 de: https://www.pueblos-originarios.ucb.edu.bo/digital/10600 1558.pdf
Camargo, J. M. F. (1988). Meliponinae (Hymenoptera, Apidae) da coleção do ‘Istituto di Entomologia Agraria’, Portici, Itália. Revista Brasileira de Entomologia, 32, 351–373.
Camargo, J. M. F. y Moure, J. S. (1994). Meliponinae neotropicais: os gêneros Paratrigona Schwarz, 1938 e Aparatrigona Moure, 1951 (Hymenoptera, Apidae), Arquivos de Zoologia, 32, 33–109. https://doi.org/10.11606/issn.2176-7793.v32i2p33-109
Camargo, J. M. F. y Moure, J. S. (1996). Meliponini Neotropicais: o gênero Geotrigona Moure, 1943 (Apinae, Apidae, Hymenoptera), com especial referência à filogenia e biogeografia. Arquivos de Zoologia, 33, 95–161. https://doi.org/10.11606/issn.2176-7793.v33i2-3p95-161
Camargo, J. M. F. y Pedro, S. R. M. (2002). Uma espécie nova de Schwarzula da Amazônia (Hymenoptera, Apidae, Meliponini). Iheringia – Serie Zoologia, 92, 101–112.
Camargo, J. M. F. y Pedro, S. R. M. (2003). Meliponini Neotropicais: o gênero Partamona Schwarz, 1939 (Hymenoptera, Apidae, Apinae) – bionomia e biogeografia. Revista Brasileira de Entomologia, 47, 311–372.
Camargo, J. M. F. y Pedro, S. R. M. (2005). Meliponini Neotropicais: o gênero Dolichotrigona Moure (Hymenoptera, Apidae, Apinae). Revista Brasileira de Entomologia, 49, 69–92.
Camargo, J. M. F. y Pedro, S. R. M. (2007). Meliponini. En J. S.Moure, D. Urban y G. A. R. Melo (Eds.), Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical Region (pp. 1058). Curitiba, Brasil: Sociedade Brasileira de Entomologia.
Camargo, J. M. F. y Pedro, S. R. M. (2009). Neotropical Meliponini: the genus Celetrigona Moure (Hymenoptera: Apidae, Apinae). Zootaxa, 2155, 37–54. https://doi.org/10.11646/zootaxa.2155.1.4
Camargo, J. M. F., Pedro, S. R. M. y Melo, A. R. (2013). Meliponini Lepeletier, 1836. Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical Region – online version., Recuperado el 27 diciembre, 2022 de: http://moure.cria.org.br/catalogue?id=83018
Camargo, J. M. F., Roubik, D. W. y Pedro, S. R. M. (2013). Historical biogeography of the Meliponini (Hymenoptera, Apidae, Apinae) of the Neotropical region. EnP. Vit, S. R. M. Pedro, & D. W. Roubik (Eds.), Pot-honey: a legacy of stingless bees (pp. 19–34). New York: Springer. https://doi.org/10.1007/978-1-4614-4960-7_2
Chang, W., Cheng, J., Allaire, J., Sievert, C., Schloerke, B., Xie, Y. et al. (2023). Shiny: web application framework for R. Recuperado el 3 enero, 2024 de: https://shiny.posit.co/
Clemente, K. J. y Lahore, J. L. (2010). Manual de meliponicultura. Asociación de apicultores del Gran Chaco (ADACHACO) – Fundación PUMA, Villa Montes. Tarija.
Cockerell, T. D. A. (1919). Bees in the collection of the United States National Museum —3. Proceedings of the United States National Museum, 55, 167–221.
Cómbita, J. L., Giraldo, C. E. y Escobar, F. (2022). Environmental variation associated with topography explains butterfly diversity along a tropical elevation gradient. Biotropica, 54, 146–156. https://doi.org/10.1111/btp.13040
Conrad, K. M., Peters, V. E. y Rehan, S. M. (2021). Tropical bee species abundance differs within a narrow elevational gradient. Nature Publishing Group UK, 11, 1–12. https://doi.org/10.1038/s41598-021-02727-9
Copa-Álvaro, M. (2016). Patrones de nidificación de Trigona (Tetragonisca) angustula y Melipona rufiventris (Hymenoptera: Meliponini) en el norte de La Paz, Bolivia. Ecología Aplicada, 3, 82. https://doi.org/10.21704/rea.v3i1-2.274
Delgado, A. y Martínez, E. (2021). Guía práctica para el manejo de abejas nativas sin aguijón, Fundación PASOS -Cooperación Alemana, Sucre, Bolivia.
De Albuquerque, P. M. C. y Camargo, J. M. F. (2007). New species of Trigonisca Moure (Hymenoptera, Apidae, Apinae). Revista Brasileira de Entomologia, 51, 160–175. https://doi.org/10.1590/s0085-56262007000200005
Engel, M. S. (2021). A key to the species of Nanoplebeia, with descriptions of four new species (Hymenoptera: Apidae). Journal of Melittology, 106, 1–14. https://doi.org/10.17161/jom.i106.15735
Engel, M. S. (2022). A second species of the stingless bee genus Plectoplebeia (Hymenoptera: Apidae). Entomologist’s Monthly Magazine, 158, 79–86. https://doi.org/10.31184/M00138908.1582.4103
Engel, M. S., Rasmussen, C., Ayala, R. y de Oliveira, F. F. (2023). Stingless bee classification and biology (Hymenoptera, Apidae): a review, with an updated key to genera and subgenera. Zookeys, 1172, 239–312. https://doi.org/10.3897/zookeys.1172.104944
Ferrufino, U. y Vit, P. (2013). Pot-honey of six meliponines from Amboró National Park, Bolivia. EnP. Vit, S. R. M. Pedro y D. W. Roubik (Eds.), Pot-honey: a legacy of stingless bees (pp. 3–17). New York: Springer. https://doi.org/10.1007/978-1-4614-4960-7_29
Ferrufino, U. y Aguilera, F. J. (2006). Producción rural sostenible con abejas melíferas sin aguijón. Asociación Ecologica del Oriente – A.S.E.O., Santa Cruz de la Sierra, Bolivia.
Freitas, B. M., Imperatriz-Fonseca, V. L., Medina, L. M., Kleinert, A. M. P., Galetto, L., Nates-Parra, G. et al. (2009). Diversity, threats and conservation of native bees in the Neotropics. Apidologie, 40, 332–346. https://doi.org/10.1051/apido/2009012
GADC/ALD. (2014). Transferencia de recursos públicos-privados en especie para el proyecto: “Implementación de capacidades productivas de miel en mujeres Yuracares- Trópico”. Gobierno Autónomo Departamental de Cochabamba – Asamblea Legislativa Departamental, Cochabamba, Bolivia.
Garcia-Rosello, E., González-Dacosta, J., Guisande, C. y Lobo, J. M. (2023). GBIF falls short of providing a representative picture of the global distribution of insects. Systematic Entomology, 48, 489–497. https://doi.org/10.1111/syen.12589
GBIF.org. (2022). GBIF Occurrence download, Global Biodiversity Information Facility, Museum National D’Histoire Naturelle. Recuperado el 03 enero, 2022 de: https://doi.org/10.5852/EJT.2014.71
González, V. H., Amith, J. D. y Stein, T. J. (2018). Nesting ecology and the cultural importance of stingless bees to speakers of Yoloxóchitl Mixtec, an endangered language in Guerrero, Mexico. Apidologie, 49, 625–636. https://doi.org/10.1007/s13592-018-0590-2
Hijmans, R. J., Guarino, L. y Mathur, P. (2012). DIVA-GIS | free, simple & effective, LizardTech, Inc. – University of California, California, USA. Recuperado el 5 agosto, 2023 de: https://www.diva-gis.org/
Hrncir, M., Jarau, S. y Barth, F. G. (2016). Stingless bees (Meliponini): senses and behavior. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology, 202, 597–601. https://doi.org/10.1007/s00359-016-1117-9
Ibisch, P. L., Beck, S. G., Gerkmann, B. y Carretero, A. (2003). Ecoregiones y ecosistemas.En P. L.Ibisch y G. Mérida (Eds.), Biodiversidad: la riqueza de Bolivia, estado de conocimiento y conservación (pp. 47–88). Santa Cruz, Bolivia: Editorial FAN.
INE (Instituto Nacional de Estadística). (2016). Localización de poblaciones del Estado Plurinacional de Bolivia, GeoBolivia, La Paz, Bolivia. Recuperado el 10 diciembre, 2022 de: http://geo.gob.bo/download/?w=igm&l=POBLACIONES
Kempf-Mercado, N. (1962). Mutualism between Trigona compressa Latr. and Crematogaster stolli Forel (Hyment- opera: Apidae). Journal of the New York Entomological Society, 70, 215–217.
Kempf-Mercado, N. (1968). Contribución al conocimiento de las abejas indígenas (Meloponidae) del departamento de Santa Cruz. La Paz, Bolivia: Editorial Don Bosco.
Layek, U., Das, A. y Karmakar, P. (2022). Supplemental stingless bee pollination in Fennel (Foeniculum vulgare Mill.): an assessment of impacts on native pollinators and crop yield. Frontiers in Sustainable Food Systems, 6, 1–13. https://doi.org/10.3389/fsufs.2022.820264
Lisperguer, A. (2015). Las abejas sin aguijón en Bolivia: conocimientos, usos y su vulnerabilidad ante efectos antropicos y el cambio climático. Informe inédito de una consultoria para la Cancilleria Boliviana. La Paz. Bolivia.
Lucia, M. y Álvarez, L. J. (2023). Tribu Meliponini, Apidae (Hymenoptera: Anthophila) species from Argentina and Uruguay. Recuperado el 20 julio, 2023 de: https://biodar.unlp.edu.ar/apidae/index-es.html
MA (Ministerio de Autonomías). (2015). Límites municipales de Bolivia (339 Municipios), La Paz, Bolivia. Recuperado el 5 agosto, 2022 de: http://geo.gob.bo/geoserver/fondos/wms/
Marchi, P. y Melo, G. A. R. (2004). Notas sobre o tipo de Trigona limao Smith (Hymenoptera, Apidae, Lestrimelitta). Revista Brasileira de Entomologia, 48, 379–382. https://doi.org/10.1590/S0085-56262004000300013
Marchi, P. y Melo, G. A. R. (2006). Revisão taxonômica das espécies brasileiras de abelhas do gênero Lestrimelitta Friese (Hymenoptera, Apidae, Meliponina). Revista Brasileira de Entomologia, 50, 6–30. https://doi.org/10.1590/S0085-56262006000100002
Martínez, T. y Cuéllar, R. L. (2004). Manejo de abejas nativas señorita (Trigona tetragonisca angustula) y abeja corta pelo (Escaptutrigona posica) en tres comunidades de Isoso. Memorias VII Congreso Internacional sobre manejo de fauna silvestre en la Amazonía y América Latina. Wildlife Conservation Society, Ilheus, Brasil.
Melo, G. A. R. (2016). Plectoplebeia, a new Neotropical genus of stingless bees (Hymenoptera: Apidae). Zoologia (Curitiba), 33, 1–8. https://doi.org/10.1590/S1984-4689zool-20150153
Melo, G. A. R., Moure, J. S., Urban, D., Maia, E, Molin, A. D. y Gonçalves, R. B. (2022). “Catálogo de abelhas Moure”. Recuperado el 15 octubre, 2022 de: http://moure.cria.org.br/catalogue.
Michener, C. D. (2013). The Meliponini. EnP. Vit, S. R. M. Pedro y D. W. Roubik (Eds.), Pot-honey: a legacy of stingless bees (pp. 3–17). New York: Springer. https://doi.org/10.1007/978-1-4614-4960-7_1
Morón, D., Adler, M., & Justiniano, H. (2023). Abejas nativas de un paisaje productivo representativo de la Chiquitania: guía ilustrada. Santa Cruz de la Sierra, Bolivia: Museo de Historia Natural Noel Kempff Mercado/ Fundación para la Conservación del bosque Chiquitano.
Montaño, M. E. (1996). La explotación de miel silvestre y su imporatancia en la comunidad indígena Sirionó de Ibiato en el Beni, Bolivia(Tesis). Universidad Autónoma Grabriel René Moreno. Santa Cruz de la Sierra. Bolivia.
Moure, J. S. (1950). Notas sobre algunos Meliponine bolivianos (Hymenoptera, Apoidea). Dusenia, 1, 70–80.
Moure, J. S. (1963). Sobre a identidade dos meliponinos descritos por Spinola em 1853 (Hymenoptera, Apoidea). Anais da Academia Brasileira de Ciencias, 35, 257–269.
Moure, J. S., Urban, D. y Melo, G. A. R. (2007). Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical Region. Sociedade Brasileira de Entomologia, Curitiba, Brasil.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. y Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853–858. https://doi.org/10.1038/35002501
Nates-Parra, G. (2001). Las abejas sin aguijón (Hymenoptera: Apidae: Meliponini) de Colombia. Biota Colombiana, 2, 233–248.
Nates-Parra, G. y Rosso-Londoño, J. M. (2013). Diversity of stingless bees (Hymenoptera:Meliponini) used in Meliponiculture in Colombia. Acta Biológica Colombiana,18, 415–426.
Nogueira, D. S. (2016). Sistemática de Scaura Schwarz, 1938 (Hymenoptera: Apidae, Meliponini), com notas biológicas (Tesis de maestría). Instituto Nacional de Pesquisas da Amazônia. Manaus, Brasil.
Nogueira, D. S. (2023). Overview of stingless bees in Brazil (Hymenoptera: Apidae: Meliponini). EntomoBrasilis, 16, 1–13. https://doi.org/10.12741/ebrasilis.v16.e1041
Paredes, E., Callisaya, C., Rodriguez, J. y Tem, S. (2022). Guía básica de meliponicultura: Área Protegida Municipal Ibare Mamoré, WWF-UABJB, Trinidad, Bolivia.
PASOS. (2015). Antecedentes. Fundación Participación y Sostenibilidad. Recuperado el 12 julio, 2022 de: http://pasosbolivia.org/index.php?option=com_content&view=section&layout=blog&id=4&Itemid=53
Pauly, A., Pedro, S. R. M., Rasmussen, C. y Roubik, D. W. (2013). Stingless bees (Hymenoptera: Apoidea: Meliponini) of French Guiana. EnP. Vit, S. R. M. Pedro y D. W. Roubik (Eds.), Pot-honey: a legacy of stingless bees (pp. 87–97). New York: Springer. https://doi.org/10.1007/978-1-4614-4960-7_5
Pedro, S. R. M. (2014). The stingless bee fauna in Brazil (Hymenoptera: Apidae). Sociobiology, 61, 348–354. https://doi.org/10.13102/sociobiology.v61i4.348-354
Pedro, S. R. M. y Camargo, J. M. F. (2003). Meliponini neotropicais: o gênero Partamona Schwarz, 1939 (Hymenoptera, Apidae). Revista Brasileira de Entomologia, 47, 1–117. https://doi.org/10.1590/S0085-56262003000500001
Pedro, S. R. M. y Camargo, J. M. F. (2009). Neotropical Meliponini: the genus Leurotrigona Moure — two new species (Hymenoptera: Apidae, Apinae). Zootaxa, 44, 37–54. https://doi.org/10.11646/zootaxa.1983.1.2
Pedro, S. R. M. y Camargo, J. M. F. (2013). Stingless bees from Venezuela. EnP. Vit, S. R. M. Pedro y D. W. Roubik (Eds.), Pot-honey: a legacy of stingless bees (pp. 73–86). New York: Springer. https://doi.org/10.1007/978-1-4614-4960-7_4
PPD/PNUD (Programa de Pequeñas Donaciones / Programa de las Naciones Unidas para el Desarrollo). (2013). Evaluando la biodiversidad en el ANMI El Palmar: identificación, caracterización y opciones de manejo recursos de flora y abejas nativas con potencial melífero para seis comunidades del área protegida. GEF Small Grants Programme, Global Environment Facility (GEF) – United Nations Development Programme (UNDP). Recuperado el 29 noviembre, 2022 de: https://sgp.undp.org/spacial-itemid-projects-landing-page/spacial-itemid-project-search-results/spacial-itemid-project-detailpage.html?view=projectdetail&id=20488
QGIS (Geographic Information System). (2020). Geospatial Foundation. QGIS Association. Recuperado el 07 octubre, 2020 de: http://www.qgis.org
Rasmussen, C. y Delgado, C. (2019). Abejas sin sguijón (Apidae: Meliponini) en Loreto. Instituto de Investigaciones de la Amazonia Peruana, Iquitos, Perú.
Rasmussen, C. y Castillo, P. S. (2003). Estudio preliminar de la meliponicultura o apicultura silvestre en el Perú (Hymenoptera: Apidae, Meliponini). Revista Peruana de Entomologia, 43, 159–164.
Rasmussen, C. y González. V. H. (2017). The neotropical stingless bee genus Nannotrigona Cockerell (Hymenoptera: Apidae: Meliponini): an illustrated key, notes on the types, and designation of lectotypes. Zootaxa, 4299, 191–220. https://doi.org/10.11646/zootaxa.4299.2.2
Ribeiro, C. F. (2021). Estudo taxonômico de Trigona Jurine, 1807 (Hymenoptera: Apidae: Meliponini) na Amazônia Brasileira (Tesis de maestría). Instituto Nacional de Pesquisas da Amazônia. Manaus, Brasil.
Rocha-Ortega, M., Rodriguez, P. y Córdoba-Aguilar, A. (2021). Geographical, temporal and taxonomic biases in insect GBIF data on biodiversity and extinction. Ecological Entomology, 46, 718–728. https://doi.org/10.1111/een.13027
Schwarz, H. F. (1932). The genus Melipona the type genus of the Meliponidae or stingless bees. Bulletin of the American Museum of Natural History, 63, 231–460.
Schwarz, H. F. (1948). Stingless bees (Meliponidae) of the Western Hemisphere: Lestrimelitta and the following subgenera of Trigona : Trigona, Paratrigona, Schwarziana, Parapartamona, Cephalotrigona, Oxytrigona, Scaura, and Mourella.Biogeography and Taxonomy of Honeybees, 90, 1–546. https://doi.org/10.1086/396840
Slaa, E. J., Sánchez Chaves, L. A., Malagodi-Braga, K. S. y Hofstede F. E. (2006). Stingless bees in applied pollination: Practice and perspectives. Apidologie, 37, 293–315. https://doi.org/10.1051/apido:2006022
Stearman, A. M. L., Stierlin, E., Sigman, M. E., Roubik, D. W. y Dorrien, D. (2008). Stradivarius in the jungle: traditional knowledge and the use of ‘black beeswax’ among the Yuquí of the Bolivian Amazon. Human Ecology, 36, 149–159. https://doi.org/10.1007/s10745-007-9153-2
Tejada, R. B. (2006). Experiencias en la crianza de abejas nativas en dos comunidades Tacanas. Wildlife Conservatión Society, Murillo, Bolivia.
Townsend, W. R. (2016). Densidad y patrón de distribución de las colmenas de abejas nativas (Meliponini) en Potrerillo del Guendá, Porongo, Santa Cruz. Boletín Tesape Arandu, 6, 2–17.
Townsend, W. R., Adler, M., Martínez, M., Cuellar, W., Rodriguez, F., Cuellar, P. et al. (2021). Explorando la relación de las abejas sin aguijón y plantas en los TCOs Sirionó y Lomerío: guía ilustrada. Museo de Historia Natural Noel Kempff – UAGRM & Fundación Noel Kempff, Santa Cruz de la Sierra, Bolivia.
Urquizo, O. N., Cardozo-Alarcón, F., Adler, M., Lozano, R., Calcina-Mamani, S., Collao-Alvarado, K. et al. (2022). Pollen preference patterns by Tetragonisca angustula (Apidae: Meliponini) in a Boliviano–Tucumano Forest. Neotropical Entomology, 51, 649–659. https://doi.org/10. 1007/s13744-022-00986-3
Vit, P., Pedro, S. R. M., Maza, F., Meléndez Ramírez, V. y Frisone, V. (2018). Diversity of stingless bees in Ecuador, pot-pollen standards, and Meliponiculture fostering a living museum Meliponini of the world. En P. Vit, S. R. M. Pedro y D. W. Roubik (Eds.), Pot-Pollen in stingless bee Melittology (pp. 207–227). Rio de Janeiro: Springer International Publishing. https://doi.org/10.1007/978-3-319-61839-5_15
Wille, A. (1960). A new species of stingless bee (Meliponini) from Bolivia. Revista de Biología Tropical, 8, 219–223. https://doi.org/10.15517/rev.biol.trop.1960.30214
Wille, A. (1962). A revision of the subgenus Nogueirapis: an archaic group of stingless bees (Hymenoptera: Apidae). Journal of the New York Entomological Society, 70, 218–234.
Wille, A. (1965). A new species of stingless bee (Meliponini) from Costa Rica. Revista de Biología Tropical, 13, 3. https://doi.org/10.15517/rev.biol.trop.1965.28774
Wille, A. (1979). Phylogeny and relationships among the genera and subgenera of the stingless bees (Meliponinae) of the world. Revista de BiologíaTropical/International Journal of Tropical Biology and Conservation, 7, 241–277.
José G. Palacios-Vargas a, Erika Rivero-Sánchez a, Yan Gao b, Margarita Ojeda a, *
a Universidad Nacional Autónoma de México, Facultad de Ciencias, Departamento de Ecología y Recursos Naturales, Laboratorio de Ecología y Sistemática de Microartrópodos, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Mexico City, Mexico
b China Shanghai Natural History Museum, Shanghai Science & Technology Museum, Shanhaiguan Road, Jing’an district, Shanghai 200041, China
As a result of 40 years of work and many projects on soil fauna and especially springtails, a collection of edaphic microarthropods has been established at the Facultad de Ciencias, UNAM, and has the name of “Colección de ácaros y colémbolos del Laboratorio de Ecología y Sistemática de Microartrópodos”. During the revision of material belonging to the subfamily Pseudachorutinae kept in the Collembola collection, we obtained 581 records of 28 species of Pseudachorutes, from 20 states and 62 localities from Mexico. Three are new records: P. ca. algidensis, P. ca. crassus, and P. reductus, and 5 new species from soil, litter and epiphytic plants in Mexico are described and illustrated herein: P.tabasquensis sp. nov., P. mexicanus sp. nov., P. chichinautzin sp. nov., P. tillandsiodes sp. nov., and P.veracruzensis sp. nov.
Cinco especies nuevas de Pseudachorutes (Collembola: Neanuridae) de México
Resumen
Como resultado del trabajo de más de 40 años y diversos proyectos sobre la fauna del suelo, especialmente colémbolos, se estableció en la Facultad de Ciencias, UNAM, una colección de microartrópodos edáficos, registrada con el nombre de “Colección de ácaros y colémbolos del Laboratorio de Ecología y Sistemática de Microartrópodos”. Durante la revisión del material depositado en ella, especialmente de la subfamilia Pseudachorutinae, se obtuvo información de 581 registros de 28 especies de Pseudachorutes distribuidos en 20 estados y 62 localidades del país. De éstas, 3 son nuevos registros (P. ca. algidensis, P. ca. crasus y P. reductus) y 5 son especies nuevas provenientes de suelo, hojarasca y plantas epífitas en México, que se describen e ilustran aquí: P.tabasquensis sp. nov., P. mexicanus sp. nov., P. chichinautzin sp. nov., P. tillandsiodes sp. nov. y P.veracruzensis sp. nov.
The genus Pseudachorutes Tullberg, 1871 (Neanuridae: Pseudachorutinae) was based on the type species Pseudachorutessubcrassus Tullberg, 1871 characterized by: 1) ocelli 8+8; 2) postantennal organ in one circle or ellipse; 3) Ant. III and IV dorsally fused, Ant. IV generally with 6 sensilla and apical bulb, Ant. III organ with 2 microsensilla in a cuticular fold, 2 guard sensilla and one microsensillum; 4) bucal cone sharp, mandible with 2 or more teeth, maxilla styliform; 5) unguiculus absent; 6) furcula usually well developed, mucro present; 7) sixth abdominal segment always visible in dorsal view, anal spines absent (Christiansen & Bellinger, 1998; Fjellberg, 1998; Palacios-Vargas, 1990).
Currently there are 119 species of Pseudachorutes in the world (Bellinger et al., 2023), and for Mexico records of 20 species from 18 states are known (Arango-Galván et al., 2007; Cutz-Pool et al., 2003, 2007a, b, 2008; Palacios-Vargas, 1997, 2005; Palacios-Vargas & Castaño-Meneses, 2003; Palacios-Vargas et al., 2000, 2007; Vázquez, 1988; Vázquez & Palacios-Vargas, 1990, 2004). The genus is found in great abundance in many ecosystems and biotopes, including soil, litter, mosses, epiphytic plants and even the forest canopy.
During a recent project to study a collection of the subfamily Pseudachorutinae housed in the Collembola collection of the Laboratorio de Ecología y Sistemática de Microartrópodos (LESM), we found 5 new species of Pseudachorutes and their descriptions and illustrations are given herein.
Materials and methods
Examination of the material deposited in the LESM scientific collection “Colección de ácaros y colémbolos” (register number D.F.-ENT-229-09-09, issued by Subsecretaría de Gestión para la Protección Ambiental, Dirección de Vida Silvestre) was made to separate all the specimens of the genus Pseudachorutes.The specimens are permanently mounted in Hoyer’s medium slides. Measurements are presented as the range with means in parentheses and expressed in micrometers (µm). Drawings were made with the aid of a phase contrast microscope Carl Zeiss Standard 3 K7, equipped with a drawing tube. The full body scales correspond to 500 µm, and the rest of the structures to 100 µm.
Type specimens are deposited in the LESM. The chaetotaxy system follows that of Jordana et al. (1997). Abbreviations used in this paper are: Ant. = antennal segment (s), Abd. = abdominal segment (s), PAO = postantennal organ, sgd = dorsal guard sensillum, sgv = ventral guard sensillum, Th. = thoracic segment (s).
Descriptions
Class Collembola Lubbock, 1870
Order Poduromorpha Börner, 1913
Family Neanuridae Börner, 1901
Subfamily Pseudachorutinae Börner, 1906
Genus Pseudachorutes Tullberg, 1871
New records
Pseudachorutes ca. algidensis Carpenter, 1925. Mexico: Hidalgo: Mineral El Chico, 2,900 m asl, 1 specimen, ex Tillandsia violacea, 10-IX-1998, J.A. Monterrubio, col. Pseudachorutes ca. crassus da Gama, 1964. Mexico: Popocatépetl, 3,800 m asl, 1 specimen, ex litter, 29-I-1983, J.G. Palacios, col. Pseudachorutes reductus Thibaud & Massoud, 1983. Mexico: Veracruz: Estación de Biología Tropical, Los Tuxtlas, 8 specimens, 20-X-1997, J. Álvarez, col. Tamaulipas: Rancho El Cielo, 975 m asl, 1 specimen, ex soil, XI-1987, F. J. Villalobos, col.
Description. Body length (n = 7): 2,625 µm (range: 1,250-4,000 µm). Body color gray-violet, with dark eyes patches. Granulations are fine and homogenous. Body setae simple and smooth, but with 2 kinds of setae, long macrosetae (M 22-24 µm) and short microsetae (m 10-12 µm), the sensorial setae relatively long (54-56 µm) (Fig. 1 A, B).
Figure 1. A-L. Pseudachorutestabasquensis sp. nov. A) Dorsal chaetotaxy from head to Th. III; B) dorsal chaetotaxy from Abd. III. to Abd. VI; C) Ant. III-IV right antenna, dorsal view; D) Ant. III-IV right antenna, ventral view; E) PAO and nearby eyes; F) mandible; G) maxilla; H) labium; I) femur, tibiotarsus, and unguis III; J) dens and mucron; K) female genital plate; L) male genital plate.
Table 1
Pseudachorutes tabasquensis sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. orghidani, P. conicus, and P. tabasquensis sp. nov.
1a.
sd
d
oc
c
p
Number of setae
5
4+1
3
1
4
Setae absent
c1, c3, c4
1b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
2
5
a5
m5
Th. III
3
2
5
a2, a5
m5
Abd. I-III
4
–
5
a2
Abd. IV
4
–
6
Abd. V
4
–
4
Abd. VI
2
2
2+1
1c.
Characters
P. conicus
P. tabasquensis sp. nov.
P. orghidani
Labium setae L
–
+
?
Ant. IV sensilla
5
6
5
Ventral tube setae
3+3
4+4
?
Mandible teeth
3
2
3
Ventral file on Ant. IV
–
25-30 short setae
25-30 cuniform
Vesicles of PAO
13-15
13-17
17
Antennae as long as head. Ant. I with 7 setae, Ant. II with 11 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II: III+IV as 1: 1; 1.8. Ant. III-organ with 2 small straight internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.4 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 cylindrical sensilla, seta “i”, and one subapical organite (Fig. 1C), ventral file with 25-30 short and strongly spine-like setae (Fig. 1D). PAO elliptical composed of 13-17 simple vesicles, 0.9 times smaller than the nearest eyes (Fig. 1E). 8+8 eyes, F, G are 0.7 times smaller than others. Buccal cone elongated. Mandible with 2 slender teeth (Fig. 1F). Maxilla styliform, with 2 blades, one has an apical tooth, another has 2 apical teeth (Fig. 1G). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae. Setae L spine-shape (Fig. 1H).
Dorsal chaetotaxy as in figure 1A-B, table 1a, b. Seta a0 on the head absent, unpaired seta d1 present, sometimes, one additional seta d3’ between the seta d3 present. Th. I with 3+3 setae, plus 1+1 lateral. Seta a2 present on Th. II, but absent from Th. III to Abd. V. Sensorial setae s on the body in position of p4 and m6 on the thoracic segments II and III, and p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 2 times as long as the macrosetae. The ratio of the largest Abd. V setae and inner unguis length is 1.0. Thoracic sterna without setae.
Legs setation from I to III is, tibiotarsi 19, 19, 18, without tenent hairs; Femora 10, 10, 11, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 7; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis wide with one inner tooth near 1/3 part from the basal, and a weakly subbasal lateral tooth. Ratio of tibiotarsus III and unguis about 1.6. Unguiculus absent (Fig. 1I).
Furcula is well developed. Dens dorsally with 6 setae, ventral with a smooth area. Mucro straight, 1.8 times shorter than dens, with granulations and 2 small lamellae (Fig. 1J). Tenaculum with 3+3 teeth. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 6-14 circumgenital setae and 1+1 eugenital setae (Fig. 1K). Male genital plate with 3+3 pregenital setae, 25 circumgenital setae and 4+4 eugenital setae (Fig. 1L).
Taxonomic summary
Type material. Holotype: male mounted on a slide (FC-UNAM: LESM-AC: 23013). 5 paratypes females and 1 juvenile mounted on slides (FC-UNAM: LESM-AC: 23014-23019), same data as holotype.
Type locality. Mexico, Tabasco, Tapijulapa, outside cave “Las Sardinas”, ex litter, 14-III-2002, D.A. Estrada col.
Etymology. The name is locative for the state of Tabasco where the type locality is.
Remarks
Pseudachorutes tabasquensis sp. nov. shares with P. orghidani Massoud & Gruia, 1973 the presence of 1 internal and 1 lateral tooth on unguis. The new species also resembles P. conicus Lee & Kim, 1994 from Korea due to the presence of 2 types of body setae. They all share a similar number of PAO vesicles (Table 1c), dens with 6 setae and tenaculum with 3+3 teeth. Main differences between them are shown in Table 1c. Additionally, P. conicus has a very long and thin unguis, but in P. tabasquensis it is short and thick.
Description. Body length (n = 17): 814 µm (range: 470-1,350 µm). Color of the body gray-violet, with a dark eyes patch. Granulations are fine and homogenous. Posterior setae of body long and capitated (Fig. 2A).
Antennae as long as head. Ant. I with 7 setae, Ant. II with 11 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1.4; 3.2. Ant. III-organ with 2 small straight internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.2 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with simple apical bulb, 6 cylindrical sensilla, seta “i”, 1 microsensillum and 1 subapical organite (Fig. 2B), ventral file poorly developed, with 20-35 short setae (Fig. 2C). PAO elliptical composed of 5-6 simple vesicles, 1.1 times as long as the nearest ocelli (Fig. 2D). 8+8 small ocelli, F, G are 0.9 times smaller than others. Buccal cone short. Mandible with 2 slender teeth (Fig. 2E). Maxilla with 2 blades, each has 2 apical teeth (Fig. 2F). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae, setae L spine-shape (Fig. 2G).
Dorsal chaetotaxy as in figure 2A, table 2b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. V. Sensory setae s on the body in position of p4 and m6 on Th. II and III, and p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensory setae longer than 1.1-1.5 times as long as body setae. Ratio of largest Abd. V setae and inner unguis length is 1.4. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 2+2 pregenital setae, 4-9 circumgenital setae and 1+1 eugenital setae (Fig. 2H). Male genital plate with 3+3 pregenital setae, 10 circumgenital setae and 4+4 eugenital setae (Fig. 2I).
Tibiotarsi I, II, III with 18, 18, 17 setae respectively, with 1 tenent hair long and capitate (Fig. B8). Femora I, II, III with 9, 9, 10 setae respectively. Trochanters with 5 setae each. Coxae I, II, III with 3, 7, 7 setae respectively. Subcoxae 2 I, II, III with 0, 2, 2 setae respectively. Subcoxae 1. I, II, III with 1, 2-3, 2-3 setae respectively. Unguis wide with 1 weakly apical inner tooth. Ratio of tibiotarsus III and unguis about 1.0. Unguiculus absent (Fig. 2J).
Furcula well developed. Dens dorsally with 5 setae. Mucro straight, 3 times shorter than dens, with two larger bladder-like swelling visible and a hook-like end (Fig. 2K). Tenaculum with 3+3 teeth.
Figure 2. A-K. Pseudachorutesmexicanus sp. nov. A) Dorsal chaetotaxy from head to abdomen VI; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO and nearby eye; E) mandible; F) maxilla; G) labium; H) female genital plate; I) male genital plate; J) femur, tibiotarsus, and unguis III; K) furcula and tenaculum.
Taxonomic summary
Type material. Holotype: female mounted on a slide (FC-UNAM: LESM-AC: 22996). 16 paratypes: 4 females, 1 male and 11 juveniles under slides (FC-UNAM: LESM-AC: 22997-23012), same data as holotype.
Type locality. Mexico, Hidalgo, Mineral El Chico, 10-IX-98, J.A. Monterrubio, col. Ex Tillandsia violacea.
Etymology. The name of the new species is after the country of the type locality: Mexico.
Remarks
This species resembles P. americanus Stach, 1949 with a similar number of vesicles in PAO (P. americanus with 5-8 vesicles), unguis with 1 minute tooth near apex, one large, clavate tenent hair, ventral tube with 4+4 setae, tenaculum with 3+3 teeth and posterior abdominal setae clearly capitate. However, dens of the new species with 5 setae, maxilla with 2 blades and each has 2 apical teeth, 6 cylindrical sensilla on Ant. IV and a poorly developed ventral file with 20-35 short setae is different from P. americanus.
Description.Body length (n = 8): 1,904 µm (range: 850-2,010 µm). Color of the body gray-violet, with a dark eyes patch. Granulations homogenous. Lateral and posterior body setae are longer and truncate (Fig. 3A).
Table 2
Pseudachorutes mexicanus sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy.
2a.
sd
d
oc
c
p
Number of setae
5
4+1
3
4
4
2b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
3
5
a4
Th. III
3
3
5
Abd. I-III
3
2
5
Abd. IV
4
–
5
Abd. V
3
–
4
Abd. VI
2
2
2+1
Antennae as long as head. Ant. I with 7 setae, Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1.2: 2.0. Ant. III-organ with 2 small internal sensilla under a cuticular fold, 2 guard sensilla (sgv about 1.1 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 cylindrical sensilla, seta “i”, and one subapical organite (Fig. 3B), ventral file with about 10 strong, spine-like setae and several slender, normal setae (Fig. 3C). Some setae on Ant. IV are blunt. PAO elliptical composed of 10-12 vesicles, sometimes, 1 or 2 of them inside in the others, subequal to nearest eyes (Fig. 3D). 8+8 eyes, F, G are 0.7 times smaller than others. Buccal cone elongated. Mandible is not clearly detected, about 2 or 3 slender teeth. Maxilla with 2 blades and one has an apical tooth, and the other styliform (Fig. 3E). Labium with normal chaetotaxy of the genus from setae A to G and 5 lateral setae, setae L spine-shape (Fig. 3F).
Dorsal chaetotaxy as in Fig. 3A, table 3a, b. Seta a0 on the head absent, unpaired seta d1 present. Th. I with 3+3 setae, plus 1+1 lateral. Setae a2 present on Th. II, but absent from Th. III to Abd. V. Sensorial setae s on the body in position of p4 and m6 on the thoracic segments II and III, and p5 from Abd. I to IV and p3 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 1.2-1.7 times as long as the normal setae. The lateral setae of the body longer and blunt. Ratio of largest Abd. V setae and inner unguis length is 1.0. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 9 circumgenital setae and 1+1 eugenital setae (Fig. 3G). Male genital plate with 3+3 pregenital setae, 8 circumgenital setae and 4+4 eugenital setae (Fig. 3H).
Leg setation from I to III, is tibiotarsi 19, 19, 18, with 1 acuminate tenent hair a little longer than others; femora 9, 9,10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 7; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis wide, with one inner tooth near 1/3 part from the basal. Sometimes, a pair of weakly subbasal lateral tooth is present. Ratio of tibiotarsus III and unguis about 1.6. Unguiculus absent (Fig. 3I).
Figure 3. A-J. Pseudachoruteschichinautzin sp. nov. A) Dorsal chaetotaxy; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO, eye patch and ocular setae; E) maxilla; F) labium; G) female genital plate; H) male genital plate; I) femur, tibiotarsus, and unguis III; J) furcula and tenaculum.
Furcula well developed. Dens dorsally with 6 setae, ventral with fine granulate. Mucro straight, 1.7 times shorter than dens, with 2 big lamella and a clearly hook-like end. Tenaculum with 3+3 teeth (Fig. 3J).
Taxonomic summary
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 22974). 7 paratypes: 4 females, 2 males and 1 juvenile under slides, same data as holotype (FC-UNAM: LESM-AC: 22975-22981).
Type locality. Mexico; Morelos, Derrame Chichinautzin. 12-IX-1976. J. Palacios, col. Ex. Tillandsia prodigiosa.
Etymology. The name is that of the Chichinautzin lava flow (state of Morelos), as a noun of type locality.
Remarks
Pseudachorutes chichinautzin sp. nov. shares with P. orghidani and P. subcrassus Tullberg, 1871 the presence of 1 internal and 1 pair of lateral teeth on unguis. The new species also resembles P. tabasquensis sp. nov. with 2 types of body setae, Ant. IV with 6 sensilla, trilobed apical bulb. Dens with 6 setae and tenaculum with 3+3 teeth. Main differences between them can be seen in table 3c.
Table 3
Pseudachorutes chichinautzin sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. orghidani, P. subcrassus, P. chichinautzin sp. nov. and P. tabasquensis sp. nov.
Description. Body length (n = 6): 2,260 µm (range: 1,750-2,860 µm). Color of the body violet, with a white strip from Th. I to Th. II and a dark eyes patch. Granulations fine and homogenous. Body with simple and spine-like setae, p row longer on last abdominal segments (24-40 µm), long sensorial setae (75-100 µm) (Fig. 4A, B).
Antennae shorter than head, 155 µm and 180 µm respectively. Ant. I with 7 setae Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1; 1.9. Ant. III-organ with 2 small curving sensilla under a cuticular fold, 2 guard sensilla (sgv is about 1.4 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV with trilobed apical bulb, 6 thin and cylindrical sensilla, seta “i”, one microsensillum and one subapical organite (Fig. 4C), ventral side with about 20 setae with some thick and spine-like (Fig. 4D). PAO elliptical composed of 14 vesicles, 1.2 times as long as the nearest eyes. 8+8 eyes, F, G 0.9 times as big as others (Fig. 4E). Buccal cone elongated. Mandible has 2 big teeth, the apical with clearly 3 small teeth (Fig. 4F). Maxilla with 2 blades, one with 2 apical teeth (Fig. 4G). Labium with normal chaetotaxy of the genus from setae A to G and 4 lateral setae, setae L reduced to a minus spine, difficult to see (Fig. 4H).
Dorsal chaetotaxy as in figure 4A, B, table 4a, b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. V. m5 present on Th. II to Th. III. Sensorial setae on body in position of p4 and m6 on thoracic segments II and III, p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 2.5-4.0 times as long as the normal setae. Ratio of largest Abd. V setae and inner unguis length is 0.6. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 9 circumgenital setae and 1+1 eugenital setae (Fig. 4I). No males were found.
Figure 4. A-K. Pseudachorutes tillandsiodes sp. nov. A) Dorsal chaetotaxy from head to Th. III; B) dorsal chaetotaxy from Abd. III. to Abd. VI; C) Ant. III-IV right antenna, dorsal view; D) Ant. III-IV right antenna, ventral view; E) PAO, eye patch and ocular setae; F) mandible; G) maxilla; H) labium; I) female genital plate; J) femur, tibiotarsus, and unguis III; K) dens and mucron.
Leg setation from I to III, is tibiotarsi 19, 19, 18, no tenent hair; femora 9, 10, 10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 5,5,5; coxae 3, 7, 8; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis with one clearly inner tooth at the basal side. Ratio of tibiotarsus III and unguis about 1.5. Unguiculus absent (Fig. 4J).
Furcula well developed. Dens dorsally with 6 setae, ventral with fine granulations. Mucro straight, 2.2 times shorter than dens, with 2 thin but long lamella, without hook-like end (Fig. 4K). Tenaculum with 3+3 teeth.
Table 4
Pseudachorutes tillandsiodes sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy, c) main characters between P. gilvusi, and P.tillandsiodes sp. nov.
4a.
sd
d
oc
c
p
Number of setae
5
4+1
3
4
4
4b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
5
2
5
m4
Th. III
4
2
5
a2
m4
Abd. I-III
4
–
5
a2
Abd. IV
4
–
5
a2
Abd. V
4
–
3
P4
Abd. VI
2
2
2+1
4c.
Characters
P. gilvus
P. tillandsiodes sp. nov.
Setae d1 on head
1+1
1
Setae number on Th. I
2+2
3+3
Setae a2 on Th. II
–
+
Ant. IV sensilla
7
6
Ventral tuve setae
3+3
4+4
Taxonomic summary
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 23172). 5 Paratypes: 4 female and 1 juvenile (FC-UNAM: LESM-AC: 23173-23177).
Type locality. Mexico; Hidalgo, Mineral El Chico. 10-IX-98. J. A. Monterrubio, col. Ex Tillandsia violacea.
Etymology. The name is taken from the epiphytic genus Tillandsia (Bromeliacea) habitat where the species was found.
Remarks
Pseudachorutes tillandsiodes sp. nov. resembles P. gilvus Oliveira & Deharveng, 1995 with white strips and long sensilla on the body, similar number of postantennal organ vesicles (P. gilvus with 11-15 vesicles), unguis with 1 inner tooth, tibiotarsi I-III with 19, 19, 18 setae and dens with 6 setae. The main differences between them are shown in table 4c. However, P. gilvus has 3 white stripes on the body: the first on the posterior part of head and the middle of Th. I, the second on the mesothorax and the third on Abd. I-II; P. tillandsiodes sp. nov. only with 1 white stripe from Th. I to Th. II.
Material examined. Holotype: female under slide (FC-UNAM: LESM-AC: 2133a). 10 paratypes: 7 females and 3 males under slides, same data as holotype (FC-UNAM: LESM-AC: 2131a-2132c, 2132a-2132e, 2133b, 2133c)
Type locality. Mexico; Veracruz, Xalapa, La Herradura. 26-IX-26-10/1998, ex Bosque Mesófilo de Montaña, J. Márquez, col.
Description. Body length (n = 11): 1,900 (range: 1,050-3,400 µm). Color body violet, with a dark eyes patch. Granulations fine and homogenous. Body with short and simple setae (10-12), some of them longer in abdomen segments, especially on 4th segment (22-26 µm), long sensorial setae (67-75 µm) (Fig. 5A).
Antennae little shorter than head, 100 µm and 105 µm, respectively. Ant. I with 7 setae, Ant. II with 12 setae. Ant. III and IV dorsally fused. Ant. segments ratio I: II; III+IV as 1: 1; 2. Ant. III-organ with 2 small straight sensilla under a cuticular fold, 2 guard sensilla (the sgv is about 1.1 times as long as sgd) and 1 microsensillum close to ventral guard sensillum. Ant. IV dorsally with trilobed apical bulb, 6 thin and cylindrical sensilla, seta “i”, 1 microsensillum and 1 subapical organite (Fig. 5B), ventral side has a distinct ventral file with about 40 short setae (Fig. 5C). PAO elliptical composed of 17-20 vesicles, 1.5 times as long as the nearest eyes. 8+8 eyes, F, G 0.8 times as big as others (Fig. 5D). Buccal cone elongated. Mandible has 2-3 big teeth (Fig. 5E). Maxilla with 1 blade and 2 apical teeth (Fig. 5F). Labium with normal chaetotaxy of the genus from setae A to G and 3 lateral setae, one longer spine-like setae present at the place of setae L (Fig. 5G).
Dorsal chaetotaxy as in figure 5A and table 5a,b. Seta a0 on head absent, unpaired seta d1 present. Th. I with 3+3 setae. Setae a2 present on Th. II, but absent from Th. III to Abd. IV. m5 present on Th. II to Th. III. Sensorial setae s on the body in position of p4 and m6 on Th. II and III, p5 from Abd. I to IV and p2 on Abd. V. Sensorial formula of the body 022/11111. Sensorial setae 6.5-7.0 times as long as the shorter setae. The ratio of the largest Abd. V setae and inner unguis length is 0.5. Thoracic sterna without setae. Ventral tube with 4+4 setae. Female genital plate with 3+3 pregenital setae, 7 circumgenital setae and 1+1 eugenital setae (Fig. 5H). Male genital plate with 2+2 pregenital setae, 16 circumgenital setae and 4+4 eugenital setae (Fig. 5I).
Leg setation from I to III, is tibiotarsi 19, 19, 18, no tenent hairs; femora 13, 11, 10, one ventro-proximal seta is an acuminate tenent hair; trochanters with 6,6,5; coxae 3, 7, 8; subcoxae 2: 0, 2, 2; subcoxae 1: 1, 2, 2. Unguis with 1 big and 1 small inner tooth together with 2 pairs of lateral teeth (Fig. 5J). Ratio of tibiotarsus III and unguis about 1.4. Unguiculus absent (Fig. 5K).
Furcula well developed. Dens dorsally with 6 setae, ventral granulate. Mucron granulated with broad and long lamella, 1 slightly hook-like end, 2.0 times shorter than dens (Fig. 5L). Tenaculum with 3+3 teeth.
Etymology. The name is a locative for the State of Veracruz where the type locality is found.
Remarks
Pseudachorutes veracruzensis sp. nov. resembles P. orghidani with granules on mucron and dens with 6 setae, mandible with 3 teeth, unguis with lateral teeth, similar number of PAO (P. orghidani with 17 vesicles) and ventral file on Ant. IV. The main difference between the species is the shape of short setae in the ventral file (P. orghidani has small setae with apex truncate) and number of sensilla on Ant. IV (P. orghidani with 5) and the teeth on unguis (P. orghidani only with 1+1 lateral teeth).
Discussion
After revision and analysis of the material deposited in the LESM collection, we were able to update the knowledge of genus Pseudachorutes in Mexico, describing 5 new species: P. tabasquensis sp. nov., P. veracrucensis sp. nov., P. tillandsiodes sp. nov., P. chichinautzin sp. nov., and P. mexicanus sp. nov. Three new records for the country are added, for the following species: P. ca. algidensis from Hidalgo, P. ca. crassus from Estado de México, these must be confirmed with the collection of more specimens that will allow the species to be fully determined. For now, this information is presented as an element to show the richness of species of the genus in the country. Pseudachorutes reductus has a distribution in the Antilles and southern Florida, the new records in Mexico extend its distribution area and confirm the affinity of the species to the Atlantic area. Total records for Pseudachorutes is increased to 28 species, from 20 states and 62 localities within the country. Quintana Roo and Hidalgo were the states with the highest number of species present (11 and 10, respectively), followed by Morelos, Estado de México, Puebla, Veracruz, and Guerrero (8, 7, 6, 6, and 4 species recorded in each one), 6 states have 3 species, 5 have 2 and Querétaro and San Luis Potosí only have 1 species recorded. Three species are widely distributed in Mexico: P. corticolus, P. simplex, and P. subcrassoides. Diversity of the genus in Mexico is around 21% of the total known worldwide, and therefore the country becomes one with the greatest number of species present.
Figure 5. A-L. Pseudachorutes veracruzensis sp. nov. A) Dorsal chaetotaxy; B) Ant. III-IV right antenna, dorsal view; C) Ant. III-IV right antenna, ventral view; D) PAO, eye patch and ocular setae; E) mandible; F) maxilla; G) labium; H) female genital plate; I) male genital plate; J) femur, tibiotarsus, and unguis III; K) unguis III, ventral view; L) dens and mucron.
Regarding biotopes where the species preferably live, we found that litter, epiphytic plants, soil, mosses and decaying wood are the most suitable habitats for this springtail. However, sand, canopy, wood, caves and anthills are also microhabitats used by some species.
Table 5
Pseudachorutes veracruzensis sp. nov. a) Head chaetotaxy, b) dorsal chaetotaxy.
5a.
sd
d
oc
c
p
Number of setae
5
4+1
3
2
4
Setae absent
c1, c3 c4
5b.
a
m
p
Setae absent
Th. I
–
3
–
m2
Th. II
4
2
5
a5
m4
Th. III
3
2
5
a2, a5
m4
Abd. I-III
3
–
5
a2, a5
Abd. IV
4
–
5
a2, a3
Abd. V
3
–
4
a4
Abd. VI
3
2
2+1
The analysis and revision of the morphology of the 5 species described, allowed us to come to the following conclusions about the morphology of the genus that needs to be taken in consideration. Size of body setae should be a characteristic to be taken into consideration for the description of the species, since we observed that it is a variable character. Antennae are a uniform structure and characters associated with the sensory organ of Ant. III have little relevance at a specific level. Sensory file of Ant. IV, present a varied shape and number, from 8 to 40 setae. Apical vesicle of Ant. lV is trilobed in most species and others have a simple shape. Number of ocelli is stable within the genus, presenting 8+8. The maxillae and mandibles vary in all species. Maxillae can be needle-shaped, crocheted or styliform, they may or may not be lamellar and the number of lamellae is variable, and in most cases, there are 1 to 2 apical teeth. The number of teeth in the mandibles in almost all species ranges from 1 to 5. The number of teeth in the retinaculum is a constant character among the species of the genus, with 3+3 teeth. The ventral tube in almost all species has 4+4 setae. The number of setae in the dens varies from three to seven, but most have six setae. The mucron has 2 lamellae in all species, except in P. reductus, where it is greatly reduced. The shape of the mucron is variable, from elliptical, triangular, elongated, short, curved, widened, or with a bladder-shaped base.
Acknowledgments
This study had the support from the International Relationships of Universidad Nacional Autónoma de México issued to the first author and partially by the National Natural Sciences Foundation of China (No. 3217047 and 32370491). Blanca E. Mejía Recamier helped remount specimens. Kenneth A. Christiansen gave criticism, suggestions, and literature. We thank Erick García and Eduardo Pacheco who assigned the catalogue numbers for the slides. Daniel A. Estrada, Jesús A. Monterrubio, Juan Márquez donated the specimens.
References
Arango-Galván, A., Cutz-Pool, L. Q., & Cano-Santana, Z. (2007). Estructura de la comunidad de colémbolos del mantillo de la Reserva Ecológica del Pedregal de San Ángel: Riqueza, composición y abundancia relativa. Entomología Mexicana, 6, 397–400.
Bellinger, P. F., Christiansen, K. A., & Janssens, F. (2023). Checklist of the Collembola of the World. Retrieved on November 11th, 2023 from: http://www.collembola.org
Christiansen, K. A., & Bellinger, P. F. (1998). The Collembola of North America, North of the Río Grande: a taxonomic analysis. Part I. Grinnell, Iowa Grinnell College.
Cutz-Pool, L. Q., Palacios-Vargas, J. G., & Vázquez, M. M. (2003). Comparación de algunos aspectos ecológicos de Collembola en cuatro asociaciones vegetales de Noh-Bec, Quintana Roo, México. Folia Entomológica Mexicana, 42, 91–101.
Cutz-Pool, L. Q., García-Gómez, A., & Palacios-Vargas, J. G. (2007a). Primer estudio de colémbolos (Hexapoda: Collembola) de la Hoya de las Guaguas en el estado de San Luis Potosí, México. Dugesiana, 14, 47–51.
Cutz-Pool, L. Q., Palacios-Vargas, J. G., Castaño-Meneses, G., & García-Calderón, N. E. (2007b). Edaphic Collembola from two agroecosystems with contrasting irrigation type in Hidalgo State, Mexico. Applied Soil Ecology, 36, 46–52. https://doi.org/10.1016/j.apsoil.2006.11.009
Cutz-Pool, L. Q., Palacios-Vargas, J. G., & Castaño-Meneses, G. (2008). Estructura de la comunidad de colémbolos (Hexapoda: Collembola) en musgos corticícolas en un gradiente altitudinal de un bosque templado subhúmedo. Revista de Biología Tropical, 56, 1–10. https://doi.org/10.15517/rbt.v56i2.5620
Fjellberg, A. (1998). Fauna entomologica scandinavica. The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. Vol. 35. Leiden: Brill Publishers.
Jordana, R., Arbea, J. I., Simón, C., & Luciáñez, M. J. (1997). Fauna ibérica. Vol. 8. Collembola, Poduromorpha. Madrid: Museo Nacional de Ciencias Naturales.
Palacios-Vargas, J. G. (1990). Manuales y guías para el estudio de microartrópodos I. Diagnosis y clave para determinar las familias de los Collembola de la Región Neotropical. México D.F.: Facultad de Ciencias, Universidad Nacional Autónoma de México.
Palacios-Vargas, J. G. (1997). Catálogo de los Collembola de México. Las Prensas de Ciencias, UNAM, México D.F.
Palacios-Vargas, J. G. (2005). Collembola. In G. Sánchez-Ramos, P. Reyes-Castillo y R. Dirzo (Eds.), Historia natural de la Reserva de la Biosfera El Cielo Tamaulipas, México (pp. 354–366). Ciudad Victoria, Tamaulipas: Universidad Autónoma de Tamaulipas.
Palacios-Vargas, J. G., & Castaño-Meneses, G. (2003). Seasonality and community composition of springtails in Mexican forests. In Y. Basset, V. Novotny, S. E. Millar, & R. L. Kitching (Eds.), Arthropods of tropical forests. Spatio-temporal dynamics and resource use in the canopy (pp. 159–169). Cambridge, UK: Cambridge University Press.
Palacios-Vargas, J. G., Castaño-Meneses, G., & Mejía-Recamier, B. E. (2000). Collembola. In J. Llorente-Bousquets, E. González-Soriano, & N. Papayero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México. Hacia una síntesis de su conocimiento II (pp. 249–273).México D.F.: Las Prensas de Ciencias, UNAM.
Palacios-Vargas, J. G., Cutz-Pool, L. Q., & Estrada-Barcenas, D. A. (2007). Collembola. In I. Luna, J. J. Morrone, & D. Espinosa (Eds.). Biodiversidad de la Faja Volcánica Transmexicana (pp. 331–344).México D.F.: Las Prensas de Ciencias, UNAM.
Tullberg, T. (1871). Förteckning öfver Svenska Podukrider. Öfversigt af Kongliga Vetenskaaps -Akademiens Förhandlingar, 28, 143–155.
Vázquez, M. M. (1988). Fauna colembológica de hojarasca y suelo. In L. Arriaga, & A. Ortega (Eds.), La sierra de la Laguna de Baja California Sur (pp. 133–148). La Paz, Baja California Sur: Centro de Investigaciones Biológicas de Baja California Sur, A.C.
Vázquez, M. M., & Palacios-Vargas, J. G. (1990). Nuevos registros y aspectos biogeográficos de los colémbolos de la sierra de la Laguna, B. C. S., México. Folia Entomológica Mexicana, 78, 5–22.
Edmundo González-Santillán a, Laura L. Valdez-Velázquez b, *, Ofelia Delgado-Hernández c, Jimena I. Cid-Uribe d, María Teresa Romero-Gutiérrez e, Lourival D. Possani d
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Colección Nacional de Arácnidos, Tercer Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Universidad de Colima, Facultad de Ciencias Químicas y Facultad de Medicina, Km 9 Carretera Colima-Coquimatlán, 28400 Coquimatlán, Colima, Mexico
c Instituto Francisco Possenti, Av. Toluca 621, Olivar de los Padres, Álvaro Obregón, 01780 Mexico City, Mexico
d Universidad Nacional Autónoma de México, Instituto de Biotecnología, Avenida Universidad 2001, Colonia Chamilpa, 62210 Cuernavaca, Morelos, Mexico
e Universidad de Guadalajara, Centro Universitario Tlajomulco, Departamento de Innovación Tecnológica, Carretera Tlajomulco – Santa Fé Km. 3.5 No.595 Lomas de Tejeda, 45641 Tlajomulco de Zúñiga, Jalisco, Mexico
Scorpion species diversity in Colima was investigated with a multigene approach. Fieldwork produced 34 lots of scorpions that were analyzed with 12S rDNA, 16S rDNA, COI, and 28S rDNA genetic markers. Our results confirmed prior phylogenetic results recovering the monophyly of the families Buthidae and Vaejovidae, some species groups, and genera. We recorded 11 described species of scorpions and found 3 putatively undescribed species of Centruroides, 1 of Mesomexovis, and 1 of Vaejovis. Furthermore, we obtained evidence that Centruroides elegans, C. infamatus,and C. limpidus do not occur in Colima, contrary to prior reports. Seven genetically different and medically relevant species of Centruroides for Colima are recorded for the first time. We used the InDRE database (Instituto de Diagnóstico y Referencia Epidemiológicos), which contains georeferenced points of scorpions, to estimate the distribution of the scorpion species found in our fieldwork. Finally, we discuss from a biogeographical, ecological, and medical point of view the presence and origin of the 14 scorpion species found in Colima.
Keywords: Barcoding; Holotype; Medical relevance; Microendemic; New species; Species group; Substrate-specialist
Una aproximación multigenes para identificar a las especies de alacranes (Arachnida: Scorpiones) de Colima, México, con cometarios sobre la diversidad de sus venenos
Resumen
La diversidad de especies de alacranes de Colima se investigó utilizando una aproximación multigenes. Del trabajo de campo se obtuvieron 34 lotes de alacranes que fueron analizados con los marcadores 12S rDNA, 16S rDNA, COI, y 28S rDNA. La comparación con trabajos de filogenia previos nos permitió confirmar la monofilia de las familias Buthidae y Vaejovidae, de algunos grupos de especies y géneros. Encontramos 11 especies de alacranes descritas, 3 putativamente nuevas de Centruroides, 1 de Mesomexovis y 1 de Vaejovis. También obtuvimos evidencia de que Centruroides elegans, C. infamatus y C. limpidus no están distribuidos en Colima, como se registró en trabajos anteriores. Reportamos 7 especies genéticamente distintas y de importancia médica para Colima. Usamos la base de datos del InDRE (Instituto de Diagnóstico y Referencia Epidemiológicos) que contiene puntos georreferenciados de alacranes para estimar la distribución de las especies que recolectamos en el campo. Finalmente, discutimos desde una perspectiva biogeográfica, ecológica y de importancia médica las 14 especies de alacranes que reportamos para Colima.
Palabras clave: Código de barras; Holotipo; Importancia médica; Microendémico; Especie nueva; Grupo de especies; Sustrato-especialista
Introduction
The knowledge of scorpion diversity in North America has improved recently (González-Santillán & Prendini, 2013; Goodman, Prendini, Francke et al., 2021; Ponce-Saavedra & Francke, 2019; Santibáñez-López et al., 2014); however, much remains to be discovered. Local-scale inventories may be a solution to unveil species communities that, in turn, can help conform regional faunas. This approach has rarely been applied to study scorpion diversity. Furthermore, few local faunal studies have been conducted in Mexico, and only a handful of them have been published; other revisionary contributions included limited fieldwork effort (Baldazo-Monsivais et al., 2012, 2016, 2017).
The International Barcode of Life (iBOL) has grown as a powerful tool for discovering biodiversity, among other applications (https://ibol.org). Scorpion barcoding studies have permitted the identification and delimitation of species in several regions of the world (Fet et al., 2014, 2016; Goodman, Prendini, & Esposito, 2021; Podnar et al., 2021). Despite the high diversity of scorpions in Mexico —an update by Ponce-Saavedra et al. (2023) comprises 311 scorpion species— only 1 mini-barcoding study has been conducted (Goodman, Prendini, & Esposito, 2021). Herein, we present a second scorpion barcoding study for this country but aim at discovering the components of a local scorpion assembly. Colima exhibits a complex topology comprising littorals with a tropical climate and extreme topological variation from sea level to mountain ranges rising to over 4,000 m in approximately 5,600 km2. Colima’s territory supports a rich local flora and fauna (Ramírez-Ruiz & Bretón-González, 2016). Colima lies between the limits of the Nearctic and Neotropical biogeographical Realms (Fig. 1B). Beyond its complexity, Colima represents an enclosed littoral surrounded by mountain ranges and 3 large rivers that divide the territory into 2 sections (Fig. 1), which makes it a well-defined and manageable geographical unit ideal for studying a unique community of scorpions. Thus far, 2 families, 3 subfamilies, 5 genera, and 12 species of scorpions have been identified in Colima (Table 1). The knowledge of scorpion diversity and evolutionary studies in Mexico has steadily been unveiling one of the vastest biodiversity hotspots in the world. For instance, 2 of the most diverse scorpion families and subfamilies, Vaejovidae (Syntropinae) and Buthidae (Centruroidinae), have been treated in recent phylogenetic and taxonomic analyses (Esposito & Prendini, 2019; González-Santillán & Prendini, 2013, 2015).
This contribution aims to survey the scorpion richness in Colima, using not only the COI-barcoding genetic marker but 2 additional mitochondrial markers and 1 nuclear marker to establish a framework to build a stable and predictable taxonomy. Taking advantage of robust phylogenies produced for the 2 families distributed in Colima, we use these topologies as a baseline for comparison to test the presence of several previously reported species and to taxonomically circumscribe our fresh samples. Unlike other barcoding studies, our approach seeks to unveil the richness within the state instead of focusing on delimiting the species of a taxonomic group of scorpions.
Materials and methods
We conducted field collections during May and September 2015-2018 in various ecosystems, including tropical deciduous forest, oak-pine, and tropical forest within the state of Colima, at elevations ranging from 47 to 2,200 m. (Table 2). Logistically, we leveraged our collection sites with the help of private landowners who gave us access to their property. Specimen collection methods included direct collection during the day by moving objects on the ground or by ultraviolet detection during the night. To preserve specimens, ethyl alcohol at 90% was used and stored at -80 °C. Each specimen lot carried a label with coordinates and locality information. We obtained scorpions from 14 localities and sequenced 18 samples of Centruroides from Colima (Table 2).
Table 1
List of families, subfamilies, genera, and species recorded in the state of Colima. *Species of Centruroides cited by Ponce-Saavedra et al. (2016). The species in bold font were not found in Colima in this study. Numbered species were reported by González-Santillán et al. (2019).
11. Vaejovis monticola Sissom, 1989 12. Vaejovis sp. mexicanus group
Figure 1. Map of the west coast of Mexico. A, Orographic and hydrographic elements of Colima (COL) and the surrounding states of Jalisco (JAL) and Michoacán (MIC). 1, Manantlán Sierra; 2, massive Cerro Grande; 3, Colima Volcano; 4, Marabasco or Cihuatlán River; 5, Armería River; 6, Coahuayana River. B, Biogeographical provinces (Morrone et al., 2017). Area within the green line corresponds to the Sierra Madre del Sur and north Colima Volcano Trans Mexican Volcanic Belt province (Morrone et al., 2017) —notice that both provinces are connected in Colima. Area outside the green line corresponds to the Pacific Lowlands province (Morrone et al., 2017). Orographic components are indicated in gray scale from light low elevation to dark high elevation.
Table 2
Collection sites of the scorpion species used in this study. The number within parenthesis after the species name is the number of samples processed from this locality and included in the phylogenetic analyses as terminals. Superscript numbers indicate sources of sequences as follows: 1Bolaños et al. (2019), 2Esposito et al. (2018), 3Esposito and Prendini (2019), 4González-Santillán and Prendini (2015). Cells filled in grey color are samples obtained from GenBank.
Species
Municipality
Locality
Latitude
Longitude
Elevation
Thorellius cristimanus (2)
Comala
La Yerbabuena
19°27′59.55″
-103°41′46.64′′
1,358 m
Centruroides ornatus (2)
Comala
Agosto
19°23′51.74′′
-103°44′03.08′′
1,076 m
Thorellius cristimanus
*Centruroides tecomanus2
Colima
Comunidad La Capacha
19°04′58.40′′
-103°41′24.67′′
656 m
*Centruroides tecomanus2
Colima
Tepames
19°06′22.8′′
-103°59′11.07′′
450 m
Thorellius intrepidus (3)
Coquimatlán
El Palapo
19°11′54.6′′
-103°54′50′′
275 m
*Centruroides tecomanus2
Cuauhtémoc
Camino a Altozano
19°18′30.19′′
-103°40′23.83′′
789 m
Thorellius cristimanus
*Centruroides tecomanus1(2)
Cuauhtémoc
Ocotillo
19°20′00.00′′
-103°39′02.00′′
895 m
*Centruroides tecomanus2
Ixtlahuacán
San Gabriel
18°54′24.48′′
-103°44′05.61′′
462 m
Mesomexovis aff. occidentalis
Centruroides hirsutipalpus
Minatitlán
Minatitlán
19°23′01.73′′
-104°03′35.19′′
703 m
Centruroides possanii (2)
Minatitlán
Terrero
19°26′35.94′′
-103°57′05.67′′
2,200 m
Centruroides possanii (2)
Minatitlán
Mirador el Filete
19°26′40′′
-103°58′10′′
2,137 m
Vaejovis sp. (mexicanus group)
Centruroides sp. 2 (2)
Manzanillo
La central
19°08′38.14′′
-104°26′04.10′′
47 m
*Centruroides tecomanus1
*Centruroides tecomanus2
Centruroides sp. 1
Tecomán
Chanchopa
18°51′58.55′′
-103°44′10.10′′
41 m
Thorellius intrepidus (2)
Villa de Álvarez
Rancho Blanco
19°14′24.71′′
-103°45′49.45′′
455 m
Thorellius intrepidus4
La Huerta (Jal.)
Estación de Biología Chamela
19°30′14.15′′
-105°2′16.50′′
33 m
Centruroides elegans (2)
Centruroides suffusus
Durango (Dgo.)
El Salto, 50 km E Durango
23°45′51.41′′
-105°19′49.16′′
2,847 m
Centruroides limpidus
Iguala (Gro.)
Iguala
18°20′11.87′′
-99°29′29.65′′
823 m
Centruroides sculpturatus
Cumpas (Son.)
18 km NE de Nacozari
30°16.473′
-109°50.070′
930 m
Centruroides noxius
Pantanal (Nay.)
Pantanal
21°25′24.42′′
-104°50′47.89′′
921 m
Centruroides huichol
Nayarit
–
–
–
–
Centruroides infamatus115scrp
Guanajuato (Gto.)
Guanajuato
–
–
–
Centruroides ornatus2LP1822
Tandamangapio (Mich.)
Los Tabanos
19.9749°
– 102.84226°
223 m
Centruroides ornatus3 2003
Michoacán
–
–
–
–
*Centruroides tecomanus11 25scrp
Comala (Col.)
–
–
–
–
*Centruroides tecomanus12 2007
Michoacán
–
–
–
–
Mesomexovis occidentalis4 LP 7056
Acapulco (Gro.)
Cumbres de Llano Largo
16°49.505
-99°49.9990
317 m
Table 2. Continued
Species
Municipality
Locality
Latitude
Longitude
Elevation
Mesomexovis spadix4LP6373
León (Gto.)
San Antonio de Padua
20°34.5170
-100°57.217
–
Mesomexovis subcristatus4LP 2049
Tehuacán (Pue.)
Tehuacán, 2 km east
18°24.0020
-97°22.8670
1435 m
Thorellius cristimanus4LP 5325
Álvaro Obregón (Mich.)
Álvaro Obregón
19°02.3100
-102°58.405
462 m
Thorellius cristimanus4LP 6551
Coquimatlán (Col.)
Road to Coquimatlán, km 71
19°06.7750
-103°51.1850
336 m
Thorellius intrepidus4LP 6377
Comala (Col.)
Comala
19°19.000
-103°45.0000
–
Thorellius intrepidus4LP 6379
Colima (Col.)
Los Ortices
19°06.0468
-103°44.0226
343 m
Vaejovis carolinianus4LP 1576
South Carolina
–
–
–
–
Vaejovis pequeno4LP 6308
Soyopa (Son.)
Sierra El Encinal, 9 km from crossroad on Highway Mex 16 to El Encinal
28°35.4120
-109°27.1480
380 m
Vaejovis rossmani4LP 2027
Hidalgo (Tams.)
Conrado Castillo
23°56.01735
-99°28.04817
–
*Genetically differentiated species.
We also included 8 species from outside the state to test the presence of C. elegans, C. infamatus, and C. limpidus reported previously in the literature (Ponce-Saavedra et al., 2016); C. noxius and C. suffusus for comparative purposes and samples of C. tecomanus and C. ornatus from GenBank with a total of 31 specimens of the genus Centruroides within 4 species groups. Unlike buthids, we obtained 12 samples of vaejovids to include in this analysis. To evaluate the identity of vaejovids, we used the BLAST® suite (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to search for similar sequences deposited in the nucleotide collection database at NCBI. Thorellius has been revised recently (González-Santillán & Prendini, 2018). Therefore, several DNA sequences are available, and fewer sequences of Mesomexovis sp. and Vaejovis sp. were available in GenBank, as they are still unrevised. Using the genetic markers as queries, we obtained 10 additional samples. The total number of taxonomic specimens used for these analyses was 53 (Tables 2, 4).
Genomic DNA was extracted from the legs or pedipalp of specimens using Qiagen Dneasy/trisol method Tissue Kits or a DNAzol Genomic DNA isolation Reagent kit (Molecular Research Center INC, Cincinnati, Oh). We amplified 3 mitochondrial markers, 12S rDNA, 16S rDNA, and the barcode COI, and the nuclear marker 28S rDNA. We performed the Polymerase Chain Reaction with the following thermal profile: an initial denaturation step (3 min at 94°C) followed by 35 cycles including denaturation at 94°C for 30 s, annealing (46-55°C) for 30 s, and extension at 72°C for 30 s, with a final extension step at 72°C for 7 min. The PCR reaction was conducted using PureTaq-Ready-To-Go PCR Beads (GE Healthcare), 2 µl of DNA template, 21 µl of DNA grade H2O, and 1 µl of each direction primer listed in Table 3. We verified PCR products with a 1% agarose-TBE electrophoresis gel stained with CYBR Safe. For purification of the amplified products, we used Ampure Magnetic Beads (Beckman-Coulter) and re-suspended in 40 µl DNA grade water by using a Beckman Coulter Biomek NX 18 robot. Each 8 µl cycle-sequenced reaction mixture included 1 µl of Big Dye, 1 µl of Big Dye Terminating buffer, 1 µl of 3.2 pm primer, and 5 µl of gene amplification product. Cycle-sequenced products were purified with CleanSeq magnetic beads on a Biomex NX robot. Products were re-suspended in EDTA, and 33 µl were processed in an Applied Biosystems, Inc. Prism 3730xl automated DNA sequencer. These products were sequenced with the same primer pairs used for amplification at the Laboratorio de Secuenciación Genómica de la Biodiversidad, at Instituto de Biología and Unidad de Síntesis y Secuenciación de DNA, Instituto de Biotecnología, UNAM. The sequences were edited using Sequencher® version 5.4.6.
Table 3
List of primers used to amplify molecular markers.
Name
Sequence
Reference
12S rDNA
12SAI
AAACTAGGATTAGATACCCTATTAT
Kocher et al. (1989)
12SBI
AAGAGCGACGGGCGATGTGT
Kocher et al. (1989)
16S rDNA
16SA
CGCCTGTTTATCAAAAACAT
Simon et al. (1994)
16SB
CTCCGGTTTGAACTCAGATCA
Simon et al. (1994)
COI
HCO
TAAACTTCAGGGTGACCAAAAAATCA
Folmer et al. (1994)
LCO1
GGTCAACAAATCATAAAGATATTGG
Folmer et al. (1994)
28S rDNA
28SA
GACCCGTCTTGAAGCACG
Nunn et al. (1996)
28SBout
CCCACAGCGCCAGTTCTGCTTACC
Prendini et al. (2005)
Each genetic fragment was aligned separately for all terminals with MAFFT using the online server (https://mafft.cbrc.jp/alignment/server/). Since the number of nucleotides per gene was similar, we used the G-ins-i iterative refinement method, as recommended elsewhere (Katoh et al., 2019; Kuraku et al., 2013), and other parameters were kept default. To select the best fit of the substitution model per partition and conduct the phylogenetic analyses, we used IQ-TREE version 2 (Kalyaanamoorthy et al., 2017; Nguyen et al., 2015) and we estimated branch support with 1,000 replicates of the ultrafast bootstrap (UFBOOT) algorithm (Hoang et al., 2018). Furthermore, each genetic marker was analyzed individually to explore its phylogenetic signal and contribution to the final topology. We conducted concatenated and partitioned analyses, handling all matrices in Mesquite (Maddison & Maddison, 2023). For the COI partition we explored the best codon partition per site, but the results had no effect on the topology. Additionally, to evaluate each marker and nucleotide site within each marker, we calculated the gene (GCF) and site (SCF) concordance factors on the topology that we emphasized in the discussion (Mihn et al., 2020).
Distributional records and maps. We obtained records with geographical coordinates of the scorpion species treated here via the Global Biodiversity Information Facility (GBIF) from the InDRE, responsible in Mexico for epidemiological vigilance (Huerta-Jiménez, 2018), and the records published by Ponce-Saavedra et al. (2015). These records were the basis for the species distribution maps depicted in figures 3 to 6. We used the program QGIS 3.16.6-Hannove (QGIS, 2021) to create the distributional maps. The topological model with the data was from Jarvis et al. (2008), and to draw the political boundaries we used shapefiles obtained from Conabio. The biogeographic regionalization of Mexico into provinces and districts follows Morrone et al. (2017) and Morrone (2019).
Results
The gene fragments that we obtained are listed in Table 4 and the main statistics of the alignment and concatenated matrix are in Table 5. Our topology produced a clade representing members of the family Buthidae and another clade representing Vaejovidae (Fig. 2). Within buthids, the first clade included C. huichol and C. noxius, component species of the bertholdii species group (Ponce-Saavedra & Francke, 2019), supported by 95% UFBOOT, 100% GCF, and 40% SCF. The next clade included C. elegans, C. limpidus, and a putative undescribed species with lower support values of 68%, 100%, and 38%, respectively, including members of the elegans group. Although with low support, the bulk of species appeared in the third clade comprising species within the infamatus group, with C. tecomanus, C. infamatus, C. possanii, C. hirsuticauda, C. ornatus, and 2 putative new species. The last clade included C. suffusus and C. sculpturatus, whichPonce-Saavedra and Francke (2019) circumscribed within the infamatus and the elegans species group, respectively (Fig. 2). However, the overall topology retrieved herein is concordant with the North American clade of the genus Centruroides (Esposito & Prendini, 2019).
Table 4
Mitochondrial genetic markers 16S, COI, and 12S and nuclear 28S information for samples analyzed in the study. Dash (-) symbols indicate unavailable sequences.
Species
NCBI:txid
16S
12S
COI
28S
Centruroides elegans
217897
Cele_30_16S (PP295377)
Cele_30_12S (PP295301)
Cele_30_COI (PP356615)
Cele_30_28S (PP295328)
Cele_31_16S (PP295378)
Cele_31_12S (PP295302)
Cele_31_COI (PP355194)
Cele_31_28S (PP295329)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Centruroides hirsutipalpus
–
Chir_06_16S (PP295353)
Chir_06_12S (PP295277)
Chir_06_COI (PP356614)
Chir_06_28S (PP295310)
Centruroides huichol
2911785
Chui_38_16S (PP295385)
Chui_38_12S (PP295309)
Chui_38_COI (PP356613)
Chui_38_28S (PP295335)
Centruroides infamatus
42200
MF134694
–
MF134798
MF134763
Centruroides limpidus
29941
Clim_34_16S (PP295381)
Clim_34_12S (PP295305)
Clim_34_COI (PP356612)
Clim_34_28S (PP295332)
Centruroides noxius
6878
Cnox_36_16S (PP295383)
Cnox_36_12S (PP295307)
Cnox_36_COI (PP356611)
Cnox_36_28S (PP295333)
Cnox_37_16S (PP295384)
Cnox_37_12S (PP295308)
Cnox_37_COI (PP356610)
Cnox_37_28S (PP295334)
Centruroides ornatus
2338500
Corn_03_16S (PP295350)
Corn_03_12S (PP295274)
Corn_03_COI (PP355195)
Corn_03_28S (PP295324)
Corn_04_16S (PP295351)
Corn_04_12S (PP295275)
Corn_04_COI (PP356609)
Corn_04_28S (PP295325)
KY981895
KY981799
–
KY982086
MK479042
MK478991
MK479195
MK479144
Centruroides possanii
–
Cpos_10_16S (PP295357)
Cpos_10_12S (PP295281)
–
Cpos_10_28S (PP295314)
Cpos_07_16S (PP295354)
Cpos_07_12S (PP295278)
Cpos_07_COI (PP355196)
Cpos_07_28S (PP295312)
Cpos_08_16S (PP295355)
Cpos_08_12S (PP295279)
Cpos_08_COI (PP356608)
Cpos_08_28S PP295313
Cpos_09_16S (PP295356)
Cpos_09_12S (PP295280)
Cpos_09_COI (PP356607)
Cpos_09_28S (PP295323)
Centruroides sculpturatus
218467
Cscu_35_16S (PP295382)
Cscu_35_12S (PP295306)
Cscu_35_COI (PP356606)
Cscu_35_28S (PP295331)
Centruroides sp. 1
3103037
Csp1_23_16S (PP295370)
Csp1_23_12S (PP295294)
Csp1_23_COI (PP356604)
Csp1_23_28S (PP295311)
Centruroides sp. 2
Csp2_11_16S (PP295358)
Csp2_11_12S (PP295282)
Csp2_11_COI (PP356603)
Csp2_11_28S (PP295327)
Csp_14_16S (PP295361)
Csp_14_12S (PP295285)
Csp_14_COI (PP356605)
Csp_14_28S (PP295326)
Centruroides suffusus
6881
Csu_33_16S (PP295380)
Csu_33_12S (PP295304)
–
Csu_33_28S (PP295330)
Centruroides tecomanus1
1028682
Cte1_12_16S (PP295359)
Cte1_12_12S (PP295283)
Cte1_12_COI (PP356602)
Cte1_12_28S (PP295315)
Cte1_17_16S (PP295364)
Cte1_17_12S (PP295288)
Cte1_17_COI (PP355197)
Cte1_17_28S (PP295320)
Cte1_18_16S (PP295365)
Cte1_18_12S (PP295289)
Cte1_18_COI (PP356601)
Cte1_18_28S (PP295318)
Centruroides tecomanus2
Cte2_13_16S (PP295360)
Cte2_13_12S (PP295284)
Cte2_13_COI (PP356600)
Cte2_13_28S (PP295316)
Cte2_15_16S (PP295362)
Cte2_15_12S (PP295286)
Cte2_15_COI (PP355198)
Cte2_15_28S (PP295317)
Table 4. Continued
Species
NCBI:txid
16S
12S
COI
28S
Cte2_19_16S (PP295366)
Cte2_19_12S (PP295290)
Cte2_19_COI (PP356599)
Cte2_19_28S (PP295319)
Cte2_20_16S (PP295367)
Cte2_20_12S (PP295291)
Cte2_20_COI (PP356598)
Cte2_20_28S (PP295321)
Cte2_21_16S (PP295368)
Cte2_21_12S (PP295292)
Cte2_21_COI (PP356597)
Cte2_21_28S (PP295322)
Centruroides tecomanus
MF134695
–
MF134799
MF134757
MK479053
MK479002
MK479206
MK479156
Mesomexovis sp.
–
Mesp_22_16S (PP295369)
Mesp_22_12S (PP295293)
Mesp_22_COI
Mesp_22_28S (PP295337)
Mesomexovis occidentalis
1532992
KM274362
KM274216
KM274800
–
Mesomexovis spadix
1532994
KM274221
KM274367
KM274805
KM274659
Mesomexovis subcristatus
1532995
KM274368
KM274222
KM274806
KM274660
Thorellius cristimanus
1533000
Tcri_01_16S (PP295348)
Tcri_01_12S (PP295272)
–
Tcri_01_28S (PP295338)
Tcri_16_16S (PP295363)
Tcri_16_12S (PP295287)
–
Tcri_16_28S (PP295336)
Tcri_02_16S (PP295349)
Tcri_02_12S (PP295273)
–
Tcri_02_28S (PP295339)
Tcri_05_16S (PP295352)
Tcri_05_12S (PP295276)
–
Tcri_05_28S (PP295340)
KM274420
KM274274
KM274858
KM274712
KM274422
KM274276
KM274860
KM274714
Thorellius intrepidus
1533001
Tint_24__16S (PP295371)
Tint_24_12S (PP295295)
Tint_24_COI (PP355193)
Tint_24_28S (PP295341)
Tint_25_16S (PP295372)
Tint_25_12S (PP295296)
Tint_25_COI (PP356616)
Tint_25_28S (PP295342)
Tint_26_16S (PP295373)
Tint_26_12S (PP295297)
Tint_26_COI (PP356617)
Tint_26_28S (PP295343)
Tint_27_16S (PP295374)
Tint_27_12S (PP295298)
Tint_27_COI (PP355192)
Tint_27_28S (PP295344)
Tint_28_16S (PP295375)
Tint_28_12S (PP295299)
Tint_28_COI (PP356618)
Tint_28_28S (PP295345)
Tint_29_16S (PP295376)
Tint_29_12S (PP295300)
Tint_29_COI (PP356619)
Tint_29_28S (PP295346)
KM274424
KM274278
KM274862
–
KM274425
KM274279
KM274863
KM274717
Vaejovis sp.
–
Vasp_32_16S (PP295379)
Vasp_32_12S (PP295303)
Vasp_32_COI (PP356620)
Vasp_32_28S (PP295347)
Vaejovis carolinianus
33322
KM274289
KM274143
KM274727
KM274581
Vaejovis pequeno
1532951
KM274293
KM274147
KM274731
KM274585
Vaejovis rossmani
1532952
KM274294
KM274148
KM274732
KM274586
Table 5
Main statistics of site information per alignment, including parsimony informative sites, (P info) and the concatenated (Conca) alignment of all partitions in a final matrix. The concatenated alignment had 6% missing data.
Partitions
Terminals/ nucleotide
Site information per alignment
Substitution
P info
Invariable
Unique
Constant
Model
16S
53/523
209
285
258
285
K3Pu+F+I+G4
28S
52/538
39
494
96
494
K2P+I
12S
51/393
153
210
212
210
TPM2+F+G4
COI
45/679
220
420
220
420
TIM2+F+G4
Conca
53/2133
621
1,409
786
1,409
simultaneous
Figure 2. Phylogenetic tree from an analysis of 4 concatenated genetic markers, 3 mitochondrial (12S, 16S, and COI) and 1 nuclear (28S). The topology shows families, subfamilies, and species groups. Numbers beside nodes indicate ultrafast bootstrap/genetic concordance factor/site concordance factor. Colored species represent the 11 species found in our collection within the study area and samples grouping with them distributed inside or outside Colima. Scorpion photos: upper right, Centruroides tecomanus from El Palapo, adult male; middle, Centruroides hirsutipalpus from Sierra de Minatitlán, adult female; lower left, Thorellius intrepidus from El Palapo, adult male.
The Vaejovid clade received full support, except SCF (60%), which included 3 genera: Vaejovis, Mesomexovis, and Thorellius (Fig. 2). The genera occurred in a topology that overall resembles that of González-Santillán and Prendini (2013, 2015); while Vaejovis is a genus within Vaejovinae, Mesomexovis and Thorellius are part of the Syntropinae subfamily. Within Syntropinae, only Thorellius was monophyletic, and T. intrepidus received full support. Additionally, we identified 2 putative new species belonging to the genera Vaejovis and Mesomexovis.
Distributional maps and species geographical ranges. We mined 6,965 records of Mexican scorpions from the InDRE database but retained only 1,000 of the species treated herein. We filtered the records using previously published identified species by specialists and the distribution of the species cited for Colima (González-Santillán & Prendini, 2013, 2018; Lourenço & Sissom, 2000; Ponce-Saavedra et al., 2016; Sissom, 2000).
Figure 3. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides elegans (circles), records north of Sierra de Manantlán may be misidentifications; B, distribution of Centruroides tecomanus (circles), records in Guerrero (GRO), Guanajuato (GTO), Jalisco (JAL), and Nayarit (NAY) may be misidentifications; Centruroides tecomanus 1 green cross, and Centruroides tecomanus 2 pink crosses. State abbreviations: AUG, Aguascalientes; COL, Colima; MEX, Estado de México; QRO, Querétaro; SLP, San Luis Potosí; ZAC, Zacatecas.
Figure 4. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database and for B additional records from Ponce-Saavedra et al. (2015). A, Distribution of Centruroides infamatus (circles); B, distribution of Centruroides ornatus (circles), our sample in Colima (cross).
Figure 5. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Centruroides hirsutipalpus (red cross), Centruroides sp. 1 (blue cross), Centruroides sp. 2 (pink cross), and Centruroides possanii (green crosses); B, distribution of Mesomexovis aff. occidentalis sequences in this work and a sample from Chamela Jalisco recorded by González-Santillán (2004), conspecific with our samples in green crosses, Vaejovis sp. (mexicanus group) (pink cross).
The distribution of Centruroides species appears to be restricted by physiographical elements. Centruroides elegans and C. tecomanus are restricted to the coastal lands of Jalisco and Colima, respectively (Fig. 3). The Sierra Madre del Sur appears to be the main barrier to the north of their distribution. The records of these species overlap broadly with a visible gap created by the Sierra de Minatitlán (Fig. 3B) either by subsampling or by an effective geographic barrier.
Centruroides ornatus and C. infamatus, on the other hand, are restricted to the Transmexican Volcanic Belt (TVB) (Fig. 4). While the former species has an entire distribution within this province (Fig. 4B), the latter seems to be distributed in patches, one to the north, along the border of the Chihuahuan desert and the TVB, and the second to the south, between the Balsas Depression and the TVB (Fig. 4A).
Centruroides possanii appears to be a microendemic scorpion species component of the fauna restricted to a massive karstic lone mountain, Cerro Grande (González-Santillán et al., 2019). Two putative undescribed species of Centruroides occupy the extreme east and west of the coastal line of Colima, and C. hirsutipalpus appears restricted to the Sierra Minatitlán (Fig. 5A).
Of the vaejovid species, thus far, the only records for Mesomexovis sp. are in Colima, albeit there is one conspecific record in Chamela, Jalisco (González-Santillán, 2004). The putatively undescribed species of Vaejovis appears restricted to Cerro Grande (Fig. 5B), like Centruroides possanii, potentially another microendemic species. Thorellius intrepidus and T. cristimanus are widely distributed within Colima and across the TVB, the Balsas Depression, and the Sierra Madre del Sur (Fig. 6).
Discussion
Of the 2 families found in Colima, Buthidae comprise 7 species of Centruroides,becoming the most diverse (Table 1, Fig. 2). Our multigene analyses suggest that the previously recorded species C. elegans, C. infamatus, and C. limpidus reported by Ponce-Saavedra et al. (2016) might not be part of the scorpion fauna of Colima, as we demonstrate in the following sections.
Figure 6. Map of the west coast of Mexico with georeferenced filtered records from the InDRE database. A, Distribution of Thorellius intrepidus greensquares, our samples in Colima pink crosses; B, distribution of Thorellius cristimanus greensquares, our samples in Colima pink crosses.
Centruroides elegans and C. limpidus were not found in Colima. Centruroides elegans has an obscure taxonomic history. Firstly, its original description is too general and never indicated a precise type locality, but “Mexico” (Fet & Lowe, 2000). Secondly, although some taxonomic works clarified its former subspecies, the nominal taxon identity of C. elegans remains ambiguous. While Lourenço and Sissom (2000) suggested that this species is distributed in Jalisco, later, González-Santillán (2004) concluded that its distributional limits have never been defined with precision. The 2 exemplars of C. elegans collected in Chamela, Jalisco, grouped with members of the elegans group, sisters to C. limpidus from Iguala, Guerrero, and these in turn, were sister to our 2 exemplars identified as Centruroides new sp. 2 11 and 14 from La Central, in the municipality of Manzanillo (Figs. 2). The distributional data from InDRE of C. elegans is limited right at the northern border of Colima, except for 9 records away from that geographic barrier (Fig. 3A), which we hypothesized as a misidentification due to their position outside the Pacific coastline. Our fieldwork produced no sample conspecific to C. elegans but did produce other genetically distant exemplars. Until denser sampling along the northern border of Colima is conducted, we conclude that C. elegans and by corollary C. limpidus might not be part of the scorpion fauna distributed in Colima as suggested by Ponce-Saavedra and Francke (2013) and Ponce-Saavedra et al. (2016).
Centruroides infamatus may not be part of the Colima scorpion fauna. Centruroides infamatus is another obscure taxon with an ambiguous distributional pattern from published data. Despite its inclusion in phylogenetic analyses (Quijano-Ravel et al., 2019; Towler et al., 2001), its taxonomic circumscription and distribution have never been clarified. Among other problems, the original description indicates “type locality unknown” (Fet & Lowe, 2000). Hoffmann (1932) studied the scorpions from Mexico and realized that the specimens from Michoacán agreed with the original description of the species and proposed that its distribution was expanded to Central Mexico from the Pacific Coast in Sinaloa and Colima to Guanajuato. We included 2 exemplars of C. infamatus, 1 from León, Guanajuato (C. infamatus 15 scrp), and 1 terminal from Tandamangapio municipality, Michoacán (C. infamatus LP1822; Esposito et al., 2018). However, the terminal LP1822 grouped with exemplars of C. ornatus (Fig. 2) and Tandamangapio is approximately 16 km from Sahuayo and Jiquilpan, 2 localities recorded in the redescription of C. ornatus (Ponce-Saavedra et al., 2015).
We plotted 612 records from the InDRE database of C. infamatus to compare the distribution of C. ornatus (Fig. 4A). Although northern Michoacán is almost exclusively occupied by C. ornatus, Jalisco and Michoacán present an overlap with C. infamatus (Fig. 4). Additionally, the InDRE database contains records of C. infamatus from Oaxaca, Puebla, Nayarit, Durango, and Sinaloa (not shown), most likely occupied by other species, implying that most records are misidentifications. The species that morphologically could be confused with C. infamatus in the states of Oaxaca and Puebla are C. baergi, C. nigrovariatus, or C. rodolfoi because the overall base color, body size and carapace pigmentation are similar among the species. The same morphological features are present in specimens from Nayarit and Durango, however, the identity of the populations in these two states have never been studied with a molecular approach and may represent a geographically distinct species.
Nevertheless, recently Ponce-Saavedra et al. (2022) described C. baldazoi for Sinaloa, related morphologically with C. infamatus, potentially a species with which it can be mistaken. Although Ponce-Saavedra and collaborators included C. infamatus in the distribution of Sinaloa and Colima, they failed to indicate precise localities. In summary, the distribution of C. infamatus remains unsolved, attested by the wide distribution reported in the InDRE database. Thus, the infamatus species complex requires comprehensive molecular analyses to delimit its taxonomic circumscription and geographical distribution. On the other hand, our samples of C. ornatus 3 and 4 grouped with C. ornatus LP1822 and with C. ornatus 2003 from Morelia, Michoacán (Esposito & Prendini, 2019), forming a monophyletic group (Fig. 2). Since the samples LP1822 and 2003 lie within the area of C. ornatus proposed by Ponce-Saavedra et al. (2015), we concluded that our exemplars are conspecific with C. ornatus and expanded the distribution southwards, drawing a distributional border in northern Colima (Fig. 4B).
In conclusion, we propose that C. infamatus might not be present in Colima, but further sampling and analyses are needed. Despite the morphological similarity between C. infamatus and C. ornatus, our molecular analysis indicates genetic differences and may present a distinctive distributional pattern yet to be drawn with more samples.
Centruroides tecomanus is a species complex. Due to their morphological similarity with C. limpidus, C. tecomanus was described as C. limpidus tecomanus (Hoffmann, 1932), and the author delimited C. tecomanus distribution to the lowlands of Colima and surmised that its distribution extended to the south along the coastline of Michoacán. But, most importantly, Hoffmann mentioned that on the northern coastline, C. elegans substitutes C. tecomanus. In contrast, the distribution recorded in the InDRE database for C. tecomanus implies that these 2 species present a wide area of sympatry on the coast of Jalisco (Fig. 3).
Ponce-Saavedra et al. (2009) proposed C. tecomanus as a “bona fide species” and assumed its distribution includes the coastline of Michoacán, following Hoffmann (1932). In their molecular and morphological analyses, the authors never included exemplars from Tecomán, Colima, the type locality for the species, assuming that the specimens from Michoacán were conspecific with the populations of Tecomán. Furthermore, Quijano-Ravell et al. (2010) extended the distribution of C. tecomanus to 4 localities in Guerrero and a similar number in Jalisco in a montane area.
The phylogenetic tree presented here substantiates 2 clades of C. tecomanus within the monophyletic infamatus species group (Fig. 2). We hypothesize that these clades may represent potentially distinct sympatric species, emphasizing that our topology obtained full support for these clades with multiple samples. Centruroides tecomanus2 appears to be more common and widely distributed in Colima, whereas C. tecomanus1 is less common, with only 2 localities grouped with Centruroides tecomanus 2007 from Michoacán and from the municipality of Comala, Colima, Centruroides tecomanus (Fig. 3B). Our results suggest that the populations distributed in Michoacán may represent a cryptic, undescribed species; consequently, the exemplars reported in Guerrero (Quijano-Ravell et al., 2010) are unlikely to be conspecific with C. tecomanus. The authors’ discovery of new populations in Jalisco and Guerrero were based entirely on their analyses of morphological characters, precisely the most common way of confusing cryptic species.
Considering that the montane border between Colima and Michoacán is occupied by Centruroides romeroi Quijano-Ravell, de Armas, Francke, Ponce-Saavedra, 2019, it is fair to assume that C. tecomanus1 inhabits coastal ranges following the coastline of Michoacán to the Lázaro Cárdenas delta, where it may be substituted by Centruroides bonito Quijano-Ravell, Teruel, Ponce-Saavedra, 2016 or even other undescribed species. The Balsas River has been proposed to be a geographical barrier for several epigean arachnids, such as Amblypygi and Theraphosidae (Mendoza & Francke, 2017; Schramm et al., 2021) and for small mammals (Ruiz-Vega et al., 2018)
Noteworthy is the locality La Central, right on the border between Colima and Jalisco, a few kilometers southeast of the Marabasco River (Fig. 1A), where we collected 3 putatively different species of Centruroides, the 2 morphotypes of C. tecomanus and Centruroides sp. 2, retrieved within the elegans group (Fig. 2). These findings illustrate the complicated patterns of diversification within this buthid genus in Colima. From a taxonomic perspective, if the identity of C. tecomanus is to be clarified, it is now imperative to analyze molecularly exemplars from Tecomán, Colima, which is the type locality of this species.
Thorellius species exhibit a more restricted distribution in Colima. Thorellius intrepidus and T. cristimanus are widely distributed in several states of the Pacific Lowlands (Fig. 6). However, in a recent revision of the genus, 2 species were described, Thorellius wixarika González-Santillán and Prendini, 2018 and Thorellius tekuani González-Santillán and Prendini, 2018 (Fig. 3 of González-Santillán and Prendini, 2018), that are relevant to these analyses. The former occupies the northwestern territory in Nayarit and Jalisco,whereas the latter inhabits the Balsas Depression of Estado de México, Guerrero, and Michoacán. This observation suggests that the InDRE database contains several misidentifications of both T. intrepidus and T. cristimanus. In fact, the T. wixarika and T. tekuani description was in 2018, and the InDRE database is several years older, hence the complete absence of records. Thus, T. intrepidus inhabits Aguascalientes, Colima, Guanajuato, Jalisco, and Michoacán (Fig. 6A); and T. cristimanus, Colima and Jalisco (González-Santillán & Prendini, 2018) (Fig. 6B). From an ecological point of view, we noticed that the InDRE database only has records with elevations below 700 m, implying that these species prefer tropical to subtropical climates in Colima.
González-Santillán (2004) reported Mesomexovis aff. occidentalis as a putative new species for the Biological Station Chamela, Jalisco. Furthermore, González-Santillán and Prendini (2015) conducted a phylogenetic analysis using morphology and mitochondrial and nuclear DNA, resulting in a topology that suggested that this species was not conspecific to Mesomexovis occidentalis, as Williams (1986)identified it. López-Granados (2019) found further morphological evidence to separate this species, but the evidence was never published. The importance of that work is that, for the first time, exemplars of Mesomexovis from Colima that were conspecific with our samples were included in a phylogenetic analysis. Once more, we demonstrate that Mesomexovis sp. 22 is not conspecific with M. occidentalis using molecular evidence, and it requires a formal separation and description (Fig. 2). Unlike Thorellius species in Colima, Mesomexovis is a widespread species inhabiting tropical to montane habitats with a wider range of elevation (Fig. 5B).
The mexicanus group was recently revisited in a monograph with the delimitation of other species groups and the description of 5 new species (Contreras-Félix & Francke, 2019). The paucity of sequences for the mexicanus group in the NCBI database only permitted retrieval of loci for Vaejovis rossmani LP 2027, a species inhabiting the Sierra Madre Oriental in the states of Tamaulipas and Nuevo León and treated in the Contreras-Félix and Francke (2019) monograph. Our analysis retrieved Vaejovis sp. 32 and Vaejovis rossmani LP 2027 together, which suggests membership of the mexicanus group of Vaejovis. Furthermore, Vaejovis sp. 32also matches the morphological diagnosis presented in Contreras-Felix and Francke (2019). Contreras-Félix et al. (2023) recorded Vaejovis santibagnezi Contreras-Felix and Francke, 2019 in Cerro Grande, where we collected our samples (Fig. 5B). However, the authors failed to present morphological evidence to justify this conclusion. We compared our specimens with the geographically closest V. monticola deposited in the CNAN and with V. santibagnezi and found significant morphological differences. Currently, we are preparing a contribution where we propose the description of a new species. Following this idea, we are inclined to think that, like C. possanii, Vaejovis sp. 32 is also microendemic because of its limitation to disperse throughout tropical valleys from the Cerro Grande massif to other mountain ranges. Finally, we submit that such an evident geographical barrier may apply to several epigean, non-volant arthropods such as the scorpions.
Finally, 2 species absent in our fieldwork are Konetontli ilitchi González-Santillán and Prendini 2015 and Vaejovis janssi Williams, 1980. The latter is endemic to the Socorro Island, part of the Revillagigedo Archipelago (Williams, 1980), and K. ilitchi has been found only inside a cave in the vicinity of Coquimatlán, Colima (González-Santillán & Prendini, 2015). These 2 features in the distribution and biology of the species hindered the possibility of collecting and studying them. Future fieldwork may illuminate the taxonomy and distribution of these elusive species. Finally, Vaejovis monticola, another high elevation dweller of the mexicanus species group (Contreras-Felix & Francke 2019) was absent in our collecting trips. Although its type locality cited by Contreras-Felix and Francke (2019) indicates: “Jalisco, northern side of Nevado de Colima” we submit that it may not be on the southern side in Colima, but extensive fieldwork is needed to confirm this hypothesis.
Biogeographical and ecological considerations
There is strong evidence that the ancestors of the North American Centruroides originated in Gondwana and have dispersed to and diversified in that territory via land bridges, vicariance, and rafting over the Atlantic Ocean, over 50 to 20 Mya (Esposito & Prendini, 2019). However, the movement of this genus towards boreal latitudes within North America is not well understood. Although one could suppose that the colonization of such territories appears to be in pulses at different periods due to the overlap in the distribution of the elegans and infamatus species groups, it is beyond the scope of our study to establish a coherent explanation; besides, our data are incomplete to that end. One startling discovery is that the Colima scorpiofauna comprises 7 distinctive, at least genetically, species of Centruroides (Fig. 2). What biotic, abiotic, or ecological factors produce such species diversity? Is it a combination of all these factors? Is it a case of sympatric speciation? Morphologically, it is sometimes difficult to distinguish some of these species. For instance, the 3 species found in La Central were identified initially as C. tecomanus because they exhibit no morphological variation, at least in traditional characters shown in identification keys (Ponce-Saavedra et al., 2016). It is fundamental to conduct morphological studies with this scorpion community to obtain independent information to precisely delimit these species. Unlike other scorpion groups, Centruroides exhibit conservative morphology. For instance, if we compared Centruroides species to species of the Syntropinae subfamily, we find that syntropines are diverse in ecomorphotypic adaptations that promoted morphological diversification (González-Santillán & Prendini, 2013), while in Centruroides, the persistent errant lifestyle maintains morphology without significant variation. It is astonishing to observe, for instance, C. tecomanus living at sea level in tropical deciduous forests and compare it to C. possanii inhabiting pine-oak forests above 2,000 m elevation; as though, the same “Centruroides bauplan” can survive diametrically different environments following a persistent lifestyle. This final assertion leads us to think that the adaptative changes are in the physiology rather than in the morphology of Centruroides.
Thorellius and Mesomexovis (Syntropinae) are substrate-specialists, since the body plan, more robust and armed with setae and spinules on the legs, allow these species to scrape and dig galleries on clayey, fine-grained substrates (González-Santillán & Prendini, 2013, 2018). Such adaptations come with a drawback because the kind of soil restricts the presence of these scorpions; for instance, rocky or shallow soils represent unhospitable habitats for these species. Like other species in the mexicanus group, Vaejovis sp. 32 and Vaejovis monticola Sissom, 1989 are inhabitants of montane habitats within the intricate topography of Colima.
In summary, the scorpion assemblage distributed in Colima includes errant species such as Centruroides without exaggerated adaptations; pelophilous/lapidicolous species such as Thorellius and Mesomexovis, powerful diggers, commonly found underneath rocks or other debris; and scorpions that can be found inside the leaf litter or under nooks of rocks or trees, such as Vaejovis sp. However, it is enigmatic why the Diplocentridae family, well-represented in Mexico, has not been found in Colima. Only in Colima, out of the 3 relatively small states in Mexico, the other 2 being Morelos (Santibáñez-López et al., 2011) and Aguascalientes (Chávez-Samayoa et al., 2022), where similar scorpiofaunistic surveys have been conducted, the genus Diplocentrus is absent. From a biogeographical point of view, if this family is absent in Colima, it is not trivial and requires further investigation.
As a final remark, we hope we have accomplished our aim of urging arachnologists to examine the benefits of local inventories and the potential use of barcodes at the forefront of discovering and documenting local arachnid diversity.
Medical and pharmacological relevance of some scorpions of Colima
From a medical point of view, it is of utmost importance to identify the species of scorpions in an area and to be able to distinguish hazardous from harmless species. Colima is one of the states with the highest incidence rates of mortality and morbidity caused by intoxication by scorpion sting (ISS) (Chowell et al., 2006; González-Santillán & Possani, 2018). By the 1940s, Colima was at the forefront of ISS and deaths in Mexico. However, with the introduction of a safe and effective antivenom in the 1970s, mortality has diminished, although morbidity is still high (González-Santillán & Possani, 2018).
Due to the medical relevance of scorpionism in Mexico, the venom of some buthid scorpions distributed in Colima has been studied extensively (Table 6). We know that mammalian sodium scorpion toxins (NaScTx) are the chief and sometimes most abundant compound responsible for the toxicity of scorpion venom, although potassium, chlorine, and calcium-gated channels, among others, are also affected by toxic peptides (Cid-Uribe et al., 2019; González-Santillán & Possani 2018). Until now, C. hirsutipalpus, C. ornatus, C. possanii, and C. tecomanus have been studied with a proteomic or transcriptomic approach, demonstrating that these venoms are powerful enough to decimate humans (García-Guerrero et al., 2020; García-Villalvazo et al., 2023, Valdez-Velázquez et al., 2016, 2018). Centruroides tecomanus venom contains at least 13 mammalian toxic compounds (Valdez-Velázquez et al., 2016), of which Ct1a is the main mammalian sodium scorpion toxin component, with a molecular weight of 7,591 Da (Martin et al., 1988). The proteomic analysis of C. ornatus toxic components identified 3 major mammalian NaScTx, CO1, CO2, and CO3, with molecular weights of 7,561.2, 7,614.3, and 7,774.9 Da, respectively (García-Guerrero et al., 2020). On the other hand, the published mass fingerprint of C. hirsutipalpus venom has 4 compounds with similar molecular weight to C. tecomanus peptides, suggesting that these species may have similar or identical NaScTx (Valdez-Velázquez et al., 2018). One of the shared compounds found in C. hirsutipalpus corresponds to the molecular weight of Ct1a (Valdez-Velázquez et al., 2013), which means that Ct1a could be responsible for, or at least contribute to, the high toxicity of this species. Although the molecular components with a weight corresponding to the toxic peptides CO1, CO2, and CO3 are not present in C. hirsutipalpus venom, the mass fingerprint of C. tecomanus reported a peptide with similar molecular weight to CO2 toxin present in C. ornatus (Valdez-Velázquez et al., 2013). Our phylogenetic results show that these species belong to the infamatus group, and it is notable that the molecular weight among some toxins is similar. This observation opens the possibility that the toxin diversity of the mammalian NaScTx may have been inherited from a common ancestor. We realize, however, the need for a more in-depth study to unveil the relationship among these toxic peptides to test such a hypothesis. In contrast, a C. possanii crude venom proteomic study resulted in the identification of 18 NaScTx, of which, CpoNatBet09 was identical to Cll2b and Cii1 from C. limpidus and C. infamatus, respectively (Valdez-Velázquez et al., 2013). This example allows us to present the counterpart of the previous one. Centruroides limpidus in our topology grouped with members of the elegans group and C. possanii within the infamatus group (Fig. 2). We can conclude that the exact sequence match between CpoNatBet09 and Cll2b is most likely due to a selective force that produced convergent evolution. Alternatively, this is a deeper inheritance within the ancestors of the infamatus and elegans group.
The comparisons and hypotheses about venom similarities and species assume that all scorpions used to conduct the experiments to obtain the crude venom were correctly identified. However, the discovery of putative species of Centruroides might question some of these results, particularly those of C. tecomanus. The first author has been collaborating to identify most samples of species used in some of the recently published investigation on venom research (Cid-Uribe et al., 2019; García-Guerrero et al., 2020; Romero-Gutiérrez et al., 2017, Valdez-Velázquez et al., 2018, among others). And in some cases, voucher specimens have been deposited in the CNAN, therefore the taxonomic identity for C. hirsutipalpus, C. ornatus, C. possanii can be readily corroborated.
Table 6
Scorpion species with reported LD50 tested on mice at μg/20g mouse weight scale with the named, most abundant, and likely responsible for high toxicity sodium toxins found in the scorpion species of Colima.
Species
LD50
NaScTx
Autor
Centruroides hirsutipalpus Ponce-Saavedra and Francke, 2009
11.7 ± 1.9
26 β-NaTx and 5 α-NaTx (proteome) 71 β-NaTx and 16 α-NaTx (transcriptome)
Valdez-Velázquez et al. (2013, 2016), Ramírez et al. (1988)
Thorellius cristimanus Pocock, 1902
N/A
10 β-NaTx and 3 α-NaTx (transcriptome)
Romero-Gutiérrez et al. (2017)
Because vaejovid scorpions exhibit lower toxin potency, proteomic studies are uncommon. However, the study of the crude venom of Thorellius cristimanus under a transcriptomic and proteomic approachidentified160 potential venom peptides (Romero-Gutiérrez et al., 2017). The authors found a great diversity of channel toxins targeting potassium and calcium ion-channels, numerous enzymes, and NDBP. Nevertheless, none of these toxins appear to affect mammals to endanger their life. Noteworthy are the NDBP compounds with antimicrobial activity (Almaaytah et al., 2014), which potentially may be a source for developing novel commercial antibiotics. More recently, Ibarra-Vega et al. (2023) reported finding the neurotransmitter serotonin and 2 derived or intermediate indoles of serotonin: N-methylserotonin, and bufotenidine. Although serotonin has been reported in other scorpions and the sea snail genus Conus, the other indoles represent the first report in scorpions. The authors proposed that the 3 components are involved in defense and probably prey submission because they produce extreme pain in the victim. Furthermore, N-methylserotonin and bufotenidine showed a similar affinity for serotonin cellular receptors, implying a role in the effect of this neurotransmitter, and thus, it showed promising medical applications.
Integrating taxonomic, distributional, and in our case venom diversity within local scorpion faunas is an interdisciplinary topic uncommon in the literature (Brito & Borges, 2015; Cao et al., 2014), although this exercise has been done in Mexico before (Santibáñez-López et al., 2015). This multidisciplinary work took advantage of a reciprocal illumination exercise. This short synthesis of the diversity of venoms of scorpion of Colima revealed the potential of toxins to be part of the evidence to identify and delimit species (Schaffrath et al., 2018). Moreover, the synthesis provided a glance of the evolution of venoms not only for the medically prominent species of Centruroides but also for vaejovid scorpion species. From the experimental discipline, we learned that it is paramount to maintain vouchers after biochemical or physiological experiments to track down the original material and keep up with the dynamics of taxonomy, a tool requiring empirical information to permit the identification of species to validate past, present, and future discoveries in other biological disciplines.
Acknowledgments
We thank the University of Colima students and Josué López Granados (Facultad de Ciencias, UNAM) for their assistance in collecting scorpions throughout the state of Colima. JICU thanks Conahcyt for granting a postdoctoral scholarship (512560). Collection permits granted by Semarnat included SGPA/DGVS/12063/15, SGPA/DGVS/02139/2022, and FAUT-0305. Finally, we are grateful to the Associate Editor and two reviewers for improving an early version of the manuscript, particularly to Andrés Ojanguren-Afilastro for his critical reading.
Baldazo-Monsivaiz, J. G., Ponce-Saavedra, J., & Flores-Moreno, M. (2012). Los alacranes (Arachnida: Scorpionida) de importancia médica del estado de Guerrero, México. Dugesiana, 19, 143−150.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., & Canché-Aguilar, I. (2017). Los escorpiones (Arachnida: Scorpiones) del municipio de Chilpancingo de los Bravo, estado de Guerrero, México Entomología Mexicana, 4, 21−27.
Baldazo-Monsivaiz, J. G., Teruel, R., Cortés-Guzmán, A. J., Sánchez-Arriaga, J., López-Flores, M., Reyes-Castelán, A. et al. (2016). Los escorpiones (Arachnida: Scorpiones) del municipio de Taxco de Alarcón, del estado de Guerrero, México. Entomología Mexicana, 3, 75−80.
Bolaños, L. M., Rosenblueth, M., de Lara, A. M., Migueles-Lozano, A., Gil-Aguillón, C., Mateo-Estrada, V. et al. (2019). Cophylogenetic analysis suggests cospeciation between the Scorpion Mycoplasma Clade symbionts and their hosts. Plos One, 14, e0209588 https://doi.org/10.1371/journal.pone.0209588
Brito, G., & Borges, A. (2015). A checklist of the scorpions of Ecuador (Arachnida: Scorpiones), with notes on the distribution and medical significance of some species. Journal of Venomous Animals and Toxins Including Tropical Diseases, 21, 1–23. https://doi.org/10.1186%2Fs40409-015-0023-x
Cao, Z., Di, Z., Wu, Y., & Li, W. (2014). Overview of scorpion species from China and their toxins. Toxins, 6, 796–815. https://doi.org/10.3390/toxins6030796
Cid-Uribe, J. I., Meneses E. P., Batista, C. V. F., Ortiz, E., & Possani, L. D. (2019). Dissecting toxicity: The venom gland transcriptome and the venom proteome of the highly venomous scorpion Centruroides limpidus (Karsch, 1879). Toxins, 247, 1−21. https://doi.org/10.3390/toxins11050247
Contreras-Félix, G. A., & Francke, O. F. (2019). Taxonomic revision of the “mexicanus” group of the genus Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae). Zootaxa, 4596, 1−100. https://doi.org/10.11646/zootaxa.4596.1.1
Contreras-Félix, G. A., del Pozo, O. G., & Navarrete-Heredia, J. L. (2023). A new species of Vaejovis from the mountains of west Mexico (Scorpiones: Vaejovidae). Dugesiana, 30, 229−245. https://doi.org/10.32870/dugesiana.v30i2.7306
Chávez-Samayoa, F., Díaz-Plascencia, J. E., & González-Santillán, E. (2022). Two new species of Vaejovis (Scorpiones: Vaejovidae) belonging to the mexicanus group from Aguascalientes, Mexico, with comments on the homology and function of the hemispermatophore. Zoologischer Anzeiger, 298, 148–169. https://doi.org/10. 1016/j.jcz.2022.04.005
Chowell, G., Díaz-Dueñas, P., Bustos-Saldaña, R., Alemán-Mireles, A., & Fet, V. (2006). Epidemiological and clinical characteristics of scorpionism in Colima, Mexico (2000-2001). Toxicon, 47, 753–758. https://doi.org/10.1016/j.toxicon.2006.02.004
Esposito, L. A., & Prendini, L. (2019). Island ancestors and New World biogeography: a case study from the scorpions (Buthidae: Centruroidinae). Scientific Reports, 9, 3500. https://doi.org/10.1038/s41598-018-33754-8
Fet, V., Graham, M. R., Blagoev, G., Karataş, A., & Karataş, A. (2016). DNA barcoding indicates hidden diversity of Euscorpius (Scorpiones: Euscorpiidae) in Turkey. Euscorpius, 216, 1–13.
Fet, V., Graham, M. R., Webber, M. M., & Blagoev, G. (2014). Two new species of Euscorpius (Scorpiones: Euscorpiidae) from Bulgaria, Serbia, and Greece. Zootaxa, 3894, 83–105. https://doi.org/10.11646/zootaxa.3894.1.7
Fet, V., & Lowe G. (2000). Family Buthidae C. L: Koch, 1837. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 54–286). New York: New York Entomological Society.
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294–299.
García-Guerrero, I. A., Cárcamo-Noriega, E., Gómez-Lagunas, F., González-Santillán, E., Zamudio, F. Z., Gurrola, G. B. et al. (2020). Biochemical characterization of the venom from the Mexican scorpion Centruroides ornatus, a dangerous species to humans. Toxicon, 15, 27–38. https://doi.org/10.1016/j.toxicon.2019.11.004
García-Villalvazo, P. E., Jiménez-Vargas, J. M., Lino-López, G. J., Meneses, E. P., Bermúdez-Guzmán, M. J., Barajas-Saucedo, C. E. et al. (2023). Unveiling the protein components of the secretory-venom gland and venom of the scorpion Centruroides possanii (Buthidae) through omic technologies. Toxins, 15, 1–20. https://doi.org/10.3390/toxins15080498
Goodman, A. M., Prendini, L., & Esposito, L. A. (2021). Systematics of the arboreal Neotropical ‘thorellii’ clade of Centruroides bark scorpions (Buthidae) and the efficacy of mini-barcodes for museum specimens. Diversity, 13, 441. https://doi.org/10.3390/d13090441
Goodman, A. M., Prendini, L., Francke, O. F., & Esposito, L. A. (2021). Systematic revision of the arboreal neotropical “thorellii” clade of Centruroides Marx, 1890, bark scorpions (Buthidae C. L. Koch, 1837) with descriptions of six new species. Bulletin of the American Museum of Natural History, 452, 1–92.
González-Santillán, E. (2004). Diversidad, taxonomía y hábitat de alacranes. In A. N. García, & R. Ayala (Eds.), Artrópodos de Chamela (pp. 25–35). México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México.
González-Santillán, E., Galán-Sánchez, M. A., & Valdez-Velázquez, L. L. (2019). A new species of Centruroides (Scorpiones, Buthidae) from Colima, Mexico. Comptes Rendus Biologies, 342, 9–10. https://doi.org/10.1016/j.crvi.2019.10.002
González-Santillán, E., & Possani, L. D. (2018). North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Tropica, 187, 264–274. https://doi.org/10.1016/j.actatropica.2018.08.002
González-Santillán, E., & Prendini, L. (2013). Redefinition and generic revision of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, with descriptions of six new genera. Bulletin of the American Museum of Natural History, 382, 1–71.
González-Santillán, E., & Prendini, L. (2015). Phylogeny of the North American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, based on morphology, mitochondrial and nuclear DNA. Cladistics, 31, 341–405. https://doi.org/10.1111/cla.12091
González-Santillán, E., & Prendini, L. (2018). Systematic revision of the North American Syntropine Vaejovid scorpion genera Balsateres, Kuarapu, and Thorellius, with descriptions of three new species. Bulletin of the American Museum of Natural History, 420, 1–81. https://doi.org/10.1206/0003-0090-420.1.1
Hoang, D. T., Chernomor, O., Haeseler, A. von, Minh, B. Q., & Vinh, L. S. (2018) UFBoot2: Improving the ultrafast bootstrap approximation. Molecular Biology and Evolution, 35, 518–522. https://doi.org/10.1093/molbev/msx281
Hoffmann, C. C. (1932). Monografías para la entomología médica de México. Monografía Núm. 2. Los scorpiones de Mexico. Segunda parte, Buthidae. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, 3, 243–361.
Huerta-Jiménez, H. (2018). Actualización de la Colección de Artrópodos con importancia médica (CAIM), Laboratorio de Entomología, InDRE. Version 1.5. Comisión Nacional para el Conocimiento y uso de la Biodiversidad.
Ibarra-Vega, R., Jiménez-Vargas, J. M., Pineda-Contreras, A., Martínez-Martínez, F. J., Barajas-Saucedo, C. E., García-Ortega, H. et al. (2023). Indolealkylamines in the venom of the scorpion Thorellius intrepidus. Toxicon, 233, 107232. https://doi.org/10.1016/j.toxicon.2023.107232
Jarvis, A., Reuter, H. I., Nelson, A. & Guevara, E. (2008). Hole-filled seamless SRTM data V4, International Centre for Tropical Agriculture (CIAT), available from http://srtm.csi.cgiar.org
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., Haeseler, A. von, & Jermiin, L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nature Methods, 14, 587–589. https://doi.org/10.1038/nmeth.4285
Katoh, K., Rozewicki, J., & Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization, Briefings in Bioinformatics, 20, 1160–1166. https://doi.org/10.1093/bib/bbx108
Kocher, T. D., Thomas, W. K., Meyer, A., Edwards, S. V., Paabo, S., Villablanca, F. X. et al. (1989). Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proceedings of the National Academy of Sciences, 86, 6196–6200. https://doi.org/10.1073%2Fpnas.86.16.6196
Kuraku, S., Zmasek, C. M., Nishimura, O., & Katoh, K. (2013). aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity, Nucleic Acids Research, 41, W22–W28. https://doi.org/10.1093/nar/gkt389
López-Granados, J. (2019). Análisis filogenético del complejo de especies occidentalis del género Mesomexovis González-Santillán y Prendini, 2013 (Scorpiones: Vaejovidae) basado en evidencia morfológica (Tesis). Facultad de Ciencias, Universidad Nacional Autónoma de México. CDMX.
Lourenço, W. R., & Sissom, W. D. (2000). Scorpiones. In B. J. Llorente, E. González, & N. Papavero (Eds.), Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia una síntesis de su conocimiento (pp. 115–135). Ciudad de México: Universidad Nacional Autónoma de México.
Maddison, W. P., & Maddison, D. R. (2023). Mesquite: a modular system for evolutionary analysis. Version 3.81. http://www.mesquiteproject.org
Martin, B. M., Carbone, E., Yatani, A., Brown, A. M., Ramírez, A. N., Gurrola, G. B. et al. (1998). Amino acid sequence and physiological characterization of toxins from the venom of the scorpion Centruroides limpidus tecomanus Hoffmann. Toxicon, 26, 785–94. https://doi.org/10.1016/0041-0101(88)90319-4
Mendoza, J., & Francke, O. (2017). Systematic revision of Brachypelma red-kneed tarantulas (Araneae: Theraphosidae), and the use of DNA barcodes to assist in the identification and conservation of CITES-listed species. Invertebrate Systematics, 31, 157–179. https://doi.org/10.1071/IS16023
Minh, B. Q., Hahn, M. W., & Lanfear, R. (2020). New methods to calculate concordance factors for phylogenomic datasets. Molecular Biology and Evolution, 37, 2727–2733. https://doi.org/10.1093/molbev/msaa106
Morrone, J. (2019). Regionalización biogeográfica y evolución biótica de México: encrucijada de la biodiversidad del Nuevo Mundo. Revista Mexicana de Biodiversidad, 90, e902980. https://doi.org/10.22201/ib.20078706e.2019.90.2980
Morrone, J., Escalante, T., & Rodríguez-Tapia, G. (2017). Mexican biogeographic provinces: map and shapefiles. Zootaxa, 4277, 277–279. https://doi.org/10.11646/zootaxa.4277.2.8
Nguyen, L. T., Schmidt, H. A., Haeseler, A. von, & Minh, B. Q. (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Molecular Biology and Evolution, 32, 268–274. https://doi.org/10.1093/molbev/msu300
Nunn, G. B., Theisen, B. F., Chirstensen, B., & Arctander, P. (1996). Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda. Journal of Molecular Evolution, 42, 211–223. https://doi.org/10.1007/bf02198847
Podnar, M., Grbac, I., Tvrtković, N., Hörweg, C., & Haring, E. (2021). Hidden diversity, ancient divergences, and tentative Pleistocene microrefugia of European scorpions (Euscorpiidae: Euscorpiinae) in the eastern Adriatic region. Journal of Zoological Systematics and Evolutionary Research, 59, 1824–1849. https://doi.org/10.1111/jzs.12562
Ponce-Saavedra, J., & Francke, O. F. (2013). Clave para la identificación de especies de alacranes del género Centruroides Marx 1890 (Scorpiones: Buthidae) en el Centro Occidente de México. Biológicas, 15, 52–62.
Ponce-Saavedra, J. y Francke, O. F. (2019). Una especie nueva de alacrán del género Centruroides (Scorpiones: Buthidae) del noroeste de México. Revista Mexicana de Biodiversidad, 90, e902660. https://doi.org/10.22201/ib.20078706e.2019. 90.2660
Ponce-Saavedra, J., Francke, O. F., Cano-Camacho, H., & Hernández-Calderón, E. (2009). Evidencias morfológicas y moleculares que validan como especie a Centruroidestecomanus (Scorpiones, Buthidae). Revista Mexicana de Biodiversidad, 80, 71–84. https://doi.org/10.22201/ib.20078706e.2009.001.585
Ponce-Saavedra, J., Francke, O. F., Quijano-Ravell, A. F., & Santillán, R. C. (2016). Alacranes (Arachnida: Scorpiones) de importancia para la salud pública de México. Folia Entomológica Mexicana, 2, 45–70.
Ponce-Saavedra, J., Jiménez, M. L., Quijano-Ravell, A. F., Vargas-Sandoval, M., Chamé-Vázquez, D., Palacios-Cardiel, C. et al. (2023). The Fauna of Arachnids in the Anthropocene of Mexico, Chapter 2(pp. 18–39) In R. W. Jones, C. P. Ornelas-García, R. Pineda-López, & F. Álvarez (Eds.), Mexican fauna in the Anthropocene. Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-031-17277-9
Ponce-Saavedra, J., Linares-Guillén, J. W., & Quijano-Ravell, A. F. (2022). Una nueva especie de alacrán del género Centruroides Marx (Scorpiones: Buthidae) de la costa Noroeste de México. Acta Zoológica Mexicana (nueva serie), 38, 1–24. https://doi.org/10.21829/azm.2022.3812517
Ponce-Saavedra, J., Quijano-Ravell, A. F., Teruel, R., & Francke, O. F. (2015). Redescription of Centruroides ornatus Pocock, 1902 (Scorpiones: Buthidae), a montane scorpion from Central Mexico. Revista Ibérica de Aracnología, 27, 81–89.
Prendini, L., Weygoldt, P., & Wheeler, W. C. (2005). Systematics of the Damon variegatus group of African whip spiders (Chelicerata: Amblypygi): evidence from behaviour, morphology and DNA. Organisms Diversity & Evolution, 5, 203–236. https://doi.org/10.1016/j.ode.2004.12.004
Quijano-Ravell, A. F., De Armas, L. F., Francke, O. F., & Ponce-Saavedra, J. (2019). A new species of the genus Centruroides Marx (Scorpiones, Buthidae) from western Michoacán state, México using molecular and morphological evidence. Zookeys, 859, 31–48. https://doi.org/10.3897/zookeys.859.33069
Quijano-Ravell, A. F., Ponce-Saavedra, J., Francke, O. F., & Villaseñor-Ramos. M. A. (2010). Nuevos registros y distribución actualizada de Centruroides tecomanus Hoffmann, 1931 (Scorpiones: Buthidae). Ciencia Nicolaita, 52, 179–189.
QGIS.org. (2021). QGIS Geographic Information System. Open-Source Geospatial Foundation Project. http://qgis.org
Ramírez-Ruiz, J. J., & Bretón-González, M. (2016). Fisiografía y geología. In La Biodiversidad en Colima. Estudio de estado (Eds.) (pp. 25–31). México D.F.: Conabio.
Romero-Gutiérrez, T., Peguero-Sánchez, E., Cevallos, M. A., Batista, C. V. F., Ortiz, E., & Possani L. D. (2017). A deeper examination of Thorellius atrox scorpion venom components with omics technologies. Toxins, 9, 399. https://doi.org/10.3390/toxins9120399
Ruiz-Vega, M. L., Hernández-Canchola, G., & León-Paniagua, L. (2018). Molecular systematics and phylogeography of the endemic Osgood’s deermouse Osgoodomys banderanus (Rodentia: Cricetidae) in the lowlands of western Mexico. Molecular Phylogenetics and Evolution, 127, 867–877. https://doi.org/10.1016/j.ympev.2018.06.034
Santibáñez-López, C. E., Francke, O. F., & Córdova-Athanasiadis, M. (2011). The genus Diplocentrus Peters (Scorpiones: Diplocentridae) in Morelos, Mexico. Revista Ibérica de Aracnología, 19, 3–3.
Santibáñez-López, C. E., Francke, O. F., & Prendini, L. (2014). Phylogeny of the North American scorpion genus Diplocentrus Peters, 1861 (Scorpiones: Diplocentridae) based on morphology, nuclear and mitochondrial DNA. Arthropod Systematics & Phylogeny, 72, 257–279. https://doi.org/10.3897/asp.72.e31789
Santibáñez-López, C., Francke, O., Ureta, C., & Possani, L. (2015). Scorpions from Mexico: from species diversity to venom complexity. Toxins, 8, 1–18. https://doi.org/10.3390/toxins8010002
Schramm, F. D., Valdez-Mondragón, A., & Prendini, L. (2021). Volcanism and palaeoclimate change drive diversification of the world’s largest whip spider (Amblypygi). Molecular Ecology, 30, 2872–2890. https://doi.org/10.1111/mec.15924
Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H., & Flook, P. K. (1994). Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Annals of the Entomological Society of America, 87, 651–700. https://doi.org/10.1093/aesa/87.6.651
Sissom, W. D. (2000). Family Vaejovidae Thorell, 1876. In V. Fet, W. D. Sissom, G. Lowe, & M. E. Braunwalder (Eds.), Catalog of the scorpions of the World (1758–1998) (pp. 503–553). New York: New York Entomological Society.
Towler, W. I. I., Ponce-Saavedra, J., Gantenbein, B., & Fet, V. (2001). Mitochondrial DNA reveals a divergent phylogeny in tropical Centruroides (Scorpiones: Buthidae) from Mexico. Biogeographica, 77, 157–172.
Valdez-Velázquez, L. L., Olamendi-Portugal, T., Restano-Cassulini, R., Zamudio, F. Z., & Possani, L. D. (2018). Mass fingerprinting and electrophysiological analysis of the venom from the scorpion Centruroides hirsutipalpus (Scorpiones: Buthidae). Journal of Venom Animal Toxins Including Tropical Diseases, 24, 17. https://doi.org/10.1186/s40409-018-0154-y
Valdez-Velázquez, L. L., Quintero-Hernández, V., Romero-Gutiérrez, M. T., Coronas, F. I., & Possani, L. D. (2013). Mass fingerprinting of the venom and transcriptome of venom gland of scorpion Centruroides tecomanus. Plos One, 8, e66486. https://doi.org/10.1371/journal.pone.0066486
Valdez-Velázquez L. L., Romero-Gutiérrez M. T., Delgado-Enciso, I., Dobrovinskaya, O., Melnikov, V., Quintero-Hernández, V. et al. (2016). Comprehensive analysis of venom from the scorpion Centruroides tecomanus reveals compounds with antimicrobial, cytotoxic, and insecticidal activities. Toxicon, 118, 95-103. https://doi.org/10.1016/j.toxicon.2016.04.046
Williams, S. C. (1980). Scorpions of Baja California, Mexico and adjacent islands. Occasional Papers of the California Academy of Sciences, 135, 1–127.
Williams, S. C. (1986). A new species of Vaejovis from Jalisco, Mexico (Scorpiones: Vaejovidae). Pan-Pacific Entomologist, 62, 355–358.